
Submitted:
28 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Suppressors of cytokine signaling (SOCS) genes are essential negative regulators that modulate cytokine signaling and play key roles in numerous biological processes, including immune responses. In this study, we cloned the complementary DNA (cDNA) sequences of two SOCS genes, designated as LvSOCS6 and LvSOCS7, from the white-leg shrimp, Litopenaeus vannamei. LvSOCS6 encoded a polypeptide of 463 amino acids (aa), spanning 1,392 base pairs (bp), while LvSOCS7 encodes a significantly larger polypeptide of 955 aa, encompassing 2,868 bp. Both LvSOCS proteins exhibited conserved domains associated with SOCS, including a centralized Src homology 2 (SH2) domain and a C-terminal SOCS box. Phylogenetic analysis revealed that the deduced aa sequences of LvSOCS6 and LvSOCS7 cluster within the invertebrate type I SOCS family, indicating their evolutionary relatedness. Tissue distribution analysis demonstrated ubiquitous expression of both LvSOCS genes across all examined tissues, with LvSOCS6 showing heightened expression in the gills and LvSOCS7 exhibiting a remarkable preference for the gills and stomach. Notably, mRNA expression patterns of LvSOCS genes following LPS and poly (I:C) stimulations exhibit significant upregulations, while PGN stimulation yields incongruous results across the experimented tissues. Interestingly, concurrent with the diminished expression of LvSOCS6 and LvSOCS7, there is a significant elevation in mRNA expression levels of LvSTAT, a vital component of the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) pathway. These findings shed light on the potential involvement of the newly discovered type I SOCS genes in regulating the JAK/STAT pathway and assuming pivotal roles in orchestrating the innate immune responses in L. vannamei defense mechanisms.
Keywords:
Litopenaeus vannamei
; Suppressor of cytokine signaling
; Gene expression
; JAK-STAT signaling pathway
1. Introduction
Cytokines, secretary proteins molecules, are pivotal players in various biological processes including proliferation, growth, and immune regulation across both vertebrates and invertebrates [1,2,3]. Most of the cytokines are involved with the Janus kinase (JAK) and the signal transducers and activators of transcription (STAT) pathways, which are known to play an essential role in immune responses [4]. Accumulating evidence suggests the presence of a number of JAKs and STATs in vertebrates including humans and fish, and their possible regulation by these molecules [5,6]. Hence, tight control of cytokines is vital for maintaining immune homeostasis. Consequently, various molecules have been identified that act as physiological suppressors and restrain the excessive activities of cytokines. Among them, the suppressor of cytokine signaling molecules (SOCS) hold particular significance as the “key negative regulators” of these molecules [7].
The SOCS family of vertebrates comprises 8 members, namely SOCS1–7 and cytokine-inducible SH2-containing protein (CISH) [8,9]. Based on the available evolutionary analysis data, two distinctive SOCS groups have been identified as type II with SOCS1–3 and CISH and type I with SOCS4–7. All SOCS members share common structural features, including a C-terminal SOCS box and a centralized SH2 domain [1,8,9]. Meanwhile, their N terminal regions exhibit variability which is believed to be associated with their specific functions. For instance, SOCS1 and SOCS3 have a unique N- terminal domain, kinase inhibitory region (KIR), which is essential for the suppression of the JAK tyrosine kinase activity [10,11]. The SOCS4 and SOCS5 contain a conserved region in their N terminal (N-terminal conserved region-NLTR) which has been shown to play an important role in the regulation of epidermal growth factor receptor signaling [12,13]. Interestingly, SOCS6 and SOCS7 do not have any of these specifications in their respective N-terminal regions [14].
Despite the well-documented immune regulatory activities of SOCS genes in vertebrates, their functions in invertebrates, particularly the type I SOCS, remain relatively undiscovered. As such, several research efforts have been directed toward understanding the role of SOCS in the immune regulation of invertebrates. The first identified invertebrate SOCS was SOCS-36E, in fruit fly (Drosophila melanogaster) revealed it has similar characteristics and functional significance to the human SOCS5, in regulating the JAK-STAT pathway [15]. Subsequent studies unrevealed 2 additional SOCS genes in D. melanogaster, named SOCS44A and SOCS16D, which displayed sequence similarities (33-34% and 45-48%) to the human SOCS6 and SOCS7, with the potential regulatory capabilities in the JAK-STAT pathway [16]. In reference to crustaceans, the role of SOCS6 in the Chinese mitten crab (Eriocheir sinensis) was recently elucidated, highlighting its involvement in the mediation of both JAK-STAT and NF-κB (nuclear factor-kappa B) pathways [14]. Additionally, in the Pacific oyster (Crassostrea gigas), three SOCS genes (SOCS2, SOCS5, SOCS7) have been identified, exhibiting potential in the regulation of NF-κB transcription [17]. Furthermore, the expression and functional implications of SOCS2 (belongs to the type II SOCS family) in the innate immune responses of red swamp crayfish and white leg shrimp (Procambarus clarkii and L. vannamei) have been investigated following lipopolysaccharide (LPS) and/or bacterial challenges [18,19]. These studies conveyed the importance of SOCS genes in the immune defense mechanisms of crustaceans.
However, despite the growing body of research on invertebrate SOCS, there is a notable knowledge gap regarding the presence and function of type I SOCS genes in shrimp. The present study aims to address this gap by investigating the presence of type I SOCS genes (LvSOCS6 and LvSOCS7) in white-leg shrimp. The mRNA expression levels of identified SOCS upon post-immune stimulations and their possible association with the JAK-STAT signaling pathway was investigated, expanding our understanding of the immune regulatory mechanisms of SOCS in crustaceans.
2. Materials and Methods
2.1. Shrimp rearing and tissue collection
One-month-old healthy juvenile white leg shrimps, L. vannamei (body weight, BW = 0.9 ± 0.1 g; body length, BL = 1.8 ± 0.2 cm), were purchased from a commercial shrimp farm in Muan-gun, Jeollanam-do, South Korea. All shrimps were transported alive to the laboratory of Pukyong National University and reared in a recirculating aquarium tank (width × depth × height = 1.0 m × 3.0 m × 0.5 m) equipped with sponge-filtered and UV-sterilized seawater. Shrimps were fed four times per day with a commercially formulated shrimp diet (Jeil Feed. Co., Ltd) on an ad libitum basis. The rearing tank was maintained under continuous aeration (dissolved oxygen, DO = 9.7± 0.2 mg/L) with an ambient temperature of 24 ± 1 °C and pH of 7.6 –7.8. Water chemical parameters were measured once a day, and the concentrations of ammonia (NH3), nitrites (NO2-), and nitrates (NO3-) were maintained at 0.25-0.5 mg/L, 0.25 mg/L, and 20-40 mg/L, respectively, to ensure the optimal rearing conditions. The white leg shrimp (L. vannamei) grown to BW of 3.0 ± 0.5 g and BL of 2.5 ± 0.3 cm were used in all experiments. To isolate hemocytes, shrimp hemolymph was drawn from the ventral region above the first abdominal segment using a sterilized syringe preloaded with a commercial anticoagulant (Alsever’s solution, A3551, Sigma Aldrich, USA) in a 1:1 ratio, and then immediately centrifuged at 8000 × g for 15 min at 4 °C. Tissues were harvested by dissection. All the tissue samples were stored in RNAlater solution (Thermo Fisher Scientific, Schwerte, Germany) at -80 °C until use. Specific approval by the local institution/ethics committee was not required for the present study using invertebrate crustaceans and all experiment procedures were strictly conducted according to the guideline for the care and use of laboratory animals by the Animal Ethics Committee of Pukyong National University.
2.2. Immune challenge
The reared shrimps were randomly distributed into four 40 L experimental tanks with 4 groups, including three immune challenge groups and a control group, each containing 30 shrimps. For immune challenge experiments, 10 µL of polyinosinic-polycytidylic acid (poly I:C) (Sigma Aldrich, St. Louis, MO, USA), lipopolysaccharide (LPS) (Escherichia coli 0111: B4, Sigma Aldrich, St. Louis, MO, USA), and peptidoglycan (PGN) (Staphylococcus aureus, Sigma Aldrich, St. Louis, MO, USA) suspended in phosphate buffer saline (PBS, pH 7.4) at 100 mg/mL were injected into the shrimp abdominal segments Ⅲ and Ⅳ. As the control group, an equal volume of PBS was injected. Tissues (gill, heart, muscle, and stomach) were pooled from three shrimps randomly sampled at 0, 6, 12, 24, 48, and 72 hours post-injections (hpi) and stored in RNAlater solution (Thermo Fisher Scientific, Schwerte, Germany) at -80 °C until use. The immune challenge experiments were performed in triplicate.
2.3. Total RNA extraction and cDNA synthesis
Total RNA was extracted from RNAlater-stored tissues using an RNA extraction kit following the manufacturer’s protocol (Bioneer, Daejeon, Korea) including the DNA elimination step. Quantity and quality of the obtained total RNA were assessed using a spectrophotometer using Nanophotomer NP 80 (Implen, Munich, Germany) and the ratios of both 260 nm/280 nm and 260 nm/230 nm were confirmed to be at least higher than 1.9. Complementary DNA was synthesized through reverse transcription with oligo dT (dT18) using AccuPower® RT PreMix (Bioneer, Daejeon, Korea) according to the manufacturer’s instructions. The synthesized cDNA was stored at -80 °C for subsequent experiments.
2.4. Cloning of LvSOCS6 and LvSOCS7 cDNA
The nucleotide sequences for L. vannamei SOCS6 and SOCS7 (designated as LvSOCS6 and LvSOCS7) were firstly obtained from a homology search against the NCBI shrimp transcriptome shotgun assembly (TSA) database using orthologs proteins, EsSOCS6 (accession number: ATW63847.1) from the Chinese mitten crab (E. sinensis) and TmSOCS7 (accession number: QDL52635.1) from the Mealworm beetle (Tenebrio molitor), as queries [14,20]. For LvSOCS6 and LvSOCS7 a single set of primers was designed based on the retrieved TSA sequences (accession numbers: GETZ01043689.1 and GETZ01051062.1). (Table S1). PCR was performed using cDNA obtained from the hepatopancreas of 3 individuals as a template according to the following procedures: a cycle of 94 °C for 3 min; 35 cycles of 94°C for 30 s, 60°C for 30, and 72°C for 1 to 3 mins; and an extension at 72 °C for 10 min. The targeted PCR product was cloned into pTOP TA V2 vector (Enzynomics, Daejeon, Korea) and verified by sequencing.
2.5. In silico sequence analysis and molecular phylogeny
The protein sequences of LvSOCS6 and LvSOCS7 were deduced from the cloned sequences using the NCBI open reading frame (ORF) finder (https://www.ncbi.nlm.nih.gov/orffinder/) [21]. Domain architectures were visualized by Simple Modular Architecture Research Tool (SMART) (http://smart.embl-heidelberg.de/) and CDD blast program (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) [22,23]. Molecular weights and theoretical isoelectric point (pI) values of each LvSOCS were identified using the ExPASy pI/Mw tool (https://web.expasy.org/compute_pi/) [24]. Multiple sequence alignments of LvSOCS6 and LvSOCS7 with ortholog SOCSs were conducted using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) and refined manually [25]. In order to elucidate the evolutionary relationships between LvSOCS6 and LvSOCS7 and other homologs, molecular phylogeny was analyzed using full-length representative protein sequences in vertebrates and invertebrates available in the NCBI database. The phylogenetic tree was reconstructed using maximum likelihood (ML) methods with MEGA software (ver. 10.0.5; https://www.megasoftware.net/). The Jones-Taylor-Thornton (JTT) model was employed as a substitution model. The confidence of tree topology was tested with 1,000 bootstrap replicates [26].
2.6. Tissue distribution and expression pattern analysis
The tissue distribution of LvSOCS mRNAs was analyzed in triplicate in 7 tissues (gill, heart, hemocytes, hepatopancreas, lymphoid organs, muscle, and stomach) that were obtained by pooling from 3 individuals. Based on the results of the tissue distribution analysis, only the tissues (muscle, stomach, heart, and gill) with relatively high mRNA expression of LvSOCS6 and LvSOCS7 were selected for analysis of expression patterns changes following immune challenge. In addition, the expression patterns of STAT gene (GeneBank accession number: HQ228176.1) in L. vannamei were also examined in order to show the possible interconnection with the JAK-STAT signaling pathway according to changes in the mRNA expression patterns of LvSOCS6 and LvSOCS7 following immune challenge. To determine tissue distribution and expression changes in response to immune challenge, quantitative real-time PCR (RT-qPCR) was carried out using a LightCycler 480 Real-Time PCR System (Roche, Germany) with SYBR green premix (TOPreal qPCR 2X PreMix, Enzynomics, Daejeon, Korea). The primer pairs used for amplifying LvSOCS6, LvSOCS7, LvSTAT, and elongation factor 1α (EF1α) cDNA as a control for normalization were listed in Table S1. Relative expression was estimated based on the normalization of the expression level of each LvSOCS6, LvSOCS7, and LvSTAT to EF1α expression using the 2-ΔΔCT method [27].
2.7. Statistical analysis
The data were presented as means ± standard deviation. The statistical analysis for RT-qPCR data was performed using a one-way analysis of variance (ANOVA) supported by Turkey multiple comparison test using SPSS software (Version 25). P values less than 0.05 (p < 0.05) were considered statistically significant.
3. Results
3.1. Molecular characteristics and phylogenetic relationships of LvSOCS6 and LvSOCS7
The initial bioinformatics analysis, employing a tblastn search utilizing reference sequences of EsSOCS6 and TmSOCS7, yielded potential TSA sequences for LvSOCS. Based on these results, 2 identified cDNAs sequences encoding LvSOCS6 and LvSOCS7 were cloned, sequenced, and deposited to the GenBank database under accession numbers OR030046 and OR030047, respectively. The cloned nucleotide sequence of LvSOCS6 contained a length of 1465 bp, in which a 1,392 bp region encoding 463 amino acid (aa) protein with a calculated molecular weight of 49.84 kDa and a theoretical isoelectric point of 8.88, while the LvSOCS7 cDNA had a nucleotide sequence of 2994 bp in which 2868 bp region encoding 955 aa protein with a molecular weight of 103.9 kDa and a theoretical isoelectric point of 7.49 (Figure 1A, Figures S1 and S2). The presence of the SH2 domain and a SOCS box domain within the LvSOCS6 and LvSOCS7 protein was confirmed through CDD blast and SMART analysis. The LvSOCS6 protein had a conserved SH2 domain (Ala301 to Tyr 400) and SOCS box (Thr423 to Tyr463), meanwhile, the LvSOCS7 protein had a SH2 domain (Ala520 to Phe618), SOCS box domain (Lys641 to Glu687) and a comparatively extended C terminal region. Following, multiple sequence alignment revealed conserved phosphotyrosine and hydrophobic binding motifs within the SH2 domain, whereas the presence of a putative elongin B/C binding motif in the SOCS box domain in both LvSOCS proteins (Figures S3 and S4). Apparently, only LvSOCS6 exhibited a conserved pY site (Arg351 and Ser352) centralized in the SH2 domain which was previously reported in E. sinensis EsSOCS6 as well [14]. The estimated percentage sequence homologies showed that LvSOCS6 and LvSOCS7 proteins were similar to the invertebrate SOCS6 (69.3%-33.5%), SOCS7 (55.2%-19%), and in comparison, their vertebrate counterparts had a range of 51%-43.9% and 26.8%-15.4% sequence similarities to the aforementioned, respectively (Table S2).
To unravel the intricate evolutionary relationships of LvSOCS proteins, a comprehensive phylogenetic tree was constructed by integrating 48 distinct SOCS protein sequences. They were thoughtfully selected to encompass a diverse array of invertebrate taxa (including crustaceans, mollusks, and insects) as well as representative vertebrate taxa (including amphibians, fish, mammalians, and birds). Employing the maximum likelihood method-based Jones-Taylor-Thornton (JTT) model, the phylogenetic tree was derived, which offered the optimal framework for discerning the evolutionary dynamics at the protein level. The results unveiled topological disparities between two major lineages, designated as type I-SOCS and type II-SOCS (Figure 1B). Within these lineages, a remarkable degree of differentiation was observed, giving rise to distinct SOCS subtypes, namely SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, and SOCS7. Notably, the identified LvSOCS6 and LvSOCS7 were assigned to the established invertebrate type I-SOCS6 and SOCS7 groups, respectively, based on their positioning within the phylogenetic tree. In contrast, the LvSOCS6 was phylogenetically rooted with the previously reported EsSOCS6 of the Chinese mitten crab, and, LvSOCS7 displayed a robust evolutionary connection to theSnow crab SOCS7, thereby corroborating their close affiliation within this taxonomic group.
3.2. Tissue distribution and mRNA expression of LvSOCS6, LvSOCS7, and LvSTAT after LPS, poly (I:C), and PGN stimulation
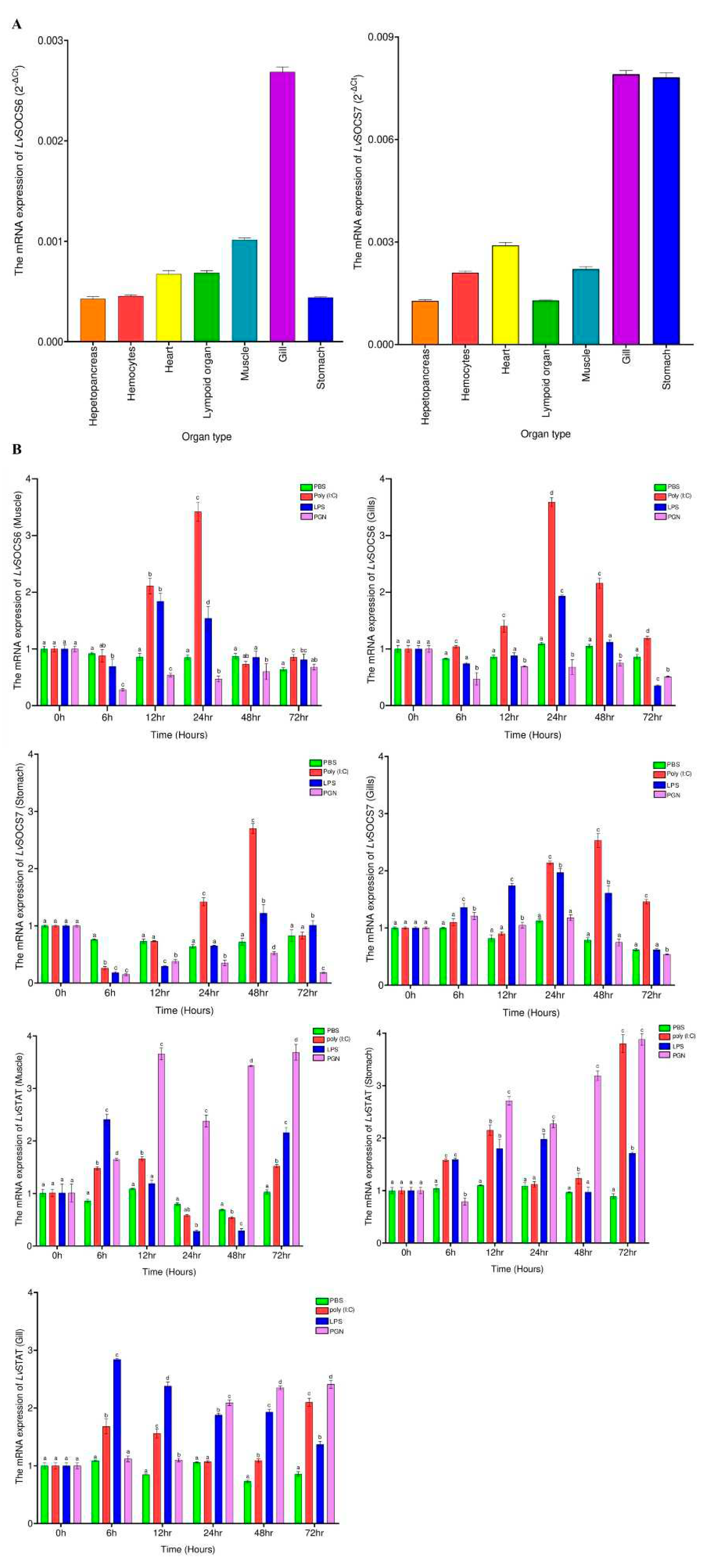
To gain insights into the intricate patterns of LvSOCS6 and LvSOCS7 expression across various tissues, quantitative reverse transcription PCR (qRT-PCR) was conducted on the hepatopancreas, hemocytes, heart, lymphoid organ, muscle, gills, and stomach (Figure 2A). Notably, both LvSOCS6 and LvSOCS7 exhibited detectable expression levels in all examined tissues. Among them, gills showed the most prominent expression of LvSOCS6, followed by elevated expression levels in muscle, lymphoid organ, heart, hemocytes, and stomach. On the other hand, LvSOCS7 exhibited distinguishable mRNA expressions in gills and stomach than that for LvSOCS6. The heart also demonstrated noteworthy expression for LvSOCS7, in comparison to its levels in hepatopancreas, hemocytes, lymphoid organ, and muscle.
The present study further investigated the mRNA expression patterns of LvSOCS6, LvSOCS7, and LvSTAT in multiple tissues following immune induction. Initially, LvSOCS6 expression exhibited a significant decrease across all experimented tissues at 6 hours post-immune stimulation in all experimental groups. However, subsequent induction with LPS and poly(I:C) led to a notable increase in LvSOCS6 expression after 6 hours in all examined tissues. The gills and muscle displayed the highest expression levels of LvSOCS6 at 24 hours, with 3.59-fold and 3.42-fold increases, and in heart, it was peaked at 48 hours (2.54-fold, P < 0.05) after poly(I:C) stimulation. LvSOCS6 also showed elevated expression in the gills, muscle, and heart at 24 hours, 12 hours, and 48 hours, respectively, following LPS induction (1.93-fold, 1.84-fold, and 1.62-fold, P < 0.05). Interestingly, the PGN induction consistently resulted in decreased LvSOCS6 expression compared to other experimental groups. Subsequently, the LvSOCS6 expression declined below 1.5-fold in LPS and poly(I:C)-stimulated groups at 72 hours. In the case of LvSOCS7, its expression significantly decreased in the heart and stomach at 6 hours post-immune stimulation, while showing an increase only in the gills following LPS and PGN induction. Following, LvSOCS7 expression reached its highest levels in the gills, stomach, and heart at 48 hours after poly(I:C) stimulation, with fold increases of 2.53, 2.7, and 2.09, respectively, compared to the control group (P < 0.05). Peak expressions of LvSOCS7 were observed at 24 hours and 48 hours in response to LPS induction, with fold increases of 1.97, 1.82, and 1.22 in the gills, heart, and stomach (P < 0.05). LvSOCS7 expression increased in the heart at 24 hours following PGN stimulation; However, overall, LvSOCS7 expression gradually decreased after 48 hours in all the experimental groups (Figure 2B and Figure S5).
Contrasting the LvSOCS genes, the LvSTAT gene, involved in the JAK-STAT immune pathway, exhibited significantly higher expression at 6 hours in all examined tissues. Stimulation with PGN resulted in a progressive increase in LvSTAT expression, peaking at 12 hours, 72 hours, and 24 hours in muscle, stomach or gills, and heart, respectively, with fold increases of 3.62, 3.88 or 2.41, and 1.47 compared to the control group (P < 0.05). In the LPS-stimulated group, the highest LvSTAT expression was observed at 6 hours in the gills, muscle, and heart, while in the stomach, it peaked at 24 hours, with fold increases of 2.84, 2.39, 1.78, and 1.98 (P < 0.05). The poly(I:C)-stimulated group exhibited higher LvSTAT expressions at 72 hours in the stomach and gills (3.8-fold and 2.1-fold, P < 0.05), while in the muscle and heart, the peaks occurred at 12 hours (1.64-fold and 2.24-fold, P < 0.05). Overall, LvSTAT demonstrated higher immune expression in all examined tissues, concomitant with a decrease in LvSOCS gene expression (Figure 2B and Figure S5).
4. Discussion
Cytokines, the secreted polypeptide molecules, exert significant influence over a myriad of biological processes encompassing cellular proliferation, growth, and immune regulation in both vertebrates and invertebrates [2,8,19,20]. Among the diverse array of molecules involved in cytokine signaling, the suppressor of cytokine signaling (SOCS) molecules possess noteworthy significance due to their crucial role as principal antagonists of these signaling mediators [7]. Within the SOCS gene family, a dichotomy emerges, comprising the type II SOCS and type I SOCS groups (consisting of SOCS1, SOCS2, SOCS3, SOCS4, SOCS5, SOCS6, and SOCS7) [1,8,12]. In contrast, the presence and comprehensive characterization of these SOCS genes, and their immunoregulatory involvement in vertebrates has reached to a well-documented stage. Nonetheless, the existence and potential functionalities of SOCS genes in invertebrates, particularly within the domain of crustaceans, lack substantive evidence. Within the confines of this investigation, two previously unreported type I SOCS genes were successfully identified in the popular food crustacean “white leg shrimps”, designated as LvSOCS6 and LvSOCS7. The characterized LvSOCS genes in the present study exhibited the archetypal, centrally localized Src homology 2 (SH2) domain region, concomitant with the C-terminal suppressor of cytokine signaling (SOCS)-box domain. Furthermore, there were 3 distinct motifs, encompassing the phosphotyrosine binding, hydrophobic binding, and putative elongin B/C binding, which are renowned for their involvement in mediating protein-protein interactions and modulating diverse signaling pathways [28,29]. Additionally, the investigation of phylogenetic relationships among SOCS proteins provided valuable insights into their evolutionary history and functional diversification of these identified LvSOCS. We employed the maximum likelihood method to construct a phylogenetic tree, which integrated a comprehensive set of SOCS protein sequences from both vertebrate and invertebrate organisms. The resulting phylogenetic tree uncovered significant topological distinctions, giving rise to 2 major lineages as type I-SOCS and type II-SOCS. Within the type II-SOCS lineage, further differentiation was observed, manifesting as distinct sub-branches that corresponded to specific members of the SOCS1, SOCS2, and SOCS3. Similarly, the type I-SOCS lineage exhibited its own differentiation, generating representative sub-branches associated with SOCS4, SOCS5, SOCS6, and SOCS7. Thus it suggests that SOCS proteins have undergone lineage-specific evolutionary changes, and emphasizes the divergent evolutionary trajectories and their unique functional attributes [8]. Importantly, our analysis also included the LvSOCS6 and LvSOCS7 identified in this study, which were assigned to the invertebrate SOCS6 and SOCS7 groups in the phylogeny. The placement of LvSOCS6 and LvSOCS7 within their respective invertebrate sub-branches of the phylogenetic tree supports their classification and highlights their evolutionary relationships with other known SOCS proteins.
In white leg shrimps, the identified LvSOCS mRNAs exhibited a pervasive presence across all analyzed tissues, showcasing noteworthy fluctuations in expression levels. These dynamic variations potentially might be aligned with the multifaceted biological functionalities intrinsic to shrimps. Comparable observations have been discerned in other invertebrate organisms, such as the Chinese mitten crab, where distinct levels of EsSOCS6 gene expression were detected in the hepatopancreas and hemopoietic tissues, displaying differential patterns of higher and lower expression [14]. Likewise, in mealworms, TmSOCS6 expression had its peak in hemocytes, whereas TmSOCS7 expression elevated in Malpighian tubules [20]. Akin to these findings, the silk moth (B. mori) exhibited heightened BmSOCS6 expression within the fat body in contrast to other tissues [30]. In the realm of vertebrates, such as fish species, the expression profiles of SOCS genes have also been observed to fluctuate across different tissue types. For example, in rainbow trout (O. mykiss), OmSOCS6 expression levels showcased variation in the skin and gills [31]. In our investigation, both LvSOCS genes manifested significant expression in the gills, with the intriguing discovery that LvSOCS7 exhibited prominent expression levels also in the stomach. These fascinating findings tentatively suggest the potential existence of distinct and organ-specific roles fulfilled by the identified LvSOCS genes, necessitating further experimental validation and confirmation.
The JAK/STAT signaling cascade plays a pivotal role in orchestrating immune responses across the animal kingdom, primarily driven by an array of cytokines [4,14]. Extensive research has elucidated the significance of type I SOCS genes in vertebrates, notable examples include the involvement of SOCS7 in the translocation of the STAT3 gene, a critical transcriptional regulator of IFN-β and interleukin 6 in humans [32]. Furthermore, investigations have unveiled potential connections between the JAK/STAT pathway and neural cell differentiation in humans, with SOCS6 emerging as an essential participant in these intricate processes [33]. In reference to invertebrates, studies involving d. melanogaster has shed light on the regulatory role of SOCS36E, SOCS44A, and SOCS16D (analogous to SOCS5, SOCS6, and SOCS7) in dampening the JAK-STAT pathway by influencing its regulation [15,16]. Nevertheless, the current body of knowledge regarding the involvement of type I SOCS genes in the JAK/STAT pathway of crustaceans remains limited, warranting further investigation, to unravel their contributions within this extensive signaling network. Hence, to further explore the regulatory association of LvSOCS and LvSTAT genes, we investigated their mRNA expression in response to the administration of pathogen-associated molecular patterns (PAMPs), including poly(I:C), LPS, and PGN. These PAMPs are known to induce inflammatory and immune regulatory responses (mimicking the activities of viruses, gram negative and/or gram positive bacteria), activating key signaling pathways such as JAK-STAT and NF-kB [19,31,34]. Additionally, they are commonly employed as stimulants to study immune response patterns. In our comprehensive investigation, intriguing observations emerged when examining the intricate interplay between LvSOCS6, LvSOCS7, and LvSTAT gene expression dynamics. First, our immune stimulation experiments revealed a significant increase in LvSOCS expression following poly(I:C) and LPS administration peaking at or after 12 hours, but not following the PGN administration. However, the highest significant increase in LvSOCS6 and LvSOCS7 expression was observed in poly(I:C) administrated group (3.59 and 2.7-fold), reinforcing that these identified genes might have enhanced sensitivity to viral infections than bacterial infections. Secondly, the conspicuous downregulation of LvSOCS expression coincided with the upregulation of LvSTAT mRNA levels. Although the precise temporal pattern of this response was not uniform across all assessed time points, it unequivocally implies a noteworthy correlation between diminished LvSOCS expression and the modulation of LvSTAT gene activity. It is important to acknowledge that other members of the SOCS gene family, such as LvSOCS2, have also been implicated in the multifaceted JAK/STAT pathway, thereby potentially contributing to the current regulatory network at play [19]. Moreover, given the existing evidence elucidating the involvement of type I SOCS genes in the NF-кB pathway, a comprehensive exploration of their sophisticated roles within the immune machinery of crustaceans becomes imperative, necessitating in-depth investigations to unravel their precise contributions [14,17]. Consequently, understanding the regulatory mechanisms of SOCS genes and their involvement in immune pathways will significantly augment our comprehension of crustacean immunology and make notable contributions to the wider domain of invertebrate immune modulation.
5. Conclusions
In synopsis, the current investigation successfully identified novel members of the type I suppressor of cytokine signaling (SOCS) gene family, namely LvSOCS6 and LvSOCS7, in white leg shrimp, thereby illuminating their involvement in the intricate orchestration of immune regulatory mechanisms. The mRNA expression patterns of LvSOCS6 and LvSOCS7 demonstrated a pervasive presence throughout diverse tissue types, with noteworthy augmented levels observed particularly within the gills, while LvSOCS7 exhibited conspicuous prominence in the stomach. Additionally, the administration of poly(I:C) and LPS elicited a significant induction of LvSOCS mRNA transcripts, underscoring their heightened responsiveness to immune stimulation mediated by viral and bacterial agents. Intriguingly, this immune activation coincided with a substantial decrement in LvSTAT expression, implying a potential intricate interplay between these genes in the context of immune modulation. However, to further unravel their genomic architectures, specific roles, and underlying mechanisms of LvSOCS within the immune pathways of white leg shrimp, investigations employing RACE PCR and RNA interference (RNAi)-based methodology are imperative. As such, we believe the findings of the present study establish a solid groundwork for prospective investigations aimed at deciphering the intricacies of immune regulation in invertebrates, particularly in crustaceans.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The qPCR data of this article can be found in supplementary files (Table S3).
Author contributions
C.W.R: Conceptualization, Methodology, Investigation, Data curation, Formal analysis, Writing—original draft, W.S.P: Investigation, Methodology, Validation, Writing. J.L: Investigation. C.H.K: Writing—review & editing, Supervision, Resources.
Institutional Review Board Statement
All experiment procedures were strictly conducted according to the guideline for the care and use of laboratory animals by the Animal Ethics Committee of Pukyong National University.
Data availability statement
The sequences of LvSOCS6 and LvSOCS7 were submitted to NCBI with GenBank accession numbers OR030046, and OR030047.
Acknowledgment
This research received support from the National research foundation of Korea [grant number: 2022R1I1A1A01073372], a grant funded by the Korean government.
Declaration of competing interest
All authors have read and agreed to the present version of the manuscript and declared no conflict of interest.
References
- Beschin, A.; Bilej, M.; Torreele, E.; De Baetselier, P. On the existence of cytokines in invertebrates. Cell. Mol. Life Sci. 2001, 58, 801–814. https://doi.org/10.1007/PL00000901. [CrossRef]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology (Basel). 2016, 5. https://doi.org/10.3390/biology5020023. [CrossRef]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. https://doi.org/10.1002/pro.3519. [CrossRef]
- Ortmann, R.A.; Cheng, T.; Visconti, R.; Frucht, D.M.; O’Shea, J.J. Janus kinases and signal transducers and activators of transcription: Their roles in cytokine signaling, development and immunoregulation. Arthritis Res. 2000, 2, 16–32. https://doi.org/10.1186/ar66. [CrossRef]
- Boudinot, P.; Bird, S.; Du Pasquier, L.; Collet, B. The repertoire of vertebrate STAT transcription factors: Origin and variations in fish. Dev. Comp. Immunol. 2021, 116, 103929. https://doi.org/10.1016/j.dci.2020.103929. [CrossRef]
- Hou, S.X.; Zheng, Z.; Chen, X.; Perrimon, N. The JAK/STAT Pathway in model organisms: Emerging roles in cell movement. Dev. Cell 2002, 3, 765–778. https://doi.org/10.1016/S1534-5807(02)00376-3. [CrossRef]
- Alexander, W.S.; Hilton, D.J. The role of suppressors of cytokine signaling (SOCS) proteins in regulation of the immune response. Annu. Rev. Immunol. 2004, 22, 503–529. https://doi.org/10.1146/annurev.immunol.22.091003.090312. [CrossRef]
- Wang, B.; Wangkahart, E.; Secombes, C.J.; Wang, T. Insights into the evolution of the suppressors of cytokine signaling (SOCS) Gene Family in vertebrates. Mol. Biol. Evol. 2019, 36, 393–411. https://doi.org/10.1093/molbev/msy230. [CrossRef]
- Delgado-Ortega, M.; Marc, D.; Dupont, J.; Trapp, S.; Berri, M.; Meurens, F. SOCS proteins in infectious diseases of mammals. Vet. Immunol. Immunopathol. 2013, 151, 1–19. https://doi.org/10.1016/j.vetimm.2012.11.008. [CrossRef]
- Yasukawa, H.; Misawa, H.; Sakamoto, H.; Masuhara, M.; Sasaki, A.; Wakioka, T.; Ohtsuka, S.; Imaizumi, T.; Matsuda, T.; Ihle, J.N.; et al. The JAK-binding protein JAB inhibits janus tyrosine kinase activity through binding in the activation loop. EMBO J. 1999, 18, 1309–1320. https://doi.org/10.1093/emboj/18.5.1309. [CrossRef]
- Sasaki, A.; Yasukawa, H.; Suzuki, A.; Kamizono, S.; Syoda, T.; Kinjyo, I.; Sasaki, M.; Johnston, J.A.; Yoshimura, A. Cytokine-inducible SH2 protein-3 (CIS3/SOCS3) inhibits janus tyrosine kinase by binding through the N-terminal kinase inhibitory region as well as SH2 domain. Genes to Cells 1999, 4, 339–351. https://doi.org/10.1046/j.1365-2443.1999.00263.x. [CrossRef]
- Dalpke, A.; Heeg, K.; Bartz, H.; Baetz, A. Regulation of innate immunity by suppressor of cytokine signaling (SOCS) proteins. Immunobiology 2008, 213, 225–235. https://doi.org/10.1016/j.imbio.2007.10.008. [CrossRef]
- Feng, Z.P.; Chandrashekaran, I.R.; Low, A.; Speed, T.P.; Nicholson, S.E.; Norton, R.S. The N-Terminal domains of SOCS proteins: A conserved region in the disordered N-termini of SOCS4 and 5. Proteins Struct. Funct. Bioinforma. 2012, 80, 946–957. https://doi.org/10.1002/prot.23252. [CrossRef]
- Qu, C.; Xu, Q.; Lu, M.; Wang, F.; Liu, Z.; Liu, D.; Yang, W.; Yi, Q.; Wang, L.; Song, L. The involvement of suppressor of cytokine signaling 6 (SOCS6) in immune response of chinese mitten crab Eriocheir Sinensis. Fish Shellfish Immunol. 2018, 72, 502–509. https://doi.org/10.1016/j.fsi.2017.11.029. [CrossRef]
- Callus, B.A.; Mathey-Prevot, B. SOCS36E, a novel Drosophila SOCS protein, suppresses JAK/STAT and EGF-R signalling in the imaginal wing disc. Oncogene 2002, 21, 4812–4821. https://doi.org/10.1038/sj.onc.1205618. [CrossRef]
- Rawlings, J.S.; Rennebeck, G.; Harrison, S.M.W.; Xi, R.; Harrison, D.A. Two Drosophila suppressors of cytokine signaling (SOCS) differentially regulate JAK and EGFR pathway activities. BMC Cell Biol. 2004, 5, 1–15. https://doi.org/10.1186/1471-2121-5-38. [CrossRef]
- Li, J.; Zhang, Y.; Zhang, Y.; Liu, Y.; Xiang, Z.; Qu, F.; Yu, Z. Cloning and characterization of three suppressors of cytokine signaling (SOCS) genes from the pacific oyster, Crassostrea Gigas. Fish Shellfish Immunol. 2015, 44, 525–532. https://doi.org/10.1016/j.fsi.2015.03.022. [CrossRef]
- Zhu, B.; Dai, L.; Yu, Y.; Wang, D.; Peng, T.; Liu, C. A role of suppressor of cytokine signaling 2 in the regulation of ecdysteroid signaling pathway in Procambarus Clarkii. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2016, 325, 441–452. https://doi.org/10.1002/jez.2029. [CrossRef]
- Wang, S.; Song, X.; Zhang, Z.; Li, H.; Lǚ, K.; Yin, B.; He, J.; Li, C. Shrimp with knockdown of LvSOCS2, a negative feedback loop regulator of JAK/STAT pathway in Litopenaeus Vannamei, exhibit enhanced resistance against WSSV. Dev. Comp. Immunol. 2016, 65, 289–298. https://doi.org/10.1016/j.dci.2016.07.021. [CrossRef]
- Patnaik, B.B.; Kim, B.B.; Jo, Y.H.; Bang, I.S. Molecular cloning and expression analysis of three suppressors of cytokine signaling genes (SOCS5, SOCS6, SOCS7) in the mealworm beetle Tenebrio Molitor. Insects 2019, 10. https://doi.org/10.3390/insects10030076. [CrossRef]
- Wheeler, D.L.; Barrett, T.; Benson, D.A.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; Dicuccio, M.; Edgar, R.; Federhen, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2008, 36, 13–21. https://doi.org/10.1093/nar/gkm1000. [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A conserved domain database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, 225–229. https://doi.org/10.1093/nar/gkq1189. [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. https://doi.org/10.1093/nar/gkaa937. [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press, 2005; pp. 571–607.
- Sievers, F.; Higgins, D.G. Clustal omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. https://doi.org/10.1002/pro.3290. [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. https://doi.org/10.1093/molbev/msab120. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. https://doi.org/10.1006/meth.2001.1262. [CrossRef]
- Kamura, T.; Sato, S.; Haque, D.; Liu, L.; Kaelin, W.G.; Conaway, R.C.; Conaway, J.W. The elongin BC complex interacts with the conserved SOCS-Box motif present in members of the SOCS, ras, WD-40 repeat, and Ankyrin repeat families. Genes Dev. 1998, 12, 3872–3881. https://doi.org/10.1101/gad.12.24.3872. [CrossRef]
- Bullock, A.N.; Rodriguez, M.C.; Debreczeni, J.É.; Songyang, Z.; Knapp, S. Structure of the SOCS4-ElonginB/C complex reveals a distinct SOCS box interface and the molecular basis for SOCS-dependent EGFR degradation. Structure 2007, 15, 1493–1504. https://doi.org/10.1016/j.str.2007.09.016. [CrossRef]
- Abbas, M.N.; Kausar, S.; Sun, Y.X.; Tian, J.W.; Zhu, B.J.; Liu, C.L. Suppressor of cytokine signaling 6 can enhance epidermal growth factor receptor signaling pathway in Bombyx Mori (Dazao). Dev. Comp. Immunol. 2018, 81, 187–192. https://doi.org/10.1016/j.dci.2017.12.003. [CrossRef]
- Wang, T.; Gao, Q.; Nie, P.; Secombes, C.J. Identification of suppressor of cytokine signalling (SOCS) 6, 7, 9 and CISH in rainbow trout Oncorhynchus Mykiss and analysis of their expression in relation to other known trout SOCS. Fish Shellfish Immunol. 2010, 29, 656–667. https://doi.org/10.1016/j.fsi.2010.06.015. [CrossRef]
- Noguchi, S.; Yamada, N.; Kumazaki, M.; Yasui, Y.; Iwasaki, J.; Naito, S.; Akao, Y. Socs7, a target gene of microRNA-145, regulates interferon-β induction through STAT3 nuclear translocation in bladder cancer cells. Cell Death Dis. 2013, 4, e482-9. https://doi.org/10.1038/cddis.2013.11. [CrossRef]
- Gupta, S.; Mishra, K.; Surolia, A.; Banerjee, K. Suppressor of cytokine signalling-6 promotes neurite outgrowth via JAK2/STAT5-mediated signalling pathway, involving negative feedback inhibition. PLoS One 2011, 6. https://doi.org/10.1371/journal.pone.0026674. [CrossRef]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. Jak-Stat signaling: A double-edged sword of immune regulation and cancer progression. Cancers (Basel). 2019, 11. https://doi.org/10.3390/cancers11122002. [CrossRef]
Figure 1.
(A) The domain architectures of LvSOCS6 and LvSOCS7. The predicted SH2 and SOCS-box positions are shown in red and blue color boxes respectively. (B) Mega software-based maximum likelihood tree of 48 SOCS protein sequences among vertebrates and invertebrates. The numbers at tree nodes refer to the percent bootstrap values following 1000 replications. The species and the GenBank accession numbers used for the phylogenetic analysis were as follows: The type I family includes SOCS1 from D. rerio (NP_001003467.1), H. sapiens (NP_003736.1), M. musculus (NP_034026.1), G. gallus (NP_001131120.1), X. tropicalis (NP_001011327.1), O. niloticus (NP_001297020.1), SOCS2 from D. rerio (XP_005164804.1), H. sapiens (NP_001257400.1), M. musculus (NP_031732.1), G. gallus (NP_989871.1), X. tropicalis (NP_001120898.1), O. niloticus (XP_036813373.1), C. gigas (EKC24772.1), L. japonicus (BAI70368.1), E. sinensis (ACU42699.1), SOCS3 from D. rerio (NP_998469.1), H. sapiens (NP_003946.3), M. musculus (NP_031733.1), G. gallus (NP_001186037.1), X. tropicalis (XP_031746340.1), O. mykiss (NP_001139640.1), SOCS4 from H. sapiens (NP_659198.1), M. musculus (NP_543119.2), G. gallus (NP_001186037.1), X. tropicalis (XP_031746340.1). The type II family includes SOCS5 from H. sapiens (NP_955453.1), M. musculus (NP_062628.2), G. gallus (NP_001120786.1), X. tropicalis (NP_001016844.1) and D. rerio (ABM68036.1). SOCS6 from H. sapiens (NP_004223.2), M. musculus (NP_061291.2), G. gallus (NP_001120784.1), X. tropicalis (NP_001096240.1), O. mykiss (NP_001182102.1), E. sinensis (ATW63847.1), L. vannamei* (SOCS6-OR030046), T. castaneum (XP_008190646.1), SOCS7 from H. sapiens (NP_055413.2), M. musculus (NP_619598.2), G. gallus (XP_040509254.1), X. tropicalis (XP_012827004.2) D. rerio (XP_009304138.1) O. mykiss (CAP17279.1), T. molitor (QDL52635.1), C. gigas (AKA59677.1), Chionoecetes opilio (KAG0725835.1) and L. vannamei* (SOCS7- OR030047). The blue colors indicate vertebrate species, purple and green colors indicate invertebrates, and the red box highlights the newly identified SOCS6/SOCS7 for L. vannamei.
Figure 1.
(A) The domain architectures of LvSOCS6 and LvSOCS7. The predicted SH2 and SOCS-box positions are shown in red and blue color boxes respectively. (B) Mega software-based maximum likelihood tree of 48 SOCS protein sequences among vertebrates and invertebrates. The numbers at tree nodes refer to the percent bootstrap values following 1000 replications. The species and the GenBank accession numbers used for the phylogenetic analysis were as follows: The type I family includes SOCS1 from D. rerio (NP_001003467.1), H. sapiens (NP_003736.1), M. musculus (NP_034026.1), G. gallus (NP_001131120.1), X. tropicalis (NP_001011327.1), O. niloticus (NP_001297020.1), SOCS2 from D. rerio (XP_005164804.1), H. sapiens (NP_001257400.1), M. musculus (NP_031732.1), G. gallus (NP_989871.1), X. tropicalis (NP_001120898.1), O. niloticus (XP_036813373.1), C. gigas (EKC24772.1), L. japonicus (BAI70368.1), E. sinensis (ACU42699.1), SOCS3 from D. rerio (NP_998469.1), H. sapiens (NP_003946.3), M. musculus (NP_031733.1), G. gallus (NP_001186037.1), X. tropicalis (XP_031746340.1), O. mykiss (NP_001139640.1), SOCS4 from H. sapiens (NP_659198.1), M. musculus (NP_543119.2), G. gallus (NP_001186037.1), X. tropicalis (XP_031746340.1). The type II family includes SOCS5 from H. sapiens (NP_955453.1), M. musculus (NP_062628.2), G. gallus (NP_001120786.1), X. tropicalis (NP_001016844.1) and D. rerio (ABM68036.1). SOCS6 from H. sapiens (NP_004223.2), M. musculus (NP_061291.2), G. gallus (NP_001120784.1), X. tropicalis (NP_001096240.1), O. mykiss (NP_001182102.1), E. sinensis (ATW63847.1), L. vannamei* (SOCS6-OR030046), T. castaneum (XP_008190646.1), SOCS7 from H. sapiens (NP_055413.2), M. musculus (NP_619598.2), G. gallus (XP_040509254.1), X. tropicalis (XP_012827004.2) D. rerio (XP_009304138.1) O. mykiss (CAP17279.1), T. molitor (QDL52635.1), C. gigas (AKA59677.1), Chionoecetes opilio (KAG0725835.1) and L. vannamei* (SOCS7- OR030047). The blue colors indicate vertebrate species, purple and green colors indicate invertebrates, and the red box highlights the newly identified SOCS6/SOCS7 for L. vannamei.
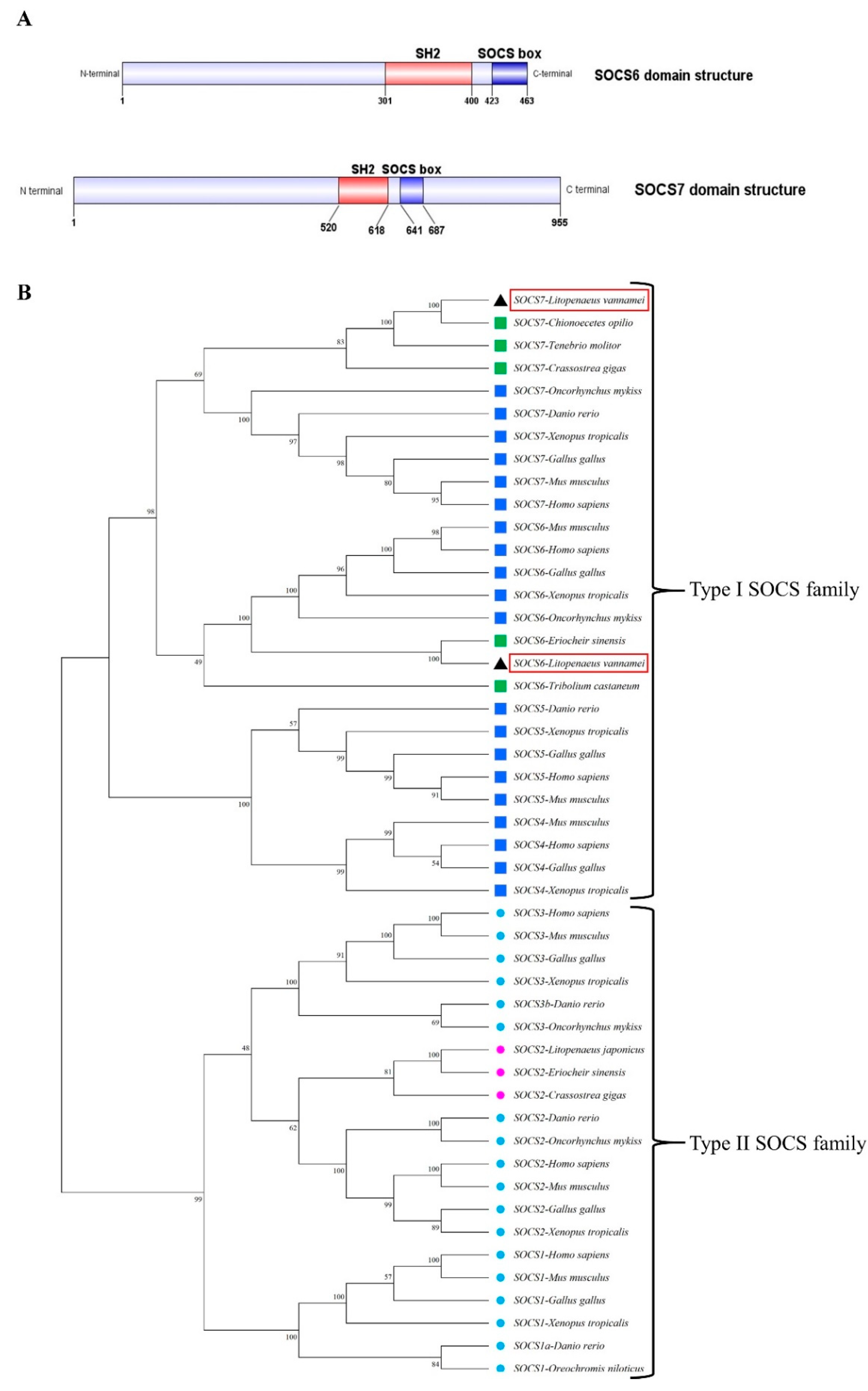
Figure 2.
(A) The tissue distribution patterns of LvSOCS6 and LvSOCS7 in different tissues following qPCR analysis. The data is expressed using the 2 (-ΔCt). (B) The temporal mRNA expression analysis of LvSOCS6 (Muscle, Gills) LvSOCS7 (Stomach, Gills), and LvSTAT (Muscle, Stomach, and Gills) following challenge experiments with LPS, poly (I:C), PGN, or phosphate-buffered saline (PBS). All the qPCR data were normalized to EF1α internal control gene. Results are represented as the mean ± S.E (N = 3). Statistically significant values (P < 0.05) are denoted with alphabets.
Figure 2.
(A) The tissue distribution patterns of LvSOCS6 and LvSOCS7 in different tissues following qPCR analysis. The data is expressed using the 2 (-ΔCt). (B) The temporal mRNA expression analysis of LvSOCS6 (Muscle, Gills) LvSOCS7 (Stomach, Gills), and LvSTAT (Muscle, Stomach, and Gills) following challenge experiments with LPS, poly (I:C), PGN, or phosphate-buffered saline (PBS). All the qPCR data were normalized to EF1α internal control gene. Results are represented as the mean ± S.E (N = 3). Statistically significant values (P < 0.05) are denoted with alphabets.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



