
Submitted:
29 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
Indole-3-acetic acid (IAA) is a key vegetative hormone that plays crucial roles in plant growth and development. In this study, the effect of the exogenous application of IAA as a biostimulant in the cultivation of onions (Allium cepa) was investigated. Different IAA concentrations were evaluated to determine their effects on onion growth and chemical parameters. Various physical and chemical parameters such as total fresh weight, plant height, chlorophyll content, nitrates, total phenols, and antioxidant capacity (DPPH) were analyzed. The results revealed that exogenous application of IAA had a significant impact on onion growth and quality. Specifically, it was found that certain concentrations of IAA fostered a greater fresh weight of the bulb, an increase in the levels of phenolic compounds and a greater DPPH. However, the onion response to IAA was dependent on the concentration. In conclusion, the present study provides evidence that the exogenous application of IAA as a biostimulant can improve onion growth and quality. These results are relevant for the development of sustainable agricultural practices and could be oriented towards crop improvement.
Keywords:
Indole-3-acetic acid
; biostimulant
; phenolic compounds
; nitrate
; chlorophylls
; exogenous application
1. Introduction
Onion (Allium cepa) is a widely produced crop throughout the world due to its nutritional importance and low cost production [1]. It is a bulb vegetable belonging to the Amaryllidaceae family and it is used in multiple culinary preparations due to its distinctive flavour and organoleptic properties [2,3]. In addition, onion is recognized for its medicinal properties and its benefits for human health. Despite its importance, onion cultivation currently faces various challenges that affect its productivity and quality. Among the main challenges are the impact of diseases caused by pathogens, such as fungus and bacteria [4,5], as well as the presence of insect pests that affect plants health and the quality of the bulb [6,7]. Moreover, abiotic stress, such as adverse weather conditions and water availability, can negatively influence the growth and development of the onion crop [8,9]. The efficiency and quality of crops are fundamental aspects to guarantee food safety [10]. Producers constantly face the challenge of optimizing production and obtain high quality crops in variable conditions according to market demand [11].
In this context, the use of biostimulant has become a promising alternative to improve crop efficiency and quality [12]. Biostimulant are natural or synthetic substances that interact with the growth and development processes of plants, influencing their physiology and metabolism [13]. They can stimulate plant growth, regulate organ development, improve resistance to abiotic and biotic stress [14], as well as increase the quality of agricultural products [15]. Among the most widely used growth regulators there is Indole-3-acetic acid (IAA), a key phytohormone in the regulation of plant growth and development [16,17]. The use of IAA as a growth regulator in various crops has shown positive effects in aspects such as germination [18], grounding [19], blossoming [20], fruit set [21] and the accumulation of secondary metabolites, as antioxidants [22,23]. In addition, the use of growth regulators can offer advantages in terms of agricultural sustainability, being able to reduce the excessive use of fertilizers, promoting good agricultural practices [24]. Likewise, its selective and dosed application can optimize the use of resources such as water and nutrients, contributing to production efficiency in cultivation [25]. However, rigorous scientific research is essential to fully understand the effects of plant growth regulators on different crops and environmental conditions. This includes evaluating their optimal concentrations, mechanisms of action, interactions with other environmental factors and possible adverse effects. The effect of IAA on crops can vary depending on the concentrations and can have different effects on parameters of growth, development and physiological response of plants.
In this sense, the present study focuses on studying the biostimulant effect of the exogenous application of different concentrations of IAA in the onion (Allium cepa) crop as a strategy to improve productivity. According to the above-mentioned facts, Allium cepa was selected due to its economic and agricultural importance in many countries of the world. Due to the advantages described for IAA, this research was designed to examine the effect of different concentrations of IAA in growth parameters, photosynthetic aspects and phenolic compounds and antioxidants contents in Allium cepa. The main aim of the research was to determine the optimal IAA concentrations that maximize these parameters without causing negative effects on the crop.
The results of this research will contribute to broaden the knowledge about the use of IAA as a biostimulant in Allium cepa and could have practical applications in agriculture to improve agricultural productivity and sustainability.
2. Materials and Methods
2.1. Greenhouse Cultivation
The experiment was carried out in the experimental field of Universidad Autònoma de Barcelona, – (Cerdanyola del Valles, Barcelona – Spain), the geographic coordinates are: latitude 41° 29’ 53.97’’ N; longitude 2° 6’ 8.30" E. The plants were grown in pots and brought into a greenhouse on September 23, 2022.
2.2. Substrate for Cultivation
The seedlings of onion (Allium cepa) were of the Hamaemi variety, obtained from the Planter Faura nursery (Castellbisbal, Barcelona, Spain), one month old. They were transferred into pots filled with a mixture of peat, sand and sterilized vermiculite (1:1:1, v/v/v) [27].
2.3. Preparation of Indole-3-Acetic Acid Solution
A 5000 ppm stock solution of IAA was prepared, from the commercial product Sigma Aldrich, product reference 1.00353 of CAS 87-51-4, with a concentration ≥ 99% and then this solution was diluted in different concentrations according to the proposed experimental design, as explained later. This solution was preserved in a dark container and stored in a cold room at 4°C.
2.4. Experimental Design
The experimental design was a Completely Random Design (CRD) with several treatments and four replications. Six treatments were implemented: without application of IAA - IA0 (control treatment) and 50, 100, 150, 200, 250 ppm of IAA [28,29], named as IA50, IA100, IA150, IA200 and IA250, respectively. All treatments were replicated 4 times for a total of 24 pots (6 concentrations of IAA biostimulant (D) for 4 replicates).
The IAA solution was sprayed on the onion leaves. The application began in the transplant by root immersion, then at 20, 45, 60, 70 and 80 days respectively, from September 23, 2022.
2.5. Agronomic Characteristics
Crop evapotranspiration (ETc) (mm/day) was determined using the FAO Penman-Monteith equation (Eq. 1);
ETc= ETo x Kc
Where Evapotranspiration (ETo) (mm/day) with limited data, and the onion crop coefficient (Kc) was calculated applying scheduled drip irrigation throughout the vegetative period.
Crop management was carried out in accordance with the guidelines of good agricultural practices for onion cultivation. Accordingly, NPK fertilizers equivalent to 140, 60 and 200 kg/ha were applied to all treatments, in accordance with the practical guide of the rational fertilization of crops in Spain and Real Decreto 47/2022 on water protection against diffuse pollution produced by nitrates from agricultural sources [30,31].
During the phenological cycle of the onion crop, chemical protection with cupric fungicide was carried out for preventive control in all treatments. Onions were harvested on March 9, 2023 (all the treatments were harvested on the same day). Immediately the fresh biomass (root, bulb, aerial biomass) and the roots were washed with distilled water and weighed the same day. The collected bulbs with the aerial part were stored in a cold room at 4°C with a ventilation system for their subsequent analysis of chemical parameters and dry material.
2.6. Greenhouse Climatic Variables
Figure 1 reports the climatic variables of the experimental site during the growth period of the onion crop. The maximum temperatures oscillated between 32.5 °C registered in September and the average minimum temperature oscillated between 6.8 °C registered in the month of January.
2.7. Determination of Growing Parameters
At harvest, the onion plants were extracted from all pots, then the total fresh biomass of the plant (FW), the root fresh weight (FR), the fresh weight of the leaves (FF) and the bulb fresh weight (FB) were weighed from all repetitions. The dry weight of the roots, leaves and bulb were determined separately, from three representative samples from each replicate, dried in an oven at 105 °C until constant weight to determine the percentage of dry matter (DM). Besides, the height of the plant and the central diameter of the bulb were also measured.
2.8. Chlorophyll, Carotenoids and Nitrates
Chlorophyll a and b, and the content of carotenoids were tested in triplicate for each plant from samples of fresh leaves. Extraction was carried out with 96% ethanol, according to the method of Lichtenhaler and Wellburn [32]; and the absorbance of the solution was measured with a spectrophotometer (HACH LANGE DR 3900) at 649, 665 and 470 nm, respectively. The total chlorophylls were calculated by the sum of chlorophylls and b.
Nitrates were tested in all the plants, using Cataldo method for the extraction [33], Then, the DR test kits for nitrate of 22-155 ppm were used. According to the manufacturer’s methodology, 100 μL of the extracted sample were taken after passing through a 0.22 µm filter and then measured using a spectrophotometer (HACH LANGE DR 3900). The concentration was subsequently calculated using the fresh weight (FW).
2.9. Total phenolic Content and Antioxidant Activity (DPPH)
The total phenolic content (TP) was determined by the Folin-Ciocalteu method [34], using gallic acid as standard equivalent. The absorbance of the blue-coloured reaction solution was read at a wavelength of 750 nm with a spectrophotometer HACH LANGE DR 3900. Extracts were obtained from the onion bulbs and from each repetition, they were measured in triplicate. The results were expressed in milligrams of Gallic Acid (AGE) equivalent per gram of bulb dry matter (DM) (mg AGE g-1 of dry weight).
A slightly modified DPPH (2,2-diphenyl-1-picrylhydrazil) free radical scavenging assay was applied to measure the antioxidant activity of methanol extracts in the onion crop [35]. An 80% methanol solution with DPPH was prepared, poured into flasks covered with metal paper, where 100 μL of the extract was used in triplicate for each repetition. 2900 μL of the DPPH solution was added, shaken for 1 minute, then left for 30 minutes at room temperature and them the absorbance of each sample was read at 517 nm using a spectrophotometer (HACH LANGE DR 3900). The results were expressed in micromoles of Trolox (TE) equivalent per gram DM of the bulb (μmol TE g-1 of dry weight).
2.10. Statistical Analysis
The average of the triplicate data was determined, then subjected to an analysis of variance (ANOVA) using the MINITAB 19 software package, InfoStat 2020, and the differences between the medians were evaluated using a post hoc LSD test (p ≤ 0.05). A principal component analysis (PCA) was also performed on the physical and chemical parameters with the effect of IAA concentrations. Hierarchical cluster analysis (HCA) analysis was performed based on physical and chemical variables and IAA concentrations. All data are presented as median ± standard error (n = 4).
3. Results
3.1. Effect of the Concentrations of IAA on the Onion Growth Physical Parameters
For the data analysis, the physical parameters were determined, corresponding to the total fresh weight per plant, fresh weight of the root per plant, bulb per plant, plant height, diameter and height of the bulb. This evaluation was obtained on the day of the harvest except for the fresh weight of the bulb that was kept in cold room until the evaluation of the chemical parameters.
The weight of the total fresh biomass (FW), which included root, bulb and leaves, did not present statistical significance between treatments concentrations of IAA (Table 1). However, according to the LSD ≤ 0.05 post hoc test, there was a statistical difference between the medians, with the AI50 dose statistically being the most effective in this variable, while the treatment with AI200 does not seem to be effective. The treatments with IA100, IA150 and IA250 are also effective but do not differ significantly from the control (Figure 1A). Therefore, the IA50 dose was the most effective in increasing the total fresh weight of the onion crop.
The fresh weight of the root (FR) did not present statistical significance between treatments, according to the LSD post hoc test (p ≤ 0.05) (Table 1). In all the treatments evaluated in relation to the fresh weight of the root, they do not present statistically significant differences among themselves. These results may be indicative that the exogenous application of IAA in the concentrations used did not significantly influence the development and growth of the root system of onion plants.
Based on the results of the FF, no significant differences were found between the treatments in relation to this parameter. The results according to the post hoc LSD test (p ≤ 0.05) indicate that the exogenous application in different IAA concentrations did not have a statistically significant effect on the FF of the onion plants.
Based on the results for the FB, it is evident that there are statistically significant differences (according to the LSD post hoc test (p ≤ 0.05) between treatment with AI50, which presented the highest FB per plant, and the rest of the treatments and control. These results could suggest that the application of IA50 can have a beneficial effect on the growth and development of the onion crop, by increasing the FB. On the contrary, the negative effect is also evident in high IAA concentrations.
In relation to the results of the PH, they show that there was no significant difference in the PH between the treatments with different concentrations of IAA (Table 2). However, in the post hoc LSD test (p ≤ 0.05), it was observed that the concentration of IA150 turned out to have a higher PH. On the other hand, the IA200 concentration resulted in a significantly lower PH compared to the IA150 concentration. In the same way, the BH and CDB did not present significant differences.
In relation to root dry weight (RDW), (Table 3) there is no significant difference in the effects of IAA concentrations. However, it is observed that there is a significant difference according to the LSD post hoc test (p ≤ 0.05). The results show that the IA200 and IA250 treatments had a significantly higher root dry weight compared to the other treatments. On the other hand, the control treatment IA0 presented a significantly lower root dry weight compared to the other treatments. The IA50, IA100 and IA150 treatments did not show significant differences between them.
Regarding the bulb dry weight (BDW) and leaf dry weight (FDW) relationship, no statistical significance was found in the effects of IAA concentrations in BDW and FDW as in the LSD post hoc test (p≤0.05) where all treatments do not present a significant difference in relation to the medians. These results suggest that different concentrations of IAA did not have a significant effect on BDW and FDW.
Summarizing, IAA concentration showed significant effect on FB where AI50 produced the highest value, while median values for FW, PH and RDW were also found statistically different in the case of AI50, AI 150 and AI200 and AI250 respectively (higher values).
3.2. Effect of the Concentrations of IAA on Chlorophyll, Carotenoids and Nitrates
Chlorophyll a and b, carotenoids and total pigments did not present statistical significance between treatments, likewise, according to the post hoc LSD test (p ≤ 0.05), no significant differences were found between the medians of the treatments.
The amount of nitrates present no statistical significance; however, the post hoc LSD test (p ≤ 0.05) demonstrated that the concentration of IA100 had a significantly higher median compared to the concentrations of IA50, IA250 and the IA0 control, which in turn had a significantly lower median than the other concentrations (Table 4).
3.3. Effect of the Concentrations of IAA on Phenolic Content (DPPH)
The results show that there was a significant difference in the levels of total phenolic content among the treatments with different concentrations of IAA. The IA200 treatment had the highest number of total phenols, followed by IAA150 and IAA250 (Table 5), while the IA100 treatment had an intermediate amount of total phenols. Treatments with IA0 and IA50 presented the lower levels of total phenols.
The results indicate that there was no statistically significant difference between the treatments in terms of their antioxidant capacity as measured by DPPH. However, by applying Duncan’s test with 90% confidence, it was observed that IA200 treatment had the highest antioxidant capacity, closely followed by IA250 treatment, while IA150 treatment presented the lowest antioxidant capacity.
3.4. Classification Analysis
In this section chemical and physiscal parameters are studied separatelly by means of a principal component analysis and afterwards in a combined using the Hierarchical Cluster Analysis.
3.4.1. Principal Component Analysis (PCA) for Chemical Parameters
In the first interaction, when the principal component analysis (PCA) was performed on the evaluated variables, a 2-component model was obtained considering 66.3% of total variance. The principal component 1 (PC1) explains 53.8% of the data variability, while component 2 (CP2) explains 19.3%, according to Figure 2a. The variables chlorophyll a, b, carotenoids, total pigments, and DPPH are in the positive quadrant of CP1, which indicates that they are strongly related. In addition, in Figure 2b, the IA200 and IA100 concentrations are close to these points, which may suggest that these concentrations are related to higher levels of chlorophylls, total pigments and carotenoids. In the same main component analysis (PC1), there are also total phenols, nitrates and antioxidant capacity measured by DPPH. These factors are slightly close to each other, indicating a relationship between them. The concentrations of IA100 and IA250 are also close to these points, which suggests that these concentrations could influence the levels of total phenols, nitrates and antioxidant capacity. Regarding PC2, the chemical parameters are observed as a separation into two groups of variables. In the positive quadrant, there are total phenols, DPPH and nitrates corresponding to the concentrations of IA100 and IA200. On the other hand, in the negative quadrant, there are chlorophyll a, chlorophyll b, total pigments and carotenoids. This separation in PC2 suggests an inverse relationship or contrast between the metabolites and characteristics associated with total phenols, DPPH, nitrates, and those associated with chlorophylls, carotenoids and total pigments. In other words, higher concentrations of IA100 and IA200 appear to have a positive effect on total phenols, DPPH and nitrates, while they have a negative effect on chlorophylls, carotenoids and nitrates.
3.4.2. Principal Component Analysis (PCA) for Physical Parameters
The principal component analysis (PCA) was performed on the evaluated physical variables, obtaining a 2-component model considering 65.3% of total variance. The principal component analysis 1 (PCA1) explains 51.2% of the data variability, while component 2 (PCA2) explains 14.1%, according to Figure 3a. The physical variables (fresh waste weight, root fresh weight, fresh weight leaves, bulb fresh weight, plant height, bulb height, bulb centre diameter, roots dry weight, bulb dry weight, leaves dry weight) are found in the positive quadrant of CP1, being the parameters strongly related. In addition, the IA50 and IA150 concentrations are close to these points, which can suggest that these values of physical parameters are related to higher levels.
On the other hand, most of the variables of the physical parameters are found in the same PCA1, which indicates that these variables are related to each other and have a lower contribution to the variability of the data. Also, IA150 and IA50 in Figure 3b, concentrations are close to these points, suggesting that these concentrations are related to root dry weight, bulb height, bulb dry weight, bulb centre diameter and leaves fresh weight. The concentrations of IA0, IA100, IA200 and IA250 are distributed on the negative side of the positive quadrant of PC1.
Regarding PC2, the variables root dry weight, bulb height, bulb dry weight, bulb centre diameter, leaves dry weight are in the positive quadrant and in the negative quadrant bulb fresh weight, fresh waste weight, fresh weight leaves, root fresh weight and plant height) are found.
PC2 explains 14.1% of the total variability of the data, which suggests that there is a moderate correlation between the variables in this component.
3.4.3. Combined Physical and Chemical Analysis
The Hierarchical Cluster Analysis (HCA) analysis was carried out on the entire data set, which corresponds to a tree diagram where the objects are grouped in rows by their similarities based on the physical variables and the IAA concentrations according to Figure 4a, which suggests that there are two groups of concentrations. They differ on the basis of their physical parameters. The first group is made up of concentrations IA50 and IA150 because they have similar growth in physical parameters, and the second group is made up of concentrations IA200, IA250, IA100 and IA0.
In the cluster analysis presented in Figure 4b, which is based on the chemical parameters and the biostimulant concentrations, it is suggested that there are two groups that differ on the basis of their chemical parameters. The first group is made up of concentrations IA50, IA100 and IA200 because they have similar levels in chemical parameters, and the second group is made up of concentrations IA250, IA150 and IA00.
In the following results, the dendrogram has grouped the treatments into two large groups (physical and chemical parameters) according to Figure 5. The first group is composed only of the treatments with concentrations IA50 and IA150, while the second group is composed of treatments with concentrations of IA0, IA100, IA200 and IA250. This result may suggest that the concentration of IA50 and IA150 may have a different effect on the physical and chemical parameters compared to the other concentrations. On the other hand, treatments with IA0, IA100, IA200 and IA250 concentrations of IAA biostimulant seem to have a similar response in terms of the physical-chemical parameters evaluated.
4. Discussion
In this study, the effect of the exogenous application of six concentrations of IAA (IAO, IA50, IA100, IA150, IA200 and IA250) as biostimulant in onion grown under greenhouse conditions were evaluated.
In terms of total fresh weight, which represents the total fresh biomass of the crop (root, bulb, leaves), this result can have interesting economic benefits, considering that more biomass is produced. The results from this study are consistent with previous studies that have demonstrated the beneficial effects of IAA on plant growth and development. Several studies have reported that IAA application can promote root growth, increase biomass accumulation and improve nutrient uptake in different crops [36,37,38]. Regarding the optimal concentration of IAA, the results of this study support the trend observed in previous research that suggests that low or moderate concentrations of IAA are more effective than higher concentrations For instance, a study conducted in maize cultivation found that the concentrations of 50 and 100 ppm of IAA had a positive effect on the yield compared to the application of 200 and 300 ppm of IAA[39]. Furthermore, our results from this study indicate that a concentration of 200 ppm of IAA may not be effective and could even have negative effects on onion cultivation. These findings are supported by previous research that has reported inhibitory or toxic effects of IAA at high concentrations [40].
The results obtained in this study agree with previous investigations that have demonstrated the significant effect of IAA on BF. Several studies have reported that the application of IAA can promote onion bulb development and increase its weight in different horticultural crops [29,41]. These results indicate that the IA50 concentration had a significant positive effect on the fresh weight of the onion bulb, being greater in comparison with the other evaluated treatments and the control. These findings are consistent with previous research that has found that low concentrations of IAA in Guizotia Abyssinica [42] can enhance growth and development in different crops [43,44]. Regarding the root dry weight (RDW), the IA200 and IA250 treatments presented better results compared to the other treatments and the control. However, it is important to note that other studies have shown that specific IAA concentrations can influence root dry weight in this crop [45]. These findings suggest that the response of root dry weight to IAA may be variable depending on the experimental conditions and the specific characteristics of each crop. Regarding the other physical parameters evaluated in this research (FR, FF, BH, CDB, BDW and FDW), they did not show significant differences between the treatments and differences in the LSD post hoc test (p ≤ 0.05). These findings are consistent with previous research that has shown that IAA concentrations do not have a significant effect on certain physical parameters in crops [46]. However, it is important to note that the scientific literature has reported some positive effects of IAA on leaf and root fresh weight in other crops, suggesting that the effects may be species-specific [47,48].
No statistical significance between treatments was observed analysing the results obtained for chlorophyll a, chlorophyll b, carotenoids and total pigments. Several previous studies have also evaluated the effect of IAA on photosynthetic pigments in different crops. Some reported results are similar to these obtained in this study, since days after the application of the IAA, the effect of these pigments begins to decrease, not having a significant impact on the levels of chlorophyll and carotenoids. For example, a study conducted by [49] on Cinnamomum camphora plants found no significant differences in the chlorophyll and carotenoid contents 60 days after IAA application. Another study carried out by [50] in Lolium perenne L. plants, showed no significant differences related to the treatment in the proportion of chlorophyll at 60 days of evaluation. These findings may suggest that the effect of the IAA concentrations used in our study at the end of the harvest may not have a direct effect on the degradation of photosynthetic pigments in the onion crop. However, it is important to highlight that the pigment content can be influenced by several factors, such as the interaction with environmental conditions, the genotype of the plant and the stage of growth. Therefore, the lack of significant effect of IAA in our results may be due to these variations and the specific sensitivity of onion to this growth regulator.
Regarding the effect of the exogenous application of IAA on the accumulation of nitrates in the onion crop, they show that there were no significant differences for nitrates. The presence of higher quantity of nitrates in the IA100 treatment may suggest that this concentration promoted the accumulation of nitrates in the onion. However, it has also been reported that IAA application can improve root architecture and nitrogen metabolism in apple trees [51]. Some studies also indicate that the presence of IAA improved the content of nitrates in wheat sprouts [52]. However, it is important to consider several explanations for these results. The response to IAA may be specific to each plant species. Different plants may have different regulatory mechanisms for nitrate accumulation and may respond differently to IAA [38]. It is possible that the onion crop is not as sensitive to IAA in terms of nitrate accumulation, which would explain the lack of significant differences observed in the present study. Furthermore, the accumulation of nitrates in plants is a complex process that is influenced by multiple factors [53,54], such as the availability of nitrates in the soil [55], the activity of enzymes related to their metabolism [56] and the response of other phytohormones [57]. These factors can interact in a complex way with IAA and nitrate signalling [58].
Our results agree with previous studies that have investigated the IAA effect on total phenol levels in onion crops. Several studies have shown that the exogenous application of IAA can positively influence the accumulation of total phenols in plants [59]. Supporting these findings, several studies have reported the role of IAA in the production of total phenols in different plant species, observing an increase in the levels of total phenols in response to the application of IAA, which is consistent with our results [60]. Furthermore, it has been reported that IAA can induce phenol synthesis in plants by acting as a regulator of secondary metabolism [61]. Furthermore, the presence of total phenols in plants is of particular interest due to their antioxidant properties and health benefits [62]. These bioactive compounds have been associated with antimicrobial, anti-inflammatory and anticancer activities, among others [63]. Therefore, the increase in the levels of total phenols in the onion crop in response to the concentrations of IAA used in our study could have positive implications from the nutritional and health point of view. However, it is important to highlight that the effect of the exogenous application of IAA may vary according to the plant species and the specific environmental conditions. Therefore, further investigations are required to better understand the underlying mechanisms involved in the regulation of total phenols in response to IAA.
The results obtained in relation to the antioxidant capacity measured by DPPH indicate that there was no statistically significant difference between the treatments in general. These results are confirmed by other studies, such as the effect of IAA on the contents and metabolites in Hibiscus sabdariffa plants, showing no considerable difference between concentrations with IAA [64]. In our case, it is suggested that the concentration of IAA may have an impact on the antioxidant capacity of onion. Treatment with IA200 showed higher antioxidant activity, which could indicate its potential to protect plant cells against oxidative stress [65]. This antioxidant capacity may be due to the presence of phenolic compounds in IAA, which are known for their antioxidant activity [60]. Importantly, antioxidant capacity measured by DPPH is only one of many ways to assess the antioxidant potential of a compound or extract [66]. Other experiments, such as free radical scavenging capacity or lipid peroxidation inhibitory activity, can provide a more comprehensive view of the antioxidant profile [67]. In the scientific literature, studies on the antioxidant activity of phenolic compounds have been reported with the presence of different biostimulants or plant extracts [68]. These studies have demonstrated the ability of phenols to neutralize free radicals and protect plants against oxidative stress [69,70]. This can be beneficial in formulating biostimulants that contain phenols that can stimulate plants against external limiting factors [71]. The results indicate that the IA200 treatment had the highest antioxidant capacity as measured by DPPH, closely followed by the IA250 treatment. These findings support the idea that IAA may contribute to the antioxidant activity in onion cultivation, although more research is necessary to fully understand the mechanisms of action and effects of antioxidant capacity in relation to the concentration of IAA and other components that are present in the biostimulant.
On the other hand, according to the PCA, these results could be interesting from the physiological and biochemical point of view of the onion crop. For example, an increase in total phenols and antioxidant capacity measured by DPPH could indicate a response to plant defense mechanisms under stress conditions [72,73] or as a form of protection against oxidative damage [74]. Also, a decrease in chlorophylls and carotenoids could be related to changes in photosynthesis or the regulation of pigment synthesis in the presence of higher IAA concentrations. This study reveals a separation between the metabolites and characteristics related to total phenols, DPPH and nitrates, and those related to chlorophylls, carotenoids and nitrates in response to IAA concentrations. These findings could provide additional information on onion responses to IAA and may be useful for future research.
The PCA results demonstrate a correlation between the variables of the physical components evaluated and the IAA concentrations in the onion crop. This is supported by previous studies that have shown the influence of IAA concentrations on plant growth and development. Regarding the physical components, such as the fresh weight of the bulb, fresh weight of the leaves, fresh weight of the root and total fresh weight and height of the plant, it has been observed that the application of IAA can promote greater development and growth of these parameters in different crops. Studies have shown that IAA can stimulate root growth and the development of the aerial part of plants, which is reflected in an increase in the fresh weight of the distinct parts of the plant [75].
Regarding the analysis of conglomerates based on the chemical parameters and the different concentrations of IAA, it reveals the existence of two distinct groups depending on their chemical characteristics. In the first group, made up of the IA50, IA100 and IA200 concentrations, a similarity in the chemical parameters evaluated, including chlorophylls a, b, carotenoids, total pigments, nitrates, phenols, and antioxidant capacity measured by DPPH, was found. These results suggest that these IAA concentrations may have a similar effect on the chemical composition of onion, which may influence its nutritional quality and antioxidant properties. The second group is made up of concentrations IA250, IA150 and IA00. These IAA concentrations show different levels in the chemical parameters evaluated, indicating a divergence in the chemical response of the onion to these IAA concentrations. These results are consistent with those that have shown that IAA concentrations can modulate the synthesis and accumulation of chemical compounds in plants, such as photosynthetic pigments, phenolic compounds, and antioxidants by DPPH, reported in this study. These findings support the importance of selecting the proper IAA concentrations to influence the chemical composition of onion and potentially improve its nutritional quality and antioxidant properties.
The cluster analysis based on the physical parameters and the different concentrations of IAA also reveals the existence of two distinct groups. In the first group, made up of the IA50 and IA150 concentrations, a similarity is observed in the growth and development of the onion in terms of fresh weight of the bulb, fresh weight of the leaves, fresh weight of the root, height of the plant, among other physical parameters. These results suggest that these IAA concentrations may have a similar effect on onion vegetative development. The second group is made up of concentrations IA200, IA250, IA100 and IA0.. It is important to highlight that this group differs from the first group in terms of the physical parameters evaluated, which suggests that these IAA concentrations may have a different impact on onion vegetative development. These findings support the importance of selecting the appropriate concentration of IAA to influence the vegetative development of onion.
The cluster analysis based on the physical and chemical parameters reveals the formation of two distinct groups depending on the IAA concentrations evaluated. In the first group, composed solely of the treatment with concentrations of IA50 and IA150, a similarity is observed in the physical and chemical parameters analysed. This suggests that these IAA concentrations may have a similar effect on onion development at both a physical and chemical level. On the other hand, the second group is made up of the treatments with concentrations of IA0, IA100, IA200 and IA250. These concentrations show differences in both physical and chemical parameters, indicating a divergent response in onion growth and development compared to the first group. These results suggest that the IAA concentrations used in this study may have a different impact on the physical and chemical aspects of the onion. These findings are consistent with previous studies that have shown that concentrations of plant hormones, such as IAA, can modulate both physical and chemical parameters in plants [37]. The exogenous application of IAA can influence the morphology and other chemical parameters of the plants [20,76]. The cluster analysis shows the formation of two distinct groups depending on the concentrations of IAA and its impact on the physical and chemical parameters of the onion [29]. These results support the importance of selecting the appropriate IAA concentration to influence onion growth and development, as well as its chemical content.
5. Conclusions
In this study, the effect of the exogenous application of IAA as a biostimulant in the onion crop was evaluated. The use of different concentrations of IAA had a significant impact on several physical and chemical parameters of the onion. Statistically significant differences were observed in the fresh weight of the bulb, fresh weight of the leaves, height of the plant, dry weight of the root and total phenol content in response to the IAA concentrations evaluated. Cluster analysis revealed the formation of distinct groups based on IAA concentrations. In the case of physical parameters, two groups were identified, with IA50 and IA150 concentrations being the most similar to each other. Regarding the chemical parameters, two groups were formed, where the concentrations of IA50, IA100 and IA200 shared similar characteristics. The results showed that the AI50 concentration was statistically the most effective on the fresh weight of the bulb, while the AI200 concentration did not seem to have a significant effect on this variable. In the case of total phenols, IA200 concentration presented the highest levels, closely followed by IA150 and IA250 concentrations. Regarding nitrates, no significant differences were found between the IAA concentrations evaluated, although the IA100 concentration showed a slightly higher median compared to other concentrations. The conclusions of this study indicate that the exogenous application of different concentrations of IAA can influence the growth and development of the onion, as well as its chemical composition. However, it is important to highlight that the onion response to IAA concentrations may vary depending on the parameters evaluated. These findings provide a basis for optimizing IAA application strategies as a biostimulant in onion crops, seeking concentrations that maximize the desired parameters without causing negative effects on the crop. Further research is needed to better understand the underlying mechanisms and the dose-response relationship of IAA in different varieties of the onion crop, with the aim of harnessing its potential as a biostimulant in agriculture.
Author Contributions
“Conceptualization, formal analysis, R.C.S.P. and AS; methodology, A.A.; validation, A.A., R.B. and C.B.M.; R.C.S.P.; investigation, R.B.; resources, C.B.M.; data curation, A.S; writing—original draft preparation, A.S; writing—review and editing, R.C.S.P; visualization, AA.; supervision, A.A, A.S; project administration, A.S. All the authors have read and agreed to the published version of the manuscript.
Funding
R.C.S.P received a grant from the National Program of Scholarships and Educational Credit – PRONABEC – Perú, resolution 2512-2021-MINEDU-VMG-PRONABEC. This research was financially supported by the Spanish Ministry of Science and Innovation in the call R+D+i projects in strategic lines 2022. Project FertiLab, (reference PLEC2022-009252).
Data Availability Statement
Presented data in this study are available on request from the corresponding author.
Acknowledgments
The secretariat of the environmental biological chemical engineering department is recognized for the administrative support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sharma, K.; Mahato, N.; Nile, S.H.; Lee, E.T.; Lee, Y.R. Economical and Environmentally-Friendly Approaches for Usage of Onion (Allium Cepa L.) Waste. Food Funct. 2016, 7, 3354–3369. [Google Scholar] [CrossRef] [PubMed]
- Marcinkowska, M.A.; Jeleń, H.H. Role of Sulfur Compounds in Vegetable and Mushroom Aroma. Molecules 2022, 27, 6116. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.; Caruso, G. Chapter 5 - Onion. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press, 2020; pp. 73–87. ISBN 978-0-12-812780-3.
- Murtado, A.; Mubarik, N.R.; Tjahjoleksono, A. Isolation and Characterization Endophytic Bacteria as Biological Control of Fungus Colletotrichum Sp. on Onion Plants (Allium Cepa L.). IOP Conf. Ser.: Earth Environ. Sci. 2020, 457, 012043. [Google Scholar] [CrossRef]
- Nepomuceno, R.A.; Brown, C.M.B.; Mojica, P.N.; Brown, M.B. Biological Control Potential of Vesicular Arbuscular Mycorrhizal Root Inoculant (VAMRI) and Associated Phosphate Solubilizing Bacteria, Pseudochrobactrum Asaccharolyticum against Soilborne Phytopathogens of Onion (Allium Cepa L. Var. Red Creole). Archives of Phytopathology and Plant Protection 2019, 52, 714–732. [Google Scholar] [CrossRef]
- Dutta, R.; K., J.; Nadig, S.M.; Manjunathagowda, D.C.; Gurav, V.S.; Singh, M. Anthracnose of Onion (Allium Cepa L.): A Twister Disease. Pathogens 2022, 11, 884. [CrossRef] [PubMed]
- Kalman, B.; Abraham, D.; Graph, S.; Perl-Treves, R.; Meller Harel, Y.; Degani, O. Isolation and Identification of Fusarium Spp., the Causal Agents of Onion (Allium Cepa) Basal Rot in Northeastern Israel. Biology 2020, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Ratnarajah, V.R.; Gnanachelvam, N.G. Effect of Abiotic Stress on Onion Yield: A Review. 2021. [Google Scholar] [CrossRef]
- Khar, A.; Singh, H.; Verma, P. Mitigating Abiotic Stresses in Allium Under Changing Climatic Scenario. In Genomic Designing for Abiotic Stress Resistant Vegetable Crops; Kole, C., Ed.; Springer International Publishing: Cham, 2022; pp. 253–278. ISBN 978-3-031-03964-5. [Google Scholar]
- Carvalho, F.P. Agriculture, Pesticides, Food Security and Food Safety. Environmental Science & Policy 2006, 9, 685–692. [Google Scholar] [CrossRef]
- Hewett, E.W. An Overview of Preharvest Factors Influencing Postharvest Quality of Horticultural Products. International Journal of Postharvest Technology and Innovation 2006, 1, 4–15. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Scientia Horticulturae 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial Plant Biostimulants: A Sustainable Way towards Improving Growth, Productivity, and Health of Crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-Acetic Acid: A Widespread Physiological Code in Interactions of Fungi with Other Organisms. Plant Signaling & Behavior 2015, 10, e1048052. [Google Scholar] [CrossRef]
- Gray, W.M. Hormonal Regulation of Plant Growth and Development. PLOS Biology 2004, 2, e311. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in Action: Signalling, Transport and the Control of Plant Growth and Development. Nat Rev Mol Cell Biol 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Hagaggi, N.Sh.A.; Mohamed, A.A.A. Enhancement of Zea Mays (L.) Growth Performance Using Indole Acetic Acid Producing Endophyte Mixta Theicola Isolated from Solenostemma Argel (Hayne). South African Journal of Botany 2020, 134, 64–71. [Google Scholar] [CrossRef]
- Zhang, Y.; Paschold, A.; Marcon, C.; Liu, S.; Tai, H.; Nestler, J.; Yeh, C.-T.; Opitz, N.; Lanz, C.; Schnable, P.S.; et al. The Aux/IAA Gene Rum1 Involved in Seminal and Lateral Root Formation Controls Vascular Patterning in Maize (Zea Mays L.) Primary Roots. Journal of Experimental Botany 2014, 65, 4919–4930. [Google Scholar] [CrossRef]
- Bommineni, V.R.; Greyson, R.I. Effect of Gibberellic Acid and Indole-3-Acetic Acid on Growth and Differentiation of Cultured Ear Inflorescences of Maize (Zea Mays L.). Plant Science 1990, 68, 239–247. [Google Scholar] [CrossRef]
- Bermejo, A.; Granero, B.; Mesejo, C.; Reig, C.; Tejedo, V.; Agustí, M.; Primo-Millo, E.; Iglesias, D.J. Auxin and Gibberellin Interact in Citrus Fruit Set. J Plant Growth Regul 2018, 37, 491–501. [Google Scholar] [CrossRef]
- Masmoudi, F.; Tounsi, S.; Dunlap, C.A.; Trigui, M. Halotolerant Bacillus Spizizenii FMH45 Promoting Growth, Physiological, and Antioxidant Parameters of Tomato Plants Exposed to Salt Stress. Plant Cell Rep 2021, 40, 1199–1213. [Google Scholar] [CrossRef]
- Mir, A.R.; Siddiqui, H.; Alam, P.; Hayat, S. Foliar Spray of Auxin/IAA Modulates Photosynthesis, Elemental Composition, ROS Localization and Antioxidant Machinery to Promote Growth of Brassica Juncea. Physiol Mol Biol Plants 2020, 26, 2503–2520. [Google Scholar] [CrossRef]
- Yadav, A.N. Plant Microbiomes for Sustainable Agriculture: Current Research and Future Challenges. In Plant Microbiomes for Sustainable Agriculture; Yadav, A.N., Singh, J., Rastegari, A.A., Yadav, N., Eds.; Desarrollo Sostenible y Biodiversidad; Springer International Publishing: Cham, 2020; pp. 475–482. ISBN 978-3-030-38453-1. [Google Scholar]
- Li, Z.; Zhang, X.; Zhao, Y.; Li, Y.; Zhang, G.; Peng, Z.; Zhang, J. Enhancing Auxin Accumulation in Maize Root Tips Improves Root Growth and Dwarfs Plant Height. Plant Biotechnology Journal 2018, 16, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-W.; Lee, S.M.; Joo, S.-H.; Yun, H.S.; Lee, Y.; Kaufman, P.B.; Kirakosyan, A.; Kim, S.-H.; Nam, K.H.; Lee, J.S.; et al. Elongation and Gravitropic Responses of Arabidopsis Roots Are Regulated by Brassinolide and IAA. Plant, Cell & Environment 2007, 30, 679–689. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Abdel-Motaal, F.; El-Sayed, M.; Jogaiah, S.; Shigyo, M.; Ito, S. ichi; Tran, L.S.P. Dissection of Trichoderma Longibrachiatum-Induced Defense in Onion (Allium Cepa L.) against Fusarium Oxysporum f. Sp. Cepa by Target Metabolite Profiling. Plant Science 2016, 246, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Jyoti, D.; Rupinder, S.; Ishita, W. Effect of Foliar Application of GA3 and NAA on Onion-a Review. Plant Archives 2018, 18, 1209–1214. [Google Scholar]
- Hye, M.; Haque, M.; Karim, M. Influence of Growth Regulators and Their Time of Application on Yield of Onion. Pakistan Journal of Biological Sciences 2002, 5. [Google Scholar] [CrossRef]
- Publicaciones fertilizantes. Available online: https://www.mapa.gob.es/es/agricultura/publicaciones/Publicaciones-fertilizantes.aspx (accessed on 9 June 2023).
- Ministerio de la Presidencia, Relaciones con las Cortes y Memoria Democrática Real Decreto 47/2022, de 18 de Enero, Sobre Protección de Las Aguas Contra La Contaminación Difusa Producida Por Los Nitratos Procedentes de Fuentes Agrarias; 2022; Vol. BOE-A-2022-860, pp. 5664–5684.
- LICHTENTHALER, H.K.; WELLBURN, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochemical Society Transactions 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Communications in Soil Science and Plant Analysis 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Cruzado, M.; Pastor, A.; Castro, N.; Cedrón, J.C. Determinación de Compuestos Fenólicos y Actividad Antioxidante de Extractos de Alcachofa (Cynara Scolymus L.). Revista de la Sociedad Química del Perú 2013, 79, 57–63. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT - Food Science and Technology 1995, 28, 25–30. [Google Scholar] [CrossRef]
- He, Y.; Zhang, T.; Sun, Y.; Wang, X.; Cao, Q.; Fang, Z.; Chang, M.; Cai, Q.; Lou, L. Exogenous IAA Alleviates Arsenic Toxicity to Rice and Reduces Arsenic Accumulation in Rice Grains. J Plant Growth Regul 2022, 41, 734–741. [Google Scholar] [CrossRef]
- Romanov; , G. A.; Aksenova, N.P.; Konstantinova, T.N.; Golyanovskaya, S.A.; Kossmann, J.; Willmitzer, L. Effect of Indole-3-Acetic Acid and Kinetin on Tuberisation Parameters of Different Cultivars and Transgenic Lines of Potato in Vitro. Plant Growth Regulation 2000, 32, 245–251. [Google Scholar] [CrossRef]
- Hu, Q.-Q.; Shu, J.-Q.; Li, W.-M.; Wang, G.-Z. Role of Auxin and Nitrate Signaling in the Development of Root System Architecture. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Darra, B.L.; Saxena, S.N. Role of IAA on the Mineral Composition of Maize (Zea Mays) Crop under Various Osmotic Stressed Conditions. Plant Soil 1973, 38, 657–661. [Google Scholar] [CrossRef]
- Serrani, J.C.; Carrera, E.; Ruiz-Rivero, O.; Gallego-Giraldo, L.; Peres, L.E.P.; García-Martínez, J.L. Inhibition of Auxin Transport from the Ovary or from the Apical Shoot Induces Parthenocarpic Fruit-Set in Tomato Mediated by Gibberellins. Plant Physiology 2010, 153, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Pflanzen, G.; Alam, M.; Khan, M.; Khan, A.; Imtiaz, M.; Khan, A.; Naeem, M.; Haq, S.; Asim Shah Bacha, S. ; Samiullah, ·; et al. Indole-3-Acetic Acid Rescues Plant Growth and Yield of Salinity Stressed Tomato (Lycopersicon Esculentum L.). 2019. [Google Scholar] [CrossRef]
- Talukdar, M.; Swain, D.K.; Bhadoria, P.B.S. Effect of IAA and BAP Application in Varying Concentration on Seed Yield and Oil Quality of Guizotia Abyssinica (L.f.) Cass. Annals of Agricultural Sciences 2022, 67, 15–23. [Google Scholar] [CrossRef]
- Gupta, S.; Stirk, W.A.; Plačková, L.; Kulkarni, M.G.; Doležal, K.; Van Staden, J. Interactive Effects of Plant Growth-Promoting Rhizobacteria and a Seaweed Extract on the Growth and Physiology of Allium Cepa L. (Onion). Journal of Plant Physiology 2021, 262, 153437. [Google Scholar] [CrossRef]
- Kondhare, K.R.; Patil, A.B.; Giri, A.P. Auxin: An Emerging Regulator of Tuber and Storage Root Development. Plant Science 2021, 306, 110854. [Google Scholar] [CrossRef]
- Ram, M.; Khan, M.A.; Jha, P.; Khan, S.; Kiran, U.; Ahmad, M.M.; Javed, S.; Abdin, M.Z. HMG-CoA Reductase Limits Artemisinin Biosynthesis and Accumulation in Artemisia Annua L. Plants. Acta Physiol Plant 2010, 32, 859–866. [Google Scholar] [CrossRef]
- Lobo, L.L.B.; de Andrade da Silva, M.S.R.; Castellane, T.C.L.; Carvalho, R.F.; Rigobelo, E.C. Effect of Indole-3-Acetic Acid on Tomato Plant Growth. Microorganisms 2022, 10, 2212. [Google Scholar] [CrossRef]
- Spaepen, S.; Dobbelaere, S.; Croonenborghs, A.; Vanderleyden, J. Effects of Azospirillum Brasilense Indole-3-Acetic Acid Production on Inoculated Wheat Plants. Plant Soil 2008, 312, 15–23. [Google Scholar] [CrossRef]
- Wang, H.; Shan, X.; Wen, B.; Owens, G.; Fang, J.; Zhang, S. Effect of Indole-3-Acetic Acid on Lead Accumulation in Maize (Zea Mays L.) Seedlings and the Relevant Antioxidant Response. Environmental and Experimental Botany 2007, 61, 246–253. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, K.; Huang, G.; Chen, G.; Zhou, S.; Huang, Y.; Zhang, J.; Duan, H.; Fan, H. Effects of Exogenous 3-Indoleacetic Acid and Cadmium Stress on the Physiological and Biochemical Characteristics of Cinnamomum Camphora. Ecotoxicology and Environmental Safety 2020, 191, 109998. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhou, J.; Chen, K.; Wang, Y.; Ai, Y.; Zhang, C.; Zhou, S. Effect of Indole-3-Acetic Acid Supplementation on the Physiology of Lolium Perenne L. and Microbial Activity in Cadmium-Contaminated Soil. Environ Sci Pollut Res 2022, 29, 52483–52492. [Google Scholar] [CrossRef]
- Qi, B.; Zhang, X.; Mao, Z.; Qin, S.; Lv, D. Integration of Root Architecture, Root Nitrogen Metabolism, and Photosynthesis of ‘Hanfu’ Apple Trees under the Cross-Talk between Glucose and IAA. Horticultural Plant Journal 2022. [Google Scholar] [CrossRef]
- Garnica, M.; Houdusse, F.; Zamarreño, A.M.; Garcia-Mina, J.M. The Signal Effect of Nitrate Supply Enhances Active Forms of Cytokinins and Indole Acetic Content and Reduces Abscisic Acid in Wheat Plants Grown with Ammonium. Journal of Plant Physiology 2010, 167, 1264–1272. [Google Scholar] [CrossRef]
- Cantliffe, D.J. Nitrate Accumulation in Table Beets and Spinach as Affected by Nitrogen, Phosphorus, and Potassium Nutrition and Light Intensity1. Agronomy Journal 1973, 65, 563–565. [Google Scholar] [CrossRef]
- Blom-Zandstra, M. Nitrate Accumulation in Vegetables and Its Relationship to Quality. Annals of Applied Biology 1989, 115, 553–561. [Google Scholar] [CrossRef]
- Chen, B.-M.; Wang, Z.-H.; Li, S.-X.; Wang, G.-X.; Song, H.-X.; Wang, X.-N. Effects of Nitrate Supply on Plant Growth, Nitrate Accumulation, Metabolic Nitrate Concentration and Nitrate Reductase Activity in Three Leafy Vegetables. Plant Science 2004, 167, 635–643. [Google Scholar] [CrossRef]
- Hageman, R.H.; Flesher, D. Nitrate Reductase Activity in Corn Seedlings as Affected by Light and Nitrate Content of Nutrient Media 12. Plant Physiol 1960, 35, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.E. Influence of Abscisic Acid on Nitrate Accumulation and Nitrate Reductase Activity in Potato Tuber Slices. Plant and Cell Physiology 1981, 22, 1541–1551. [Google Scholar] [CrossRef]
- Wang, B.; Zhu, X.; Guo, X.; Qi, X.; Feng, F.; Zhang, Y.; Zhao, Q.; Han, D.; Sun, H. Nitrate Modulates Lateral Root Formation by Regulating the Auxin Response and Transport in Rice. Genes 2021, 12, 850. [Google Scholar] [CrossRef]
- Atif, M.; Perveen, S.; Parveen, A.; Mahmood, S.; Saeed, M.; Zafar, S. Thiamine and Indole-3-Acetic Acid Induced Modulations in Physiological and Biochemical Characteristics of Maize (Zea Mays L.) under Arsenic Stress. Sustainability 2022, 14, 13288. [Google Scholar] [CrossRef]
- Kecis, H.; Bagues, M.; Abdelouhab, Y.; Mekircha, F.; Gali, L.; Kadi, K.; Addad, D.; Nagaz, K.; Brahmi, F.; Kouba, Y. Different Indole-3-Acetic Acid and 6 Benzyl Amino Purine Concentrations Affect Biomass, Phenolic Profile, and Bioactivity in Mentha Rotundifolia L. Food Measure 2023. [Google Scholar] [CrossRef]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased Resistance of Drought by Trichoderma Harzianum Fungal Treatment Correlates with Increased Secondary Metabolites and Proline Content. Journal of Integrative Agriculture 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Arias, J.P.; Zapata, K.; Rojano, B.; Arias, M. Effect of Light Wavelength on Cell Growth, Content of Phenolic Compounds and Antioxidant Activity in Cell Suspension Cultures of Thevetia Peruviana. Journal of Photochemistry and Photobiology B: Biology 2016, 163, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Oueslati, S.; Ksouri, R.; Falleh, H.; Pichette, A.; Abdelly, C.; Legault, J. Phenolic Content, Antioxidant, Anti-Inflammatory and Anticancer Activities of the Edible Halophyte Suaeda Fruticosa Forssk. Food Chemistry 2012, 132, 943–947. [Google Scholar] [CrossRef]
- Mirheidari, F.; Hatami, M.; Ghorbanpour, M. Effect of Different Concentrations of IAA, GA3 and Chitosan Nano-Fiber on Physio-Morphological Characteristics and Metabolite Contents in Roselle (Hibiscus Sabdariffa L.). South African Journal of Botany 2022, 145, 323–333. [Google Scholar] [CrossRef]
- Esan, A.M.; Masisi, K.; Dada, F.A.; Olaiya, C.O. Comparative Effects of Indole Acetic Acid and Salicylic Acid on Oxidative Stress Marker and Antioxidant Potential of Okra (Abelmoschus Esculentus) Fruit under Salinity Stress. Scientia Horticulturae 2017, 216, 278–283. [Google Scholar] [CrossRef]
- Lee, S.K.; Mbwambo, Z.H.; Chung, H.; Luyengi, L.; Gamez, E.J.; Mehta, R.G.; Kinghorn, A.D.; Pezzuto, J.M. Evaluation of the Antioxidant Potential of Natural Products. Comb Chem High Throughput Screen 1998, 1, 35–46. [Google Scholar] [CrossRef]
- Subhasree, B.; Baskar, R.; Laxmi Keerthana, R.; Lijina Susan, R.; Rajasekaran, P. Evaluation of Antioxidant Potential in Selected Green Leafy Vegetables. Food Chemistry 2009, 115, 1213–1220. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Kocira, A.; Czerwińska, E.; Wójtowicz, A.; Bronowicka-Mielniczuk, U.; Koszel, M.; Findura, P. Modeling Biometric Traits, Yield and Nutritional and Antioxidant Properties of Seeds of Three Soybean Cultivars Through the Application of Biostimulant Containing Seaweed and Amino Acids. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The Role of Plant-Derived Natural Antioxidants in Reduction of Oxidative Stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef]
- Andre, C.M.; Larondelle, Y.; Evers, D. Dietary Antioxidants and Oxidative Stress from a Human and Plant Perspective: A Review. Current Nutrition & Food Science 2010, 6, 2–12. [Google Scholar] [CrossRef]
- Kisiriko, M.; Anastasiadi, M.; Terry, L.A.; Yasri, A.; Beale, M.H.; Ward, J.L. Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants. Molecules 2021, 26, 6343. [Google Scholar] [CrossRef]
- Báidez, A.G.; Gómez, P.; Del Río, J.A.; Ortuño, A. Dysfunctionality of the Xylem in Olea Europaea L. Plants Associated with the Infection Process by Verticillium Dahliae Kleb. Role of Phenolic Compounds in Plant Defense Mechanism. J. Agric. Food Chem. 2007, 55, 3373–3377. [Google Scholar] [CrossRef]
- Schauffler, G.P.; dos Anjos Verzutti Fonseca, J.; Di Piero, R.M. Defense Mechanisms Involved in the Resistance of Maize Cultivars to Bipolaris Maydis. Eur J Plant Pathol 2022, 163, 269–277. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A.; Saha, P.P.; Sengupta, D.N. Differential Antioxidative Responses of Indica Rice Cultivars to Drought Stress. Plant Growth Regul 2010, 60, 51–59. [Google Scholar] [CrossRef]
- Figueredo, E.F.; Cruz, T.A. da; Almeida, J.R. de; Batista, B.D.; Marcon, J.; Andrade, P.A.M. de; Hayashibara, C.A. de A.; Rosa, M.S.; Azevedo, J.L.; Quecine, M.C. The Key Role of Indole-3-Acetic Acid Biosynthesis by Bacillus Thuringiensis RZ2MS9 in Promoting Maize Growth Revealed by the IpdC Gene Knockout Mediated by the CRISPR-Cas9 System. Microbiological Research 2023, 266, 127218. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, Y.J.; Morgan, A.M.A.; Valan Arasu, M.; Al-Dhabi, N.A.; Park, S.U. Influence of Indole-3-Acetic Acid and Gibberellic Acid on Phenylpropanoid Accumulation in Common Buckwheat (Fagopyrum Esculentum Moench) Sprouts. Molecules 2017, 22, 374. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The maximum monthly temperature (°C, orange line) and average monthly temperature (blue line) during the growing season of the onion crop (September 2022-March 2023).
Figure 1.
The maximum monthly temperature (°C, orange line) and average monthly temperature (blue line) during the growing season of the onion crop (September 2022-March 2023).
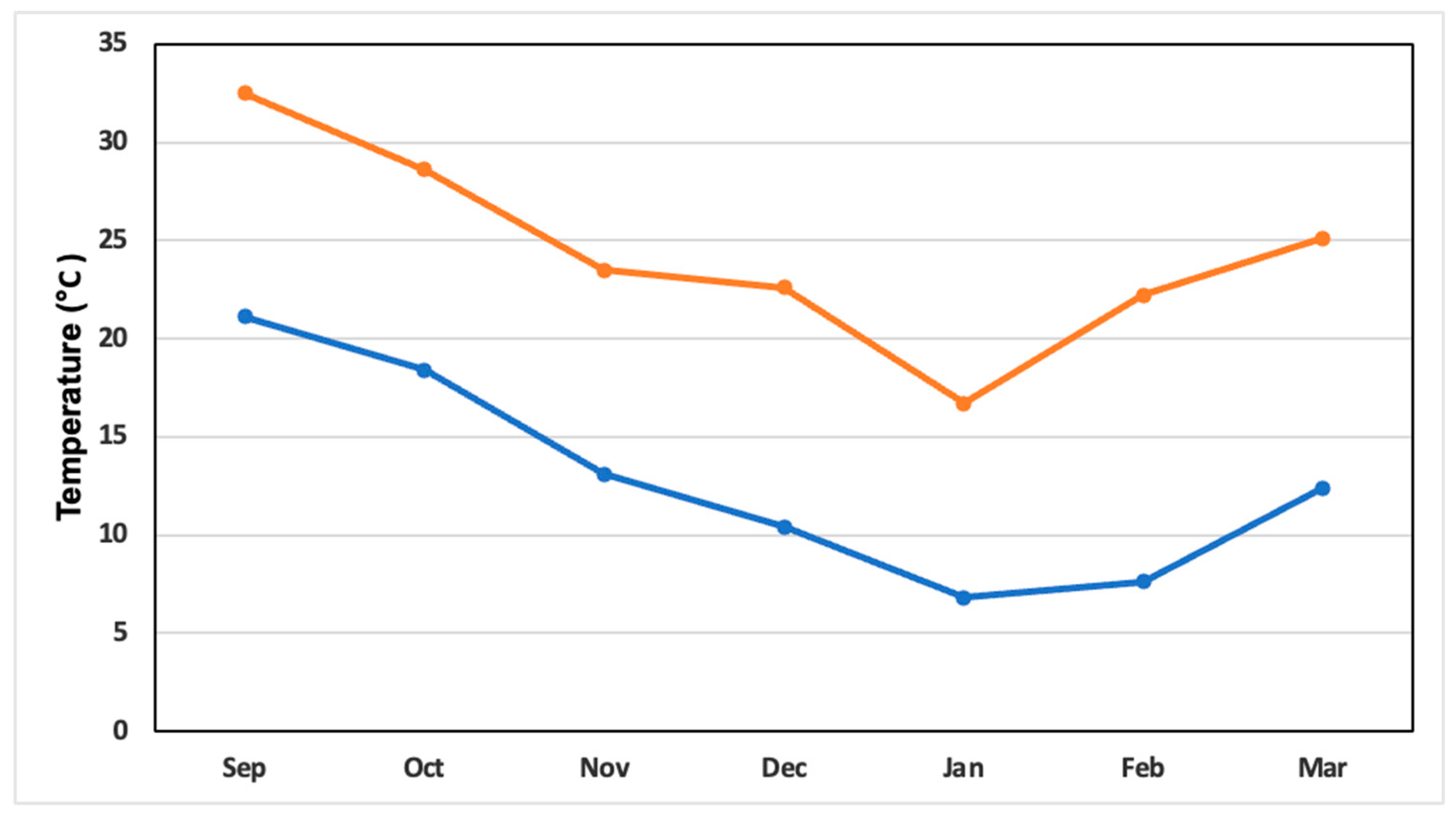
Figure 2.
a. Principal Component Analysis (PCA) of the relationship between chemical parameters; 2b. IAA concentrations and their relationship with chemical parameters.
Figure 2.
a. Principal Component Analysis (PCA) of the relationship between chemical parameters; 2b. IAA concentrations and their relationship with chemical parameters.
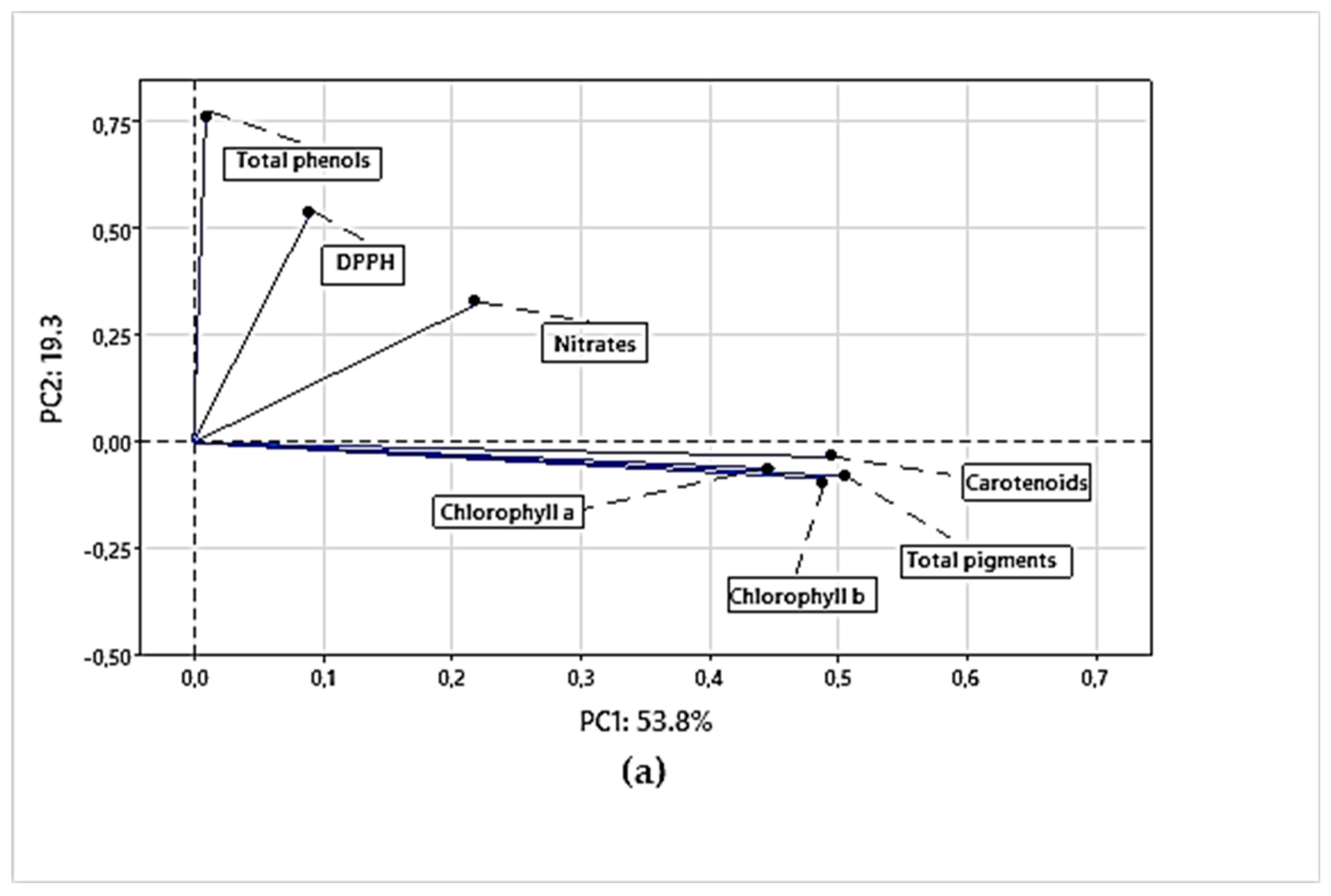
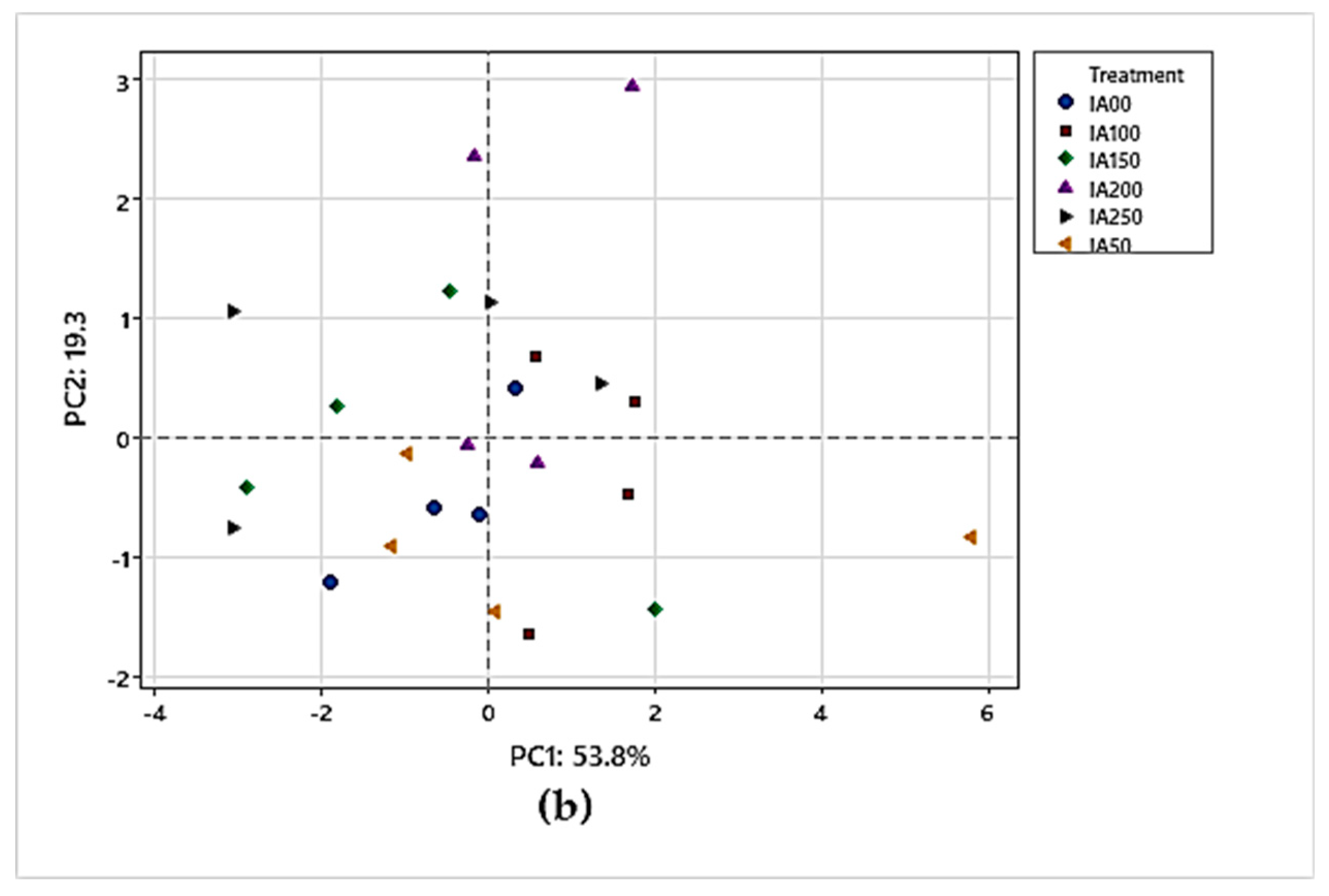
Figure 3.
a. Principal Component Analysis (PCA) of the relationship between physical parameters; 3b. IAA concentrations and their relationship with physical parameters.
Figure 3.
a. Principal Component Analysis (PCA) of the relationship between physical parameters; 3b. IAA concentrations and their relationship with physical parameters.

Figure 4.
a) Cluster dendrogram based on the biostimulant effect on the physical parameters of the onion crop, b) Cluster dendrogram based on the biostimulant effect on the chemical parameters of the onion crop.
Figure 4.
a) Cluster dendrogram based on the biostimulant effect on the physical parameters of the onion crop, b) Cluster dendrogram based on the biostimulant effect on the chemical parameters of the onion crop.
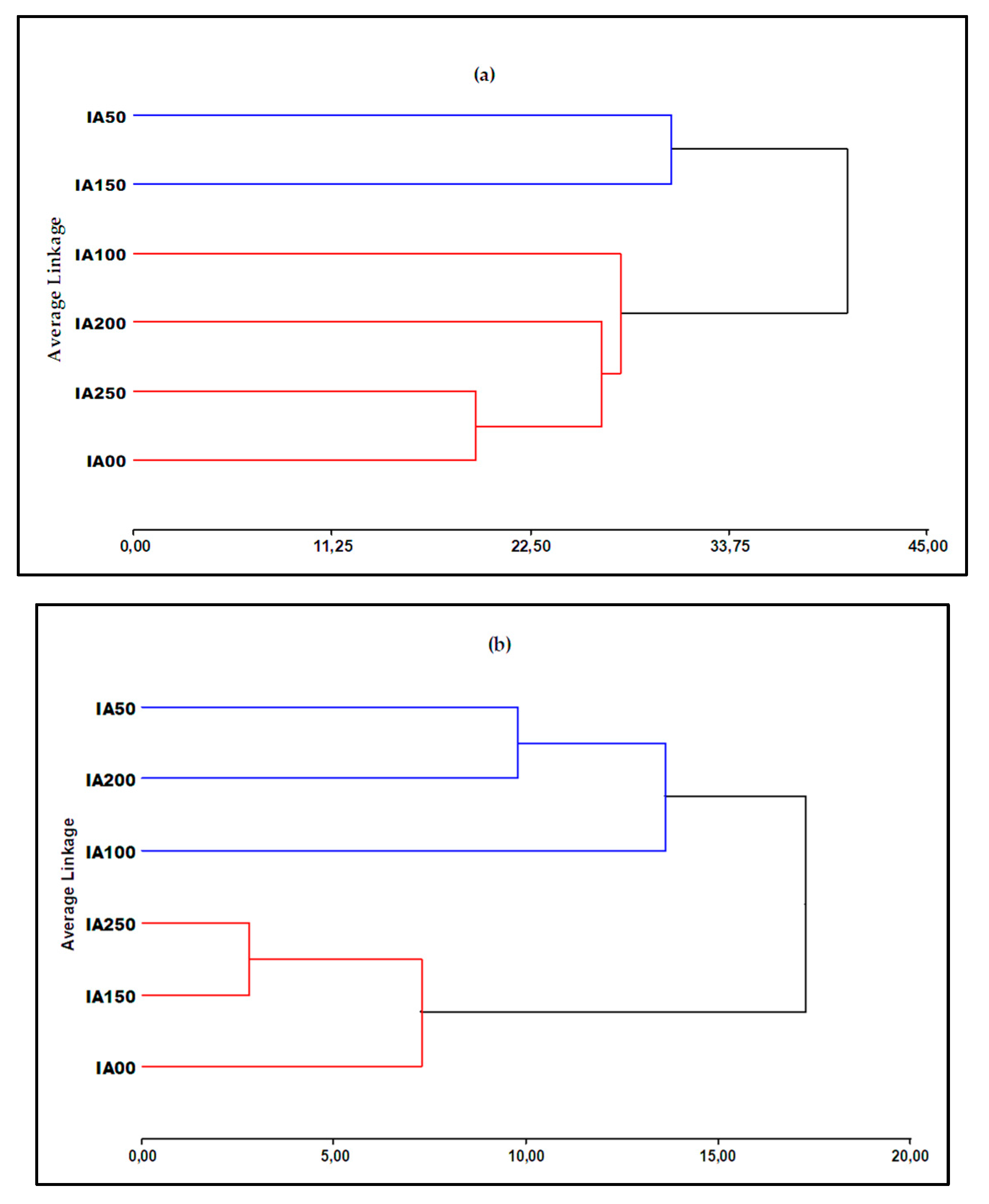
Figure 5.
Cluster dendrogram of physical and chemical parameters in the application of biostimulant in onion cultivation.
Figure 5.
Cluster dendrogram of physical and chemical parameters in the application of biostimulant in onion cultivation.
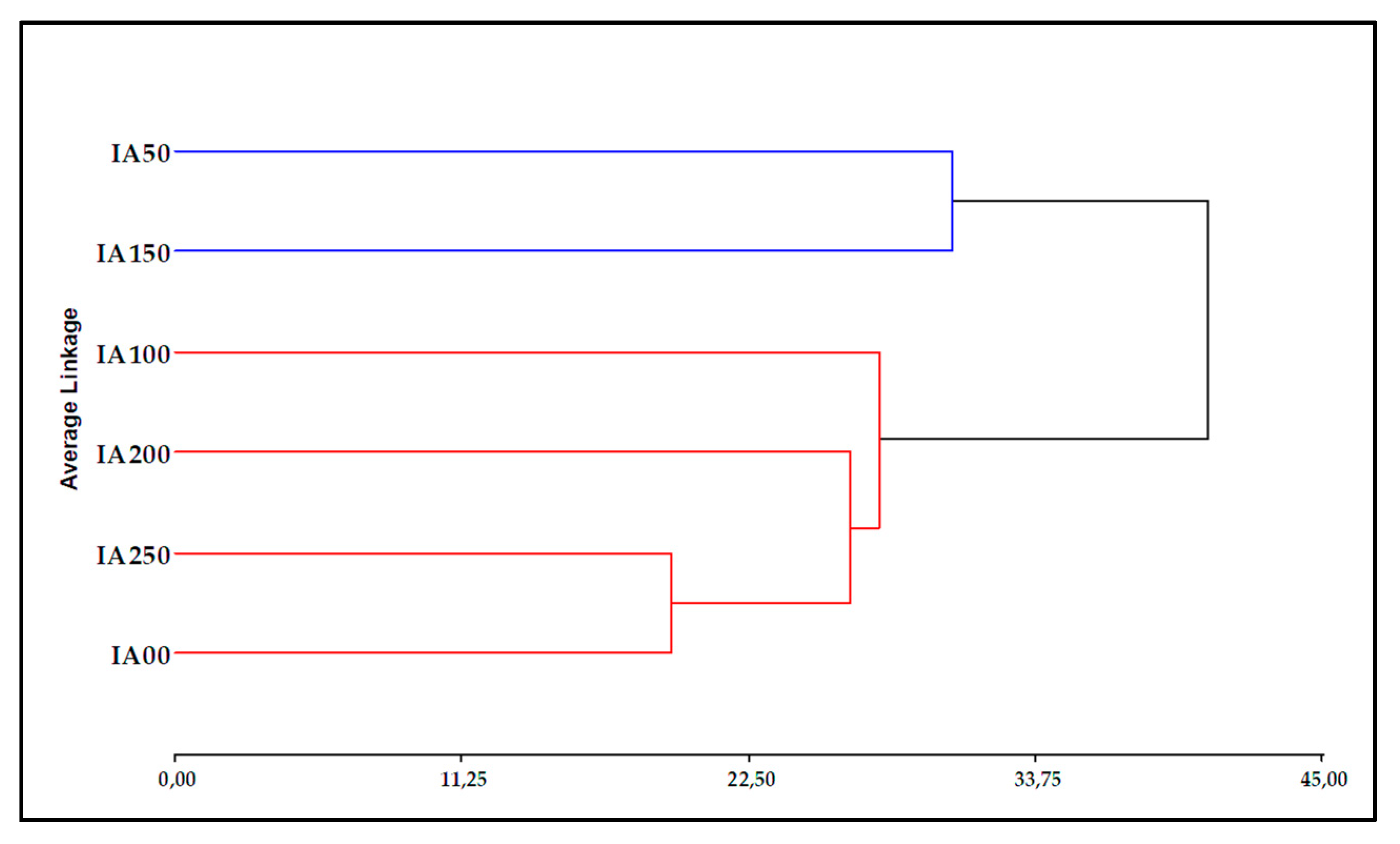

Table 1.
Effect of IAA concentrations by exogenous application on the total fresh biomass of the plant (FW), root fresh weight (FR), bulb fresh weight (FB) and leaf fresh weight (FF) in the pots test of Allium cepa.
Table 1.
Effect of IAA concentrations by exogenous application on the total fresh biomass of the plant (FW), root fresh weight (FR), bulb fresh weight (FB) and leaf fresh weight (FF) in the pots test of Allium cepa.
| Treatments | FW (gr) |
FR (gr) |
FF.(gr) | FB (gr) |
|---|---|---|---|---|
| IAA | ||||
| IA50 | 92.0 ± 11.6 a | 13.63 ±1.39 a | 36.47 ± 3.79 a | 3.688 ± 0.180 a |
| IA100 | 63.88 ± 3.02 ab | 9.500 ± 0.354 a | 26.25 ± 3.49 a | 3.3292 ± 0.0712 ab |
| IA150 | 79.4 ± 16.0 ab | 13.25 ± 2.89 a | 31.87 ± 7.57 a | 3.476 ± 0.186 ab |
| IA200 | 53.00 ± 8.37 b | 9.38 ± 1.01 a | 27.21 ± 6.57 a | 2.7915 ± 0.0678 c |
| IA250 | 62.5 ± 15.4 ab | 12.00 ± 3.23 a | 25.23 ± 6.75 a | 3.127 ± 0.257 bc |
| Control | ||||
| IA0 | 57.5 ± 13.4 ab | 7.75 ± 1.53 a | 24.13 ± 5.67 a | 3.090 ± 0.206 bc |
| Significance | ||||
| biostimulant concentration (B) | ns | ns | ns | * |
The results in Table 1 represent medians ± standard error of the median, while letters (a, ab, b, c) of each column indicate significant differences according to the LSD post hoc test (p ≤ 0.05) in total fresh biomass of the plant (FW), root weight (FR) and bulb weight (FB). ns, not significant; * significant p ≤ 0.05. The FB data did not meet the normality test and were transformed using the natural logarithm LN (X).
Table 2.
Effect of IAA concentrations by exogenous application on plant height (PH), bulb height (BH) and central bulb diameter (CDB) in the Allium cepa pots trial.
Table 2.
Effect of IAA concentrations by exogenous application on plant height (PH), bulb height (BH) and central bulb diameter (CDB) in the Allium cepa pots trial.
| Treatments | PH (cm) |
BH (cm) |
CDB (cm) |
|---|---|---|---|
| IA50 | 56.00 ±2.61 ab | 5.000 ±0.449 a | 4.025 ±0.315 a |
| IA100 | 56.50 ±2.25 ab | 4.625 ±0.492 a | 3.225 ±0.131 a |
| IA150 | 58.25 ±2.78 a | 4.800 ±0.394 a | 3.850 ±0.497 a |
| IA200 | 51.00 ±2.45 b | 5.250 ±0.250 a | 3.200 ±0.618 a |
| IA250 | 54.50 ±1.94 ab | 4.675 ±0.287 a | 3.050 ±0.429 a |
| Control | |||
| IA0 | 52.25 ±1.03 ab | 4.425 ±0.417 a | 3.075 ±0.180 a |
| Significance | |||
| biostimulant concentration (B) | ns | ns | ns |
The results represent medians ± standard error of the median, letters (a, ab, b, c) of each column indicate significant differences according to the LSD post hoc test (p ≤ 0.05) in plant height (AT), bulb height (AB) and bulb central diameter (DCB). ns, not significant (p ≤ 0.05), significant *.
Table 3.
Effect of IAA concentrations by exogenous application on root dry weight (RDW), bulb dry weight (BDW), leaf dry weight (FDW) in the Allium cepa pots trial.
Table 3.
Effect of IAA concentrations by exogenous application on root dry weight (RDW), bulb dry weight (BDW), leaf dry weight (FDW) in the Allium cepa pots trial.
| Treatments | RDW (gr) |
BDW (gr) |
FDW (gr) |
|---|---|---|---|
| IAA | |||
| IA50 | 0.4582 ± 0.0807 ab | 0.532 ± 0.176 a | 0.793 ±0.200 a |
| IA100 | 0.4546 ± 0.0918 ab | 0.5092 ± 0.0480 a | 0.631 ± 0.106 a |
| IA150 | 0.4364 ± 0.0751 ab | 0.797 ± 0.301 a | 0.749 ± 0.190 a |
| IA200 | 0.5508 ± 0.0626 a | 0.448 ± 0.233 a | 0.4419 ± 0.0950 a |
| IA250 | 0.5338 ± 0.0818 a | 0.498 ± 0.148 a | 0.6107 ± 0.0718 a |
| Control | |||
| IA0 | 0.3178 ± 0.0346 b | 0.517 ± 0.175 a | 0.458 ± 0.159 a |
| Significance | |||
| biostimulant concentration (B) | ns | ns | ns |
The results represent medians ± standard error of the median, letters (a, ab, b, c) of each column indicate significant differences according to the LSD post hoc test (p ≤ 0.05) in root dry weight (RDW), weight bulb dry weight (BDW), leaf dry weight (FDW). Ns: not significant (p ≤ 0.05). The BDW was transformed using the square root of (X).
Table 4.
Effect of IAA concentrations by exogenous application on photosinthetic indicators chlorophyll a, chlorophyll b, carotenoids, total pigments and nitrates in the Allium cepa pots trial.
Table 4.
Effect of IAA concentrations by exogenous application on photosinthetic indicators chlorophyll a, chlorophyll b, carotenoids, total pigments and nitrates in the Allium cepa pots trial.
| Treatments | Chlorophyll a | Chlorophyll b | Carotenoids | Total pigments | Nitrates |
|---|---|---|---|---|---|
| (mg/ fw) | (mg/ fw) | (mg/ fw) | (mg/ fw) | (mg/ kg fw) | |
| IAA | |||||
| IA50 | 0.445 ± 0.120 a | 0.409 ± 0.131 a | 0.2006 ± 0.0587 a | 0.4473 ± 0.0384 a | 1424 ± 174 b |
| IA100 | 0.3706 ± 0.0286 a | 0.4532 ± 0.0275 a | 0.1987 ± 0.0230 a | 0.5869 ± 0.0320 a | 3734 ± 763 a |
| IA150 | 0.2941 ± 0.0595 a | 0.2983 ± 0.0739 a | 0.1253 ± 0.0533 a | 0.4193 ± 0.0943 a | 2422 ± 552 ab |
| IA200 | 0.4135 ± 0.0580 a | 0.3598 ± 0.0414 a | 0.1557 ± 0.0165 a | 0.5565 ± 0.0447 a | 2278 ± 366 ab |
| IA250 | 0.2669 ± 0.0551 a | 0.2405 ± 0.0816 a | 0.1270 ±0.0565 a | 0.3627 ± 0.0942 a | 2104 ± 619 b |
| Control | |||||
| IA0 | 0.2592 ± 0.0133 a | 0.3561 ± 0.0578 a | 0.1291 ±0.0231 a | 0.4473 ± 0.0384 a | 2072 ±371 b |
| Significance | |||||
| biostimulant concentration (B) | ns | ns | ns | ns | ns |
The results represent medians ± standard error of the median, letters (a, ab, b, c) of each column indicate significant differences according to the LSD post hoc test (p ≤ 0.05), in chlorophyll a (CHA) chlorophyll b (CHB), carotenoids (CR), total pigments (TP) and nitrates (NT). Ns: not significant (p≤ 0.05). CHA, CHB, TP data were transformed using square root (X).
Table 5.
Effect of IAA concentrations by exogenous application on total phenols (TF), and antioxidant capacity (DPPH), in the Allium cepa pot assay.
Table 5.
Effect of IAA concentrations by exogenous application on total phenols (TF), and antioxidant capacity (DPPH), in the Allium cepa pot assay.
| Treatments | FT (mg AGE/g dm) |
DPPH (μmol TROLOX/g dm) |
|---|---|---|
| IAA | ||
| IA50 | 0.690 ± 0.269 c | 7.577 ± 0.413 ab |
| IA100 | 1.600 ± 0.597 bc | 6.007 ± 0.272 bc |
| IA150 | 2.692 ± 0.364 abc | 5.421 ± 0.925 c |
| IA200 | 3.989 ± 0.766 a | 7.94 ± 1.00 a |
| IA250 | 2.973 ±0.394 ab | 6.662 ± 0.546 abc |
| Control | ||
| IA0 | 0.635 ± 0.449 c | 7.498 ± 0.361 ab |
| Significance | ||
| biostimulant concentration (B) | * | ns |
The results represent medians ± standard error of the median, letters (a, ab, b, c) of each column indicate significant differences according to the LSD post hoc test (p ≤ 0.05) in total phenols (FT), DPPH, ns not significant; significant * p ≤ 0.05. FT, DPPH data was transformed using natural logarithm LN(X).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



