
Submitted:
27 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
The outermost layer of the heart, the epicardium, is an essential cell population that contributes, through epithelial-to-mesenchymal transition (EMT), to form different cell types and provide paracrine signals to the developing heart. Despite its quiescent state during adulthood, in response to cardiac injury, adult epicardium reactivates and recapitulates many aspects of embryonic cardiogenesis supporting cardiac tissue remodeling. Thus, the epicardium has been considered as a crucial source of cell progenitors with important contribution to developing and injured heart. Although several studies have provided evidence about cell fate determination in the epicardium, to date it is unclear whether epicardial-derived cells (EPDCs) come from specific, and previously predetermined, epicardial cell subpopulations or if they are derived from a common progenitor. In recent years, different approaches have been used to study cell heterogeneity within the epicardial layer by using different experimental models. However, generated data are still insufficient to understand the complexity of this epithelial layer. In this review, we summarize the previous works supporting the cellular composition, molecular signatures and diversity within the developing and adult epicardium.
Keywords:
epicardium
; heterogeneity
; cardiac development
; cardiac repair
1. Introduction
The epicardium is an epithelial layer covering the myocardium which plays a key role in the developing heart, being a crucial source of cells and signaling. During cardiac development, epicardial cells undergo epithelial-to-mesenchymal transition (EMT), giving rise to epicardial-derived cells (EPDCs) that internalize into the myocardium and differentiate into distinct cardiac cell types, such as coronary vascular smooth muscle cells (vSMCs), cardiac fibroblasts (CFs) or endothelial cells (ECs) [1,2,3,4]. Although the epicardium remains a quiescent layer during adulthood, it can be reactivated in response to cardiac injury by upregulating its embryonic developmental genetic program [5,6]. Thus, the epicardium constitutes an intriguing cell population of multipotent progenitors to study and may have a potential contribution to cardiac wound healing by regulating tissue remodeling after heart damage.
However, despite its essential role during development, maintenance of cardiac homeostasis and after cardiac injury, fundamental insights into the epicardium cell heterogeneity and functional crosstalk with other cell types are currently lacking. Furthermore, little is known about how different cell types emerge from EPDCs or the molecular mechanisms underlying EPDCs cell fate during development and reactivation after heart damage. In recent years, different approaches have been used to study cell heterogeneity and molecular signatures within the epicardial cell layer in embryonic and adult hearts. In spite of this knowledge, there are many open questions regarding epicardial cell behavior, the resolution of which are necessary to better understand the role of this epithelial layer during development and heart diseases. Herein, we provide a state-of-the-art review of current studies on epicardial cell heterogeneity in different experimental models during embryonic development and adulthood.
2. Epicardium during cardiac development and regeneration
An epithelial layer of mesothelial-derived cells covering the outermost of the heart can be observed in the vertebrate heart. This evolutionarily conserved structure, called the epicardium, harbors a population of progenitor cells that undergo EMT prior to differentiating into distinct cardiac lineages [7,8,9]. During this transition, epithelial cells lose their cell-cell adhesions and their apical-basal polarity acquiring migratory and invasive characteristics of mesenchymal stem cells that allow its internalization [9,10,11]. Once in the myocardium, the delaminated EPDCs differentiate into specialized cells, including coronary vascular smooth muscle cells, cardiac fibroblasts, endothelial cells and presumably a subpopulation of cardiomyocytes [2,3,4,12]. However, the role of the epicardium during development is not limited to serve as a progenitor source, contributing multipotent cells that give rise to cardiac mesenchyme. This epithelial layer is also a source of paracrine cues and ECM components that are essential for fetal cardiac growth, coronary vessel patterning and regenerative heart repair [13]. After birth, the postnatal epicardium enters a quiescent state, being a simple barrier between the myocardium and the pericardial cavity.
However, in response to cardiac damage, the adult epicardium reactivates an embryonic-like response in which developmental gene programs are upregulated to modulate tissue repair [6,14,15,16]. Although during the cardiac regenerative process, epicardial cells undergo EMT and migrate into the underlying tissue, the adult epicardium is thought to serve more as a source of paracrine signals rather than a source of “reparative” cells [15,17,18]. In recent years, the use of lineage tracing models has revealed that the majority of newly differentiated cells that appear after cardiac injury arise from preexisting cardiac cell subpopulations, such as smooth muscle cells, endothelial cells or cardiomyocytes [19,20,21,22,23]. Thus, the contribution of EPDCs post-injury appears to be less efficient compared to its role during development [14,24].
Apart from this fact, it should not be forgotten that the epicardium is a crucial player involved in cardiac development and tissue remodeling after heart damage. Although in recent years, the epicardium has been proposed as a potential target in the treatment of cardiovascular disease, a deeper understanding is needed to unlock its full potential as a source of cells and signals that satisfactorily complete the reparative process.
3. Determining epicardial heterogeneity: do different cell types constitute the epicardial layer?
Identification of epicardial cell heterogeneity and cellular intercommunication can play a vital role in differentiating healthy from diseased hearts and it may predict future outcomes with superior precision and at earlier stages of cardiac failure. Fluorescence-activated cell sorting (FACS) is a powerful tool that allows the identification of specific cells; however, it affords some limitations in sample preparation or a paucity of markers that can be assayed simultaneously [25]. Therefore, FACS is not suitable for the subclassification of unknown cells and the identification of their potential functions. Although, in recent years, Cre-based lineage tracing models have been used to understand EPDCs cell fate, most of them are not sufficiently specific to the epicardial layer and have been supplemented or replaced by single-cell RNA sequencing (scRNA-seq) techniques [5]. These methods have enabled us to gain a high-power view of novel individual cellular phenotypes, identifying new cell types and subpopulations with distinct cellular expression patterns by integrating the transcripts of different genes even within an apparently homogeneous cell population [26,27]. In this context, by using scRNA-seq and transcriptomic analyses, Weinberger et al. (2020) have described three epicardial cell subpopulations with distinctive spatial distribution during cardiac development in zebrafish embryos, concretely on day 5 post-fertilization [28]. Interestingly, only one of these transcriptionally distinct epicardial cell subpopulations contained cells co-expressing the prototypical epicardial signature genes tcf21, tbx18 and wt-1b. In addition, this cell cluster expressed high levels of tgm2b (transglutaminase 2b), which acts to ensure cell-cell contact and plays a critical role in maintaining the integrity of the forming epicardium. The second subpopulation was enriched for genes associated with the regulation of epicardial cell contribution to the smooth muscle layer of the outflow tract such as tbx18, acta2 (actin alpha 2) and mylka (myosin light chain kinase a). This scRNA-seq analyses defined a third cell cluster spatially restricted to an area between the BA and the atrium that was characterized by the high expression of genes involved in leukocyte chemotaxis and guidance cues [28]. Altogether, the appearance of three distinct cell subpopulations, with different localizations and functions, suggests that the epicardium in the zebrafish developing heart is a heterogenous cell layer.
In developing mammals, such as mice and humans, epicardial cells heterogeneously express the transcription factors TCF21 and WT1, both are also expressed in PE cells and downregulated in the adult epicardium but reactivated after myocardial ischemic injury [29,30]. However, little is known about the molecular regulation of epicardial cell heterogeneity and its different functions. In this context, scRNA-seq assays of EPDCs derived from human pluripotent stem cells have identified basonuclin (BNC1) as an upstream regulator of a transcriptional hierarchy regulating cell identity in the developing epicardium. This transcription factor is expressed in the adult epicardium and downregulated after myocardium injury. BNC1 has been identified, together with the membrane protein THY1 (Thy-1 cell surface antigen), as a marker of different epicardial subpopulations [29,31]. Thus, TCF21high/THY1+ (Thy-1 cell surface antigen) cell population had a higher propensity to become CFs whereas BNC1high cell cluster expressed genes involved in muscle differentiation, migration and cell-cell interaction [29].
As mentioned above, scRNA-seq is a powerful tool for studying the cellular mechanisms involved in heart development, allowing us to characterize different cell types and generating hypotheses about their origins and fates [32]. Therefore, scRNA-seq enables the dissection of cellular heterogeneity in an unbiased manner with no need for any prior knowledge of the cell population [27]. However, this recent approach does not preserve spatial information about tissue morphology and cellular interactions [32]. Thus, to obtain more detailed analyses that can provide us a more global view on epicardial development, scRNA-seq studies were combined with spatiotemporal transcriptomics, in which several timepoints along the studied period were considered. In this context, two heterogeneous epicardial cell subpopulations expressing genes that encodes integral membrane proteins such as Upk1b (uroplakin-1b) and Upk3b (uroplakin-3b) have been identified in the outflow tract region between E11.5 to E13 stages in mice [33,34]. In a similar study, integrating scRNA-seq and spatial RNA-seq data, epicardial cells were identified in 5 main cell clusters based on their position within different Hamburger-Hamilton ventricular development stages in the chicken embryo. The timepoints analyzed represented key steps of epicardial development ranging from epicardium prior EMT (HH24), during EMT (HH31) and during/after epicardial differentiation (HH36 and HH40). Thus, at HH24, an early epicardial progenitor cell cluster, characterized by the expression of canonical epicardial progenitor markers such as TCF21, TBOX and WT1 was identified restricted to the epicardial layer of the ventricular walls. In this region, another subpopulation enriched in BMP4 (bone morphogenetic protein 4) and LUM (lumican) was described at HH31, when EMT is taking place. In the third cell cluster described within the myocardium, high levels of extracellular matrix transcripts implicated in cell migration were observed at the same stage. Interestingly, as Mantri et al. (2021) have suggested, this subpopulation may represent a mesenchymal phenotype of EPDCs that are undergoing EMT and migrating into the myocardium. At HH36, a fibroblast-like cell cluster, that expressed COL3A1 (collagen type III), and a mural cell cluster enriched, among others, in ACTA2 (actin alpha 2) and MYH11 (myosin heavy chain 11) were present within the myocardium. Furthermore, at the same stage, some cells of the outermost epicardial layer still maintain an undifferentiated phenotype [35].
Although no functional epicardial cell heterogeneity was observed in Mantri et al. (2021) study, a variation in the differentiation state was described, as cells may continue to stay in the epicardium in an undifferentiated intermediate phenotype during later stages of development, where EMT have been initiated. Thus, their data hypothesized that EPDCs, which undergo EMT, maintain a progenitor-like transcriptional profile before fate specification in the myocardium [35]. These results suggest that the observed heterogeneity does not stem from differences in the initial cell population. Although what triggers EPCDs to undergo EMT at specific developmental times and locations remains unknown, this study suggests that extracellular matrix (ECM) cues are significantly involved in this process [35]. Similarly, recent studies have demonstrated that in the developing mouse heart, ECM is a key player during EMT. Thus, in those regions near epicardial cells undergoing EMT a variation in ECM composition has been described, characterized by a less amount of integrin α4 and laminin. Additionally, other ECM components such as the basement membrane-associated proteoglycan agrin were upregulated in EPDCs during EMT, enhancing their migration by decreasing β-catenin and promoting pFAK (phospho-focal adhesion kinase) localization at focal adhesions. In both mouse and human models, agrin promoted epicardial EMT through the aggregation of dystroglycans to the Golgi apparatus, connecting extracellular signals from ECM with intracellular pathways [36]. Interestingly, these data suggest that EPDCs differentiation trajectories do not only depend on epicardial cell transcriptome but also on extrinsic cues such as the ECM formed by epicardium and EPDCs during cardiac development or induced by paracrine factors [37].
Even though cellular heterogeneity is a general feature of biological tissues that it has been identified after sequencing analysis in multiple organisms, even within an apparently homogeneous cell population; some controversial results have been observed linked to cell heterogeneity within epicardial layer [27]. Thus, the conversion of early epicardial Wt1high/Tcf21high cells to Wt1low/Tcf21high mesenchymal cells during EMT reflects a developmental transition rather than heterogeneity of the starting population in the developing murine heart. These results suggest that the different subpopulations that have been described are more likely a result of developmental progression and that the fate of EPDCs is specified after EMT, potentially in response to extrinsic cues like paracrine factors or ECM [37]. In contrast, similar analyses in the human heart at different time points during development (4.5-5, 6.5 and 9 post-conception weeks) showed that epicardium display less gene expression heterogeneity compared to more undifferentiated mesenchymal. However, the authors argued that the low number of sequenced cells and the limited number of genes analyzed were insufficient for a detailed comprehension of what is happening in epicardial cells throughout the period under study [38].
In contrast to the developing epicardium, in the postnatal period, this epithelial layer maintains a quiescent state characterized by reduced expression of developmental genes such as Wt1 and Tbx18. Furthermore, no cellular contribution to the myocardium has been observed in this quiescent state [15,16,39]. Nevertheless, in response to cardiac damage, the epicardium suffers a reactivation initiating an embryonic-like response [15,16]. Thus, reactivated epicardial cells can form a multi-cell layer of epicardial stromal cells (EpiSc) on the surface of the heart that secrete paracrine factors to stimulate cardiomyocyte growth, angiogenesis and adaptive immune regulation [15,16,40]. In this context, FGF2 and VEGFA secreted by EPDCs have been described to increase vessel density after myocardial infarction in a mouse model [15]. Hence, the epicardium plays a crucial role as a signaling center being a resident population of progenitor cell involved in cardiac repair, accomplishing cell repopulation of the damaged area and promoting the regenerative responses against cardiac injury [39,41]. For instance, in the mice model WT1CreERT2/+; ROSA26-tdT+/- of atrial cardiomyopathy, reactivated epicardial cells differentiate into myofibroblasts during tissue remodeling [42].
Although it is generally assumed that the adult activated epicardium recapitulates an embryonic-like response, characterized by the generation of mesenchymal progenitor cells, after ischemic insults or cardiac injury there may be important differences concerning their embryonic phenotype [43]. In the adult heart, the reparative process appears to be less efficient at EPDCs migration and differentiation, producing signals that activate the proliferation of resident CFs and fibrosis induction [44]. Furthermore, it remains unknown whether this disparity in regenerative capacity could be derived from differences in the cellular composition of the epicardium, due to the presence of subsets of cells that contribute more to cardiac repair. In this context, several groups have made significant efforts to better understand cell heterogeneity and molecular signatures within the epicardial layer of the adult heart. For instance, Cao et al. (2016) have reported three different cell subsets defined by distinct gene expression signatures in the tcf21+ epicardial population isolated from adult zebrafish ventricles. One subpopulation represents the outermost epithelial epicardial cells; the second cell cluster, composed of internal cells ubicated in the middle of the ventricular wall, was characterized by likely containing perivascular components and the third one formed an innermost layer of EPDCs [45].
In mammals, epicardium activation after MI induction and thymosin beta 4 (Tβ4) administration revealed the presence of distinct subpopulations of EPDCs in the activated epicardium with variated cardiovascular potential and molecular phenotypes distinct from embryonic EPDCs (E12.5) [43,46]. However, it is important to highlight that the origin of the different cell populations observed in these studies is unclear and remain to be elucidate if such cell heterogeneity is already present within the “quiescent” epicardial layer.
In recent years, scRNA-seq combined with RNA in situ hybridization and lineage tracing has allowed the identification of 11 transcriptionally distinct EpiSC populations isolated from mouse hearts after MI, which can be classified into three independent groups [47]. Based on their data, it has been hypothesized that these populations are, respectively, involved in the secretion of paracrine factors, the attraction of monocytes and neutrophils during the modulation of the innate immune response post-MI and exhibit a fibroblast-like phenotype [47]. However, despite analysis using a high-resolution technique, their results did not assess the function of the identified cell subpopulations and came from a singular timepoint analysis. Even though, these results bear some resemblance to the functionally heterogeneous epicardial subpopulations described in zebrafish by Weinberger et al. [28,47].
4. Does the embryonic origin of the epicardium underlie its cell heterogeneity?
As mentioned above, during development, a subset of epicardial cells undergoes EMT and migrates into the myocardium to give rise several cardiac cell types. This fact, by which epicardial cells in the developing heart can differentiate into several distinct cell subpopulations, suggests that this epithelial layer is not composed of only one specific cell type. The hypothesis of the presence of different cell clusters along the epicardial layer can be supported by the notion that multiple tissues contribute to its embryonic origin. In this context, it has been proposed that the epicardial heterogeneity may be attributed, at least in part, to the origin and cellular composition of the proepicardium (PE) or proepicardial organ (PEO), a heterogeneous cell mass from which most epicardial cells originate, which could influence the multiple cell fates of EPDCs during heart development [48]. Therefore, the PE is an extracardiac primitive organ formed as an outgrowth of the coelomic mesothelium located between the heart and the liver [49]. In the mouse, at embryonic day 8.5 (E8.5), the PE is situated at the base of the venous inflow tract of the developing primitive heart and does not interact directly with the myocardium [50,51,52]. The PE has an outer layer of epithelial cells expressing well-characterized proepicardial markers such as Wilms´ Tumor-1 (WT-1) or Tbx-18 (T-box transcription factor 18). Those cells, which originate from precursors of the early cardiac progenitor fields that express the transcription factors Nkx2.5 (Homeobox protein Nkx2.5) and Isl-1 (Insulin gene enhancer protein ISL-1), overlie an inner core of several mesenchymal cell types and endothelial cells [53]. Thus, despite PE seams morphologically homogeneous, those different molecular markers have a heterogeneous spatiotemporal expression in the PE suggesting the existence of several subtypes of cells with distinct roles that divide the PE into genetically distinguishable cell subcompartments. Therefore, apart from Tbx18- and WT-1-expressing cell populations, lineage tracing experiments in mice and chicken have shown that some proepicardial cells from the mesenchymal core partially express other markers such as Scx (Scleraxis) and Sema3D (Semaphorin 3D) [37,54]. Curiously, at early stages, Scx+ cell population contributes to the formation of the endocardium and Sema3D+ cells to the endothelium of the sinus venosus. It is important to note that both tissues are linked to the formation of the endothelium of coronary vasculature at later stages [54].
In addition, it has been shown that the PE is an important transient structure that contributes to different types of cells from various cardiac lineages; for instance, studies in avian and mammalian models have established that this compartmentalized structure is a source of vSMCs, myocytes and CFs [53,54,55]. In this context, Cossett & Misra (2011) have identified three different populations of endothelial cell precursors within the PE that appear to have distinct origins, such as the developing liver bud or the sinus venosus, suggesting that the PE “per se” may also acts as a source of endothelial cells [56]. Thus, there are previous evidences highlighting the possibility that the PE primitive organ might include distinct cell subcompartments containing different cardiac cell precursors populations that differ in both the routes and timing of migration to the heart; to first constitute the epicardium and, after, to give rise to distinct, albeit overlapping, cell fates [54]. However, in recent years, new findings contrast with the idea of proepicardial subcompartments reported by Katz et al. (2012), suggesting that, in mice, all PE and epicardial cells co-express Wt1, Tbx18, Tcf21 (Transcription factor 21), Scx and Sema3D until the embryonic stage E13.5 (Figure 1). Additionally, the minimal variation in the expression levels of those canonical markers between cells does not provide evidence for the existence of distinct areas in the PE or epicardial layer [37].
Once the embryonic heart has looped, clusters of PE cells begin to proliferate and spread, covering the bare heart tube with an epithelial layer, the epicardium [57]. When the epicardium completely covers the heart, around E11.5 in mice and week 5 in human embryos, it constitutes a heterogeneous multicellular epithelium lining the ventricles, expressing different specific proteins such as WT1, TBX18, TCF21, GATA5 (GATA binding protein 5) and cytokeratin [53,55,58,59,60] (Figure 1). Furthermore, other clusters such as CD45+ cells have been identified within the epicardium. However, whether the different cell populations identified in the PE represent distinct stages of cell differentiation or if they constitute different cell precursors that give rise to specific cell populations remains to be investigated [61].
Although most epicardial cells originate from a heterogeneous cell mass called the PE, part of the epicardium covering the bulbus arteriosus (BA) in zebrafish was found to be derived from the pericardial sac [62,63,64,65]. In the developing mouse heart, Tyser et al. (2021) have described a novel source of proepicardial cells derived from the juxta-cardiac field (JCF); a population of progenitor cells located rostral to the cardiac crescent which represents the earliest known progenitors of the epicardium. Using single-cell resolution time-lapse imaging and genetic lineage labeling, this group established that the JCF, characterized by the Mab21l2 (male-abnormal 21-like 2) expression, constitutes a cardiac progenitor cell pool that is spatially and transcriptionally distinct from the sinus venosus progenitors. Thus, JCF progenitor cells may have specific abilities to differentiate into cardiomyocytes within the linear heart tube and, also, contribute to the PE. However, it is still unclear whether the JCF population is composed of unipotent progenitors of cardiomyocytes and proepicardial cells or contains bipotent cells capable of giving rise to both cell types [32] (Figure 1).
Once the epicardium layer is formed, during heart development, a subset of epicardial cells loses their apical-basal polarity and cell-cell adhesions undergoing an EMT, which allows the formation of EPDCs [10,11]. Thereby, these epicardial cells can migrate and invade the myocardium and differentiate into various cell types such as fibroblasts or smooth muscle cells [2,3]. However, it remains unclear whether pre-migratory EPDCs are a homogeneous group of multipotent cell progenitors or if they are somehow specified as epicardial subpopulations within the epicardium before migration. In addition, many other questions are open regarding epicardial cell behavior during EMT process. For instance, why do some epicardial cells undergo EMT while others remain as epithelial cells? Which are the molecular mechanisms that drive epicardial cell dynamic during the process of EMT?
4. Reflections and future perspectives
Far from being just the outermost layer of the heart, the epicardium plays an essential role during cardiac development, being a source of cardiac cells, secretory and paracrine factors. After a cardiac injury, this epithelial layer, which remains in a quiescent state during adulthood, recapitulates embryonic capabilities, providing the epicardium the ability to contribute to cardiac repair. Although it contributes in a less efficient manner compared to its role during development, the epicardium has been considered a very appealing actor in endogenous cardiac remodeling after damage. However, to date, it is still unclear whether specific epicardial cell subpopulations can participate in cardiac remodeling or whether distinct cell types residing within the epicardial layer have determined capabilities during the reparative response [5].
In this context, several studies over the last few years have been focused on understanding epicardial composition as well as the molecular mechanisms underlying epicardial cell fate decisions to identify targets to optimize the post-injury response in the adult heart. However, it is common for the potential heterogeneity of the epicardium to be overlooked due to a low number of epicardial cells, few developmental stages under study or incorrect markers used to identify these novel subpopulations [66,67]. These factors, along with fluctuations in the abundance of diverse cellular lineages, spatial organization, molecular composition and interactions, as well as interspecies differences complicate the interpretation of the results and its extrapolation to humans. Moreover, it is important to note that some of the aforementioned studies were performed in different organisms and were confined to a limited developmental timeframe. However, in those approaches, in which different time points have been studied, the dataset was not able to confirm functional heterogeneity, suggesting that this divergence seems to be due to a varying differentiation state or a reflection of the transcriptional changes rather than the origin of the distinct subpopulations of cells [35,38]. In the adult heart, scRNA-seq analysis after cardiac injury has suggested the presence of diverse clusters of EPDCs during the reparative response. Furthermore, in most of these studies, epicardial cell characterization was based on known markers and was not further scrutinized; thus, novel subpopulations are potentially not identified [5].
Nevertheless, if there is anything evident regarding the epicardial heterogeneity and its composition, it is the need for further analysis to distinguish epicardial cells from EPDCs, in which the transcriptional signature is altered. Although in recent years, scRNA-seq and high-resolution analysis have postulated that there is a heterogenic environment within the epicardial layer, thousands of generated data are still unanalyzed, harboring rare cell populations and interactions within the epicardium waiting to be discovered. It is also important to pay special attention to the developmental stage by performing different analyses at multiple time points during embryonic development and in the adult epicardium after cardiac injury. Thus, studies during embryonic development and cardiac disease can help us to have a more comprehensive view of the heterogeneity of the epicardium, identifying epicardial subpopulations and their role during heart formation and in the repair process, either through cellular contributions or via paracrine signaling. Those findings could shed more light on the optimization of the post-injury response, leading to the identification of mechanisms that contribute to cardiac repair and regeneration.
Author Contributions
Conceptualization, CS-F and AA.; CS-F and AA.; writing—original draft preparation, CS-F and AA.; writing—review and editing, CS-F and AA.; review the manuscript critically for important intellectual content, LR-O, LM-V, FR, FH-T, EL-V, JND and DF. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Spanish Ministry of Science and Innovation, grant number PID2019-107492GB-I00; and by Regional Government of Andalucía, Department of University, Research and Innovation, grant number ProyExcel_00513.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trembley, M.A.; Velasquez, L.S.; Bentley, K.L.d.M.; Small, E.M. Myocardin-related transcription factors control the motility of epicardium-derived cells and the maturation of coronary vessels. Development 2015, 142, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Grieskamp, T.; Rudat, C.; Lüdtke, T.H.-W.; Norden, J.; Kispert, A.; P, Q.; M, T.; E, S.; G, L.; G, D.; et al. Notch Signaling Regulates Smooth Muscle Differentiation of Epicardium-Derived Cells. Circ. Res. 2011, 108, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Moore-Morris, T.; Cattaneo, P.; Puceat, M.; Evans, S.M. Origins of cardiac fibroblasts. J. Mol. Cell. Cardiol. 2015, 91, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Carmona, R.; Barrena, S.; Gambero, A.J.L.; Rojas, A.; Muñoz-Chápuli, R. Epicardial cell lineages and the origin of the coronary endothelium. FASEB J. 2020, 34, 5223–5239. [Google Scholar] [CrossRef] [PubMed]
- Streef, T.J.; Smits, A.M. Epicardial Contribution to the Developing and Injured Heart: Exploring the Cellular Composition of the Epicardium. Front. Cardiovasc. Med. 2021, 8. [Google Scholar] [CrossRef]
- Redpath, A.N.; Smart, N. Recapturing embryonic potential in the adult epicardium: Prospects for cardiac repair. STEM CELLS Transl. Med. 2020, 10, 511–521. [Google Scholar] [CrossRef]
- Zhou, B.; Ma, Q.; Rajagopal, S.; Wu, S.M.; Domian, I.; Rivera-Feliciano, J.; Jiang, D.; von Gise, A.; Ikeda, S.; Chien, K.R.; et al. Epicardial progenitors contribute to the cardiomyocyte lineage in the developing heart. Nature 2008, 454, 109–113. [Google Scholar] [CrossRef]
- Carmona, R.; Guadix, J.A.; Cano, E.; Ruiz-Villalba, A.; Portillo-Sánchez, V.; Pérez-Pomares, J.M.; Muñoz-Chápuli, R. The embryonic epicardium: an essential element of cardiac development. J. Cell. Mol. Med. 2010, 14, 2066–2072. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef]
- von Gise, A.; Pu, W.T.; H, Z.; K, L.; B, Z.; L, Z.; M, O.; L, Y.; Q, M.; N, S.; et al. Endocardial and Epicardial Epithelial to Mesenchymal Transitions in Heart Development and Disease. Circ. Res. 2012, 110, 1628–1645. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed]
- del Moral, S.D.; Barrena, S.; Hernández-Torres, F.; Aránega, A.; Villaescusa, J.M.; Doblas, J.J.G.; Franco, D.; Jiménez-Navarro, M.; Muñoz-Chápuli, R.; Carmona, R. Deletion of the Wilms’ Tumor Suppressor Gene in the Cardiac Troponin-T Lineage Reveals Novel Functions of WT1 in Heart Development. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Masters, M.; Riley, P.R. The epicardium signals the way towards heart regeneration. Stem Cell Res. 2014, 13, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Poss, K.D. The epicardium as a hub for heart regeneration. Nat. Rev. Cardiol. 2018, 15, 631–647. [Google Scholar] [CrossRef]
- Tricarico, D.; Servidei, S.; Tonali, P.; Jurkat-Rott, K.; Camerino, D.C. Impairment of skeletal muscle adenosine triphosphate–sensitive K+ channels in patients with hypokalemic periodic paralysis. J. Clin. Investig. 1999, 103, 675–682. [Google Scholar] [CrossRef]
- van Wijk, B.; Gunst, Q.D.; Moorman, A.F.M.; Hoff, M.J.B.v.D. Cardiac Regeneration from Activated Epicardium. PLOS ONE 2012, 7, e44692. [Google Scholar] [CrossRef] [PubMed]
- Stevens, S.M.; von Gise, A.; VanDusen, N.; Zhou, B.; Pu, W.T. Epicardium is required for cardiac seeding by yolk sac macrophages, precursors of resident macrophages of the adult heart. Dev. Biol. 2016, 413, 153–159. [Google Scholar] [CrossRef]
- Huang, G.N.; Thatcher, J.E.; McAnally, J.; Kong, Y.; Qi, X.; Tan, W.; DiMaio, J.M.; Amatruda, J.F.; Gerard, R.D.; Hill, J.A.; et al. C/EBP Transcription Factors Mediate Epicardial Activation During Heart Development and Injury. Science 2012, 338, 1599–1603. [Google Scholar] [CrossRef]
- Cai, W.; Tan, J.; Yan, J.; Zhang, L.; Cai, X.; Wang, H.; Liu, F.; Ye, M.; Cai, C.-L. Limited Regeneration Potential with Minimal Epicardial Progenitor Conversions in the Neonatal Mouse Heart after Injury. Cell Rep. 2019, 28, 190–201. [Google Scholar] [CrossRef]
- Sereti, K.-I.; Nguyen, N.B.; Kamran, P.; Zhao, P.; Ranjbarvaziri, S.; Park, S.; Sabri, S.; Engel, J.L.; Sung, K.; Kulkarni, R.P.; et al. Analysis of cardiomyocyte clonal expansion during mouse heart development and injury. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Kanisicak, O.; Khalil, H.; Ivey, M.J.; Karch, J.; Maliken, B.D.; Correll, R.N.; Brody, M.J.; Lin, S.-C.J.; Aronow, B.J.; Tallquist, M.D.; et al. Genetic lineage tracing defines myofibroblast origin and function in the injured heart. Nat. Commun. 2016, 7, 12260. [Google Scholar] [CrossRef] [PubMed]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.-D.; Guerquin-Kern, J.-L.; Lechene, C.P.; Lee, R.T. Mammalian heart renewal by pre-existing cardiomyocytes. Nature 2013, 493, 433–436. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Huang, X.; Kanisicak, O.; Li, Y.; Wang, Y.; Li, Y.; Pu, W.; Liu, Q.; Zhang, H.; Tian, X.; et al. Preexisting endothelial cells mediate cardiac neovascularization after injury. J. Clin. Investig. 2017, 127, 2968–2981. [Google Scholar] [CrossRef]
- Moerkamp, A.T.; Lodder, K.; van Herwaarden, T.; Dronkers, E.; Dingenouts, C.K.E.; Tengström, F.C.; van Brakel, T.J.; Goumans, M.-J.; Smits, A.M. Human fetal and adult epicardial-derived cells: a novel model to study their activation. Stem Cell Res. Ther. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Menasche, B.L.; Crisman, L.; Gulbranson, D.R.; Davis, E.M.; Yu, H.; Shen, J. Fluorescence Activated Cell Sorting (FACS) in Genome-Wide Genetic Screening of Membrane Trafficking. Curr. Protoc. Cell Biol. 2018, 82, e68. [Google Scholar] [CrossRef] [PubMed]
- Packer, J.; Trapnell, C. Single-Cell Multi-omics: An Engine for New Quantitative Models of Gene Regulation. Trends Genet. 2018, 34, 653–665. [Google Scholar] [CrossRef]
- Wen, L.; Tang, F. Single-cell sequencing in stem cell biology. Genome Biol. 2016, 17, 71–71. [Google Scholar] [CrossRef]
- Weinberger, M.; Simões, F.C.; Patient, R.; Sauka-Spengler, T.; Riley, P.R. Functional Heterogeneity within the Developing Zebrafish Epicardium. Dev. Cell 2020, 52, 574–590. [Google Scholar] [CrossRef]
- Gambardella, L.; McManus, S.A.; Moignard, V.; Sebukhan, D.; Delaune, A.; Andrews, S.; Bernard, W.G.; Morrison, M.A.; Riley, P.R.; Göttgens, B.; et al. BNC1 regulates cell heterogeneity in human pluripotent stem cell derived-epicardium. Development 2019, 146. [Google Scholar] [CrossRef]
- Braitsch, C.M.; Combs, M.D.; Quaggin, S.E.; Yutzey, K.E. Pod1/Tcf21 is regulated by retinoic acid signaling and inhibits differentiation of epicardium-derived cells into smooth muscle in the developing heart. Dev. Biol. 2012, 368, 345–357. [Google Scholar] [CrossRef]
- Bochmann, L.; Sarathchandra, P.; Mori, F.; Lara-Pezzi, E.; Lazzaro, D.; Rosenthal, N. Revealing New Mouse Epicardial Cell Markers through Transcriptomics. PLOS ONE 2010, 5, e11429. [Google Scholar] [CrossRef] [PubMed]
- Tyser, R.C.V.; Ibarra-Soria, X.; McDole, K.; Jayaram, S.A.; Godwin, J.; Brand, T.A.H.v.D.; Miranda, A.M.A.; Scialdone, A.; Keller, P.J.; Marioni, J.C.; et al. Characterization of a common progenitor pool of the epicardium and myocardium. Science 2021, 371. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, W.; Li, W.; Li, Y.; Priest, J.R.; Zhou, B.; Wang, J.; Zhou, Z. Single-Cell RNA-Seq of the Developing Cardiac Outflow Tract Reveals Convergent Development of the Vascular Smooth Muscle Cells. Cell Rep. 2019, 28, 1346–1361. [Google Scholar] [CrossRef] [PubMed]
- Rudat, C.; Grieskamp, T.; Röhr, C.; Airik, R.; Wrede, C.; Hegermann, J.; Herrmann, B.G.; Schuster-Gossler, K.; Kispert, A. Upk3b Is Dispensable for Development and Integrity of Urothelium and Mesothelium. PLOS ONE 2014, 9, e112112–e112112. [Google Scholar] [CrossRef]
- Mantri, M.; Scuderi, G.J.; Abedini-Nassab, R.; Wang, M.F.Z.; McKellar, D.; Shi, H.; Grodner, B.; Butcher, J.T.; De Vlaminck, I. Spatiotemporal single-cell RNA sequencing of developing chicken hearts identifies interplay between cellular differentiation and morphogenesis. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Sun, X.; Malandraki-Miller, S.; Kennedy, T.; Bassat, E.; Klaourakis, K.; Zhao, J.; Gamen, E.; Vieira, J.M.; Tzahor, E.; Riley, P.R. The extracellular matrix protein agrin is essential for epicardial epithelial-to-mesenchymal transition during heart development. Development 2021, 148. [Google Scholar] [CrossRef]
- Lupu, I.-E.; Redpath, A.N.; Smart, N. Spatiotemporal Analysis Reveals Overlap of Key Proepicardial Markers in the Developing Murine Heart. Stem Cell Rep. 2020, 14, 770–787. [Google Scholar] [CrossRef]
- Asp, M.; Giacomello, S.; Larsson, L.; Wu, C.; Fürth, D.; Qian, X.; Wärdell, E.; Custodio, J.; Reimegård, J.; Salmén, F.; et al. A Spatiotemporal Organ-Wide Gene Expression and Cell Atlas of the Developing Human Heart. Cell 2019, 179, 1647–1660. [Google Scholar] [CrossRef]
- Smart, N.; Bollini, S.; Dubé, K.N.; Vieira, J.M.; Zhou, B.; Davidson, S.; Yellon, D.; Riegler, J.; Price, A.N.; Lythgoe, M.F.; et al. De novo cardiomyocytes from within the activated adult heart after injury. Nature 2011, 474, 640–644. [Google Scholar] [CrossRef]
- Ramjee, V.; Li, D.; Manderfield, L.J.; Liu, F.; Engleka, K.A.; Aghajanian, H.; Rodell, C.B.; Lu, W.; Ho, V.; Wang, T.; et al. Epicardial YAP/TAZ orchestrate an immunosuppressive response following myocardial infarction. J. Clin. Investig. 2017, 127, 899–911. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Suffee, N.; Moore-Morris, T.; Jagla, B.; Mougenot, N.; Dilanian, G.; Berthet, M.; Proukhnitzky, J.; Le Prince, P.; Tregouet, D.A.; Pucéat, M.; et al. Reactivation of the Epicardium at the Origin of Myocardial Fibro-Fatty Infiltration During the Atrial Cardiomyopathy. Circ. Res. 2020, 126, 1330–1342. [Google Scholar] [CrossRef] [PubMed]
- Bollini, S.; Vieira, J.M.N.; Howard, S.; Dubè, K.N.; Balmer, G.M.; Smart, N.; Riley, P.R. Re-Activated Adult Epicardial Progenitor Cells Are a Heterogeneous Population Molecularly Distinct from Their Embryonic Counterparts. Stem Cells Dev. 2014, 23, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Bertaud, A.; Joshkon, A.; Heim, X.; Bachelier, R.; Bardin, N.; Leroyer, A.S.; Blot-Chabaud, M. Signaling Pathways and Potential Therapeutic Strategies in Cardiac Fibrosis. Int. J. Mol. Sci. 2023, 24, 1756. [Google Scholar] [CrossRef]
- Cao, J.; Navis, A.; Cox, B.D.; Dickson, A.L.; Gemberling, M.; Karra, R.; Bagnat, M.; Poss, K.D. Single epicardial cell transcriptome sequencing identifies Caveolin-1 as an essential factor in zebrafish heart regeneration. Development 2015, 143, 232–243. [Google Scholar] [CrossRef]
- Owenier, C.; Hesse, J.; Alter, C.; Ding, Z.; Marzoq, A.; Petzsch, P.; Köhrer, K.; Schrader, J. Novel technique for the simultaneous isolation of cardiac fibroblasts and epicardial stromal cells from the infarcted murine heart. Cardiovasc. Res. 2019. [Google Scholar] [CrossRef]
- Hesse, J.; Owenier, C.; Lautwein, T.; Zalfen, R.; Weber, J.F.; Ding, Z.; Alter, C.; Lang, A.; Grandoch, M.; Gerdes, N.; et al. Single-cell transcriptomics defines heterogeneity of epicardial cells and fibroblasts within the infarcted murine heart. eLife 2021, 10. [Google Scholar] [CrossRef]
- Simões, F.C.; Riley, P.R. The ontogeny, activation and function of the epicardium during heart development and regeneration. Development 2018, 145, dev155994. [Google Scholar] [CrossRef]
- Smits, A.M.; Dronkers, E.; Goumans, M.-J. The epicardium as a source of multipotent adult cardiac progenitor cells: Their origin, role and fate. Pharmacol. Res. 2018, 127, 129–140. [Google Scholar] [CrossRef]
- Maya-Ramos, L.; Cleland, J.; Bressan, M.; Mikawa, T. Induction of the Proepicardium. J. Dev. Biol. 2013, 1, 82–91. [Google Scholar] [CrossRef]
- Bressan, M.; Liu, G.; Mikawa, T. Early Mesodermal Cues Assign Avian Cardiac Pacemaker Fate Potential in a Tertiary Heart Field. Science 2013, 340, 744–748. [Google Scholar] [CrossRef] [PubMed]
- 52. Wei, K.; Díaz-Trelles, R.; Liu, Q.; Diez-Cuñado, M.; Scimia, M.C.; Cai, W.; Sawada, J.; Komatsu, M.; Boyle, J.J.; Zhou, B.; Ruiz-Lozano, P.; Mercola, M. Developmental origin of age-related coronary artery disease. Cardiovascular Research 2015, 107, 107–294. [Google Scholar]
- Wei, K.; Díaz-Trelles, R.; Liu, Q.; Diez-Cuñado, M.; Scimia, M.C.; Cai, W.; Sawada, J.; Komatsu, M.; Boyle, J.J.; Zhou, B.; Ruiz-Lozano, P.; Mercola, M. Nkx2-5- andIsl1-expressing cardiac progenitors contribute to proepicardium. Biochem.Biophys. Res. Commun. 2008, 375, 450–453. [Google Scholar] [CrossRef]
- Katz, T.C.; Singh, M.K.; Degenhardt, K.; Rivera-Feliciano, J.; Johnson, R.L.; Epstein, J.A.; Tabin, C.J. Distinct Compartments of the Proepicardial Organ Give Rise to Coronary Vascular Endothelial Cells. Dev. Cell 2012, 22, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.-L.; Martin, J.C.; Sun, Y.; Cui, L.; Wang, L.; Ouyang, K.; Yang, L.; Bu, L.; Liang, X.; Zhang, X.; et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature 2008, 454, 104–108. [Google Scholar] [CrossRef]
- Cossette, S.; Misra, R. The identification of different endothelial cell populations within the mouse proepicardium. Dev. Dyn. 2011, 240, 2344–2353. [Google Scholar] [CrossRef]
- Vicente-Steijn, R.; Scherptong, R.W.C.; Kruithof, B.P.T.; Duim, S.N.; Goumans, M.J.T.H.; Wisse, L.J.; Zhou, B.; Pu, W.T.; Poelmann, R.E.; Schalij, M.J.; et al. Regional differences in WT-1 and Tcf21 expression during ventricular development: implications for myocardial compaction. PLOS ONE 2015, 10, e0136025. [Google Scholar] [CrossRef]
- Kikuchi, K.; Gupta, V.; Wang, J.; Holdway, J.E.; Wills, A.A.; Fang, Y.; Poss, K.D. tcf21+ epicardial cells adopt non-myocardial fates during zebrafish heart development and regeneration. Development 2011, 138, 2895–2902. [Google Scholar] [CrossRef]
- MacNeill, C.; French, R.; Evans, T.; Burch, A.W.A.J.B. Modular Regulation of cGATA-5 Gene Expression in the Developing Heart and Gut. Dev. Biol. 2000, 223, 463. [Google Scholar] [CrossRef]
- Peeters, M.-. .-P.F.M.V.; Mentink, M.M.T.; Poelmann, R.E.; Groot, A.C.G.-D. Cytokeratins as a marker for epicardial formation in the quail embryo. Anat. Embryol. 1995, 191, 503–508. [Google Scholar] [CrossRef]
- Balmer, G.M.; Bollini, S.; Dubé, K.N.; Martinez-Barbera, J.P.; Williams, O.; Riley, P.R. Dynamic haematopoietic cell contribution to the developing and adult epicardium. Nat. Commun. 2014, 5, 4054. [Google Scholar] [CrossRef]
- Niderla-BieliŃska, J.; Jankowska-Steifer, E.; Flaht-Zabost, A.; Gula, G.; Czarnowska, E.; Ratajska, A. Proepicardium: Current Understanding of its Structure, Induction, and Fate. Anat. Rec. 2018, 302, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Plavicki, J.S.; Hofsteen, P.; Yue, M.S.; A Lanham, K.; E Peterson, R.; Heideman, W. Multiple modes of proepicardial cell migration require heartbeat. BMC Dev. Biol. 2014, 14, 18–18. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; Phelps, A.; Sedmerova, M.; Wessels, A. Epicardial-like cells on the distal arterial end of the cardiac outflow tract do not derive from the proepicardium but are derivatives of the cephalic pericardium. Dev. Dyn. 2003, 227, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Peralta, M.; González-Rosa, J.M.; Marques, I.J.; Mercader, N. The Epicardium in the Embryonic and Adult Zebrafish. J. Dev. Biol. 2014, 2, 101–116. [Google Scholar] [CrossRef]
- Cui, Y.; Zheng, Y.; Liu, X.; Yan, L.; Fan, X.; Yong, J.; Hu, Y.; Dong, J.; Li, Q.; Wu, X.; et al. Single-Cell Transcriptome Analysis Maps the Developmental Track of the Human Heart. Cell Rep. 2019, 26, 1934–1950. [Google Scholar] [CrossRef] [PubMed]
- Farbehi, N.; Patrick, R.; Dorison, A.; Xaymardan, M.; Janbandhu, V.; Wystub-Lis, K.; Ho, J.W.; E Nordon, R.; Harvey, R.P. Single-cell expression profiling reveals dynamic flux of cardiac stromal, vascular and immune cells in health and injury. eLife 2019, 8. [Google Scholar] [CrossRef]
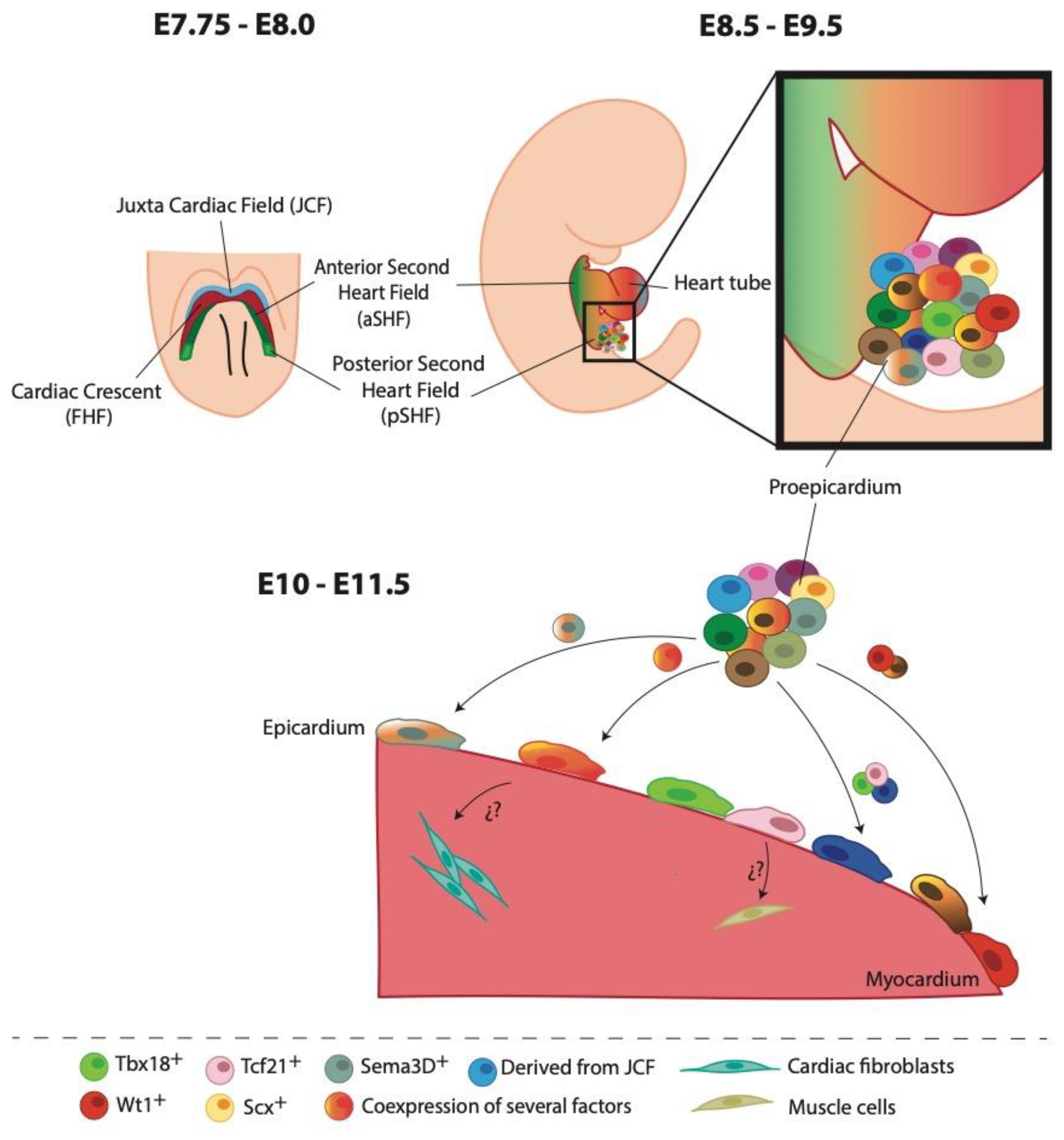
Figure 1.
Cell heterogeneity of the epicardium during heart development. At E7.75-8.0 cardiac progenitor cells extend towards the midline to form the cardiac crescent or first heart field (FHF, shown in red) caudal to the headfolds. Rostral to the FHF, a population of progenitor cells constituted the juxta cardiac field (JCF, shown in light blue). The second heart field (SHF) is composed of two subdomains, anterior (shown in dark green) and posterior (illustrated in light green). At E8.5-9.5, cardiac crescent fusion and posterior looping at the midline form the early cardiac tube. Proepicardium (PE) is situated in the venous inflow tract in the developing primitive heart. Around E10-11.5, clusters of PE cells begin to proliferate and spread covering the heart tube with an epithelial layer, the epicardium. New experimental data have proposed that EPDCs with specific cell fate (fibroblasts or muscle cells) might be originated from different epicardial cell subpopulations.
Figure 1.
Cell heterogeneity of the epicardium during heart development. At E7.75-8.0 cardiac progenitor cells extend towards the midline to form the cardiac crescent or first heart field (FHF, shown in red) caudal to the headfolds. Rostral to the FHF, a population of progenitor cells constituted the juxta cardiac field (JCF, shown in light blue). The second heart field (SHF) is composed of two subdomains, anterior (shown in dark green) and posterior (illustrated in light green). At E8.5-9.5, cardiac crescent fusion and posterior looping at the midline form the early cardiac tube. Proepicardium (PE) is situated in the venous inflow tract in the developing primitive heart. Around E10-11.5, clusters of PE cells begin to proliferate and spread covering the heart tube with an epithelial layer, the epicardium. New experimental data have proposed that EPDCs with specific cell fate (fibroblasts or muscle cells) might be originated from different epicardial cell subpopulations.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



