
Submitted:
27 July 2023
Posted:
28 July 2023
You are already at the latest version
Abstract
Hepatocellular carcinoma (HCC) is one of the most frequent cancers in humans, characterised by high resistance to conventional chemotherapy, late diagnosis, and high mortality rate. It is necessary to elucidate the molecular mechanisms involved in hepatocarcinogenesis to improve diagnosis and treatment outcomes. The Runt-related (RUNX) family of transcription factors (RUNX1, RUNX2, and RUNX3) participates in cardinal biological processes and play paramount roles in the pathogenesis of numerous human malignancies. Their role is often controversial as they can act as oncogenes or tumour suppressors, dependent on cellular context. Evidence shows that deregulated RUNX genes may be involved in hepatocarcinogenesis from the earliest to the latest stages. In this review, we summarise the topical evidence on the roles of RUNX gene family members in HCC. We discuss their possible application as non-invasive molecular markers for early diagnosis, prognosis, and development of novel treatment strategies in HCC patients.
Keywords:
RUNX
; hepatocellular carcinoma
; oncogenes
; tumour suppressors
; biomarkers
1. Introduction
Hepatocellular carcinoma is a primary liver cancer and one of the leading causes of mortality with wide geographic variation [1]. Any factor that leads to chronic liver injury and cirrhosis can be considered an oncogenic agent. Prevalent HCC risk factors are Hepatitis viruses B and C infection, excessive alcohol intake, nonalcoholic steatohepatitis (NASH), and aflatoxin B1 exposure [2,3].
The major problem with the HCC treatment is the diagnosis is usually made in progressed disease stadiums when conventional systemic chemotherapy is ineffective [4]. To overcome this problem, researchers are developing molecularly targeted treatments that represent a more promising approach for advanced HCC. Several such drugs are clinically available, but their efficacy is limited [5]. Therefore, elucidating the molecular processes at the basis of hepatocarcinogenesis is critical for improving therapy outcomes and diagnosis.
The family of Runt-related (RUNX) genes (RUNX1, RUNX2, and RUNX3) is crucial for the different tumour types' development and progression. RUNX genes are transcription factors that behave in opposing ways, promoting or suppressing tumourigenesis [6,7,8].
1.1. RUNX genes’ and proteins’ structure
Human RUNX genes participate in neuro-, blood, and bone development [7,8]. The role in developmental processes implies tight control of RUNX genes at transcriptional and posttranscriptional levels. RUNX genes' chromosome location differs depending on the species, and human RUNX1, RUNX2, and RUNX3 genes’ locations are on chromosomes 21, 6, and 1, respectively. Two alternative promoters of RUNX genes, P1 and P2, generate two dominant isoforms with differing 5'-untranslated regions (5'-UTRs) (Figure 1A). These transcripts give two polypeptides with different N-terminal sequences, distal and proximal [9]. The diversity of RUNX transcripts is the result of alternative splicing. RUNX1 and RUNX2 have nine exons and twelve isoforms, whereas RUNX3 has six exons and two isoforms [7,10]. RUNX proteins' molecular weight is 44, 50 and 57 KDa [7].
RUNX proteins got their name from the Runt homology domain (RHD) on the protein N-terminus, which has approximately 90% sequence homology in all three proteins [6]. The RHD domain binds to the target genes' DNA through the consensus sequence of seven nucleotides, 'PyGPyGGTPy'. Nuclear localisation and interaction with other proteins are also the functions of the RHD domain [10]. The RUNX proteins’ C-terminus encompasses the transactivation domain (TAD), and inhibitory domain (ID), with the consensus sequence of five amino acids, valine-tryptophan-arginine-proline-tyrosine (VWRPY). This sequence recruits Groucho/Transducin-like enhancer protein (TLE) corepressors [7,10,11]. TLE corepressors further control several target genes’ transcription. The TAD domain is the place of RUNX proteins’ interactions with the other regulatory proteins and control of their transcriptional activity [12]. In complex with the coregulators, RUNX proteins influence various cell processes [10] (Figure 1B).
1.2. The role of the RUNX genes in normal development
RUNX genes engage in normal cell development and differentiation processes, playing a tissue-specific role [14]. RUNX1 is crucial for cell growth and differentiation of immune cells, epithelial stem and epithelial cells, and neurodevelopment [11,15]. RUNX1 acts in the development of hematopoietic cells in vertebrates [13,16]. Consequently, the chromosomal rearrangements and somatic and germline mutations involving RUNX1 lead to various leukaemia types [13]. RUNX2 is a factor in bone generation [11]. The RUNX2 knockout mice lack osteoblast differentiation, leading to osteoporosis [17]. RUNX3 is essential for embryogenesis, and RUNX3 knockout mice die soon [18]. Additional studies have shown that RUNX3 participated in nervous and gastrointestinal system development, bone, and immune cells [11,19]. We used multi-omics datasets from the atlas of the healthy human liver [20] to examine RUNX-specific tissue expression in different liver cell compartments. According to the Liver Cell Atlas datasets, the overall RUNX1, RUNX2, and RUNX3 expression levels were 14.9%, 7.6%, and 25.1%, respectively [21].
1.3. The role of RUNX genes in cancer
RUNX genes’ mutations and abnormal expression lead to various cancer types. RUNX genes can hinder or activate tumourigenesis [12]. A recent study by Pan and colleagues [22] suggests that RUNX genes’ aberrant expression causes disparate cancer types and influences disease prognosis. Also, the co-expression of RUNX genes with epigenetic regulators could affect the onset of some cancer types. The researchers should pay attention to epigenetic mechanisms of RUNX genes’ regulation, including DNA methylation and miRNAs [22]. As epigenetic regulators, RUNX proteins cooperate with diverse coregulators and involve many signal transduction processes. Besides, they participate in chromatin landscape remodeling [10].
The previously mentioned RUNX genes’ functions indicate that they are of great importance for further investigation of HCC. With this in mind, this review collects the existing data and questions the possibility of including RUNX genes as the diagnostic, predictive or prognostic biomarkers in HCC treatment.
2. RUNX1 IN HCC
The RUNX1 gene belongs to a Runt-related gene family on chromosome 21, coding for the RUNX1 protein [23]. The researchers characterised the RUNX1 gene for the first time in chromosomal translocation t(8;21) of the acute myeloid leukaemia gene 1 (AML1) in AML cancer patients [24]. Two promoters, P1 (distal) and P2 (proximal) regulate the transcription of the RUNX1 gene, forming two isoforms which differ in the first exon [25]. The researchers have discovered twelve transcription variants of the RUNX1 mRNA [7].
2.1. RUNX1 role in HCC
The RUNX1 role in solid tumours is controversial, acting in two opposite ways. It could impede or promote cancerogenesis, as reviewed in [7,26]. Databases that collect transcriptomic studies and link data on the genomic and clinical parameters with various cancer groups show controversial results on the RUNX1 expression. According to the Gent2 and TIMER2.0 databases, there was a significantly higher expression of RUNX1 transcript in hepatocellular carcinoma compared to normal tissue [27,28]. However, the UCSC database shows the opposite results [29]. It has an upgraded version [30]. The RUNX1 expression was decreased in hepatocellular carcinoma. That was consistent with the work of Miyagawa and colleagues, who noticed that RUNX1 mRNA was 76% and 47% lower in HCC and cirrhotic tissue than in normal tissue. Also, there was a significant decrease of RUNX1 mRNA in HCC compared to cirrhotic liver samples [31]. Liu and colleagues also noticed the RUNX1 transcript and protein expression decreased in HCC patients’ samples and cell culture compared to paratumor controls [32]. By transfecting RUNX1-expressing vectors into liver cancer cells, Liu and colleagues found that RUNX1 negatively influenced tumour cell potential of metastasis and proliferation. They assumed that RUNX1 influenced epithelial-mesenchymal transition (EMT) and its factors. In RUNX1 overexpressed cells, they noticed Vimentin and MMP2 expression decreased, and E cadherin increased, indicating an inhibitory role of RUNX1 in the EMT process [32]. On the other hand, RUNX1 influence the collagen type IV alpha 1 chain (COL4A1) upregulation. The COL4A1 stimulated the growth and expansion of HCC cells, involving the Fak-Src signalling pathway [33].
One of the crucial processes in cancer progression is angiogenesis. In hematopoietic cell differentiation from hemogenic endothelium cells, RUNX1 has a vital role. The RUNX1-deficient mice lack hematopoiesis and angiogenesis [34]. Added exogenously, IGFBP-3 inhibited RUNX1-promoted angiogenesis dose-dependently [35]. Thus, we can conclude that RUNX1 has a cardinal role in angiogenic differentiation and vascularisation. Vascular endothelial growth factor (VEGF) is an angiogenesis modulator in a cancer cell environment and a negative prognostic factor for acute myeloid leukaemia. In the HCC cell culture, Elst and colleagues found that RUNX1 inhibited VEGF expression [36]. Liu and colleagues confirmed these results on HCC patients’ samples and cell lines. They observed Vegf expression decreased and RUNX1 expression increased in HCC patients’ samples. The HCC cell lines with increased RUNX1 expression exhibited VEGF expression decline, too [32]. They concluded that both molecules, VEGF and RUNX1, could be the candidates for the molecularly targeted HCC treatment. Figure 2 shows the impact of RUNX1 on proliferation, metastasis and angiogenesis in HCC.
2.2. RUNX1 and miRNAs
There are not many studies of epigenetic processes involving the RUNX1 gene. Tuo and colleagues noticed the hypomethylation of the RUNX1 promoter in hepatocellular carcinoma [37]. Some studies show the association between several miRNAs and RUNX1 in HCC (Table 1). Transcript 1 of RUNX1 (RUNX1-IT1) is a long noncoding sequence of an RNA transcript of the RUNX1 gene [38]. Yan and colleagues noticed the RUNX1 expression decrease in the HCC patients’ samples, and the knocking-down of RUNX1-IT1 increased the proliferation and reduced apoptosis in HCC cells [39]. Sun and colleagues observed the association of decreased RUNX1-IT1 expression with shorter DFS and OS. RUNX1-IT1 binds mir-632, competing with the other RNAs in HCC cells for target gene GSK-3β binding and modulating the WNT/β-catenin signalling cascade. Added hypoxia-prompted histone deacetylase 3 (HDAC3) in HCC cells reduced the RUNX1-IT1 expression. They concluded that the goal of HCC therapy should be to activate RUNX1-IT1 [40]. On the other hand, Vivacqua and colleagues noticed that oestrogen receptor agonists, such as the G protein-coupled oestrogen receptor agonist (G-1) and 17β-oestradiol (E2), increased miR-144 expression in HepG2 hepatocarcinoma cells, via the G protein-coupled oestrogen receptor 1 (GPER) and the PI3K/ERK1/2/Elk1 pathway. miR-144 then downregulates RUNX1 promoting the cell cycle [41], (Table 1).
Liand colleagues found that the molecule Pam3CSK4, an agonist of Toll-like receptor 2 (TLR2), injected into mice inhibited tumour growth and reduced myeloid-derived suppressor cells (MDSCs), thereby attenuating HCC progression [42]. MDSCs participate in the formation of an immune microenvironment of the tumour. Pam3CSK4 targets RUNX1 and promotes the MDSCs polarisation. On the contrary, inhibiting RUNX1 resulted in tumour enlargement and shortened overall survival. Their results indicated the role of RUNX1 and TLR2 in the MDSCs' formation, function, and polarity. Considering all, RUNX1 and TLR2 targeting could lead to a potential mechanism of HCC immunotherapy [42].
In conclusion, RUNX1 binds target genes (VEGF, COL4A1) and involves signalling pathways of cancer proliferation, metastasis, and angiogenesis. RUNX1 also interacts with several miRNAs, for example, mir-632 and mir-144. Given the importance of RUNX1 in hepatocellular carcinoma, its potential suitability as a treatment target requires additional studies. Researchers should pay particular attention to the binding of RUNX1 to the other genes and miRNAs and its involvement in signalling pathways.
3. RUNX2 IN HCC
The second member of Runt-related family genes, RUNX2, is located on the 6p21 chromosomal region, with the function of a transcriptional regulator in human osteoblast differentiation and chondrocyte maturation [43,44]. Further experimental data indicate that RUNX2 also has a carcinogenic function in various human malignancies [45]. As a regulatory molecule, RUNX2 has been a part of molecular networks that promote the invasive behaviour of tumours [45].
3.1. General role of RUNX2 in HCC
According to literature data, the expression of RUNX2 on mRNA and/or protein level is elevated in HCC cell lines, as well as in liver tumour tissue [46,47,48], suggesting that this transcription factor has a role in hepatocarcinogenesis. Previous findings confirmed higher RUNX2 expression in HCC patients than expression detected in non-tumour tissues or healthy controls [46,47,49]. Wang and colleagues noticed that increased expression of RUNX2 significantly correlates with unfavourable clinicopathological features in HCC. These adverse features included the onset of multiple tumour nodes, higher histological grades and TNM stages, and venous invasion presence [47]. Moreover, aberrant RUNX2 expression acts as an independent prognostic factor since hepatocellular carcinoma patients with high RUNX2 expression demonstrated shorter 5-year disease-free and overall survival [47,48]. Additionally, RUNX2 contributed to the HCC development regardless of the presence of HBV or HCV infections. Besides, the measured level of RUNX2 expression was not significantly impaired by the HCV or HBV existence [46].
3.2. RUNX2 tumour invasion activity in HCC
The general mechanisms underlying the role of RUNX2 in various tumour types provide directions for detailed studies on the impact of RUNX2 on the pathogenesis of HCC. Many reports showed that increased RUNX2 expression enhances the migration of tumour cells migration and invasive properties [50,51,52,53,54,55,56]. Previous studies revealed the crucial role of the RUNX2 in the regulation of the epithelial-to-mesenchymal transition (EMT) process in many tumours [48,57], which is the first step toward tumour invasion and metastatic potential (Figure 3).
Cao and colleagues found that RUNX2 overexpression can promote EMT in HCC [48]. Elevated RUNX2 expression can also trigger vasculogenic mimicry (VM), providing a direct metastatic route to distant sites [58,59]. Experiments on HCC cell lines revealed that RUNX2 is associated with EMT and VM processes by regulating the expression of adhesion molecules such as VE-cadherin and galectin-3, which indirectly contribute to tumour cell migration and enhanced metastatic potential [48,60,61]. Moreover, RUNX2 is implicated in tissue microenvironment regulation and extracellular matrix reshaping. A previous report demonstrated that RUNX2 acted as an initiator of migration and invasion of the HCC cells in vitro by enhancing the expression of the matrix metalloproteinase 9 (MMP9) [47].
The level of RUNX2 expression significantly correlates with the expression level of MMP9 in hepatocellular carcinoma [47]. This association between RUNX2 and MMP9 expression levels was also detected in breast cancer [53]. Moreover, the results of two experimental studies have clarified the RUNX2 indirect oncogenic role in hepatocarcinogenesis. Using a gain/loss-of-function study approach, Yang and colleagues demonstrated that zinc finger protein 521 (ZNF521) strongly repressed the transcriptional activity of RUNX2 and affected RUNX2-related PI3K/AKT signalling pathways, significantly inhibiting HCC growth [62]. Moreover, ZNF521-mediated downregulation of RUNX2 also suppresses tumorigenic processes in HCC cells [62]. Moreover, the downregulated RUNX2 gene notably decreased the HCC cells’ propagation, migration, and chemoresistance [49]. This study specifically examined the role of RUNX2 in the NUPR1/RELB/IER3 signalling cascade as a suggested molecular mechanism underlying HCC development and response to sorafenib treatment [49].
3.3. RUNX2 and non-coding RNAs in HCC
Previous studies showed that multiple microRNAs could be differentially expressed in HCC, directly or indirectly affecting RUNX2 expression and activity (Table 1). RUNX2 might be directly repressed by the miR-455 molecule in human HCC samples [63], which has already demonstrated tumour-suppressive properties [64,65]. Further gain/loss-of-function analyses showed that in HCC cells, miR-455 regulates the process of RUNX2 accumulation in vitro, which significantly suppresses cell migration abilities [63]. Additionally, several miRNAs can regulate RUNX2 expression by direct binding to the RUNX2 gene 30-UTR region [66,67]. Wang and colleagues suggested that miR-196a could have a significant role in HCC development through the RUNX2 upregulation, which in HCC cell lines produced higher osteopontin levels as a consequence [68]. Osteopontin is a well-known bone marrow-produced protein that regulates bone regeneration, although previous reports indicate that this protein also contributes to cancer metastasis [69]. On the other hand, RUNX2 may entangle in the HCC development by directing the expression level regulation of several miRNAs. Wang and colleagues investigated the mechanism underlying the increased level of O-GlcNAc transferase, which enhances tumour cell migratory abilities and HCC invasive capacities [70]. Their results showed that RUNX2 indirectly affects OGT expression via transcriptional activation of miR-24 by binding to its promoter [70]. In another study on mouse hepatoma cells, RUNX2 binds to the miR-23a gene’s promoter and indirectly promotes lymphatic metastasis by targeting the Mgat3 glycosyltransferase affecting directly the glycosylation process on the cell surface [71].
Increasing evidence suggests that RUNX2 can interact with long non-coding RNAs (lncRNAs), which contribute to hepatocellular carcinogenesis (Table1.). For example, the lncRNA called HAND2-AS plays a tumour-suppressive role in liver cancer and prohibits hepatoma cancer cell proliferation by decreasing the expression level of RUNX2 [46]. In another study, RUNX2 and transcriptional regulator YAP inhibit the expression level of lncRNA annotated MT1DP, demonstrating tumour-suppressive behaviour in hepatocellular carcinoma [72]. However, detailed analyses are necessary to clarify the correlation between RUNX2 and different lncRNAs, and their synergistic effect on liver carcinogenesis.
RUNX2, a unique transcription factor, exhibits a crucial oncogenic role in hepatocellular carcinoma. Moreover, we should consider RUNX2 aberrant expression as a novel prognostic indicator in HCC. Studies on RUNX2-related regulatory mechanisms hint at its pro-invasive functions in HCC by reshaping the tumour microenvironment, making RUNX2 a potential therapeutic target for blocking metastasis and further disease progression. Since the RUNX2 transcription regulator is implicated in many signalling pathways and interacts with multiple regulatory molecules like microRNAs and lncRNAs, more in-depth studies for a better understanding of its role in the molecular pathology of hepatocellular carcinoma are needed.
4. RUNX3 IN HCC
The third member of the Runt-related gene family, RUNX3, is located in the chromosomal region 1p36–35 [73]. RUNX3 has initially been reported as a tumour suppressor in gastric cancer [74]. Subsequent studies confirmed its tumour-suppressive role in some of the most common cancer types in humans, including colorectal [75], prostate [76], breast [77], lung cancer [78], and melanoma [79]. On the other hand, RUNX3 has been shown to act as an oncogene and promote tumour development in ovarian [80], head and neck [81], and pancreatic carcinoma [82]. This dualistic function of RUNX3 is cell-context dependent [83].
RUNX3 is required for normal liver development, while its loss is associated with hepatocellular carcinogenesis, where it acts as a tumour suppressor [18,84]. RUNX3 gene expression is decreased in up to 80% of HCCs, predominantly due to promoter methylation. The loss of heterozygosity (LOH) was also observed in several cases [85,86]. In two meta-studies, RUNX3 hypermethylation has been shown to occur early in hepatocarcinogenesis, including premalignant conditions, like liver fibrosis and cirrhosis, with the highest frequencies being reported in HCC [87,88].
4.1. General role of RUNX3 in HCC
Several studies have shown that RUNX3 inactivation is cardinal for the initiation and progression of HCC [87,88,89,90,91,92]. As a multifunctional transcription factor, RUNX3 is implicated in diverse signalling pathways and cellular processes, thereby exerting multiple effects on tumour suppression [93,94]. According to current knowledge, RUNX3 participates in the regulation of the cell cycle [95], proliferation and apoptosis [96], angiogenesis [18], and EMT [90,97]. Its loss is also related to chemoresistance [98,99] (Figure 4).
4.2. RUNX3 regulates cell cycle, proliferation, and apoptosis
Dysregulation of the cell cycle is a prime event in hepatocarcinogenesis. RUNX3 may play a pivotal role in this process by employing diverse mechanisms [96]. Earlier studies on gastric epithelial cells demonstrated that RUNX3 regulates the cell cycle by interacting with p21, p27, and cyclin D1 proteins [100,101,102]. Further research revealed that RUNX3 induce the expression of the ARF and CDKN1A cell cycle regulators by interaction with BRD2 and pRB proteins [78,95]. Moreover, it has recently been shown that RUNX3 activates the cell cycle restriction (R)-point-associated genes by recruitment of chromatin-remodelling complex, histone modifiers and cell-cycle regulators to form RUNX3-containing activator complex, which opens chromatin structure in the vicinity of target genes [103]. Whether RUNX3 exerts such a function in HCC remains to be elucidated.
As the major component of the transforming growth factor-beta signalling (TGF-β) pathway [8,104], RUNX3 can stop cell proliferation, inducing p21 cell-cycle inhibitor [102]. Similarly, it can suppress apoptosis by inducing apoptosis initiator Bim, as it has been shown on gastric cancer cell lines [105].
Another study on human HCC cell lines demonstrated that RUNX3 could induce apoptosis through the Bim-caspase pathway, even in the TGF-β absence [84]. RUNX3 also regulates the TGF-β-mediated growth arrest through the induction of CDK inhibitors and/or the repression of the c-Myc proto-oncogene [106,107].
Another study in mice showed that proliferation marker Ki67 was more frequently observed in the RUNX3 knock-out liver cells than in wild-type cells, which further confirmed the role of RUNX3 in hepatocyte proliferation regulation [18]. RUNX3 has been reported to control cellular senescence, a potent anti-cancer mechanism that prevents the proliferation of potentially cancerous cells [108]. A recent study on human HCC samples and cell lines demonstrated that RUNX3 could modulate the expression of key markers of cellular senescence, p53 and p21, via the circLARP4/miR-761/RUNX3 signalling axis [109], (Table 1). As a competing endogenous RNA, the circLARP4 harbours miR-761, abrogating its inhibitory effect on the RUNX3 gene. RUNX3 subsequently activates the p53/p21 signalling pathway and enhances the downstream senescence phenotype in HCC [109].
In addition, evidence suggests that RUNX3 can regulate cell cycle and apoptosis through the Wnt/β-catenin signalling pathway, whose oncogenic activation is a usual event in HCC [110,111,112]. RUNX3 directly interacts with the Wnt transcription factor, TCF4-β-catenin complex, and thus inhibits the expression of Wnt target genes, c-Myc and cyclin D, regulators of apoptosis and cell cycle, respectively [93,113,114].
Oncogenic activation of the Notch signalling pathway is also implicated in hepatocyte growth and proliferation [115,116]. Gao and colleagues have shown that RUNX3 can suppress oncogenic Notch signalling through direct interaction with the intracellular domain of the Notch1 protein in HCC cell lines [94]. Further studies revealed that RUNX3 decreases jagged-1 (JAG1) mRNA and thus inhibits JAG1-mediated Notch signalling in HCC [115,117]. Moreover, RUNX3 has been reported to inhibit the transcription of HES1, the Notch target gene, implicated in the stemness, metastasis, and chemoresistance regulation in cancer [94]. Given that, affecting Notch1 signalling by RUNX3 reactivation might be a promising therapeutic approach for the HCC treatment.
4.3. RUNX3 in the angiogenesis regulation
A crucial tumour-suppressive role of RUNX3 is angiogenesis prevention and tumour invasion. A recent study revealed that after the HCC therapeutic drug’s application, sorafenib, RUNX3 suppressed VEGF expression in HCC, which was associated with reduced tumour growth [118]. A previous study on gastric cancer cells demonstrated that RUNX3 destabilised hypoxia-inducible factor HIF-1α in the hypoxic microenvironment, thus inhibiting angiogenesis [119]. Additional research is necessary to confirm whether this mechanism exists in HCC, as inhibition of angiogenesis is an important therapeutic strategy for the prevention of HCC progression [120].
4.4. RUNX3 and epithelial-mesenchymal transition
Previous studies have shown that the loss of RUNX3 contributes to EMT, a crucial process related to metastasis, chemoresistance, and tumour stemness [121,122]. In vitro experiments demonstrated that RUNX3 repressed tumour metastasis and invasion by upregulating E-cadherin through the miR-186/E-cadherin/EMT axis [90,123] (Table 1.). In addition, experiments on human HCC cell lines revealed that the loss of RUNX3 supports pro-oncogenic TGF-β signalling through the upregulation of EMT genes and that RUNX3 can also suppress EMT via the inhibition of Wnt signaling [97]. Given its crucial role in carcinogenesis, targeting EMT by re-expressing RUNX3 could be another potential therapeutic approach for treating HCC patients.
4.5. RUNX3 and chemoresistance
A study on human HCC samples and cell lines demonstrated that RUNX3 could be downregulated by overexpression of miR-130 through the miR-130a/RUNX3/Wnt signalling pathway. This mechanism was associated with increased chemoresistance to cisplatin [99]. Studies in gastric [124] and cervical cancer [125] demonstrated that miR-130 directly binds to the RUNX3 and thus inhibits its expression. Accordingly, restoration of RUNX3 expression by targeting miR-130 could be a potential approach to overcome chemotherapy resistance in HCC patients.
Researchers also demonstrated that the loss of RUNX3 contributes to 5-fluorouracil (5-FU) and cisplatin (CDDP) resistance in HCC cell lines and patients through increased expression of multidrug resistance-associated proteins (MRP) [98]. Drug resistance is an extensive obstacle to the successful treatment of HCC. Therefore, additional research is necessary to address this issue and develop more efficient treatment approaches.
All concerning, RUNX3 appears to be involved in hepatocarcinogenesis at distinct stages, from initiation to progression and metastasis. Thus, its potential clinical application might have a wide range. However, given that many results are on cell lines, further studies on HCC patients are needed for a complete understanding of the significance of RUNX3 in HCC.
5. Conclusion and Future Directions
Despite considerable advances in cancer diagnosis and treatment, HCC remains one of the most common and hard-to-treat human cancers. Revealing the essential molecular processes underlying hepatocarcinogenesis is crucial for establishing reliable diagnostic, prognostic, and therapeutic markers. RUNX genes are often deregulated in HCC, exerting complex and conflicting functions. The role of RUNX1 is still contradictory, as there are reports of its tumour-suppressive but also an oncogenic role in HCC. According to the current knowledge, RUNX2 acts as an oncogene and is related to the more aggressive forms of the disease, whereas RUNX3 exerts tumour suppressive role and could be used as a biomarker for early HCC detection. All three genes could serve as therapeutic targets. However, a deeper understanding of the relationship between different RUNX family members and the signalling pathways they are involved in, considering the cell-specific microenvironment, is necessary for effective HCC therapeutic strategy development.
Author Contributions
Conceptualization, M.K. and S.J.C.; writing - original draft: M.K., B.K., A.B. and S.J.C.; writing - review and editing: M.K., B.K. and A.B.; figure preparation: B.K., and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development, and Innovations, of the Republic of Serbia (Reg. No: 451-03-9/2021-14/200017), under Research Theme "Molecular alterations as prognostic and predictive markers in human malignant tumours" – No.0802303.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| HCC | Hepatocellular Carcinoma; |
| RUNX | Runt-related transcription factors;; |
| HBV | Hepatitis B Virus; |
| HCV | Hepatitis C Virus; |
| RHD | Runt homology domain; |
| miRNA | Micro ribonucleic acids; |
| lncRNAs | Long non-coding ribonucleic acids. |
References
- Asafo-Agyei, K.O.; Samant, H. Hepatocellular Carcinoma. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2023. [Google Scholar]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular Carcinoma. Nat Rev Dis Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Balogh, J.; Victor, D.; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, M.; Monsour, H. Hepatocellular Carcinoma: A Review. JHC 2016, Volume 3, 41–53. [Google Scholar] [CrossRef]
- Sim, H.-W.; Knox, J. Hepatocellular Carcinoma in the Era of Immunotherapy. Current Problems in Cancer 2018, 42, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Suresh, D.; Srinivas, A.N.; Prashant, A.; Harikumar, K.B.; Kumar, D.P. Therapeutic Options in Hepatocellular Carcinoma: A Comprehensive Review. Clin Exp Med 2023. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.S.H.; Ito, K.; Ito, Y. RUNX Family: Regulation and Diversification of Roles through Interacting Proteins. Int. J. Cancer 2013, 132, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Otalora-Otalora, B.; Henriquez, B.; Lopez-Kleine, L.; Rojas, A. RUNX Family: Oncogenes or Tumor Suppressors (Review). Oncol Rep 2019. [Google Scholar] [CrossRef]
- Ito, Y.; Bae, S.-C.; Chuang, L.S.H. The RUNX Family: Developmental Regulators in Cancer. Nat Rev Cancer 2015, 15, 81–95. [Google Scholar] [CrossRef]
- Levanon, D.; Groner, Y. Structure and Regulated Expression of Mammalian RUNX Genes. Oncogene 2004, 23, 4211–4219. [Google Scholar] [CrossRef]
- Yi, H.; He, Y.; Zhu, Q.; Fang, L. RUNX Proteins as Epigenetic Modulators in Cancer. Cells 2022, 11, 3687. [Google Scholar] [CrossRef]
- Mevel, R.; Draper, J.E.; Lie-a-Ling, M.; Kouskoff, V.; Lacaud, G. RUNX Transcription Factors: Orchestrators of Development. Development 2019, 146, dev148296. [Google Scholar] [CrossRef]
- Darnell, J.E. Transcription Factors as Targets for Cancer Therapy. Nat Rev Cancer 2002, 2, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Kamikubo, Y.; Liu, P. Role of RUNX1 in Hematological Malignancies. Blood 2017, 129, 2070–2082. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y. Oncogenic Potential of the RUNX Gene Family: ‘Overview. ’ Oncogene 2004, 23, 4198–4208. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M. RUNX Family in Hypoxic Microenvironment and Angiogenesis in Cancers. Cells 2022, 11, 3098. [Google Scholar] [CrossRef]
- Yamagata, T.; Maki, K.; Mitani, K. Runx1/AML1 in Normal and Abnormal Hematopoiesis. International Journal of Hematology 2005, 82, 1–8. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Innamorati, G.; Valenti, M.T. Transcription Factor Runx2 and Its Application to Bone Tissue Engineering. Stem Cell Rev and Rep 2012, 8, 891–897. [Google Scholar] [CrossRef]
- Lee, J.-M.; Lee, D.-J.; Bae, S.-C.; Jung, H.-S. Abnormal Liver Differentiation and Excessive Angiogenesis in Mice Lacking Runx3. Histochem Cell Biol 2013, 139, 751–758. [Google Scholar] [CrossRef]
- Wang, J.W.; Stifani, S. Roles of Runx Genes in Nervous System Development. In RUNX Proteins in Development and Cancer; Groner, Y., Ito, Y., Liu, P., Neil, J.C., Speck, N.A., Van Wijnen, A., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2017; ISBN 978-981-10-3231-8. [Google Scholar]
- Guilliams, M.; Bonnardel, J.; Haest, B.; Vanderborght, B.; Wagner, C.; Remmerie, A.; Bujko, A.; Martens, L.; Thoné, T.; Browaeys, R.; et al. Spatial Proteogenomics Reveals Distinct and Evolutionarily Conserved Hepatic Macrophage Niches. Cell 2022, 185, 379–396. [Google Scholar] [CrossRef]
- Human All Liver Cells. Available online: https://www.livercellatlas.org/umap-humanAll.php (accessed on 3.11.22).
- Pan, S.; Sun, S.; Liu, B.; Hou, Y. Pan-Cancer Landscape of the RUNX Protein Family Reveals Their Potential as Carcinogenic Biomarkers and the Mechanisms Underlying Their Action. Journal of Translational Internal Medicine 2022, 10, 156–174. [Google Scholar] [CrossRef]
- National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/gene/861 (accessed on 3.18.23).
- Miyoshi, H.; Shimizu, K.; Kozu, T.; Maseki, N.; Kaneko, Y.; Ohki, M. T(8;21) Breakpoints on Chromosome 21 in Acute Myeloid Leukemia Are Clustered within a Limited Region of a Single Gene, AML1. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 10431–10434. [Google Scholar] [CrossRef]
- Martinez, M.; Hinojosa, M.; Trombly, D.; Morin, V.; Stein, J.; Stein, G.; Javed, A.; Gutierrez, S.E. Transcriptional Auto-Regulation of RUNX1 P1 Promoter. PLoS ONE 2016, 11, e0149119. [Google Scholar] [CrossRef]
- Lin, T.-C. RUNX1 and Cancer. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2022, 1877, 188715. [Google Scholar] [CrossRef] [PubMed]
- TIMER2.0. Available online: http://timer.cistrome.org/ (accessed on 4.3.23).
- GENT2. Available online: http://gent2.appex.kr/gent2/ (accessed on 4.3.23).
- Zhu, J.; Sanborn, J.Z.; Benz, S.; Szeto, C.; Hsu, F.; Kuhn, R.M.; Karolchik, D.; Archie, J.; Lenburg, M.E.; Esserman, L.J.; et al. The UCSC Cancer Genomics Browser. Nat Methods 2009, 6, 239–240. [Google Scholar] [CrossRef] [PubMed]
- UCSC Xena. Available online: http://xena.ucsc.edu/welcome-to-ucsc-xena/ (accessed on 4.3.23).
- Miyagawa, K.; Sakakura, C.; Nakashima, S.; Yoshikawa, T.; Kin, S.; Nakase, Y.; Ito, K.; Yamagishi, H.; Ida, H.; Yazumi, S.; et al. Down-Regulation of RUNX1, RUNX3 and CBFbeta in Hepatocellular Carcinomas in an Early Stage of Hepatocarcinogenesis. Anticancer Res 2006, 26, 3633–3643. [Google Scholar] [PubMed]
- Liu, C.; Xu, D.; Xue, B.; Liu, B.; Li, J.; Huang, J. Upregulation of RUNX1 Suppresses Proliferation and Migration through Repressing VEGFA Expression in Hepatocellular Carcinoma. Pathol. Oncol. Res. 2020, 26, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jin, H.; Hu, J.; Li, X.; Ruan, H.; Xu, H.; Wei, L.; Dong, W.; Teng, F.; Gu, J.; et al. COL4A1 Promotes the Growth and Metastasis of Hepatocellular Carcinoma Cells by Activating FAK-Src Signaling. J Exp Clin Cancer Res 2020, 39, 148. [Google Scholar] [CrossRef] [PubMed]
- Takakura, N.; Watanabe, T.; Suenobu, S.; Yamada, Y.; Noda, T.; Ito, Y.; Satake, M.; Suda, T. A Role for Hematopoietic Stem Cells in Promoting Angiogenesis. Cell 2000, 102, 199–209. [Google Scholar] [CrossRef]
- Iwatsuki, K.; Tanaka, K.; Kaneko, T.; Kazama, R.; Okamoto, S.; Nakayama, Y.; Ito, Y.; Satake, M.; Takahashi, S.-I.; Miyajima, A.; et al. Runx1 Promotes Angiogenesis by Downregulation of Insulin-like Growth Factor-Binding Protein-3. Oncogene 2005, 24, 1129–1137. [Google Scholar] [CrossRef]
- Ter Elst, A.; Ma, B.; Scherpen, F.J.G.; De Jonge, H.J.M.; Douwes, J.; Wierenga, A.T.J.; Schuringa, J.J.; Kamps, W.A.; De Bont, E.S.J.M. Repression of Vascular Endothelial Growth Factor Expression by the Runt-Related Transcription Factor 1 in Acute Myeloid Leukemia. Cancer Research 2011, 71, 2761–2771. [Google Scholar] [CrossRef]
- Tuo, Z.; Zhang, Y.; Wang, X.; Dai, S.; Liu, K.; Xia, D.; Wang, J.; Bi, L. RUNX1 Is a Promising Prognostic Biomarker and Related to Immune Infiltrates of Cancer-Associated Fibroblasts in Human Cancers. BMC Cancer 2022, 22, 523. [Google Scholar] [CrossRef]
- National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=80215 (accessed on 3.18.23).
- Yan, P.-H.; Wang, L.; Chen, H.; Yu, F.-Q.; Guo, L.; Liu, Y.; Zhang, W.-J.; Bai, Y.-L. LncRNA RUNX1-IT1 Inhibits Proliferation and Promotes Apoptosis of Hepatocellular Carcinoma by Regulating MAPK Pathways. European Review for Medical and Pharmacological Sciences 2019, 23, 8287–8294. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, L.; Chen, T.; Shi, Y.; Yao, B.; Liu, Z.; Wang, Y.; Li, Q.; Liu, R.; Niu, Y.; et al. LncRNA RUNX1-IT1 Which Is Downregulated by Hypoxia-Driven Histone Deacetylase 3 Represses Proliferation and Cancer Stem-like Properties in Hepatocellular Carcinoma Cells. Cell Death Dis 2020, 11, 95. [Google Scholar] [CrossRef]
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic Gper Signaling Regulates Mir144 Expression in Cancer Cells and Cancer-Associated Fibroblasts (Cafs). Oncotarget 2015, 6, 16573–16587. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, F.; Xu, L.; Liu, X.; Zhu, X.; Gao, W.; Shen, X. TLR2 Agonist Promotes Myeloid-Derived Suppressor Cell Polarization via Runx1 in Hepatocellular Carcinoma. International Immunopharmacology 2022, 111, 109168. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.P.; Später, D.; Taketo, M.M.; Birchmeier, W.; Hartmann, C. Canonical Wnt/β-Catenin Signaling Prevents Osteoblasts from Differentiating into Chondrocytes. Developmental Cell 2005, 8, 727–738. [Google Scholar] [CrossRef]
- Zhang, Y.-W.; Yasui, N.; Ito, K.; Huang, G.; Fujii, M.; Hanai, J.; Nogami, H.; Ochi, T.; Miyazono, K.; Ito, Y. A RUNX2/PEBP2 AA/ CBFA1 Mutation Displaying Impaired Transactivation and Smad Interaction in Cleidocranial Dysplasia. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 10549–10554. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, H.; Chai, J.; Xing, L. RUNX2 as a Promising Therapeutic Target for Malignant Tumors. CMAR 2021, Volume 13, 2539–2548. [Google Scholar] [CrossRef]
- Jing, G.; Zheng, X.; Ji, X. LncRNA HAND2-AS1 Overexpression Inhibits Cancer Cell Proliferation in Hepatocellular Carcinoma by Downregulating RUNX2 Expression. J Clin Lab Anal 2021, 35. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, W.; Huang, T.; Zhu, Y.; Huang, C. RUNX2 Promotes Hepatocellular Carcinoma Cell Migration and Invasion by Upregulating MMP9 Expression. Oncology Reports 2016, 36, 2777–2784. [Google Scholar] [CrossRef]
- Cao, Z.; Sun, B.; Zhao, X.; Zhang, Y.; Gu, Q.; Liang, X.; Dong, X.; Zhao, N. Correction: Cao, Z., et al. The Expression and Functional Significance of Runx2 in Hepatocellular Carcinoma: Its Role in Vasculogenic Mimicry and Epithelial—Mesenchymal Transition. Int. J. Mol. Sci. 2017, 18, 500. IJMS 2020, 22, 77. [Google Scholar] [CrossRef]
- Emma, M.R.; Iovanna, J.L.; Bachvarov, D.; Puleio, R.; Loria, G.R.; Augello, G.; Candido, S.; Libra, M.; Gulino, A.; Cancila, V.; et al. NUPR1, a New Target in Liver Cancer: Implication in Controlling Cell Growth, Migration, Invasion and Sorafenib Resistance. Cell Death Dis 2016, 7, e2269–e2269. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L.; Wu, Y.; Zhang, R.; Zhang, M.; Liao, D.; Wang, G.; Qin, G.; Xu, R.; Kang, T. CBX4 Suppresses Metastasis via Recruitment of HDAC3 to the Runx2 Promoter in Colorectal Carcinoma. Cancer Research 2016, 76, 7277–7289. [Google Scholar] [CrossRef] [PubMed]
- Sase, T.; Suzuki, T.; Miura, K.; Shiiba, K.; Sato, I.; Nakamura, Y.; Takagi, K.; Onodera, Y.; Miki, Y.; Watanabe, M.; et al. Runt-Related Transcription Factor 2 in Human Colon Carcinoma: A Potent Prognostic Factor Associated with Estrogen Receptor. Int. J. Cancer 2012, 131, 2284–2293. [Google Scholar] [CrossRef]
- Komori, T. Runx2, A Multifunctional Transcription Factor in Skeletal Development. J. Cell. Biochem. 2002, 87, 1–8. [Google Scholar] [CrossRef]
- Pratap, J.; Javed, A.; Languino, L.R.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. The Runx2 Osteogenic Transcription Factor Regulates Matrix Metalloproteinase 9 in Bone Metastatic Cancer Cells and Controls Cell Invasion. Molecular and Cellular Biology 2005, 25, 8581–8591. [Google Scholar] [CrossRef] [PubMed]
- Boregowda, R.K.; Olabisi, O.O.; Abushahba, W.; Jeong, B.-S.; Haenssen, K.K.; Chen, W.; Chekmareva, M.; Lasfar, A.; Foran, D.J.; Goydos, J.S.; et al. RUNX2 Is Overexpressed in Melanoma Cells and Mediates Their Migration and Invasion. Cancer Letters 2014, 348, 61–70. [Google Scholar] [CrossRef]
- Li, X.-Q.; Du, X.; Li, D.-M.; Kong, P.-Z.; Sun, Y.; Liu, P.-F.; Wang, Q.-S.; Feng, Y.-M. ITGBL1 Is a Runx2 Transcriptional Target and Promotes Breast Cancer Bone Metastasis by Activating the TGFβ Signaling Pathway. Cancer Research 2015, 75, 3302–3313. [Google Scholar] [CrossRef]
- El-Gendi, S.M.; Mostafa, M.F. Runx2 Expression as a Potential Prognostic Marker in Invasive Ductal Breast Carcinoma. Pathol. Oncol. Res. 2016, 22, 461–470. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Khalid, O.; Gabet, Y.; Shah, R.R.; Purcell, D.J.; Mav, D.; Kohn-Gabet, A.E.; Shi, Y.; Coetzee, G.A.; Frenkel, B. Runx2 Transcriptome of Prostate Cancer Cells: Insights into Invasiveness and Bone Metastasis. Mol Cancer 2010, 9, 258. [Google Scholar] [CrossRef]
- Meng, J.; Sun, B.; Zhao, X.; Zhang, D.; Zhao, X.; Gu, Q.; Dong, X.; Zhao, N.; Liu, P.; Liu, Y. Doxycycline as an Inhibitor of the Epithelial-to-Mesenchymal Transition and Vasculogenic Mimicry in Hepatocellular Carcinoma. Molecular Cancer Therapeutics 2014, 13, 3107–3122. [Google Scholar] [CrossRef]
- Sun, T.; Zhao, N.; Zhao, X.; Gu, Q.; Zhang, S.; Che, N.; Wang, X.; Du, J.; Liu, Y.; Sun, B. Expression and Functional Significance of Twist1 in Hepatocellular Carcinoma: Its Role in Vasculogenic Mimicry. Hepatology 2010, 51, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Funasaka, T.; Raz, A.; Nangia-Makker, P. Galectin-3 in Angiogenesis and Metastasis. Glycobiology 2014, 24, 886–891. [Google Scholar] [CrossRef]
- Vestweber, D. VE-Cadherin: The Major Endothelial Adhesion Molecule Controlling Cellular Junctions and Blood Vessel Formation. ATVB 2008, 28, 223–232. [Google Scholar] [CrossRef]
- Yang, N.; Wang, L.; Chen, T.; Liu, R.; Liu, Z.; Zhang, L. ZNF521 Which Is Downregulated by MiR-802 Suppresses Malignant Progression of Hepatocellular Carcinoma through Regulating Runx2 Expression. J. Cancer 2020, 11, 5831–5839. [Google Scholar] [CrossRef]
- Qin, L.; Zhang, Y.; Lin, J.; Shentu, Y.; Xie, X. MicroRNA-455 Regulates Migration and Invasion of Human Hepatocellular Carcinoma by Targeting Runx2. Oncology Reports 2016, 36, 3325–3332. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; Ping, C.; Tang, J.; Zhang, W. MicroRNA-455 Suppresses Non-Small Cell Lung Cancer through Targeting ZEB1: The Role of MiRNA-455 in NSCLC. Cell Biol Int 2016, 40, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Wang, S.; Han, D.; Dong, W.; Xie, C.; Guo, H. MicroRNA-455 Inhibits Proliferation and Invasion of Colorectal Cancer by Targeting RAF Proto-Oncogene Serine/Threonine-Protein Kinase. Tumor Biol. 2015, 36, 1313–1321. [Google Scholar] [CrossRef]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H.; et al. MiRTarBase Update 2018: A Resource for Experimentally Validated MicroRNA-Target Interactions. Nucleic Acids Research 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, S.; Wang, B.; Huang, J.; Lu, W.W.; Chen, D. Runx2 and MicroRNA Regulation in Bone and Cartilage Diseases: Runx2 and MiRNAs in Bone and Cartilage. Ann. N.Y. Acad. Sci. 2016, 1383, 80–87. [Google Scholar] [CrossRef]
- Wang, S.-Y.; Chen, C.-L.; Hu, Y.-C.; Chi, Y.; Huang, Y.-H.; Su, C.-W.; Jeng, W.-J.; Liang, Y.-J.; Wu, J.-C. High Expression of MicroRNA-196a Is Associated with Progression of Hepatocellular Carcinoma in Younger Patients. Cancers 2019, 11, 1549. [Google Scholar] [CrossRef]
- Wai, P.Y.; Kuo, P.C. The Role of Osteopontin in Tumor Metastasis. Journal of Surgical Research 2004, 121, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Y.; Zhang, C.; Chen, X.; Huang, H.; Li, W.; Zhang, J.; Liu, Y. Upregulation of OGT by Caveolin-1 Promotes Hepatocellular Carcinoma Cell Migration and Invasion. Cell Biology International 2021, 45, 2251–2263. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, Y.; Yu, P.; Qu, J.; Guo, Y.; Li, W.; Wang, S.; Zhang, J. MiR-23a Transcriptional Activated by Runx2 Increases Metastatic Potential of Mouse Hepatoma Cell via Directly Targeting Mgat3. Sci Rep 2018, 8, 7366. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Qiao, Y.; Tang, X.; Ma, L.; Wang, Y.; Zhang, X.; Weng, W.; Pan, Q.; Yu, Y.; Sun, F.; et al. Tumor Suppressor Long Non-Coding RNA, MT1DP Is Negatively Regulated by YAP and Runx2 to Inhibit FoxA1 in Liver Cancer Cells. Cellular Signalling 2014, 26, 2961–2968. [Google Scholar] [CrossRef] [PubMed]
- Lund, A.H.; Van Lohuizen, M. RUNX: A Trilogy of Cancer Genes. Cancer Cell 2002, 1, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-L.; Ito, K.; Sakakura, C.; Fukamachi, H.; Inoue, K.; Chi, X.-Z.; Lee, K.-Y.; Nomura, S.; Lee, C.-W.; Han, S.-B.; et al. Causal Relationship between the Loss of RUNX3 Expression and Gastric Cancer. Cell 2002, 109, 113–124. [Google Scholar] [CrossRef]
- Goel, A.; Arnold, C.N.; Tassone, P.; Chang, D.K.; Niedzwiecki, D.; Dowell, J.M.; Wasserman, L.; Compton, C.; Mayer, R.J.; Bertagnolli, M.M.; et al. Epigenetic Inactivation OfRUNX3 in Microsatellite Unstable Sporadic Colon Cancers. Int. J. Cancer 2004, 112, 754–759. [Google Scholar] [CrossRef]
- Kang, G.H.; Lee, S.; Lee, H.J.; Hwang, K.S. Aberrant CpG Island Hypermethylation of Multiple Genes in Prostate Cancer and Prostatic Intraepithelial Neoplasia: CpG Island Methylation in Prostate Cancer and PIN. J. Pathol. 2004, 202, 233–240. [Google Scholar] [CrossRef]
- Lau, Q.C.; Raja, E.; Salto-Tellez, M.; Liu, Q.; Ito, K.; Inoue, M.; Putti, T.C.; Loh, M.; Ko, T.K.; Huang, C.; et al. RUNX3 Is Frequently Inactivated by Dual Mechanisms of Protein Mislocalization and Promoter Hypermethylation in Breast Cancer. Cancer Research 2006, 66, 6512–6520. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, J.-W.; Jang, J.-W.; Chi, X.-Z.; Kim, J.-H.; Li, Y.-H.; Kim, M.-K.; Kim, D.-M.; Choi, B.-S.; Kim, E.-G.; et al. Runx3 Inactivation Is a Crucial Early Event in the Development of Lung Adenocarcinoma. Cancer Cell 2013, 24, 603–616. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, G.; Cheng, Y.; Martinka, M.; Li, G. Prognostic Significance of RUNX3 Expression in Human Melanoma: RUNX3 in Melanoma Prognosis. Cancer 2011, 117, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Nevadunsky, N.S.; Barbieri, J.S.; Kwong, J.; Merritt, M.A.; Welch, W.R.; Berkowitz, R.S.; Mok, S.C. RUNX3 Protein Is Overexpressed in Human Epithelial Ovarian Cancer. Gynecologic Oncology 2009, 112, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, T.; Kudo, Y.; Iizuka, S.; Ogawa, I.; Fujita, T.; Kurihara, H.; Abiko, Y.; Takata, T. RUNX3 Has an Oncogenic Role in Head and Neck Cancer. PLoS ONE 2009, 4, e5892. [Google Scholar] [CrossRef] [PubMed]
- Whittle, M.C.; Izeradjene, K.; Rani, P.G.; Feng, L.; Carlson, M.A.; DelGiorno, K.E.; Wood, L.D.; Goggins, M.; Hruban, R.H.; Chang, A.E.; et al. RUNX3 Controls a Metastatic Switch in Pancreatic Ductal Adenocarcinoma. Cell 2015, 161, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sundaram, S.; Rayala, S.K.; Venkatraman, G. UnPAKing RUNX3 Functions–Both Sides of the Coin. Small GTPases 2017, 1–7. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Shiraha, H.; Nishina, S.; Tanaka, S.; Matsubara, M.; Horiguchi, S.; Iwamuro, M.; Takaoka, N.; Uemura, M.; Kuwaki, K.; et al. Loss of Runt-Related Transcription Factor 3 Expression Leads Hepatocellular Carcinoma Cells to Escape Apoptosis. BMC Cancer 2011, 11, 3. [Google Scholar] [CrossRef]
- Mori, T.; Nomoto, S.; Koshikawa, K.; Fujii, T.; Sakai, M.; Nishikawa, Y.; Inoue, S.; Takeda, S.; Kaneko, T.; Nakao, A. Decreased Expression and Frequent Allelic Inactivation of the RUNX3 Gene at 1p36 in Human Hepatocellular Carcinoma. Liver Int 2005, 25, 380–388. [Google Scholar] [CrossRef]
- Xiao, W.-H.; Liu, W.-W. Hemizygous Deletion and Hypermethylation of RUNX3 Gene in Hepatocellular Carcinoma. WJG 2004, 10, 376. [Google Scholar] [CrossRef]
- Zhang, X.; He, H.; Zhang, X.; Guo, W.; Wang, Y. RUNX3 Promoter Methylation Is Associated with Hepatocellular Carcinoma Risk: A Meta-Analysis. Cancer Investigation 2015, 33, 121–125. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Z.; Zou, Z.; Xiao, G.; Luo, G.; Yang, H. Clinicopathological Significance of RUNX3 Gene Hypermethylation in Hepatocellular Carcinoma. Tumor Biol. 2014, 35, 10333–10340. [Google Scholar] [CrossRef]
- El-shaarawy, F.; Abo ElAzm, M.M.; Mohamed, R.H.; Radwan, M.I.; Abo-Elmatty, D.M.; Mehanna, E.T. Relation of the Methylation State of RUNX3 and P16 Gene Promoters with Hepatocellular Carcinoma in Egyptian Patients. Egypt J Med Hum Genet 2022, 23, 48. [Google Scholar] [CrossRef]
- Gou, Y.; Zhai, F.; Zhang, L.; Cui, L. RUNX3 Regulates Hepatocellular Carcinoma Cell Metastasis via Targeting MiR-186/E-Cadherin/EMT Pathway. Oncotarget 2017, 8, 61475–61486. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Zhang, C.; Feng, M.; Liu, W.; Xie, H.; Qin, Q.; Zhao, E.; Wan, L. Methylation Analysis of P16, SLIT2, SCARA5, and Runx3 Genes in Hepatocellular Carcinoma. Medicine 2017, 96, e8279. [Google Scholar] [CrossRef] [PubMed]
- El-Bendary, M.; Nour, D.; Arafa, M.; Neamatallah, M. Methylation of Tumour Suppressor Genes RUNX3, RASSF1A and E-Cadherin in HCV-Related Liver Cirrhosis and Hepatocellular Carcinoma. British Journal of Biomedical Science 2020, 77, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, X.; Bai, J.; Pei, D.; Zheng, J. The Emerging Role of RUNX3 in Cancer Metastasis (Review). Oncology Reports 2016, 35, 1227–1236. [Google Scholar] [CrossRef]
- Gao, J.; Chen, Y.; Wu, K.-C.; Liu, J.; Zhao, Y.-Q.; Pan, Y.-L.; Du, R.; Zheng, G.-R.; Xiong, Y.-M.; Xu, H.-L.; et al. RUNX3 Directly Interacts with Intracellular Domain of Notch1 and Suppresses Notch Signaling in Hepatocellular Carcinoma Cells. Experimental Cell Research 2010, 316, 149–157. [Google Scholar] [CrossRef]
- Chi, X.-Z.; Lee, J.-W.; Lee, Y.-S.; Park, I.Y.; Ito, Y.; Bae, S.-C. Runx3 Plays a Critical Role in Restriction-Point and Defense against Cellular Transformation. Oncogene 2017, 36, 6884–6894. [Google Scholar] [CrossRef]
- Shiraha, H.; Nishina, S.; Yamamoto, K. Loss of Runt-Related Transcription Factor 3 Causes Development and Progression of Hepatocellular Carcinoma. J. Cell. Biochem. 2011, 112, 745–749. [Google Scholar] [CrossRef]
- Tanaka, S.; Shiraha, H.; Nakanishi, Y.; Nishina, S.-I.; Matsubara, M.; Horiguchi, S.; Takaoka, N.; Iwamuro, M.; Kataoka, J.; Kuwaki, K.; et al. Runt-Related Transcription Factor 3 Reverses Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma. Int. J. Cancer 2012, 131, 2537–2546. [Google Scholar] [CrossRef]
- Kataoka, J.; Shiraha, H.; Horiguchi, S.; Sawahara, H.; Uchida, D.; Nagahara, T.; Iwamuro, M.; Morimoto, H.; Takeuchi, Y.; Kuwaki, K.; et al. Loss of Runt-Related Transcription Factor 3 Induces Resistance to 5-Fluorouracil and Cisplatin in Hepatocellular Carcinoma. Oncology Reports 2016, 35, 2576–2582. [Google Scholar] [CrossRef]
- Xu, N.; Shen, C.; Luo, Y.; Xia, L.; Xue, F.; Xia, Q.; Zhang, J. Upregulated MiR-130a Increases Drug Resistance by Regulating RUNX3 and Wnt Signaling in Cisplatin-Treated HCC Cell. Biochemical and Biophysical Research Communications 2012, 425, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gao, N.; Shen, Y.; Cen, J. Hypermethylation Downregulates Runx3 Gene Expression and Its Restoration Suppresses Gastric Epithelial Cell Growth by Inducing P27 and Caspase3 in Human Gastric Cancer. Journal of Gastroenterology and Hepatology 2010, 25, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Gong, W.; Oh, S.C.; Li, Q.; Kim, W.D.; Wang, L.; Le, X.; Yao, J.; Wu, T.T.; Huang, S.; et al. Loss of RUNX3 Expression Significantly Affects the Clinical Outcome of Gastric Cancer Patients and Its Restoration Causes Drastic Suppression of Tumor Growth and Metastasis. Cancer Research 2005, 65, 4809–4816. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.-Z.; Yang, J.-O.; Lee, K.-Y.; Ito, K.; Sakakura, C.; Li, Q.-L.; Kim, H.-R.; Cha, E.-J.; Lee, Y.-H.; Kaneda, A.; et al. RUNX3 Suppresses Gastric Epithelial Cell Growth by Inducing P21 WAF1 / Cip1 Expression in Cooperation with Transforming Growth Factor β-Activated SMAD. Molecular and Cellular Biology 2005, 25, 8097–8107. [Google Scholar] [CrossRef]
- Lee, J.-W.; Kim, D.-M.; Jang, J.-W.; Park, T.-G.; Song, S.-H.; Lee, Y.-S.; Chi, X.-Z.; Park, I.Y.; Hyun, J.-W.; Ito, Y.; et al. RUNX3 Regulates Cell Cycle-Dependent Chromatin Dynamics by Functioning as a Pioneer Factor of the Restriction-Point. Nat Commun 2019, 10, 1897. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Miyazono, K. RUNX Transcription Factors as Key Targets of TGF-β Superfamily Signaling. Current Opinion in Genetics & Development 2003, 13, 43–47. [Google Scholar] [CrossRef]
- Yano, T.; Ito, K.; Fukamachi, H.; Chi, X.-Z.; Wee, H.-J.; Inoue, K.; Ida, H.; Bouillet, P.; Strasser, A.; Bae, S.-C.; et al. The RUNX3 Tumor Suppressor Upregulates Bim in Gastric Epithelial Cells Undergoing Transforming Growth Factorβ-Induced Apoptosis. Molecular and Cellular Biology 2006, 26, 4474–4488. [Google Scholar] [CrossRef]
- Bae, S.-C.; Choi, J.-K. Tumor Suppressor Activity of RUNX3. Oncogene 2004, 23, 4336–4340. [Google Scholar] [CrossRef]
- Zaidi, S.K.; Sullivan, A.J.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Integration of Runx and Smad Regulatory Signals at Transcriptionally Active Subnuclear Sites. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 8048–8053. [Google Scholar] [CrossRef]
- Zeng, S.; Shen, W.; Liu, L. Senescence and Cancer. Cancer Transl Med 2018, 4, 70. [Google Scholar] [CrossRef]
- Chen, Z.; Zuo, X.; Pu, L.; Zhang, Y.; Han, G.; Zhang, L.; Wu, J.; Wang, X. Circ LARP 4 Induces Cellular Senescence through Regulating MiR-761/ RUNX 3/P53/P21 Signaling in Hepatocellular Carcinoma. Cancer Sci 2019, 110, 568–581. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, K.; Cameron, E.R.; Blyth, K. Complex Interplay between the RUNX Transcription Factors and Wnt/β-Catenin Pathway in Cancer: A Tango in the Night. Mol Cells 2020, 43, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, W.; Cheng, N.; Wang, K.; Li, B.; Jiang, X.; Sun, S. Hepatitis C Virus-Induced up-Regulation of MicroRNA-155 Promotes Hepatocarcinogenesis by Activating Wnt Signaling. Hepatology 2012, 56, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Steinhart, Z.; Angers, S. Wnt Signaling in Development and Tissue Homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Chuang, L.S.H.; Ito, T.; Chang, T.L.; Fukamachi, H.; Salto–Tellez, M.; Ito, Y. Loss of Runx3 Is a Key Event in Inducing Precancerous State of the Stomach. Gastroenterology 2011, 140, 1536–1546. [Google Scholar] [CrossRef]
- Ito, K.; Lim, A.C.-B.; Salto-Tellez, M.; Motoda, L.; Osato, M.; Chuang, L.S.H.; Lee, C.W.L.; Voon, D.C.-C.; Koo, J.K.W.; Wang, H.; et al. RUNX3 Attenuates β-Catenin/T Cell Factors in Intestinal Tumorigenesis. Cancer Cell 2008, 14, 226–237. [Google Scholar] [CrossRef]
- Giovannini, C.; Fornari, F.; Piscaglia, F.; Gramantieri, L. Notch Signaling Regulation in HCC: From Hepatitis Virus to Non-Coding RNAs. Cells 2021, 10, 521. [Google Scholar] [CrossRef]
- Ahn, S.; Hyeon, J.; Park, C.-K. Notchl and Notch4 Are Markers for Poor Prognosis of Hepatocellular Carcinoma. Hepatobiliary & Pancreatic Diseases International 2013, 12, 286–294. [Google Scholar] [CrossRef]
- Nishina, S.-I. Restored Expression of the Tumor Suppressor Gene RUNX3 Reduces Cancer Stem Cells in Hepatocellular Carcinoma by Suppressing Jagged1-Notch Signaling. Oncol Rep 2011. [Google Scholar] [CrossRef]
- Chai, M.Y.; Kou, B.X.; Fu, Z.; Wei, F.L.; Dou, S.S.; Chen, D.X.; Liu, X.N. [Sorafenib regulates vascular endothelial growth factor by runt-related transcription factor-3 to inhibit angiogenesis in hepatocellular carcinoma]. Zhonghua Gan Zang Bing Za Zhi 2022, 30, 770–776. [Google Scholar] [CrossRef]
- Lee, S.H.; Bae, S.C.; Kim, K.W.; Lee, Y.M. RUNX3 Inhibits Hypoxia-Inducible Factor-1α Protein Stability by Interacting with Prolyl Hydroxylases in Gastric Cancer Cells. Oncogene 2014, 33, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Galle, P.R.; Forner, A.; Llovet, J.M.; Mazzaferro, V.; Piscaglia, F.; Raoul, J.-L.; Schirmacher, P.; Vilgrain, V. EASL Clinical Practice Guidelines: Management of Hepatocellular Carcinoma. Journal of Hepatology 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular Mechanisms of Epithelial–Mesenchymal Transition. Nat Rev Mol Cell Biol 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Voon, D.C.-C.; Wang, H.; Koo, J.K.W.; Nguyen, T.A.P.; Hor, Y.T.; Chu, Y.-S.; Ito, K.; Fukamachi, H.; Chan, S.L.; Thiery, J.P.; et al. Runx3 Protects Gastric Epithelial Cells Against Epithelial-Mesenchymal Transition-Induced Cellular Plasticity and Tumorigenicity. Stem Cells 2012, 30, 2088–2099. [Google Scholar] [CrossRef]
- Loh, C.-Y.; Chai, J.; Tang, T.; Wong, W.; Sethi, G.; Shanmugam, M.; Chong, P.; Looi, C. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, Y.D.; Choi, Y.S.; Lee, Y.M. Targeting of RUNX3 by MiR-130a and MiR-495 Cooperatively Increases Cell Proliferation and Tumor Angiogenesis in Gastric Cancer Cells. Oncotarget 2015, 6, 33269–33278. [Google Scholar] [CrossRef]
- Wang, M.; Wang, X.; Liu, W. MicroRNA-130a-3p Promotes the Proliferation and Inhibits the Apoptosis of Cervical Cancer Cells via Negative Regulation of RUNX3. Mol Med Rep 2020. [Google Scholar] [CrossRef]
Figure 1.
(A) RUNX1, RUNX2, and RUNX3 genes’ structure. The rectangles-exons, the lines-introns, the ATG-start codon; P1 and P2-promoters; the RHD-Runt homology domain; the TAD-transactivation domain. Grey rectangles-untranslated regions. (B) RUNX1, RUNX2, and RUNX3 proteins’ structure. The rectangles-protein domains. The numbers-the amino acids’ numbers. NLS-nuclear localisation signal; QA-the glutamine/alanine-rich signal, the RUNX2 specific; the ID-inhibitory domain; VWRPY-Groucho/TLE binding site. The figure is a not-to-scale drawing. We modified the figure, under CC BY NC, based on Yi et al. 2022 [10].
Figure 1.
(A) RUNX1, RUNX2, and RUNX3 genes’ structure. The rectangles-exons, the lines-introns, the ATG-start codon; P1 and P2-promoters; the RHD-Runt homology domain; the TAD-transactivation domain. Grey rectangles-untranslated regions. (B) RUNX1, RUNX2, and RUNX3 proteins’ structure. The rectangles-protein domains. The numbers-the amino acids’ numbers. NLS-nuclear localisation signal; QA-the glutamine/alanine-rich signal, the RUNX2 specific; the ID-inhibitory domain; VWRPY-Groucho/TLE binding site. The figure is a not-to-scale drawing. We modified the figure, under CC BY NC, based on Yi et al. 2022 [10].
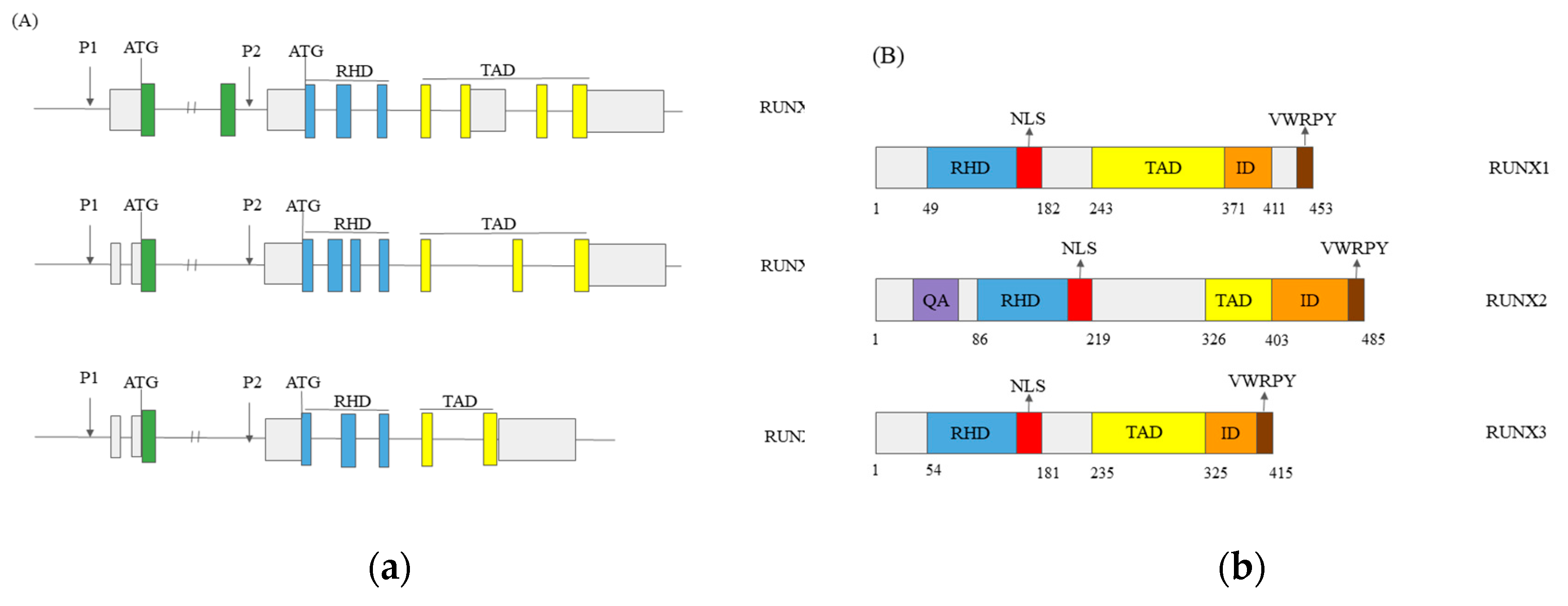
Figure 2.
Roles of RUNX1 in hepatocellular carcinoma. COL4A1↑- Collagen type IV alpha 1 chain increases; VEGF↓-Vascular endothelial growth factor decreases.
Figure 2.
Roles of RUNX1 in hepatocellular carcinoma. COL4A1↑- Collagen type IV alpha 1 chain increases; VEGF↓-Vascular endothelial growth factor decreases.
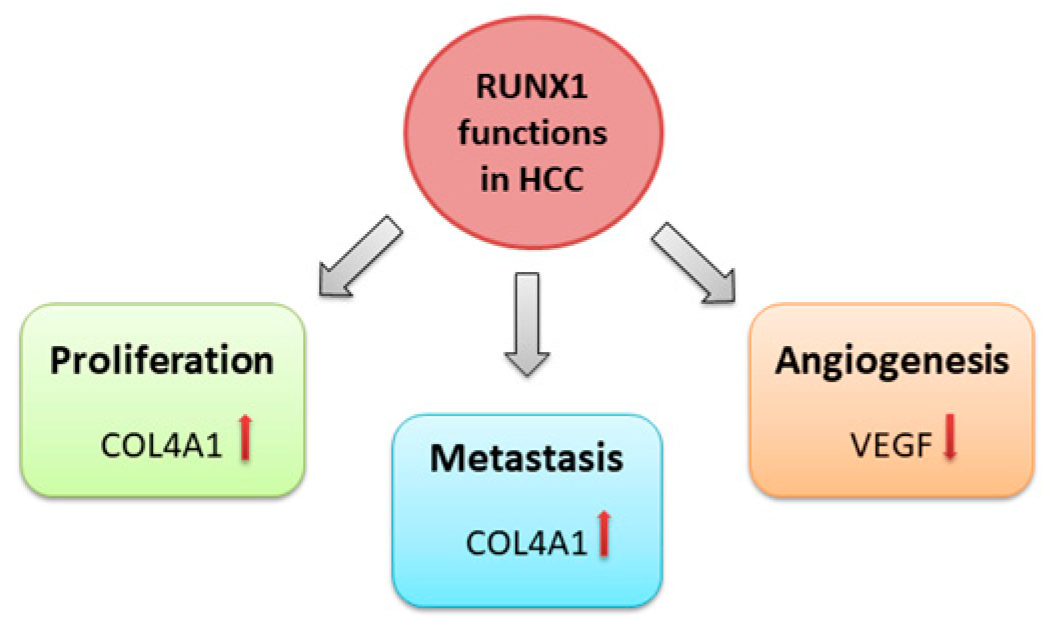
Figure 3.
RUNX2 oncogenic mechanisms in hepatocellular carcinoma.
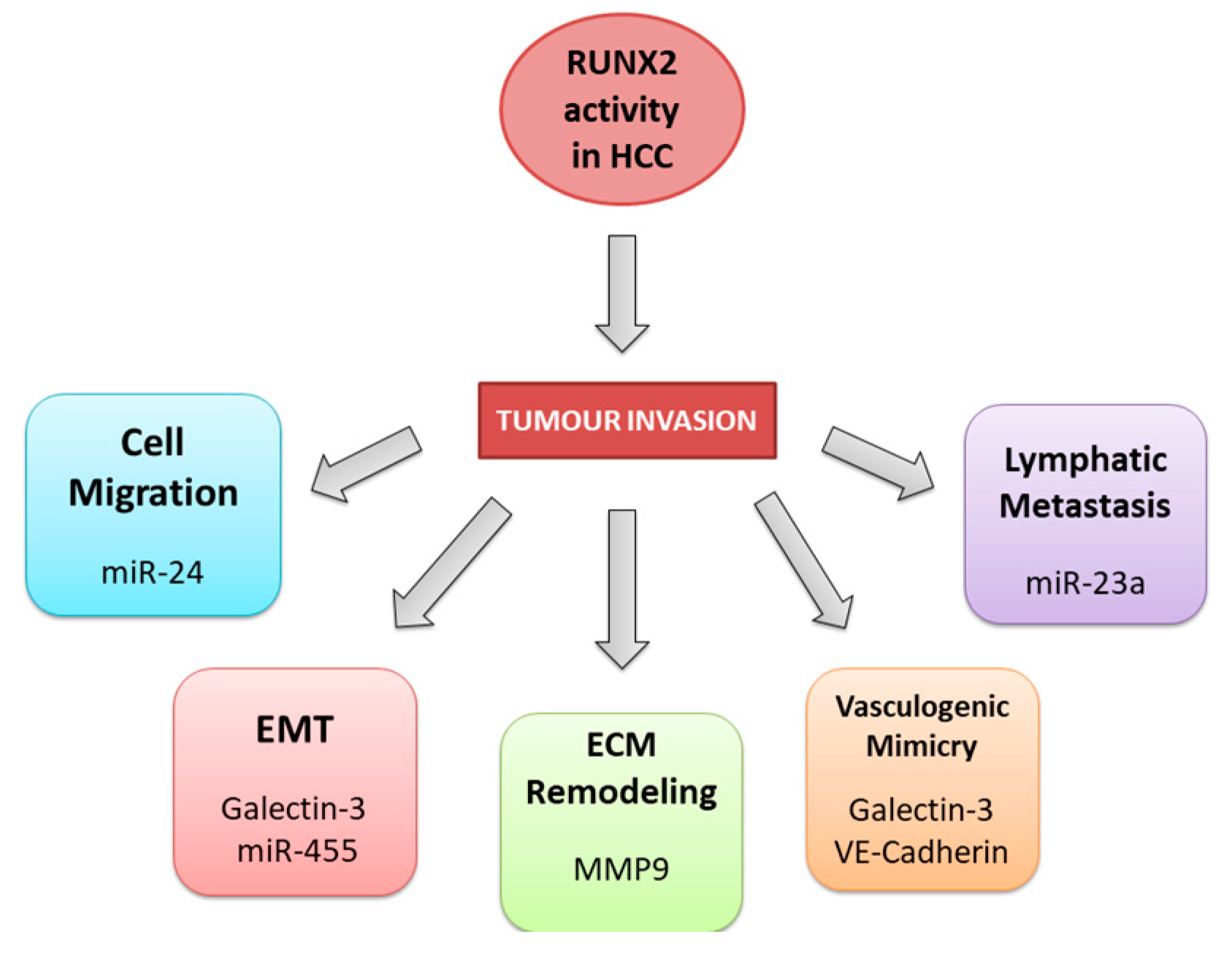
Figure 4.
The downregulation of RUNX3 in HCC.

Table 1.
Link between miRNAs or lncRNAs and RUNX genes in HCC.
| Gene | miRNA(s) or lncRNA(s) | References |
| RUNX1 | miR-632 | Sun et al., 2020 |
| miR-144 | Vivacqua et al., 2015 | |
| RUNX2 | miR-455 | Qin et al., 2016 |
| miR-196 | Wang et al., 2019 | |
| miR-24 | Wang et al., 2021 | |
| miR-23a | Huang et al., 2018 | |
| lncRNA HAND2 antisense RNA 1 (HAND2-AS1) | Jing et al., 2021 | |
| lncRNAmetallothionein 1D, pseudogene (MT1DP) | Yu et al., 2014 | |
| RUNX3 | miR-186 | Chen et al., 2019 |
| miR-130 | Gou et al., 2017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



