
Submitted:
27 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Neurodegenerative diseases, such as Alzheimer’s disease and Parkinson’s disease, are caused by a combination of multiple events that damage neuronal function. A well-characterized biomarker of neurodegeneration is proteinaceous aggregates accumulation in the brain. However, the gradually worsening symptoms of neurodegenerative diseases are unlikely to be solely due to the result of a mutation in a single gene, but rather a multi-step process involving epigenetic changes. Multiple epigenetic studies have revealed that modification of histones accompanied by both the local and global remodeling of the chromatin structure and alternations in transcriptional patterns are closely associated with the pathogenesis of neurodegenerative diseases. Unlike DNA mutations, epigenetic alterations are reversible, and therefore raise the possibilities for therapeutic intervention including dietary modifications. Additionally, reactive oxygen species may contribute to the pathogenesis of Alzheimer’s disease and Parkinson’s disease. Given that the antioxidant properties of plant-derived phytochemicals are likely to exhibit pleiotropic effects against ROS-mediated epigenetic alternation, dietary intervention may be promising for the management of neurodegeneration in these diseases. In this review, the state-of-the-art applications using single-cell multi-modal omics approaches including genetics, and epigenetics, and dietary approaches for the identification of novel biomarkers and therapeutic approaches for the treatment of neurodegenerative diseases are discussed.
Keywords:
Alzheimer’s disease
; Parkinson’s disease
; phytochemicals
; dietary intervention
; epigenetics
; transcriptome
; epigenome
; transcriptomics
; epigenomics
1. Introduction
Chronic neurodegenerative disorders, with the most common being Alzheimer’s disease (AD) and Parkinson’s disease (PD), are believed to be caused by a combination of multiple events that damage neuronal function [1]. They are predicted to affect public health worldwide, are predominantly observed in the elderly population, and are estimated to increase threefold by the middle of the 21st century [2]. Therefore, several efforts have focused on pursuing the identification of biomarkers and causative factors in the environment, lifestyle including diet, and genetic changes including epigenetics [3]. The most well-characterized biomarker for neurodegeneration is the accumulation of proteinaceous aggregates in the brain [4]. Accordingly, the definition of neurodegenerative diseases is descriptive, in that, the typical pathological feature of aggregation of specific proteins is their hallmark and is also used for their diagnosis and pathological classification [5]. However, the slowly progressing symptoms of neurodegenerative diseases are unlikely to be solely due to the result of a mutation in a single gene, but rather as a consequence of a multi-step process involving epigenetic changes [1].
Epigenetics is defined as the stable inheritance of a phenotype resulting from changes in the chromosomal DNA without mutations in the nucleotide sequence [6]. A number of epigenetic researches have revealed that the modification of histones accompanied by both a local and global remodeling of the chromatin structure and alternations in the transcriptional patterns are closely associated with the pathogenesis of neurodegenerative diseases including AD and PD [7], in particular, DNA methylation, gain or loss of heterochromatin, and nucleolar reorganization [8]. Thousands of epigenetic changes throughout the lifetime affect various cells and tissues and are therefore considered the key hallmarks of aging [9]. They involve alterations in the sites of DNA methylation by DNA methyltransferases; modification of histone proteins by histone acetylases, deacetylases, methylases, and demethylases; and structural remodeling of the chromatin [10]. In contrast to DNA mutations, epigenetic changes are theoretically reversible and therefore raise the possibility of targeting them for designing novel therapeutic interventions including dietary alterations. The typical, age-associated epigenetic changes such as an increase in H4K16 acetylation, and H4K20 and H3K4 methylation, or a decrease in H3K9 and H3K27 methylation, constitute the characteristic age-associated epigenetic markers [10]. In this review, the state-of-the-art applications using single-cell multi-omics approaches including genetics and epigenetics, and dietary approaches to identify novel biomarkers and therapeutic approaches for the management of neurodegeneration.
2. Epigenetics and Alzheimer’s disease
AD is a common type of neurodegenerative disorder, the typical diagnostic phenotypes of which are the enhanced deposition of senile plaques composed of insoluble, neurofibrillary proteins in the brain. These aggregates mainly consist of an early accumulation amyloid-β (Aβ) protein with an abnormal form of aggregation and hyperphosphorylated tau proteins in the hippocampus that eventually led to cognitive impairment over time [11]. The established biomarkers used at present, are Aβ1-42, total-tau, and phosphorylated-tau in the cerebrospinal fluid (CSF). Actual therapeutic and pharmacological interventions effective in the management of AD that impede the progress of the disease have not been developed, with the only intervention being restricted to symptomatic treatments that retard the progress of the disease [12]. Therefore, the establishment of new methods effective in treating AD and the unraveling of the neurodegenerative mechanism is of significance for biomedical researchers [13]. Though the stage of the disease and the mechanism by which neurodegeneration occurs during the pathogenesis of AD have remained unresolved, many trials have been performed to clarify the pathophysiological features of neuroinflammation in AD based on the multiple parameters concerning inflammatory mediators [14]. Additionally, oxidative stress may contribute to the progression and pathogenesis of AD [15]. Oxidative stress induced in the neuronal cells was mainly attributed to excess production of reactive oxygen species (ROS), which plays a key role in the progression of AD [16]. ROS refers to a family of ionic species continuously generated from O2 and scavenged within the cells. The major ROS include H2O2 (hydrogen peroxide), O3 (ozone), O2-•(superoxide anion radical), and OH• (hydroxyl radical) [14]. Dysregulation of ROS contributed to the pathogenesis of neurodegenerative diseases [17]. Increased ROS production adversely affected DNA function including its epigenetic modification related to AD [18]. An accumulating body of evidence implied that oxidative stress is the main AD risk factor for by inducing apoptosis of neurons and dysfunction of the brain at the initiation stage and throughout AD progression [19], and the DNA of hyper-methylated nucleotides was easily influenced by β-amyloid-promoted oxidative damage as methyl-cytosines restricted the repair of the adjacent hydroxy-guanosines [20] (Figure 1).
3. Epigenetics and Parkinson’s disease
PD is the most prevalent neurodegenerative disease with the characteristics of neuropsychiatric symptoms including depression and anxiety existing before the onset of symptoms related to motor and movement illnesses [21]. Most patients clinically present with motor disorder and suffer from slowness of movement, rest tremors, rigidity, and disturbances in balance; the major features include the substantia nigra (SN)’s dopaminergic neuron loss and Lewy body depositions [21]. Dysfunction of mitochondria, oxidative stress may cause the progression of PD [22]. The dopaminergic neuron activities in the SN is critical for striatal synaptic plasticity and positive learning, and their degeneration led to an enhancement of the subthalamic nucleus, that in turn amplified the excitation signals relayed to the SN [23]. Exposure to harmful mediators in the environment, such as ROS, might cause to the enhancement of neurodegeneration with clinical symptoms similar to those of PD [24]. Therefore, it is necessary to establish methods for ensuring the longevity of healthy neurons following the attack by ROS without using specific medications. A close association between genetics and the environmental factors may determine the fate of PD, and the participation of many networks involving DNA, proteins, organelles, and neural networks may contribute to the complexity in the manifestation of symptoms [25]. Many pathogenesis-related association have been focused for the management of symptoms and progression of PD [25]. While dopaminergic-based treatments have been the gold standard for the symptomatic control of PD, a few requirements for the management of dopaminergic-resistant motor/non-motor symptoms remain unaddressed and for treatments altering the normal clinical course of PD [25].
Recently, much efforts have been made for elucidating the epigenetics driving alternations of gene expression associated with the pathogenesis of PD. The alternations in gene expression are well known cause of PD, and epigenetics is likely to play a critical role in its regulation [26]. Therefore, an imbalanced epigenetic alternation may cause harmful effects. Environments may raise an objection for the formation and maintenance of epigenetics and may thereby fill a gap between the deeper understanding of the origin and pathogenesis of neurodegenerative diseases [27]. A recent study on genome-wide DNA methylation in brain and blood samples of PD patients revealed a characteristic methylated pattern involving a number of genes implicated in PD; thus the functional significance of epigenetics as a regulatory factor in PD is evident, implying that peripheral blood may be a promising source for detecting DNA methylation levels in addition to brain tissue for the discovery of biomarkers associated with PD [28] (Figure 1). Given that PD is a disease with many related disorders including a variety of clinical symptoms, epigenetic regulation, treatment response, and survival, possibly reflecting multimodal changes, i.e., genetic, epigenetic, proteinaceous, and organellar contributions; thus, much improvement of diagnostic procedures may improve the success of therapeutic methods, the mechanisms of action of which may beneficial for PD patients [29].
4. Multimodal genomic studies on neurodegenerative diseases
Considering the recent evolution in data analysis methods with the development of automated cell population identification software and machine learning algorithms, a proposal to set up a practical intelligent imaging flow cytometry system as a high-throughput biosensing tool with sustainable manufacturing was presented [30]. Flow cytometry is a commonly used analytical tool in the fields of biology and medicine; it utilizes an optical laser system to profile cells and rapidly accumulates data concerning multiple parameters, including cell morphology and patterns of protein expression from a heterogeneous mixture of live cells at the single-cell level. For example, in immunology, flow cytometry is used to identify and characterize a certain population or subpopulation of immune cell types [31]. This methodology may also be advantageous if applied to the field of epigenetics related to neurodegenerative diseases, to implement novel processes for the development of sustainable production strategies toward achieving an integrative healthcare approach that combines nutrigenetics and translational medicine. Flow cytometry measures the biochemical and morphological characters of a mass of cells in the form of a suspension, which are often labeled with fluorescence dyes. The suspension consisting of saline is injected and funneled through a nozzle in the flow cytometer, thereby focusing the particles or cells into a stream of single cells at the center of the sample flow stream [32]. This is accomplished through a hydrodynamic focusing effect caused by laminar flow resulting from a sample and sheath flow that forces the cells to pass through a laser beam inside the instrument. Using the technology, millions of particles or cells can be rapidly counted, and the acquired parameters are automatically saved on a computer. Flow cytometry involves photonic components, such as tightly focused laser beams that can illuminate a single particle or a cell, the cell scatters the laser light, and the ratio of the cell size to the wavelength of the laser measured by optical detectors determines their scattering behaviors. Accordingly, the light scatter ascertained on one of the two detectors measures the light scattered along the path of the laser beam, which is referred to as forward scatter (FSC). The intensity of FSC is proportional to the diameter of the cell. The light scatter in vertical orientation compared to the FSC is the side scatter (SSC) and reflects the internal complexity of a cell.
In the field of molecular biology, the fourth paradigm enables a comprehensive understanding of the biochemical processes conducted by complex networks involving biomolecular associations and genetically regulated patterns at the single cell level [33] (Figure 2). Thus, single-cell technologies can be applied for transcriptomic, epigenetic, and proteomic profiling which may contribute to the wider areas of healthcare-related studies, especially cancer, as in the case of the present review. The main focus would be on researches related to single cell/nucleus transcriptome and epigenome in humans and address the emerging techniques that bridge transcriptome and metabolome at the single cell level. Such single cell, multiomics methods are closely related to individual phenotype analysis and diagnosis and can be used in designing precision nutrition and functional medicine therapies to prevent and treat neurodegenerative diseases. This field of bioinformatics has emerged as a result of the human genome project and has enabled the sequencing of DNA at a large-scale employing high-throughput sequence methodologies [34]. This methodologies have opened the door for the systematic understanding of gene expression profiles of not only coding regions but also the non-coding RNAs (ncRNAs) including microRNA (miRNA) or long non-coding RNAs (lncRNAs); the regulatory elements in the 5’- and 3’- untranslated regions (UTRs); and post-transcriptional RNA modification/editing events such as nucleobase modifications, including the deamination of “cytidine to uridine” and “adenosine to inosine,” additions/insertions of nucleotide(s) [35]. These herald the next-generational shift in system biology, focusing on reconstruction of the associations for explaining the synaptic network functioning by employing machine learning techniques [36].
Many inventions and advancements have been accomplished in multiomics approaches. Single-Cell Assay for Transposase-Accessible Chromatin sequencing (scATAC-seq) method is the methods used to explore the accessibility to the genome-wide chromatin network in many cells at a single cell resolution [35] (Table 1). Cellular Indexing of Transcriptomes and Epitopes by sequencing (CITE-seq) protocol is a single cell technique that integrates protein measurements and transcriptomic analysis by high-throughput single-cell RNA sequencing (scRNA-seq) into an efficient, single cell readout [37]. RNA Expression And Protein sequencing (REAP-seq) assay is a recently developed multimodal single cell technique, that enables fine characterization of cell subtypes and functional states by measuring the expression levels of RNA and proteins [38].
These emerging techniques can bridge human transcriptome and proteome at the single cell level and these multimodal omics methodologies can be introduced for specific personal phenotyping and diagnosis for designing precise nutritional medicine. Flow cytometry has the potential to be extended on a single cell basis to a wide range of studies, including the determination of antigen expression either on the cell surface or within cells, DNA/RNA content analysis, and functional analysis. Although flow cytometry enables the high-throughput measurement of the characteristics of one cell at a time, a high signal to noise ratio is a critical requirement [39].
Powerful methods for the characterization and sorting of cells by employing spatially and high time-resolved data including imaged flow cytometer have been designed [40]. However, conventional flow cytometry-based medical diagnostics is seldom used in the clinical setting currently. The advancements in image-based cell sorting technology significantly expand its applicability in functional genomics. This methodology enabled image-based high-throughput cell sorting and multicolor imaging, ensuring a broader availability in various fields, including basic biological research and medical diagnostics, with significant potential applicability in multimodal omics readouts [41]. Flow cytometry imaging obtains multi-channel image characteristics of cells through the detectors located in image platforms that are equipped with a complementary metal oxide semiconductor sensors to precisely capture images of individual cells as they flow through [41]. This intelligent, image-activated cell sorter can be integrated seamlessly with high-throughput, single-cell multi-omics analysis tools. Combining these analytical techniques with computational approaches for the analyses and integration of single cell multiomics data across multiple modalities can open new doors for the precise reconstruction of the genetic regulation and signal networks driving the identity and function of specific cells.
Machine learning, the utilization of trainable statistical models used to recognize patterns and predict future behavior, is a promising methodology for analyzing the data accumulated from the end-users of newly developed flow cytometry technologies, which are applicable for research into neurodegenerative diseases [42]. Machine learning methods range from simple linear regression to deep neural networks used for microfluidic applications in biology. Machine learning is often employed for the analysis of flow cytometric images to isolate from the mixture cells to single cells. In image-based cell sorting, certain populations of cells are sorted according to their morphological characters and thereby characterized for the genomic, epigenomic, transcriptomic, proteomic, and other morphological properties.
Numerous fluorochromes are available for staining the cells before flow cytometric analysis; thus, multicolor, that is, multi-parameter, flow cytometry has become a powerful tool capable of generating large amounts of data. Therefore, automated tools are being designed for the analysis of big data sets. One of the most widely used tools in flow cytometry is t-stochastic neighbor embedding (t-SNE) [43]. t-SNE scoring is useful for visualizing high dimensional big datasets. t-SNE scoring enables conversion of the similarity between each point into joint probabilities and then minimizes the divergence (Kullback–Leibler divergence) among the joint probabilities of low dimensional embedding and high dimensional data [40]. Another dimension reduction tool is uniform manifold approximation and projection which can be used for visualization like t-SNE but is more useful in general non-linear dimension reduction [44].
Single cell-imaged sorting by flow cytometry is indispensable techniques in molecular biology that can be used synergistically with fluorescence markers, microfluidics, and machine learning [45]. Machine learning is an application of artificial intelligence (AI) that includes comprehensive, automated analytical modeling approaches that enable machines to execute decisions by automatically learning from the available data. AI is more multifaceted than the simple statistical models being used for biological datasets. The field of bioinformatics has also emerged and gained prominence along with the human genome project and has realized the fine reading of large DNA molecules using high-throughput sequencing techniques that have opened the door for the systematic understanding of not only protein-coding gene expression profiles, but also of ncRNAs including miRNAs and lncRNAs; the regulatory elements in the 5′- and 3′- UTRs; and post-transcriptional RNA modification/editing events, including nucleobase modifications, including the deamination of cytidine and adenosine. This shift to the next generation in systems biology aims to reconstruct the interactions for explaining the synergistic networks by introducing machine learning [46].
5. Dietary approaches to impede neurodegeneration
The mutual relation between diet/nutrition and the immune system plays an important role in maintenance of human body. Systematic determination of the full range/variety of cells has been the challenge due to limitations of analytical methods that have restricted the parameters that can be determine. Recently, the single cell techniques have overwhelmed these limitations by their high-throughputness [47]. The interactions also contribute to the progression of diseases including neurodegenerative diseases. Therefore, along with these big international projects, a healthcare-based approach integrating the single cell techniques may offer deeper insights into the current state of studies on nutritional and molecular medicine and serve to discover and develop new clinical interventions for the treatments of diseases shortly [48]. For example, coffee consumption ameliorated cognitive impairment induced by AD [49]. Similarly, a negative association between the moderate consumption of coffee and the risk of age-related cognitive disorders and PD was reported [50]. Coffee consumption has been also linked to potential health benefits owing to its chemopreventive and anti-inflammatory effects. It has been suggested that a reduction in inflammation by coffee was attributable to the antioxidative features of certain ingredients.
6. Conclusions and future directions
Neurodegeneration during the course of AD and PD pathogenesis are caused by a combination of multiple events that damage neuronal function. Although a well-characterized biomarker of such neurodegeneration is proteinaceous aggregates accumulation in the brain, the gradually worsening symptoms of neurodegenerative diseases are unlikely to be solely due to the result of a mutation in a single gene, but rather a multi-step process involving epigenetic changes.
Technology serves as a strong partner in biochemistry and molecular medicine, allowing discoveries that were previously beyond the reach of researchers. Path-breaking exploration is possible with the improvement and innovation of novel techniques/tools. Advances in technology have also been inspired by the systems and processes of organisms or the need to establish a novel methodology to solve a biological problem. Such a relationship is powerful when applied to the synergistic exploration of fundamental problems in biochemistry and molecular medicine. Currently, improvements in the visualization and manipulation of biological systems as well as analyzing and integrating big data and the scope of the discovery are important. Studies on neurodegenerative diseases have developed through the use of advanced technologies; recently, improved sequencing techniques and single-cell/single-nucleus technology enabled the analysis of multiple human-associated species for transcriptomic studies. These methodologies have become routine in recent studies concerning neurodegenerative diseases; single-cell technologies have revolutionized the studies with much improvement in the throughput. These analytical methods used for neurodegenerative diseases overcome one of the challenges that differ among individuals and epigenetic complexity. Unlike DNA mutations, epigenetic alterations are reversible, and therefore raise the possibilities for therapeutic intervention including dietary modifications. Additionally, reactive oxygen species may contribute to the pathogenesis of Alzheimer’s disease and Parkinson’s disease. Given that the antioxidant properties of plant-derived phytochemicals are likely to exhibit pleiotropic effects against ROS-mediated epigenetic alternation, dietary intervention may be promising for the management of neurodegeneration in these diseases.Thus, the recent innovations including single-cell methods have much potential for applicability in the analysis of neurodegeneration.
Author Contributions
Conceptualization, T.M., S.M.; writing—original draft preparation, T.M.; supervision, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were obtained or analyzed in this study. Data sharing does not apply to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Aβ | amyloid-β |
| AD | Alzheimer’s disease |
| AI | artificial intelligence |
| ATAC-seq | Assay for Transposase-Accessible Chromatin sequencing |
| CITE-seq | Cellular Indexing of Transcriptomes and Epitopes by sequencing |
| CSF | erebrospinal fluid |
| lncRNA | long non-coding RNA |
| miRNA | microRNA |
| ncRNA | non-coding RNA |
| PD | Parkinson’s disease |
| REAP-seq | RNA Expression And Protein sequencing |
| ROS | reactive oxygen species |
| scRNA-seq | single-cell RNA sequencing |
| scATAC-seq | Single-Cell Assay for Transposase-Accessible Chromatin sequencing |
| SN | substantia nigra |
| t-SNE | t-stochastic neighbor embedding |
| UTR | untranslated region |
References
- Bossy-Wetzel, E.; Schwarzenbacher, R.; Lipton, SA. Molecular pathways to neurodegeneration. Nat. Med. 2004, 10, S2–9. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Hansson, O. Biomarkers for neurodegenerative diseases. Nat. Med. 2021, 27, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Teunissen, C.E.; Verberk, I.M.W.; Thijssen, E.H.; Vermunt, L.; Hansson, O.; Zetterberg, H.; van der Flier, W.M.; Mielke, M.M.; Del Campo, M. Blood-based biomarkers for Alzheimer’s disease: towards clinical implementation. Lancet Neurol. 2022, 21, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Dubois, B.; Villain, N.; Frisoni, G.B.; Rabinovici, G.D.; Sabbagh, M.; Cappa, S.; Bejanin, A.; Bombois, S.; Epelbaum, S.; Teichmann, M.; Habert, M.O.; Nordberg, A.; Blennow, K.; Galasko, D.; Stern, Y.; Rowe, C.C.; Salloway, S.; Schneider, L.S.; Cummings, J.L.; Feldman, H.H. Clinical diagnosis of Alzheimer’s disease: recommendations of the International Working Group. Lancet Neurol. 2021, 20, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef]
- Sen, P.; Shah, P.P.; Nativio, R.; Berger, S.L. Epigenetic mechanisms of longevity and aging. Cell 2016, 166, 822–839. [Google Scholar] [CrossRef]
- Wilson, D.M. 3rd; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. 3rd; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Hallmarks of neurodegenerative diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–217. [Google Scholar] [CrossRef]
- Migliore, L.; Coppedè, F. Gene-environment interactions in Alzheimer disease: the emerging role of epigenetics. Nat. Rev. Neurol. 2022, 18, 643–660. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Goedert, M. Tau pathology and neurodegeneration. Lancet Neurol. 2013, 12, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.; Nakanishi, A.; Murai, T. Implications of PI3K/AKT/PTEN signaling on superoxide dismutases expression and in the pathogenesis of Alzheimer’s disease. Diseases 2018, 6, E28. [Google Scholar] [CrossRef] [PubMed]
- Murai, T.; Matsuda, S. Therapeutic implications of probiotics in the gut microbe-modulated neuroinflammation and progression of Alzheimer’s disease. Life 2023, 13, 1466. [Google Scholar] [CrossRef]
- Halliwell, B. Oxidative stress and neurodegeneration: where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Contestabile, A. Oxidative stress in neurodegeneration: mechanisms and therapeutic perspectives. Curr. Top. Med. Chem. 2001, 1, 553–568. [Google Scholar] [CrossRef]
- Murai, T.; Matsuda, S. Pleiotropic signaling by reactive oxygen species concerted with dietary phytochemicals and microbial-derived metabolites as potent therapeutic regulators of the tumor microenvironment. Antioxidants 2023, 12, 1056. [Google Scholar] [CrossRef]
- Ischiropoulos, H.; Beckman, J.S. Oxidative stress and nitration in neurodegeneration: cause, effect, or association? J. Clin. Invest. 2003, 111, 163–169. [Google Scholar] [CrossRef]
- Baker, D.J.; Petersen, R.C. Cellular senescence in brain aging and neurodegenerative diseases: evidence and perspectives. J. Clin. Invest. 2018, 128, 1208–1216. [Google Scholar] [CrossRef]
- Zawia, N.H.; Lahiri, D.K.; Cardozo-Pelaez, F. Epigenetics, oxidative stress, and Alzheimer disease. Free Radic. Biol. Med. 2009, 46, 1241–1249. [Google Scholar] [CrossRef]
- Dauer, W.; Przedborski, S. Parkinson’s disease: mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed]
- Borsche, M.; Pereira, S.L.; Klein, C.; Grünewald, A. Mitochondria and Parkinson’s disease: Clinical, molecular, and translational aspects. J. Parkinsons Dis. 2021, 11, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Kang, X.; Hu, J.; Zhang, D.; Liang, Z.; Meng, F.; Zhang, X.; Xue, Y.; Maimon, R.; Dowdy, S.F.; Devaraj, N.K.; Zhou, Z.; Mobley, W.C.; Cleveland, D.W.; Fu, X.D. Reversing a model of Parkinson’s disease with in situ converted nigral neurons. Nature 2020, 582, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Nakano, N.; Matsuda, S.; Ichimura, M.; Minami, A.; Ogino, M.; Murai, T.; Kitagishi, Y. PI3K/AKT signaling mediated by G,protein-coupled receptors is involved in neurodegenerative Parkinson’s disease. Int. J. Mol. Med. 2017, 39, 253–260. [Google Scholar] [CrossRef]
- Elkouzi, A.; Vedam-Mai, V.; Eisinger, R.S.; Okun, M.S. Emerging therapies in Parkinson disease - repurposed drugs and new approaches. Nat. Rev. Neurol. 2019, 15, 204–223. [Google Scholar] [CrossRef]
- Labbé, C.; Lorenzo-Betancor, O.; Ross, O.A. Epigenetic regulation in Parkinson’s disease. Acta Neuropathol. 2016, 132, 515–530. [Google Scholar] [CrossRef]
- Pavlou, M.A.S.; Outeiro, T.F. Epigenetics in Parkinson’s Disease. Adv Exp Med Biol. 2017, 978, 363–390. [Google Scholar]
- Masliah, E.; Dumaop, W.; Galasko, D.; Desplats, P. Distinctive patterns of DNA methylation associated with Parkinson disease: identification of concordant epigenetic changes in brain and peripheral blood leukocytes. Epigenetics 2013, 8, 1030–1038. [Google Scholar] [CrossRef]
- Espay, A.J.; Brundin, P.; Lang, A.E. Precision medicine for disease modification in Parkinson disease. Nat. Rev. Neurol. 2017, 13, 119–126. [Google Scholar] [CrossRef]
- Schraivogel, D.; Kuhn, T.M.; Rauscher, B.; Rodríguez-Martínez, M.; Paulsen, M.; Owsley, K.; Middlebrook, A.; Tischer, C.; Ramasz, B.; Ordoñez-Rueda, D.; Dees, M.; Cuylen-Haering, S.; Diebold, E.; Steinmetz, L.M. High-speed fluorescence image-enabled cell sorting. Science 2022, 375, 315–320. [Google Scholar] [CrossRef]
- Filby, A.; Carpenter, A.E. A new image for cell sorting. N. Engl. J. Med. 2022, 386, 1755–1758. [Google Scholar] [CrossRef] [PubMed]
- LaBelle, C.A.; Massaro, A.; Cortés-Llanos. B.; Sims, C.E.; Allbritton, N,L. Image-based live cell sorting. Trends Biotechnol. 2021, 39, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sun, Z.; Yin, H.; Liu, X.; Wang, J.; Zhao, H.; Pang, C.H.; Wu, T.; Li, S.; Yin, Z.; Yu, X.F. Data-driven materials innovation and applications. Adv. Mater. 2022, 34, e2104113. [Google Scholar] [CrossRef] [PubMed]
- Steponaitis, G.; Stakaitis, R.; Valiulyte, I.; Krusnauskas, R.; Dragunaite, R.; Urbanavičiūtė, R.; Tamasauskas, A.; Skiriute, D. Transcriptome-wide analysis of glioma stem cell specific m6A modifications in long-non-coding RNAs. Sci. Rep. 2022, 12, 5431. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro, J.D.; Wu, B.; Litzenburger, U.M.; Ruff, D.; Gonzales, M.L.; Snyder, M.P.; Chang, H.Y.; Greenleaf, W.J. Single-cell chromatin accessibility reveals principles of regulatory variation. Nature 2015, 523, 486–490. [Google Scholar] [CrossRef]
- Narayanan, H.; Dingfelder, F.; Butté, A.; Lorenzen, N.; Sokolov, M.; Arosio, P. Machine learning for biologics: opportunities for protein engineering, developability, and formulation. Trends Pharmacol. Sci. 2021, 42, 151–165. [Google Scholar] [CrossRef]
- Stoeckius, M.; Hafemeister, C.; Stephenson, W.; Houck-Loomis, B.; Chattopadhyay, P.K.; Swerdlow, H.; Satija, R.; Smibert, P. Simultaneous epitope and transcriptome measurement in single cells. Nat. Methods 2017, 14, 865–868. [Google Scholar] [CrossRef]
- Peterson, V.M.; Zhang, K.X.; Kumar, N.; Wong, J.; Li, L.; Wilson, D.C.; Moore, R.; McClanahan, T.K.; Sadekova, S.; Klappenbach, J.A. Multiplexed quantification of proteins and transcripts in single cells. Nat. Biotechnol. 2017, 35, 936–939. [Google Scholar] [CrossRef]
- McIntyre, D.; Lashkaripour, A.; Fordyce, P.; Densmore, D. Machine learning for microfluidic design and control. Lab Chip 2022, 22, 2925–2937. [Google Scholar] [CrossRef]
- Hudson, W.H.; Wieland, A. Technology meets TILs: deciphering T cell function in the -omics era. Cancer Cell 2023, 41, 41–57. [Google Scholar] [CrossRef]
- Ahmad, A.; Sala, F.; Paiè, P.; Candeo, A.; D’Annunzio, S.; Zippo, A.; Frindel, C.; Osellame, R.; Bragheri, F.; Bassi, A.; Rousseau, D. On the robustness of machine learning algorithms toward microfluidic distortions for cell classification via on-chip fluorescence microscopy. Lab Chip 2022, 22, 3453–3463. [Google Scholar] [CrossRef] [PubMed]
- Myszczynska, M.A.; Ojamies, P.N.; Lacoste, A.M.B.; Neil, D.; Saffari, A.; Mead, R.; Hautbergue, G.M.; Holbrook, J.D.; Ferraiuolo, L. Applications of machine learning to diagnosis and treatment of neurodegenerative diseases. Nat. Rev. Neurol. 2020, 16, 440–456. [Google Scholar] [CrossRef] [PubMed]
- van der Maaten, L.J.P.; Hinton, G.E. Visualizing high-dimensional data using t-SNE. J. Machine Learning Res. 2008, 9, 2579–2605. [Google Scholar]
- Zhao, K.; Grayson, J.M.; Khuri, N. Multi-objective genetic algorithm for cluster analysis of single-cell transcriptomes. J. Pers. Med. 2023, 13, 183. [Google Scholar] [CrossRef]
- Ganesh, G.; Kolusu, A.S.; Prasad, K.; Samudrala, P.K.; Nemmani, K.V.S. Advancing health care via artificial intelligence: from concept to clinic. Eur. J. Pharmacol. 2022, 934, 175320. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Teichmann, SA. Computational methods for single-cell omics across modalities. Nat. Methods 2020, 17, 14–17. [Google Scholar] [CrossRef]
- Zare Harofte, S.; Soltani, M.; Siavashy, S.; Raahemifar, K. Recent advances of utilizing artificial intelligence in lab on a chip for diagnosis and treatment. Small 2022, 18, e2203169. [Google Scholar] [CrossRef]
- Pratapa, A.; Doron, M.; Caicedo, J.C. Image-based cell phenotyping with deep learning. Curr. Opin. Chem. Biol. 2021, 65, 9–17. [Google Scholar] [CrossRef]
- Murai, T.; Matsuda, S. The chemopreventive effects of chlorogenic acids, phenolic compounds in coffee, against inflammation, cancer, and neurological diseases. Molecules 2023, 28, 2381. [Google Scholar] [CrossRef]
- Murai, T.; Matsuda, S. Fatty acid metabolites and the tumor microenvironment as potent regulators of cancer stem cell signaling. Metabolites 2023, 13, 709. [Google Scholar] [CrossRef]
Figure 1.
Multimodal genomics and dietary approaches for the management of Neurodegenerative diseases including Alzheimer’s disease (AD) and Parkinson’s disease (PD). Multiple epigenetic studies have revealed that modification of histones accompanied by both the local and global remodeling of the chromatin structure and alternations in transcriptional patterns are closely associated with the pathogenesis of neurodegenerative diseases. ROS is the main AD risk factor for by inducing apoptosis of neurons and dysfunction of the brain at the initiation stage and throughout AD progression, and the DNA of hyper-methylated nucleotides was easily influenced by β-amyloid-promoted oxidative damage. Methylation of a number of genes are also associated with PD. Epigenetic changes involve alterations in the sites of DNA methylation by DNA methyltransferases; modification of histone proteins by histone acetylases, deacetylases, methylases, and demethylases; and structural remodeling of the chromatin. Unlike DNA mutations, epigenetic alterations are thought to be reversible, and therefore raise the possibilities for therapeutic intervention including dietary modifications.
Figure 1.
Multimodal genomics and dietary approaches for the management of Neurodegenerative diseases including Alzheimer’s disease (AD) and Parkinson’s disease (PD). Multiple epigenetic studies have revealed that modification of histones accompanied by both the local and global remodeling of the chromatin structure and alternations in transcriptional patterns are closely associated with the pathogenesis of neurodegenerative diseases. ROS is the main AD risk factor for by inducing apoptosis of neurons and dysfunction of the brain at the initiation stage and throughout AD progression, and the DNA of hyper-methylated nucleotides was easily influenced by β-amyloid-promoted oxidative damage. Methylation of a number of genes are also associated with PD. Epigenetic changes involve alterations in the sites of DNA methylation by DNA methyltransferases; modification of histone proteins by histone acetylases, deacetylases, methylases, and demethylases; and structural remodeling of the chromatin. Unlike DNA mutations, epigenetic alterations are thought to be reversible, and therefore raise the possibilities for therapeutic intervention including dietary modifications.
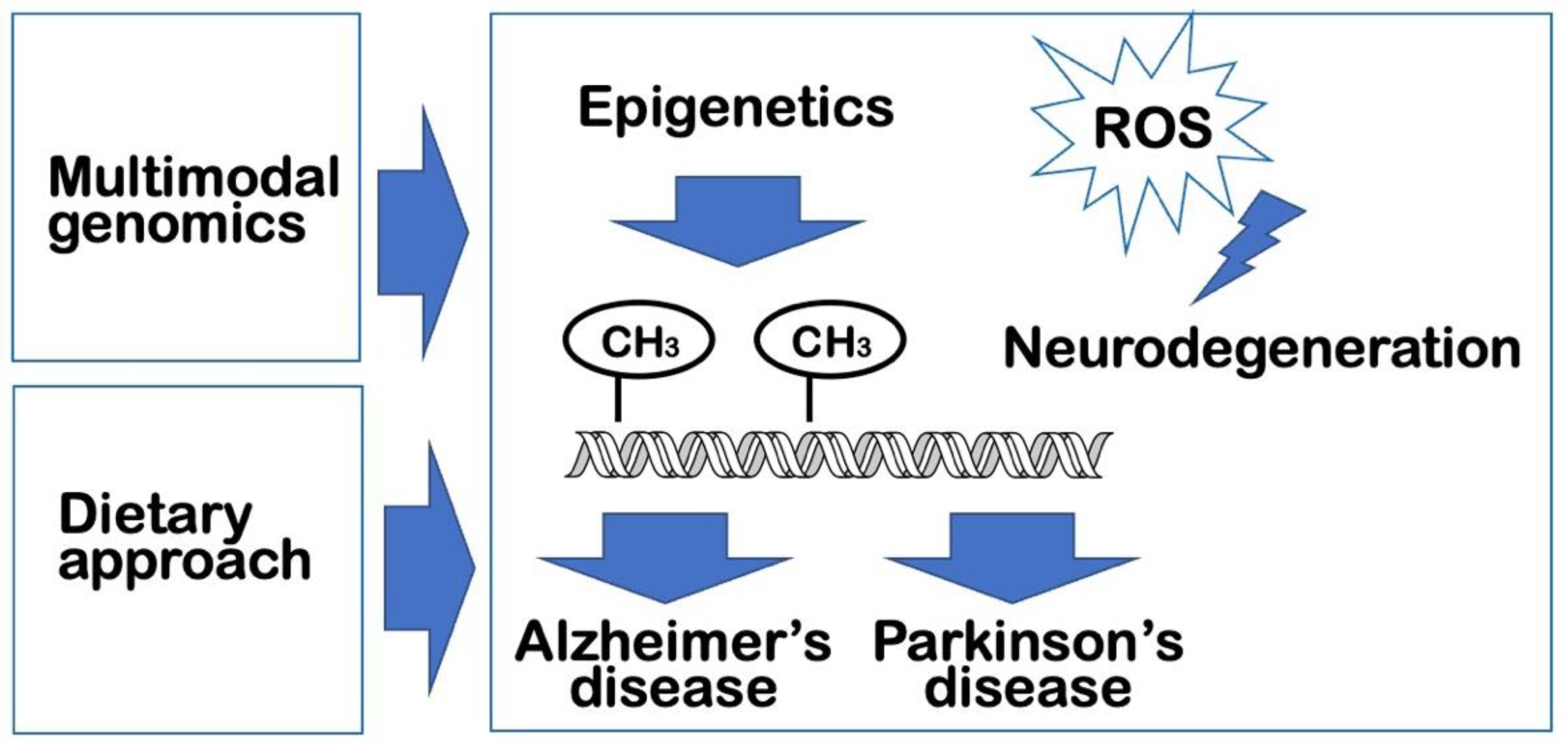
Figure 2.
Multimodal approaches to studies in neurodegeneration utilizing the emerging technologies including transcriptome, epigenome, and proteome analyses. Recently evolved single-cell technologies have enabled a comprehensive understanding of the biological processes orchestrated by complex networks involving biomolecular associations and genetically regulated patterns at the single cell level. Thus, single-cell technologies can be applied for transcriptomic, epigenetic, and proteomic profiling which may contribute to the wider areas of healthcare-related studies, especially neurodegenerative diseases including AD and PD. The single-cell or single-nucleus transcriptome, epigenome, and proteome in humans can be harnessed into multi-modal omics analyses that are closely related to individual phenotype analysis and diagnosis and can be used in designing precision nutrition and functional medicine therapies to prevent and treat neurodegenerative diseases. These herald the next-generational shift in system biology, focusing on reconstruction of the associations for explaining the synaptic network functioning in neurodegenerative diseases.
Figure 2.
Multimodal approaches to studies in neurodegeneration utilizing the emerging technologies including transcriptome, epigenome, and proteome analyses. Recently evolved single-cell technologies have enabled a comprehensive understanding of the biological processes orchestrated by complex networks involving biomolecular associations and genetically regulated patterns at the single cell level. Thus, single-cell technologies can be applied for transcriptomic, epigenetic, and proteomic profiling which may contribute to the wider areas of healthcare-related studies, especially neurodegenerative diseases including AD and PD. The single-cell or single-nucleus transcriptome, epigenome, and proteome in humans can be harnessed into multi-modal omics analyses that are closely related to individual phenotype analysis and diagnosis and can be used in designing precision nutrition and functional medicine therapies to prevent and treat neurodegenerative diseases. These herald the next-generational shift in system biology, focusing on reconstruction of the associations for explaining the synaptic network functioning in neurodegenerative diseases.
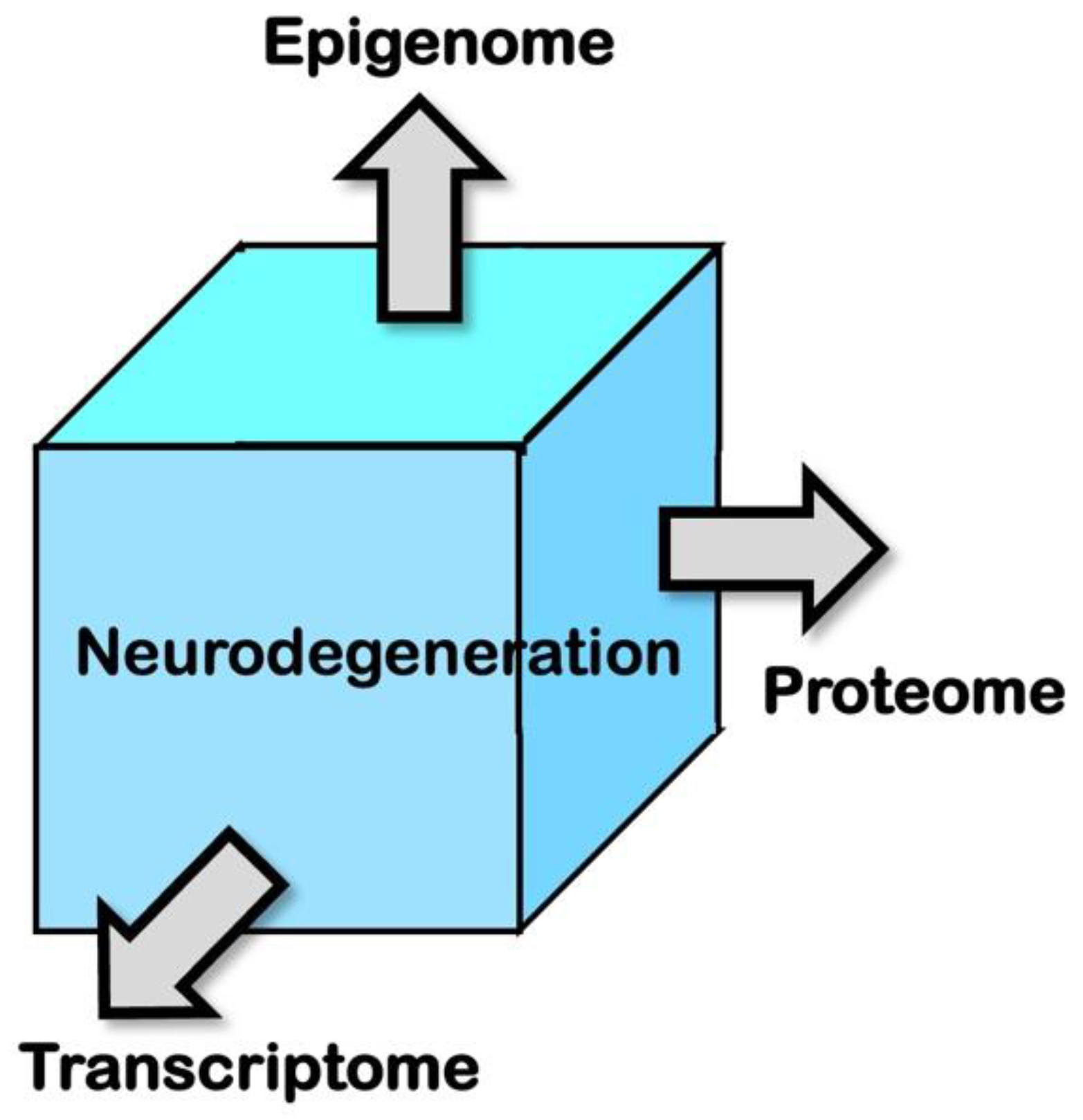

Table 1.
Representative single-cell approaches applicable to multimodal analysis of neurodegenerative diseases.
Table 1.
Representative single-cell approaches applicable to multimodal analysis of neurodegenerative diseases.
| Methods | Targets | References |
|---|---|---|
| scATAC-seq1 | Epigenome | 35 |
| CITE-seq2 | Transcriptome and proteome | 37 |
| REAP-seq3 | Transcriptome and proteome | 38 |
1scATAC-seq, Single-Cell Assay for Transposase-Accessible Chromatin sequencing. 2CITE-seq, Cellular Indexing of Transcriptomes and Epitopes by sequencing. 3REAP-seq, RNA Expression And Protein sequencing.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



