
Submitted:
28 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
The non-coding RNAs (ncRNAs) are indispensable for controlling genes and genetic programming during development, as well as for health and cardiovascular diseases. Cardiac arrhythmia is a frequent cardiovascular disease, which has a complex pathology. Recent studies have shown that ncRNAs are also associated with cardiac arrhythmias. Many non-coding RNAs and/or genomes have been reported as a genetic background for cardiac arrhythmias. Arrhythmias may be affected by several functional and structural changes in the myocardium. Therefore, ncRNAs might be indispensable regulators of gene expression in cardiomyocytes, which could play a dynamic role in regulating the stability of cardiac conduction and/or in the remodeling process. Although it remains almost unclear how ncRNAs regulate the expression of molecules for controlling the cardiac conduction and/or in the remodeling process, gut microbiota and immune system within the intricate networks might be involved in the regulatory mechanisms. This study would discuss them and provide a research basis for ncRNAs modulation, which might support the development of emerging innovative therapies against cardiac arrhythmias.
Keywords:
ncRNA
; lncRNA
; miRNA
; cardiac arrhythmia
; atrial fibrillation
; gut microbiota
; APRO family protein
1. Introduction
Cardiac arrhythmias are a set of heterogenic dysfunctions in the heart rhythm, which may be often defined as any variations in the rate of the normal heart. Abnormal impulse formations and disturbances in conduction may be two major reasons responsible for arrhythmias. However, the physiological dysfunctions linked to the specific arrhythmias are very intricate. The primary pace-making activity of the heart is determined by a spontaneous action potential within atrial node cells [1]. At present, it has been suggested that the distinctive pace generation might characteristically rely on probable membrane clock mechanism. The membrane clock consists of plasma membrane ion channels carrying an inward current [2]. Therefore, the rhythm-associated ion channels implicated in the initiation or progression of the action potential have been well documented. Most likely, arrhythmias are mainly caused by imbalances of these calcium ion channels and dysregulations of conduction in cardiac muscles. The identification of genetic components underlying the cardiac arrhythmias have also highlighted the specific role of ion channels. Most ion channels are protein complexes that are positioned in the cardiomyocyte sarcolemma, and they might play a role in calcium ion flow conduction [3].
Non-coding RNAs (ncRNAs) have been shown to regulate a variety of ion channels and/or intercellular linking proteins/molecules, suggesting that ncRNA may be an important regulator of cardiovascular diseases [4]. Therefore, we deliberate recent findings on the implication of ncRNAs in the development of the most common forms of arrhythmias with a focus on therapeutic and/or clinical application. In recent years, it has been revealed that ncRNA could regulate disease development [5]. The ncRNAs are around 200-nt base sequences that might regulate the genetic, epigenetic gene expression as well as intra-cellular signaling mechanisms [6]. In general, the ncRNAs have been used as biomarkers for diagnosis and/or treatment due to their involvement in the disease commencement. Hence, ncRNA has been also investigated in cardiovascular diseases [7]. (Figure 1) The ncRNAs have enormous importance in clinical applications, which are usually categorized into several categories such as miRNAs, circRNAs, and/or lncRNAs [8]. Recent studies have shown that the association between lncRNAs and protein may be related to the cardiac arrhythmias. Short interference RNA (siRNA) could be often used to target the specific ncRNA. RNAi-mediated siRNAs are highly adjustable, which could be used to suppress their mRNA’s encoding protein. The miRNAs are approximately 22-nucleotide RNA molecules that could regulate cell signaling and/or decrease the expression of specific genes by modifying the translation procedure [9].
Although more than a few arrhythmogenic mechanisms such as alterations in calcium ion channels function and/or oxidative stresses in cardiac cells have been widely studied, there are still some vacancies to be clarified. Somewhat, the understanding of pathogenesis for the prevention and/or treatment of cardiac arrhythmias without any side effects has become an important issue to be solved urgently. In this regards, the gut microbiota might have a significant impact on cardiac arrhythmias through a variety of mechanisms, suggesting a broad range of conceivable treatment options [10]. In addition, a related study suggest that cardiac arrhythmias may associate with nutrient intake and/or some metabolisms [11]. The exploration of the related mechanisms of dysbiosis of the gut microbiota including the increase of harmful metabolites has been increasingly discovered. On this basis, it has been shown that abnormal metabolites of gut dysbiosis might be linked to cardiac arrhythmias [12]. Here, a comprehensive arrhythmogenicity study have been briefly summarized, which might provide theoretical basis for novel treatment strategies inspiring the prevention and/or treatment of cardiac arrhythmias.

Figure 1.
Schematic representation of the players of ncRNAs involved in the cardiac arrhythmias. Example ncRNA molecules are shown for each cardiac arrhythmia.
Figure 1.
Schematic representation of the players of ncRNAs involved in the cardiac arrhythmias. Example ncRNA molecules are shown for each cardiac arrhythmia.
2. Atrial Fibrillation
Atrial fibrillation is an asymmetrical heart rhythm, which might commonly lead to heart palpitations. Interestingly, individuals carrying the non-coding 4q25 locus near the PITX2 gene were 60% more susceptible to this abnormal heart rhythm [13]. Similarly, lncRNAs may be potentially associated with the development of Atrial fibrillation. Several lncRNAs including LINC00844, RP11-532N4.2, UNC5B-AS1, RP3-332B22.1, RP11-432J24.5, and RP11-557H15.4 have been shown to be differentially expressed in patients of atrial fibrillation compared to normal individuals, whose target might be related to the calcium signaling pathway and/or toll-like receptor signaling pathway [14, 15]. In addition, several miRNAs including miR-135a might be involved in the process of atrial fibrillation.
Atrial fibrillation is the most prevalent type of arrhythmia worldwide [16 ]. Atrial fibrosis is a hallmark feature of atrial structural remodeling in atrial fibrillation, which is regulated by the TGF-β1/Smad3 pathway. The dysregulation of tissue blockers of matrix metalloproteinases including MMPs and TIMPs linked to collagen upregulation is also a key factor in the development of atrial fibrillation. The miR-146b-5p might be an inhibitor of TIMPs protein expression, which may be increased in atrial tissue with atrial fibrillation. Furthermore, some profibrotic markers, such as MMP9, TGFβ1, and COL1A1, were downregulated upon miR-146b-5p knockdown, whereas TIMP4 caused inversed expression patterns [17]. Another mechanism underlying atrial fibrillation is ferroptosis, which is defined as iron-dependent cell death due to excessive accumulation of peroxidized polyunsaturated fatty acids. Interestingly, the occurrence of ferroptosis in atrial fibrillation has been shown to be promoted by miR-23a-3p. In addition, SLC7A11 is a direct target of miR-23a-3p, which might be also associated with ferroptosis [18]. Alternative miR-dependent gene regulation could elicit the initiation and progression of atrial fibrillation. For example, miR-34a could upregulate the expression of tandem of P domains in a fragile inner repairing K+ channel-linked acid-sensitive K+ channel 1 (TASK-1), which might promote a reduction in the resting membrane potential [19].
3. Bradycardia and Tachycardia
Bradycardia is a heart dysfunction described by a decreased heart rate mostly lower than 50 beats/min. The main etiological factors of bradycardia may include the dysregulation of sinus, atrial or junctional bradycardia, and/or an intricate transmission system with atrioventricular block. The most frequent form of bradycardia asymptomatically takes place during sleep and/or in athletes [20]. Bradycardia is usually regulated by a range of molecules including ncRNAs at a molecular levels. Transcription factors also control the expression of genes involved in BA by modulating a variety of effectors including miRNAs. For example, it has been demonstrated that the function of miR-370-3p could contribute to the development of bradycardia [21], which might be triggered via the reduced function of sinus node pacemaker channels with a related reduction of the ionic current. The miR-370-3p could directly bind to the mRNA of HCN4 to suppress the activity of HCN4 [21]. Similarly, the miR-486-3p could suppress the HCN4 inducing a sinus node dysregulation such as bradycardia [22, 23]. Supraventricular arrhythmia is an example of tachycardia that starts in the upper compartments of the heart.Tachycardia is known to be considered as a fast heartbeat, which might be also regulated by a range of molecules including ncRNAs at a molecular levels. For example, the expression of miR-1183 may be considerably upregulated in tachycardia. Elevated expression of miR-1183 might serve as a tissue biomarker for atrial remodeling, which might be of potential significance in cardiac disease [24]. Similarly, transcriptomic data from the urine sample have revealed significantly reduced levels of miRNAs including miR-423, miR-1180, miR-3162, miR-319,7 miR-3613, miR-6511, miR-6763. The changes in these miRs were associated with increased expression of profibrotic markers, such as Col I, Col III, fibronectin, and TGF-β. MiR-423 specifically was shown to regulate calcium-handling proteins, such as the phosphorylated calmodulin-dependent protein kinase II, which might regulate the development of tachycardia [25].
4. Other cardiac rhythm disorders
Ventricular arrhythmias are described as irregular heartbeats that derive from the ventricles. The origin of ventricular arrhythmias might be sympathetic remodeling that originates in myocardial infarction. Various ncRNAs are important regulators of inflammation and sympathetic remodeling following the myocardial infarction. Several lncRNAs are potentially implicated in ventricular arrhythmias following myocardial infarction. For example, the lncRNA LOC100911717 (LOC10) might be increased in cardiac cells and macrophages at the infarcted heart area [26]. Another important factor of ventricular arrhythmias may be NLRP3 inflammasomes. It has been demonstrated that the role of SOX2-OT lncRNA in regulating NLRP3 inflammasome-mediated ventricular arrhythmias. The levels of SOX2-OT and NLRP3 inflammasomes could be increased after ventricular arrhythmias [27]. Similarly, the miR-2355-3p and miR-1231 may play key roles in the regulation of ventricular arrhythmias [28, 29].
Long QT and short QT syndrome are known as the entity of Brugada syndrome [30]. Risk stratification for sudden cardiac death in patients with Brugada syndrome remains a major challenge. Brugada patients display a distinct miRNA expression profile compared with unaffected control individuals. For example, the miR-145-5p and miR-585-3p are associated with the standing position of Brugada patients, which might be the potential utility of leucocyte-derived miRNAs as prognostic biomarkers for Brugada syndrome [31, 32]. Arrhythmogenic cardiomyopathy is a genetically determined disorder caused by mutations in proteins constituting desmosomes, which is an inherited cardiomyopathy histologically characterized by the replacement of myocardium by fibrofatty infiltration, cardiomyocyte loss, and inflammation [33]. Inflammation might be also involved in the pathogenesis of arrhythmogenic cardiomyopathy with various inflammatory cytokines [34]. Consistently, it has been suggested that circulating levels of cytokine receptors (IL1-R1, IL6-R, TNF-R1/R2) may be elevated and associated with the risk of arrhythmias in arrhythmogenic cardiomyopathy [35-37]. In particular, myocardial expression of IL-17 and/or TNF is substantially enhanced in all cases of arrhythmogenic cardiomyopathy [38]. In a large cohort of samples, the miR-122-5p, miR-133b, miR-133a-3p, miR-142-3p, miR-183-5p and miR-182-5p could possess high diagnostic potential in arrhythmogenic cardiomyopathy patients [39, 40]. The implication of miRNAs in the pathogenesis of arrhythmogenic cardiomyopathy has been the objective of multiple studies.
5. Gut microbiota and cardiac arrhythmia
Disruption of gut microbiota could induce cardiorespiratory morbidity. Consistently, modulation of autonomic homeostasis via the gut microbiota-brain axis could control the heart rate [41]. The adult gut system might be an extremely diverse and dynamic ecosystem [42, 43], which exert specific physiological functions including immune regulation, gut mucosal protection, and conservation of nutritional metabolism in addition to the development of several diseases [44]. Microbiota elements can directly or indirectly trigger vagus nerve afferent fibers through the gut endocrine cells, which could stimulate the central nervous system (CNS). Depending on the substance, the different SCFAs produced by microbiota maybe activate vagal afferent fibers in different ways. For example, butyric acid, a short fatty acid, directly affects afferent terminals [45, 46]. An increase in the concentration of butyric acid at the colon may produce a significant hypotensive effect which depends on the afferent colonic vagus nerve signaling and GPR41/43 receptors [46]. Several studies have investigated the effects of food components on gut microbiota, which could be an imperative target for the forthcoming treatment of arrhythmias through the modification of gut microbiota. For example, patients with arrhythmias may prefer to take further energy from animal fat [47]. Therefore, patients with cardiac arrhythmias are often found with atherosclerosis. In addition, diabetes may be also a frequent co-morbidity in individuals with arrhythmias. Furthermore, the interaction between some drugs and their impact on gut microbiota may result in uncontrolled arrhythmias [48-50]. Recent increasing data suggest that regulatory non-coding RNAs such as miRNAs, circular RNAs, and lncRNA may affect host-microbe interactions. These ncRNAs have been also suggested as potential biomarkers in microbiome-associated disorders with a direct cross-talk between microbiome composition and ncRNAs [51]. Epigenetically, miR-23a-3p could lead to Th17/Treg imbalance and participate in the progression of Graves' disease [52], which might be also involved in the mechanism underlying atrial fibrillation. Gut microbiota plays a vital role in the pathogenesis both of atrial fibrillation and Graves' disease [52]. The imbalance of Th17/Treg cells induced by the alteration of gut microbiota plays a vital role in the pathogenesis of arrhythmias [53, 54]. Interestingly, altered composition of gut microbiota could change the expression of hepatic miR-34a [55], which might be also involved in the development of atrial fibrillation. A negative correlation between miR-122-5p and the intestinal bacteria including Bacteriodes. uniformis and Phascolarctobacterium. Faecium has been revealed, suggesting that the crosstalke between miRNA and gut microbiota could regulate the intracellular signal transduction by controlling key genes [56]. As mentioned before, the miR-122-5p could possess great diagnostic potential in arrhythmogenic cardiomyopathy patients [40, 57]. (Figure 2)
In these ways, evidence from numerous studies in both humans and animals suggests that the role of the gut microbiota and its metabolites in arrhythmia has been well established. Through a variety of pathways, the gut microbiota significantly affects cardiac arrhythmia, which may provide a wide range of potential therapeutic approaches [58, 59]. Linking the possible interfering role of gut microbiota in the pharmacokinetics of drugs used in management of cardiac arrhythmias would help to better understand the potential of probiotics [60, 61].
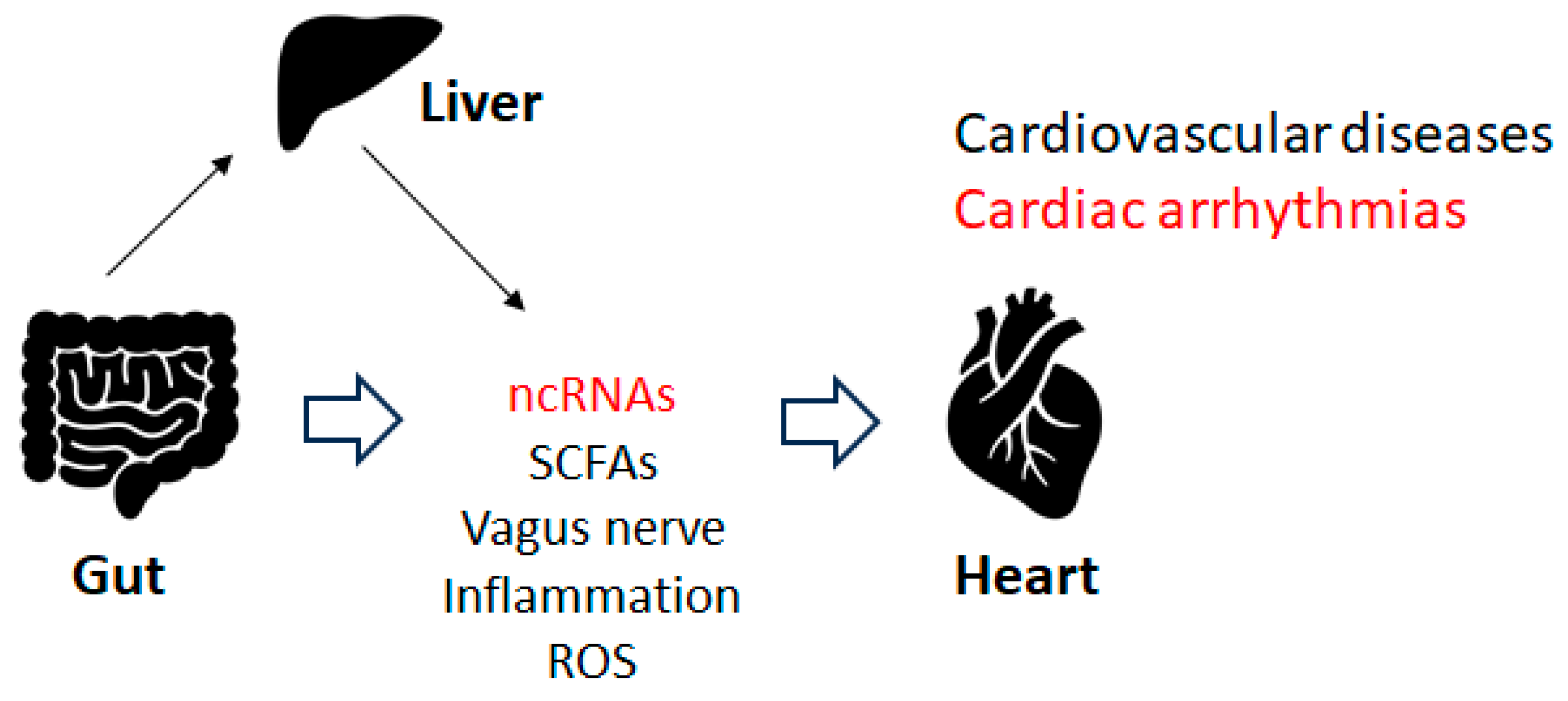
Figure 2.
Several factors and/or inflammation with ROS may affect the development of cardiovascular diseases including cardiac arrhythmias. Note that some critical pathways have been omitted for clarity.
Figure 2.
Several factors and/or inflammation with ROS may affect the development of cardiovascular diseases including cardiac arrhythmias. Note that some critical pathways have been omitted for clarity.
6. Immune pathway and cardiac arrhythmias
There is increasing attention on the molecular mechanisms controlling the onset and the progression points of cardiac arrhythmias to the complex cell to cell interplay that triggers immune cells enhancing atrial fibrosis [62]. The dysregulation of immune pathway might meaningfully contribute to ion channel dysfunction and the initiation of cardiac arrhythmias [63]. In turn, the interaction between immune cells and atrial myocytes may enhance atrial electrical and functional remodeling, which is responsible for the progression of cardiac arrhythmias [64]. Interestingly, some ncRNAs are involved in the ribonucleolytic cleavage with the target of mRNA [65], in which several members of the APRO family have been shown to be associated with the cytoplasmic mRNA deadenylation [66, 67]. Likewise, some of the APRO family proteins could interact with the poly (A) ribonuclease complex [68, 69]. These APRO family proteins (Btg1, Btg2, Tob1 and Tob2) might be a key modulator of microRNAs [70], which might be deeply involved in the regulation of immune cells [71]. Therefore, APRO family proteins have been shown to be involved in immune-related disorders including ulcerative colitis and cancers [72-74]. Interestingly, it has been shown that one of the APRO family member, BTG1, may be involved in the regulation of ion channel related gene expression [75, 76]. Remarkably, it has been shown that the BTG1 could promote the deadenylation and/or degradation of mRNA to secure T cell quiescence [77], suggesting that BTG1 might be involved in a key mechanism underlying T cell quiescence. In addition, another APRO family member, Tob1, might be a key regulatory molecule in the process of endothelialization in the repair progress of atrial septal defects [78]. Tob1 might also play a fundamental role in keeping cells to a quiescent position, thereby obstructing cellular proliferation of normal and/or cancer cells [79]. TOB1 is inactivated in many human cancers. Therefore, TOB1 has been considered as a tumor-suppressor protein [80].
7. Future perspectives
Nevertheless, the immune-regulating potential of GM mediated some anti-cancer therapeutic functions, specifically anti-CTLA-4 and anti-PD-L1 targeting drugs that showed better outcomes in patients with some species of Bacteroides and Bifidobacterium, respectively [81, 82]. These actions are hypothesized to be related to the action of GM on DCs which are essential to trigger proper T-cells response [83]. Additionally, short-chain fatty acids (SCFAs) produced in the gut may modulate anti-CTLA-4 and anti-PD-1 stimulated immune responses and their anti-tumor efficacy [84]. SCFAs are characterized by containing fewer than six carbons, such as acetate, propionate, and butyrate, which are mainly produced by the gut microbiota as fermentation products. The human gut microbiota and/or their metabolites including SCFAs have also become a potential therapeutic target for the development of interventions for the prevention of cardio-metabolic disorders [85]. There are noteworthy associations between gut microbiota and the microglia in the brain, suggesting that the crosstalk of gut microbiota and nerve function, based on the roles of microglia [86]. Therefore, some modifications in the composition of gut microbiota might play a critical role in the pathogenesis of several diseases. Remarkably, we presume that cardiovascular diseases including cardiac arrhythmia might also relate to the pathogenesis that is based on the engram memory system [87-89]. (Figure 3) The engram memory system in the brain could retain the information of a certain inflammation in the body which might be involved in the pathogenesis of various diseases including cardiovascular diseases, in which the immunity-linked processes might be associated with the neuronal responses to memory engrams [90]. Future work should define at the molecular level how this pathway could precisely interfere to regulate the cardiac arrhythmias. In particular, forthcoming research should focus on the identification of disease-specific engram retention over time during the latent period of cardiovascular diseases.
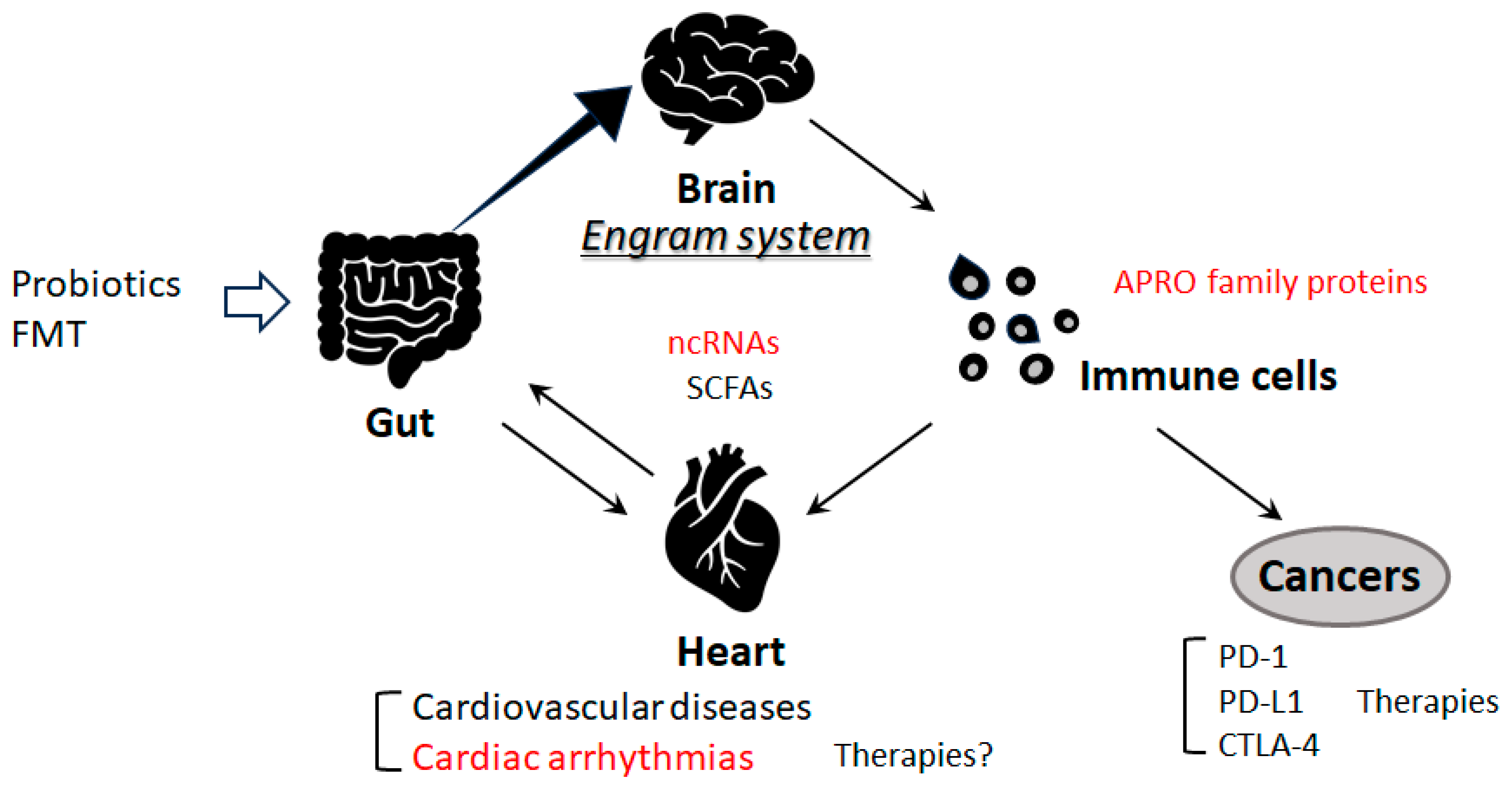
Figure 3.
Schematic representation of the potential inhibitory tactics against the pathogenesis of cardiovascular diseases including cardiac arrhythmias. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) could contribute to the alteration of gut microbial community for the alteration of ncRNA and/or SCFAs production, which could be beneficial for the treatment of cardiovascular diseases. Note that several important activities such as inflammatory reaction, autophagy initiation, and ROS production have been omitted for clarity.
Figure 3.
Schematic representation of the potential inhibitory tactics against the pathogenesis of cardiovascular diseases including cardiac arrhythmias. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) could contribute to the alteration of gut microbial community for the alteration of ncRNA and/or SCFAs production, which could be beneficial for the treatment of cardiovascular diseases. Note that several important activities such as inflammatory reaction, autophagy initiation, and ROS production have been omitted for clarity.
8. Conclusion
Several ncRNAs could be involved in the development of certain cardiac arrhythmias, in which the association between gut and heart might play an important role. In addition, the correlation between gut and brain as well as the correlation between brain and immunity might also influence the development of cardiovascular diseases including cardiac arrhythmias. An in-depth knowledge of the role of ncRNAs in cardiac arrhythmias may be valuable for progressing new clinical diagnosis and treatment.
Author Contributions
Conceptualization: NS, YI, and SM; original draft preparation and editing, NS, YI, SY, KT, HS, and SM; visualization, NS, YI and SM; supervision, SM. Each author (NS, YI, SY, KT, HS, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing financial interests.
Abbreviations
| APROs | anti-proliferative proteins |
| BTG1 | B cell translocation gene 1 |
| CNS | central nervous system |
| circRNA | circular RNA |
| CTLA-4 | cytotoxic T lymphocyte-associated protein-4 |
| FMT | fecal microbiota transplantation |
| LOC10 | LOC100911717 |
| lncRNAs | long non-coding RNAs |
| mRNA | messenger RNA |
| ncRNA | non-coding RNA |
| PD-1 | programmed cell death protein 1 |
| PD-L1 | programmed cell death ligand 1 |
| QOL | quality of life |
| ROS | reactive oxygen species |
| SCFAs | short-chain fatty acids |
| siRNA | short interference RNA |
| TASK-1 | tandem of P domains in a fragile inner repairing K+ channel-linked acid-sensitive K+ channel 1 |
| Tob1 | transducer of ErbB2 1 |
| UTR | untranslated region |
References
- Fan, W.; Sun, X.; Yang, C.; Wan, J.; Luo, H.; Liao, B. Pacemaker activity and ion channels in the sinoatrial node cells: MicroRNAs and arrhythmia. Prog. Biophys. Mol. Biol. 2023, 177, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Yaniv, Y.; Lakatta, E.G.; Maltsev, V.A. From two competing oscillators to one coupled-clock pacemaker cell system. Front. Physiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Wilde, A.A.M.; Bezzina, C.R. Genetics of cardiac arrhythmias. Hear. 2005, 91, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, S.; Moukette, B.; Hayasaka, T.; Haskell, A.K.; Mah, J.; Sepúlveda, M.N.; Tang, Y.; Kim, I.-M. Noncoding RNAs as Key Regulators for Cardiac Development and Cardiovascular Diseases. J. Cardiovasc. Dev. Dis. 2023, 10, 166. [Google Scholar] [CrossRef]
- Sallam, T.; Sandhu, J.; Tontonoz, P. Long Noncoding RNA Discovery in Cardiovascular Disease: Decoding Form to Function. Circ. Res. 2018, 122, 155–166. [Google Scholar] [CrossRef]
- Zhang, C.; Han, B.; Xu, T.; Li, D. The biological function and potential mechanism of long non-coding RNAs in cardiovascular disease. J. Cell. Mol. Med. 2020, 24, 12900–12909. [Google Scholar] [CrossRef]
- Correia, C.C.M.; Rodrigues, L.F.; Pelozin, B.R.d.A.; Oliveira, E.M.; Fernandes, T. Long Non-Coding RNAs in Cardiovascular Diseases: Potential Function as Biomarkers and Therapeutic Targets of Exercise Training. Non-Coding RNA 2021, 7, 65. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef]
- Lu, P.; Ding, F.; Xiang, Y.K.; Hao, L.; Zhao, M. Noncoding RNAs in Cardiac Hypertrophy and Heart Failure. Cells 2022, 11, 777. [Google Scholar] [CrossRef]
- Fan, H.; Liu, X.; Ren, Z.; Fei, X.; Luo, J.; Yang, X.; Xue, Y.; Zhang, F.; Liang, B. Gut microbiota and cardiac arrhythmia. Front. Cell. Infect. Microbiol. 2023, 13. [Google Scholar] [CrossRef]
- Nalliah, C.J.; Sanders, P.; Kalman, J.M. The Impact of Diet and Lifestyle on Atrial Fibrillation. Curr. Cardiol. Rep. 2018, 20, 137. [Google Scholar] [CrossRef] [PubMed]
- Svingen, G.F.; Zuo, H.; Ueland, P.M.; Seifert, R.; Løland, K.H.; Pedersen, E.R.; Schuster, P.M.; Karlsson, T.; Tell, G.S.; Schartum-Hansen, H.; et al. Increased plasma trimethylamine- N -oxide is associated with incident atrial fibrillation. Int. J. Cardiol. 2018, 267, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Gudbjartsson, D.F.; Arnar, D.O.; Helgadottir, A.; Gretarsdottir, S.; Holm, H.; Sigurdsson, A.; Jonasdottir, A.; Baker, A.; Thorleifsson, G.; Kristjansson, K.; et al. Variants conferring risk of atrial fibrillation on chromosome 4q25. Nature 2007, 448, 353–357. [Google Scholar] [CrossRef]
- Ruan, Z.-B.; Wang, F.; Gongben, B.-D.; Chen, G.-C.; Zhu, L. Identification of Circulating lncRNA Expression Profiles in Patients with Atrial Fibrillation. Dis. Markers 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Xie, L.; Huang, G.; Gao, M.; Huang, J.; Li, H.; Xia, H.; Xiang, X.; Wu, S.; Ruan, Y. Identification of Atrial Fibrillation-Related lncRNA Based on Bioinformatic Analysis. Dis. Markers 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Chugh, SS.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, EJ.; Gillum, RF.; Kim, YH.; McAnulty, JH Jr.; Zheng, ZJ.; et al. Worldwide epidemiology of atrial fibrillation: a Global Burden of Disease 2010 Study. Circulation. 2014, 129(8), 83. [CrossRef]
- Ye, Q.; Liu, Q.; Ma, X.; Bai, S.; Chen, P.; Zhao, Y.; Bai, C. ; Liu, Y, J Cell Mol Med. 2021, 25(22), 10543. [Google Scholar] [CrossRef]
- Liu, D.; Yang, M.; Yao, Y.; He, S.; Wang, Y.; Cao, Z.; Chen, H.; Fu, Y.; Liu, H.; Zhao, Q. Cardiac Fibroblasts Promote Ferroptosis in Atrial Fibrillation by Secreting Exo-miR-23a-3p Targeting SLC7A11. Oxid Med Cell Longev. 2022, 29; 3961495. [CrossRef]
- Wiedmann, F.; Kraft, M.; Kallenberger, S.; Büscher, A.; Paasche, A.; Blochberger, P.L.; Seeger, T.; Jávorszky, N.; Warnecke, G.; Arif, R.; et al. MicroRNAs Regulate TASK-1 and Are Linked to Myocardial Dilatation in Atrial Fibrillation. J. Am. Hear. Assoc. 2022, 11, e023472. [Google Scholar] [CrossRef] [PubMed]
- Barstow, C.; McDivitt, J.D. Cardiovascular Disease Update: Bradyarrhythmias. FP essentials 2017, 454, 18–23. [Google Scholar]
- Yanni, J.; D’souza, A.; Wang, Y.; Li, N.; Hansen, B.J.; Zakharkin, S.O.; Smith, M.; Hayward, C.; Whitson, B.A.; Mohler, P.J.; et al. Silencing miR-370-3p rescues funny current and sinus node function in heart failure. Sci. Rep. 2020, 10, 1–23. [Google Scholar] [CrossRef]
- Petkova, M.; Atkinson, A.J.; Yanni, J.; Stuart, L.; Aminu, A.J.; Ivanova, A.D.; Pustovit, K.B.; Geragthy, C.; Feather, A.; Li, N.; et al. Identification of Key Small Non-Coding MicroRNAs Controlling Pacemaker Mechanisms in the Human Sinus Node. J. Am. Hear. Assoc. 2020, 9, e016590. [Google Scholar] [CrossRef]
- Aminu, A.J.; Petkova, M.; Atkinson, A.J.; Yanni, J.; Morris, A.D.; Simms, R.T.; Chen, W.; Yin, Z.; Kuniewicz, M.; Holda, M.K.; et al. Further insights into the molecular complexity of the human sinus node – The role of ‘novel’ transcription factors and microRNAs. Prog. Biophys. Mol. Biol. 2021, 166, 86–104. [Google Scholar] [CrossRef]
- Djalinac, N.; Kolesnik, E.; Maechler, H.; Scheruebel-Posch, S.; Pelzmann, B.; Rainer, P.P.; Foessl, I.; Wallner, M.; Scherr, D.; Heinemann, A.; et al. miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium. Int. J. Mol. Sci. 2022, 23, 6962. [Google Scholar] [CrossRef]
- Park, H.; Park, H.; Park, J. Circulating microRNA 423 attenuates the phosphorylation of calcium handling proteins in atrial fibrillation. Mol Med Rep. 2022, 25(5), 186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tan, Z.; Zhou, J.; Wu, Y.; Hu,Q.; Ling, Q.; Ling, J.; Liu, M.; Ma, J.; Zhang, D.; et al. The regulation of circRNA and lncRNAprotein binding in cardiovascular diseases: Emerging therapeutic targets. Biomed Pharmacother. 2023, 165, 115067.
- Liang, Y.; Wang, B.; Huang, H.; Wang, M.; Wu, Q.; Zhao, Y.; He, Y. Silenced SOX2-OT alleviates ventricular arrhythmia associated with heart failure by inhibiting NLRP3 expression via regulating miR-2355-3p. Immunity, Inflamm. Dis. 2020, 9, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Mao, R.; Zhang, L.; Zhao, R.; Tan, K.; Liu, C.; Xu, J.; Du, G.; Zhang, T. CircNPHP4 in monocyte-derived small extracellular vesicles controls heterogeneous adhesion in coronary heart atherosclerotic disease. Cell Death Dis. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Qiao, L.; Han, F.; Xie, X.; Wang, W. MiR-1231 regulates L-calcium in ventricular arrhythmia in chronic heart failure. 2021, 112, 305–306. [CrossRef]
- Sarquella-Brugada, G.; Cesar, S.; Zambrano, M.D.; Fernandez-Falgueras, A.; Fiol, V.; Iglesias, A.; Torres, F.; Garcia-Algar, O.; Arbelo, E.; Brugada, J.; et al. Electrocardiographic Assessment and Genetic Analysis in Neonates: a Current Topic of Discussion. Curr. Cardiol. Rev. 2018, 15, 30–37. [Google Scholar] [CrossRef]
- Steinberg, C.; Gaudreault, N.; I Papadakis, A.; Henry, C.; Champagne, J.; Philippon, F.; O’hara, G.; Blier, L.; Plourde, B.; Nault, I.; et al. Leucocyte-derived micro-RNAs as candidate biomarkers in Brugada syndrome. Eur. 2023, 25. [Google Scholar] [CrossRef]
- Scumaci, D.; Oliva, A.; Concolino, A.; Curcio, A.; Fiumara, CV.; ammè, L.; Campuzano, O.; Pascali, VL.; Coll, M.; Iglesias, A.; et al. Integration of "Omics" Strategies for Biomarkers Discovery and for the Elucidation of Molecular Mechanisms Underlying Brugada Syndrome. Proteomics Clin Appl. 2018, 12(6), e1800065. [Google Scholar] [CrossRef] [PubMed]
- Piquer-Gil, M.; Domenech-Dauder, S.; Sepúlveda-Gómez, M.; Machí-Camacho, C.; Braza-Boïls, A.; Zorio, E. Non Coding RNAs as Regulators of Wnt/β-Catenin and Hippo Pathways in Arrhythmogenic Cardiomyopathy. Biomedicines 2022, 10, 2619. [Google Scholar] [CrossRef] [PubMed]
- Asatryan, B.; Asimaki, A.; Landstrom, A.P.; Khanji, M.Y.; Odening, K.E.; Cooper, L.T.; Marchlinski, F.E.; Gelzer, A.R.; Semsarian, C.; Reichlin, T.; et al. Inflammation and Immune Response in Arrhythmogenic Cardiomyopathy: State-of-the-Art Review. Circulation 2021, 144, 1646–1655. [Google Scholar] [CrossRef]
- Broch, K.; Leren, I.S.; Saberniak, J.; Ueland, T.; Edvardsen, T.; Gullestad, L.; Haugaa, K.H. Soluble ST2 is associated with disease severity in arrhythmogenic right ventricular cardiomyopathy. Biomarkers 2017, 22, 367–371. [Google Scholar] [CrossRef]
- Christensen, A.H.; Platonov, P.G.; Jensen, H.K.; Chivulescu, M.; Svensson, A.; Dahlberg, P.; Madsen, T.; Frederiksen, T.C.; Heliö, T.; Lie. H.; et al. Genotype–phenotype correlation in arrhythmogenic right ventricular cardiomyopathy—risk of arrhythmias and heart failure. J. Med Genet. 2021, 59, 858–864. [Google Scholar] [CrossRef]
- Cappelletto, C.; Gregorio, C.; Barbati, G.; Romani, S.; De Luca, A.; Merlo, M.; Mestroni, L.; Stolfo, D.; Sinagra, G. Antiarrhythmic therapy and risk of cumulative ventricular arrhythmias in arrhythmogenic right ventricle cardiomyopathy. Int. J. Cardiol. 2021, 334, 58–64. [Google Scholar] [CrossRef]
- Asimaki, A.; Tandri, H.; Duffy, ER.; Winterfield, JR.; Mackey-Bojack, S.; Picken, MM.; Cooper, LT.; Wilber, DJ.; Marcus, FI.; Basso, C.; et al. Altered desmosomal proteins in granulomatous myocarditis and potential pathogenic links to arrhythmogenic right ventricular cardiomyopathy. Circ Arrhythm Electrophysiol. 2011, 4(5):, 743. [CrossRef]
- Bueno Marinas, M.; Celeghin, R.; Cason, M.; Thiene, G.; Basso, C.; Pilichou, K. The Role of MicroRNAs in Arrhythmogenic Cardiomyopathy: Biomarkers or Innocent Bystanders of Disease Progression? Int J Mol Sci. 2020, 21(17), 6434. [Google Scholar] [CrossRef]
- Marinas, M.B.; Celeghin, R.; Cason, M.; Bariani, R.; Frigo, A.C.; Jager, J.; Syrris, P.; Elliott, P.M.; Bauce, B.; Thiene, G.; et al. A microRNA Expression Profile as Non-Invasive Biomarker in a Large Arrhythmogenic Cardiomyopathy Cohort. Int. J. Mol. Sci. 2020, 21, 1536. [Google Scholar] [CrossRef]
- Lucking, E.F.; O'Connor, K.M.; Strain, C.R.; Fouhy, F.; Bastiaanssen, T.F.; Burns, D.P.; Golubeva, A.V.; Stanton, C.; Clarke, G.; Cryan, J.F.; et al. Chronic intermittent hypoxia disrupts cardiorespiratory homeostasis and gut microbiota composition in adult male guinea-pigs. EBioMedicine 2018, 38, 191–205. [Google Scholar] [CrossRef]
- Anthony, W.E.; Wang, B.; Sukhum, K.V.; D’souza, A.W.; Hink, T.; Cass, C.; Seiler, S.; Reske, K.A.; Coon, C.; Dubberke, E.R.; et al. Acute and persistent effects of commonly used antibiotics on the gut microbiome and resistome in healthy adults. Cell Rep. 2022, 39, 110649–110649. [Google Scholar] [CrossRef]
- Tierney, BT.; Yang, Z.; Luber, JM.; Beaudin, M.; Wibowo, MC,.; Baek, C.; Mehlenbacher, E.; Patel, CJ.; Kostic, AD. The Landscape of Genetic Content in the Gut and Oral Human Microbiome. Cell Host Microbe. 2019, 26(2), 283. [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef]
- Lal, S.; Kirkup, A.J.; Brunsden, A.M.; Thompson, D.G.; Grundy, D.; Park, S.J.; Yu, Y.; Wagner, B.; Valinsky, W.C.; Lomax, A.E.; et al. Vagal afferent responses to fatty acids of different chain length in the rat. Am. J. Physiol. Liver Physiol. 2001, 281, G907–G915. [Google Scholar] [CrossRef]
- Onyszkiewicz, M.; Gawrys-Kopczynska.; M, Konopelski, P.; Aleksandrowicz, M.; Sawicka, A.; Koźniewska, E.; Samborowska, E.; Ufnal, M. Butyric acid, a gut bacteria metabolite, lowers arterial blood pressure via colon-vagus nerve signaling and GPR41/43 receptors. Pflugers Arch. 2019, 471(11-12), 1441. [CrossRef]
- Tabata, T.; Yamashita, T.; Hosomi, K.; Park, J.; Hayashi, T.; Yoshida, N.; Saito, Y.; Fukuzawa, K.; Konishi, K.; Murakami, H.; et al. Gut microbial composition in patients with atrial fibrillation: effects of diet and drugs. Hear. Vessel. 2020, 36, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Celikyurt, I.; Meier, C.R.; Kühne, M.; Schaer, B. Safety and Interactions of Direct Oral Anticoagulants with Antiarrhythmic Drugs. Drug Saf. 2017, 40, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Aliabadi, T.; Saberi, E.A.; Tabatabaei, A.M.; Tahmasebi, E. Antibiotic use in endodontic treatment during pregnancy: A narrative review. Eur. J. Transl. Myol. 2022, 32. [Google Scholar] [CrossRef]
- Valdivielso, JM.; Balafa, O.; Ekart, R.; Ferro, CJ.; Mallamaci, F.; Mark, PB.; . Rossignol, P.; Sarafidis, P.; Del Vecchio, L. Hyperkalemia in Chronic Kidney Disease in the New Era of Kidney Protection Therapies. Ortiz A.Drugs. 2021, 81(13), 1467. [Google Scholar] [CrossRef] [PubMed]
- Fardi, F.; Khasraghi, L.B.; Shahbakhti, N.; Naseriyan, A.S.; Najafi, S.; Sanaaee, S.; Alipourfard, I.; Zamany, M.; Karamipour, S.; Jahani, M.; et al. An interplay between non-coding RNAs and gut microbiota in human health. Diabetes Res. Clin. Pr. 2023, 201, 110739. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Wang, X.; Wang, L.; Sun, X.; Tan, G.; Wei, W.; Zheng, G.; Ma, X.; Tian, D.; Yu, H. Genetics, Epigenetics, Cellular Immunology, and Gut Microbiota: Emerging Links With Graves' Disease. Front Cell Dev Biol. 2022, 9, 794912. [Google Scholar] [CrossRef]
- Wang, X.; Fan, H.; Wang, Y.; Yin, X.; Liu, G.; Gao, C.; Li, X.; Liang, B. Elevated Peripheral T Helper Cells Are Associated With Atrial Fibrillation in Patients With Rheumatoid Arthritis. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- He, Y.; Chen, X.; Guo, X.; Yin, H.; Ma, N.; Tang, M.; Liu, H.; Mei, J. Th17/Treg Ratio in Serum Predicts Onset of Postoperative Atrial Fibrillation After Off-Pump Coronary Artery Bypass Graft Surgery. Heart Lung Circ. 2018, 27(12), 1467. [Google Scholar] [CrossRef]
- Jia, N.; Lin, X.; Ma, S.; Ge, S.; Mu, S.; Yang, C.; Shi, S.; Gao, L.; Xu, J.; Bo, T.; et al. Amelioration of hepatic steatosis is associated with modulation of gut microbiota and suppression of hepatic miR-34a in Gynostemma pentaphylla (Thunb.) Makino treated mice. Nutr. Metab. 2018, 15, 86. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Lv, M.; Hu, Q.; Guo, L.; Xiong, D. Correlation between alterations of gut microbiota and miR-122-5p expression in patients with type 2 diabetes mellitus. Ann. Transl. Med. 2020, 8, 1481–1481. [Google Scholar] [CrossRef]
- Khudiakov, A.A.; Panshin, D.D.; Fomicheva, Y.V.; Knyazeva, A.A.; Simonova, K.A.; Lebedev, D.S.; Mikhaylov, E.N.; Kostareva, A.A. Different Expressions of Pericardial Fluid MicroRNAs in Patients With Arrhythmogenic Right Ventricular Cardiomyopathy and Ischemic Heart Disease Undergoing Ventricular Tachycardia Ablation. Front. Cardiovasc. Med. 2021, 8. [Google Scholar] [CrossRef]
- Kassan, A.; Ait-Aissa, K.; Kassan, M. Hypothalamic miR-204 Induces Alteration of Heart Electrophysiology and Neurogenic Hypertension by Regulating the Sympathetic Nerve Activity: Potential Role of Microbiota. Cureus 2021, 13. [Google Scholar] [CrossRef]
- Pastori, D.; Carnevale, R.; Nocella, C.; Novo, M.; Santulli, M.; Cammisotto, V.; Menichelli, D.; Pignatelli, P.; Violi, F. Gut-Derived Serum Lipopolysaccharide is Associated With Enhanced Risk of Major Adverse Cardiovascular Events in Atrial Fibrillation: Effect of Adherence to Mediterranean Diet. J. Am. Hear. Assoc. 2017, 6, e005784. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zuo, K.; Zhang, J.; Hu, C.; Wang, P.; Jiao, J.; Liu, Z.; Yin, X.; Liu, X.; Li, K.; et al. Shifts in gut microbiome and metabolome are associated with risk of recurrent atrial fibrillation. J. Cell. Mol. Med. 2020, 24, 13356–13369. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.-X.; Wang, S.; Wei, L.; Cui, Y.-Y.; Chen, Y.-H. Proanthocyanidins: Components, Pharmacokinetics and Biomedical Properties. Am. J. Chin. Med. 2020, 48, 813–869. [Google Scholar] [CrossRef]
- Desantis, V.; Potenza, M.A.; Sgarra, L.; Nacci, C.; Scaringella, A.; Cicco, S.; Solimando, A.G.; Vacca, A.; Montagnani, M. microRNAs as Biomarkers of Endothelial Dysfunction and Therapeutic Target in the Pathogenesis of Atrial Fibrillation. Int. J. Mol. Sci. 2023, 24, 5307. [Google Scholar] [CrossRef]
- Hao, H.; Dai, C.; Han, X.; Li, Y. A novel therapeutic strategy for alleviating atrial remodeling by targeting exosomal miRNAs in atrial fibrillation. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2022, 1869, 119365. [Google Scholar] [CrossRef]
- Tan, A.Y.; Zimetbaum, P. Atrial Fibrillation and Atrial Fibrosis. J. Cardiovasc. Pharmacol. 2011, 57, 625–629. [Google Scholar] [CrossRef]
- Lee, Y.; Choe, J.; Park, O.H.; Kim, Y.K. Molecular Mechanisms Driving mRNA Degradation by m6A Modification. Trends Genet. 2020, 36, 177–188. [Google Scholar] [CrossRef]
- Winkler, G.S. The mammalian anti-proliferative BTG/Tob protein family. J. Cell. Physiol. 2010, 222, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Rouault, J.; Magaud, J.; Berthet, C. In search of a function for the TIS21/PC3/BTG1/TOB family. FEBS Lett. 2001, 497, 67–72. [Google Scholar] [CrossRef]
- Tirone, F. The gene PC3TIS21/BTG2, prototype member of the PC3/BTG/TOB family: Regulator in control of cell growth, differentiation, and DNA repair? J. Cell. Physiol. 2001, 187, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Ezzeddine, N.; Chang, T.-C.; Zhu, W.; Yamashita, A.; Chen, C.-Y.A.; Zhong, Z.; Yamashita, Y.; Zheng, D.; Shyu, A.-B. Human TOB, an Antiproliferative Transcription Factor, Is a Poly(A)-Binding Protein-Dependent Positive Regulator of Cytoplasmic mRNA Deadenylation. Mol. Cell. Biol. 2007, 27, 7791–7801. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Taniguchi, K.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Reactive oxygen species may influence on the crossroads of stemness, senescence, and carcinogenesis in a cell via the roles of APRO family proteins. Explor. Med. 2021, 2, 443–454. [Google Scholar] [CrossRef]
- Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Yoshikawa, S.; Tsuji, A.; Matsuda, S. Presumed Roles of APRO Family Proteins in Cancer Invasiveness. Cancers 2022, 14, 4931. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, W. Expression of B Cell Translocation Gene 1 Protein in Colon Carcinoma and its Clinical Significance. Recent Patents Anti-Cancer Drug Discov. 2020, 15, 78–85. [Google Scholar] [CrossRef]
- Zhao, S.; Xue, H.; Hao, C.-L.; Jiang, H.-M.; Zheng, H.-C. BTG1 Overexpression Might Promote Invasion and Metastasis of Colorectal Cancer via Decreasing Adhesion and Inducing Epithelial–Mesenchymal Transition. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Sung, J.-Y.; Kim, J.-Y.; Kim, H.-S. Down-regulation of B-Cell Translocation Gene 1 by Promoter Methylation in Colorectal Carcinoma. Anticancer. Res. 2018, 38, 691–697. [Google Scholar] [CrossRef]
- Ishida, Y.; Kawakami, H.; Kitajima, H.; Nishiyama, A.; Sasai, Y.; Inoue, H.; Muguruma, K. Vulnerability of Purkinje Cells Generated from Spinocerebellar Ataxia Type 6 Patient-Derived iPSCs. Cell Rep. 2016, 17, 1482–1490. [Google Scholar] [CrossRef]
- Luan, S.-H.; Yang, Y.-Q.; Ye, M.-P.; Liu, H.; Rao, Q.-F.; Kong, J.-L.; Wu, F.-R. ASIC1a promotes hepatic stellate cell activation through the exosomal miR-301a-3p/BTG1 pathway. Int. J. Biol. Macromol. 2022, 211, 128–139. [Google Scholar] [CrossRef]
- Hwang, S.S.,; Lim, J.; Yu, Z.; Kong, P.; Sefik, E.; Xu, H.; Harman, C.C.D.; Kim, .LK.; Lee, G.R.; Li, H.B.; Flavell, R.A. mRNA destabilization by BTG1 and BTG2 maintains T cell quiescence. Science. 2020, 367, 1255-1260. [CrossRef]
- Li, B.-N.; Tang, Q.-D.; Tan, Y.-L.; Yan, L.; Sun, L.; Guo, W.-B.; Qian, M.-Y.; Chen, A.; Luo, Y.-J.; Zheng, Z.-X.; et al. Key Regulatory Differentially Expressed Genes in the Blood of Atrial Septal Defect Children Treated With Occlusion Devices. Front. Genet. 2021, 12, 790426. [Google Scholar] [CrossRef]
- Tzachanis, D.; Boussiotis, V.A. Tob, a member of the APRO family, regulates immunological quiescence and tumor suppression. Cell Cycle 2009, 8, 1019–1025. [Google Scholar] [CrossRef]
- Lee, H.S.; Kundu, J.; Kim, R.N.; Shin, Y.K. Transducer of ERBB2.1 (TOB1) as a Tumor Suppressor: A Mechanistic Perspective. Int. J. Mol. Sci. 2015, 16, 29815–29828. [Google Scholar] [CrossRef]
- Szczyrek, M.; Bitkowska, P.; Chunowski, P.; Czuchryta, P. ; Krawczyk, P, Diet, Microbiome, and Cancer Immunotherapy-A Comprehensive Review. Milanowski J.Nutrients. 2021, 13(7), 2217. [Google Scholar] [CrossRef]
- Grenda, A.; Krawczyk, P. Cancer trigger or remedy: two faces of the human microbiome. Appl. Microbiol. Biotechnol. 2021, 105, 1395–1405. [Google Scholar] [CrossRef]
- Rescigno, M. The microbiota revolution: Excitement and caution. Eur. J. Immunol. 2017, 47, 1406–1413. [Google Scholar] [CrossRef]
- Sawamura, H.; Taniguchi, K.; Ikeda, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Gut Microbiota Potentiates the Effect of Immune Checkpoint Therapy against Cancers. Recent Prog. Nutr. 2021, 2, 1–1. [Google Scholar] [CrossRef]
- Asai, T.; Yoshikawa, S.; Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Tsuji, A.; Matsuda, S. Encouraging Tactics with Genetically Modified Probiotics to Improve Immunity for the Prevention of Immune-Related Diseases including Cardio-Metabolic Disorders. Biomolecules 2022, 13, 10. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wan, X.; Tan, Y.; Qu, Y.; Shan, J.; Yang, Y.; Ma, L.; Hashimoto, K. (R)-ketamine ameliorates demyelination and facilitates remyelination in cuprizone-treated mice: A role of gut–microbiota–brain axis. Neurobiol. Dis. 2022, 165, 105635. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. A New Concept of Associations between Gut Microbiota, Immunity and Central Nervous System for the Innovative Treatment of Neurodegenerative Disorders. Metabolites 2022, 12, 1052. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. Encouraging probiotics for the prevention and treatment of immune-related adverse events in novel immunotherapies against malignant glioma. Explor. Target. Anti-tumor Ther. 2022, 3, 817–827. [Google Scholar] [CrossRef]
- Tsuji, A.; Ikeda, Y.; Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Morikawa, S.; Nakashima, M.; Asai, T.; Matsuda, S. The Tryptophan and Kynurenine Pathway Involved in the Development of Immune-Related Diseases. Int. J. Mol. Sci. 2023, 24, 5742. [Google Scholar] [CrossRef]
- Håvik, B.; Røkke, H.; Dagyte, G.; Stavrum, A.-K.; Bramham, C.; Steen, V. Synaptic activity-induced global gene expression patterns in the dentate gyrus of adult behaving rats: Induction of immunity-linked genes. Neuroscience 2007, 148, 925–936. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



