
Submitted:
27 July 2023
Posted:
31 July 2023
You are already at the latest version
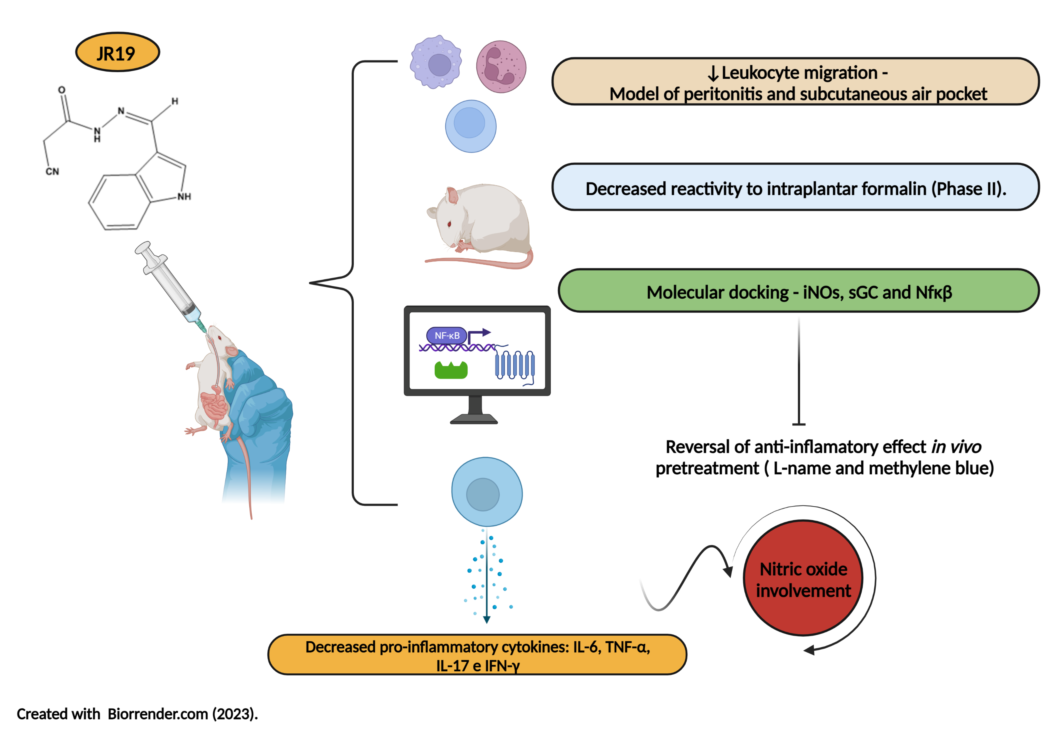
Abstract
The N-acylhydrazone function has been reported as a pharmacophore group of molecules with diverse pharmacological activities, including anti-inflammatory effects. Thus, this study evaluated the anti-inflammatory potential of the compound N'-(3-(1H-indol-3-yl)benzylidene)-2-cyanoacetohydrazide (JR19) in vivo. The study started with the carrageenan-induced peritonitis model, followed by an investigation of leukocyte migration using the subcutaneous air pouch test and an assessment of the antinociceptive profile using formalin-induced pain. To find out the likely mechanism of action, a preliminary molecular docking study was performed focusing on the crystallographic structures of NFκB, iNOS, and sGCs. The computational study revealed satisfactory interaction energies with the selected targets, and the same peritonitis model was used to validate the involvement of the nitric oxide pathway and cytokine expression in the peritoneal exudate of mice pretreated with L-NAME or methylene blue. In the peritonitis assay, JR19 (10 and 20 mg/kg) reduced leukocyte migration by 59% and 52%, respectively, compared to the control group, with the 10 mg/kg dose used in subsequent assays. In the subcutaneous air pouch assay, the reduction in cell migration was 66%, and the response to intraplantar formalin was reduced by 39%, particularly during the inflammatory phase, suggesting that the compound lacks central analgesic activity. In addition, a reversal of the anti-inflammatory effect was observed in mice pretreated with L-NAME or methylene blue, indicating the involvement of iNOS and sGC in the anti-inflammatory response of JR19. The compound effectively and significantly decreased the levels of IL-6, TNF-α, IL-17, and IFN-γ, and this effect was reversed in animals pretreated with L-NAME, supporting a NO-dependent anti-inflammatory effect. In contrast, pretreatment with methylene blue only reversed the reduction in TNF-α levels. Therefore, these results demonstrate the pharmacological potential of the novel N-acylhydrazone derivative, which acts through the nitric oxide pathway and cytokine signaling, making it a strong candidate as an anti-inflammatory and immunomodulatory agent.
Keywords:
cytokines
; leukocyte migration
; N-acylhydrazones
; nitric oxide
1. Introduction
The inflammatory process is defined as a natural response of the immune system to noxious stimuli affecting cells or tissues, triggered by chemical, physical or biological agents [1,2]. When activated excessively and persistently, this process can lead to organ and system damage associated with the pathogenesis of various diseases, including autoimmune diseases, cancers, musculoskeletal, gastrointestinal, and psychoemotional disorders, as well as worsen the prognosis of viral infections, thus contributing to healthcare costs [3,4,5].
The pathophysiology of inflammation involves vasodilation resulting from the release of mediators by leukocytes that migrate from the blood to the site of injury, leading to characteristic symptoms such as edema, pain, and redness. If left unchecked, this process can lead to loss of organ or tissue function [6]. Effector mechanisms of the immune system include the expression of cytokines, eicosanoids, vasoactive amines, and nitric oxide, which play a role in signaling and activating vascular endothelial cells and higher regulatory adhesion molecules that mediate leukocyte rolling, cellular adhesion, and extravasation to the site of inflammation, actively contributing to the cellular response [2].
As mentioned above, an important mediator involved in the genesis of inflammation is nitric oxide (NO), a small signaling molecule whose functions are primarily regulated by the expression and activity of nitric oxide synthase (NOS) enzymes: neuronal (nNOS), endothelial (eNOS), or inducible (iNOS), which synthesize NO by converting L-arginine to L-citrulline [7,8]. Due to its anti-inflammatory and pro-inflammatory properties, the role of NO remains unclear; however, iNOS expression is known to result from inflammatory responses [8].
In localized infections and sepsis, iNOS expression is associated with the inhibition of pathogen growth and platelet aggregation and the NO pathway is involved in peripheral analgesia in primary sensory neurons [7,8]. In addition, iNOS can be transcriptionally stimulated by the activation of TLRs in response to PAMPs, where biosynthesized NO exerts its vasoactive function in acute inflammatory responses accompanied by leukocyte migration throughout the process [9].
Another critical event in the pathophysiology of inflammation is the biosynthesis and release of prostaglandins (PGs). The pharmacological action of classical nonsteroidal anti-inflammatory drugs (NSAIDs) is to suppress the biosynthesis of these mediators from arachidonic acid by selective or non-selective inhibition of cyclooxygenase (COX) enzymes [2,10]. Compared with steroidal anti-inflammatory drugs (SAIDs), long-term use of NSAIDs is considered safer, and they are among the most commonly used drugs, with a place in the World Health Organization Model List of Essential Medicines [11].
Non-selective NSAIDs in conventional therapy, although effective, have undesirable adverse effects on the gastrointestinal (GI) tract, renal and hepatic systems due to the constitutive expression of cyclooxygenase 1 (COX-1) in the body, which serves as a source of cytoprotective PGs for the GI tract [8,11]. Therefore, the development of drugs with selective inhibition of cyclooxygenase 2 (COX-2) has been proposed, assuming greater therapeutic advantages since it is an isoform induced in response to the onset of an inflammatory process and associated with various pathological dysfunctions [12]. However, these drugs may have serious cardiovascular side effects due to the suppression of cardioprotective PGs derived from COX-2 [13].
Thus, the critical role of iNOS and COX-2 enzymes and cytokines in the initiation and progression of inflammation is evident. Therefore, drugs capable of modulating these targets and associated with a low toxicity profile are essential as potential pharmacological agents for new anti-inflammatory therapies [14,15]. In this context, acylhydrazone derivatives stand out as a class of synthetic chemical structures with diverse bioactive activities, among which the N-acylhydrazone function has shown relevant anti-inflammatory and analgesic responses [14]. Therefore, this study aims to evaluate the in vivo anti-inflammatory potential of the compound N'-(3-(1H-indol-3-yl)benzylidene)-2-cyanoacetohydrazide (JR19) and to perform in silico and in vivo investigations of possible mechanisms of action involved in the anti-inflammatory response regarding cellular behavior and inflammatory cytokine expression.
2. Materials and Methods
2.1. Substances
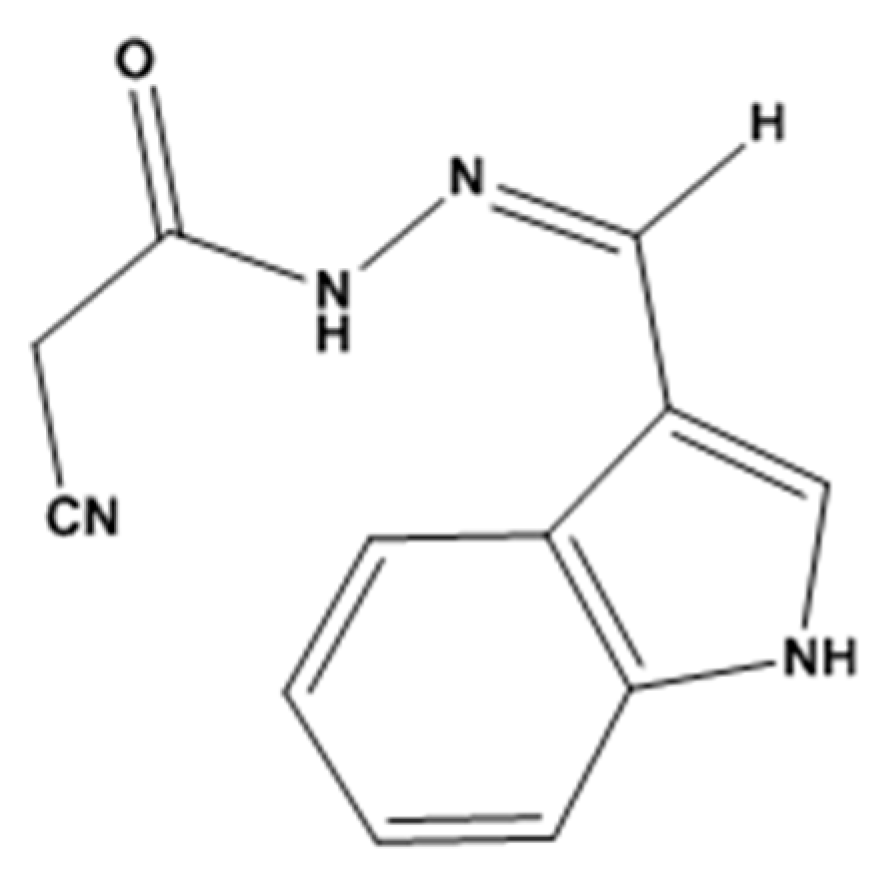
Distilled water; Sterile saline solution (NaCl 0.9%); Indomethacin (Sigma Al-dricht®, USA); Carrageenan (Sigma Aldricht®, USA); Formalin; Sodium heparin 5,000 IU/ml (Cristália®); Ketamine hydrochloride (Vetbrands®); Xylazine Hydrochloride (Vetbrands®); Turk's solution; Phosphate Buffer Saline (PBS); NG-Nitro-L-arginine Methyl Ester (L-NAME) (Sigma Aldri-cht®, USA), Metilene Blue and JR19. N' - (1H-indol-3yl) benzylidene) - 2 cyanoacetrohydrazide (JR19) – synthesized and identified – was provided by the Laboratory of Molecular Synthesis and Vectorization (LSVM) linked to the State University of Paraíba (UEPB), under the responsibility of Prof. Dr. Ricardo Olímpio de Moura [16].
Figure 1.
JR19 - N'-(3-(1H-indol-3-yl)benzylidene)-2-cyanoacetohydrazide.
2.2. Biological activity
2.2.1. Animals
Adult male Swiss Mus musculus mice weighing between 25 and 35g, obtained from the Keizo Asami Immunopathology (LIKA) Animal Facilities of the Federal University of Pernambuco (UFPE) were used. In the vivarium, the animals were kept in plastic cages, under room temperature and humidity (23 ± 2ºC), respecting a 12h light-dark cycle, and fed with feed and water ad libitum. All studies were performed between 08:00 and 17:00 p.m. The animal care and handling procedures were followed in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and the Brazilian College of Animal Experimentation. Previously, the project was approved by the Ethics Committee on Animal Use under number nº 5905022016 (Centro de Ensino Superior e Desenvolvimento – CESED). All procedures from the beginning of the study until the time of euthanasia were performed to avoid suffering, reduce discomfort and pain of the animals.
2.2.2. Inflammatory models
Carrageenan-induced peritonitis in mice
Mice, divided into groups: positive control, negative control and test groups, were respectively treated orally with saline (10 mL/kg), indomethacin (10 mg/kg) and with JR19 at doses of (10 and 20 mg/kg). After 30 minutes of the treatments, 0.25 mL of 1% carrageenan was injected intraperitoneally. Four hours after induction of inflammation, the animals were euthanized, administering 2 mL of heparinized phosphate buffered saline into the intraperitoneal cavity. At the end, an incision was made, collecting the exudate, whose cells were resuspended in 500 μL of PBS and 10 μL of Turk's fluid (1:20). To count the leukocytes, a Neubauer chamber was used under light microscopy, examining the four external quadrants [17].
Subcutaneous air pocket
The induction of an air pocket on the back of mice (group n=6) was performed by injecting sterile air. On the first day, 2.5 mL of sterile air was injected into the animal's back, repeating the procedure after 72 hours. On the seventh day of the trial, the animals received, orally, vehicle (saline, 10 mL/kg), indomethacin (10 mg/kg) and JR19 (10 mg/kg). To induce inflammation, 1.0 mL of a 1% carrageenan solution was injected into the air bag one hour after the administration of the compound. Six hours after the application of the phlogistic agent, the animals were euthanized and the bags were washed with 3.0 mL of phosphate buffer solution (PBS pH 7.2) containing heparin as a carrier liquid. The total leukocyte count was performed in a Neubauer chamber, under optical microscopy. The mean total leukocyte numbers/mm³ of each experimental group and the percentage of inhibition of leukocyte migration compared to the control experimental group were expressed as results [18,19].
Formalin-induced nociception
Mice (group n=6) were treated orally with saline (10 mL/kg), indomethacin and JR19 (10 mg/kg). After 60 minutes, 20 μL of 2.0% formalin solution was administered under the plantar pad of the right hind paw. Immediately afterwards, the time, in seconds, that the animals manifested painful signs with different behaviors was timed: paw elevation, licking, biting, swinging the injected paw, reducing the weight on the paw [20]. The reduction of these behaviors is interpreted as an antinociceptive effect. Based on the pattern of responses, it was possible to establish two periods: first phase, first five minutes and second phase, fifteen to thirty minutes after injection, which were respectively related to the neurogenic phase and inflammatory phase. Results were expressed as mean ± standard deviation of each experimental group, in seconds [21,22].
Investigation of the sGC/NOs pathway in the anti-inflammatory effect
The animals were divided into 6 groups (n=6). Group 1 received saline p.o, group 2 received compound JR19 (p.o, 10 mg/kg, groups 3 and 4, respectively, received i.p. L-NAME (non-specific NOs blocking compounds at a dose of 20 mg/kg) or Methylene Blue, a sGC blocker at a dose of 1 mg/kg, groups 5 and 6 were first pre-treated with the respective blockers and after 30 min they received by p.o the compound JR19. After 30 minutes after each treatment, 0.25mL of 1% carrageenan was injected into the intraperitoneal cavity. Four hours after the induction of inflammation, the animals were euthanized and 2 mL of phosphate buffer solution (PBS pH 7.2) was injected into the peritoneal cavity. The animal's abdomen was lightly massaged and, through an incision, peritoneal fluids were collected to perform the total leukocyte count in a Neubauer chamber, under optical microscopy. The mean total leukocyte numbers/mm³ of each experimental group and the percentage of inhibition of leukocyte migration compared to the control experimental group were expressed as results [17].
Dosage of cytokines in peritoneal exudate
Peritoneal exudate was collected and centrifuged at 7000 rpm at 4°C for 5 minutes. The supernatant was removed to carry out the cytokine experiments (TNF-α, IFN-γ, IL-2, IL-4, IL-6 and IL-17). Flow cytometry tubes, 1 mL multichannel micropipettor, 20% tween PBS and kit for each cytokine for mouse models (BDTM) Cytrometric Bead Array (CBA) mouse Th1/ Th2/ Th17 CBA were used to measure cytokines. Kit. The standard procedure protocol for cytokine assays provided by the manufacturer was followed [23].
2.3. Docking studies
Regarding the target selection and preparation, 3D-structures of Inducible nitric oxide synthase cocrystallized with AR-C95791 (PDB ID: 3E7G), IkBb/NF-kB p65 homodimer complex (PDB ID: 1K3Z) and soluble guanylate cyclase 1 (PDB ID: 3UVJ) were obtained from the Research Collaboratory for Structural Bioinformatics Protein Data Bank database (RCSB PDB - https://www.rcsb.org/). The cocrystallized ligand, ions, and water molecules were initially removed, and hydrogens were added for the structure. Next, these were submitted to the re-docking procedure using the GOLD® v. 5.8.1 software. For the re-docking, all four algorithms (Chemical Piecewise Linear Potential (ChemPLP), GoldScore, ChemScore, and Astex Statistical Potential - ASP) were applied to obtain FitScores and binding modes. The best binding pose was chosen with Root-Mean-Square Deviation (RMSD) value of ≤ 2Ǻ.
The structures of the JR19, nigakinone, N(gamma)-nitro-L-arginine methyl ester (L-NAME) and methylene blue were built with Chemdraw professional 3D 15.0 software. Additionally, they were minimized using the ArgusLab® software by applying the semi-empirical AM1 (Austin Model 1) and then saved as MOL2 files. Molecular docking studies were performed using the GOLD® v. 5.8.1 software by employing the ChemPLP scoring function. A 6 Å region around the cocrystallized ligand was selected using the maximum efficiency of the genetic algorithm (GA). Thus, 100 binding poses were generated for each ligand, and the highest FitScore value was further analyzed for their interactions with the key residues using the Discovery Studio® software and the 3D illustrations were generat-ed using the Chimera® software.
2.4. Statistical Analysis
Data are presented as means± SEM of measurements made on 6 animals in each group. Comparisons across three or more treatments were made using one-way ANOVA with Dunnett post hoc test or repeated measures two-way ANOVA with Bonferroni's post hoc test, when appropriate. All data were analyzed using the Prism 5.01 computer software (GraphPad, San Diego, USA). Statistical differences were considered to be significant at p < 0.05.
3. Results
3.1. Carrageenan-induced peritonitis
This study aimed to evaluate the anti-inflammatory potential of the N-acylhydrazone derivative JR19. For this purpose, the carrageenan-induced peritonitis test was performed. Carrageenan is a polysaccharide of plant origin isolated from red algae and widely used as a inflammatory agent; thus, it chemically activates the inflammatory process by triggering different pathways of the inflammatory cascade [24]. In this experimental model, oral administration of JR19 (at doses of 10 and 20 mg/kg) significantly inhibited leukocyte migration into the peritoneal cavity by 59% and 52%, respectively. The standard treatment with indomethacin (10 mg/kg) resulted in an inhibition of 40%. All groups were compared with the control. Since there was no significant difference between the 10 mg and 20 mg doses of JR19, the 10 mg/kg dose was selected for further pharmacological testing (Figure 2).
3.2. Subcutaneous air pouch
The subcutaneous air pouch experimental model is considered an important tool for the evaluation of acute inflammation. Administration of carrageenan into the air pouch induces a rapid inflammatory response with high levels of PGs and LTs in the exudate [25]. This model mimics rheumatoid arthritis and is used as a screening tool for potential drug candidates for the treatment of arthritis because the air pouch formed on the animal’s back closely resembles the inflamed synovium in patients with the disease [26,27]. In this model, the compound JR19 demonstrated anti-inflammatory activity, as indicated by a significant 66% reduction in cell migration compared to the control group. The standard drug (indomethacin) used in this model was tested at its therapeutic dose of 10 mg/kg and showed inhibition of 55% (Figure 3).
3.3. Formalin-induced nociception
The formalin-induced nociception model was used as a starting point for the study of antinociceptive effects. This model allows the identification of drug candidates with analgesic effects dependent on anti-inflammatory action and involves two phases of nociceptive behavior that indicate the involvement of different chemical mediators. The first phase is characterized by intense neurogenic pain and involves chemical mediators such as substance P, glutamate, and bradykinin. The second phase appears to be caused by tissue and functional changes in the dorsal horn of the spinal cord, involving mediators such as histamine, serotonin, prostaglandins, and bradykinin. Importantly, the second phase is not a consequence of the first. There is a period of quiescence between the two phases in which no stimulus is manifested [28,29]. Oral administration of JR19 or indomethacin at a dose of 10 mg/kg did not affect the first phase of nociception (neurogenic pain) in this model (Figure 4A). In the second phase of nociception (inflammatory pain), both JR19 and the positive control, indomethacin, significantly reduced the animals’ response time to intraplantar formalin injection by 39% and 93%, respectively, compared to the vehicle control group (Figure 4B).
3.4. Docking molecular
Given the results of in vivo studies presented previously, the possible involvement of JR19’s mechanism of action in the oxide nitrergic pathway was investigated through in silico analysis initially. In this way, the selected targets were inducible nitric oxide syn-thase cocrystallized with AR-C95791 (PDB ID: 3E7G), IkBb/NF-kB p65 homodimer com-plex (PDB ID: 1K3Z) and soluble guanylate cyclase 1 (PDB ID: 3UVJ). The scores achieved by each ligand analyzed are described in the Table 1, where higher values of score means better interactions inside the respective binding site, which are demonstrated in the Table 2.
Regarding the target sGC, which is a nitric oxide sensor that when activated perform the catalytic conversion of GTP to cGMP, propagating downstream signals. The reference compound, methylene blue, presented important interactions with Asp530 and Asp486 residues that are essential for metal coordination surrounding the ATP binding site, however JR19 demonstrated another approach of inhibition, given the interaction with Arg552 (Figure 5), which is responsible for the stabilization of γ-phosphate from the substrate [30]. Moreover, JR19 exhibited higher score compared to methylene blue, suggesting that the additional interactions through hydrogen bonds with Arg552 and Thr527 were involved in that achievement.
For the transcription factor, NfκB, there were two different poses of JR19 inside the binding site (Figure 5), which possessed equivalent scores in comparison to the reference compound nigakinone mainly through hydrogen bonds and transfer charge interactions, where JR19 interacted with Arg302 and Arg304 (also observed for nigakinone) that are two basic residues from the p65 NLS for ion pairing interactions with acidic residues in IκBβ, contributing to maintain the stability of NfκB-Iκβ complex, therefore preventing the NfκB’s activation [31]. Additionally, in the studies reported by Qu and collaborators [32] constituents from an ethyl acetate extract that significantly suppressed LPS-induced increases in Nf-κB p65 in the nucleus, also interacted with Arg302, Arg304 and Phe309 residues in a similar way as JR19.
Concerning the iNOS target, which is crucial to the inflammatory process due to the stimulation of immune cells by pro-inflammatory cytokines and endotoxins, its inhibition controls the deleterious effects mediated by NO [33]. Moreover, JR19 presented higher score value compared to the inhibitor of iNOS, L-NAME, and was close to the result achieved by the cocrystallized inhibitor (IC50= 0.35 μM). According to Garcin and collaborators [34] the amino acid residues Gln263 and Glu377 are essential for the inhibitory activity of the target, given that substitutions lead to binding affinity reductions (Kd), mostly Glu 377, whose Kd ranged from 0.4 µM to above 100 µM. In this way, besides the interactions with Gln263 and Glu377, JR19 established a parallel position in relation to the heme group (Figure 5), favoring the interaction with the cofactor Hem901, additionally, interacted with Pro350 that was also observed for the ligand cocrystallized and L-NAME.
3.5. Investigation of the participation of the oxidonitrergic pathway in the anti-inflammatory effect of JR19
To evaluate the involvement of NO in the anti-inflammatory mechanism of compound JR19, the carrageenan-induced peritonitis experiment was performed using an iNOS inhibitor. The results obtained showed that pretreatment of animals with L-NAME reversed the anti-inflammatory effect of compound JR19 on leukocyte migration (15.6%). Thus, the animals treated with L-NAME and the compound did not show a decrease in the number of leukocytes similar to the group treated with the compound alone (48.3%), indicating that the maintenance of NO synthesis is a crucial factor in the anti-inflammatory activity of JR19 (Figure 6). However, the treatment of animals with L-NAME alone did not show a statistically significant difference compared to the control group in this experiment. These results suggest that NO produced by the activation of iNOS is associated with the suppression of leukocyte migration induced by the compound.
Concerning experimental pretreatment with the non-specific soluble guanylate cyclase inhibitor methylene blue (1 mg/kg, i.p.), a significant reversal of the inhibitory effect on leukocyte migration promoted by the compound JR19 (10 mg/kg, p.o.) was observed. Moreover, the administration of the inhibitor alone did not produce any significant modifications in the response to carrageenan when compared to the control group (Figure 7). This suggests that when used in isolation, this agent does not elicit changes in the animals’ baseline inflammatory response to carrageenan. These results support the involvement of NO in the activity of compound JR19 and suggest that the mechanism of action of JR19 may involve the maintenance of NO levels in the acute inflammation model tested in this study.
3.6. Cytokine dosage
Cytokines are essential mediators that direct the inflammatory response to sites of infection and injury. At elevated concentrations, cytokines lead to the activation of nuclear transcription factors such as NFκB, which is associated with inflammation and progression of the process [35]. Therefore, we evaluated the effects of compound JR19 on the production of pro-inflammatory cytokines (TNF-α, IL-2, IFN-γ, IL-6, and IL-17), which play a crucial role in leukocyte recruitment, as well as the anti-inflammatory cytokine (IL-4) in the carrageenan-induced peritonitis model in the presence or absence of L-NAME or methylene blue.
The compound JR19 (at a dose of 10 mg/kg) significantly decreased the levels of the pro-inflammatory cytokines IL-6 (Figure 8A), TNF-α (Figure 8B), IL-17 (Figure 8C), and IFN-γ (Figure 8D). This effect was reversed in groups of animals pretreated with L-NAME, supporting the NO-dependent anti-inflammatory effect of the compound, which to some extent, contributes to the reduction in leukocyte numbers. In contrast, a reversal of the reduction in TNF-α levels was observed in the groups pretreated with methylene blue (Figure 8B). The compound did not affect the levels of IL-2 (Figure 8E) and IL-4 (Figure 8F). We also observed that L-NAME or methylene blue administered alone did not cause significant changes in the levels of any of the cytokines studied. These data suggest that the activity of JR19 in reducing the levels of these cytokines may depend on maintaining NO levels but may be independent of sGC enzyme activation.
4. Discussion
Initially, the peritonitis model was used to study cellular behavior in response to the inflammatory stimulus, using carrageenan as the inflammatory agent. This polypeptide promotes capillary vasodilation in the peritoneal membrane, leading to increased blood flow, structural changes in the microcirculation, allowing extravasation of plasma proteins into the interstitium as inflammatory exudate, and migration of leukocytes from the microcirculation that accumulate at the site of initial injury [36,37]. Substances with anti-inflammatory activity can reduce leukocyte migration into the peritoneal cavity by two mechanisms: by inhibiting the synthesis and/or release of chemotactic mediators or by inhibiting the expression of adhesion molecules, since the presence of chemotactic substances is necessary to facilitate their migration to the site of injury and to trigger their effects in an attempt to eliminate the aggressive agent [38,39].
Leukocytes play a critical role in the defense and repair provided by the inflammatory response, although platelets and erythrocytes also participate. Through the release of chemical mediators of inflammation, leukocytes migrate across interendothelial junctions (diapedesis) and move toward sites of inflammation [40]. In addition to their chemotactic effect, these mediators can trigger a cascade capable of amplifying and releasing other stimulatory factors. Leukocyte activation results in the production of arachidonic acid (AA) metabolites, degranulation and secretion of lysosomal enzymes, cytokine secretion, and increased adhesion molecule expression and integrin exposure [40,41].
Thus, animal models of acute inflammation that allow the quantitative assessment of leukocyte migration have been widely used, making it possible to quantitatively measure cellular migration, inflammatory mediators, and plasma extravasation after an acute inflammatory process induced by various irritants applied in the cavity [42,43,44]. The results obtained in this screening suggest that JR19 has anti-inflammatory activity, possibly related to its ability to suppress the action and/or release of vasoactive amines, NO, and PGs. These results are consistent with those described by other authors, such as Silva (2022), who tested a ferrocenyl-N-acylhydrazone derivative (100 μmol/kg) and found it to be active in this model.
The second model used to investigate the cellular migration profile of JR19 was the subcutaneous air pouch model. The main difference between the air pouch and carrageenan-induced peritonitis models is the predominant cell type in the exudate. In the subcutaneous air pouch model, neutrophils are the most abundant cells responsible for chemotaxis. In contrast, carrageenan-induced peritonitis shows a high amount of macrophages and mast cells attracted to the site of inflammation by pro-inflammatory cytokines, particularly IL-1β and TNF-α [45,46]. Based on the results obtained in this study, it can be concluded that the tested compounds specifically inhibit the production of mediators by neutrophils and macrophages, considering the significant inhibition observed in the air pouch and peritonitis assays, respectively.
Salomé et al. [47] tested a series of naftyl-N-acylhydrazone derivatives and also obtained satisfactory results in the subcutaneous air pouch model. At the doses tested (1, 10, and 30 μmol/kg), most of the compounds showed statistically similar effects to the standard drug dexamethasone (6.5 μmol/kg). Similarly, Cordeiro [48] tested doses of 30 and 100 μmol/kg of a new N-acylhydrazone derivative and its corresponding hydrochloride, using dexamethasone (65 μmol/kg) as a standard.
Considering the anti-inflammatory potential of JR19, it has been suggested that this derivative may also have an antinociceptive effect induced by formalin through a biphasic response. The two phases of the formalin test have different characteristics, making it useful for evaluating analgesic substances and elucidating the mechanism of analgesia. The first phase is characterized by the immediate transmission of impulses to the central nervous system and occurs during the first 5 minutes after the application of the inflammatory agent. It is sensitive to drugs that interact with the opioid system, such as morphine. This pain is caused by the direct action of formalin, especially on the C-afferent sensory fibers and partially on the Aδ afferent fibers [49,50]. The second phase is prolonged, occurring 15-30 minutes after formalin injection, and is associated with the development of an inflammatory response. It can be inhibited by non-steroidal anti-inflammatory drugs (NSAIDs) such as indomethacin. Thus, centrally-acting drugs (opioids) can inhibit both phases, whereas peripherally-acting drugs inhibit only the second phase [28,29,51].
Compound JR19 was active only in the second phase of formalin administration, confirming its lack of central analgesic effect and possibly its retention of anti-inflammatory activity. This effect may be attributed to the inhibition of the synthesis of inflammatory mediators such as PGs, TXs, and LTs [46]. From these results, it can be concluded that the observed antinociceptive effect involves peripheral mechanisms acting on inflammatory pain. These results support the hypothesis that the N-acylhydrazone derivative JR19 may have peripheral antinociceptive activity, possibly by inhibiting PG synthesis or by inhibiting the release and/or production of inflammatory mediators, such as cytokines, histamine, and serotonin, among others. In a study by Silva (2015) [52] the antinociceptive activity of a series of new cyclohexyl-N-acylhydrazone derivatives was evaluated. Most of the compounds tested showed a relevant antinociceptive profile in the formalin-induced inflammatory response (second phase). The compounds active in the neurogenic phase were evaluated in the hot plate test, where the hypothesis of central antinociceptive activity was ruled out.
As demonstrated by molecular docking, the involvement of the enzymes “soluble guanylate cyclase” and “inducible nitric oxide synthase” was investigated using the carrageenan-induced peritonitis model. As mentioned above, this model induces acute inflammation in mice, and in response to the injury, the body mobilizes immune system cells, mainly neutrophils, to the site of injury. A fundamental step in this response is the adhesion of circulating leukocytes to the endothelial cells of blood vessels, allowing their subsequent migration through the barrier of these cells to the site of inflammation. Leukocyte migration from the circulation to the injured tissue is a critical event in the development of the inflammatory process [40,46]. As leukocytes migrate to the tissue, they release a number of chemical mediators that can amplify and prolong the inflammatory process. Therefore, compounds capable of inhibiting the activation of these cells can reduce the inflammatory process [53].
The presence of NO in biological systems is often determined based on physiological effects such as vasodilation, activation of sGC, increased cGMP concentration, citrulline production, or inhibition of platelet aggregation [54]. It can also be assessed using NO synthesis inhibitors such as L-arginine analogs or hemoglobin and by measuring nitrite and nitrate concentrations. All of these methods have varying degrees of specificity and provide indirect information on NO production [55].
Our results suggest that JR19 may exert anti-inflammatory effects by modulating NO levels. NO plays a crucial role in various aspects of the inflammatory response, but its role in leukocyte migration is controversial in the literature, with evidence supporting both anti-inflammatory and pro-inflammatory effects [56]. Studies suggest that NO exerts anti-inflammatory effects by interfering with leukocyte migration. An important piece of evidence for the anti-inflammatory effects of NO is related to the activation of the second messenger cGMP [57,58].
NO promotes the modulation of P-selectin (adhesion protein) expression in vitro induced by IL-1β produced in the endothelium through activation of the GCs enzyme, thereby reducing leukocyte adhesion to the vascular wall and neutrophil aggregation and secretion [57]. Elevated cGMP levels are also associated with a reduction in the expression of glycoprotein IIb/IIIa, another important leukocyte adhesion molecule [59,60]. To support the experimental findings, we also investigated whether the anti-inflammatory activity represented by the reduction of leukocytes exerted by the compound JR19 would involve the GC-cGMP pathway through blockade of the GCs enzyme by methylene blue, a nonspecific inhibitor of this enzyme.
Iwata et al. [61] and Florentino et al. [62] demonstrated, in carrageenan-induced pleurisy models, that administration of a NO donor (L-arginine) reduces inflammatory cell migration and edema formation. This model, similar to ours, mimics an acute inflammatory process. The anti-inflammatory effect of NO has been attributed to the blockade of transcription factor binding to nuclear DNA (NFκB), thereby inhibiting the synthesis of chemokine IL-8 (a chemotactic factor secreted by activated monocytes and macrophages that promotes the coordinated and directional migration of immune cells such as neutrophils, basophils, and T lymphocytes), thereby compromising the chemotaxis process [57,63].
There is also an important relationship between NO and O2- levels associated with leukocyte migration, which concerns the enzymatic system responsible for O2- synthesis, which possesses a heme prosthetic group to which NO can bind, inhibiting the generation of this reactive oxygen species [57,64]. The reduction in O2- availability prevents its dismutation to hydrogen peroxide (H2O2). This chemical species is responsible for inducing leukocyte adhesion by generating PAF and increasing the expression of other molecules involved in this process [65,66].
In addition, elevated levels of O2- can activate NFκB, leading to increased production of cytokines and chemokines and stimulating mast cells to release pro-adhesive agents such as cytokines s [67,68,69]. Therefore, we conclude that the anti-inflammatory effect of compound JR19, as observed by the reduction of peritoneal leukocytes, is due in part to the acute maintenance of NO levels.
Finally, peritonitis was used to study cytokine expression by JR19 through an sGC/NOS-dependent mechanism using the blockers L-NAME or methylene blue. JR19 (at a dose of 10 mg/kg) was effective in significantly reducing the levels of the pro-inflammatory cytokines IL-6, TNF-α, IL-17, and IFN-γ. IL-6 is a cytokine secreted by various cell types such as macrophages, monocytes, eosinophils, and hepatocytes, and TNF-α is a potent inducer of IL-6 [43]. It is a pro-inflammatory cytokine that promotes the maturation and activation of neutrophils and macrophages as well as the differentiation/maintenance of cytotoxic T lymphocytes and NK cells. IL-6 is one of the earliest and most important mediators in the induction and control of acute phase protein synthesis and release by hepatocytes during painful stimuli such as infection [65,70] It is also involved in the metabolic regulation of C-reactive protein (CRP), an acute-phase protein that increases rapidly during inflammatory processes [71,72]. Elevation of serum IL-6 and CRP levels may cause downregulation of NO production by inhibiting the enzyme nitric oxide synthase [73,74].
TNF-α is mainly produced by macrophages. It acts on endothelial cells to promote vasodilation and stimulates chemokines to facilitate the chemotaxis of leukocytes, mainly neutrophils and monocytes [75]. In addition to stimulating their activation, TNF-α also stimulates the production of acute-phase proteins and fibrinogen during the inflammatory process [76]. Even at low concentrations, TNF-α induces the expression of adhesion molecules in endothelial cells and stimulates macrophages and other cells to secrete chemokines [77,78].
IFN-γ is primarily produced by NK cells and T lymphocytes and acts synergistically with TNF-α to stimulate chemokine secretion [79]. In addition, IFN-γ induces the expression of intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1), promoting better leukocyte adhesion to the endothelium and facilitating their diapedesis to the site of inflammation [79,80]. It has been reported that this cytokine is involved in neutrophil infiltration in malignant tumors, which typically contain various types of immune cells in addition to cancer cells [81].
IL-17 is a pro-inflammatory cytokine produced by T cells with the ability to activate and mature neutrophils [82]. It also stimulates the production of IL-6 and IL-8 (responsible for the migratory stimulus of immune cells, mainly neutrophils, and increases the expression of adhesion molecules on endothelial cells) [82,83]. These pro-inflammatory cytokines, among others, stimulate the production of other cytokines and other inflammatory mediators such as inflammatory prostanoids via COX-2 [84].
Freitas [85] has classified N-acylhydrazone derivatives as inhibitors of p38 MAPK. Additionally, Cordeiro [48] demonstrated that these derivatives inhibit the expression of mediators involved in this pathway, such as IL-1β, TNF-α, and NO (at doses of 10, 30, and 100 μmol/kg). Furthermore, Cordeiro [48] found that at the same doses, there was a significant reduction in pain induced by formalin during the second phase, compared to the control group using morphine.
Since cytokines play a critical role in the inflammatory response and are produced by various cell types at the site of injury and by cells of the immune system during the inflammatory process, compounds that reduce the levels of pro-inflammatory cytokines may help to control inflammation. Thus, JR19 shows great promise in controlling inflammation by acting on key inflammatory pathways, making it a potential candidate for a new anti-inflammatory drug.
5. Conclusions
Our results showed that JR19 exhibited activity in reducing leukocyte migration and inflammation-related nociception in the inflammatory phase, suggesting that its effects are likely related to the regulation of pro-inflammatory mediators such as nitric oxide and cytokines (IL-6, IL-17, TNF-α, IFN-γ). This hypothesis was supported by in vivo assays and further substantiated by in silico assays where JR19 showed affinity, particularly for iNOS, sGCs, and the transcription factor NFκB. Taken together, the data obtained in the screening indicated that the novel N-acylhydrazone derivative showed promising anti-inflammatory activity, which opens favorable prospects for further complementary studies to ensure its safety and pharmacological efficacy.
Author Contributions
Participated in research design: P.R.d.S., V.L.d.S., R.O.d.M., N.d.M.A., T.G.d.S and S.A.d.S, performed the experiments and the data analysis: P.R.d.S., N.d.M.A., S.A.d.S., M.E.C.A., T.B.C., Y.M.S.d.M.e.S.; wrote or contributed to the writing of the manuscript: P.R.d.S., N.d.M.A., S.A.d.S., M.E.C.A., T.B.C., Y.M.S.d.M.e.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed in part by Paraiba State University, grant #02/2023.
Acknowledgments
Paraiba State University, Fundação de Amparo à Pesquisa do Estado da Paraiba (FAPESQ), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes).
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Fikry, E.M.; Hasan, W.A.; Mohamed, E.G. Rutin and meloxicam attenuate paw inflammation in mice: Affecting sorbitol dehydrogenase activity. J. Biochem. Mol. Toxicol. 2018, 32, e22029. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. FASEB J. 2017, 31, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Song, L.; Wang, X.; Xu, Z.; Wang, S.; Wang, J.; Xu, H.; Zheng, Y.; Wang, Y. Cardiac injury prediction and lymphocyte immunity and inflammation analysis in hospitalized patients with coronavirus disease 2019 (COVID-19). Int. J. Cardiol. 2021, 326, 237–242. [Google Scholar] [CrossRef] [PubMed]
- El Menyiy, N.; El Allam, A.; Aboulaghras, S.; Jaouadi, I.; Bakrim, S.; El Omari, N.; Shariati, M.A.; Miftakhutdinov, A.; Wilairatana, P.; Mubarak, M.S.; et al. Inflammatory auto-immune diseases of the intestine and their management by natural bioactive compounds. BioMedicine 2022, 151, 113158. [Google Scholar] [CrossRef]
- da Silva, P.R.; NunesPazos, N.D.; Gonzaga, T.K.S.D.N.; de Andrade, J.C. ; Monteiro,.B.; Portela, A.C.R.; Pires, H.F.O.; Maia, M.d.S.; da Fonsêca, D.V.; Scotti, M.T.; et al. Anxiolytic and antidepressant-like effects of monoterpene tetrahydrolinalool and in silico approach of new potential targets. Curr. Top. Med. Chem. 1530. [Google Scholar] [CrossRef]
- Alessandri, A.L.; Sousa, L.P.; Lucas, C.D.; Rossi, A.G.; Pinho, V.; Teixeira, M.M. Resolution of inflammation: Mechanisms and opportunity for drug development. Pharmacol. Ther. 2013, 139, 189–212. [Google Scholar] [CrossRef]
- Gomes, F.I.F.; Cunha, F.Q.; Cunha, T.M. Peripheral nitric oxide signaling directly blocks inflammatory pain. Biochem. Pharmacol. 2020, 176, 113862. [Google Scholar] [CrossRef]
- Sansbury, B.E.; Cummins, T.D.; Tang, Y.; Hellmann, J.; Holden, C.R.; Harbeson, M.A.; Chen, Y.; Patel, R.P.; Spite, M.; Bhatnagar, A.; et al. Overexpression of Endothelial Nitric Oxide Synthase Prevents Diet-Induced Obesity and Regulates Adipocyte Phenotype. Circ. Res. 2012, 111, 1176–1189. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E. Nitric oxide signaling in health and disease. Cell 2022, 185, 2853–2878. [Google Scholar] [CrossRef]
- Buckley, C.D.; Gilroy, D.W.; Serhan, C.N. Proresolving Lipid Mediators and Mechanisms in the Resolution of Acute Inflammation. Immunity 2014, 40, 315–327. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147–114147. [Google Scholar] [CrossRef]
- Patrignani, P.; Patrono, C. Cyclooxygenase inhibitors: From pharmacology to clinical read-outs. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2015, 1851, 422–432. [Google Scholar] [CrossRef]
- Kowalski, M.L.; Makowska, J.S. Seven Steps to the Diagnosis of NSAIDs Hypersensitivity: How to Apply a New Classification in Real Practice? Allergy, Asthma Immunol. Res. 2015, 7, 312–320. [Google Scholar] [CrossRef]
- Nakhostin, A.; Mirseyyedhosein, S.; Toolabi, M.; Khodabakhsh, P.; Aghamiri, H.; Ghaffari, S.; Shafaroodi, H.; Shayesteh, A.; Amini, M.; Shafiee, A.; et al. Synthesis, conformational assignment, and anti-inflammatory activities of N-arylidene-2-(4-chloro-2-(2-substituted phenoxy)phenyl)acetic acid hydrazides. Med. Chem. Res. 2016, 25, 2220–2236. [Google Scholar] [CrossRef]
- Kumar, A.; Alam, M.S.; Hamid, H.; Chugh, V.; Tikla, T.; Kaul, R.; Dhulap, A.; Sharma, S.K. Design and synthesis of anti–inflammatory 1,2,3–triazolylpyrrolobenzodiazepinone derivatives and impact of molecular structure on COX–2 selective targeting. J. Mol. Struct. 2023, 1272. [Google Scholar] [CrossRef]
- Moraes, A.D.T.d.O.; de Miranda, M.D.S.; Jacob. T.T.; Amorim, C.A.d.C.; de Moura, R.O.; da Silva, S..S.; Soares, M.B.P.; de Almeida, S.M.V.; Souza, T.R.C.d.L.; de Oliveira, J.F.; et al. Synthesis, in vitro and in vivo biological evaluation, COX-1/2 inhibition and molecular docking study of indole-N-acylhydrazone derivatives. Bioorganic Med. Chem. 2018, 26, 5388–5396. [Google Scholar] [CrossRef]
- Oliveira-Tintino, C.D.d.M.; Pessoa, R.T.; Fernandes, M.N.M.; Alcântara, I.S.; da Silva, B.A.F.; de Oliveira, M.R.C.; Martins, A.O.B.P.B.; da Silva, M.D.S.; Tintino, S.R.; Rodrigues, F.F.G.; et al. Anti-inflammatory and anti-edematogenic action of the Croton campestris A. St.-Hil (Euphorbiaceae) essential oil and the compound β-caryophyllene in in vivo models. Phytomedicine 2018, 41, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Sarmento-Filha, M.J.; Torres-Rêgo, M.; Daniele-Silva, A.; de Queiroz-Neto, M.F.; Rocha, H.A.O.; Camara, C.A.; Araújo, R.M.; da Silva-Júnior, A.A.; Silva, T.M.S.; Fernandes-Pedrosa, M.d.F. Phytochemical analysis by UPLC-QTOF-MS/MS and evaluation of antioxidant and anti-inflammatory activities of the extract and fractions from flowers of Cochlospermum vitifolium. South Afr. J. Bot. 2022, 148, 293–306. [Google Scholar] [CrossRef]
- Torres-Rêgo, M.; Furtado, A.A.; Bitencourt, M.A.O.; Lima, M.C.J.d.S.; de Andrade, R.C.L.C.; de Azevedo, E.P.; Soares, T.d.C.; Tomaz, J.C.; Lopes, N.P.; da Silva-Júnior, A.A.; et al. Anti-inflammatory activity of aqueous extract and bioactive compounds identified from the fruits of Hancornia speciosa Gomes (Apocynaceae). BMC Complement. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, L.J.; Guilhon, C.C.; Alviano, D.S.; Matheus, M.E.; Antoniolli, A.R.; Cavalcanti, S.C.; Alves, P.B.; Alviano, C.S.; Fernandes, P.D. Characterisation of the anti-inflammatory and antinociceptive activities of the Hyptis pectinata (L.) Poit essential oil. J. Ethnopharmacol. 2011, 134, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.S.H.d.S.; Malta, D.J.D.N.; Laranjeira, L.P.M.; Maia, M.B.S.; Colaço, N.C.; de Lima, M.D.C.A.; Galdino, S.L.; Pitta, I.d.R.; Gonçalves-Silva, T. Anti-inflammatory and antinociceptive activities of indole–imidazolidine derivatives. Int. Immunopharmacol. 2011, 11, 1816–1822. [Google Scholar] [CrossRef]
- The Formalin Test in Mice: Dissociation Between;
- Guerra, A.S.H.d.S.; Malta, D.J.D.N.; Laranjeira, L.P.M.; Maia, M.B.S.; Colaço, N.C.; de Lima, M.D.C.A.; Galdino, S.L.; Pitta, I.d.R.; Gonçalves-Silva, T. Anti-inflammatory and antinociceptive activities of indole–imidazolidine derivatives. Int. Immunopharmacol. 2011, 11, 1816–1822. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.; Bomfim, R.; Jesus, H.; Alves, P.; Blank, A.; Estevam, C.; Antoniolli, A.; Thomazzi, S. Evaluation of the analgesic and anti-inflammatory effects of the essential oil of Lippia gracilis leaves. J. Ethnopharmacol. 2010, 129, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Dissertação Fernanda Virginia Barreto Mota.
- Jain, M.; Parmar, H.S. Evaluation of antioxidative and anti-inflammatory potential of hesperidin and naringin on the rat air pouch model of inflammation. Inflamm. Res. 2011, 60, 483–491. [Google Scholar] [CrossRef]
- Vandooren, J.; Berghmans, N.; Dillen, C.; Van Aelst, I.; Ronsse, I.; Israel, L.L.; Rosenberger, I.; Kreuter, J.; Lellouche, J.-P.; Michaeli, S.; et al. Intradermal air pouch leukocytosis as an in vivo test for nanoparticles. Int. J. Nanomed. 2013, 8, 4745–4756. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and Molecular Mechanisms of Pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, S.; Migliorati, G.; Delfino, D. Association of inflammatory mediators with pain perception. BioMedicine 2017, 96, 1445–1452. [Google Scholar] [CrossRef]
- Allerston, C.K.; von Delft, F.; Gileadi, O. Crystal Structures of the Catalytic Domain of Human Soluble Guanylate Cyclase. PLOS ONE 2013, 8, e57644. [Google Scholar] [CrossRef]
- Malek, S.; Huang, D.-B.; Huxford, T.; Ghosh, S.; Ghosh, G. X-ray Crystal Structure of an IκBβ·NF-κB p65 Homodimer Complex. J. Biol. Chem. 2003, 278, 23094–23100. [Google Scholar] [CrossRef]
- Qu, B.; Wang, S.; Zhu, H.; Yin, T.; Zhou, R.; Hu, W.; Lu, C. Core Constituents of Caragana sinica Root for Rheumatoid Arthritis Treatment and the Potential Mechanism. ACS Omega 2022, 8, 2586–2595. [Google Scholar] [CrossRef]
- Singh, M.; Padhy, G.; Vats, P.; Bhargava, K.; Sethy, N.K. Hypobaric hypoxia induced arginase expression limits nitric oxide availability and signaling in rodent heart. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2014, 1840, 1817–1824. [Google Scholar] [CrossRef]
- Garcin, E.D.; Arvai, A.S.; Rosenfeld, R.J.; Kroeger, M.D.; Crane, B.R.; Andersson, G.; Andrews, G.; Hamley, P.J.; Mallinder, P.R.; Nicholls, D.J.; et al. Anchored plasticity opens doors for selective inhibitor design in nitric oxide synthase. Nat. Chem. Biol. 2008, 4, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F. Targeting the interleukin pathway in the treatment of asthma. Lancet 2015, 386, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Paulino, N.; Abreu, S.R.L.; Uto, Y.; Koyama, D.; Nagasawa, H.; Hori, H.; Dirsch, V.M.; Vollmar, A.M.; Scremin, A.; Bretz, W.A. Anti-inflammatory effects of a bioavailable compound, Artepillin C, in Brazilian propolis. Eur. J. Pharmacol. 2008, 587, 296–301. [Google Scholar] [CrossRef]
- Francischetti, I.; Bitu MORENO, J.; Scholz, M.; Bonetti YOSHIDA, W.; Paulo, S.; Francischetti Rua Doutor Augusto Barreto, I. Os Leucócitos e a Resposta Inflamatória Na Lesão de Isquemia-Reperfusão Leukocytes and the Inflammatory Response in Ischemia-Reperfusion Injury; 2010; Vol. 25;
- A Muller, W. Leukocyte-Endothelial Cell Interactions in the Inflammatory Response. Lab. Investig. 2002, 82, 521–533. [Google Scholar] [CrossRef]
- Sherwood, E.R.; Toliver-Kinsky, T. Mechanisms of the inflammatory response. Best Pr. Res. Clin. Anaesthesiol. 2004, 18, 385–405. [Google Scholar] [CrossRef] [PubMed]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. Mech. Dis. 2019, 15, 493–518. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Oncotarget 7204 Www.Impactjournals.Com/Oncotarget Inflammatory Responses and Inflammation-Associated Diseases in Organs; 2018; Vol. 9;
- Sedgwick, A.D.; Lees, P. A Comparison of Air Pouch, Sponge and Pleurisy Models of Acute Carrageenan Inflammation in the Rat; 1986; Vol. 18;
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: a critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- da Silva, P.R.; Santo, R.F.D.E.; Melo, C.d.O.; Cavalcante, F.E.P.; Costa, T.B.; Barbosa, Y.V.; Silva, Y.M.S.d.M.e.; de Sousa, N.F.; Villarreal, C.F.; de Moura, R.O.; et al. The Compound (E)-2-Cyano-N,3-diphenylacrylamide (JMPR-01): A Potential Drug for Treatment of Inflammatory Diseases. Pharmaceutics 2022, 14, 188. [Google Scholar] [CrossRef]
- Da, E.H.; Ramos, S. Avaliação Das Atividades Biológicas Do Óleo Essencial Do Látex de Mangifera Indica L. (Var. 2014. [Google Scholar]
- Baral, P.; Udit, S.; Chiu, I.M. Pain and immunity: implications for host defence. Nat. Rev. Immunol. 2019, 19, 433–447. [Google Scholar] [CrossRef]
- Salomé, D.d.C.; de Freitas, R.H.C.N.; Fraga, C.A.M.; Fernandes, P.D. Novel Regioisomeric Analogues of Naphthyl-N-Acylhydrazone Derivatives and Their Anti-Inflammatory Effects. Int. J. Mol. Sci. 2022, 23, 13562. [Google Scholar] [CrossRef]
- Cordeiro, N.d.M.; Freitas, R.H.; Fraga, C.A.M.; Fernandes, P.D. New 2-amino-pyridinyl-N-acylhydrazones: Synthesis and identification of their mechanism of anti-inflammatory action. BioMedicine 2020, 123, 109739. [Google Scholar] [CrossRef] [PubMed]
- Ha Yoon, M.; il Choi, J. Pharmacologic Interaction between Cannabinoid and Either Clonidine or Neostigmine in the Rat Formalin Test; 2003; Vol. 99;
- Kang, S.; Kim, C.H.; Lee, H.; Kim, D.Y.; Han, J.I.; Chung, R.K.; Lee, G.Y. Antinociceptive Synergy Between the Cannabinoid Receptor Agonist WIN 55,212-2 and Bupivacaine in the Rat Formalin Test. Obstet. Anesthesia Dig. 2007, 104, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, H.A.; Nascimento, M.V.M.; Tavares, A.; Galdino, P.M.; de Paula, J.R.; Costa, E.A. Effects of ethanolic extract of leaves of Lafoensia pacari A. St.-Hil., Lythraceae (pacari), in pain and inflammation models. Rev. Bras. de Farm. 2010, 20, 328–333. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Júnior, W.B.; Alexandre-Moreira, M.S.; Costa, F.N.; Monteiro, C.E.d.S.; Ferreira, F.F.; Barroso, R.C.R.; Noël, F.; Sudo, R.T.; Zapata-Sudo, G.; et al. Novel Orally Active Analgesic and Anti-Inflammatory Cyclohexyl-N-Acylhydrazone Derivatives. Molecules 2015, 20, 3067–3088. [Google Scholar] [CrossRef]
- Domínguez-Luis, M.; Herrera-García, A.; Arce-Franco, M.; Armas-González, E.; Rodríguez-Pardo, M.; Lorenzo-Díaz, F.; Feria, M.; Cadenas, S.; Sánchez-Madrid, F.; Díaz-González, F. Superoxide anion mediates the L-selectin down-regulation induced by non-steroidal anti-inflammatory drugs in human neutrophils. Biochem. Pharmacol. 2013, 85, 245–256. [Google Scholar] [CrossRef]
- Ghasemi, M. Ghasemi, M. Nitric Oxide: Antidepressant Mechanisms and Inflammation. In Advances in Pharmacology; Academic Press Inc., 2019; Vol. 86, pp. 121–152 ISBN 9780128166680.
- Biava, M.; Porretta, G.C.; Poce, G.; Battilocchio, C.; Alfonso, S.; Rovini, M.; Valenti, S.; Giorgi, G.; Calderone, V.; Martelli, A.; et al. Novel Analgesic/Anti-Inflammatory Agents: Diarylpyrrole Acetic Esters Endowed with Nitric Oxide Releasing Properties. J. Med. Chem. 2011, 54, 7759–7771. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Rao, C. v. INOS-Selective Inhibitors for Cancer Prevention: Promise and Progress. Future Med Chem 2012, 4, 2193–2204. [Google Scholar] [CrossRef]
- Król, M.; Kepinska, M. Human Nitric Oxide Synthase—Its Functions, Polymorphisms, and Inhibitors in the Context of Inflammation, Diabetes and Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 56. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.; de Almeida, M.; Silva, J.; Albino, S.; Espírito-Santo, R.; Lima, M.; Villarreal, C.; Moura, R.; Santos, V. (E)-2-Cyano-3-(1H-Indol-3-yl)-N-Phenylacrylamide, a Hybrid Compound Derived from Indomethacin and Paracetamol: Design, Synthesis and Evaluation of the Anti-Inflammatory Potential. Int. J. Mol. Sci. 2020, 21, 2591. [Google Scholar] [CrossRef]
- Ahluwalia, A.; Foster, P.; Scotland, R.S.; Mclean, P.G.; Mathur, A.; Perretti, M.; Moncada, S.; Hobbs, A.J. Antiinflammatory Activity of Soluble Guanylate Cyclase: CGMP-Dependent down-Regulation of P-Selectin Expression and Leukocyte Recruitment; 2004; Vol. 101;
- Florentino, I.F.; Galdino, P.M.; De Oliveira, L.P.; Silva, D.P.; Pazini, F.; Vanderlinde, F.A.; Lião, L.M.; Menegatti, R.; Costa, E.A. Involvement of the NO/cGMP/KATP pathway in the antinociceptive effect of the new pyrazole 5-(1-(3-fluorophenyl)-1H-pyrazol-4-yl)-2H-tetrazole (LQFM-021). Nitric Oxide 2015, 47, 17–24. [Google Scholar] [CrossRef]
- Iwata, M.; Suzuki, S.; Asai, Y.; Inoue, T.; Takagi, K. Involvement of Nitric Oxide in a Rat Model of Carrageenin-Induced Pleurisy. Mediat. Inflamm. 2010, 2010, 1–11. [Google Scholar] [CrossRef]
- Florentino, I.F.; Silva, D.P.; Silva, D.M.; Cardoso, C.S.; Moreira, A.L.; Borges, C.L.; Soares, C.M.d.A.; Galdino, P.M.; Lião, L.M.; Ghedini, P.C.; et al. Potential anti-inflammatory effect of LQFM-021 in carrageenan-induced inflammation: The role of nitric oxide. Nitric Oxide 2017, 69, 35–44. [Google Scholar] [CrossRef]
- Carreau, A.; Kieda, C.; Grillon, C. Nitric oxide modulates the expression of endothelial cell adhesion molecules involved in angiogenesis and leukocyte recruitment. Exp. Cell Res. 2011, 317, 29–41. [Google Scholar] [CrossRef]
- Schioppa, T.; Sozio, F.; Barbazza, I.; Scutera, S.; Bosisio, D.; Sozzani, S.; Del Prete, A. Molecular Basis for CCRL2 Regulation of Leukocyte Migration. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Secco, D.D.; Paron, J.A.; de Oliveira, S.H.; Ferreira, S.H.; Silva, J.S.; Cunha, F.d.Q. Neutrophil migration in inflammation: nitric oxide inhibits rolling, adhesion and induces apoptosis. Nitric Oxide 2003, 9, 153–164. [Google Scholar] [CrossRef]
- Matsui, T.C.; Coura, G.M.; Melo, I.S.; Batista, C.R.; Augusto, P.S.A.; Godin, A.M.; Araújo, D.P.; César, I.C.; Ribeiro, L.S.; Souza, D.G.; et al. Nicorandil inhibits neutrophil recruitment in carrageenan-induced experimental pleurisy in mice. Eur. J. Pharmacol. 2015, 769, 306–312. [Google Scholar] [CrossRef]
- Suk, K.; Kim, S.Y.; Kim, H. Regulation of IL-18 Production by IFNg and PGE 2 in Mouse Microglial Cells: Involvement of NF-KB Pathway in the Regulatory Processes; 2001; Vol. 77;
- Santos, L.A.M.; Ribeiro, E.L.; Barbosa, K.P.S.; Fragoso, I.T.; Gomes, F.O.d.S.; Donato, M.A.M.; Silva, B.S.; Silva, A.K.S.; Rocha, S.W.S.; França, M.E.R.; et al. Diethylcarbamazine inhibits NF-κB activation in acute lung injury induced by carrageenan in mice. Int. Immunopharmacol. 2014, 23, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Albensi, B.C. What Is Nuclear Factor Kappa B (NF-ΚB) Doing in and to the Mitochondrion? Front Cell Dev Biol 2019, 7. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in Inflammation, Autoimmunity and Cancer. Int Immunol 2021, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.C.; Lopes, A.L.; Macedo, R.C.O.; Correa, C.S.; Ramis, T.R.; Ribeiro, J.L.; Reischak-Oliveira, A. Marcadores Inflamatórios, Função Endotelial e Riscos Cardiovasculares. J Vasc Bras 2014, 13, 108–115. [Google Scholar] [CrossRef]
- Le, K.T.T.; Chu, X.; Jaeger, M.; Plantinga, J.A.; Matzaraki, V.; Withoff, S.; Joosten, L.A.B.; Netea, M.G.; Wijmenga, C.; Li, Y.; et al. Leukocyte-Released Mediators in Response to Both Bacterial and Fungal Infections Trigger IFN Pathways, Independent of IL-1 and TNF-α, in Endothelial Cells. Front. Immunol. 2019, 10, 2508. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.M.; Campos Pellanda, L.; Gus, I.; Portal, V.L. Inflammatory Markers of Cardiovascular Disease in the Elderly;
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and Atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin Rev Allergy Immunol 2020, 58, 342–365. [Google Scholar]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A Road to Inflammation. Nat Rev Immunol 2023, 23, 289–303. [Google Scholar] [PubMed]
- Schurgers, E.; Billiau, A.; Matthys, P.; Dolati, S.; Sadreddini, S.; Rostamzadeh, D.; Ahmadi, M.; Jadidi-Niaragh, F.; Yousefi, M.; Rosillo, M.A.; et al. Collagen-Induced Arthritis as an Animal Model for Rheumatoid Arthritis: Focus on Interferon-γ. J. Interf. Cytokine Res. 2011, 31, 917–926. [Google Scholar] [CrossRef]
- Larkin, J.; Ahmed, C.M.; Wilson, T.D.; Johnson, H.M. Regulation of Interferon Gamma Signaling by Suppressors of Cytokine Signaling and Regulatory T Cells. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef]
- Ohms, M.; Möller, S.; Laskay, T. An Attempt to Polarize Human Neutrophils Toward N1 and N2 Phenotypes in vitro. Front. Immunol. 2020, 11, 532. [Google Scholar] [CrossRef]
- Mills, K.H.G. IL-17 and IL-17-producing cells in protection versus pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef]
- Schaper, F.; Rose-John, S. Interleukin-6: Biology, signaling and strategies of blockade. Cytokine Growth Factor Rev. 2015, 26, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Stenken, J.A.; Poschenrieder, A.J. Bioanalytical chemistry of cytokines – A review. Anal. Chim. Acta 2015, 853, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.H.C.N.; Cordeiro, N.M.; Carvalho, P.R.; Alves, M.A.; Guedes, I.A.; Valerio, T.S.; Dardenne, L.E.; Lima, L.M.; Barreiro, E.J.; Fernandes, P.D.; et al. Discovery of naphthyl-N -acylhydrazone p38α MAPK inhibitors with in vivo anti-inflammatory and anti-TNF-α activity. Chem. Biol. Drug Des. 2018, 91, 391–397. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Influence of JR19 (10 and 20 mg/kg) treatment on total leukocyte count in the peritonitis induced by carrageenan 1%. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
Figure 2.
Influence of JR19 (10 and 20 mg/kg) treatment on total leukocyte count in the peritonitis induced by carrageenan 1%. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
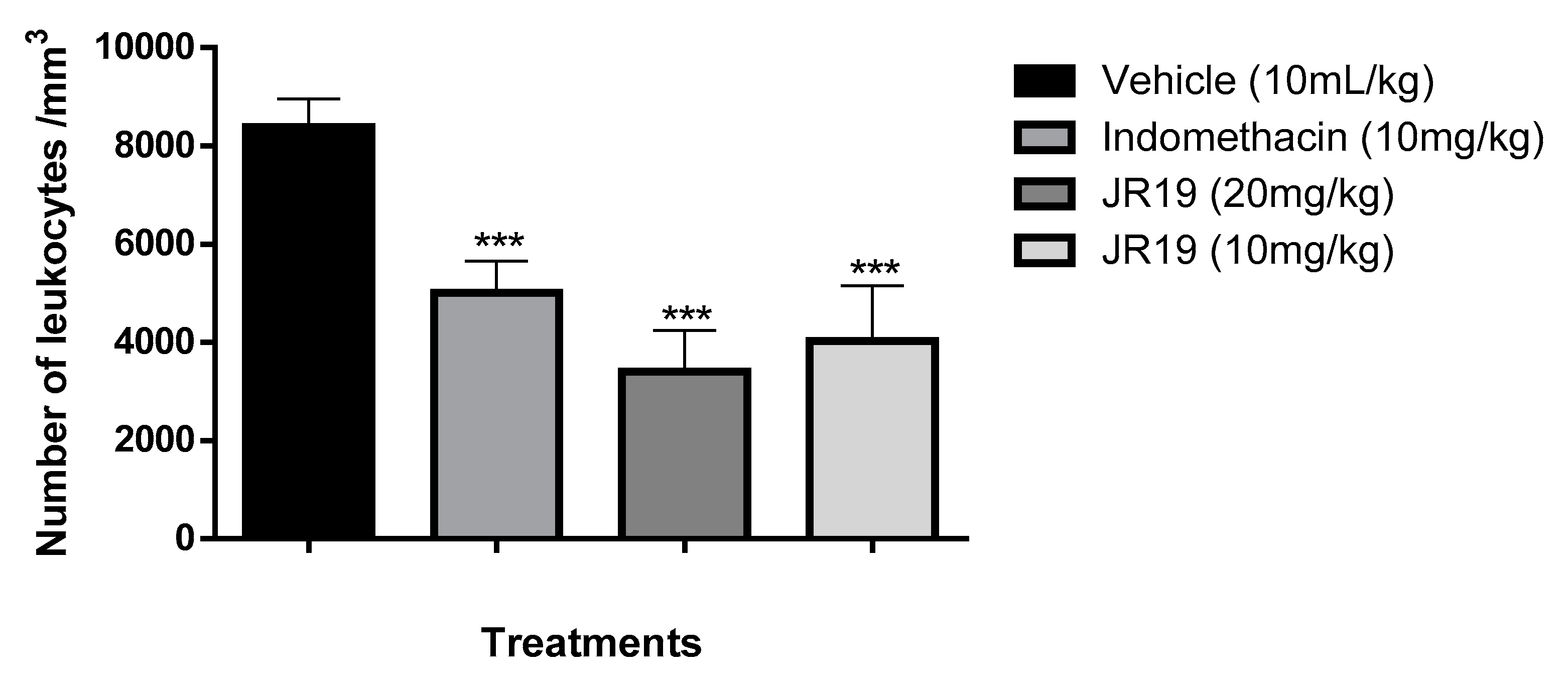
Figure 3.
Influence of JR19 (10 mg/kg) treatment on total leukocyte count in the subcutaneous air pocket induced by carrageenan 1%. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
Figure 3.
Influence of JR19 (10 mg/kg) treatment on total leukocyte count in the subcutaneous air pocket induced by carrageenan 1%. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
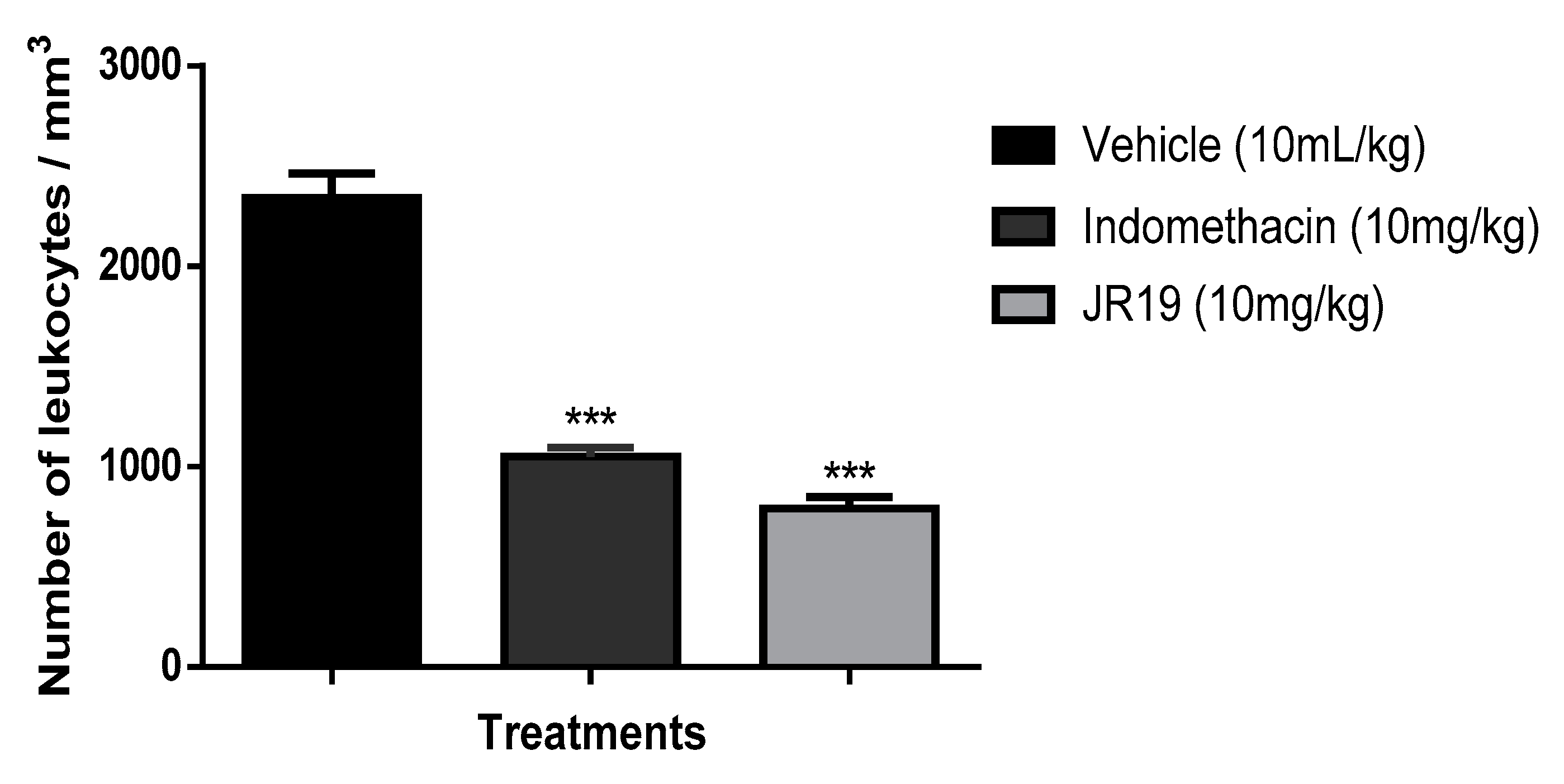
Figure 4.
Influence of treatment with the compound JR19 on the time of reactivity to pain in phase I (A) and phase II (B) of the intraplantar formalin test. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
Figure 4.
Influence of treatment with the compound JR19 on the time of reactivity to pain in phase I (A) and phase II (B) of the intraplantar formalin test. Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group (ANOVA followed by Dunnett's test).
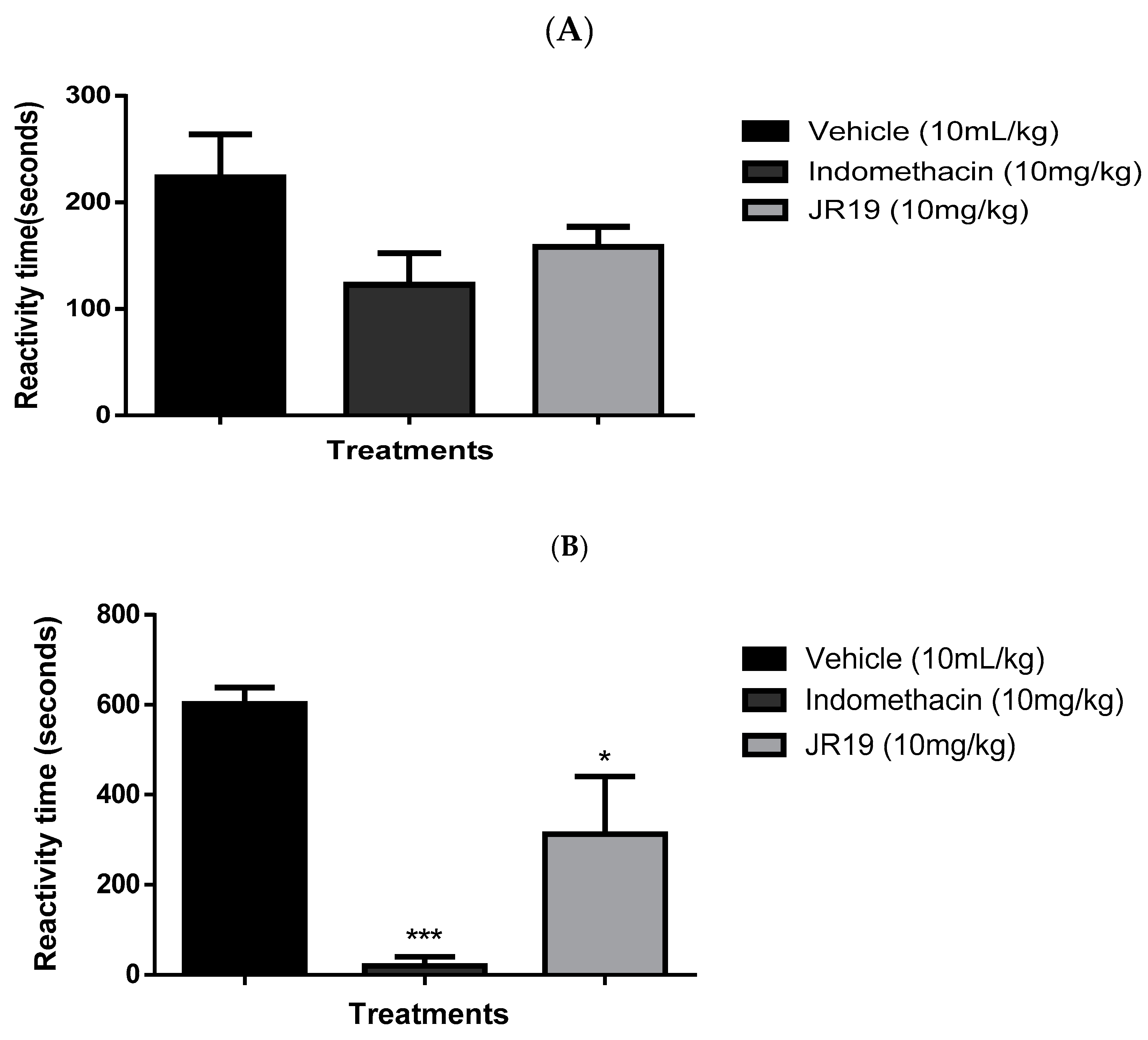
Figure 5.
3D representation of JR19 in sGC (A), NfκB (B-pose 1), NfκB (C-pose 2) and iNOS (D) binding site.
Figure 5.
3D representation of JR19 in sGC (A), NfκB (B-pose 1), NfκB (C-pose 2) and iNOS (D) binding site.
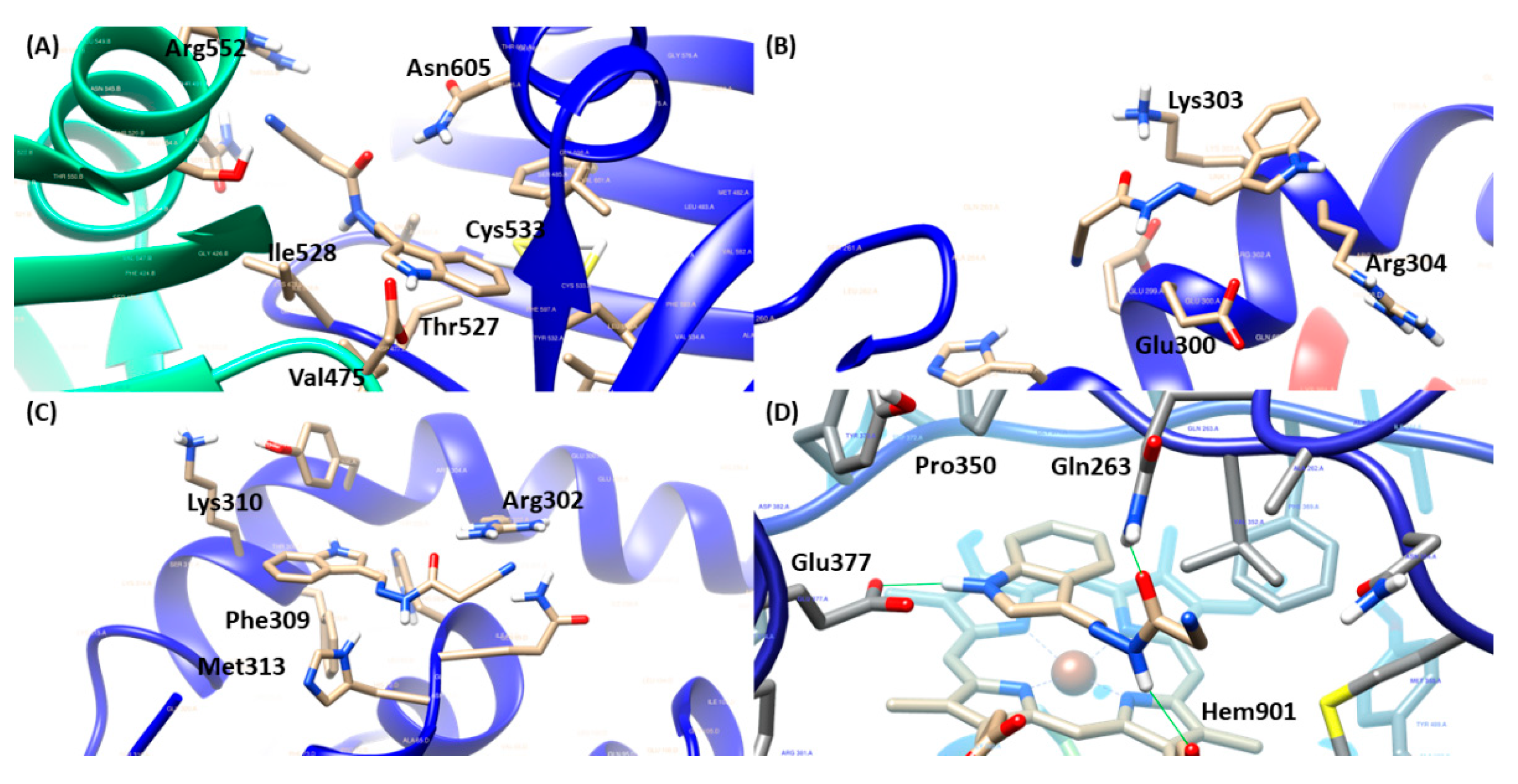
Figure 6.
Influence of treatment with the compounds JR19 and L-NAME on the total leukocyte count in the model of peritonitis induced by carrageenan (1%). Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group, ### p < 0,001 JR19 versus o JR19 + L-NAME (ANOVA followed by Dunnett's test).
Figure 6.
Influence of treatment with the compounds JR19 and L-NAME on the total leukocyte count in the model of peritonitis induced by carrageenan (1%). Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group, ### p < 0,001 JR19 versus o JR19 + L-NAME (ANOVA followed by Dunnett's test).
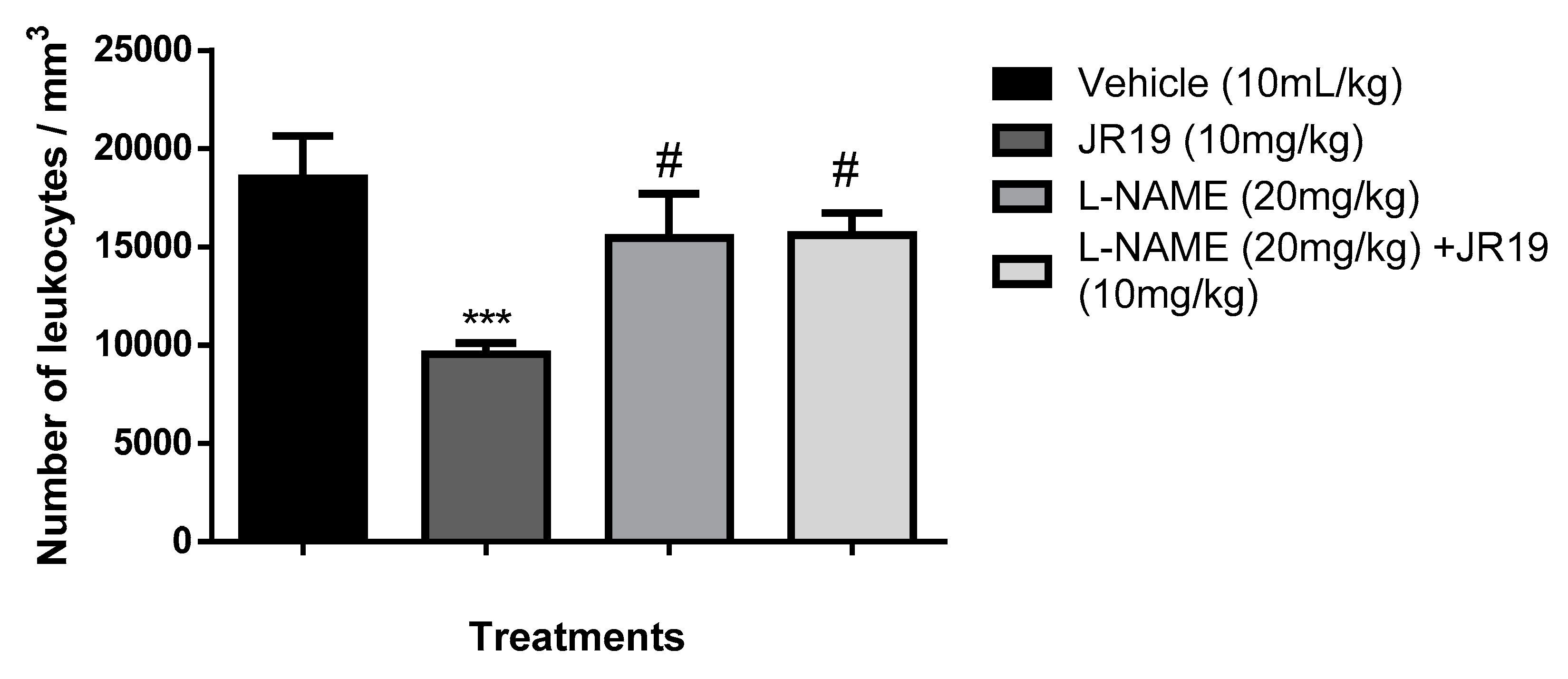
Figure 7.
Influence of treatment with the compounds JR19 and Methylene Blue on the total leukocyte count in the model of peritonitis induced by carrageenan (1%). Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group, ### p < 0,001 JR19 versus o JR19 + L-NAME (ANOVA followed by Dunnett's test).
Figure 7.
Influence of treatment with the compounds JR19 and Methylene Blue on the total leukocyte count in the model of peritonitis induced by carrageenan (1%). Values are expressed as mean ± standard deviation of mean, n = 6. *** p < 0.001 significantly different from control group, ### p < 0,001 JR19 versus o JR19 + L-NAME (ANOVA followed by Dunnett's test).
Figure 8.
Effect of JR19 compound and pretreatment with L-NAME or methylene blue (i.p.) administered 30 min before JR19 compound (p.o.) on the concentrations of cytokines (IL-6, TNF-α, IL-17, IFN-γ, IL-2, and IL-4) in the peritoneal exudate of mice. Values are expressed as mean ± SD, and asterisks indicate statistically significant differences compared with the negative control (saline): ***p<0.001, **p<0.01, *p<0.05. Analysis of variance (ANOVA) followed by Bonferroni test.
Figure 8.
Effect of JR19 compound and pretreatment with L-NAME or methylene blue (i.p.) administered 30 min before JR19 compound (p.o.) on the concentrations of cytokines (IL-6, TNF-α, IL-17, IFN-γ, IL-2, and IL-4) in the peritoneal exudate of mice. Values are expressed as mean ± SD, and asterisks indicate statistically significant differences compared with the negative control (saline): ***p<0.001, **p<0.01, *p<0.05. Analysis of variance (ANOVA) followed by Bonferroni test.
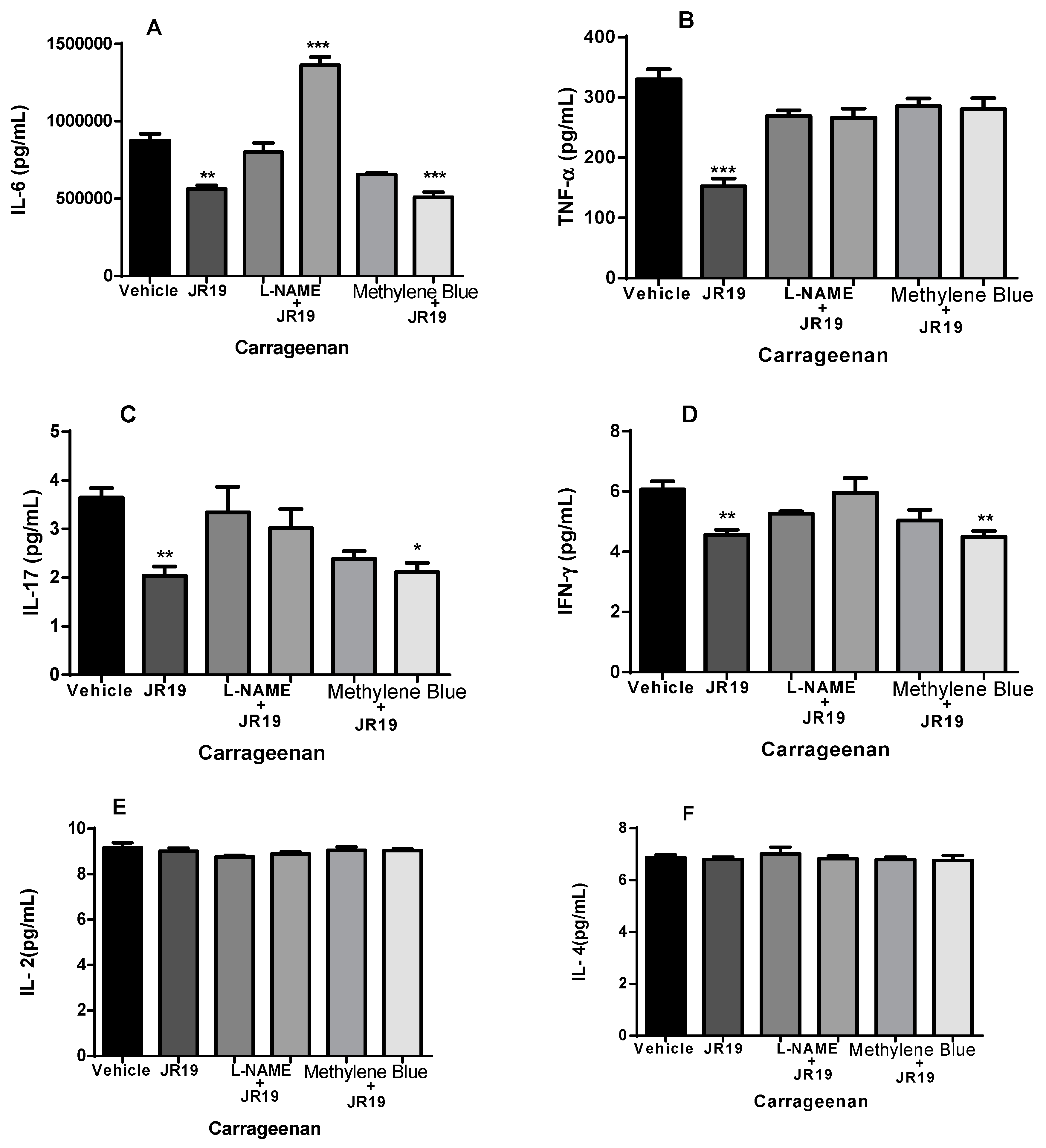

Table 1.
FitScore values of target-ligand complexe:.
| Molecular Targets | ||||
|---|---|---|---|---|
| Compounds | sGC | NFκB/p65 | iNOS | RMSD (Å ) |
| JR19 | 55.2887 | 32.0025 (pose 1) 42.3261 (pose 2) |
74.6536 | - |
| AR-C95791 | 82.3597 | 0.3297 | ||
| L-NAME | 66.1212 | - | ||
| Nigakinone | 32.5710 (pose 1) 43.4235 (pose 2) |
- | ||
| Methylene blue | 46.9947 | - | ||
Table 2.
Interactions inside the binding site of the targets.
| Molecular Targets | ||||
|---|---|---|---|---|
| Compounds | sGC | NFκB/p65 (pose 1) | NF-κB/p65 (pose 2) | iNOS |
| JR-19 | Arg5521, Ile5281,3, Thr5271, Asn6051, Val4752,3 and Cys5332 | Glu3001, Lys3031,2 and Arg3043 | Arg3021, Phe3092, Met3132 and Lys3103 | Gln2631, Hem9011,2, Glu3771 and Pro3503 |
| AR-C95791 | Glu3771, Tyr3471, Asp3821, Trp3721, Hem9012,3, Phe3693, Val3523 and Pro3503 | |||
| L-NAME | Glu3771,2, Gly3711, Tyr3731, Tyr3471, Trp3721, Phe3691,3, Pro3501,3, Hem9011,2, Val3523, Asp3823 | |||
| Nigakinone | Glu3002, Lys3031,2 and Arg3043 | Arg3021, Tyr3062 and His681 | ||
| Methylene blue | Asp5301, Asp4861, Asn6051, Ile5281, Glu5261, Leu5961, Ala5313, Cys5332, Phe4842 and Val4753 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



