
Submitted:
28 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Spinal cord injury is a traumatic injury that causes a catastrophic state in patients due to neuronal deficits, loss of motor and sensory function, and this is due to secondary deleterious events subsequent to mechanical damage that expands the death of neural cells beyond the initial trauma. One of the most important events is inflammation, which activates molecules like pro-inflammatory cytokines (IL-1beta, IFN-gamma, TNF-alpha) that provoke a toxic environment, inhibiting axonal growth and exacerbating CNS damage. As there is no effective treatment, one of the strategies developed is neuroprotection to preserve healthy neural tissue and reduce neuroinflammation. Some of the neuroprotective treatments that have been practiced are the use of cell therapy, the administration of peptides and molecules or supplements that have been shown to favor an anti-inflammatory environment, and help to preserve the cells and tissues surrounding the injured area, favoring axonal growth and improving locomotor function. In this review we will explain some of these strategies used in different animal models of spinal cord injury, their activity as modulators of the immune system and the benefits they have shown.
Keywords:
spinal cord injury
; cell therapy
; neuroprotection
; supplements
; peptides
; inflammation
1. Introduction
Spinal cord injury (SCI) can occur at any level or segment [1]. The pathological events originated after injury cause two types of damage, the first, refers to direct mechanical injury of the spinal cord that anatomically, causes contusion, compression, hemisection or complete transection of the spinal cord. These inductors of damage have similar self-destructive mechanisms, but their progression and complications are different [2]. In general the primary injury, directly imparts force to the spinal cord, disrupting axons, blood vessels, and cell membranes. This primary damage eventually triggers a cascade of harmful events like vascular dysfunction, edema, ischemia, excitotoxicity, free radical production, inflammation, and delayed apoptotic cell death that, on a period of time extend tissue damage causing the secondary injury. Whereas neurological deficits are, present immediately following the initial injury, the secondary injury results in a protracted period of tissue destruction.
On the other hand and based on time after-injury and pathologic mechanisms, this injury process can be divided into acute, subacute (or intermediate) and chronic phases. In each one, there are diverse destructive mechanisms with a different degree of damage that, determine neuronal destruction and allow the possibility of finding therapeutic targets to promote neuroprotection or neuroregeneration [3]. One of the most deleterious phenomena after injury is the inflammation. The early acute phase involves infiltration of inflammatory cells and continue activation of resident microglia. The inflammatory process following SCI is highly complex and involves numerous cellular populations, including astrocytes, microglia, T cells, neutrophils, and peripheral monocytes that are known to participate in the major inflammatory reactions. In this acute phase of injury, various inflammatory events contribute to neuronal and glial destruction. These phenomena are initiated by the primary insult, which leads to the release of significant amounts of DAMPs (damage-associated molecular patterns) and continue into the secondary phase of damage. This inflammatory reaction involved the activation of both resident and peripheral cells, such as systemic macrophages and neutrophils. These cells access the site of the lesion through locally produced chemokines and ICAM (intercellular adhesion molecules). Additionally, resident cells such as microglia and astrocytes play a role in the acute and subacute phases, through their activation towards inflammatory phenotypes, which is characterized by the production of proinflammatory molecules [proteases, myeloperoxidase, reactive oxygen (ROS) and nitrogen (RNS) species, inducible nitric oxide synthase (iNOS), nitric oxide (NO), macrophage inflammatory protein 1 (MIP 1 α, γ, β), monocyte pro-chemoattractant-1 (MCP-1), C-X-C motif chemokine ligand-10 (CXCL-10), IL1α/β, tumor necrosis factor- alpha (TNFα), IL-6] and the presentation -by major histocompatibility complex II (MHC II)- of antigens derived from the central nervous system (CNS) like myelin basic protein (MBP). This presentation phase exacerbates the inflammatory response by promoting the activation of helper T cells to the Th1 phenotype -which release other additional proinflammatory cytokines (TNFβ, IFNγ, IL-12) and B cells, leading to the production of IgG and IgM antibodies. Both T cells and B cells will favor the increase in the inflammatory response through the release of inflammatory cytokines, which further differentiate microglia and macrophage cells into M1 inflammatory phenotypes. These cells, promotes lipid peroxidation due to excessive NO and superoxide anion production, resulting in the generation of a neurotoxic compound called peroxynitrite, responsible, in part of Wallerian degeneration[4,5,6].
In relation to this, chemokines direct immune cells to the site of injury and activate a signaling cascade that increases the permeability of neuronal cell membranes and the release of glutamate. Over activation by glutamate, also known as "excitotoxicity," results in increased calcium influx, triggering free radical production and cell death by pro-apoptotic reactions. Cell death is not limited to damaged neurons but can also affect surrounding myelin-producing oligodendrocytes and healthy neurons, affecting up to four segments of the trauma site [7].
The physiological response after injury intends to resolve the inflammatory response through the release of anti-inflammatory cytokines such as IL-10 and TGF betta, by reactive astrocytes. Nevertheless, far from solving the problem, these cytokines stimulate the proliferation of astrocytes, leading to the formation of a glial scar. Additionally, these cytokines could contribute to fibroblast proliferation and collagen production. During the chronic phase, the glial scar indirectly forms a barrier at the lesion site preventing neuronal reconnection and axonal conduction. In this conditions, other molecules like growth factors [brain-derived growth factor (BDNF), neural growth factor (NGF), epidermal growth factor (EGF), insulin growth factor (IGF)] and neurotrophins (NT3, 4 and 5), are produced with the aim of promoting neural restoration, they are mainly produced by the M2 phenotype of macrophages and microglia and the Th2 lymphocytes. Unfortunately, these cells are not stimulated at the optimal time and quantity to promote their beneficial effects. In contrast, in the center of the injury, the M1 and Th1 phenotypes predominate activated causing more tissue damage [4,5,6] ( Figure 1).
Recently, dual effects (detrimental and beneficial) of neuroinflammation have been reported [4,8]. Inflammation is a process of enormous complexity, with some mechanisms detrimentally contributing to further secondary damage, and others -if it is modulated- contributing beneficially to the protection and restoration of SCI [9,10]. For this reason, diverse therapeutic strategies are directed to modulate or even inhibit this inflammatory response.
Various pharmacological strategies have been developed to reduce, modulate or inactivate inflammation after injury; however, the results have not been totally satisfactory in inducing a better functional recovery. For these reason, other therapeutic alternatives continue to be sought. In this line, non-pharmacological strategies have also been a promising alternative, in order to modulate the inflammatory response on the different phases of SCI (see Figure 1). Some of the most studied strategies include the cell therapy, the use of immunomodulatory peptides and supplements (including vitamins, minerals, and probiotics). In this review we analyzed the most important of them.
2. Cells Useful in Therapy for Spinal Cord Injury
Cell transplantation has been used to repair the injured spinal cord both in the acute and chronic phases, and the use of allogeneic transplantation including different cell types is of great interest, as it is a positive alternative for axonal regeneration. In line with this, Schwann cells (SCs), olfactory glia ensheathing cells (OECs), hematopoietic embryonic stem cells, neural stem cells and bone marrow stromal cells [11] have been used. The importance of this therapy is the different mechanisms that favour an improvement in function after damage, including neuroprotection, immunomodulation, axonal branching, axonal regeneration, and axonal re-myelination [12].
2.1. Schwann Cells
Schwann cells (SCs) are part of the peripheral nervous system (PNS), which is responsible for forming the myelin sheaths on peripheral axons [13], unlike oligodendrocytes in the CNS, SCs have a role in the repair of nervous tissue. After an injury; they can change their phenotype from myelinating to phagocytic and recruit neutrophils and macrophages through chemotactic signaling, which is key to the regenerative capacity of PNS [14]. This is why SCs have a strong relationship with the immune system [15], since they are modulated through an exchange of different signals.
In PNS injuries SCs cells have been observed to be able to modulate macrophages and mediate their transition from M1 to M2, although not through classical M2-mediated cytokines [16], as during the peak of debris clearance this macrophage phenotype is increasing due to elevated arginine expression [17].
The SCs are capable of both recognizing and presenting antigens, as well as expressing many cytokines, chemokines and immune modulatory factors and responding to them through the expression of their receptors [15,18]. The Toll-Like Receptors (TLRs) enable the expression of TNF-α, iNOS and MCP1, which promotes macrophage recruitment. Pattern Recognition Receptors (PRR) have been shown to induce the immune response by modulating T lymphocytes through the presentation of antigens in the context of MHC II. On the other hand, factors such as NGF that are secreted by SCs through the p75/AMPK/mTOR receptor promote increased autophagy of myelin debris [19].
Transplantation of SCs can depend on the stage of spinal injury, and achieve immunomodulation and neuroprotection of the resident tissue [20,21]. In animal models with a spinal contusion, SCs transplantation has been shown to reduce cystic cavities and increase neuronal marker (NeuN+) [22]. In the subacute phase, SCs also decrease the number of cells expressing CD11b+, CD68+ and Iba1+ cells, reducing the number of proinflammatory macrophages/microglia [23]. Mousavi et al. tested the neuroprotective potential of SCs by combating inflammasome activation, demonstrating improved motor function and re-myelination, because SCs reduced levels of NOD-like receptor Pyrin domain containing 1a (NLRP1a) and NLRP3 (inflammasome activators), caspase 1 (Casp1), TNF-α, and IL-1β [24].
SCs have a role in remyelination and neurogenesis during SCI repair. Isolated SCs retain the functional capacity for nerve repair and remyelination [25]. After SCs are transplanted into the injured CNS, they can reduce the size of lesions, attract and guide descending and ascending axons to the implant, and improve locomotor function without additional interventions [25]. In addition, it has been shown that co-transplantation of SCs with neural precursor cells, derived from human embryonic stem cells, could offer a synergistic effect and promote neuronal differentiation and functional recovery [26].
2.2. Olfactory Ensheathing Cells
OECs are glial cells that envelop bundles of olfactory axons, both peripherally in the olfactory nerve and within the olfactory nerve layer (ONL) of the olfactory bulb [27]. The OECs play a critical role in the growth of neurites and the establishment of functional connections along the olfactory plaque where new olfactory sensory neurons are generated from olfactory epithelial stem cells [28]. These cells can respond to invasion or damage and modulate the immune system, releasing IL-10 and TGF-β promoting phagocytosis via integrin receptors [29], and having receptors that help mediate the release of cytokines such as IL-6, ciliary neurotrophic factor (CNTF) and leukemia inhibitory factor (LIF) [29,30].
Several studies are using OECs as a treatment for SCI, in different animal models, not only because they can modulate the inflammation of the injury, but also due to their regenerative capacity [31]. In transection models of SCI, transplantation of OECs decreases immune cell infiltration by interacting with astrocytes via the signal transducer and activator of transcription 3 (STAT3) pathway, reducing the cystic cavity and protecting and preserving axons [32]. They can also induce the polarization of microglia from M1 to M2 by inhibiting the JAK/STAT3 pathway, and they reduce the population of the proinflammatory iNOS+/CD86+ phenotype and increase the anti-inflammatory microglia/macrophage phenotype Arginase+/CD206+ [29]. Moreover, they can reduce TNF-α, IL-1β, and oxidative stress and elevate IL-10 to protect nerve tissue [33]. In addition, OECs can phagocytose myelin debris, increasing neuronal survival via the p38/MAPK pathway [34], and even transplantation of their exosomes alone promotes the M1/M2 switch of microglia/macrophages inhibiting the nuclear factor kappa beta (NFkB) pathway [35]. Lopez-Vales et al. observed that transplanted OECs modulated the early astrocyte response from A1 to A2 astroglial phenotype. The transplanted OECs reduced the inflammatory peak to stop the earlier inflammatory phase and glial scarring, thus slowing down degeneration and preventing the spread of spinal cord damage [36].
In the subacute phase, studies have shown that they can reduce the number of astrocytes and macrophages by decreasing infiltration, reducing CCL2/3 chemokine levels, modifying cytokine levels by increasing anti-inflammatory cytokines such as IL-10/13 and decreasing inflammatory IL-6 and TNF-α. Additionally, OECs are capable of modifying the phenotype of astrocytes and macrophages, down-regulating the number of iNOS+/CD16+/32+ macrophages and increasing Arginina+/CD206+ through the secretion of IFN-γ to IL-4 [37].
2.3. NG2 Glia or Oligodendrocyte Precursor Cells
NG2 glia, also known as oligodendrocyte progenitor cells (OPCs), are located throughout the CNS and serve as a reservoir of progenitors to differentiate into oligodendrocytes. In response to SCI, NG2 cells increase their proliferation and differentiation into myelinating oligodendrocytes [38]. However, post-traumatic endogenous remyelination is rarely complete, and a better understanding of the characteristics of OPCs and their manipulation is critical for the development of new therapies [39,40]. The role of these cells in neuroprotection and modulation of the immune system in injury has been studied, but only their endogenous effect.
In IL-1β -/- Knockout mice a demyelination process came up and OPCs are not capable to differentiate and maturate into myelinating oligodendrocytes, what means that IL-1β is required for OPCs differentiation, also in contusion models, (C-X-C chemokine receptor type 4) CXCR4 and C-X-C Motif Chemokine Ligand 12 (CXCL12) expression is upregulated in astrocyte-like cells [41]. Although activation of CXCR4 by CXCL12 promotes differentiation of OPCs under demyelinating conditions [42], despite this result, the functional significance of these changes following injury is still unknown. Finally, IL-17 increases the production of IL- 6 and matrix metalloproteases (MMP3, MMP9) by OPCs in vitro, suggesting that OPCs may be directly involved in regulating the inflammatory response to injury [43].
The reduction in the accumulation of activated microglia and macrophages at the site of injury decreases the proliferative response of OPCs to the site of injury by eliminating β-catenin specifically in OPCs [44], raising the possibility that can attract leukocytes and monocytes to the site of injury. The OPCs were also found to be important for the maintenance of microglial homeostasis, because in their absence microglia was driven towards a reactive phenotype [45]. In addition, they regulate innate immunity as they can secrete TGF-2 which inhibits microglia activation via the SMAD2 (mothers against DPP homolog 2) pathway [37].
2.4. Bone Marrow-Derived Mesenchymal Stem Cells
The bone marrow-derived mesenchymal stem cells (BMSCs) are adult multipotent cells that have the capacity for self-renewal, proliferation, and differentiation. BMSCs are an alternative for the experimental treatment of SCI (contusions, complete sections, or ischemia). They have been shown to promote axonal regeneration and improvement in locomotor function; [46,47,48,49]. They also can form bundles of cells that serve as a bridge at the epicentre of the SCI [46,50,51]. In consonance with the context, they can differentiate both in vitro and in vivo into cells that express markers related to: astrocytes, oligodendrocytes, SCs, microglia and neurons [52,53]. In its niche it integrates endocrine, autocrine, and paracrine signals by secreting factors that control hematopoietic cells, keeping them in an undifferentiated state by secreting stem cell trophic factor (SCF), stromal cell factor (SDF-1 or CXCL12), bone morphogenetic protein 4 (BMP-4), LIF and granulocyte monocyte colony-stimulating factor (GM-CSF) [54].
In contusions and compressions models of SCI, the use of BMSc favors the decrease of proinflammatory cytokines such as TNF-α and IL-6; and promotes the secretion of anti-inflammatory cytokines such as IL-4, IL-10 and IL-13, which favors the activation of M2 macrophages, supporting a neuroprotective environment [55,56]. They can increase the number of M2 macrophages and decrease M1 macrophages at the site of injury, which may contribute to improved function by protecting healthy axons at the site of injury [13].
BMSCs have been shown to inhibit T-cell activation division by down-regulating cyclin D2, promoting the expression of p27Kip1 in the G1 phase of the cell cycle. The inhibitory effect on T cells is mediated by the secretion of TGF-β, hepatocyte growth factor and prostaglandin E2. In addition, BMSCs secrete soluble factors as inflammatory mediators such as indoleamine 2-3 dioxygenase (IDO), iNOS, and homo-oxygenase 1; they can also secrete human leukocyte antigen G, and are involved in the inhibition of CD4+ CD8+ T cells, by inhibition of STAT5 phosphorylation [13,54,57]. They can not only influence the switch from Th1 to Th2, but also decrease the Th17/Treg ratio, accompanied by a decrease in TNF-α and IL-1β, increasing TGF-β levels [13]. Finally, BMSCs have been reported as a neuroprotective capacity following injury as a positive strategy for CNS repair.
2.5. Neural Stem Cells
Neural stem cells (NSCs) are progenitor cells located in the lateral ventricle of the brain, dentate gyrus of the hippocampus and central canal of the spinal cord [58]. They are capable of self-renewal and differentiation into oligodendrocytes, astrocytes and neurons [59]. The main mechanism of these cells in neurodegenerative diseases is the modulation of astrocytes on glial scar formation, promoting differentiation to oligodendrocytes and neuronal differentiation, nerve cell replacement after SCI and secretion of pro-regenerative factors to protect damaged tissue by promoting neuritic growth [58].
The NSCs can secrete multiple growth factors, such as BDNF, CNTF, glial derived neurotrophic factor (GDNF), NGF and IGF-1, which contribute to the survival and growth of neuronal cells. After injury, they regulate T cells and macrophages to inhibit demyelination by reducing the number of CD4+ T cells, favoring a regenerative phenotype shift [58,59].
In addition, they have been shown to suppress the accumulation of neutrophils and macrophages at the site of injury, causing M1 macrophage activation to be blocked and a decrease in the secretion of pro-inflammatory cytokines such as TNF-α, IL-6, IL-1β and iNOS. This change in M1 macrophage polarization is due to the secretion of factors such as type 1 metalloproteinase (TIMP-1), vascular endothelial growth factor (VEGF), TGF-β, MMP9 and haptoglobin [59]. They have also been found to affect neuronal apoptosis by reducing glutamate exposure, decreasing pro-apoptotic markers and increasing anti-apoptotic markers such as B-cell lymphoma 2 (Bcl-2) [60].
2.6. Neural Precursor Cells: Aldynoglia
In the spinal cord and brain, we can find aldynoglia cells that make up the ependymal layer of the spinal cord. Within these, it could present tanycytes, ependymal cells and central canal neurons in an ependymal layer of the spinal cord, called spinal ependymal layer (SEL) [61].
In the subacute phase, SEL migrates to the site of injury to begin proliferation and differentiation and may also interact with immune response cells. The polarization of M1/M2 macrophages/microglia at the site of injury has been shown to directly affect the growth and differentiation of ependymal stem cells (EpSCs); specifically, M2 polarization promoting neuronal differentiation [62].
The mechanism by which M2 regulates the differentiation of EpSCs is through (Sirtuin-2) SIRT2. SIRTs, NAD+-dependent diacetylated histone class III, are involved in catalysing several biological processes, such as metabolism and gene expression [61,62]. In vivo and in vitro studies have shown that M2 microglia promote SIRT2 upregulation of EpSCs, directly affecting their differentiation; this change acts directly on microtubule dynamism thus promoting their differentiation into neurons. This change is produced by growth factors such as BDNF, which is secreted by M2 microglia, activating the BDNF/tropomyosin receptor kinase B (TrkB) signalling cascade, when the TrkB tyrosinase B on the surface of EpSCs is activated, inducing the expression of SIRT2 [61,62].
The compound of immune responses and effects after SCI of all the types of cells mentioned are summarized on Table 1.
The neuroprotective and neurorestorative effects shown by cell therapy after SCI are encouraging, however each type of cell needs a specific microenvironment to exert its beneficial effects or induce changes of immune cell phenotypes as mentioned in this section, however, to enhance this attribute the use of immunomodulatory peptides, supplements, probiotics or prebiotics could also be considered in a combined therapy.
3. Immunomodulatory Peptides after Spinal Cord Injury
The use of peptides as a therapeutic strategy after SCI has shown encouraging results. The findings have encourage the search for a different innovative therapy where the inflammatory response and the promotion of neuroregeneration are the main blank. Peptides, currently have demonstrated high activity, safety, low cost, and easy production [63]. An increasing number of active peptides have been studied, including antioxidants, analgesics, immunomodulators, among others. For the purpose of this review, we focus on some non-myelin or myelin-related peptides.
3.1. Non-Myelin Related Peptides
3.1.1. Glutathione Monoethyl Ester (GME)
Glutathione is a tripeptide formed by the amino acids glycine, cysteine and glutamic acid. It is produced by the liver and is involved in many bodily processes. Glutathione functions as a cell reducer, as a catalyst in various biological and metabolic reactions, and in the protection against ROS and toxic compounds of endogenous and exogenous origin [64]. The possibility that an increase in cellular glutathione may be beneficial under certain conditions in order to protect cells against oxidative stress considers it a potent antioxidant, however its depletion is significant when used and its recovery and transport to cells is limited. Glutathione monoethyl ester (L-γ-glutamyl-L-cysteinyl glycine ethyl ester) is efficiently transported to many cells, and glycine carboxylic ester is deesterified intracellularly in cells and rapidly converted to glutathione using approximately 90% of glutathione monoethyl ester administered and subjected to deesterification after 30 minutes, obtaining a high bioavailability as an antioxidant in this [64]. Glutathione in its reduced form (GSH) is quantitatively the most important endogenous rechargeable antioxidant. It has also been shown that it acts as a vasodilator under conditions of oxidative stress that alters endothelial function, thus improving cell ischemia [65]. In addition, it has several important functions related to amino acid transport across membranes, protein synthesis and degradation, gene regulation, and cellular redox regulation [64]; by the state of oxidative stress that occurs after CNS trauma [66]. Reduced glutathione monoethyl ester (GSHE) is permeable to cells and efficiently transported into the cerebrospinal fluid [64], thus providing the most direct and convenient means available for increasing GSH concentration intracellular [67]. Some studies have evaluated the significant effect of GSHE on lipid peroxidation (LPO) after SCI [68] and on neuroprotection after transient focal cerebral ischemia. GSHE is capable of reducing infarct size from 46% to 16% [64].
After SCI, the use of GSHE was compared with methylprednisolone (MP). MP is currently the only approved acute treatment available for patients with SCI. The use of corticosteroids in the treatment of SCI has been widely studied. It is currently known that their administration could be beneficial due to the anti-inflammatory properties of these drugs, reducing the formation of edema generated in turn by the local inflammatory process [69,70]. MP exerts a neuroprotective effect by suppressing the tissue inflammatory response through inhibition of inflammatory cell function, including chemotaxis, phagocytosis, synthesis of inflammatory mediators, and lysosomal enzyme release [71,72,73]. Several clinical studies on the use of MP question the effectiveness of this treatment and the safety of long-term side events in patients, however, it is currently still used in common practice and for legal medical reasons it is the appropriate treatment [69]. To determine if GSHE has more beneficial effects than MP, GSHE was administered intraperitoneally to rats subjected to moderate SCI. Animals receiving GSHE showed significant recovery of motor function, increased postoperative body weight and survival of red nucleus neurons compared to MP or vehicle [74].
3.1.2. Monocyte Locomotion Inhibitory Factor
Another peptide used like a novel strategy is monocyte locomotion inhibitory factor (MLIF), which is an exogenous pentapeptide (Met-Gln-Cys-Asn-Ser) produced by Entamoeba histolytica that has a potent anti-inflammatory effect (Silva-Garcia & Rico-Rosillo, 2011). In vitro studies in peripheral blood cells (monocytes, neutrophils), cell lines of human promonocytes (U937) and fibroblasts (MRC-5), treated with MLIF, demonstrated that, the factor decreases or inhibits: 1) the expression of the chemokines CCL3, CCL4, CCL1 and the CCR1 receptor, which are mainly involved in the chemotaxis of monocytes, 2) the synthesis of ROS and RNS and 3) the production of IL-1β, IL-6, IL-12 and IFN-γ [75,76]. On the other hand, MLIF favors the synthesis of IL-10 and TGF-β and the translocation to the nucleus of the p50/p50 homodimer of NF- κB, suggesting that this translocation reduces inflammatory gene expression [76].
The neuroprotective effect of MLIF has been demonstrated in the C57BL/6 mouse model infected with Plasmodium berghei. In this case, the use of this peptide prevented the increase in TNF-α and IFN-γ, protecting the integrity of the blood-brain barrier and providing an increase in the survival of animals without showing signs of brain damage [77].
Lipid peroxidation is a neurodegenerative phenomenon that occurs after SCI, caused by increased production of ROS an RNS such as NO. Recently, in an experimental model of traumatic SCI with the application of intraspinal MLIF, the peptide caused a downregulation of the expression of the iNOS gene that was correlated with a lower systemic production of NO and lipid peroxidation. MLIF also increased the expression of neuroprotective anti-inflammatory cytokines such as IL-10 and TGF-𝛽, these findings correlated with the preservation of neurons of the ventral horn and rubrospinal tract and a significant improvement in neurological recovery [78].
3.2. Myelin Related Peptides
Several neuroprotective or neurorestorative strategies focus on preserving myelinated axons or inducing axonal growth to promote axonal reconnection and then motor and functional recovery. MBP is produced by oligodendrocytes and by SC. This protein is localized in the myelin sheath, which is a unique multilayered membrane that surrounds the axon of neurons in which, the cytoplasmic and extracellular regions accumulate alternately and compactly [79]. This unique myelin structure provides several neurological functions, including saltatory conduction, nerve metabolism, ion regulation, and water homeostasis [80,81]. After SCI, the process of lipid peroxidation and T-cell recognition of the MBP leads to axon demyelination. This canonic recognition of self-constituents, allows the participation of an autorreactive response in the neurodegenerative process observed after injury. The activation of T cells against neural antigens -specially against MBP- intensify the inflammatory response and thereby, the destruction of neural tissue. That is why, several therapeutic strategies were directed to inhibit these autorreactive and inflammatory responses. Nevertheless, as immune cells are necessary for protecting and remodeling tissues, the elimination of the immune function at the site of injury, was not the best strategy [82]. It was more realistic to look for the modulation instead of the elimination of immune function. For this reason, different neural-derived peptides were generated to modulate the autorreactive response. [83,84,85,86].
It is now accepted that a well-controlled inflammatory reaction after injury is essential for neuroprotection and regeneration. After SCI, a prolonged and uncontrolled response ensues, leading to a vicious cycle of glial priming that promotes neuronal damage. In 1999, the laboratory of Dr. Michal Schwartz reported that autoimmunity in the CNS could, under certain circumstances, protect injured neurons from the propagation of damage and facilitate regeneration processes in the injured spinal cord [85,86]. They proposed the new concept of protective autoimmunity (PA) to refer to the "autoimmune response that elicits CNS injury when local nonimmunologic protective mechanisms fail to adequately buffer the injury-induced toxicity" [85]. Later studies showed that the immunization with myelin-derived proteins or modified neural-derived peptides, could render significant effects in improving motor function after SCI [87,88].
Immunization with a weaker version of the autoantigen could generate adequate PA that, modulates inflammation and exerts beneficial actions. Much research has been done in recent years to develop these immunomodulatory peptides and their potential therapeutic applications. Glatiramer acetate (GA) and A91 have been used to induce PA in animal models after SCI with encouraging results alone or in combination with other treatments.
3.2.1. Glatiramer Acetate
Glatiramer acetate (GA), also called copolymer 1 (Cop-1), is a random copolymer of glutamic acid, lysine, alanine, and tyrosine with an average molar fraction of 0.141, 0.427, 0.095 and 0.0338, respectively. GA has been approved by the Food and Drug Administration (FDA) for the treatment of relapsing-remitting multiple sclerosis [89,90]. Its mechanism of action has not been fully elucidated, but it seems that GA has immunomodulatory effects and neuroprotective properties [91]. GA has been shown to exert an immunomodulatory effect on cells of the innate and adaptive immune systems by inhibiting the activation of MBP-reactive T cells and inducing an anti-inflammatory T cell milieu. Repeated immunization with glatiramer acetate may modulate the adaptive immune system by shifting from a proinflammatory Th1 immune response to an anti-inflammatory Th2 phenotype characterized by IL-4 secretion [90].
GA has been assumed to be an immunomodulatory agent with neuroprotective properties in various neurodegenerative diseases, including Alzheimer's disease, Parkinson's disease, and amyotrophic lateral sclerosis [92,93,94,95]. In addition, it is known that GA exerts an inhibitory effect on the microglia M1 phenotype and stimulate the M2. GA has been shown to protect neurons and oligodendrocytes and affect three characteristic processes of neurogenesis: neuronal progenitor cell proliferation, migration, and differentiation [91,96,97,98,99].
The effectiveness of GA in SCI seems to depend on the dose used, the time of administration, and the intensity of the lesion. The effect of GA treatment at low doses for 2 weeks (0.5 mg/animal/day) was studied in rats subjected to ventral lumbar root avulsion. The results indicated that GA treatment induced a 40% increase in neuronal survival in the motor nucleus of the spinal cord together with a reduction in astroglial reactivity at the site of injury [100].
On the other hand, the immunization with GA showed that it reduces lipid peroxidation. It is possible that GA could be interfering with NO production. To test this hypothesis, the effect of GA on the amount of NO and iNOS expression were evaluated on glial cells when co cultured with autoreactive T cells and cells from spinal cord of injured animals (mice and rats). In vitro studies showed that GA significantly reduced the production of NO by glial cells. This observation was substantiated by in vivo experiments demonstrating that immunization with GA decreases the amount of NO and iNOS gene expression at the site of injury, these results clarified a possible mechanism by which protective autoimmunity promotes neuroprotection [101].
In a more recent study, GA treatment was performed in female Sprague-Dawley rats using a high dose (2 mg/kg) for 28 consecutive days after injury. In contrast to other studies, the authors observed impaired locomotor recovery, increased neuronal loss in the acute phase after SCI, and a detrimental response against MBP [102].
The controversial results observed in the former studies could be caused by diverse factors. For instance, the intensity of injury plays a crucial role in the final outcome of injury. In line with this, a study with lesions of different intensities (moderate and severe), changes in the expression of eight different genes were shown: IL-6, IL-12, IL-1β, IFNγ, TNFα, IL-10, IL-4 and IGF-1. Sprague Dawley females were immunized with Cop-1 (150 μg/animal) after injury and changes in gene expression were obtained at 7 days [103]. It was shown that a moderate lesion allows Cop-1 to create a microenvironment where cytokines such as IL-4 and IL-10 prevail, which could play an important role in the protection and restoration of neural tissue [104,105]. Aside from this, a significant reduction of inflammatory cytokines such as TNFα, was observed [106,107]. In contrast, after a severe contusion, Cop-1 failed to induce the same effect.
In this context, compared to moderate SCI, severe injury causes a more pronounced release of DAMPsand neural components, so that the high concentration of these molecules could be modifying the immune response to a Th1 encephalitogenic phenotype [108]. This predominant phenotype , which could be directed against other immunogenic determinants, could also inhibit the proliferation of protective Th2 lymphocytes elicited by Cop-1 and, therefore, their beneficial actions [101].
3.2.2. A91 Peptide
A91 is a peptide derived from the immunogenic sequence (87–99) of MBP, in which the lysine residue at position 91 has been replaced with alanine. Inoculation of peptide A91 induces the proliferation of CNS antigen-specific T cells that exert protective actions through several mechanisms that promote neuroprotection [83,109].
To establish prophylactic therapy with A91, in a well-designed study, rats were immunized with the peptide prior to SCI. Immunization induced a significant increase in the survival of rubrospinal and ventral horn neurons. These results were correlated with a significant improvement in motor recovery [88].
Some of the mechanisms by which A91 immunization exerts its neuroprotective and neurorestorative properties have already been reported. Once presented to helper T lymphocytes (CD4+), A91 moderates the inflammatory process in the subacute phase of SCI by inducing Th2 lymphocytes, an anti-inflammatory phenotype [101]. The cytokines secreted by A91-activated lymphocytes are mainly IL-4 and IL-10, which reduce ROS levels. High levels of IL-10 not only reduce the synthesis of TNF-α in macrophages, but also participate in the inhibition of lipid peroxidation. On the other hand, IL-4 decreases the synthesis of IFN-γ, which is a cytokine that stimulates macrophages and microglia to acquire a proinflammatory M1 phenotype. Furthermore, IL-4 promotes arginase production, which removes arginine and thus reduces NO production by iNOS. Therefore, there is a lower number of nitrates in the region of the lesion, which inhibits the formation of RNS [8,110].
Another beneficial effect of A91 immunization after SCI has already been reported. Immunization with A91-peptide reduces apoptosis caused by traumatic injury. This reduction was accompanied by a significant decrease in caspase-3 (Casp3) activity and TNF-α concentrations [111].
In addition, it has been observed that A91-immunization stimulates the production of neurotrophic factors that are essential for the proper functioning of the CNS. As well, these molecules contribute to tissue protection, regeneration, neurogenesis and stimulation for the formation of new synaptic connections [112,113]. A91 induced a significant production of anti-inflammatory proteins associated with regeneration and a significant increase in neurogenesis in the chronic stages of the lesion [114]. It also generates an improvement in the long-term production of BDNF, NT3 and GAP-43. This favorable effect was associated with better motor and sensory recovery in the chronic stage of SCI [112].
This therapeutic strategy has yielded encouraging results in both acute and chronic SCI, the latter being a difficult condition to treat successfully. Despite the promising results previously described for moderate lesions, these beneficial effects were not observed in animals with severe SCI [103,112]. Previous research has shown that severe injury or excessive administration of A91 inhibits the beneficial action of protective autoimmunity [101,115]. Furthermore, A91-immunized animals subjected to severe SCI did not show significant IL-10 production. These results support the hypothesis that the beneficial effects induced by A91-immunization are avoided and an inflammatory microenvironment predominate in severe SCI. [103]. Immunization with A91 has been shown to have promising effects, however the final result can be potentiated by combining it with other therapeutic strategies.
3.2.2.1. Synergism of A91-Immunization with Other Strategies
Given the promising results of A91-immunization, it was interesting to find out if this potential therapy would be beneficial in people treated with MP. Investigations on the topic indicated that MP given immediately after SCI, if given in conjunction with vaccination, overrides the benefit of vaccination. However, if MP is administered immediately after injury and vaccination 48 hours later, MP does not reduce the beneficial effect of A91-immunization [74].
In another study, A91 immunization was combined with GSHE, an antioxidant agent that itself has been shown to exert neuroprotective effects after SCI [74]. In this case, the experiments demonstrated that after SCI, combination therapy induces better motor recovery, a greater number of myelinated axons, and better survival of rubrospinal neurons than immunization alone. GSHE contributes to the elimination of ROS, reducing lipid peroxidation levels without interfering in the therapeutic effect of A91 [109]. Consequently, the combination of A91 and GSHE was shown to have superior effects when administered immediately after SCI or even within the first 72 hours post-injury [115,116].
Recently, it was shown in this same SCI model that the use of MLIF in combination with other neuroprotective peptides, such as A91 and GSHE, contributes to promote a better neuroprotective effect by preserving the medullary parenchyma and axonal fibers. The combined strategy increased the number of motor neurons and decreased the presence of collagen promoting a better motor recovery in rats after SCI [117].
3.2.2.2. Use of A91- Immunization Combined with Cell Therapy
Immunization with A91 has been shown to provide neuroprotection in SCI [88,111]. However, the neuroprotective effect achieved by the immunization strategy is related to the genetic background of the animals (strains), the type of adjuvant, and the intensity of the lesion [118], important characteristic that could be consider for use of combination therapies.
Dendritic cell (DC) strategies help reduce the required antigen dose, while further limiting cross-presentation of other cell types for their antigen-presenting ability. For stimulation of an immune response, DC migrates to the spleen or draining lymph nodes. There, DC mount the immune response to antigens and can also mount the autoantigen-specific responses seen in autoimmunity [119,120]. Vaccination with spinal cord homogenate-A91-pulsed dendritic cells (hpDC) was reported to increase levels of BDNF, NT-3, IL-4, and IFN-γ at the injured site, as well as the supernatant of cultured T cells of the hpDC group which was significantly higher than that of the control groups [121] also promotes functional motor recovery [122] A91-pulsed dendritic cells improved the level of BDNF and NT-3 expression and exerted a neuroprotective effect and possible regeneration in an SCI mouse model [110]. Evidence has shown that a high concentration of neurotrophic factors in the SCI is beneficial for axonal growth and neuronal survival [123,124]. Under pathological conditions, T cells are a source of neurotrophic factors, and the release of these molecules can be significantly increased by antigen activation [125]. Some CNS growth factors have been reported to be continuously produced and released at the site of injury by activated T cells [126,127]. Further studies are needed to test this strategy in SCI rat models to compare the beneficial effect with another combination.
In the chronic phases, cell necrosis, glial reaction, and inflammation induce the appearance of glial cavities, cysts, and scars that interrupt the descending and ascending axonal tracts, preventing possible axon regeneration[122]. In the chronic phase of SCI, there is scar tissue that prevents the correct reconnection of axons by forming a physical and chemical barrier of sulfated proteoglycans, thus inhibiting the formation of growth cones and axonal prolongation [6,128,129,130]. Additionally, chronic SCI is considered a period of low activity with progressive decline, so a possible alternative to achieve axonal regeneration could be scar removal (SR), with the aim of restoring axonal connections and consequently synapses, in addition to restoring the conditions of an acute injury, such as the activation of protective autoimmunity, as well as the release of cytokines and neurotrophic factors [122,127,131]. Previous research has shown that SR or a bone marrow mesenchymal stem cell implant, used separately, promotes significant tissue restoration and motor recovery after SCI [55,130]. Considering the results obtained, the effect of combining A91-immunization with these alternative strategies to achieve a more efficient therapy was investigated. In a first approach, it was explored both immunization with the A91 peptide alone or in combination with RS, on locomotor recovery, gene expression associated with regeneration and cytokines, as well as the number of regenerating axons, in a model of chronic SCI. The results suggested that both treatments could substantially modify the non-permissive microenvironment that prevails in the chronic phase of SCI, providing the opportunity to promote greater motor recovery [132].
Subsequently, the investigation was directed to design a combination therapy that integrated, a fibrin glue matrix with mesenchymal stem cells, and A91-immunization 72 hours after SCI or in chronic phase (60 days after). The intervention on acute phases (72 hr) of combination strategy after moderate SCI, was the best strategy to promote motor and sensibility recovery at 60 days after treatment. In addition, significant increases in tissue preservation and axonal density were observed, suggest that these therapy exhibits potential effects on the protection and regeneration of neural tissue after acute spinal cord injury [133]. On chronic phases after a complete SCI transection model, the combination therapy promoted motor and electrophysiological recovery. In the same way the combined therapy was capable of inducing a permissive microenvironment for neuroregeneration [103].
In a subsequent study, the same combination was evaluated also in the chronic phase but in a SCI contusion model. The results in this case showed that the combination therapy was able to modify the non-permissive post-SCI microenvironment but was unable to induce adequate axonal regeneration or neurogenesis as observed after treatment with A91-immunization alone [132].
A91-immunization combined with other therapeutic strategies could be useful for other acute or chronic neurodegenerative and neuroinflammatory pathologies. However, at present, there is a lack of evidence on the beneficial effect of A91 for pathologies other than SCI. The chronology of A91-immunization research is summarized on Table 2.
The effectiveness of any therapeutic strategy also depends on other factors such as the nutritional status and the balance of intestinal microbiota. These elements could also determine the permissible microenvironment that will favor the protection and/or restoration of tissue after SCI. It is worth mentioning that the CNS, due to its metabolic role, added to the impossibility for controlling the inflammatory response, would require other external agents that would promote changes in the microbiota and its metabolites involved in neuroprotective and neurodegenerative processes. Various strategies have implemented the use of supplements and probiotics in animal models and in patients, which are described below.
4. Supplements and Probiotics as Therapeutic Strategy after Spinal Cord Injury-Inflammation
4.1. Supplements
The European Food Safety Authority (EFSA) defines food supplements as a “concentrated source of nutrients or other substances with nutritional or physiological effect that are marketed in dose form. A wide range of nutrients and other ingredients may be present in food supplements, including, but not limited to, vitamins, minerals, amino acids, essential fatty acids, fiber, and various plant and herbal extracts” [134].
Dietary supplements have been proposed for a variety of neurodegenerative diseases and post-traumatic conditions, as they have been studied for their anti-inflammatory and neuroprotective effects on the brain.
Supplement use is common in people with SCI, with multivitamins, calcium, and vitamin D being the most commonly used [135]. Currently, supplementation has also been tested to reduce inflammation induced by SCI. There are several animal and human reports exploring the beneficial effects of various supplements.
4.1.1. Animal Studies
4.1.1.1. Vitamins
Vitamin C (ascorbic acid): is a water-soluble vitamin and an essential nutrient found in high concentrations in the brain. It is well known for its antioxidant role in the brain and its catalytic effect on various enzymatic reactions [136].
Intracellular vitamin C plays important roles in the central nervous system, such as myelin formation, neurotransmission modulation, and antioxidant protection [137]. Part of the protective action of this vitamin in the CNS is because it promotes the removal of glutamate from the synaptic cleft and inhibits calcium channels. Regarding neuroinflammation, it reduces the production of inflammatory cytokines, IL-1B and TNF-a, which in turn increase glutamatergic neurotransmission [138].
In a comparative study in rats evaluating inflammatory markers after SCI, serum levels of TNF-alpha, IL-6, and nitrite were significantly reduced compared with the control group with administration of 100 mg/kg ascorbic acid for 45 consecutive days [139].
Vitamin E (α-tocopherol): It is known as the main fat-soluble antioxidant in the body. At the same that vitamin C, humans must obtain this vitamin through the diet since we are not capable of biosynthesizing it [140]. Vitamin E has the ability to reduce the neuroinflammatory responses of microglial cells, increases the number of oligodendrocytes [141] and decreases IL-1beta and TNF-alpha in the hippocampus of animal models [142]. After vitamin E supplementation for 14 days after SCI, there was an improvement in BBB scores (Basso, Beattie and Bresnahan scale), acting as inhibition of lipid peroxidation products such as thiobarbituric acid reactive substances and malondialdehyde [142]. The anti-inflammatory actions of vitamin C and E as antioxidants have been shown in several animal studies to lead to improved recovery of motor function after SCI. Although oral intake was not associated with a significantly different effect compared to no treatment, favorable results were observed with intraperitoneal administration of vitamin C. Simultaneous administration of combined vitamins C and E did not provide any additional therapeutic effect. The pharmacological action of these vitamins against free radicals and oxidative stress makes them potential candidates for pharmacological treatment of SCI, possibly limiting damage in the first phase and inhibiting the spread of damage during the second phase [138].
Supplementation with trace elements and vitamin E, maintains the number and function of T-lymphocytes after acute SCI. This therapeutic strategy also promotes motor recovery, which is probably due to its antioxidant effect [143].
4.1.1.2. Minerals
Zinc is present as a cofactor in more than 300 enzymes and is involved in remodeling mechanisms after injury in many types of tissue. Copper/zinc-dependent superoxide dismutase decreases oxidative stress and attenuates blood-brain barrier mediated by MMP9 matrix metalloproteinases. This is important, because ischemic and damaged neuronal tissue is affected by lipid peroxidation of the cell membrane leading to edema and local inflammation [144]. Adequate zinc concentrations can inhibit inflammation, promote autophagy, inhibit oxidative stress, protect injured neurons, and treat spinal cord injuries. However, if administered in high concentrations, zinc can impair neurons through excitotoxicity, inducing oxidative stress and impairing cellular energy production. Therefore, the appropriate concentration of zinc could be used in SCI, but this dose has yet to be further defined [145].
4.1.1.3. Trace Elements
Certain trace elements are involved in development and regeneration processes [144].
Selenium is an essential trace element, which participates in the structure of glutathione peroxidase and is widely known for its harmful effects in case of deficiency [147]. The selenocysteine (sec) containing group of selenoproteins contributes to protecting cells from oxidative damage, especially through the selenium-dependent families of glutathione peroxidases and thioredoxin reductases. Nanoparticles in rats with SCI have a neuroprotective effect and accelerate neuronal function by controlling the inflammatory response [148]. The essentiality of the selenoproteins GPX4, thioredoxin reductase 1 (TXNRD1) and thioredoxin reductase 2 (TXNRD2) for normal development and survival of neurons became well understood after studying the phenotypes of respective gene knockout mice, which resulted in failed embryonic development [149].
Copper is an essential trace element; it is an important catalyst for heme synthesis and iron absorption. After zinc and iron, copper is the third most abundant trace element in the body [150]. It serves as a catalytic factor in redox chemistry for biologically active proteins (e.g., superoxide dismutase, cytochrome c oxidase, and lysyl oxidase). All these proteins function to regulate growth, metabolism, and oxidative stress, aiding in the pathophysiology of SCI and possible neuroregeneration after injury [151].
Seeling et al 2020; in their 2019 study suggest a potentially favorable association between the use of copper, selenium in damaged cells and the potential for a better anti-inflammatory response. Also allowing a more reliable prognosis after SCI [151].
4.1.1.4. Omega-3 Fatty Acids
Polyunsaturated fatty acids (PUFA) include α-linolenic acid (ALA), stearidonic acid (SDA), eicosapentaenoic acid (EPA), docosapentaenoic acid (DPA) and docosahexaenoic acid (DHA) oils that contain these fatty acids (FA). These FA originate mainly from certain plant, algae and unicellular sources [152]. Essential FA have been reported to be important in membrane fluidity, inflammatory eicosanoids, and neural membrane oxidation [153].
DHA in a mouse model study by Marinelli et al (2019) confirmed and extended the beneficial effects of repeated systemic administration of DHA in the primary and secondary phases of SCI. DHA treatment was shown to improve motor and sensory function, exert neuroprotective action, modulate cell response to injury (apoptosis and survival) and inflammation in astrocytes and microglia, ultimately promoting spinal regeneration. In recent years, there has been a debate about whether the action of omega-3 fatty acids, mainly DHA, in neurodegenerative disorders exerts an anti-inflammatory, neuroprotective and pro-regenerative action. In line with this, it has already been shown that DHA is capable of exerting an anti-inflammatory effect not only in the acute phase of SCI, but also in the intermediate/chronic phases of SCI [154].
In SCI, apoptosis of oligodendrocytes and neurons leads to increased oxidative stress and inflammation. A common explanation for the anti-inflammatory effects of omega-3 PUFAS is that they compete with arachidonic acid (AA), the main representative of the omega-6 family of PUFAs. AA is the precursor of prostaglandins, thromboxanes, and leukotrienes, which have proinflammatory effects [155].
4.1.1.5. Antioxidants
Oxygen radical-induced lipid peroxidation is perhaps the most important deleterious phenomenon that develops due to inflammation after SCI. Therefore, various therapeutic strategies are aimed at neutralizing the harmful effect of this process [156]. Resveratrol is a naturally occurring polyphenol found in grapes and is effective in preventing redox spoilage. It has anti-inflammatory properties by altering arachidonic acid metabolism and inhibiting protein kinase, making it a possible therapeutic strategy in SCI [156].
4.1.1.6. Botanicals
Beta-carotene is a tetraterpenoid consisting of a C40 structure that includes two β-ionone rings. Along with lycopene, it is among the most frequently consumed dietary carotenoids in humans and also among the highest in plasma concentrations [159]. Beta-carotene could improve functional recovery and spinal histological changes, as well as inhibit oxidative stress and inflammation by inhibiting NF-KB in the spinal cord of SCI rats. Further clinical experiments or human studies are suggested [160].
Spirulina platensis is a microalgae that is most often used as a supplement. It contains carotenoids, phycocyanin, xanthophylls, and phycobilins that have antioxidant activity [161]. Spirulina platensis supplementation after induction of SCI significantly improves functional recovery. Significant ultrastructural improvement was observed with reduced progression of morphological damage by preserving the ultrastructure of the spinal cord from secondary injury. This suggests that Spirulina platensis could be used in SCI patients to induce functional recovery [1].
4.1.2. Human Studies
4.1.2.1. Vitamins
Vitamin D: is a generic term, since it refers to a group of fat-soluble compounds with a main chain of cholesterol rings [162]. Despite our growing understanding of the extraskeletal functions of vitamin D, this vitamin has been poorly studied in SCI. Low calcifediol (25-OHD) levels might be a marker of the severity of the inflammatory process rather than an accurate reflection of vitamin D status. Shortly after injury, people with SCI show acute inflammation as a result of trauma, which then becomes chronic. Although the impact of chronic inflammation on vitamin D status has never been reported, interpretation of 25-OHD levels in an SCI setting should consider the inflammatory status of the patient [163].
4.1.2.2. Omega-3 Fatty Acids
The use of Omega-3 PUFA could suppress the production of proinflammatory cytokines such as IL-6 and IL-1β and TNF-α in human models, in addition to suppressing the production of bone absorption in patients with SCI, however, until now there is not enough conclusive information available in patients with SCI. Sabour et al., in their study, demonstrated inconsistency in the reported effects of Omega-3 PUFA on proinflammatory cytokines, reporting that no benefit was found in reducing these cytokines [164].
4.2. Probiotics
The intestinal flora includes a heterogeneous group of bacteria that live in the gastrointestinal system and influence human health, especially through interaction with the immune system, skin, lungs, and brain. The microbiome-gut-brain axis is a well-established research term that includes connections of afferent and efferent neurons, endocrine and metabolic signaling that make the brain and gut connected. Modulation of neurotransmission in gut dysbiosis has been postulated to play a key role in the pathogenesis of CNS disorders. An alteration in the intestinal microbiome causes an increase in proinflammatory molecules, which translates into an alteration in the permeability of the blood-brain barrier. Spinal cord injuries disrupt the autonomic nervous system, they also cause dysbiosis, which is essentially an imbalance in the gastrointestinal tract that affects the gut microbiota [165].
Probiotics, which are regulated as dietary supplements and foods, consist of yeast or bacteria. They come in various presentations such as powder, tablets, and capsules, in fermented foods, dairy products, or drinks such as kombucha. Probiotics are available as a single microorganism or as a mixture of several species [166].
The use of probiotics as therapeutic strategies after SCI is also a topic under study.
4.2.1. Animal Studies
An animal study demonstrated that SCI in mice affected gastric permeability and induced gut dysbiosis, which in turn led to delayed locomotor recovery. Therapeutic management with commercially available probiotics resulted in improved locomotor recovery [167,168].
Recent data from rodents indicate that SCI causes gut dysbiosis that exacerbates intraspinal inflammation and injury pathology leading to poor recovery of motor function. Post-injury administration of probiotics containing various types of beneficial bacteria may partially overcome the pathophysiological effects of gut dysbiosis. Immune function, locomotor recovery, and spinal cord integrity are partially restored by a sustained regimen of oral probiotics. Recently, using a mouse model, Phylum Bacteroidetes and Phylum Firmicutes, the 2 major bacterial orders in the gut, were revealed to be inversely regulated by SCI 3 weeks after injury. Phylum Bacteroidetes decreased 30% and Phylum Firmicutes increased 250% relative to pre-injury values. Given that the bacterial orders Bacteroidales and Clostridiales together constitute >80% of all species residing in the gut, significant and long-lasting changes in their relative population densities after SCI are likely to influence numerous physiological processes. For instance, communication between the gut microbiota and gut-associated lymphoid tissue (GALT) immune cells produce cytokines and other metabolites that circulate and affect CNS function. Dysbiosis is associated with marked changes in the relative proportion of immune cells found in mesenteric lymph nodes and Peyer's patches. There was also an increase in the synthesis of inflammatory and immunoregulatory cytokines in GALT in parallel with changes in immune cell populations. Gut microbes also produce neuroactive metabolites (short-chain fatty acids) and neurotransmitters, which can affect central nervous system function by activating vagal afferent nerve fibers in the gut. There is scarce information about whether probiotics can confer neuroprotection or ameliorate a variety of comorbidities and neurological complications caused by traumatic SCI [167,169,170].
Studies that have sought to correct the gastrointestinal microbiota (GIM) have seen a decrease in the levels of inflammatory cytokines. GIM also plays an essential role in the production of neurotransmitters. Lactobacillus and Bifidobacterium, two genera of bacteria that decline after injury, play important roles in the production of neurotransmitters such as serotonin, dopamine, and 𝛄-aminobutyric acid [171].
Gut microbiota depletion significantly reduced glial cell line-derived neurotrophic factor (GDNF) expression in mice. Mice deficient in TLR-2 increased susceptibility to inflammation. This highlights the importance of the gut microbiota-TLR-2-GDNF axis in modulating the enteral nervous system and inflammation [172].
In a rat study by Lin et al 2021; early administration of Lactobacillus rhamnoides GG conditioned medium (LGG-CM) immediately after SCI reduced the extent of post-traumatic inflammation near the injury site and promoted recovery of locomotor function after SCI. This is because LGG-CM inhibits the NK-𝜅B pathway by reducing I𝜅Bⲁ phosphorylation, which promotes microglia/macrophage polarization to M2 and inhibits polarization to M1. In this way, LGG-CM may be a possible therapeutic supplement for neuroprotection after a SCI event [173].
4.2.2. Studies in Humans
Gungor et al 2016; conducted a clinical study to show that the gut microbiome in SCI patients was different compared to healthy adults. They tested 30 SCI patients with different types of bowel dysfunction and 10 healthy controls. Microbial patterns were taken from stool samples. Firmicutes and Bacteroides spp they are the most predominant phylum in the intestine. They ferment indigestible polysaccharides and generate metabolites that the host can use for energy. Among these, butyrate is the most pronounced single-chain fatty acid with modulatory effects on epithelial cell growth and differentiation and immune function, has a potent anti-inflammatory effect on macrophages, and suppresses ongoing inflammation in the CNS. In this study, there was a significant reduction in butyrate in SCI patients, suggesting that reduced butyrate levels may contribute to microglia-mediated neurotoxicity in these patients, implying that low butyrate levels may have an impact in long-term recovery after SCI [174]. Therefore, supplementation with butyrate-increasing probiotics could be a therapeutic strategy for SCI.
5. Conclusions
Spinal cord injury continues being a health problem for which, there is no fully effective treatment. Pharmacological and non-pharmacological strategies have been evaluated and have provided different expectations to improve the neurological recovery of injured individuals. Nevertheless, due to the complexity of the events generated after injury -particularly the one presented by the inflammatory response- it has not been easy to establish an effective therapy that may help neuroprotection or promote neuronal regeneration. The approach for modulating the inflammatory response has been a hard topic of investigation. The use of cell therapy, immunomodulatory peptides or even diverse supplements as modulators of immune response, has provided a hopeful light in the way of finding the best strategy to neutralize the harmful effects of inflammation. Nonetheless, the way to find the final goal is still long and needs more investigation in order to obtain better results at a baseline level that could scale to clinical application.
Author Contributions
EG: Conceptualization, writing—original draft preparation, writing—review and editing, visualization, supervision; VBA: Conceptualization, writing—original draft preparation , writing—review and editing, visualization, supervision; RSG: Conceptualization, writing—review and editing, FES: writing—review and editing; AAC: writing—review and editing; AE: writing—review and editing; EDP: Conceptualization, writing—original draft preparation, review and editing, project administration, funding acquisition; AI: Conceptualization, writing—original draft preparation, writing-review and editing, project administration, funding acquisition. All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding and the APC was funded by Universidad Anáhuac México.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not aplicable.
Acknowledgments
This review is in accordance with the Framework Agreement for Research Collaboration, signed between the Castilla-La Mancha Health Service (SESCAM), Spain; and Universidad Anáhuac, México (July, 2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abdullahi, D.; Ahmad Annuar, A.; Sanusi, J. Improved Spinal Cord Gray Matter Morphology Induced by Spirulina Platensis Following Spinal Cord Injury in Rat Models. Ultrastruct. Pathol. 2020, 44, 359–371. [Google Scholar] [CrossRef]
- Oyinbo, C.A. Secondary Injury Mechanisms in Traumatic Spinal Cord Injury: A Nugget of This Multiply Cascade. Acta Neurobiol. Exp. (Wars). 2011, 71, 281–299. [Google Scholar]
- Rowland, J.W.; Hawryluk, G.W.J.; Kwon, B.; Fehlings, M.G. Current Status of Acute Spinal Cord Injury Pathophysiology and Emerging Therapies: Promise on the Horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed]
- Beck, K.D.; Nguyen, H.X.; Galvan, M.D.; Salazar, D.L.; Woodruff, T.M.; Anderson, A.J. Quantitative Analysis of Cellular Inflammation after Traumatic Spinal Cord Injury: Evidence for a Multiphasic Inflammatory Response in the Acute to Chronic Environment. Brain 2010, 133, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Popovich, P.G. Inflammation and Its Role in Neuroprotection, Axonal Regeneration and Functional Recovery after Spinal Cord Injury. Exp. Neurol. 2009, 209, 378–388. [Google Scholar] [CrossRef]
- Fitch, M.T.; Silver, J. CNS Injury, Glial Scars, and Inflammation: Inhibitory Extracellular Matrices and Regeneration Failure. Exp. Neurol. 2008, 209. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after Spinal Cord Injury: A Review of the Critical Timeline of Signaling Cues and Cellular Infiltration. J. Neuroinflammation 2021, 18, 284. [Google Scholar] [CrossRef]
- Dietrich, W.D.; Chatzipanteli, K.; Vitarbo, E.; Wada, K.; Kinoshita, K. The Role of Inflammatory Processes in the Pathophysiology and Treatment of Brain and Spinal Cord Trauma. Acta Neurochir. Suppl. 2004, 69–74. [Google Scholar] [CrossRef]
- Schiwy, N.; Brazda, N.; Müller, H.W. Enhanced Regenerative Axon Growth of Multiple Fibre Populations in Traumatic Spinal Cord Injury Following Scar-Suppressing Treatment. Eur. J. Neurosci. 2009, 30, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, A.; Olivas, A.D.; Noble-Haeusslein, L.J. Inflammation and Spinal Cord Injury: Infiltrating Leukocytes as Determinants of Injury and Repair Processes. Clin. Neurosci. Res. 2006, 6, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Watt, F.M.; Driskell, R.R. The Therapeutic Potential of Stem Cells. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2010, 365, 155–163. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell Transplantation Therapy for Spinal Cord Injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.M.; Chen, S.Y.; Xu, Q.J.; Fu, S.P.; Yang, Y.C.; Zou, W.H.; Zhang, M.; Liu, J.; Wan, W.H.; Peng, J.C.; et al. Neuroinflammation and Scarring After Spinal Cord Injury: Therapeutic Roles of MSCs on Inflammation and Glial Scar. Front. Immunol. 2021, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- El Seblani, N.; Welleford, A.S.; Quintero, J.E.; van Horne, C.G.; Gerhardt, G.A. Invited Review: Utilizing Peripheral Nerve Regenerative Elements to Repair Damage in the CNS. J. Neurosci. Methods 2020, 335. [Google Scholar] [CrossRef]
- Tzekova, N.; Heinen, A.; Küry, P. Molecules Involved in the Crosstalk Between Immune- and Peripheral Nerve Schwann Cells. J. Clin. Immunol. 2014, 34, 86–104. [Google Scholar] [CrossRef] [PubMed]
- Painter, M.W.; Brosius Lutz, A.; Cheng, Y.-C.; Latremoliere, A.; Duong, K.; Miller, C.M.; Posada, S.; Cobos, E.J.; Zhang, A.X.; Wagers, A.J.; et al. Diminished Schwann Cell Repair Responses Underlie Age-Associated Impaired Axonal Regeneration. Neuron 2014, 83, 331–343. [Google Scholar] [CrossRef]
- Stratton, J.; Shah, P. Macrophage Polarization in Nerve Injury: Do Schwann Cells Play a Role? Neural Regen. Res. 2016, 11, 53. [Google Scholar] [CrossRef]
- Meyer Zu Hörste, G.; Hu, W.; Hartung, H.-P.; Lehmann, H.C.; Kieseier, B.C. The Immunocompetence of Schwann Cells. Muscle Nerve 2008, 37, 3–13. [Google Scholar] [CrossRef]
- Li, R.; Li, D.; Wu, C.; Ye, L.; Wu, Y.; Yuan, Y.; Yang, S.; Xie, L.; Mao, Y.; Jiang, T.; et al. Nerve Growth Factor Activates Autophagy in Schwann Cells to Enhance Myelin Debris Clearance and to Expedite Nerve Regeneration. Theranostics 2020, 10, 1649–1677. [Google Scholar] [CrossRef]
- Orr, M.B.; Gensel, J.C. Spinal Cord Injury Scarring and Inflammation: Therapies Targeting Glial and Inflammatory Responses. Neurotherapeutics 2018, 15, 541–553. [Google Scholar] [CrossRef]
- Fu, H.; Hu, D.; Chen, J.; Wang, Q.; Zhang, Y.; Qi, C.; Yu, T. Repair of the Injured Spinal Cord by Schwann Cell Transplantation. Front. Neurosci. 2022, 16. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Funk, L.; Lee, J.; Bunge, M. Macrophage Depletion and Schwann Cell Transplantation Reduce Cyst Size after Rat Contusive Spinal Cord Injury. Neural Regen. Res. 2018, 13, 684. [Google Scholar] [CrossRef]
- Pearse, D.; Bastidas, J.; Izabel, S.; Ghosh, M. Schwann Cell Transplantation Subdues the Pro-Inflammatory Innate Immune Cell Response after Spinal Cord Injury. Int. J. Mol. Sci. 2018, 19, 2550. [Google Scholar] [CrossRef]
- Mousavi, M.; Hedayatpour, A.; Mortezaee, K.; Mohamadi, Y.; Abolhassani, F.; Hassanzadeh, G. Schwann Cell Transplantation Exerts Neuroprotective Roles in Rat Model of Spinal Cord Injury by Combating Inflammasome Activation and Improving Motor Recovery and Remyelination. Metab. Brain Dis. 2019, 34, 1117–1130. [Google Scholar] [CrossRef]
- Monje, P. V.; Deng, L.; Xu, X.-M. Human Schwann Cell Transplantation for Spinal Cord Injury: Prospects and Challenges in Translational Medicine. Front. Cell. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Niapour, A.; Karamali, F.; Nemati, S.; Taghipour, Z.; Mardani, M.; Nasr-Esfahani, M.H.; Baharvand, H. Cotransplantation of Human Embryonic Stem Cell-Derived Neural Progenitors and Schwann Cells in a Rat Spinal Cord Contusion Injury Model Elicits a Distinct Neurogenesis and Functional Recovery. Cell Transplant. 2012, 21, 827–843. [Google Scholar] [CrossRef]
- Barraud, P.; Seferiadis, A.A.; Tyson, L.D.; Zwart, M.F.; Szabo-Rogers, H.L.; Ruhrberg, C.; Liu, K.J.; Baker, C.V.H. Neural Crest Origin of Olfactory Ensheathing Glia. Proc. Natl. Acad. Sci. 2010, 107, 21040–21045. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; He, B.-R.; Hao, D.-J. Biological Roles of Olfactory Ensheathing Cells in Facilitating Neural Regeneration: A Systematic Review. Mol. Neurobiol. 2015, 51, 168–179. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, J.; Tang, X.; Wang, X.; Hao, D.; Yang, H. The Immunological Roles of Olfactory Ensheathing Cells in the Treatment of Spinal Cord Injury. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.W.; Roskams, A.J. Olfactory Ensheathing Cell Transplantation Following Spinal Cord Injury: Hype or Hope? Exp. Neurol. 2008, 209, 353–367. [Google Scholar] [CrossRef]
- Chuah, M.I.; Hale, D.M.; West, A.K. Interaction of Olfactory Ensheathing Cells with Other Cell Types in Vitro and after Transplantation: Glial Scars and Inflammation. Exp. Neurol. 2011, 229, 46–53. [Google Scholar] [CrossRef]
- Khankan, R.R.; Griffis, K.G.; Haggerty-Skeans, J.R.; Zhong, H.; Roy, R.R.; Reggie Edgerton, V.; Phelps, P.E. Olfactory Ensheathing Cell Transplantation after a Complete Spinal Cord Transection Mediates Neuroprotective and Immunomodulatory Mechanisms to Facilitate Regeneration. J. Neurosci. 2016, 36, 6269–6286. [Google Scholar] [CrossRef]
- Pourkhodadad, S.; Oryan, S.; Kaka, G.; Sadraie, S.H. Neuroprotective Effects of Combined Treatment with Minocycline and Olfactory Ensheathing Cells Transplantation against Inflammation and Oxidative Stress after Spinal Cord Injury. Cell J. 2018, 21. [Google Scholar] [CrossRef]
- He, B.-R.; Xie, S.-T.; Wu, M.-M.; Hao, D.-J.; Yang, H. Phagocytic Removal of Neuronal Debris by Olfactory Ensheathing Cells Enhances Neuronal Survival and Neurite Outgrowth via P38MAPK Activity. Mol. Neurobiol. 2014, 49, 1501–1512. [Google Scholar] [CrossRef]
- Fan, H.; Chen, Z.; Tang, H. Bin; Shan, L.Q.; Chen, Z.Y.; Wang, X.H.; Huang, D.G.; Liu, S.C.; Chen, X.; Yang, H.; et al. Exosomes Derived from Olfactory Ensheathing Cells Provided Neuroprotection for Spinal Cord Injury by Switching the Phenotype of Macrophages/Microglia. Bioeng. Transl. Med. 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- LóPEZ-VALES, Rub; GARCíA-ALíAS, G.; FORéS, J.; VELA, Jos.M.; NAVARRO, X.; VERDú, E. Transplanted Olfactory Ensheathing Cells Modulate the Inflammatory Response in the Injured Spinal Cord. Neuron Glia Biol. 2004, 1, 201–209. [CrossRef] [PubMed]
- Zhang, S.; Wang, Q.; Yang, Q.; Gu, H.; Yin, Y.; Li, Y.; Hou, J.; Chen, R.; Sun, Q.; Sun, Y.; et al. NG2 Glia Regulate Brain Innate Immunity via TGF-Β2/TGFBR2 Axis. BMC Med. 2019, 17, 204. [Google Scholar] [CrossRef]
- Hackett, A.R.; Lee, J.K. Understanding the NG2 Glial Scar after Spinal Cord Injury. Front. Neurol. 2016, 7. [Google Scholar] [CrossRef]
- Li, N.; Leung, G.K.K. Oligodendrocyte Precursor Cells in Spinal Cord Injury: A Review and Update. Biomed Res. Int. 2015, 2015, 1–20. [Google Scholar] [CrossRef]
- Levine, J. The Reactions and Role of NG2 Glia in Spinal Cord Injury. Brain Res. 2016, 1638, 199–208. [Google Scholar] [CrossRef]
- Knerlich-Lukoschus, F.; von der Ropp-Brenner, B.; Lucius, R.; Mehdorn, H.M.; Held-Feindt, J. Chemokine Expression in the White Matter Spinal Cord Precursor Niche after Force-Defined Spinal Cord Contusion Injuries in Adult Rats. Glia 2010, NA–NA. [Google Scholar] [CrossRef]
- Williams, J.L.; Patel, J.R.; Daniels, B.P.; Klein, R.S. Targeting CXCR7/ACKR3 as a Therapeutic Strategy to Promote Remyelination in the Adult Central Nervous System. J. Exp. Med. 2014, 211, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Wang, C.; Zepp, J.; Wu, L.; Sun, K.; Zhao, J.; Chandrasekharan, U.; DiCorleto, P.E.; Trapp, B.D.; Ransohoff, R.M.; et al. Act1 Mediates IL-17–Induced EAE Pathogenesis Selectively in NG2+ Glial Cells. Nat. Neurosci. 2013, 16, 1401–1408. [Google Scholar] [CrossRef]
- Rodriguez, J.P.; Coulter, M.; Miotke, J.; Meyer, R.L.; Takemaru, K.-I.; Levine, J.M. Abrogation of -Catenin Signaling in Oligodendrocyte Precursor Cells Reduces Glial Scarring and Promotes Axon Regeneration after CNS Injury. J. Neurosci. 2014, 34, 10285–10297. [Google Scholar] [CrossRef]
- Liu, Y.; Aguzzi, A. NG2 Glia Are Required for Maintaining Microglia Homeostatic State. Glia 2020, 68, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Suzuki, Y.; Ejiri, Y.; Noda, T.; Bai, H.; Kitada, M.; Kataoka, K.; Ohta, M.; Chou, H.; Ide, C. Bone Marrow Stromal Cells Enhance Differentiation of Cocultured Neurosphere Cells and Promote Regeneration of Injured Spinal Cord. J. Neurosci. Res. 2003, 72, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Neuhuber, B.; Timothy Himes, B.; Shumsky, J.S.; Gallo, G.; Fischer, I. Axon Growth and Recovery of Function Supported by Human Bone Marrow Stromal Cells in the Injured Spinal Cord Exhibit Donor Variations. Brain Res. 2005, 1035, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Ohta, M.; Suzuki, Y.; Noda, T.; Ejiri, Y.; Dezawa, M.; Kataoka, K.; Chou, H.; Ishikawa, N.; Matsumoto, N.; Iwashita, Y.; et al. Bone Marrow Stromal Cells Infused into the Cerebrospinal Fluid Promote Functional Recovery of the Injured Rat Spinal Cord with Reduced Cavity Formation. Exp. Neurol. 2004, 187, 266–278. [Google Scholar] [CrossRef]
- Lu, P.; Jones, L.L.; Tuszynski, M.H. BDNF-Expressing Marrow Stromal Cells Support Extensive Axonal Growth at Sites of Spinal Cord Injury. Exp. Neurol. 2005, 191, 344–360. [Google Scholar] [CrossRef]
- Hofstetter, C.P.; Schwarz, E.J.; Hess, D.; Widenfalk, J.; El Manira, a; Prockop, D. J.; Olson, L. Marrow Stromal Cells Form Guiding Strands in the Injured Spinal Cord and Promote Recovery. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 2199–2204. [Google Scholar] [CrossRef]
- Gu, W.; Zhang, F.; Xue, Q.; Ma, Z.; Lu, P.; Yu, B. Transplantation of Bone Marrow Mesenchymal Stem Cells Reduces Lesion Volume and Induces Axonal Regrowth of Injured Spinal Cord. Neuropathol. Off. J. Japanese Soc. Neuropathol. 2010, 30, 205–217. [Google Scholar] [CrossRef]
- Li, Y.; Chopp, M. Marrow Stromal Cell Transplantation in Stroke and Traumatic Brain Injury. Neurosci. Lett. 2009, 456, 120–123. [Google Scholar] [CrossRef]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult Rat and Human Bone Marrow Stromal Cells Differentiate into Neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, S.L.; Barnett, S.C. Therapeutic Potential of Niche-specific Mesenchymal Stromal Cells for Spinal Cord Injury Repair. Cells 2021, 10. [Google Scholar] [CrossRef]
- Nakajima, H.; Uchida, K.; Guerrero, A.R.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.T.; Johnson, W.E.B.; et al. Transplantation of Mesenchymal Stem Cells Promotes an Alternative Pathway of Macrophage Activation and Functional Recovery after Spinal Cord Injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Urdzíková, L.; Růžička, J.; LaBagnara, M.; Kárová, K.; Kubinová, Š.; Jiráková, K.; Murali, R.; Syková, E.; Jhanwar-Uniyal, M.; Jendelová, P. Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat. Int. J. Mol. Sci. 2014, 15, 11275–11293. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, C. Mesenchymal Stem Cells Immunosuppressive Properties: Is It Specific to Bone Marrow-Derived Cells? Stem Cell Res. Ther. 2010, 1, 15. [Google Scholar] [CrossRef] [PubMed]
- Shao, A.; Tu, S.; Lu, J.; Zhang, J. Crosstalk between Stem Cell and Spinal Cord Injury: Pathophysiology and Treatment Strategies. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhu, W.; Cao, K.; Wu, F.; Li, J.; Wang, G.; Li, H.; Lu, M.; Ren, Y.; He, X. Anti-Inflammatory Mechanism of Neural Stem Cell Transplantation in Spinal Cord Injury. Int. J. Mol. Sci. 2016, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Liu, W.; Wang, J.; Fan, J.; Luo, Y.; Li, L.; Kong, F.; Chen, J.; Tang, P.; Cai, W. Neural Stem Cell-Derived Small Extracellular Vesicles Attenuate Apoptosis and Neuroinflammation after Traumatic Spinal Cord Injury by Activating Autophagy. Cell Death Dis. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Buzoianu-Anguiano, V.; Torres-Llacsa, M.; Doncel-Pérez, E. Role of Aldynoglia Cells in Neuroinflammatory and Neuroimmune Responses after Spinal Cord Injury. Cells 2021, 10, 2783. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Deng, M.; Zhao, X.Q.; Liu, M. Alternatively Polarized Macrophages Regulate the Growth and Differentiation of Ependymal Stem Cells through the SIRT2 Pathway. Exp. Neurobiol. 2020, 29, 150–163. [Google Scholar] [CrossRef]
- Kaspar, A.A.; Reichert, J.M. Future Directions for Peptide Therapeutics Development. Drug Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E.; Powrie, F.; Puri, R.N.; Meister, A. Glutathione Monoethyl Ester: Preparation, Uptake by Tissues, and Conversion to Glutathione. Arch. Biochem. Biophys. 1985, 239, 538–548. [Google Scholar] [CrossRef]
- Arosio, E.; De Marchi, S.; Zannoni, M.; Prior, M.; Lechi, A. Effect of Glutathione Infusion on Leg Arterial Circulation, Cutaneous Microcirculation, and Pain-Free Walking Distance in Patients With Peripheral Obstructive Arterial Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Mayo Clin. Proc. 2002, 77, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.H.; Wheeler, D.G.; Guan, Z.; Suntres, Z.; Stokes, B.T. Effect of Glutathione Augmentation on Lipid Peroxidation after Spinal Cord Injury. J. Neurotrauma 2002, 19, 763–775. [Google Scholar] [CrossRef]
- Wernerman, J.; Hammarqvist, F. Modulation of Endogenous Glutathione Availability. Curr. Opin. Clin. Nutr. Metab. Care 1999, 2, 487–492. [Google Scholar] [CrossRef]
- Santoscoy, C.; Rı́os, C.; Franco-Bourland, R.E.; Hong, E.; Bravo, G.; Rojas, G.; Guízar-Sahagún, G. Lipid Peroxidation by Nitric Oxide Supplements after Spinal Cord Injury: Effect of Antioxidants in Rats. Neurosci. Lett. 2002, 330, 94–98. [Google Scholar] [CrossRef]
- Bracken, M.B. Steroids for Acute Spinal Cord Injury. Cochrane Database Syst. Rev. 2012, 2018. [Google Scholar] [CrossRef]
- Sultan, I.; Lamba, N.; Liew, A.; Doung, P.; Tewarie, I.; Amamoo, J.J.; Gannu, L.; Chawla, S.; Doucette, J.; Cerecedo-Lopez, C.D.; et al. The Safety and Efficacy of Steroid Treatment for Acute Spinal Cord Injury: A Systematic Review and Meta-Analysis. Heliyon 2020, 6, e03414. [Google Scholar] [CrossRef]
- Fischer, H.J.; Finck, T.L.K.; Pellkofer, H.L.; Reichardt, H.M.; Lühder, F. Glucocorticoid Therapy of Multiple Sclerosis Patients Induces Anti-Inflammatory Polarization and Increased Chemotaxis of Monocytes. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Hanada, M.; Sugiura, Y.; Shinjo, R.; Masaki, N.; Imagama, S.; Ishiguro, N.; Matsuyama, Y.; Setou, M. Spatiotemporal Alteration of Phospholipids and Prostaglandins in a Rat Model of Spinal Cord Injury. Anal. Bioanal. Chem. 2012, 403, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Resnick, D.K.; Nguyen, P.; Cechvala, C.F. Regional and Temporal Changes in Prostaglandin E 2 and Thromboxane B 2 Concentrations after Spinal Cord Injury. Spine J. 2001, 1, 432–436. [Google Scholar] [CrossRef] [PubMed]
- GuÍzar-Sahagún, G.; Ibarra, A.; Espitia, A.; MartÍnez, A.; Madrazo, I.; Franco-Bourland, R.E. Glutathione Monoethyl Ester Improves Functional Recovery, Enhances Neuron Survival, and Stabilizes Spinal Cord Blood Flow after Spinal Cord Injury in Rats. Neuroscience 2005, 130, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Silva-García, R.; Estrada-García, I.; Ramos-Payán, R.; Torres-Salazar, A.; Morales-Martínez, M.E.; Arenas-Aranda, D.; Giménez-Scherer, J.A.; Blanco-Favela, F.; Rico-Rosillo, M.G. The Effect of an Anti-Inflammatory Pentapeptide Produced by Entamoeba Histolytica on Gene Expression in the U-937 Monocytic Cell Line. Inflamm. Res. 2008, 57, 145–150. [Google Scholar] [CrossRef]
- Silva-García, R.; Rico-Rosillo, G. Anti-Inflammatory Defense Mechanisms of Entamoeba Histolytica. Inflamm. Res. 2011, 60, 111–117. [Google Scholar] [CrossRef]
- Galán-Salinas, A.; Corral-Ruíz, G.; Pérez-Vega, M.J.; Fabila-Castillo, L.; Silva-García, R.; Marquina-Castillo, B.; León-Contreras, J.C.; Barrios-Payán, J.; Francisco-Cruz, A.; Montecillo-Aguado, M.; et al. Monocyte Locomotion Inhibitory Factor Confers Neuroprotection and Prevents the Development of Murine Cerebral Malaria. Int. Immunopharmacol. 2021, 97. [Google Scholar] [CrossRef]
- Bermeo, G.; Ibarra, A.; García, E.; Flores-Romero, A.; Rico-Rosillo, G.; Marroquín, R.; Mestre, H.; Flores, C.; Blanco-Favela, F.; Silva-García, R. Monocyte Locomotion Inhibitory Factor Produced by E. Histolytica Improves Motor Recovery and Develops Neuroprotection after Traumatic Injury to the Spinal Cord. Biomed Res. Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Aggarwal, S.; Yurlova, L.; Simons, M. Central Nervous System Myelin: Structure, Synthesis and Assembly. Trends Cell Biol. 2011, 21, 585–593. [Google Scholar] [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the Central Nervous System: Structure, Function, and Pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef]
- Stassart, R.M.; Möbius, W.; Nave, K.-A.; Edgar, J.M. The Axon-Myelin Unit in Development and Degenerative Disease. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Monroy, G.R.; Murguiondo Pérez, R.; Weintraub Ben Zión, E.; Vidal Alcántar-Garibay, O.; Loza-López, E.C.; Tejerina Marion, E.; Blancarte Hernández, E.; Navarro-Torres, L.; Ibarra, A. Immunization with Neural-Derived Peptides in Neurodegenerative Diseases: A Narrative Review. Biomedicines 2023, 11, 919. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.; Boehme, S.A.; Chalmers, D.; Crowe, P.D.; Pahuja, A.; Ling, N.; Brocke, S.; Steinman, L.; Conlon, P.J. Amelioration of Relapsing Experimental Autoimmune Encephalomyelitis with Altered Myelin Basic Protein Peptides Involves Different Cellular Mechanisms. J. Neuroimmunol. 1997, 74, 149–158. [Google Scholar] [CrossRef]
- Liu, H.; Shiryaev, S.A.; Chernov, A. V.; Kim, Y.; Shubayev, I.; Remacle, A.G.; Baranovskaya, S.; Golubkov, V.S.; Strongin, A.Y.; Shubayev, V.I. Immunodominant Fragments of Myelin Basic Protein Initiate T Cell-Dependent Pain. J. Neuroinflammation 2012, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.; Kipnis, J. Protective Autoimmunity: Regulation and Prospects for Vaccination after Brain and Spinal Cord Injuries. Trends Mol. Med. 2001, 7, 252–258. [Google Scholar] [CrossRef]
- Schwartz, M.; London, A.; Shechter, R. Boosting T-Cell Immunity as a Therapeutic Approach for Neurodegenerative Conditions: The Role of Innate Immunity. Neuroscience 2009, 158, 1133–1142. [Google Scholar] [CrossRef]
- Hauben, E.; Butovsky, O.; Nevo, U.; Yoles, E.; Moalem, G.; Agranov, E.; Mor, F.; Leibowitz-Amit, R.; Pevsner, E.; Akselrod, S.; et al. Passive or Active Immunization with Myelin Basic Protein Promotes Recovery from Spinal Cord Contusion. J. Neurosci. 2000, 20, 6421–6430. [Google Scholar] [CrossRef]
- Ibarra, A.; Sosa, M.; García, E.; Flores, A.; Cruz, Y.; Mestre, H.; Martiñón, S.; Pineda-Rodríguez, B.A.; Gutiérrez-Ospina, G. Prophylactic Neuroprotection with A91 Improves the Outcome of Spinal Cord Injured Rats. Neurosci. Lett. 2013, 554, 59–63. [Google Scholar] [CrossRef]
- Johnson, K.P.; Brooks, B.R.; Cohen, J.A.; Ford, C.C.; Goldstein, J.; Lisak, R.P.; Myers, L.W.; Panitch, H.S.; Rose, J.W.; Schiffer, R.B.; et al. Copolymer 1 Reduces Relapse Rate and Improves Disability in Relapsing-Remitting Multiple Sclerosis: Results of a Phase III Multicenter, Double-Blind, Placebo-Controlled Trial. Neurology 1995, 45, 1268–1276. [Google Scholar] [CrossRef]
- Prod’homme, T.; Zamvil, S.S. The Evolving Mechanisms of Action of Glatiramer Acetate. Cold Spring Harb. Perspect. Med. 2019, 9, a029249. [Google Scholar] [CrossRef]
- De Kleijn, K.M.A.; Martens, G.J.M. Molecular Effects of FDA-Approved Multiple Sclerosis Drugs on Glial Cells and Neurons of the Central Nervous System. Int. J. Mol. Sci. 2020, 21, 4229. [Google Scholar] [CrossRef] [PubMed]
- Angelov, D.N.; Waibel, S.; Guntinas-Lichius, O.; Lenzen, M.; Neiss, W.F.; Tomov, T.L.; Yoles, E.; Kipnis, J.; Schori, H.; Reuter, A.; et al. Therapeutic Vaccine for Acute and Chronic Motor Neuron Diseases: Implications for Amyotrophic Lateral Sclerosis. Proc. Natl. Acad. Sci. 2003, 100, 4790–4795. [Google Scholar] [CrossRef]
- Benner, E.J.; Mosley, R.L.; Destache, C.J.; Lewis, T.B.; Jackson-Lewis, V.; Gorantla, S.; Nemachek, C.; Green, S.R.; Przedborski, S.; Gendelman, H.E. Therapeutic Immunization Protects Dopaminergic Neurons in a Mouse Model of Parkinson’s Disease. Proc. Natl. Acad. Sci. 2004, 101, 9435–9440. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Koronyo-Hamaoui, M.; Kunis, G.; Ophir, E.; Landa, G.; Cohen, H.; Schwartz, M. Glatiramer Acetate Fights against Alzheimer’s Disease by Inducing Dendritic-like Microglia Expressing Insulin-like Growth Factor 1. Proc. Natl. Acad. Sci. 2006, 103, 11784–11789. [Google Scholar] [CrossRef] [PubMed]
- Incontri-Abraham, D.; Esparza-Salazar, F.J.; Ibarra, A. Copolymer-1 as a Potential Therapy for Mild Cognitive Impairment. Brain Cogn. 2022, 162, 105892. [Google Scholar] [CrossRef]
- Arnon, R.; Aharoni, R. Neurogenesis and Neuroprotection in the CNS — Fundamental Elements in the Effect of Glatiramer Acetate on Treatment of Autoimmune Neurological Disorders. Mol. Neurobiol. 2007, 36, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Dhib-Jalbut, S. Mechanisms of Action of Interferons and Glatiramer Acetate in Multiple Sclerosis. Neurology 2002, 58, S3–S9. [Google Scholar] [CrossRef]
- Liblau, R. Glatiramer Acetate for the Treatment of Multiple Sclerosis: Evidence for a Dual Anti-Inflammatory and Neuroprotective Role. J. Neurol. Sci. 2009, 287, S17–S23. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, M.L.; Trinchero, M.F.; Zorrilla-Zubilete, M.A.; Schinder, A.F.; Genaro, A.M. Glatiramer Acetate Reverts Stress-Induced Alterations on Adult Neurogenesis and Behavior. Involvement of Th1/Th2 Balance. Brain. Behav. Immun. 2012, 26, 429–438. [Google Scholar] [CrossRef]
- Scorisa, J.M.; Zanon, R.G.; Freria, C.M.; de Oliveira, A.L.R. Glatiramer Acetate Positively Influences Spinal Motoneuron Survival and Synaptic Plasticity after Ventral Root Avulsion. Neurosci. Lett. 2009, 451, 34–39. [Google Scholar] [CrossRef]
- Martiñón, S.; García, E.; Gutierrez-Ospina, G.; Mestre, H.; Ibarra, A. Development of Protective Autoimmunity by Immunization with a Neural-Derived Peptide Is Ineffective in Severe Spinal Cord Injury. PLoS One 2012, 7, e32027. [Google Scholar] [CrossRef]
- Askarifirouzjaei, H.; Khajoueinejad, L.; Salek Farrokhi, A.; Tahoori, M.-T.; Fazeli, M.; Tiraihi, T.; Pourfathollah, A.A. Implications of Immunotherapy with High-Dose Glatiramer Acetate in Acute Phase of Spinal Cord Injury in Rats. Immunopharmacol. Immunotoxicol. 2019, 41, 150–162. [Google Scholar] [CrossRef]
- Ibarra, A.; Mendieta-Arbesú, E.; Suarez-Meade, P.; García-Vences, E.; Martiñón, S.; Rodriguez-Barrera, R.; Lomelí, J.; Flores-Romero, A.; Silva-García, R.; Buzoianu-Anguiano, V.; et al. Motor Recovery after Chronic Spinal Cord Transection in Rats: A Proof-of-Concept Study Evaluating a Combined Strategy. CNS Neurol. Disord. - Drug Targets 2018, 18, 52–62. [Google Scholar] [CrossRef]
- Walsh, J.T.; Hendrix, S.; Boato, F.; Smirnov, I.; Zheng, J.; Lukens, J.R.; Gadani, S.; Hechler, D.; Gölz, G.; Rosenberger, K.; et al. MHCII-Independent CD4+ T Cells Protect Injured CNS Neurons via IL-4. J. Clin. Invest. 2015, 125, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. IL-10 Promotes Neuronal Survival Following Spinal Cord Injury. Exp. Neurol. 2009, 220, 183–190. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. TNF-Alpha as a Therapeutic Target in Inflammatory Diseases, Ischemia- Reperfusion Injury and Trauma. Curr. Med. Chem. 2009, 16, 3152–3167. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. Anti-TNF Therapy in the Injured Spinal Cord. Trends Pharmacol. Sci. 2011, 32, 107–115. [Google Scholar] [CrossRef]
- Fan, H.; Tang, H. Bin; Chen, Z.; Wang, H.Q.; Zhang, L.; Jiang, Y.; Li, T.; Yang, C.F.; Wang, X.Y.; Li, X.; et al. Inhibiting HMGB1-RAGE Axis Prevents pro-Inflammatory Macrophages/Microglia Polarization and Affords Neuroprotection after Spinal Cord Injury. J. Neuroinflammation 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; García, E.; Flores, N.; Martiñón, S.; Reyes, R.; Campos, M.G.; Maciel, M.; Mestre, H. Immunization with Neural-Derived Antigens Inhibits Lipid Peroxidation after Spinal Cord Injury. Neurosci. Lett. 2010, 476, 62–65. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Kong, P.; Zhao, S.; Yang, H.; Chen, C.; Yan, J. Enhanced Expression of Neurotrophic Factors in the Injured Spinal Cord Through Vaccination With Myelin Basic Protein-Derived Peptide Pulsed Dendritic Cells. Spine (Phila. Pa. 1976). 2015, 40, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Barrera, R.; Fernández-Presas, A.M.; García, E.; Flores-Romero, A.; Martiñón, S.; González-Puertos, V.Y.; Mestre, H.; Flores-Dominguez, C.; Rodriguez-Mata, V.; Königsberg, M.; et al. Immunization with a Neural-Derived Peptide Protects the Spinal Cord from Apoptosis after Traumatic Injury. Biomed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Martiñón, S.; García-Vences, E.; Toscano-Tejeida, D.; Flores-Romero, A.; Rodriguez-Barrera, R.; Ferrusquia, M.; Hernández-Muñoz, R.E.; Ibarra, A. Long-Term Production of BDNF and NT-3 Induced by A91-Immunization after Spinal Cord Injury. BMC Neurosci. 2016, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-D.; Wu, C.-L.; Hwang, W.-C.; Yang, D.-I. More Insight into BDNF against Neurodegeneration: Anti-Apoptosis, Anti-Oxidation, and Suppression of Autophagy. Int. J. Mol. Sci. 2017, 18, 545. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Barrera, R.; Flores-Romero, A.; Fernández-Presas, A.M.; García-Vences, E.; Silva-García, R.; Konigsberg, M.; Blancas-Espinoza, L.; Buzoianu-Anguiano, V.; Soria-Zavala, K.; Suárez-Meade, P.; et al. Immunization with Neural Derived Peptides plus Scar Removal Induces a Permissive Microenvironment, and Improves Locomotor Recovery after Chronic Spinal Cord Injury. BMC Neurosci. 2017, 18. [Google Scholar] [CrossRef]
- Martiñon, S.; García, E.; Flores, N.; Gonzalez, I.; Ortega, T.; Buenrostro, M.; Reyes, R.; Fernandez-Presas, A.M.; Guizar-Sahagún, G.; Correa, D.; et al. Vaccination with a Neural-Derived Peptide plus Administration of Glutathione Improves the Performance of Paraplegic Rats. Eur. J. Neurosci. 2007, 26, 403–412. [Google Scholar] [CrossRef]
- del Rayo Garrido, M.; Silva-García, R.; García, E.; Martiñón, S.; Morales, M.; Mestre, H.; Flores-Domínguez, C.; Flores, A.; Ibarra, A. Therapeutic Window for Combination Therapy of A91 Peptide and Glutathione Allows Delayed Treatment After Spinal Cord Injury. Basic Clin. Pharmacol. Toxicol. 2013, 112, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Parra-Villamar, D.; Blancas-Espinoza, L.; Garcia-Vences, E.; Herrera-García, J.; Flores-Romero, A.; Toscano-Zapien, A.; Villa, J.; Barrera-Roxana, R.; Karla, S.; Ibarra, A.; et al. Neuroprotective Effect of Immunomodulatory Peptides in Rats with Traumatic Spinal Cord Injury. Neural Regen. Res. 2021, 16, 1273. [Google Scholar] [CrossRef]
- HENDRIX, S.; NITSCH, R. The Role of T Helper Cells in Neuroprotection and Regeneration. J. Neuroimmunol. 2007, 184, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.; Delamarre, L. Dendritic Cell-Targeted Vaccines. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Trumpfheller, C.; Longhi, M.P.; Caskey, M.; Idoyaga, J.; Bozzacco, L.; Keler, T.; Schlesinger, S.J.; Steinman, R.M. Dendritic Cell-Targeted Protein Vaccines: A Novel Approach to Induce T-Cell Immunity. J. Intern. Med. 2012, 271, 183–192. [Google Scholar] [CrossRef]
- Wang, K.; Chao, R.; Guo, Q.-N.; Liu, M.-Y.; Liang, H.-P.; Liu, P.; Zhao, J.-H. Expressions of Some Neurotrophins and Neurotrophic Cytokines at Site of Spinal Cord Injury in Mice after Vaccination with Dendritic Cells Pulsed with Homogenate Proteins. Neuroimmunomodulation 2013, 20, 87–98. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, J.; Liang, H.; Bian, X. Vaccination with Dendritic Cells Pulsed with Homogenate Protein of Spinal Cord Promotes Functional Recovery from Spinal Cord Injury in Mice. Spinal Cord 2009, 47, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.H.; Rosenzweig, E.S.; Blesch, A.; Moseanko, R.; Havton, L.A.; Edgerton, V.R.; Tuszynski, M.H. Local and Remote Growth Factor Effects after Primate Spinal Cord Injury. J. Neurosci. 2010, 30, 9728–9737. [Google Scholar] [CrossRef] [PubMed]
- Franz, S.; Ciatipis, M.; Pfeifer, K.; Kierdorf, B.; Sandner, B.; Bogdahn, U.; Blesch, A.; Winner, B.; Weidner, N. Thoracic Rat Spinal Cord Contusion Injury Induces Remote Spinal Gliogenesis but Not Neurogenesis or Gliogenesis in the Brain. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Moalem, G.; Gdalyahu, A.; Shani, Y.; Otten, U.; Lazarovici, P.; Cohen, I.R.; Schwartz, M. Production of Neurotrophins by Activated T Cells: Implications for Neuroprotective Autoimmunity. J. Autoimmun. 2000, 15, 331–345. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Gallmeier, E.; Behrens, L.; Leal, V.V.; Misgeld, T.; Klinkert, W.E.F.; Kolbeck, R.; Hoppe, E.; Oropeza-Wekerle, R.-L.; Bartke, I.; et al. Activated Human T Cells, B Cells, and Monocytes Produce Brain-Derived Neurotrophic Factor In Vitro and in Inflammatory Brain Lesions: A Neuroprotective Role of Inflammation? J. Exp. Med. 1999, 189, 865–870. [Google Scholar] [CrossRef]
- Silver, J.; Miller, J.H. Regeneration beyond the Glial Scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Coulson-Thomas, V.J.; Lauer, M.E.; Soleman, S.; Zhao, C.; Hascall, V.C.; Day, A.J.; Fawcett, J.W. Tumor Necrosis Factor-Stimulated Gene-6 (TSG-6) Is Constitutively Expressed in Adult Central Nervous System (CNS) and Associated with Astrocyte-Mediated Glial Scar Formation Following Spinal Cord Injury. J. Biol. Chem. 2016, 291, 19939–19952. [Google Scholar] [CrossRef]
- Niclou, S.P.; Ehlert, E.M.E.; Verhaagen, J. Chemorepellent Axon Guidance Molecules in Spinal Cord Injury. J. Neurotrauma 2006, 23, 409–421. [Google Scholar] [CrossRef]
- Hu, R.; Zhou, J.; Luo, C.; Lin, J.; Wang, X.; Li, X.; Bian, X.; Li, Y.; Wan, Q.; Yu, Y.; et al. Glial Scar and Neuroregeneration: Histological, Functional, and Magnetic Resonance Imaging Analysis in Chronic Spinal Cord Injury. J. Neurosurg. Spine 2010, 13, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the Glial Scar for Spinal Cord Repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Barrera, R.; Flores-Romero, A.; Buzoianu-Anguiano, V.; Garcia, E.; Soria-Zavala, K.; Incontri-Abraham, D.; Garibay-López, M.; Juárez-Vignon Whaley, J.J.; Ibarra, A. Use of a Combination Strategy to Improve Morphological and Functional Recovery in Rats With Chronic Spinal Cord Injury. Front. Neurol. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- García, E.; Rodríguez-Barrera, R.; Buzoianu-Anguiano, V.; Flores-Romero, A.; Malagón-Axotla, E.; Guerrero-Godinez, M.; De La Cruz-Castillo, E.; Castillo-Carvajal, L.; Rivas-Gonzalez, M.; Santiago-Tovar, P.; et al. Use of a Combination Strategy to Improve Neuroprotection and Neuroregeneration in a Rat Model of Acute Spinal Cord Injury. Neural Regen. Res. 2019, 14, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Ciulla, M.; Marinelli, L.; Cacciatore, I.; Stefano, A. Di Role of Dietary Supplements in the Management of Parkinson’s Disease. Biomolecules 2019, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Opperman, E.A.; Buchholz, A.C.; Darlington, G.A.; Martin Ginis, K.A. Dietary Supplement Use in the Spinal Cord Injury Population. Spinal Cord 2010, 48, 60–64. [Google Scholar] [CrossRef]
- Hong, J.Y.; Davaa, G.; Yoo, H.; Hong, K.; Hyun, J.K. Ascorbic Acid Promotes Functional Restoration after Spinal Cord Injury Partly by Epigenetic Modulation. Cells 2020, 9, 1310. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Michels, A.J.; Frei, B. Vitamin C. Adv. Nutr. 2014, 5, 16–18. [Google Scholar] [CrossRef]
- Holton, K.F. Micronutrients May Be a Unique Weapon Against the Neurotoxic Triad of Excitotoxicity, Oxidative Stress and Neuroinflammation: A Perspective. Front. Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- Chen, C.; Yang, Q.; Ma, X. Synergistic Effect of Ascorbic Acid and Taurine in the Treatment of a Spinal Cord Injury-Induced Model in Rats. 3 Biotech 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Barbas, C. Vitamin E: Action, Metabolism and Perspectives. J. Physiol. Biochem. 2001, 57, 43–56. [Google Scholar] [CrossRef]
- Cordero, K.; Coronel, G.; Serrano-Illán, M.; Cruz-Bracero, J.; Figueroa, J.; De León, M. Effects of Dietary Vitamin E Supplementation in Bladder Function and Spasticity during Spinal Cord Injury. Brain Sci. 2018, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Sarveazad, A.; Babahajian, A.; Baikpour, M.; Vaccaro, A.R.; Chapman, J.R.; Yousefifard, M.; Rahimi-Movaghar, V. Effect of Vitamins C and E on Recovery of Motor Function after Spinal Cord Injury: Systematic Review and Meta-Analysis of Animal Studies. Nutr. Rev. 2020, 78, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Hernández-Ayvar, F.; Rodríguez-Barrera, R.; Flores-Romero, A.; Borlongan, C.; Ibarra, A. Supplementation With Vitamin E, Zinc, Selenium, and Copper Re-Establishes T-Cell Function and Improves Motor Recovery in a Rat Model of Spinal Cord Injury. Cell Transplant. 2022, 31, 096368972211098. [Google Scholar] [CrossRef] [PubMed]
- Heller, R.A.; Seelig, J.; Bock, T.; Haubruck, P.; Grützner, P.A.; Schomburg, L.; Moghaddam, A.; Biglari, B. Relation of Selenium Status to Neuro-Regeneration after Traumatic Spinal Cord Injury. J. Trace Elem. Med. Biol. 2019, 51, 141–149. [Google Scholar] [CrossRef]
- Wen, S.; Li, Y.; Shen, X.; Wang, Z.; Zhang, K.; Zhang, J.; Mei, X. Protective Effects of Zinc on Spinal Cord Injury. J. Mol. Neurosci. 2021, 71, 2433–2440. [Google Scholar] [CrossRef]
- Li, D.; Tian, H.; Li, X.; Mao, L.; Zhao, X.; Lin, J.; Lin, S.; Xu, C.; Liu, Y.; Guo, Y.; et al. Zinc Promotes Functional Recovery after Spinal Cord Injury by Activating Nrf2/HO-1 Defense Pathway and Inhibiting Inflammation of NLRP3 in Nerve Cells. Life Sci. 2020, 245, 117351. [Google Scholar] [CrossRef] [PubMed]
- Neve, J.; Vertongen, F.; Molle, L. 6 Selenium Deficiency. Clin. Endocrinol. Metab. 1985, 14, 629–656. [Google Scholar] [CrossRef] [PubMed]
- Javdani, M.; Habibi, A.; Shirian, S.; Kojouri, G.A.; Hosseini, F. Effect of Selenium Nanoparticle Supplementation on Tissue Inflammation, Blood Cell Count, and IGF-1 Levels in Spinal Cord Injury-Induced Rats. Biol. Trace Elem. Res. 2019, 187, 202–211. [Google Scholar] [CrossRef]
- Santesmasses, D.; Gladyshev, V.N. Selenocysteine Machinery Primarily Supports TXNRD1 and GPX4 Functions and Together They Are Functionally Linked with SCD and PRDX6. Biomolecules 2022, 12, 1049. [Google Scholar] [CrossRef] [PubMed]
- Barceloux, D.G.; Barceloux, D. Copper. J. Toxicol. Clin. Toxicol. 1999, 37, 217–230. [Google Scholar] [CrossRef]
- Seelig, J.; Heller, R.A.; Hackler, J.; Haubruck, P.; Moghaddam, A.; Biglari, B.; Schomburg, L. Selenium and Copper Status - Potential Signposts for Neurological Remission after Traumatic Spinal Cord Injury. J. Trace Elem. Med. Biol. 2020, 57, 126415. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Bi, J.; Chen, C.; Sun, P.; Tan, H.; Feng, F.; Shen, J. Neuroprotective Effect of Omega-3 Fatty Acids on Spinal Cord Injury Induced Rats. Brain Behav. 2019, 9. [Google Scholar] [CrossRef]
- Marinelli, S.; Vacca, V.; De Angelis, F.; Pieroni, L.; Orsini, T.; Parisi, C.; Soligo, M.; Protto, V.; Manni, L.; Guerrieri, R.; et al. Publisher Correction: Innovative Mouse Model Mimicking Human-like Features of Spinal Cord Injury: Efficacy of Docosahexaenoic Acid on Acute and Chronic Phases. Sci. Rep. 2019, 9, 17043. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Delattre, A.M.; Carabelli, B.; Pudell, C.; Bortolanza, M.; Staziaki, P.V.; Visentainer, J.V.; Montanher, P.F.; Del Bel, E.A.; Ferraz, A.C. Neuroprotective Effect of Omega-3 Polyunsaturated Fatty Acids in the 6-OHDA Model of Parkinson’s Disease Is Mediated by a Reduction of Inducible Nitric Oxide Synthase. Nutr. Neurosci. 2018, 21, 341–351. [Google Scholar] [CrossRef]
- Ibarra, A.; Martinon, S. Pharmacological Neuroprotective Therapy for Acute Spinal Cord Injury: State of the Art. Mini-Reviews Med. Chem. 2008, 8, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Senturk, S.; Yaman, M.E.; Aydin, H.E.; Guney, G.; Bozkurt, I.; Paksoy, K.; Abdioglu, A.A. Effects of Resveratrol on Inflammation and Apoptosis after Experimental Spinal Cord Injury. Turk. Neurosurg. 2017, 28. [Google Scholar] [CrossRef]
- Zhao, H.; Mei, X.; Yang, D.; Tu, G. Resveratrol Inhibits Inflammation after Spinal Cord Injury via SIRT-1/NF-ΚB Signaling Pathway. Neurosci. Lett. 2021, 762, 136151. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; El, S.N.; Keijer, J.; van Schothorst, E.; Rühl, R.; Borel, P. β-Carotene in the Human Body: Metabolic Bioactivation Pathways – from Digestion to Tissue Distribution and Excretion. Proc. Nutr. Soc. 2019, 78, 68–87. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ouyang, L.; Lin, S.; Chen, S.; Liu, Y.; Zhou, W.; Wang, X. Protective Role of β-Carotene against Oxidative Stress and Neuroinflammation in a Rat Model of Spinal Cord Injury. Int. Immunopharmacol. 2018, 61, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Seyidoğlu, N.; Köşeli, E.; Gurbanlı, R.; Aydın, C. The Preventive Role of Spirulina Platensis ( Arthrospira Platensis ) in Immune and Oxidative Insults in a Stress-Induced Rat Model. J. Vet. Res. 2021, 65, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.J.; Farruggia, M.; Veronese, N.; Barbagallo, M. Vitamin D Sources, Metabolism, and Deficiency: Available Compounds and Guidelines for Its Treatment. Metabolites 2021, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, J.; Mailhot, G. Vitamin D and Spinal Cord Injury: Should We Care? Spinal Cord 2016, 54, 1060–1075. [Google Scholar] [CrossRef]
- Sabour⁎, H.; Norouzi, A.; Shidfar, F.; Vafa, M.R. The Effects of N-3 Fatty Acids on Inflammatory Cytokines in Osteoporotic Spinal Cord Injured Patients: A Randomized Clinical Trial. Bone 2012, 50, S156–S157. [Google Scholar] [CrossRef]
- Glisic, M.; Flueck, J.L.; Ruettimann, B.; Hertig-Godeschalk, A.; Valido, E.; Bertolo, A.; Stucki, G.; Stoyanov, J. The Feasibility of a Crossover, Randomized Controlled Trial Design to Assess the Effect of Probiotics and Prebiotics on Health of Elite Swiss Para-Athletes: A Study Protocol. Pilot Feasibility Stud. 2022, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.T. Probiotics. Am. J. Heal. Pharm. 2010, 67, 449–458. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Hall, J.C.E.; Wang, L.; Mo, X.; Yu, Z.; Popovich, P.G. Gut Dysbiosis Impairs Recovery after Spinal Cord Injury. J. Exp. Med. 2016, 213, 2603–2620. [Google Scholar] [CrossRef]
- Jing, Y.; Bai, F.; Yu, Y. Spinal Cord Injury and Gut Microbiota: A Review. Life Sci. 2021, 266, 118865. [Google Scholar] [CrossRef]
- Panther, E.J.; Dodd, W.; Clark, A.; Lucke-Wold, B. Gastrointestinal Microbiome and Neurologic Injury. Biomedicines 2022, 10, 500. [Google Scholar] [CrossRef]
- Wong, S.; Jamous, A.; O’Driscoll, J.; Sekhar, R.; Weldon, M.; Yau, C.Y.; Hirani, S.P.; Grimble, G.; Forbes, A. A Lactobacillus Casei Shirota Probiotic Drink Reduces Antibiotic-Associated Diarrhoea in Patients with Spinal Cord Injuries: A Randomised Controlled Trial. Br. J. Nutr. 2014, 111, 672–678. [Google Scholar] [CrossRef]
- Bannerman, C.A.; Douchant, K.; Sheth, P.M.; Ghasemlou, N. The Gut-Brain Axis and beyond: Microbiome Control of Spinal Cord Injury Pain in Humans and Rodents. Neurobiol. Pain 2021, 9, 100059. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cao, S.; Zhang, X. Modulation of Gut Microbiota-Brain Axis by Probiotics, Prebiotics, and Diet. J. Agric. Food Chem. 2015, 63, 7885–7895. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhang, B.; Shi, Q.; Liang, J.; Wang, X.; Lian, X.; Xu, J. The Conditioned Medium of Lactobacillus Rhamnoides GG Regulates Microglia/Macrophage Polarization and Improves Functional Recovery after Spinal Cord Injury in Rats. Biomed Res. Int. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Gungor, B.; Adiguzel, E.; Gursel, I.; Yilmaz, B.; Gursel, M. Intestinal Microbiota in Patients with Spinal Cord Injury. PLoS One 2016, 11, e0145878. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Self-destructive mechanisms and therapeutic targets in different phases of SCI. In the acute and subacute phases, resident and peripheral cells determine neural destruction through pro-inflammatory cytokines (IL-1β, IL-6, TNF-α, IFN-γ, IL-12), chemokines (MCP-1, CXCL10, CCL2, CXCL2), free radicals (ROS and NO produced by iNOS) and other molecules (proteases, DAMPs, IgG, and IgM). For this reason, it is imperative to develop strategies that act on different molecular mechanisms for reducing damage. In the chronic phase, the therapeutic strategies should directed to limit the neurodegenerative process and promote a favorable microenvironment for neural restoration. In line with this, glial scar removal, release of anti-inflammatory cytokines (IL-10, IL-13, IL-4, TGF-β), growth factors (BDNF, NGF, EGF, IGF-1), and neurotrophins (NT3, 4, and 5) could play a significant role.
Figure 1.
Self-destructive mechanisms and therapeutic targets in different phases of SCI. In the acute and subacute phases, resident and peripheral cells determine neural destruction through pro-inflammatory cytokines (IL-1β, IL-6, TNF-α, IFN-γ, IL-12), chemokines (MCP-1, CXCL10, CCL2, CXCL2), free radicals (ROS and NO produced by iNOS) and other molecules (proteases, DAMPs, IgG, and IgM). For this reason, it is imperative to develop strategies that act on different molecular mechanisms for reducing damage. In the chronic phase, the therapeutic strategies should directed to limit the neurodegenerative process and promote a favorable microenvironment for neural restoration. In line with this, glial scar removal, release of anti-inflammatory cytokines (IL-10, IL-13, IL-4, TGF-β), growth factors (BDNF, NGF, EGF, IGF-1), and neurotrophins (NT3, 4, and 5) could play a significant role.
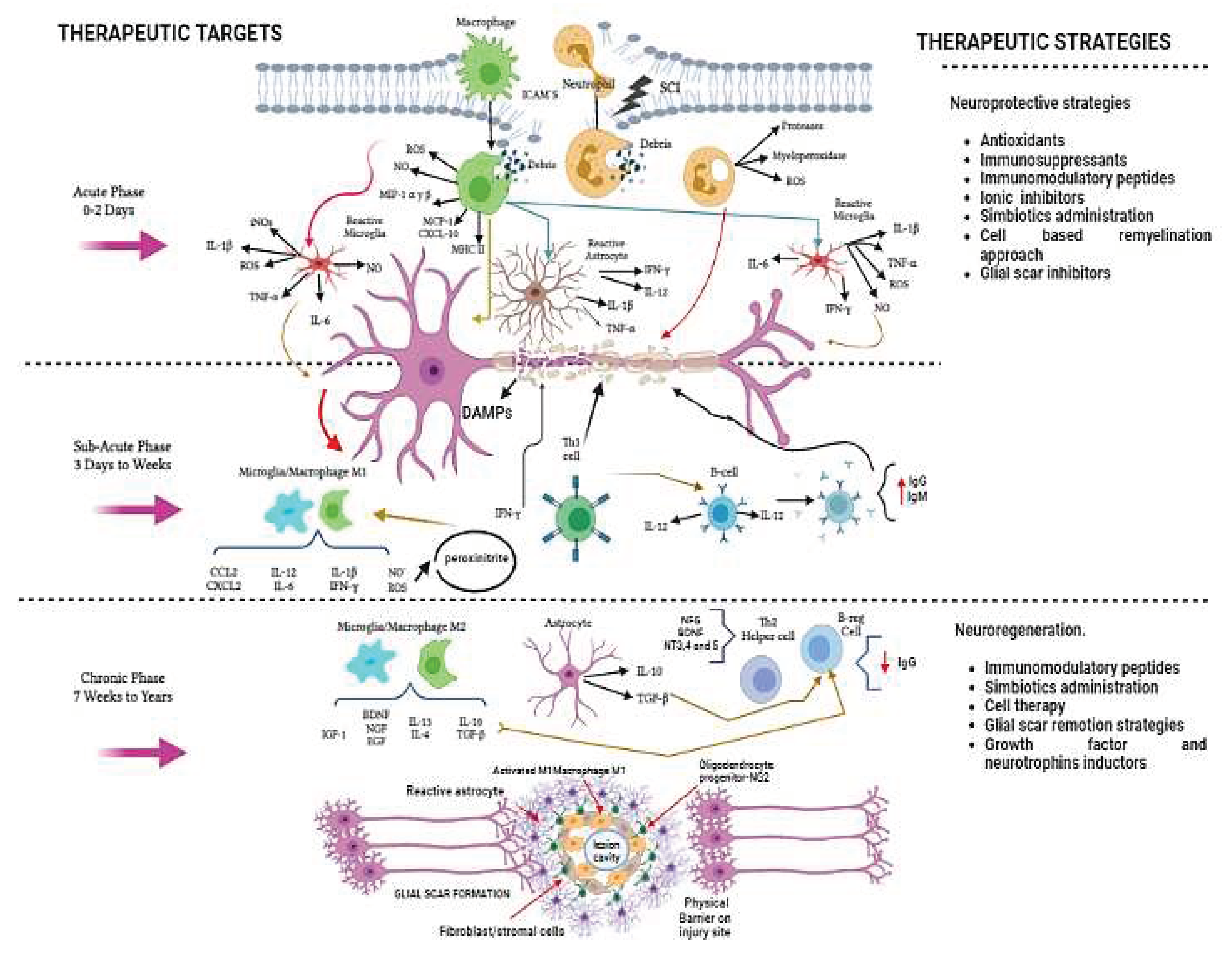

Table 1.
Immune responses in different cell types.
| Cells | Immune Responses | Effect of SCI | References |
|---|---|---|---|
| SCs |
|
Neuroprotection and neuroregeneration | Painter et al., 2014; Meyer et al., 2008; Tzekova et al., 2014; Monje et al., 2021; Niapour et al., 2012 |
| OECS |
|
Neuroprotection |
Jiang et al. 2022; Khankan et al., 2016; Lopez-Valej et al., 2004; Zhang et al., 2019. |
| NG2 |
|
Neuroprotection |
Knerlich-Lukoschus et al., 2010; Kang et al., 2013. |
| BMSCs |
|
Neuroprotection and neuroregeneration | Pang et al., 2021; Nakajima et al., 2012; Urdzíková et al., 2014; Jorgensen 2010; Lindsay and Barnett 2021. |
| NSCs |
|
Neuroprotection. | Shao et al., 2019; Cheng et al., 2016; Rong et al., 2019. |
| Aldynoglia |
|
Neuroprotective and neuroregeneration | Buzoianu-Anguiano et al., 2021; Ma et al., 2020. |
Table 2.
Chronology of A91 Immunization results.
| References. | A91 immunization alone. |
|---|---|
| Ibarra et al., 2004 | The beneficial effect of immunization with A91 was demonstrated if applied alone compared with the administration of methylprednisolone, reflecting an increase in motor recovery and rubrospinal neurons. |
| Ibarra et al.,2010 | A91-immunization reduces lipid peroxidation levels. |
| Martiñon et al.,2012 | SCI inflicted by moderate contusion or incomplete transection showed significant anti-A91 T cell proliferation that was mainly characterized by predominant IL-4 production and BDNF release. It was not the same for severe contusion or complete transection. |
| García E et al 2012 | Immunization with A91 peptide or copolymer-1 reduces NO production and iNOS gene expression in rat and mouse SCI models. |
| Ibarra et al., 2013 | Prophylactic neuroprotection with A91 vaccination induces significant survival of rubrospinal neurons and ventral horn neurons after injury. |
| Rodríguez-Barrera et al., 2013 | A91-immunization reduces apoptosis caused by SCI by decreasing Casp3 activity and TNF-α levels. |
| Martiñon et al.,2016 | A91-immunization induces a long-term production of BDNF and NT-3 and a substantial improvement in motor recovery in chronic stages of SCI. |
| Garcia E et al, 2018 | A91 or Cop-1-immunization significantly reduce IL-6, IL-1β, TNFα and increase IL-10, IL-4 and IGF-1 gene expression after moderate concussion. In contrast, the opposite pattern was seen in severe spinal cord contusion. |
| García E. et al 2019 | Combination therapy that integrated surgical removal of glial scars, an adhesive fibrin matrix with MSCs, and A91 was the best strategy to promote motor and sensibility recovery. In addition, significant increases on tissue preservation and axonal density were observed suggest that these exhibits potential effects on the protection and regeneration of neural tissue after acute spinal cord injury |
| A91-immunization combined with other strategies | |
| Martiñon et al., 2007 | A91-immunization plus GSHE induces better motor recovery, a greater number of myelinated axons, and better survival of rubrospinal neurons than immunization alone. |
| Del Rayo Garrido et al., 2013 and Martiñon et al., 2007. | The combination of A91-immunization and GSHE was shown to have superior effects when given immediately after SCI or even within the first 72 hours after injury. |
| Parra-Villamar et al., 2021 | A91-immunization combined with MLIF and GSHE, induced a better preservation of the spinal cord parenchyma and axonal fibers, increased the number of motor neurons, and reduced the presence of collagen. |
| A91- immunization with another regeneration strategies | |
| Wang Y. et al., 2015 | A91-immunization and DC enhanced the expression level of BDNF and NT-3 and exerted a neuroprotective effect and possible regeneration in an SCI mouse model. |
| Rodríguez-Barrera et al., 2017, | The effect of A91-immunization alone versus A91-immunization with scar removal on motor recovery, could modify the non-permissive microenvironment prevailing at the chronic phase of SCI, providing the opportunity of improving motor recovery. |
| Ibarra A. et al 2019 | Combination therapy promotes motor and electrophysiological recovery in the chronic phase of SC injury subsequent to a complete transection. Likewise, it is capable of inducing the permissive microenvironment to promote axonal regeneration. |
| Rodríguez-Barrera et al., 2020 | Combination therapy that integrated surgical removal of glial scars, an adhesive fibrin matrix with mesenchymal stem cells, and A91 modifies the non-permissive microenvironment post SCI, but it is not capable of inducing an appropriate axonal regeneration or neurogenesis when compared to the treatment with A91alone |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



