Submitted:
27 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Cancer cells exhibit aberrant extracellular matrix mechanosensing due to altered expression of mechanosensory cytoskeletal proteins. Aberrant mechanosensing of the tumor microenvironment (TME) by cancer cells has been shown to be associated with cancer development and progression. Recent studies show that altered mechanosensing changes the mechanobiological properties of cells, and, in turn, cells become susceptible to mechanical perturbations. Due to an increasing understanding of cell biomechanics and cellular machinery, several approaches have emerged to target the mechanobiological properties of cancer cells and cancer-associated cells to inhibit cancer growth and progression. In this review, we summarize the progress that happened in the development of mechano-based approaches to target cancer by interfering with the cellular mechanosensing machinery and overall TME.
Keywords:
mechanical force
; apoptosis
; tumor
; tumor microenvironment
; mechanomedicine
1. Introduction
1.1. Mechanobiology of the Tumor Microenvironment
Contemporary cancer research has confirmed that the tumor microenvironment (TME) is a key driver of tumor development and progression. On examination, TME has often displayed unique characteristics, such as dense extracellular matrix (ECM), increased stiffness, leaky vasculature, and inflammation. These features have been known to be associated with tumor growth.[1] Owing to the dense microenvironment and the fact that TME harbors other types of cells apart from cancer cells, such as fibroblasts, epithelial cells, immune cells, and stem cells, the cancer cells constantly interact with other cancer cells, cancer-associated cells, and ECM. This interaction often occurs through mechanical forces generated in the pushing and pulling of the ECM and the surrounding cells. Additionally, cancer cells experience mechanical forces due to interstitial fluid pressure and vascular flow during the event of invasion and metastasis.
The studies published mainly in the last two decades have demonstrated that these mechanical forces generated in TME contribute to the onset of cancer and its subsequent progression. These forces have been placed in three broad categories: compressive stress, tensile stress, and fluid shear stress (Figure 1).[2] Compressive stress is the compression caused in the dense tumor interior due to the indefinite growth of cancer cells at the primary site. Corresponding to this, reports show that a 35-142 mm Hg value of mechanical loading on human tumors induces cancer cell proliferation and invasion.[3,4,5] Tensile stress is the force generated by the push and pull of the cells on crosslinked ECM fibers. It results in an increase in tissue stiffness. For instance, the stiffness of breast cancer tissue is in the range of 4-12 kPa, while the stiffness of normal breast tissues is 0.4-2 kPa. Likewise, other tumor tissues, such as the lung, brain, bone, and liver, exhibit higher stiffness than matched normal tissues.[6] Fluid shear stress is the force exerted by interstitial fluid and blood flow on cancer cells. This mechanical force is a significant cue in cancer progression. During the invasion process, cancer cells migrate through confined tissue interstitial spaces, dense ECM fibers, and blood capillary networks associated with TME. In this case, the fluid shear forces, as well as the fluid viscosity experienced during migration, have been shown to facilitate cancer cell dissemination from the primary site.[7,8]
1.2. The Impact of TME on Cancer Cell Behavior
In the preceding section, we discussed the major types of mechanical forces and their effects on the ECM. In this section, we will focus on how changes in ECM architecture determine cancer cell behavior (Figure 2).
Cancer cells sense the ECM stiffness by the transmembrane receptor integrin that induces downstream cellular changes. Consequently, a high number of focal adhesion and membrane protrusions are formed in the cancer cell. This, in turn, leads to the emergence of large lamellipodia and the recruitment of non-muscle myosin II, which results in directional cancer cell migration.[9,10] Apart from the stiffness of the ECM, their spatial molecular arrangement also plays a pivotal role in cancer cell adhesion and proliferation. Repetitive binding domains, such as the RGD domain, are present in ECM fibers that provide a binding site to the cell.[11] Many studies have revealed that this inter-ligand spacing should be less than 70nm to form stable focal adhesion.[12]
Other characteristics of the ECM that decide the phenotypic fate of cancer cells are the presence of ligand density, degradability, and composition. The ECM with high ligand density and degradability enhances tumor growth, while that with reduced ligand density (adhesivity) and degradability promotes balanced cellular dormancy. These features play a coordinated role with ECM stiffness and induce invasiveness of cancer cells at the intermediate-stiffness substrate.[13,14] Adding to this, the size, alignment, spaces, and gaps in the collagen bundle of ECM determine cancer cell invasion. Studies have identified that ECM pore size also controls cancer cell migration. For instance, the HT-1080 fibrosarcoma, a common 3D tumor cell motility model, cannot migrate from less than 3 µm width pores.[15,16,17] When the cancer cells move out from the primary site, they migrate through confined spaces in the ECM that are 3 to 30 µm wide and up to 600 µm long.[18] Physical confinement, such as this, has been shown to regulate several biomechanical properties of cancer cells. This includes the upregulation of mechanosensitive channels (Piezo 1), suppression of focal adhesion, and disassembly of microtubules. It has also been observed that confined cancer cells are resistant to therapeutics and display cancer stem cell-like properties.[19] Additionally, physical confinement also affects the migratory behavior of cancer cells. In this regard, it has been noted that confined cancer cell migration depends on microtubule dynamics rather than cell contractility.[20,21]
In recent years, viscoelasticity and viscosity of both ECM and cancer cells have been found to affect tumor progression. For example, the latest report revealed that ECM viscoelastic properties promote cancer cell proliferation as an added function of ECM stiffness.[22] Conversely, cancer cells show more deformability and less viscoelastic properties than normal cells. This nature of cancer cells helps in its metastatic progression. In fact, different cancer cell states (epithelial and mesenchymal) express different levels of viscoelasticity. Such properties have the potential to become reliable mechanical biomarkers for cancer.[23,24] Konstantopoulos's lab and others have recently shown that the high viscosity of matrix fluid increases the invasion and metastasis of various cancer cell types using 3D microfluidic and in vivo assays. Briefly, viscosity enhances the integrin-dependent cell spreading and actin cytoskeleton re-arrangement, further enhancing cell migration and mechanosensing properties.[8,25]
2. Mechanobiological Strategies Targeting Tme
2.1. Extracellular Matrix (ECM)
As described in the earlier sections, ECM stiffness plays a crucial role in altering the mechanical properties of TME and, thus, promoting tumor growth, metastasis, drug resistance, and evading the immune system. The ECM protein expression is upregulated in tumors, and their crosslinking causes an increase in tissue stiffening. Thus, targeting ECM synthesis, ECM crosslinking, ECM mechanosensory and mechanotransducers, and other physical means of ECM degradation have been considered strategies to alleviate tumor stiffness (Table 1). Here, we have discussed some of these targets and mechanobiological strategies investigated at various levels.
2.2. Targeting ECM Protein Synthesis and Stiffening
TGF-β is an important target to curb ECM stiffening due to the versatility of its function. TGF-β signaling activates collagen synthesis (a significant contributor to the ECM stiffening), heat shock protein 47 (a collagen chaperon that promotes collagen folding and organization), and lysyl oxidase enzyme LOX (causes collagen-elastin crosslinking). In addition, TGF-β promotes T-reg cell differentiation to induce immunosuppression in TME.[77] LH2 and FKBP10 also contribute to ECM crosslinking. TGF-β acts as proapoptotic in the initial stages of cancer, while in the later stages, it shows oncogenic function.[78] TGF-βR is downregulated in many cancers[79,80]; thus, the proapoptotic pathways are not activated. Hence targeting TGF-β and its receptor should be wisely done depending on the cancer stage and considering its diverse roles. A clinical trial (NCT01401062) to study the efficiency and feasibility of a monoclonal antibody Fresolimumab (GC1008) to block TGF-β has shown a longer median overall survival at higher doses with an increased level of tumor-specific CD8 T cells.[26] Another monoclonal antibody (NCT02581787)[81] targeting TGF-β in non-small cell lung cancer patients is currently under clinical trials. Halofuginone (anticoccidial drug) inhibits TGF-β signaling and decreases collagen synthesis in cancer animal models.[28,82] LOX inhibitors like Pirfenidone and ND-L02-s0201 injection (a lipid nanoparticle containing siRNA against Hsp47) demonstrated antifibrotic activity in lung and hepatic fibrosis, respectively. PXS-5505, a pan-LOX inhibitor, was found to be safe in clinical trials.[37]
Along with collagen, fibronectin is also upregulated in many types of cancer and contributes to increased stiffness. Extra domain A and additional domain B (EDB) of fibronectin were found to be upregulated in tumors.[83,84,85] Hence, EDB has been targeted in several strategies. A clinical trial of huBC-1-mIL-12 (a murine monoclonal antibody against the cryptic domain adjacent to human fibronectin EDB, BC-1 was fused with murine IL12) showed ~46% of patients with the stable condition after at least six cycles of treatments.[34] Another antibody targeting EDB, L19, was fused with IL-2 (L19-IL-2) and significantly improved the tumor inhibitory efficiency of IL-2 in patients showing stable condition without treatment-related death during its clinical trial in renal cell carcinoma and melanoma patients.[86]
2.3. Physical Disruption of ECM
Increased production of ECM proteins promotes chemoresistance by inhibiting drug molecules' entry inside TME. Several strategies have been exploited to disrupt collagen, enhancing drug penetration into the TME. Apart from matrix metalloproteases (MMPs) and collagenases enzyme, pulsed high-intensity focused ultrasound (HIFU) is a promising strategy to degrade ECM by ablation. An animal model study with A549 tumor tissues has shown ECM structural remodeling upon treatment with low power of Pulse-HIFU (20 W/cm2), leading to decreased collagen fibers, increased blood flow, and enhanced penetration of nanoparticles without acute tissue damage.[40]
2.4. Targeting Mechanosensory and Mechanotransducers of ECM Stiffness
In the past, efforts have been made to target mechanosensory proteins, such as integrins, mechanosensitive channels (Piezo and TRPV4), and Rho-ROCK pathways (Figure 3). Integrin is an important receptor that mediates tumor progression effects of ECM stiffness. Although targeting integrin seemed to be promising in ECM stiffness-related tumor-promoting pathways, and despite the positive outcomes in preclinical trials with integrin inhibitor, Cilengitide, several other antibodies, such as ATN-161, anti-αVβ3 antibody (etaracizumab), anti-α5β1 antibody (volociximab), anti-αV antibodies (intetumumab and abituzumab) failed in the clinal trials. [37,87]
Recent studies show that mechanosensitive channels Piezo1 acts as a mechanosensor and contribute to ECM stiffness. Thus, inhibitors of Piezo1 and other cationic mechanosensitive ion channels need further studies to understand their inhibitory effect on tumor progression.[49]
TRPV4, another mechanosensitive ion channel, contributes to tumor progression. Several antagonists have been developed in recent years[88,89], of which GSK2798745 is the first TRPV4 blocker in clinical trial studies and was tolerated by patients in the early phases.[50] YAP/TAZ are transcription coactivators activated by mechanical stress and other types of signals, which act as signal transducers to the nucleus. The constitutive activation of YAP/TAZ was responsible for the uncontrolled growth of tumors. Cancer progression gets promoted by YAP/TAZ in multiple ways through malignancy, metastasis, and chemoresistance. Although YAP/TAZ are undruggable molecules[90], RNA interference methods reduce their expression. ION537, an anti-YAP DNA antisense oligonucleotide, inhibits YAP expression and suppresses the growth of tumor xenografts.[91] Verteporfin restrains YAP/TAZ binding to TEADs which leads reduction in YAP/TAZ-induced transcription and activity[92], although its cytotoxic effects may be YAP independent[93,94], and both of these are in phase I clinical trials (NCT04659096, NCT04665206).
3. Cancer Cells
Cancer cells vary in their biomechanical properties compared to normal cells, such as stiffness, contractility, and mechanosensing, where the nucleus contributes to higher degrees of alteration in these properties as reviewed by Liu S et al.[95] Such mismatched mechanical properties are exploited in several mechanotherapy strategies to kill cancer cells selectively. Different modes of ultrasound have been used to target cancer cells (Figure 3). Due to the added advantage of its non-invasiveness and ability to penetrate deep tissues, ultrasound has been widely explored for developing cancer mechanotherapy. High-frequency Low-Intensity Pulsed Ultrasound (LIPUS) has been found to selectively ablate melanoma and breast cancer cells by triggering the intracellular explosion of nanobubbles with negligible damage to normal cells.[58] Low-frequency LIPUS has been found to produce nonthermal mechanical effects[96,97], such as stretch, compression, and shear stress[94,95] through cavitation and acoustic streaming.[96] In vitro studies using low-frequency LIPUS (33 kHz, ISPTA=7.7 mW/cm2) on breast cancer cells, melanoma cells, and breast epithelial cells found a significant increase in the death rate of cancer cells (52 %) compared to the normal cells 18 % when applying ultrasound.[67] Mouse cervical cancer models have shown an increase in survival rate to 52% from 16% in control upon therapeutic ultrasound irradiation and folate-conjugated nanobubbles.[59] On the contrary, the High-Intensity Pulsed Ultrasound (HIPU) showed nonselective thermal ablation of cancer and normal cells.
Shock wave therapy has also shown potential by killing 30% - 50% of bladder and prostate cancer cells and 10% of normal cells.[60] Other studies found smaller deformation and more damage to cancer cells than normal cell types after the shock wave treatment.[100] Surprisingly, similar shock wave treatment caused over 90% reduction of the local melanoma tumors in Hamsters.[62]
Recent studies found that mechanical stretching caused the selective apoptosis of many cancer cell types under cyclic stretch (5% strain and 0.5 Hz amplitude). The molecular mechanistic studies showed that mechanical stretching activates Piezo1 channels, causing an influx of calcium ions and subsequently activating the calpain-dependent apoptotic pathway.[63] The authors termed such mechanical force-induced cancer cell apoptosis as mechanoptosis.
4. Immune Cells
Although recent developments in cancer immunotherapy have yielded promising results, many solid tumors have failed to respond effectively. Owing to low immunogenicity, lack of universal tumor-specific antigens, poor infiltration of immune cells, and immunosuppressive effects of TME.[101,102,103,104] Investigating the underlying mechanical characteristics of cancer cells, TME, and immune cells that can be used as a treatment strategy is essential, given the growing understanding of the significance of mechanical forces in relation to cancer and immunological response. Thus, conditioning immune cells using mechanical forces is covered in this section.
4.1. T Cells
The ability of T-cells to recognize and destroy cancer cells has been extensively used in cancer immunotherapy. T-cell dysfunction brought on by tumor-induced immunosuppression, however, limits the immunological response in the TME. The mechanobiology of T cells, which is becoming better understood, strongly suggests that these cells can detect and respond to mechanical stimuli that modify their behavior.[105] As a result, directing mechanical forces to modulate T-cell function may represent a new immunotherapy approach. [105]
T-cell mechanics plays a critical role during cognate peptide presentation by APCs to the TCR, immunological synapse formation, and downstream transmission of the mechanical signals.[106,107,108] The T-cell generates force through the T-cell receptor (TCR) via cytoskeletal rearrangement, modulating T-cell activation. Interestingly, providing external cyclical forces to dysfunctional T-cells (that cannot generate their own forces) was able to re-activate these T cells.[106,108]During immunological synapse formation, the mechanosensor channels Piezo1 sense a vertical mechanical force in the form of cell membrane stretch. The downstream signaling involves calpain activation and actin assembly organization, thereby generating optimal T-cell activation. It has been shown that Piezo1-deficient T-cells failed to achieve optimal TCR activation.[109] These studies provide insight into mechanical forces and mechanosensor as an immune checkpoint for cancer immunotherapy.
T-cell migration requires mechanosensitive cytoskeletal elements, including actin filaments and microtubules, is another T-cell feature essential for immune response. Microtubule stability controls the transition between the amoeboid and mesenchymal modes of T-cell migration. According to studies, unstable microtubules cause contractility by the Rho-dependent pathway, which improves T-cell migration on an ICAM1-nanotextured surface.[110] On the other hand, T-cell migration ex vivo in 3D collagen matrices is inhibited by microtubule-stabilizing drugs. To mechanically optimize T-cells for efficient navigation through the TME, mechanosensitive cytoskeletal components offer a unique platform.
Hydrogels that resemble the body's ECM are another intriguing technique for T-cell-based immunotherapy. Hickey et al. (2019) investigated the role of an artificial T-cell stimulating matrix (aTM). The studies showed that adjusting aTM's stiffness can modulate T cell activation. These studies suggested promising results to enhance the effectiveness of T-cell treatment in vitro and extend to ex vivo T-cell stimulation that can be injected into mice for effective tumor suppression.[111] Recently, nanoparticles coated with engineered T-cell membranes (with specific receptors against targeted cells) have been employed in a murine model to eliminate glioblastoma and glioblastoma stem cells using photothermal therapy. The surface membrane on the NPs helped in penetrating through the blood-brain barrier.[112] Other studies have demonstrated optogenetic activation of bulk immune cells, including T-cells and dendritic cells, using wide-field illumination and calcium actuators. [113,114,115]
The spatiotemporal control of T-cells in the tumor region is crucial for effective cancer therapy. Recently, mechano-genetically engineered CAR-T systems have been mechanically sensitized to guide and control CAR-T cell activities spatiotemporally. In another study, ultrasound, used for external sources for activating CAR-T cells, has the additional advantage of guiding engineered T cells into deep tissues. Thermal induction of CAR-T cells by Focused Ultrasound showed a significant reduction in tumor growth rate in mice with prostate cancer.[116] The non-thermal induction of CAR-T cells using ultrasound waves is through the mechanosensitive PIEZO1 channel.[117,118] The mechanical signal then transduces via calcineurin-mediated NFAT (nuclear factor of activated T cells) transcription factor, which subsequently transcribes anti-CD19 CAR genes in T-cells.[119] It has been envisioned that these mechano-genetic CAR-T systems will be the future generation tool for cancer treatment which can be controlled by wireless devices.[119] Although ultrasound is a promising option in CAR-T therapy, further research and technological advancements are needed for clinical use. Overall, an in-depth understanding of T-cells' mechanobiology and mechanosensitive machinery can give insight into novel strategies for cancer treatment by overcoming the present limitation in the TME.
4.2. Dendritic Cells (DCs)
The majority of research on cancer immunobiology focuses on T-cells’ actin and actin-binding proteins, although cytoskeletal remodeling in APCs (DCs and macrophages) also plays a role in antigen presentation and immunological synapse formation, which are crucial for effective immune response.[120,121,122,123] In bone marrow-derived macrophages, Jonsson et al. (2012) demonstrated that the actin-binding protein n-cofilin (F-actin depolymerizing factor) regulates various immune response-related events. In addition to mediating macrophage motility and cell polarity, n-cofilin-mediated F-actin remodeling is essential for T-cell antigen presentation. [123]
DCs, which are potent antigen-presenting cells, greatly aid primary T-cell activation. On DCs, a structure known as a podosome rich in actin senses the substrate rigidity. It has been demonstrated that podosomes apply mechanical stresses and locally break down the ECM at substrate-soft areas (low physical resistance). When the pore size is >1µm, podosomes develop into a protruding structure where pathogen recognition receptors (PRR) occupy the region and mediate active antigen uptake followed by antigen presentation to T-cells.[124] An effective immune system depends on the navigation of these cells across endothelium and ECM cells. A greater infiltration of DCs and other immune cells at the TME may therefore be facilitated by reducing the ECM stiffness.
Activated DCs releases effector molecules that kill cancer cells, but in the TME, immunosuppressive cytokines co-opt dendritic cells to withstand cancer. Therefore, promoting dendritic cell (DC) activation by utilizing mechanical forces is one promising strategy. It has been observed that after HIFU therapy on breast cancer, a significant amount of tumor debris and heat shock proteins accumulate in situ. This debris not only induces local infiltration of activated DCs, in addition, triggers more significant lymphocyte proliferation.[125,126,127] HIFU therapy has been clinically to treat cancer.[128] However, HIFU has been linked to tissue necrosis near the tumor due to heat dissipation.[129] Therefore, research into low-frequency ultrasound is being investigated. In a study focusing on angiogenesis and DCs-related immunotherapy, low-frequency ultrasound combined with microbubbles in murine prostate cancer cells reduced VEGF expression and significant DCs differentiation, which then activated T-cell mediated immune response in vitro.[130] Thus, DCs are potent targets for cancer immunotherapy. However, further studies are necessary to comprehensively understand mechanical force sensing, induction, and transmission in DCs.
4.3. Tumor-Associated Macrophages (TAMs)
A wide range of mechanical signals has been demonstrated to control macrophage polarization and determine their anti-tumor (M1) or pro-tumor (M2) nature. For instance, reduced actin polymerization brought on by spatial confinement and decreased substrate stiffness cause macrophages to change from the M1 to the M2 phenotype.[131,132,133] YAP detects the substrate stiffness and causes the macrophages in the TME to exhibit an anti-tumoricidal M2 phenotype.[134,135] Thus, changing the phenotype of M2 macrophages to M1 is beneficial, as these cells are responsible for building an immunosuppressive environment for the cancer cells. Inhibitors of class IIa histone deacetylase (HDAC) in breast cancer patients [136,137] and both IL-12 and IL-37 in hepatocellular carcinoma patients were able to reverse the phenotype of TAMs in TME. The FAK/NF-κB signaling pathway, as demonstrated by Shan et al. (2019), facilitates the M1 polarization of macrophages upon mechanical stretch.[138,139] TAMs have been shown to be more effectively reprogrammed from the M2 to the M1 (anti-tumor) phenotype in vitro when ultrasound-targeted nanobubble destruction and nanobubbles containing drugs (Low Molecular Weight Hyaluronic Acid) are combined. The potential cause of the enhanced reeducation effect on UTND may be attributed to oxidative stress.[140] In addition, studies showed that TAMs also interfere with immune checkpoint inhibitors; as a result, therapeutic targeting of TAMs may be more advantageous for immunotherapy. Their mechanobiological aspects need further focus in this regard.
4.4. Cancer-Associated Fibroblasts (CAFs)
CAFs play a crucial role in cancer development by directly exerting mechanical forces. These forces are transferred by a cadherin-composed adhesion structure that bridges CAF with the cell membrane of cancer cells.[141] CAFs exert a compressive force on the cancer cells that activate YAP-mediated cancer cell proliferation.[142] CAFs also play a significant role in drug resistance in several cancer types through mechanisms including immunological regulation, cancer metabolism, desmoplasia, and angiogenesis.[143,144,145,146,147] It has been well documented that the increased ECM stiffness brought on by CAFs presents a considerable barrier to immune cell migration.[148] Therefore, researchers are focusing on preventing CAF-induced ECM-stiffening events.
Zhang et al. (2019) demonstrated the feed-forward cycle between stiff ECM and CAFs. During the onstage of cancer, the ECM stiffness is maintained by increasing the activity of the mechanical responsive transcriptional regulator SNAIL1 in the CAFs. Depletion of SNAIL1 resulted in altered CAFs function, including tumor fibrosis.[149] Moreover, the mechanics of CAFs in blood vessel growth in the TME has recently been explored. CAF generates mechanical forces that induce large deformations in the ECM, subsequently causing blood vessel growth in the TME.[150] Sewell-Loftin et al. (2017) also showed the involvement of Rho-ROCK and YAP signaling in the CAF-mediated vascularization, providing insights into targeting these signaling pathways for anti-cancer therapeutics.[150] These studies suggest that targeting the mechanosensitive factors and pathways in CAFs could result in better therapeutic outcomes. In vitro studies of prostrate CAFs showed that the treatment of patient-derived CAFs to Extracorporeal Shock Waves (ESWs), a form of acoustic waves, reduced gene expression and protein level of mesenchymal markers of CAFs α-smooth muscle actin and type I collagen. Lower levels of these markers consequently reduced the growth of prostate cancer epithelial cells.[151] Thus, targeting CAF may improve treatment outcomes given the prevalence of CAFs and their pro-tumorigenic role.
CONCLUSION AND OUTLOOK
It has become increasingly clear that biophysical properties of TME, including ECM characteristics, play a dominant role in cancer onstage and progression. For example, increased ECM stiffness is associated with cancer progression and is widely used for breast cancer detection. Mechanical forces generated in TME due to cell-cell or cell-ECM interaction, mainly compressive, tensile and shear forces, also contribute to cancer progression. Thus, targeting stiffness and biophysical forces associated with TME by targeting ECM has been considered in several preclinical/clinical studies. In recent years, several mechanical force-based strategies have emerged that target the mechanosensory machinery of the cancer cells and cancer-associated cells to induce selective killing (mechanoptosis). Although these mechano-based treatments look promising due to their selectivity in killing cancer cells over normal cells, the comprehensive knowledge of the molecular mechanism of the killing is not yet fully understood. Further studies in this direction will help to harness the mechanical forces to develop a mechano-based treatment for personalized use.
Author Contributions
Alka, Manasa, and Rashmita contributed equally.
Acknowledgment
We acknowledge the Science Engineering Research Board (SERB), India, for the funding.
Conflicts of Interest
The authors declare no competing financial interest.
Biographies
Alka Kumari is a graduate student in the Centre for Biosystems Science and Engineering at the Indian Institute of Science, Bangalore. She works at the centre in Dr. Ajay Tijore’s ‘Mechanobiologics lab.’ Her research focuses on developing PDMS elastomer-based microfluidic platforms to study cancer cell growth under mechanical force treatment.
S Manasa Veena is a graduate student in the Centre for Biosystems Science and Engineering at the Indian Institute of Science, Bangalore. She works at the centre in Dr. Ajay Tijore’s ‘Mechanobiologics lab.’ Her research focuses on developing nanopatterned platforms to study cancer cell growth under mechanical force treatment.
Rashmita Luha is a graduate student in the Centre for Biosystems Science and Engineering at the Indian Institute of Science, Bangalore. She works at the centre in Dr. Ajay Tijore’s ‘Mechanobiologics lab.’ Her research focuses on studying the effect of ultrasound-mediated mechanical forces on patient-derived oral cancer cell survival.
Ajay Tijore received a Ph.D. in stem cell bioengineering from Nanyang Technological University, Singapore, in 2016. He then joined the Mechanobiology Institute National University of Singapore as a postdoctoral research fellow to study the work in the field of cancer mechanobiology. In Nov 2021, he joined the Centre for Biosystems Science and Engineering (BSSE), where his lab works on investigating the effect of mechanical forces on cancer cell growth and in regulating stem cell fate using custom-built microfluidic devices and micro/nanoscale biomaterials.
References
- Shi, X.; Zhang, L.; Li, B.; Feng, X. The Mechanical Problems in Tumor and Tumor Microenvironment. Advances in Mechanics 2018. [CrossRef]
- Liu, Q.; Luo, Q.; Ju, Y.; Song, G. Role of the Mechanical Microenvironment in Cancer Development and Progression. Cancer Biology and Medicine 2020. [CrossRef] [PubMed]
- Stylianopoulos, T.; Martin, J. D.; Chauhan, V. P.; Jain, S. R.; Diop-Frimpong, B.; Bardeesy, N.; Smith, B. L.; Ferrone, C. R.; Hornicek, F. J.; Boucher, Y.; Munn, L. L.; Jain, R. K. Causes, Consequences, and Remedies for Growth-Induced Solid Stress in Murine and Human Tumors. Proc Natl Acad Sci U S A 2012, 109. [Google Scholar] [CrossRef]
- Luo, M.; K. Y. Ho, K.; Tong, Z.; Deng, L.; P. Liu, A. Compressive Stress Enhances Invasive Phenotype of Cancer Cells via Piezo1 Activation. bioRxiv 2019. [Google Scholar] [CrossRef]
- Stylianopoulos, T. The Solid Mechanics of Cancer and Strategies for Improved Therapy. J Biomech Eng 2017, 139. [Google Scholar] [CrossRef]
- Yu, H.; Mouw, J. K.; Weaver, V. M. Forcing Form and Function: Biomechanical Regulation of Tumor Evolution. Trends in Cell Biology 2011. [CrossRef]
- Northcott, J. M.; Dean, I. S.; Mouw, J. K.; Weaver, V. M. Feeling Stress: The Mechanics of Cancer Progression and Aggression. Frontiers in Cell and Developmental Biology 2018. [CrossRef]
- Bera, K.; Kiepas, A.; Godet, I.; Li, Y.; Mehta, P.; Ifemembi, B.; Paul, C. D.; Sen, A.; Serra, S. A.; Stoletov, K.; Tao, J.; Shatkin, G.; Lee, S. J.; Zhang, Y.; Boen, A.; Mistriotis, P.; Gilkes, D. M.; Lewis, J. D.; Fan, C. M.; Feinberg, A. P.; Valverde, M. A.; Sun, S. X.; Konstantopoulos, K. Extracellular Fluid Viscosity Enhances Cell Migration and Cancer Dissemination. Nature 2022, 611. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, Z.; He, Y.; Li, P.; Chen, Y.; Chen, X.; Jiang, Y.; Qin, X.; Li, S.; Li, T.; Wu, C.; Yang, H.; You, F.; Liu, Y. Non-Muscle Myosin II Isoforms Orchestrate Substrate Stiffness Sensing to Promote Cancer Cell Contractility and Migration. Cancer Lett 2022, 524. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, Z.; Chen, Y.; Li, S.; Jiang, Y.; Yang, H.; Wu, C.; You, F.; Zheng, C.; Zhu, J.; Tan, Y.; Qin, X.; Liu, Y. ROCK Isoforms Differentially Modulate Cancer Cell Motility by Mechanosensing the Substrate Stiffness. Acta Biomater 2019, 88. [Google Scholar] [CrossRef]
- Ruoslahti, E. RGD and Other Recognition Sequences for Integrins. Annual Review of Cell and Developmental Biology 1996. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhao, C.; Spatz, J. P.; Wei, Q. Nanopatterned Adhesive, Stretchable Hydrogel to Control Ligand Spacing and Regulate Cell Spreading and Migration. ACS Nano 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Farino Reyes, C. J.; Pradhan, S.; Slater, J. H. The Influence of Ligand Density and Degradability on Hydrogel Induced Breast Cancer Dormancy and Reactivation. Adv Healthc Mater 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, O.; Koshy, S. T.; Branco Da Cunha, C.; Shin, J. W.; Verbeke, C. S.; Allison, K. H.; Mooney, D. J. Extracellular Matrix Stiffness and Composition Jointly Regulate the Induction of Malignant Phenotypes in Mammary Epithelium. Nat Mater 2014, 13. [Google Scholar] [CrossRef]
- Koorman, T.; Jansen, K. A.; Khalil, A.; Haughton, P. D.; Visser, D.; Rätze, M. A. K.; Haakma, W. E.; Sakalauskaitè, G.; van Diest, P. J.; de Rooij, J.; Derksen, P. W. B. Spatial Collagen Stiffening Promotes Collective Breast Cancer Cell Invasion by Reinforcing Extracellular Matrix Alignment. Oncogene 2022, 41. [Google Scholar] [CrossRef]
- Wolf, K.; te Lindert, M.; Krause, M.; Alexander, S.; te Riet, J.; Willis, A. L.; Hoffman, R. M.; Figdor, C. G.; Weiss, S. J.; Friedl, P. Physical Limits of Cell Migration: Control by ECM Space and Nuclear Deformation and Tuning by Proteolysis and Traction Force. Journal of Cell Biology 2013, 201. [Google Scholar] [CrossRef]
- Harada, T.; Swift, J.; Irianto, J.; Shin, J. W.; Spinler, K. R.; Athirasala, A.; Diegmiller, R.; Dingal, P. C. D. P.; Ivanovska, I. L.; Discher, D. E. Nuclear Lamin Stiffness Is a Barrier to 3D-Migration, but Softness Can Limit Survival. In Proceedings of the IEEE Annual Northeast Bioengineering Conference, NEBEC; 2014; Vol. 2014-December. [Google Scholar] [CrossRef]
- Weigelin, B.; Bakker, G.-J.; Friedl, P. Intravital Third Harmonic Generation Microscopy of Collective Melanoma Cell Invasion. Intravital 2012, 1. [Google Scholar] [CrossRef]
- Shen, Q.; Hill, T.; Cai, X.; Bui, L.; Barakat, R.; Hills, E.; Almugaiteeb, T.; Babu, A.; Mckernan, P. H.; Zalles, M.; Battiste, J. D.; Kim, Y. T. Physical Confinement during Cancer Cell Migration Triggers Therapeutic Resistance and Cancer Stem Cell-like Behavior. Cancer Lett 2021, 506, 142. [Google Scholar] [CrossRef]
- Balzer, E. M.; Tong, Z.; Paul, C. D.; Hung, W. C.; Stroka, K. M.; Boggs, A. E.; Martin, S. S.; Konstantopoulos, K. Physical Confinement Alters Tumor Cell Adhesion and Migration Phenotypes. FASEB Journal 2012, 26. [Google Scholar] [CrossRef]
- Hung, W. C.; Yang, J. R.; Yankaskas, C. L.; Wong, B. S.; Wu, P. H.; Pardo-Pastor, C.; Serra, S. A.; Chiang, M. J.; Gu, Z.; Wirtz, D.; Valverde, M. A.; Yang, J. T.; Zhang, J.; Konstantopoulos, K. Confinement Sensing and Signal Optimization via Piezo1/PKA and Myosin II Pathways. Cell Rep 2016, 15. [Google Scholar] [CrossRef]
- Elosegui-Artola, A.; Gupta, A.; Najibi, A. J.; Seo, B. R.; Garry, R.; Tringides, C. M.; de Lázaro, I.; Darnell, M.; Gu, W.; Zhou, Q.; Weitz, D. A.; Mahadevan, L.; Mooney, D. J. Matrix Viscoelasticity Controls Spatiotemporal Tissue Organization. Nat Mater 2023, 22. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, M.; Cheng, M.; Gao, Z.; Wang, G. The Viscoelastic Behaviors of Several Kinds of Cancer Cells and Normal Cells. J Mech Behav Biomed Mater 2019, 91. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, B.; Meng, S.; Li, S.; Yang, X. Electrical and Viscoelastic Measurement of Cancer Cells in Epithelial-Mesenchymal Transition Process on a Microfluidic Device. In 2019 8th International Symposium on Next Generation Electronics, ISNE 2019; 2019. [CrossRef]
- Gonzalez-Molina, J.; Zhang, X.; Borghesan, M.; Mendonça da Silva, J.; Awan, M.; Fuller, B.; Gavara, N.; Selden, C. Extracellular Fluid Viscosity Enhances Liver Cancer Cell Mechanosensing and Migration. Biomaterials 2018, 177, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Formenti, S. C.; Hawtin, R. E.; Dixit, N.; Evensen, E.; Lee, P.; Goldberg, J. D.; Li, X.; Vanpouille-Box, C.; Schaue, D.; McBride, W. H.; Demaria, S. Baseline T Cell Dysfunction by Single Cell Network Profiling in Metastatic Breast Cancer Patients. J Immunother Cancer 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Zion, O.; Genin, O.; Kawada, N.; Yoshizato, K.; Roffe, S.; Nagler, A.; Iovanna, J. L.; Halevy, O.; Pines, M. Inhibition of Transforming Growth Factor β Signaling by Halofuginone as a Modality for Pancreas Fibrosis Prevention. Pancreas 2009, 38, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Yi, S.; Gong, L.; Liu, W.; Wang, P.; Liu, N.; Zhao, L.; Wang, P. Inhibition of TGF-β Signaling with Halofuginone Can Enhance the Antitumor Effect of Irradiation in Lewis Lung Cancer. Taylor & Francis 2015, 8, 3549–3559. [Google Scholar] [CrossRef]
- Juárez, P.; Mohammad, K.; Yin, J.; research, P. F.-C. 2012, undefined. Halofuginone Inhibits the Establishment and Progression of Melanoma Bone MetastasesHalofuginone Decreases Melanoma Bone Metastases. AACR.
- Juárez, P.; Fournier, P.; Oncotarget, K. M. 2017, undefined. Halofuginone Inhibits TGF-β/BMP Signaling and in Combination with Zoledronic Acid Enhances Inhibition of Breast Cancer Bone Metastasis. ncbi.nlm.nih.gov.
- Zhang, B.; Halder, S.; Zhang, S.; letters, P. D.-C. 2009, undefined. Targeting Transforming Growth Factor-β Signaling in Liver Metastasis of Colon Cancer. Elsevier.
- Melisi, D.; Ishiyama, S.; Sclabas, G.; … J., F.-M. cancer; 2008, undefined. LY2109761, a Novel Transforming Growth Factor β Receptor Type I and Type II Dual Inhibitor, as a Therapeutic Approach to Suppressing Pancreatic Cancer Metastasis. AACR.
- Korpal, M.; Yan, J.; Lu, X.; Xu, S.; Lerit, D.; medicine, Y. K.-N. 2009, undefined. Imaging Transforming Growth Factor-β Signaling Dynamics and Therapeutic Response in Breast Cancer Bone Metastasis. nature.com.
- Rudman, S. M.; Jameson, M. B.; McKeage, M. J.; Savage, P.; Jodrell, D. I.; Harries, M.; Acton, G.; Erlandsson, F.; Spicer, J. F. A Phase 1 Study of AS1409, a Novel Antibody-Cytokine Fusion Protein, in Patients with Malignant Melanoma or Renal Cell Carcinoma. Clinical Cancer Research 2011, 17. [Google Scholar] [CrossRef]
- Hisatomi, K.; Mukae, H.; Sakamoto, N.; Ishimatsu, Y.; Kakugawa, T.; Hara, S.; Fujita, H.; Nakamichi, S.; Oku, H.; Urata, Y.; Kubota, H.; Nagata, K.; Kohno, S. Pirfenidone Inhibits TGF-Β1-Induced over-Expression of Collagen Type I and Heat Shock Protein 47 in A549 Cells. BMC Pulm Med 2012, 12. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Quimbo, A.; Xia, F.; Yao, J.; Clamme, J.-P.; Zabludoff, S.; Zhang, J.; Ying, W. Anti-HSP47 SiRNA Lipid Nanoparticle ND-L02-S0201 Reverses Interstitial Pulmonary Fibrosis in Preclinical Rat Models. Eur Respiratory Soc. [CrossRef]
- Jiang, Y.; Zhang, H.; Wang, J.; Liu, Y.; Luo, T.; Hua, H. Targeting Extracellular Matrix Stiffness and Mechanotransducers to Improve Cancer Therapy. J Hematol Oncol 2022, 15, 1–15. [Google Scholar] [CrossRef]
- Benson, A. B.; Wainberg, Z. A.; Hecht, J. R.; Vyushkov, D.; Dong, H.; Bendell, J.; Kudrik, F. A Phase II Randomized, Double-Blind, Placebo-Controlled Study of Simtuzumab or Placebo in Combination with Gemcitabine for the First-Line Treatment of Pancreatic Adenocarcinoma. The Oncologist 2017, 241-e15. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J. R.; Benson, A. B.; Vyushkov, D.; Yang, Y.; Bendell, J.; Verma, U. A Phase II, Randomized, Double-Blind, Placebo-Controlled Study of Simtuzumab in Combination with FOLFIRI for the Second-Line Treatment of Metastatic KRAS Mutant Colorectal Adenocarcinoma. The Oncologist 2017, 243-e23. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Han, H.; Koo, H.; Na, J. H.; Yoon, H. Y.; Lee, K. E.; Lee, H.; Kim, H.; Kwon, I. C.; Kim, K. Extracellular Matrix Remodeling in Vivo for Enhancing Tumor-Targeting Efficiency of Nanoparticle Drug Carriers Using the Pulsed High Intensity Focused Ultrasound. Journal of Controlled Release 2017, 263, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Fink, K.; Scott, B.; Health, W.; Ravin, P.; Raizer, J. J. Randomized Phase II Study of Cilengitide, an Integrin-Targeting Arginine-Glycine-Aspartic Acid Peptide, in Recurrent Glioblastoma Multiforme. researchgate.net 2008. [Google Scholar] [CrossRef]
- Cianfrocca, M. E.; Kimmel, K. A.; Gallo, J.; Cardoso, T.; Brown, M. M.; Hudes, G.; Lewis, N.; Weiner, L.; Lam, G. N.; Brown, S. C.; Shaw, D. E.; Mazar, A. P.; Cohen, R. B. Phase 1 Trial of the Antiangiogenic Peptide ATN-161 (Ac-PHSCN-NH2), a Beta Integrin Antagonist, in Patients with Solid Tumours. British Journal of Cancer 2006, 94, 1621–1626. [Google Scholar] [CrossRef]
- Delbaldo, C.; Raymond, E.; Vera, K.; Hammershaimb, L.; Kaucic, K.; Lozahic, S.; Marty, M.; Faivre, S. Phase I and Pharmacokinetic Study of Etaracizumab (AbegrinTM), a Humanized Monoclonal Antibody against Avβ3 Integrin Receptor, in Patients with Advanced Solid Tumors. Invest New Drugs 2008, 26, 35–43. [Google Scholar] [CrossRef]
- Hersey, P.; Sosman, J.; Steven O’day, ; Richards, J.; Bedikian, A.; Gonzalez, R.; Sharfman, W.; Weber, R.; Logan, T.; Buzoianu, M.; Hammershaimb, ; Luz; Kirkwood, J. M. A Randomized Phase 2 Study of Etaracizumab, a Monoclonal Antibody against Integrin Avβ3, ± Dacarbazine in Patients with Stage IV Metastatic Melanoma. Wiley Online Library 2010, 116, 1526–1534. [CrossRef]
- O’day, S.; Pavlick, A.; Loquai, C.; … D., L.-B. journal of; 2011, undefined. A Randomised, Phase II Study of Intetumumab, an Anti-Av-Integrin MAb, Alone and with Dacarbazine in Stage IV Melanoma. nature.com.
- Heidenreich, A.; Rawal, S.; Szkarlat, K.; oncology, N. B.-A. 2013, undefined. A Randomized, Double-Blind, Multicenter, Phase 2 Study of a Human Monoclonal Antibody to Human Aν Integrins (Intetumumab) in Combination with Docetaxel. Elsevier.
- Élez, E.; Kocáková, I.; Höhler, T.; Martens, U. M.; Bokemeyer, C.; Van cutsem, E.; Melichar, B.; Smakal, M.; Csoszi, T.; Topuzov, E.; Orlova, R.; Tjulandin, S.; Rivera, F.; Straub, J.; Bruns, R.; Quaratino, S.; Tabernero, J. Abituzumab Combined with Cetuximab plus Irinotecan versus Cetuximab plus Irinotecan Alone for Patients with KRAS Wild-Type Metastatic Colorectal Cancer: The Randomised Phase I/II POSEIDON Trial. Ann Oncol 2015, 26, 132–140. [Google Scholar] [CrossRef]
- Hussain, M.; Le Moulec, S.; Gimmi, C.; Bruns, R.; Straub, J.; Miller, K. Differential Effect on Bone Lesions of Targeting Integrins: Randomized Phase II Trial of Abituzumab in Patients with Metastatic Castration-Resistant Prostate Cancer. Clinical Cancer Research 2016, 22, 3192–3200. [Google Scholar] [CrossRef]
- Bae, C.; Sachs, F.; Gottlieb, P. A. The Mechanosensitive Ion Channel Piezo1 Is Inhibited by the Peptide GsMTx4. Biochemistry 2011, 50. [Google Scholar] [CrossRef]
- Dayaram, V.; Malloy, C.; Martha, S.; Alvarez, B.; Chukwudolue, I.; Dabbain, N.; Dlovan, D.; Goleva, S.; Hickey, T.; Ho, A.; Kington, P.; Mattingly, M.; Potter, S.; Simpson, L.; Uradu, H.; Doorn, J. Van; Cooper, R. L. Stretch Activated Channels in Proprioceptive Organs of Crab and Crayfish Are Sensitive to Gadolinium but Not Amiloride, Ruthenium Red or Low PH. Impulse (Sydney) 2017, 1–17. [Google Scholar]
- Coste, B.; Xiao, B.; Santos, J. S.; Syeda, R.; Grandl, J.; Spencer, K. S.; Kim, S. E.; Schmidt, M.; Mathur, J.; Dubin, A. E.; Montal, M.; Patapoutian, A. Piezo Proteins Are Pore-Forming Subunits of Mechanically Activated Channels. Nature 2012, 483, 176–181. [Google Scholar] [CrossRef]
- Goyal, N.; Skrdla, P.; Schroyer, R.; Kumar, S.; Fernando, D.; Oughton, A.; Norton, N.; Sprecher, D. L.; Cheriyan, J. Clinical Pharmacokinetics, Safety, and Tolerability of a Novel, First-in-Class TRPV4 Ion Channel Inhibitor, GSK2798745, in Healthy and Heart Failure Subjects. American Journal of Cardiovascular Drugs 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Research, A. M.-C. 2021, undefined. Abstract ND11: The Discovery and Characterization of ION-537: A next Generation Antisense Oligonucleotide Inhibitor of YAP1 in Preclinical Cancer Models. AACR.
- Chapeau, E.; Schmelzle, T. IAG933, an Oral Selective YAP1-TAZ/Pan-TEAD Protein-Protein Interaction Inhibitor (PPIi) with Pre-Clinical Activity in Monotherapy and Combinations with MAPK Inhibitors. 2023. [Google Scholar]
- Vincent, P., Maeder, M.E., Hunt, B., Linn, B., Mangels-Dick, T., Hasan, T., Wang, K.K. and Pogue, B.W. 2021, undefined. CT Radiomic Features of Photodynamic Priming in Clinical Pancreatic Adenocarcinoma Treatment. iopscience.iop.org 2021. [CrossRef]
- Hanada, Y.; Pereira, S.; Pogue, B.; … E., M.-G. 2021, undefined. EUS-Guided Verteporfin Photodynamic Therapy for Pancreatic Cancer. Elsevier.
- Huggett, M.; Jermyn, M.; Gillams, A.; … R., I.-B. journal of; 2014, undefined. Phase I/II Study of Verteporfin Photodynamic Therapy in Locally Advanced Pancreatic Cancer. nature.com.
- Lejbkowicz, F.; Zwiran, M.; biology, S. S.-U. in medicine &; 1993, undefined. The Response of Normal and Malignant Cells to Ultrasound in Vitro. Elsevier.
- Shen, S.; Li, Y.; Xiao, Y.; Zhao, Z.; Zhang, C.; Wang, J.; Biomaterials, H. L.-. 2018, undefined. Folate-Conjugated Nanobubbles Selectively Target and Kill Cancer Cells via Ultrasound-Triggered Intracellular Explosion. Elsevier.
- Nicolai, H.; Steinbach, P.; Knuechel-Clarke, R.; Grimm, D.; Roessler, W.; Wieland, W. F.; Hofstaedter, F. Proliferation of Tumor Spheroids after Shock-Wave Treatment. J Cancer Res Clin Oncol 1994, 120, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Pellegrino, A.; Hallack, A.; Petrinic, N.; Jérusalem, A.; Cleveland, R. O. Response of Single Cells to Shock Waves and Numerically Optimized Waveforms for Cancer Therapy. Biophys J 2018, 114, 1433–1439. [Google Scholar] [CrossRef]
- Gamarra, F.; Spelsberg, F.; Dellian, M.; Goetz, A. E. Complete Local Tumor Remission after Therapy with Extra-corporeally Applied High-energy Shock Waves (HESW). Int J Cancer 1993, 55, 153–156. [Google Scholar] [CrossRef]
- Tijore, A.; Margadant, F.; Yao, M.; Hariharan, A.; Chew, C. A. Z.; Powell, S.; Bonney, G. K.; Sheetz, M. Ultrasound-Mediated Mechanical Forces Selectively Kill Tumor Cells. bioRxiv 2020. [Google Scholar]
- Berrueta, L.; Bergholz, J.; Munoz, D.; Muskaj, I.; Badger, G. J.; Shukla, A.; Kim, H. J.; Zhao, J. J.; Langevin, H. M. Stretching Reduces Tumor Growth in a Mouse Breast Cancer Model. Sci Rep 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Azagury, A.; Amar-Lewis, E.; Yudilevitch, Y.; Isaacson, C.; Laster, B.; Kost, J. Ultrasound Effect on Cancerous versus Non-Cancerous Cells. Ultrasound Med Biol 2016, 42, 1560–1567. [Google Scholar] [CrossRef]
- Lejbkowicz, F. Sensitivity of Normal. 1997, 105 (December), 1575–1578.
- Singh, A.; Tijore, A.; Margadant, F.; Simpson, C.; Chitkara, D.; Low, B. C.; Sheetz, M. Enhanced Tumor Cell Killing by Ultrasound after Microtubule Depolymerization. Bioeng Transl Med 2021, 6, e10233. [Google Scholar] [CrossRef]
- Los, U. M. D. E. C. D. E. No 主観的健康感を中心とした在宅高齢者における 健康関連指標に関する共分散構造分析Title.
- Sultan, L. R.; Karmacharya, M. B.; Hunt, S. J.; Wood, A. K. W.; Sehgal, C. M. Subsequent Ultrasound Vascular Targeting Therapy of Hepatocellular Carcinoma Improves the Treatment Efficacy. mdpi.com 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Y.; Li, R.; Jia, W.; Zhang, F.; Hu, X.; Reports, L. C.-S. 2022, undefined. High Speed Photo-Mediated Ultrasound Therapy Integrated with OCTA. nature.com.
- Xu, Z. L.; Zhu, X. Q.; Lu, P.; Zhou, Q.; Zhang, J.; Wu, F. Activation of Tumor-Infiltrating Antigen Presenting Cells by High Intensity Focused Ultrasound Ablation of Human Breast Cancer. Ultrasound Med Biol 2009, 35, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, L.; Chen, W. R. Host Antitumor Immune Responses to HIFU Ablation. International Journal of Hyperthermia 2007, 23, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhang, Y.; Feng, J.; Wu, F. Dendritic Cells Loaded with Ultrasound-Ablated Tumour Induce in Vivo Specific Antitumour Immune Responses. Ultrasound Med Biol 2010, 36, 441–448. [Google Scholar] [CrossRef]
- Zhang, W.; Shou, W. De; Xu, Y. J.; Bai, W. K.; Hu, B. Low-Frequency Ultrasound-Induced VEGF Suppression and Synergy with Dendritic Cell-Mediated Anti-Tumor Immunity in Murine Prostate Cancer Cells in Vitro. Sci Rep 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.; Shang, M.; Shi, D.; Liang, P.; Jing, X.; Meng, D.; Liu, X.; Zhou, X.; Zhao, Y.; Li, J. Ultrasound Mediated Destruction of Lmw-Ha-Loaded and Folate-Conjugated Nanobubble for Tam Targeting and Reeducation. Int J Nanomedicine 2020, 15, 1967–1981. [Google Scholar] [CrossRef]
- Rinella, L.; Pizzo, B.; Frairia, R.; Delsedime, L.; Calleris, G.; Gontero, P.; Zunino, V.; Fortunati, N.; Arvat, E.; Catalano, M. G. Modulating Tumor Reactive Stroma by Extracorporeal Shock Waves to Control Prostate Cancer Progression. Prostate 2020, 80, 1087–1096. [Google Scholar] [CrossRef]
- Wan, Y. Y.; Flavell, R. A. “Yin-Yang” Functions of Transforming Growth Factor-β and T Regulatory Cells in Immune Regulation. Immunological Reviews 2007. [CrossRef]
- Chen, Y.; Di, C.; Zhang, X.; Wang, J.; Wang, F.; Yan, J. fang; Xu, C.; Zhang, J.; Zhang, Q.; Li, H.; Yang, H.; Zhang, H. Transforming Growth Factor β Signaling Pathway: A Promising Therapeutic Target for Cancer. Journal of Cellular Physiology 2020. [CrossRef]
- Johannsen, M.; Spitaleri, G.; Curigliano, G.; Roigas, J.; Weikert, S.; Kempkensteffen, C.; Roemer, A.; Kloeters, C.; Rogalla, P.; Pecher, G.; Miller, K.; Berndt, A.; Kosmehl, H.; Trachsel, E.; Kaspar, M.; Lovato, V.; González-Iglesias, R.; Giovannoni, L.; Menssen, H. D.; Neri, D.; De Braud, F. The Tumour-Targeting Human L19-IL2 Immunocytokine: Preclinical Safety Studies, Phase i Clinical Trial in Patients with Solid Tumours and Expansion into Patients with Advanced Renal Cell Carcinoma. Eur J Cancer 2010, 46. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, L.; Wan, D.; Zhou, L.; Zheng, S.; Lin, S.; Qiao, Y. Extracellular Matrix and Its Therapeutic Potential for Cancer Treatment. Signal Transduct Target Ther 2021, 6. [Google Scholar] [CrossRef]
- Liu, S.; Ren, J.; ten Dijke, P. Targeting TGFβ Signal Transduction for Cancer Therapy. Signal Transduction and Targeted Therapy 2021, 6, 1–20. [Google Scholar] [CrossRef]
- Zion, O.; Genin, O.; Kawada, N.; Yoshizato, K.; Pancreas, S. R.-. 2009, undefined. Inhibition of Transforming Growth Factor β Signaling by Halofuginone as a Modality for Pancreas Fibrosis Prevention. journals.lww.com.
- Castellani, P.; Viale, G.; Dorcaratto, A.; Nicolo, G.; Kaczmarek, J.; Querze, G.; Zardi, L. Erratum: The Fibronectin Isoform Containing the ED-B Oncofetal Domain: A Marker of Angiogenesis (Int. J. Cancer, 59, 612-618 (1994)). International Journal of Cancer. 1995. [CrossRef]
- Glukhova, M. A.; Frid, M. G.; Shekhonin, B. V.; Balabanov, Y. V.; Koteliansky, V. E. Expression of Fibronectin Variants in Vascular and Visceral Smooth Muscle Cells in Development. Dev Biol 1990, 141. [Google Scholar] [CrossRef] [PubMed]
- Rybak, J. N.; Roesli, C.; Kaspar, M.; Villa, A.; Neri, D. The Extra-Domain A of Fibronectin Is a Vascular Marker of Solid Tumors and Metastases. Cancer Res 2007, 67. [Google Scholar] [CrossRef] [PubMed]
- Eigentler, T. K.; Weide, B.; De Braud, F.; Spitaleri, G.; Romanini, A.; Pflugfelder, A.; Gonzaĺez-Iglesias, R.; Tasciotti, A.; Giovannoni, L.; Schwager, K.; Lovato, V.; Kaspar, M.; Trachsel, E.; Menssen, H. D.; Neri, D.; Garbe, C. A Dose-Escalation and Signal-Generating Study of the Immunocytokine L19-IL2 in Combination with Dacarbazine for the Therapy of Patients with Metastatic Melanoma. Clinical Cancer Research 2011, 17. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M. E.; Gorlia, T.; Erridge, S. C.; Perry, J.; Hong, Y. K.; Aldape, K. D.; Lhermitte, B.; Pietsch, T.; Grujicic, D.; Steinbach, J. P. eter; Wick, W.; Tarnawski, R.; Nam, D. H.; Hau, P.; Weyerbrock, A.; Taphoorn, M. J. B.; Shen, C. C.; Rao, N.; Thurzo, L.; Herrlinger, U.; Gupta, T.; Kortmann, R. D.; Adamska, K.; McBain, C.; Brandes, A. A.; Tonn, J. C. hristian; Schnell, O.; Wiegel, T.; Kim, C. Y.; Nabors, L. B. urt; Reardon, D. A.; van den Bent, M. J.; Hicking, C.; Markivskyy, A.; Picard, M.; Weller, M. Cilengitide Combined with Standard Treatment for Patients with Newly Diagnosed Glioblastoma with Methylated MGMT Promoter (CENTRIC EORTC 26071-22072 Study): A Multicentre, Randomised, Open-Label, Phase 3 Trial. Lancet Oncol 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Lawhorn, B. G.; Brnardic, E. J.; Behm, D. J. TRPV4 Antagonists: A Patent Review (2015–2020). Expert Opinion on Therapeutic Patents 2021. [CrossRef] [PubMed]
- Doñate-Macian, P.; Duarte, Y.; Rubio-Moscardo, F.; Pérez-Vilaró, G.; Canan, J.; Díez, J.; González-Nilo, F.; Valverde, M. A. Structural Determinants of TRPV4 Inhibition and Identification of New Antagonists with Antiviral Activity. Br J Pharmacol 2022, 179. [Google Scholar] [CrossRef] [PubMed]
- Article, R. YAP / TAZ as Master Regulators in Cancer : Modulation, Function and Therapeutic Approaches Check for Updates. 2023, 4 (January), 9–26.
- Barry, E. R.; Simov, V.; Valtingojer, I.; Venier, O. Recent Therapeutic Approaches to Modulate the Hippo Pathway in Oncology and Regenerative Medicine. Cells 2021. [Google Scholar] [CrossRef]
- Liu-Chittenden, Y.; Huang, B.; Shim, J. S.; Chen, Q.; Lee, S. J.; Anders, R. A.; Liu, J. O.; Pan, D. Genetic and Pharmacological Disruption of the TEAD-YAP Complex Suppresses the Oncogenic Activity of YAP. Genes Dev 2012, 26. [Google Scholar] [CrossRef]
- Dasari, V. R.; Mazack, V.; Feng, W.; Nash, J.; Carey, D. J.; Gogoi, R. Verteporfin Exhibits YAP-Independent Anti-Proliferative and Cytotoxic Effects in Endometrial Cancer Cells. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Zhang, H.; Ramakrishnan, S. K.; Triner, D.; Centofanti, B.; Maitra, D.; Gyorffy, B.; Sebolt-Leopold, J. S.; Dame, M. K.; Varani, J.; Brenner, D. E.; Fearon, E. R.; Omary, M. B.; Shah, Y. M. Tumor-Selective Proteotoxicity of Verteporfin Inhibits Colon Cancer Progression Independently of YAP1. Sci Signal 2015, 8. [Google Scholar] [CrossRef]
- Liu, S.; Li, Y.; Hong, Y.; Wang, M.; Zhang, H.; Ma, J.; Qu, K.; Huang, G.; Lu, T. J. Mechanotherapy in Oncology: Targeting Nuclear Mechanics and Mechanotransduction. Adv Drug Deliv Rev 2023, 194. [Google Scholar] [CrossRef] [PubMed]
- Collis, J.; Manasseh, R.; Liovic, P.; Tho, P.; Ultrasonics, A. O.-. 2010, undefined. Cavitation Microstreaming and Stress Fields Created by Microbubbles. Elsevier.
- Krasovitski, B.; Frenkel, V.; Shoham, S.; Kimmel, E. Intramembrane Cavitation as a Unifying Mechanism for Ultrasound-Induced Bioeffects. Proc Natl Acad Sci U S A 2011, 108, 3258–3263. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Gerriets, T.; Walberer, M.; Mueller, C.; Rolke, R.; Eicke, B. M.; Bohl, J.; Kempski, O.; Kaps, M.; Bachmann, G.; Dieterich, M.; Nedelmann, M. Brain Edema and Intracerebral Necrosis Caused by Transcranial Low-Frequency 20-KHz Ultrasound: A Safety Study in Rats. Stroke 2006, 37, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Burleson, A.; Nusstein, J.; Reader, A.; endodontics, M. B.-J. of; 2007, undefined. The in Vivo Evaluation of Hand/Rotary/Ultrasound Instrumentation in Necrotic, Human Mandibular Molars. Elsevier.
- Li, D.; Pellegrino, A.; Hallack, A.; Petrinic, N.; … A., J.-B. 2018, undefined. Response of Single Cells to Shock Waves and Numerically Optimized Waveforms for Cancer Therapy. Elsevier.
- Hato, T.; Goyal, L.; Greten, T. F.; Duda, D. G.; Zhu, A. X. Immune Checkpoint Blockade in Hepatocellular Carcinoma: Current Progress and Future Directions. Hepatology 2014. [CrossRef]
- Ribas, A.; Wolchok, J. D. Cancer Immunotherapy Using Checkpoint Blockade. Science 2018. [CrossRef]
- Fu, Y.; Peng, Y.; Zhao, S.; Mou, J.; Zeng, L.; Jiang, X.; Yang, C.; Huang, C.; Li, Y.; Lu, Y.; Wu, M.; Yang, Y.; Kong, T.; Lai, Q.; Wu, Y.; Yao, Y.; Wang, Y.; Gou, L.; Yang, J. Combination Foretinib and Anti-PD-1 Antibody Immunotherapy for Colorectal Carcinoma. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Xiong, G.; Cao, Z.; Yang, G.; Zheng, S.; Song, X.; You, L.; Zheng, L.; Zhang, T.; Zhao, Y. PD-1/PD-L1 and Immunotherapy for Pancreatic Cancer. Cancer Letters 2017. [CrossRef]
- Harrison, D. L.; Fang, Y.; Huang, J. T-Cell Mechanobiology: Force Sensation, Potentiation, and Translation. Frontiers in Physics 2019. [CrossRef]
- Hu, K. H.; Butte, M. J. T Cell Activation Requires Force Generation. Journal of Cell Biology 2016, 213. [Google Scholar] [CrossRef]
- Le Floc’H, A.; Huse, M. Molecular Mechanisms and Functional Implications of Polarized Actin Remodeling at the T Cell Immunological Synapse. Cellular and Molecular Life Sciences 2015, 72. [Google Scholar] [CrossRef]
- Tabdanov, E.; Gondarenko, S.; Kumari, S.; Liapis, A.; Dustin, M. L.; Sheetz, M. P.; Kam, L. C.; Iskratsch, T. Micropatterning of TCR and LFA-1 Ligands Reveals Complementary Effects on Cytoskeleton Mechanics in T Cells. Integrative Biology (United Kingdom) 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, C. S. C.; Raychaudhuri, D.; Paul, B.; Chakrabarty, Y.; Ghosh, A. R.; Rahaman, O.; Talukdar, A.; Ganguly, D. Cutting Edge: Piezo1 Mechanosensors Optimize Human T Cell Activation. The Journal of Immunology 2018, 200. [Google Scholar] [CrossRef] [PubMed]
- Tabdanov, E. D.; Rodríguez-Merced, N. J.; Cartagena-Rivera, A. X.; Puram, V. V.; Callaway, M. K.; Ensminger, E. A.; Pomeroy, E. J.; Yamamoto, K.; Lahr, W. S.; Webber, B. R.; Moriarity, B. S.; Zhovmer, A. S.; Provenzano, P. P. Engineering T Cells to Enhance 3D Migration through Structurally and Mechanically Complex Tumor Microenvironments. Nat Commun 2021, 12. [Google Scholar] [CrossRef]
- Hickey, J. W.; Dong, Y.; Chung, J. W.; Salathe, S. F.; Pruitt, H. C.; Li, X.; Chang, C.; Fraser, A. K.; Bessell, C. A.; Ewald, A. J.; Gerecht, S.; Mao, H. Q.; Schneck, J. P. Engineering an Artificial T-Cell Stimulating Matrix for Immunotherapy. Advanced Materials 2019, 31. [Google Scholar] [CrossRef]
- Wang, W.; Wu, F.; Mohammadniaei, M.; Zhang, M.; Li, Y.; Sun, Y.; Tang, B. Z. Genetically Edited T-Cell Membrane Coated AIEgen Nanoparticles Effectively Prevents Glioblastoma Recurrence. Biomaterials 2023, 293. [Google Scholar] [CrossRef]
- Kim, K. Do; Bae, S.; Capece, T.; Nedelkovska, H.; De Rubio, R. G.; Smrcka, A. V.; Jun, C. D.; Jung, W.; Park, B.; Kim, T. Il; Kim, M. Targeted Calcium Influx Boosts Cytotoxic T Lymphocyte Function in the Tumour Microenvironment. Nat Commun 2017, 8. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, Y.; Tan, X.; Zheng, X.; Wang, F.; Ke, K.; Zhang, C.; Liao, N.; Dang, Y.; Shi, Y.; Zheng, Y.; Gao, Y.; Li, Q.; Liu, X.; Liu, J. An Optogenetic Controllable T Cell System for Hepatocellular Carcinoma Immunotherapy. Theranostics 2019, 9. [Google Scholar] [CrossRef]
- Tan, P.; He, L.; Han, G.; Zhou, Y. Optogenetic Immunomodulation: Shedding Light on Antitumor Immunity. Trends in Biotechnology 2017. [CrossRef]
- Wu, Y.; Liu, Y.; Huang, Z.; Wang, X.; Jin, Z.; Li, J.; Limsakul, P.; Zhu, L.; Allen, M.; Pan, Y.; Bussell, R.; Jacobson, A.; Liu, T.; Chien, S.; Wang, Y. Control of the Activity of CAR-T Cells within Tumours via Focused Ultrasound. Nat Biomed Eng 2021, 5. [Google Scholar] [CrossRef]
- Liao, D.; Li, F.; Lu, D.; Zhong, P. Activation of Piezo1 Mechanosensitive Ion Channel in HEK293T Cells by 30 MHz Vertically Deployed Surface Acoustic Waves. Biochem Biophys Res Commun 2019, 518, 541–547. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, J.; Kala, S.; Zhu, J.; Xian, Q.; Qiu, W.; Li, G.; Zhu, T.; Meng, L.; Zhang, R.; Chan, H. C.; Zheng, H.; Sun, L. The Mechanosensitive Ion Channel Piezo1 Significantly Mediates In Vitro Ultrasonic Stimulation of Neurons. iScience 2019, 21. [Google Scholar] [CrossRef]
- Pan, Y.; Yoon, S.; Sun, J.; Huang, Z.; Lee, C.; Allen, M.; Wu, Y.; Chang, Y. J.; Sadelain, M.; Kirk Shung, K.; Chien, S.; Wang, Y. Mechanogenetics for the Remote and Noninvasive Control of Cancer Immunotherapy. Proc Natl Acad Sci U S A 2018, 115. [Google Scholar] [CrossRef] [PubMed]
- Billadeau, D. D.; Nolz, J. C.; Gomez, T. S. Regulation of T-Cell Activation by the Cytoskeleton. Nature Reviews Immunology 2007. [CrossRef] [PubMed]
- Al-Alwan, M. M.; Liwski, R. S.; Haeryfar, S. M. M.; Baldridge, W. H.; Hoskin, D. W.; Rowden, G.; West, K. A. Cutting Edge: Dendritic Cell Actin Cytoskeletal Polarization during Immunological Synapse Formation Is Highly Antigen-Dependent. The Journal of Immunology 2003, 171. [Google Scholar] [CrossRef] [PubMed]
- Gombos, I.; Detre, C.; Vámosi, G.; Matkó, J. Rafting MHC-II Domains in the APC (Presynaptic) Plasma Membrane and the Thresholds for T-Cell Activation and Immunological Synapse Formation. Immunol Lett 2004, 92, 117–124. [Google Scholar] [CrossRef]
- Jönsson, F.; Gurniak, C. B.; Fleischer, B.; Kirfel, G.; Witke, W. Immunological Responses and Actin Dynamics in Macrophages Are Controlled by N-Cofilin but Are Independent from ADF. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Baranov, M. V.; Beest, T. M.; Reinieren-Beeren, I.; Cambi, A.; Figdor, C. G.; Den Bogaart, G. Van. Podosomes of Dendritic Cells Facilitate Antigen Sampling. J Cell Sci 2014, 127. [Google Scholar] [CrossRef]
- Xu, Z. L.; Zhu, X. Q.; Lu, P.; Zhou, Q.; Zhang, J.; Wu, F. Activation of Tumor-Infiltrating Antigen Presenting Cells by High Intensity Focused Ultrasound Ablation of Human Breast Cancer. Ultrasound Med Biol 2009, 35. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, L.; Chen, W. R. Host Antitumor Immune Responses to HIFU Ablation. International Journal of Hyperthermia 2007, 23. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, Y.; Feng, J.; Wu, F. Dendritic Cells Loaded with Ultrasound-Ablated Tumour Induce in Vivo Specific Antitumour Immune Responses. Ultrasound Med Biol 2010, 36. [Google Scholar] [CrossRef]
- Marinova, M.; Huxold, H. C.; Henseler, J.; Mücke, M.; Conrad, R.; Rolke, R.; Ahmadzadehfar, H.; Rauch, M.; Fimmers, R.; Luechters, G.; Cuhls, H.; Radbruch, L.; Schild, H. H.; Strunk, H. Clinical Effectiveness and Potential Survival Benefit of US-Guided High-Intensity Focused Ultrasound Therapy in Patients with Advanced-Stage Pancreatic Cancer. Ultraschall in der Medizin 2019, 40. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yu, Z.; Liu, N.; Huang, J.; Liang, X.; Liang, X.; Liang, M.; Li, M.; Ni, J. The Efficacy of Low-Frequency Ultrasound as an Added Treatment for Chronic Wounds: A Meta-Analysis. Int Wound J 2023, 20. [Google Scholar] [CrossRef]
- Zhang, W.; Shou, W. De; Xu, Y. J.; Bai, W. K.; Hu, B. Low-Frequency Ultrasound-Induced VEGF Suppression and Synergy with Dendritic Cell-Mediated Anti-Tumor Immunity in Murine Prostate Cancer Cells in Vitro. Sci Rep 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Vogel, V. Spatial Confinement Downsizes the Inflammatory Response of Macrophages. Nat Mater 2018, 17. [Google Scholar] [CrossRef]
- Okamoto, T.; Takagi, Y.; Kawamoto, E.; Park, E. J.; Usuda, H.; Wada, K.; Shimaoka, M. Reduced Substrate Stiffness Promotes M2-like Macrophage Activation and Enhances Peroxisome Proliferator-Activated Receptor γ Expression. Exp Cell Res 2018, 367. [Google Scholar] [CrossRef]
- Sridharan, R.; Cavanagh, B.; Cameron, A. R.; Kelly, D. J.; O’Brien, F. J. Material Stiffness Influences the Polarization State, Function and Migration Mode of Macrophages. Acta Biomater 2019, 89. [Google Scholar] [CrossRef]
- Meli, V. S.; Atcha, H.; Veerasubramanian, P. K.; Nagalla, R. R.; Luu, T. U.; Chen, E. Y.; Guerrero-Juarez, C. F.; Yamaga, K.; Pandori, W.; Hsieh, J. Y.; Downing, T. L.; Fruman, D. A.; Lodoen, M. B.; Plikus, M. V.; Wang, W.; Liu, W. F. YAP-Mediated Mechanotransduction Tunes the Macrophage Inflammatory Response. Sci Adv 2020, 6. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage Polarization: Tumor-Associated Macrophages as a Paradigm for Polarized M2 Mononuclear Phagocytes. Trends in Immunology 2002. [CrossRef]
- Guerriero, J. L.; Sotayo, A.; Ponichtera, H. E.; Castrillon, J. A.; Pourzia, A. L.; Schad, S.; Johnson, S. F.; Carrasco, R. D.; Lazo, S.; Bronson, R. T.; Davis, S. P.; Lobera, M.; Nolan, M. A.; Letai, A. Class IIa HDAC Inhibition Reduces Breast Tumours and Metastases through Anti-Tumour Macrophages. Nature 2017, 543. [Google Scholar] [CrossRef]
- Heusinkveld, M.; van der Burg, S. H. Identification and Manipulation of Tumor Associated Macrophages in Human Cancers. Journal of Translational Medicine 2011. [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; He, P.; Han, J.; Sun, C. Interleukin-37 Suppresses Hepatocellular Carcinoma Growth through Inhibiting M2 Polarization of Tumor-Associated Macrophages. Mol Immunol 2020, 122. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Fang, B.; Zhang, Y.; Wang, C.; Zhou, J.; Niu, C.; Gao, Y.; Zhao, D.; He, J.; Wang, J.; Zhang, X.; Li, Q. Mechanical Stretch Promotes Tumoricidal M1 Polarization via the FAK/NF-ΚB Signaling Pathway. FASEB Journal 2019, 33. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Guo, L.; Shang, M.; Shi, D.; Liang, P.; Jing, X.; Meng, D.; Liu, X.; Zhou, X.; Zhao, Y.; Li, J. Ultrasound Mediated Destruction of Lmw-Ha-Loaded and Folate-Conjugated Nanobubble for Tam Targeting and Reeducation. Int J Nanomedicine 2020, 15. [Google Scholar] [CrossRef]
- Labernadie, A.; Kato, T.; Brugués, A.; Serra-Picamal, X.; Derzsi, S.; Arwert, E.; Weston, A.; González-Tarragó, V.; Elosegui-Artola, A.; Albertazzi, L.; Alcaraz, J.; Roca-Cusachs, P.; Sahai, E.; Trepat, X. A Mechanically Active Heterotypic E-Cadherin/N-Cadherin Adhesion Enables Fibroblasts to Drive Cancer Cell Invasion. Nat Cell Biol 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Barbazan, J.; Pérez-González, C.; Gómez-González, M.; Dedenon, M.; Richon, S.; Latorre, E.; Serra, M.; Mariani, P.; Descroix, S.; Sens, P.; Trepat, X.; Vignjevic, D. M. Cancer-Associated Fibroblasts Actively Compress Cancer Cells and Modulate Mechanotransduction. bioRxiv 2021. [CrossRef]
- Johansson, A. C.; Ansell, A.; Jerhammar, F.; Lindh, M. B.; Grénman, R.; Munck-Wikland, E.; Östman, A.; Roberg, K. Cancer-Associated Fibroblasts Induce Matrix Metalloproteinase-Mediated Cetuximab Resistance in Head and Neck Squamous Cell Carcinoma Cells. Molecular Cancer Research 2012, 10. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Ren, Y.; Geng, H.; Zhang, Q.; Cao, L.; Meng, Z.; Wu, X.; Xu, M.; Xu, K. Cancer-Associated Fibroblasts Contribute to Cisplatin Resistance by Modulating ANXA3 in Lung Cancer Cells. Cancer Sci 2019, 110. [Google Scholar] [CrossRef]
- Long, X.; Xiong, W.; Zeng, X.; Qi, L.; Cai, Y.; Mo, M.; Jiang, H.; Zhu, B.; Chen, Z.; Li, Y. Cancer-Associated Fibroblasts Promote Cisplatin Resistance in Bladder Cancer Cells by Increasing IGF-1/ERβ/Bcl-2 Signalling. Cell Death Dis 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yue, J.; Jiang, Z.; Zhou, R.; Xie, R.; Xu, Y.; Wu, S. CAF-Secreted CXCL1 Conferred Radioresistance by Regulating DNA Damage Response in a ROS-Dependent Manner in Esophageal Squamous Cell Carcinoma. Cell Death Dis 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gan, G.; Wang, B.; Wu, J.; Cao, Y.; Zhu, D.; Xu, Y.; Wang, X.; Han, H.; Li, X.; Ye, M.; Zhao, J.; Mi, J. Cancer-Associated Fibroblasts Promote Irradiated Cancer Cell Recovery Through Autophagy. EBioMedicine 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Nicolas-Boluda, A.; Vaquero, J.; Vimeux, L.; Guilbert, T.; Barrin, S.; Kantari-Mimoun, C.; Ponzo, M.; Renault, G.; Deptula, P.; Pogoda, K.; Bucki, R.; Cascone, I.; Courty, J.; Fouassier, L.; Gazeau, F.; Donnadieu, E. Tumor Stiffening Reversion through Collagen Crosslinking Inhibition Improves t Cell Migration and Anti-Pd-1 Treatment. Elife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Grither, W. R.; Van Hove, S.; Biswas, H.; Ponik, S. M.; Eliceiri, K. W.; Keely, P. J.; Longmore, G. D. Mechanical Signals Regulate and Activate SNAIL1 Protein to Control the Fibrogenic Response of Cancer-Associated Fibroblasts. J Cell Sci 2016, 129. [Google Scholar] [CrossRef]
- Sewell-Loftin, M. K.; Bayer, S. V. H.; Crist, E.; Hughes, T.; Joison, S. M.; Longmore, G. D.; George, S. C. Cancer-Associated Fibroblasts Support Vascular Growth through Mechanical Force. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Rinella, L.; Pizzo, B.; Frairia, R.; Delsedime, L.; Calleris, G.; Gontero, P.; Zunino, V.; Fortunati, N.; Arvat, E.; Catalano, M. G. Modulating Tumor Reactive Stroma by Extracorporeal Shock Waves to Control Prostate Cancer Progression. Prostate 2020, 80. [Google Scholar] [CrossRef]
Figure 1.
The types of mechanical forces experienced by cancer cells during the onstage of cancer and its progression. Compressive forces are generated due to the indefinite growth of cancer cells in the dense tumor interior. The pushing and pulling by the cancer cells on a dense network of crosslinked ECM fibers generates tensile forces. Cancer cells experience fluid-generated shear forces during their migration through interstitial fluid and blood flow.
Figure 1.
The types of mechanical forces experienced by cancer cells during the onstage of cancer and its progression. Compressive forces are generated due to the indefinite growth of cancer cells in the dense tumor interior. The pushing and pulling by the cancer cells on a dense network of crosslinked ECM fibers generates tensile forces. Cancer cells experience fluid-generated shear forces during their migration through interstitial fluid and blood flow.
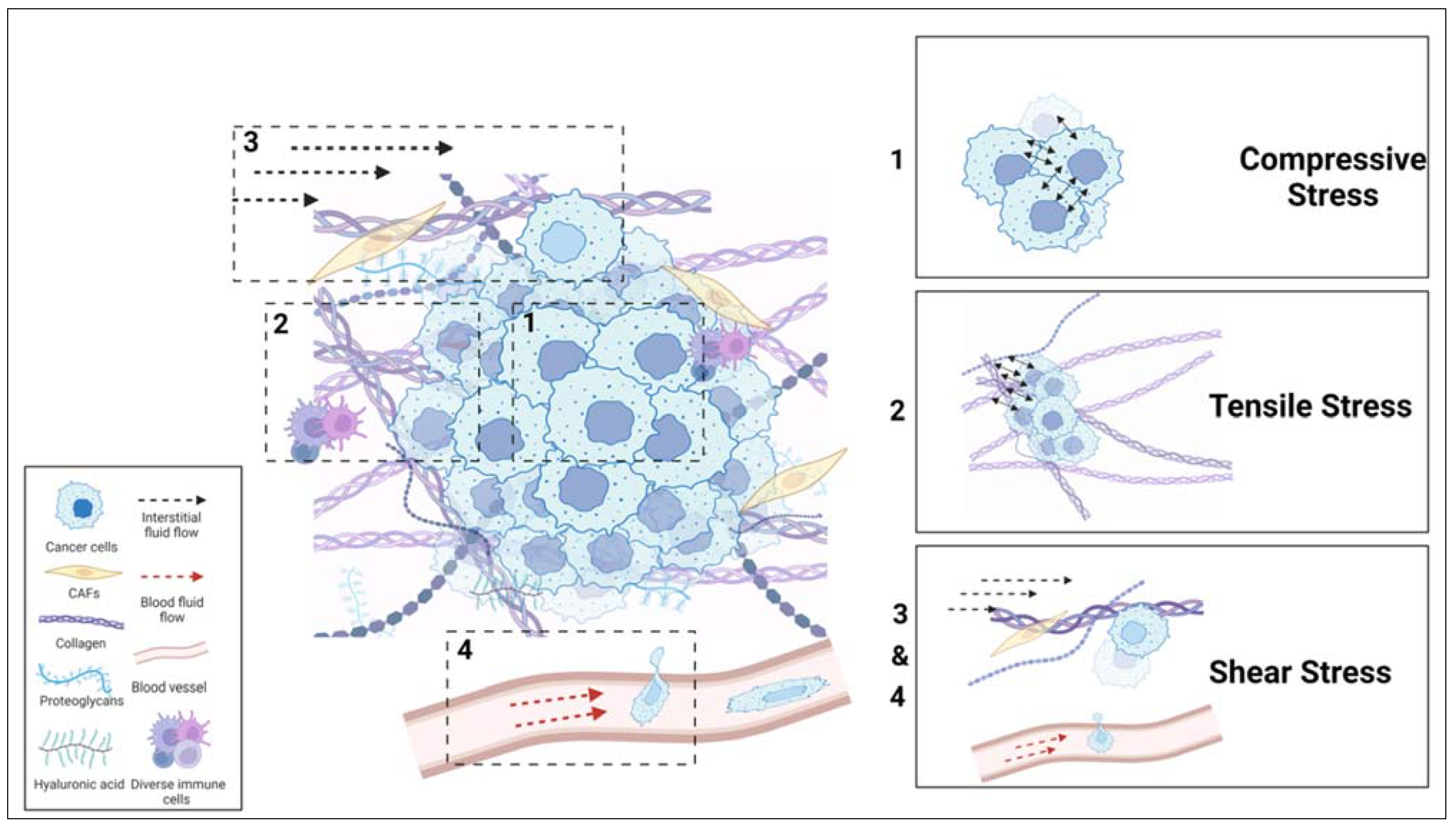
Figure 2.
Cancer cell-TME interactions in 3D. Cancer cells are mainly surrounded by a dense network of ECM, cancer-associated cells and vasculature within the tumor site. Cancer cells sense biomechanical properties of TME, such as ECM characteristics (stiffness, ligand density, confinement, degradability, porosity, viscoelasticity), RGD nanospacing, fluid viscosity and then activate intracellular signaling pathways, which in turn regulates cancer progression.
Figure 2.
Cancer cell-TME interactions in 3D. Cancer cells are mainly surrounded by a dense network of ECM, cancer-associated cells and vasculature within the tumor site. Cancer cells sense biomechanical properties of TME, such as ECM characteristics (stiffness, ligand density, confinement, degradability, porosity, viscoelasticity), RGD nanospacing, fluid viscosity and then activate intracellular signaling pathways, which in turn regulates cancer progression.
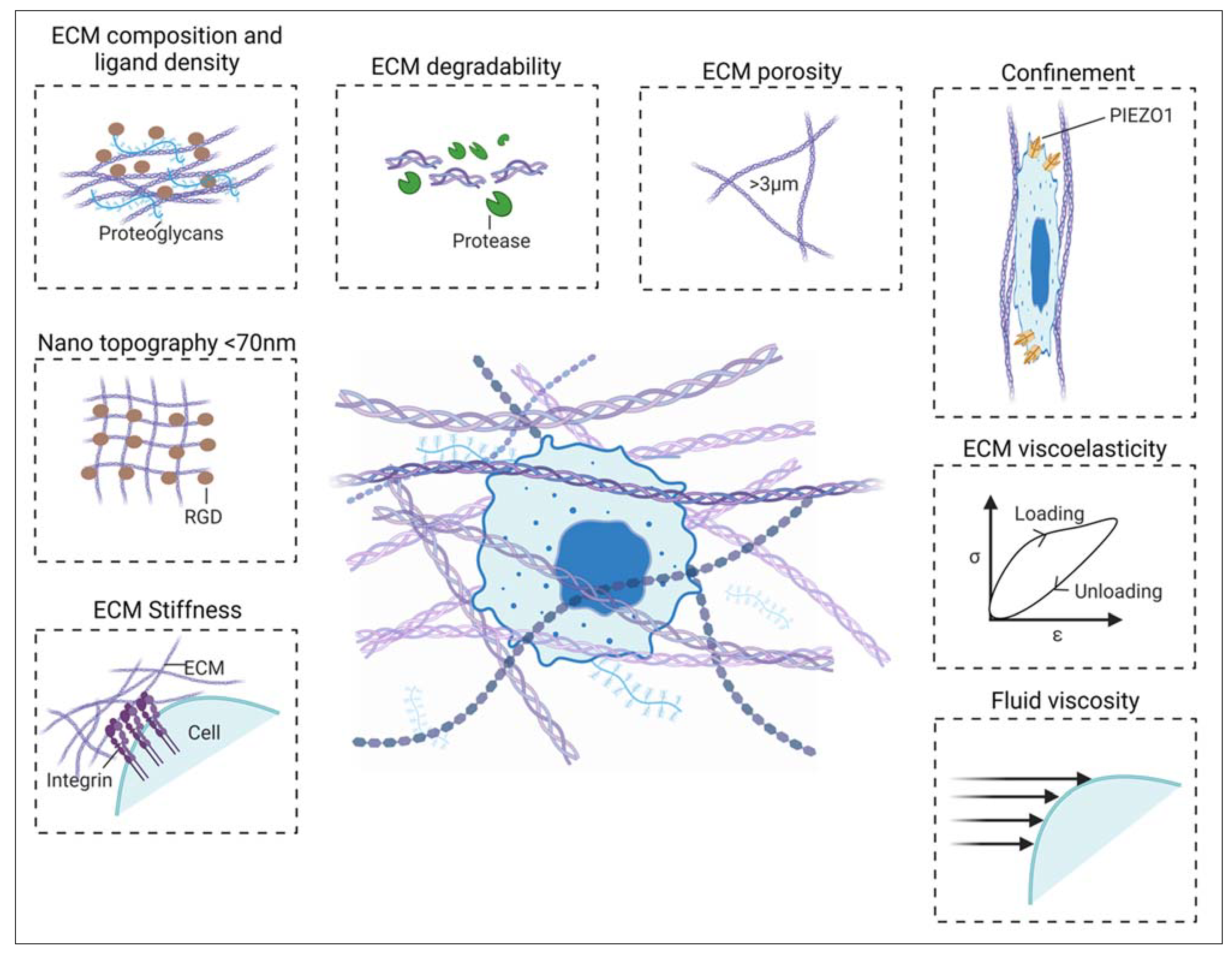
Figure 3.
Schematic representation of mechanobiology-based therapies. Targeting cancer cells by mechanical perturbations like (a) shock wave [61,62,63,76] (b) mechanical stretch [63,64,138,139] (c) High-Intensity Ultrasound Frequency [40,72,125,126,127](d) Low-Intensity Pulsed Ultrasound [67,96,97,98,99](e) targeting cells associated with tumors associated T-cells, [107,112] dendritic cells,[121,123,126] tumor-associated macrophages, [136] cancer-associated fibroblasts[147] and (f) targeting mechanosensitive channels, mechanosensors and mechanotransducers of the cancer cell including TRPV4,[50,85] Piezo1,[49,63,105,113,114] Rho/ROCK, [10,146] YAP/TAZ [51,52,53,54,55] and TGF-β [28,30,35,77].
Figure 3.
Schematic representation of mechanobiology-based therapies. Targeting cancer cells by mechanical perturbations like (a) shock wave [61,62,63,76] (b) mechanical stretch [63,64,138,139] (c) High-Intensity Ultrasound Frequency [40,72,125,126,127](d) Low-Intensity Pulsed Ultrasound [67,96,97,98,99](e) targeting cells associated with tumors associated T-cells, [107,112] dendritic cells,[121,123,126] tumor-associated macrophages, [136] cancer-associated fibroblasts[147] and (f) targeting mechanosensitive channels, mechanosensors and mechanotransducers of the cancer cell including TRPV4,[50,85] Piezo1,[49,63,105,113,114] Rho/ROCK, [10,146] YAP/TAZ [51,52,53,54,55] and TGF-β [28,30,35,77].
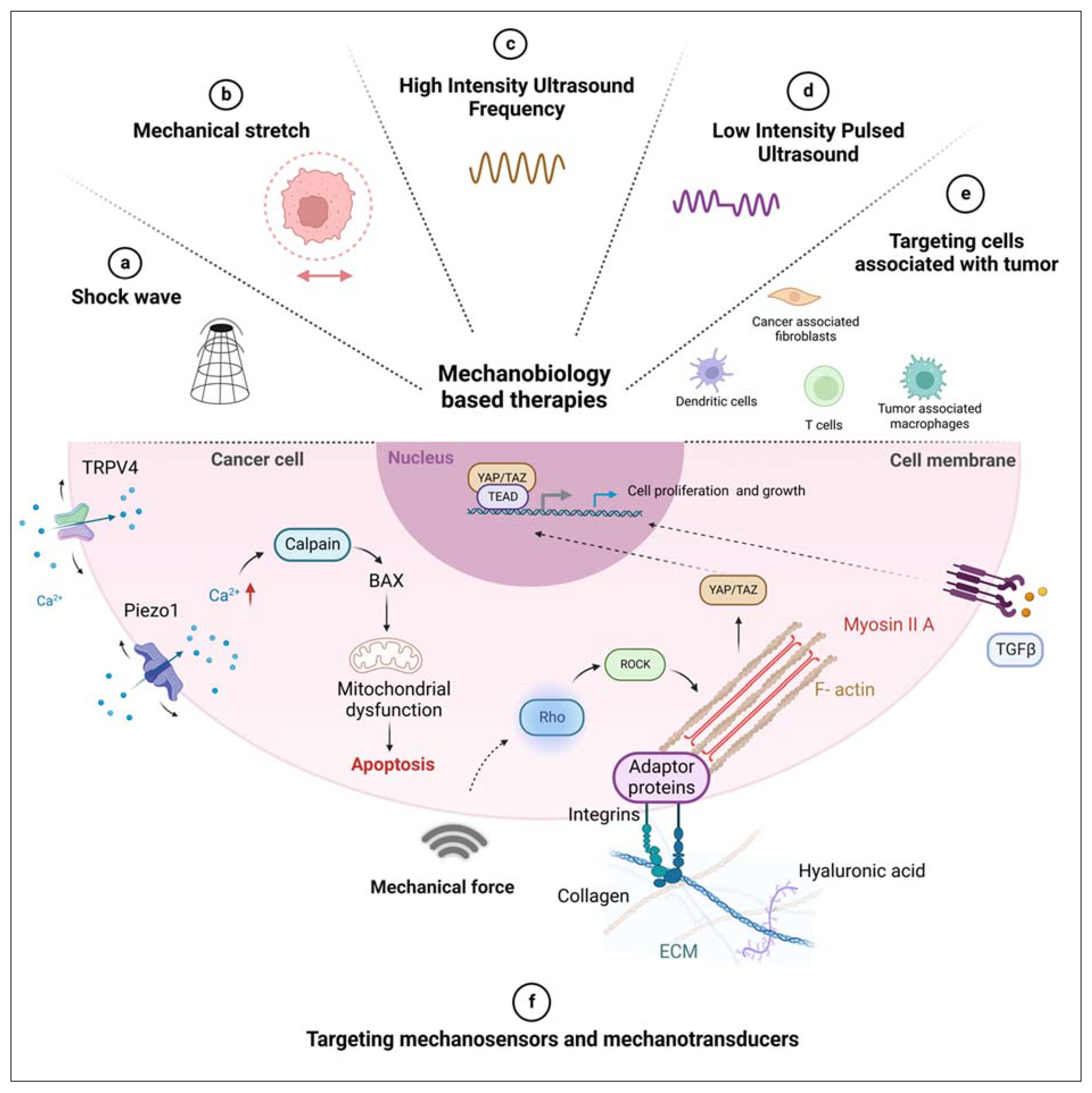

Table 1.
Existing mechanobiological-based strategies for cancer treatment
| Aim | Targeting | Targets | Targeting agents | Stage of clinical trials and cancer type |
|---|---|---|---|---|
| ECM stiffness reduction | Collagen production | TGF-β | Fresolimumab (NCT01401062), (NCT02581787) | I/II, metastatic breast cancer, early-stage non-small cell lung cancer[26] |
| Halofuginone | Animal models of pancreatic, lung, melanoma, and breast cancer[27,28,29,30] | |||
| LY-2109761 | In vitro (liver metastasis of colon cancer, pancreatic cancer metastasis), and animal model (breast cancer bone metastasis)[31,32,33] | |||
| Fibronectin production | Extra domain B (EDB) | AS1409 | I, malignant melanoma or renal cell carcinoma[34] | |
| Collagen chaperons | HSP7 | Pirfenidone, | In vitro, lung fibrosis[35] | |
| ND-L02-s0201 (NCT03241264) | I, fibrosis[36] | |||
| ECM crosslinking | Pan LOX, LOX, LOXL2 | PXS-5505 (NCT04676529) | II, myelofibrosis[37] | |
| PXS-5382A (NCT04183517) | I, healthy individuals | |||
| (NCT02852551) | I, healthy individuals | |||
| Simtuzumab (NCT01472198), (NCT01479465) |
Phase II, pancreatic adenocarcinoma[38], colorectal adenocarcinoma[39] | |||
| ECM structural disruption | Collagen | low power of Pulse-HIFU (20 W/cm2) | Animal model[40] | |
| Mechano-sensors (Stiffness sensors) and Transducers | Integrins | Cilengitide (NCT00093964) | III, glioblastoma[41] | |
| ATN-161 (NCT00352313) | II, malignant glioma[42] | |||
| anti-αVβ3 Ab -Etaracizumab (MEDI-522), | I/II, metastatic melanoma, renal cell, prostate cancer, lymphoma, small intestine cancer, colorectal cancer[43,44] | |||
| anti-α5β1 integrin Ab - Volociximab | II, metastatic pancreatic cancer, ovarian cancer peritoneal neoplasms, melanoma | |||
| anti-αV Ab -Intetumumab (NCT00246012), (NCT00537381) |
II, melanoma[45], prostate cancer[46] | |||
| anti-αV Ab -Abituzumab (NCT01008475), (NCT01360840) |
II, colorectal cancer[47], prostate cancer[48] | |||
| Piezo1 | GsMTx4 | In vitro, transfected HEK293 cells[49] | ||
| Gadolinium and ruthenium red | In vitro on non-cancerous cells, sensory cells[50], Piezo-expressing HE293T cells[51] | |||
| TRPV4 | GSK2798745 (NCT02119260) | II, Healthy subjects and Patients[52] | ||
| YAP/TAZ | ION537 (anti-YAP DNA antisense oligonucleotide) (NCT04659096) |
I, advanced solid tumors[53] | ||
| IAG933(NCT04857372) | I, ongoing, mesothelioma and other solid tumors[54] | |||
| VT398 (NCT04665206) | ||||
| Verteporfin (NCT04590664), (NCT03067051), (NCT03033225) |
I/II, glioblastoma, prostate cancer, pancreatic cancer[55,56,57] | |||
| To utilize mismatching of mechano-phenotype of cancerous and normal cells | Nuclear mechano-transduction | High frequency LIPU |
In vitro, breast carcinoma and a malignant melanoma[58], mice cervical cancer (HeLa cell)[59] |
|
| Shock wave therapy |
In vitro, bladder cancer cell and prostate cancer cell[60], human renal epithelial, cancer cell[61], hamster melanomas[62] |
|||
| Mechanical stretch therapy | In vitro and in vivo, breast cancer cells[63], p53PTEN-/- mice breast cancer model[64] | |||
| Low frequency LIPU |
In vitro and in vivo, breast cancer cell, chick embryo grafted tumors [63], murine mammary sarcoma and murine mammary sarcoma[65], Human foreskin fibroblasts[66], Breast cancer cells, malignant melanoma, breast epithelial cell[67], Hacat and Cal33, In vivo mice injected with Cal33 HNSCC cell line[68] |
|||
| Depletion of blood flow to tumor | Tumor blood vessels | Low intensity ultrasound | Animal models, hepatocellular carcinoma[69], rabbit[70] | |
| Dendritic cells | High intensity-focused ultrasound |
In vitro, primary effector CD4+ T cells obtained from TCR-transgenic OT-II mice[71], primary human CD4+ T cells[72], human breast cancer sample[73] |
||
| Low frequency ultrasound |
In vitro, murine prostate cancer cells (RM-1) and bone marrow derived DCs from BALB/c mice[74] |
|||
| TAM | Ultrasound targeted nanobubble destruction (UTND) | In vitro, mouse macrophage cell line RAW264.7 (M0) and Lewis lung carcinoma (LLC) cell line[75] | ||
| CAFs | Shock wave therapy | In vitro, CAFs extracted from tumors of prostate cancer patients[76] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



