
Submitted:
29 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
The etiological agent of various viral diseases has been identified, and clinical characteristics and complications of each disease are well known. In general, a viral disease appears as acute systemic inflammatory disease, but after acute infection, some patients remain asymptomatic carriers or suffer chronic viral diseases, suggesting that the phenomena are unavoidable processes for viruses’ coexistence with their hosts as virome in microbiota of human species. Although each viral disease has been described as having different pathophysiology, the host's control systems, including immune systems, against the insults from any viral infections, have the same components in previously healthy hosts. Therefore, considering the characteristics of the life cycle of viruses being dependent on the host cell and the same immune components of the host, it is reasonable assumption that there is a common immune mechanism to respond to the insults from all viral diseases. The authors discuss on the characteristics of virus infection and unresolved issues in viral diseases and propose a rationale of early immune modulators for acute insults from viral diseases based on our clinical experiences and the protein-homeostasis-system hypothesis.
Keywords:
virus infection
; virome
; microbiota
; pathophysiology
; immunology
; protein-homeostasis-system hypothesis
1. Introduction
The life cycle of viruses is dependent on the host cells, and viruses have an affinity to certain host cells, where viruses perform their replication, production of progeny, and residency. Initially infected cells (these are named the focus in this article) may locate in the environmental interface of the host such as oropharynx, intestine, skin, and urogen-ital tract, and along with those cells, secondary lymphoid tissues are distributed as protective walls against invading pathogens. However, identifying the focus as the origin of disease-onset is difficult in most viral infections, and all injured organ cells during viral disease processes have not affinity to the virus.
The infectious disease has clinical characteristics, including discovery of etiologic agent, self-limited natural course, main involved organ, and complications. It has been believed that pathophysiology of each viral disease is different according to species; for example, the pathogenesis of pneumonia/acute respiratory distress syndrome (ARDS) in coronavirus disease 2019 (COVID 19) and that of influenza or other respiratory pathogens differ. That concept may be based on the notion that viruses themselves are responsible for lung cell injuries causing pneumonia/ARDS, and that a virus-induced cytopathy is main pathophysiological role on viral infections. However, there are many enigmas that cannot be explained by virus-induced cytopathy, and it is undoubtful that all diseases involve etiological or inflammation-inducing substances for disease-onset [1,2,3,4].
Human viral diseases are caused by the viruses of their own species although external viruses belonged to other species or places can affect human beings on occasion. Accordingly, most of pathogenic agents in viral infections as well as those in bacterial infections may be the strains in microbiota of human species. After initial infection some patients remain asymptomatic carriers (or reservoirs) or are affected with chronic viral diseases in cases of human immunodeficiency virus (HIV) and hepatitis B virus (HBV) infections.
The host's control systems against the insults from any viral infections, including the components of immune system, might be the same in individual hosts of a species, although severity of the disease can differ across infected individuals. The immune system of mammals, including human beings, reacts not only to pathogens themselves, but also to substances derived from pathogen-infected cells, including fragments of pathogens, pathogen-associated molecular patterns (PAMPs), damage (or danger)-associated molecular patterns (DAMPs), pathogenic proteins such as toxins, pathogenic peptides, and other smaller substances being not yet identified. Since current immunological concepts have limitations to explain the unresolved issues in infectious diseases, new concepts or hypotheses are needed for solving the enigmas. It is proposed that the immune system of organisms evolves to protect their cells, and there is an integrated control system for well-being of organisms; the host’s immune system controls toxic substances for protecting the cells according to size and biochemical property of the substances (the protein-homeostasis-system (PHS) hypothesis) [5,6,7,8,9].
In this paper, we further discuss on the unresolved issues in viral diseases and propose a common pathophysiology of viral diseases through the PHS hypothesis. Because the PHS hypothesis broadly integrates of the pathogenesis of virus diseases, known pathophysiological or clinical characteristics on a specific virus disease based on the current immunological concepts or understanding are omitted or reinterpreted.
2. The nature of virus infections
Viruses are extremely small infectious agents that replicate inside the living cells of an organism. Viruses are found in almost every ecosystem on Earth and infect all life forms, from animals and plants to microorganisms, including fungi, bacteria and archaea [10]. In addition, viruses can be regarded as life forms that carry genetic material, reproduce, and evolve through natural selection, although they lack criteria for defining life, such as cell structure and self-reproduction [11]. Viruses are the most numerous types of biological entities. Since the first discovery of the tobacco mosaic virus by Martinus Beijerinck in 1898, more than 9,000 of the millions of virus species have been described in detail [12,13].
The morphology and characteristic of viruses, such as shape, size, and type and amount of genetic material, differ by species, and viruses are classified according to the characteristic of their genomes [14]. Generally, virion is a form of infectious viral particle consisting of 1) genetic material, that is, deoxyribonucleic acid (DNA) or ribonucleic acid (RNA) that encodes the structural and enzymatic proteins by which the virus acts, 2) the capsid, which is a protein coat that surrounds and protects the genetic material, and 3) an outside envelope of lipids in some species. The host immune system recognizes virions as a whole and controls them using phagocytes, while fragments of virions produced during replication processes, such as viral-origin proteins, RNAs, and DNAs are controlled by other innate and adaptive immune systems. Classically, antigen-presenting cells (APCs) first engulfs virions and then presents processed viral-origin proteins and peptides to B and T cells, respectively. On the other hand, viral proteins, RNAs, DNAs, and envelope lipids within infected cells can elicit immune reactions directly if the substances are released into the systemic circulation.
The life processes of viruses, including influenza viruses, hepatitis C virus (HCV), and HIV, have been studied [15,16,17]. In general, there are six stages in the life cycle of viruses: 1) attachment, 2) penetration, 3) uncoating, 4) replication, 5) assembly, and 6) release. These processes are complex at each stage and may require the cooperation of host cells for the production of viral progeny since viruses have only limited genes that cannot perform the complex tasks, and mechanisms for attachment, replication, and assembly may require precise control systems that are associated with host cells. The development of antivirals has targeted blocking each process of the viral life cycle and host cell interactions, and these drugs include monoclonal antibodies against host cell receptors and peptide-based inhibitors against viral proteins for blocking of attachment or penetration stage, and inhibitors that target viral enzymes such as nucleoside reverse transcriptase, polymerase, or protease for blocking replication process [18,19].
Viruses attach to complementary host cell receptors and enter host cells. During the replication processes, various types of viral proteins and nucleic acids, including viral-specific messenger RNAs (mRNA) for structural and nonstructural proteins, including polyproteins and RNA-dependent RNA polymerase, are produced with the assistance or control of host cells because they provide materials such as nucleotides and phospholipids for viral nucleic acids and envelope and biological factories such as ribosomes. Viroids are RNA molecules that lack a structural protein coat and can infect cells in the form of genetic materials [20]. Viral genetic material and structural proteins are assembled into progeny viruses, and the virions are released from the infected cells. It is believed that virus-infected cells contain larger numbers of viral proteins and nucleic acids than the number of virions by a factor of 103–104 [21]. On the other hand, a host cell as an independent biosystem can operate protective mechanisms against external insults such as virus invasion or physical stresses, including heat and trauma. When viruses invade cells, the affected cells produce interferons and proteins associated with interferon-related pathways, and other immune proteins and peptides are activated. It has been suggested that acute systemic manifestations of a viral infection, such as measles, are associated with viremia from a secondary focus, including cells of the regional lymph nodes [22]. Additionally, a variety of viruses can induce a “viral sepsis” although definition, epidemiology, and pathogenesis of the condition remains to be further evaluated [23]. When viremia occurs, virus-origin and host cell-origin substances spread systemically and locally through the vascular system. Moreover, it is possible that the load and type of substances, including intact virions, differ according to the number or type of infected cells, and that insults from various virus infections could induce the same organ cell injury of the host. Although the cytopathic effects of viruses have been observed in vitro, some viruses do not show cytopathic effects, and intact virions are rarely found in injured cells in acute viral infections, including lungs, central nervous system (CNS), or other affected organ lesions in influenza and COVID-19 [1,24,25,26]. The efficacy of viral progeny production is dependent, at least in part, on host factors, and the severity of viral infections depends on the loads of substances derived from the focus.
2.1. Viruses and microbiota
Because viruses live with any life on Earth, they may have existed since living cells first appeared and would have depended on cellular life. The genetic material of some viruses can be integrated into the genome of host organisms, and a part of the human genome contains ancient viral DNA sequences [27]. DNA sequences can be passed to the host offspring for many generations. These findings suggest that viruses have evolved with the cells that first appeared on Earth over billions of years [11].
The microbiota (or microbiome) is an ecosystem found in all multicellular organisms, and the microbiota and host have co-evolved in a collaborative partnership. Microbiota include bacteria, archaea, protists, fungi, and viruses, and in human species, they reside on or in the host cells of the oropharynx and lower respiratory tract, gastrointestinal tract, skin, urogenital tract, and possibly other undetected sites [28]. Although viruses in the human microbiota have received less research attention, there has been a recent increasing interest in the virome in the gut and other sites [29,30,31,32]. Studies on the human virome have revealed that bacteria in the microbiota of the host live with viruses, that is, bacteriophages. Bacteriophages constitute most of the virome, and endogenous retroviruses, eukaryotic viruses, and viruses associated with archaea are small portions [29].
Dysbiosis, the disruption of the collaborative relationship between the microbiota and the host, has become an important subject in various biomedical fields. Researchers have reported the relationships between altered microbiota and diseases in their fields, including obesity, autoimmune diseases, cancers, and neuropsychiatric diseases [33,34,35]. The mechanisms by which dysbiosis induces disease remain elusive. Overproduction of toxic materials by dysbiotic microbes, vulnerability to invasion by microbes through weakened mucosal barriers, and disruption of the homeostatic relationship between microbiota and the host immune system, including the development and function of the immune system, have been suggested to be associated with disease [35,36,37]. Dysbiotic states of viruses in the microbiota may also play crucial roles in host immunological, biological, and metabolic homeostasis, and may be related to the development and progression of some diseases [29,30,31,32].
2.2. Species-specificity and adaptation of viruses
Infectious diseases have a strict species-specific relationship between the pathogen strains in the microbiota and the host species. Each animal species has had its characteristic influenza viruses, coronaviruses, and bacteria strains, including mycoplasma strains, for a long time [38,39,40]. For example, most bacterial infections are caused by bacterial strains in the host itself or other hosts in humans, including Escherichia coli and Streptococcus pneumoniae. Thus, all pathogenic agents that are species-specific may be regarded as strains in the microbiota of a species, although viruses, bacterial and fungal strains of other species can infect hosts of a species. It is possible that viruses adapted or specific to human species, such as human influenza viruses and coronaviruses, including severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2), could easily produce their progeny and permit the existence of asymptomatic carriers or reservoirs to spread progeny. By contrast, viruses in avian influenza, SARS, and Middle East respiratory syndrome (MERS) from other animal species may produce their progeny less effectively, and there are no human carriers for existence in the human species [3]. In addition, viruses from cold-blooded animals, plants, and oceans may not be permitted to attach to or produce progeny through the unknown control systems of the host or virus-host interaction. A close interaction between the species and adapted strains in the microbiota might manifest as low virulence in the host of the species. It is now known that nearly all children and adults worldwide have been infected with SARS-CoV-2 during the COVID-19 pandemic, and infants, neonates, and even fetuses of infected mothers have had uneventful outcomes from the early stage of the pandemic [3,41]. Most of the deaths in the pandemic are in older hosts with underlying diseases or immune-compromised hosts, as observed in other infections caused by strains in the microbiota, including seasonal influenza, common cold, and outbreaks of E. coli [42].
It is natural that increasing of susceptible hosts that are not infected with pre-existing strains are responsible for outbreaks or epidemics as shown aseptic meningitis or hepatitis A caused by enteric viruses [43,44]. On the other hand, it is possible that a new strain appears and substitutes pre-existing strains in microbiota for maintenance of life with their hosts in a species when most hosts have immunity to pre-existing strains in microbiota, as observed in the 2009 H1N1 pandemic and COVID-19 pandemic [3]. Thus, it is not a strange prediction that a new epidemic or pandemic of adapted viruses, including enteroviruses and other respiratory viruses, will come in the near future. Although the disease caused by the new strain appears more severe or virulent than that caused by pre-existing microbiota strains in the early stages of an epidemic or pandemic, the strains of the viruses have soon changed to less virulent forms, as observed during the 2009 H1N1 and COVID-19 pandemics [3,45]. This adaptation may be established and strengthened by the passage number of infected persons, such that many times passaged viruses show attenuated virulence in vaccine development and animal study. In addition, because antimicrobial-resistant strains, such as the extended-spectrum beta-lactamase (ESBL) producing strains in acute pyelonephritis and macrolide-resistant Mycoplasma pneumoniae (MP), may appear by the passage through infected hosts, they may be less virulent in immune-competent hosts over time [46,47].
Since close species-specific interactions between the host (or host cells) and microbiota are in partnership, some viruses from animal species, including influenza viruses and HIV, could adapt to human species over time. It is possible that some pathogenic viruses, bacteria, mycoplasmas, and fungi at the present time are adapting or departing from human species. Similarly, severe infectious diseases, including pandemic influenza, scarlet fever, acquired immunodeficiency syndrome (AIDS), and infection-related immune-mediated diseases, such as Kawasaki disease (KD), acute poststreptococcal glomerulonephritis (APSGN), and Henoch-Schönlein purpura, have become less severe over time in Korea [48,49,50]. These phenomena suggest that disease phenotypes can change as pathogens adapt to the human species over time. Although emerging and re-emerging viruses, such as SARS-CoV, MERS-CoV, avian influenza A viruses, Zika virus, and enteroviruses, including hepatitis A virus, have posed significant global public health problems, among the pathogenic strains, those that cannot adapt as microbiota might be eventually extracted from human species over time.
2.3. Microbiota, infectious disease, and immune-mediated diseases
Although a homeostatic balance between the microbiota and the host is important for the well-being of the host, some strains in the microbiota can invade the host. Commensals, including E. coli and S. pyrogens, lead to infectious diseases, such as acute pyelonephritis, otitis media, and pneumonia, and postinfectious immune-mediated diseases, such as acute rheumatic fever, APSGN, and possibly KD [46,51]. In addition, viruses in chronic viral infections such as herpes viruses, HIV, and hepatitis viruses can be reactivated in unknown-residing cells after the initial infection.
The microbiota composition in individual hosts changes continuously after birth, together with immune maturation [52,53]. In addition, the strains of microbiota are influenced by environmental factors such as diet and socioeconomic status, including hygiene and antibiotic use [54]. Thus, the composition of the microbiota differs between ethnic groups or populations and can be changed by changing the environment. The prevalence or incidence rates of infectious diseases, including HIV/AIDS, hepatitis B and C, and infection-associated immune mediated diseases, including KD, a multisystem inflammatory syndrome in childhood (MIS-C), and juvenile idiopathic arthritis (JIA), differ widely among populations [8,55,56,57]. In addition, some childhood immune-mediated diseases, including KD and some subgroups of JIA, have a predilection for age or sex [8]. Although susceptibility to diseases, especially immune-mediated diseases such as KD, MIS-C, and JIA, has been explained by genetic factors such as race and ethnicity, the clinical manifestations of the disease and immune function against infectious insults may be similar across populations. Because the strains in the microbiota colonize first and then invade the host and/or host cells, the different incidences among ethnic groups and age or sex prediction could be explained by the colonization state of the etiological agents in the microbiota. Patients or animals living in higher prevalence areas may have a higher chance of being exposed to pathogens originating from the microbiota, while genetic differences affecting immune functions in individual hosts may be involved in disease outcomes [8,9,58].
3. Pathophysiology of viral infections
3.1. Immune reactions to viral infection
The host immune system controls the viral infection. Current immunological concepts have been developed and established mainly through studies on invading pathogens. Immunopathogenesis of viral diseases is based on the notion that the virus plays a major role in cell injury in various organs. In previously immune-competent patients, virions exposed in the bloodstream may be effectively removed by phagocytes such as neutrophils and monocytes because polymerase chain reaction (PCR) assays in the blood are commonly negative except for chronic viral infections, regardless of the severity and stage of the disease and virus-specific antibodies. There are few intact virions in extensively injured tissues and in the blood as previously discussed. Thus, isolation of blood-borne virions as HIV and HCV might take a long time or be difficult.
Phagocytes express receptors for immunoglobulins and complement, and other receptors, and phagocytic activity is more effectively achieved with the assistance of pathogen-specific antibodies and complement components [59]. Innate immune cells express the pattern recognition receptors (PRRs) such as toll-like receptors (TLRs) or other intracellular sensors for binding to PAMPs or DAMPs. Activated cells via these receptors produce immune proteins, including proinflammatory cytokines, and control the interferon-mediated immune pathway and subsequent adaptive immune cells fighting against infectious insults [60,61]. Also, other innate immune components such as interferons, DNA traps of neutrophils, and activation of complement pathway may be helpful to removal of virions [62,63]. The basic innate immune reactions may be ineffective to virions within cells because of no direct contact to virions. The virions within cells could produce and release infective particles and nucleic materials without cytopathic effect, and they could adapt to the insults such as antivirals and produce variants against them. Although only adaptive immune system has been believed to have an ability of immunological memory on previous infection, now it is known that innate immune system in mammals has an inheritance of acquired immune resistance, named trained immunity, as like in plants and insects [64]. The progeny of previously infected mammals has a memory of infection that confers enhanced protection against infection, and that is like that provided by the adaptive immune system. The innate memory in natural killer (NK) cells and phagocytes may be associated with epigenetic reprogramming of transcriptional pathways rather than gene recombination [64,65].
T cells and B cells in adaptive immune system cannot directly control virions but they are only associated with protein fragments of virion. Since a B cell clone produces a specific antibody against a pathogenic protein, likewise a pathogenic peptide induces a specific T cell clone against the peptide, there are many virus-specific B cell and T cell clones in a viral infection. It is possible that B cells and T cells react to not only viral proteins and peptides presented by APCs, but also those are presented by or bound to other cells when viremia occurs as previously mentioned. Virus-specific antibodies induce a virus neuralization and can prevent viral entry process, and they can help a cytopathic effect via antibody dependent cellular cytotoxicity (ADCC) to target cells [66]. It has been suggested that cytotoxic T cells (CD8+) that target viral peptides bounded in major histocompatibility complex (MHC) I molecules on infected cells may be main effectors against viral infections, and helper T cells (CD4+), including Th1, Th2, and regulatory T cells, are involved in control of antibody production and immune homeostasis against disease insults. However, the modes of T cell function remain to be further evaluated [5,6].
On the other hand, it is worth note that injured infected cells by cytotoxic T cells or other immunologic insults such as cytokine storm, NK cells, or activated compliment pathways could release virions and other substances [67,68,69]. All immune cells in innate and adaptive immune systems, including CD4 T cells, CD8 T cells, and NK cells, and immune proteins, including immunoglobulins and complements, perform their functions during noninfectious events, such as trauma and wound healing, transplantation rejection, allergy, intoxication, and cancer. Thus, each component in both systems may have same functions in these conditions [5,6]. Moreover, not only immune system but also various intracellular or systemic biological and metabolic systems, including inflammasome with pyroptosis, apoptosis, autophagy, proteosome, thromboembolic pathway and glycolysis pathway, and the epigenetic change, including micro-RNAs, are activated in systemic inflammation of infectious diseases, including COVID-19 [70,71,72,73]. It is a reasonable assumption that there are substances eliciting these diverse biological changes and corresponding control systems against the substances. In addition, majority of the biological changes are observed not only in acute infectious diseases but also in other acute or chronic immune-mediated disease, such as KD, macrophage activation syndrome (MAS), MIS-C, and systemic JIA. The biological changes may be useful for detecting diagnostic or prognostic biomarkers and developing targeted therapies for each disorder. Biological alterations observed under various conditions could contribute to the de-velopment or progression of the disease; otherwise, they could also be secondary or adaptive phenomena, indicating immune reactions to insults caused by the disease.
3.2. Common immunopathogenesis of infectious diseases through the PHS hypothesis
We have proposed the PHS hypothesis for further understanding the pathophysiology of diseases, and the PHS hypothesis has already been introduced for infectious diseases, including influenza, MP pneumonia, COVID-19, and ARDS; immune-mediated diseases such as KD and MIS-C; kidney diseases, including genetic diseases and cancers; and CNS diseases, including prion diseases and Alzheimer disease [1,2,3,4,5,6,7,8,9]. Briefly, life as a biosystem has an integrated system, named the PHS, to maintain a state of well-being. Every disease involves etiological or inflammation-inducing substances that are toxic or signaling to their cells. These substances have variable sizes and biological forms, such as extremely small materials, including elements, monoamines, neuropeptides, biochemicals and chemicals (drugs), peptides, and proteins, and larger complex materials, such as virions and bacteria. The small substances can be classified as protein and nonprotein substances, and even the smallest substances, such as elements and monoamines, can bind affinitive receptors on and in host cells. Innate immune systems control small nonproteins and larger complex materials, such as bacteria, virions, apoptotic and necrotic bodies, and transformed cells, while adapted immune systems control protein substances; B cells control pathogenic proteins, and T cells control pathogenic peptides. In addition, the PHS controls in part regarding protein deficiency in cells and the host, as shown in Table 1 [9], and the schematic diagram of pathophysiology of virus diseases is shown in Figure 1.

Table 1.
Etiological substances and corresponding immune effectors in the PHS hypothesis.
| Etiological substances (or events) | Corresponding immune effectors |
| Pathogenic proteins (BCR-associated) | B cells: antibodies against pathogenic proteins |
| Pathogenic peptides (TCR-associated) | T cells: peptides or cytokines against pathogenic peptides |
| Pathogenic small peptides, monoamines, their metabolites (especially in CNS) | Immune proteins such as PrP gene products and other amyloid proteins, mast cell-associated immune responses |
| Non-protein materials such as LPS, RNAs, DNAs, chemicals, biochemicals | TLR-associated immune responses, natural antibodies, other immune systems such as complements and other proteins |
| Large complex substances; viruses, bacteria, parasite, apoptotic & necrotic bodies, and transformed cells | Phagocytes (neutrophils and macrophages), eosinophils (in case of large parasites), and natural killer cells |
| A protein deficiency or malfunctioning protein in organ tissues or within a cell | Production of alternative proteins in genetic diseases and cancers |
Abbreviations. BCR, B cell receptor; TCR, T cell receptor; PrP, prion protein; CNS, central nervous system; LPS, lipopolysaccharide; TLR, Toll-like receptor (from Ref. 9).
Figure 1.
Schematic diagram of pathophysiology of virus infections.
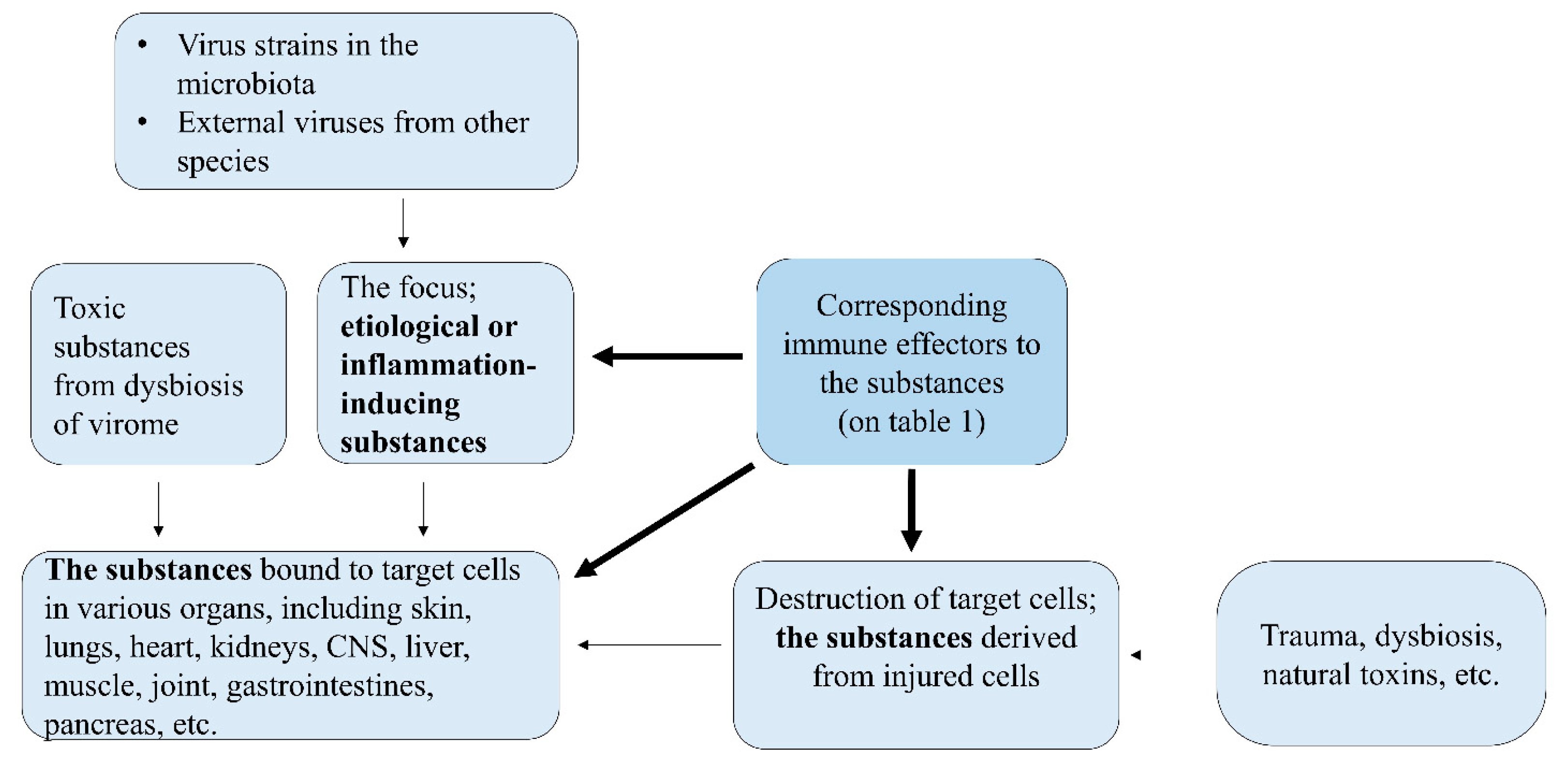
The injury of organ cells in every disease, including viral infections, may begin with the binding of toxic substances to affinitive receptors of organ cells (target cells), and each component in the immune system is activated to protect the cells. The host immune system controls toxic substances not only those originating from pathogens, including toxins and PAMPs, but also those originating from injured or infected-host cells, including DAMPs, pathogenic proteins, pathogenic peptides, and other smaller biochemical substances, especially in intracellular pathogen infections such as virus, chlamydia, rickettsia, and legionella and salmonella species. Therefore, it is not strange that pathogens and/or pathogen fragments cannot be detected in extensively injured cells in pathologic lesions or remote lesions in postinfectious immune-mediated diseases that may result from bacteremia or viremia.
Infectious and infection-related immune-mediated diseases are caused by substances derived from the focus in the PHS hypothesis. The load of substances determines the severity of the disease, and the host’s control system against the substances, that is, the host’s immune status, is responsible for the initiation of inflammation, progression, chronicity, and determining the prognosis of the disease. Clinical symptoms such as fever, myalgia, and pneumonia in COVID-19, influenza, or MP infection begin when toxic substances from the focus abruptly spread systemically and locally and bind to target organ cells and induce immediate immune reactions. The released toxic substances can differ according to the type of infected cells (focus) of the host, and the same host cells can release different toxic substances depending on the pathogen species. Thus, the main target cells and incubation period for infectious diseases can differ between individuals and animal species. Moreover, substances derived from injured target cells (lung cells in the case of pneumonia) caused by initial immunological insults can induce further inflammation if released into the systemic circulation or near local lesions. Extrapulmonary manifestations such as skin rashes, myositis, encephalopathy, and other organ cell injuries occurring in the acute and convalescent stages are also caused by substances derived from the focus or injured target cells. Severe pneumonia or ARDS due to viral infection tends to induce subsequent bacterial invasion in patients, which add to the workload of immune cells.
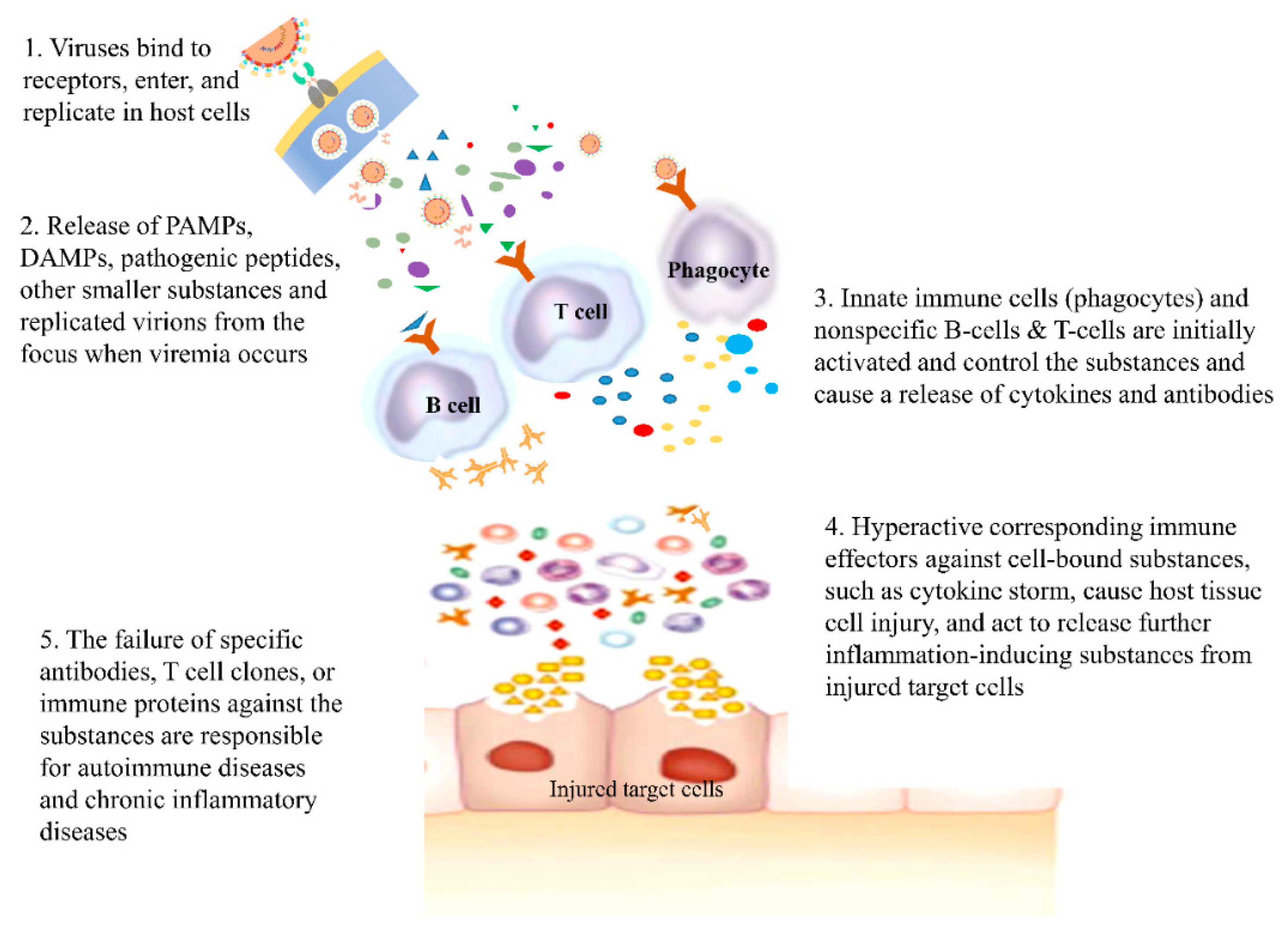
As immune reactions occur through networks across immune components, various immune substances, including proinflammatory cytokines and proteolytic proteins, are also activated. It has been suggested that hyperimmune or aberrant immune reactions, such as cytokine storms, are related to the target cell injury [67,74]. Moreover, it is proposed that autoimmune diseases, including systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA), and chronic inflammatory diseases, including Alzheimer’s disease and amyloidosis, are caused by the failure of the control systems, including specific antibodies, specific T cell clones, and specific immune proteins such as prions, against the etiological substances derived from injured host cells [9]. Therefore, early control of target cell injuries from initial hyperactive immune reactions, performed by initially acting nonspecific adaptive immune cells, is crucial for reducing morbidity and preventing disease progression in patients with severe pneumonia and ARDS, as well as in those with acute infection-related whole-organ diseases, including fulminant hepatitis, myocarditis, necrotizing pancreatitis, rapidly progressive glomerulonephritis, extensive epidermolysis, Water-house-Friderichsen syndrome (adrenal glands), and encephalopathy (Figure 2).
Figure 2.
The pathogenesis of virus infection & infection-related autoimmune diseases.
3.3. Clinical aspects of viral infections
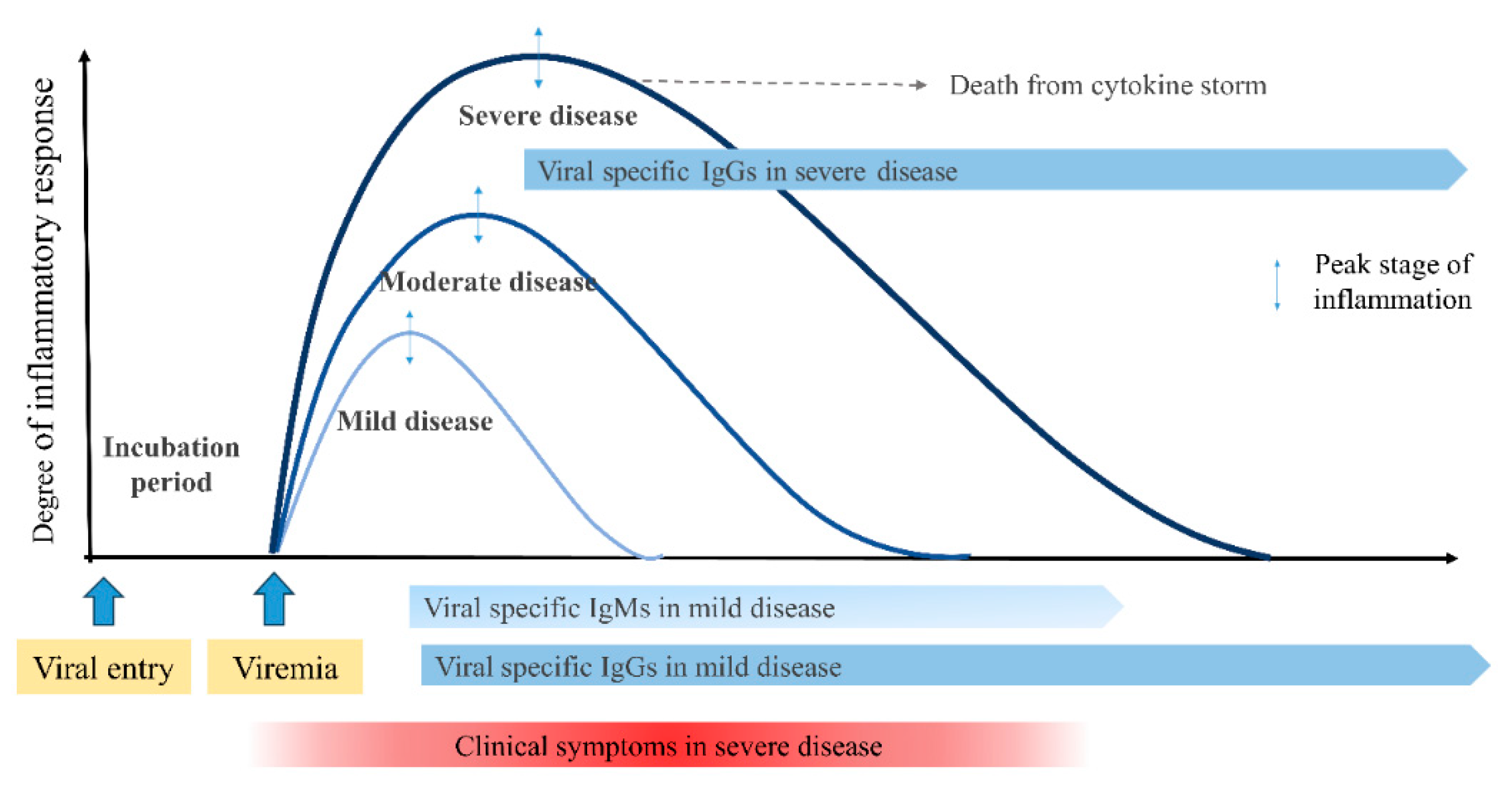
Generally, a viral disease begins as an acute systemic inflammatory process, reaches the peak stage of inflammation, and resolves the inflammation. Inflammation triggered by insults from any infections is a protective host response crucial to restrain pathogen and induce repair responses. Most patients recover from the disease through the immune system. During this process, some patients are severely affected or have fatal outcomes. Systemic inflammatory processes are reflected by inflammatory biomarkers associated with the activation of various immune components, including activated immune cells, immunoglobulins, cytokines, pathogen-specific antibodies, and T-cell clones. Because most patients with infectious and infection-related immune-mediated diseases are self-limiting, immune reactions before the peak of the inflammation process (proinflammatory biomarkers may be involved in this stage) may be involved in target cell injury. Immune reactions after the peak are involved in tissue cell repair (anti-inflammatory biomarkers) and induce the convalescent stage [5]. In addition, the intensity of systemic inflammation during this process is reflected in laboratory parameters, such as white blood cell count and differential (lymphopenia), C-reactive protein (CRP), procalcitonin, lactate dehydrogenase, and immune proteins, including immunoglobulins, cytokines, chemokines, and other biomarkers. All severe pneumonia cases from any pathogen infections showed similar clinical and laboratory findings, suggesting that common pathogenesis may involve severe insults [8]. Thus, the early control of the inflammatory process before the peak stage is critical for infectious diseases, including COVID-19, influ-enza, and MP infection, and infection-related immune-mediated diseases, including KD, MIS-C, and systemic JIA [5,8] (Figure 3).
Figure 3.
Clinical sevrity and antibody responces from viral entry to the convalescent stage. Clinical symptoms of a systemic viral infection begin to appear when toxic substances from the focus abruptly spread systemically via viremia. Inflammatory intensities during the clinical course vary among individuals presented as mild, moderate, and severe disease. Patients with severe disease have a higher degree of inflammatory response and may reach the threshold for target organ destruction faster, but reach the peak stage of inflammation later than milder diseases. Total imunoglobulins, including virus specific IgM and IgG, begin to increase at the peak stage of the disease, suggesting that immunoglobulins work for recovery reaction of the disease (see 4.3 Antibodies in viral infections).
Figure 3.
Clinical sevrity and antibody responces from viral entry to the convalescent stage. Clinical symptoms of a systemic viral infection begin to appear when toxic substances from the focus abruptly spread systemically via viremia. Inflammatory intensities during the clinical course vary among individuals presented as mild, moderate, and severe disease. Patients with severe disease have a higher degree of inflammatory response and may reach the threshold for target organ destruction faster, but reach the peak stage of inflammation later than milder diseases. Total imunoglobulins, including virus specific IgM and IgG, begin to increase at the peak stage of the disease, suggesting that immunoglobulins work for recovery reaction of the disease (see 4.3 Antibodies in viral infections).
After the initial infection, viruses can reside for a long time or persistently in certain cells in some patients as asymptomatic carriers or reservoirs. For example, coronavirus or influenza virus outbreaks occur every year with seasonal predominance, and the main affected patients are young children who were not infected previously [75]. Each outbreak may be initiated by carriers whose viruses have adapted to the host. The components, including virions and inflammation-inducing substances, in infected upper respiratory epithelial cells (the focus) are mainly expelled from the host. In contrast, those components directed into the internal side of the host or the components from the infected internal cells, such as lymphoid tissue or other organ tissue cells, might induce more severe clinical manifestations, as previously mentioned. This assumption could explain the differences in clinical phenotypes and target cells in animal coronavirus, influenza virus, and mycoplasma infections [3 8]. Also, chronic viral infections, including HIV, HBV, HCV, and herpes virus groups, including Epstein–Barr virus, cytomegalovirus, and varicella zoster virus, begin with acute systemic inflammatory phenotypes, although clinicians may not be familiar with the acute form of the infections [76,77,78]. Acute hepatitis B has rarely been observed in Korea, probably because of the successful vaccination program and/or adaptation of HBV with improved hygiene over time [79]. Most patients infected with these viruses recover from the diseases, and a small proportion of patients become chronic carriers who have latently infected cells and the stable set point of viral load. The cells in carriers can be intermittently reactivated and produce infective virus particles or possibly viroids and inflammation-inducing substances when infected cells are injured. Some carriers that undergo repeated activation can develop organ failure and/or cancer. The injured cells in the relapsed host may not necessarily be latently infected. For example, HBV could reside in other liver cells or places other than hepatocytes, and hepatocytes can be injured by substances derived from these cells. In addition, hepatocytes injured by immunological insults can release toxic substances that affect neighboring hepatocytes without infection. This phenomenon can be applied to the pathophysiology of chronic viral infections. These acute and chronic natural courses of viruses may represent persistent viruses existing in humans and may be unavoidable processes for the coexistence of viruses with the hosts as strains of the microbiota of humans.
4. Unresolved issues and the PHS hypothesis
The PHS hypothesis has provided new interpretations of unresolved issues in infectious and other diseases. Herein, we discuss more on infectious diseases using the PHS hypothesis.
4.1. Pathologic findings
The nature of disease at the molecular level can be defined as an injury or malfunction of cells in specific organs. The processes of cell injury from initiation to recovery appear as histopathological findings in the tissues of the affected organs. Pathological findings are essential to diagnose the disease, and there is a characteristic infiltration of immune cells, immune proteins, including immunoglobulins and complements, and undetermined immune materials, including amyloid proteins and peptides. Pathological findings from affected organs at any stage of the disease can provide information about the state of inflammation that reflects the relationship between etiological or inflammation-inducing substances and the corresponding immune reaction. Thus, we inferred the etiological substances from the pathological findings based on the PHS hypothesis. For example, if T cells are the predominant cells in early pathologic lesions, the etiologic substances refer to pathogenic peptides, and in IgA-infiltrated pathologic lesions, such as IgA nephropathy, the pathogenic proteins may have a specific size and biochemical characteristics that are suitable for control by IgA but not by IgG or IgM [7]. There are diseases whose pathologic lesions do not show immune cells or immunoglobulins, including minimal change nephrotic syndrome, amyloidosis, Reye syndrome, and chronic neurodegenerative diseases such as prion diseases and Alzheimer’s disease. The etiological substances in these diseases may be very small materials, such as toxic monoamine metabolites or neuropeptides, and are controlled by immune proteins such as prions and amyloid proteins [6,7,9].
In general, there are few intact virions, and occasional positive PCR or monoclonal antibodies acting on fragment of virions are seen in most pathologic findings of severely affected cases as previously mentioned. Moreover, there are similar pathologic findings in fatal cases of ARDS caused by various respiratory viruses, including influenza viruses and corona viruses in SARS, MERS, and COVID-19, but also those caused by nonpathogen-origin insults such as blunt chest trauma, inhalation of toxic gas, severe burn, and amniotic fluid embolism [2]. The findings suggest that inflammation-inducing substances are associated with injured cells caused by infection or physical stress such as trauma, and a common control system of the host act on these conditions [2,3,9].
4.2. Protein homeostasis (proteostasis), proteasome, and lymphopenia in viral infections
All human cells have a genome that potentially produces any protein; however, cells in different organs produce different proteins and express different receptors, as determined by cell fate [80]. Cells in multicellular organisms have structures and functions like those in unicellular eukaryotic organisms, and similar peptidomes and proteosomes exist in the cells of all organisms, including unicellular organisms [81,82]. These findings suggest that peptides are important effectors of cellular biological functions. Protein homeostasis (proteostasis) is important for maintaining cellular metabolic processes and survival. Within cells, proteins are produced and undergo folding into activated forms and degradation as needed. The mechanisms of protein folding, conformational changes, and degradation through the ubiquitin–proteasome system and autophagy-lysosomal pathways have been well described [83,84]. Proteasomes degrade unneeded, misfolded, or damaged proteins by proteolysis and regulate the concentration of proteins using ubiquitin, ubiquitin ligases, chaperones, and co-chaperones. Growing evidence suggests that dysregulation of the ubiquitin–proteasome system or autophagy is related to diseases, including several neurodegenerative, infectious, and inflammatory disorders, and cancers [85,86,87]. Proteasomes are protein complexes found in the nucleus and cytoplasm of eukaryotes, archaea, and some bacteria. Multiple proteasomes are present in a single cell type, and there is variation between proteasome types and tissue cells, suggesting that the proteasome is optimized for each cell function by generating distinct sets of peptides [86]. Since a cell is too small to store all the proteins required to maintain homeostasis, and peptides are not encoded in the genome but play crucial roles in organisms, peptides may be the main effectors of biological tasks in a steady cell environment. Thus, proteosomes may be manufactured to produce peptides required for cellular homeostasis [9]. In addition, cells affected by trauma, infection, or other insults contain numerous peptides that can induce inflammation if released and bind to target cells.
Lymphopenia may be a characteristic of patients with severe pneumonia infected with respiratory pathogens, including influenza viruses, coronaviruses, measles virus, and MP [88,89,90,91]. The severity of lymphopenia correlates with target cell injury in these diseases and chronic viral infections, including AIDS and hepatitis virus infections, when relapsed patients are severely affected [92,93]. Earlier experimental animal studies on respiratory pathogens, including influenza viruses and MP, have reported that extensive infiltration of small lymphocytes is observed in early lung lesions within 2–3 days after inoculation, before the appearance of virus-specific antibodies, irrespective of inoculation sites, such as intraperitoneal, oral, or intranasal routes [1]. In addition, autopsy findings of patients with severe ARDS and experimental animals infected with influenza viruses show lymphocyte depletion in whole lymphoid tissues [1,94]. These findings suggest that lymphopenia results from extensive and/or persistent lymphocyte use in peripheral pathologic lesions, and that T cells may be associated with toxic peptide substances that bind to the receptors of target cells in the lungs in the PHS hypothesis. Lymphopenia in chronic viral infections, including HIV, may also be associated with T cell consumption against other tissue cell injuries caused by substances derived from the focus or latently infected cells. Although HIV is believed to reside in CD4+ T cells, causing lymphopenia in AIDS, T cell dynamics occur during the clinical course of HIV/AIDS [17,95], and other conditions can show persistent CD4+ lymphopenia without AIDS [96]. There may be a limitation in the numerical capacity of the host immune system to mobilize immune cells against these substances to counter extensive or persistent target cell injury in immune-competent patients. Naturally, patients with underlying diseases, malnutrition, or immunodeficiency have a limited repertoire or function of immune cells. In addition, because certain underlying diseases in these hosts can be controlled by immune cells, it is possible that a new infection can aggravate underlying organ diseases, organ failure, and even death [3].
4.3. Antibodies in viral infections
The types of pathogen-origin substances, including pathogenic proteins and peptides, differ according to the pathogen species. In viral infections, they are significantly different according to the presenting methods, whether they are produced through the classical APC-presenting pathway or directly released from infected cells. For example, markedly increased levels of immunoglobulins, especially IgG, are observed at the convalescent stage of systemic infectious diseases, including scarlet fever and tuberculosis, and immune-mediated diseases including acute rheumatic fever and KD [97,98]. However, the proportion of pathogen-specific antibodies among the increased immunoglobulins is very low. Additionally, one or more pathogen-specific antibodies are used to diagnose diseases. Generally, pathogen-specific IgM antibodies are produced first, followed by specific IgG and IgA antibodies in viral infections and intracellular infections such as chlamydia, rickettsia, possibly mycoplasma, and intracellular bacteria. In addition, there is a time gap of several days or longer between the onset of symptoms and the appearance of pathogen-specific antibodies. It was reported that some patients who had pre-existing neutralizing antibodies could be infected with the H1N1 pandemic influenza virus [3], and those who received two-dosed measles-mumps-rubella (MMR) vaccines could be infected with measles or mumps and had neutralizing antibodies at the time of the symptom onset, suggesting that not only pathogen-specific antibodies but also other immune components may be needed to gain long-term immunity [3]. The theory of antibody-dependent enhancement (ADE) has been proposed to play a role in the pathogenesis of viral infections, including dengue fever and COVID-19 [3,99]. Patients with pre-existing or higher levels of pathogen-specific antibodies seem to have a more severe clinical course. This phenomenon may be explained that substances that exacerbate clinical symptoms are not associated with the viruses, and virus-specific antibodies do not control toxic substances. Toxic substances may not be pathogen-origin proteins but other substances, including pathogenic proteins or peptides from injured or infected-host cells [3]. Patients with more severe pneumonia are more lately seroconverted to intracellular pathogen infections, including MERS, COVID-19, and MP infection [100,101,102]. This finding also suggests that the pathogen may not directly induce inflammation during the early stages of severe infection. Thus, it is possible that clinical improvement of the disease begins at the peak stage of inflammation, and immunoglobulins, together with pathogen-specific antibodies, also begin to increase at the peak stage. These findings suggest that each class of immunoglobulin may be involved in recovery reactions by acting against diverse pathogenic proteins produced during the early stage of disease (Figure 3).
Organ-specific autoimmune or autoantibody-related diseases involve various organs, including the CNS, liver, and kidneys [9,103,104]. Antibodies that react to components within self-cells, the autoantibodies, may be produced against protein antigens from injured self-cells and detected in the blood and/or pathological lesions. Autoantibodies are commonly observed in immune-mediated diseases, including SLE, RA, and JIA, as well as in autoantibody-related diseases. By contrast, autoantibodies can appear in healthy individuals, patients in long-term remission, and patients with cancers such as teratoma or lung carcinoma. Moreover, some autoantibodies are detected in clinically diverse immune-mediated diseases, and multiple autoantibodies can appear in a single autoimmune disease. Although autoantibodies are believed to be associated with the pathogenesis of diseases, according to the PHS hypothesis, specific antibodies or T cell clones acting against antigens derived from self-cells are not the cause of the disease; instead, they represent a preventive immune response to possibly toxic substances derived from self-cells [6]. The appearance of autoantibodies could be related to target cell injury associated with unknown pathogenic infections or conditions.
4.4. Unresolved issues in the diagnosis of infectious disease
Rapid diagnosis of infectious diseases, particularly bacterial diseases, is crucial for deciding the treatment options for patient care. The culture method is the “confirmative method” for pathogen infection; however, obtaining results takes a long time. Thus, various PCR assays have replaced culture methods. Pathogen culture and PCR assays depend on the notion that pathogens are responsible for tissue cell injury during infectious diseases. Although PCR assays are used in clinical practice for the early diagnosis of the disease, there are some confounding factors. The results of PCR assays are influenced by disease stage, patient age, sampling site, causative strain variants, and technical errors [8,105]. Indeed, patients with severe pneumonia with MP infection, influenza, or COVID-19 have shown negative PCR results [8,105,106,107], suggesting that the viral load can be low or absent at upper respiratory tract in severely affected patients even in the early stages of the disease. Moreover, a PCR-positive result obtained from the upper respiratory tract or even bronchoalveolar lavage fluids does not confirm the causal agent of pneumonia, especially bacterial infections such as S. pneumonia and H. influenzae, because there are many asymptomatic carriers during the epidemic of these pathogens. Accordingly, pneumonia caused by other pathogens or events can be diagnosed as an infection with epidemic pathogens during epidemics of influenza, COVID-19, and MP infection (false PCR positive).
Currently, there are no reliable serologic methods for the early confirmative diagnosis of infectious diseases because pathogen-specific antibodies are detected at least 3–4 days after disease onset. In addition, patients with severe pneumonia tend to show late seroconversion (false negatives), as previously discussed. In addition, the levels of patho-gen-specific IgM (and IgG) can vary among patients and can be detected long after the initial infection, as shown in MP infection (false positive). Additionally, the results of serological diagnostics may differ if a diagnostic kit targets different antigen(s) among the pathogen fragments. Although there are limitations to PCR assays and serologic tests, paired or repeated serologic tests in the short-term period (during hospitalization) confirming seroconversion (IgM-negative to IgM-positive) and/or increase in pathogen-specific IgM and IgG, including IgG seroconversion, is a reliable test for confirmation of intracellular infectious pathogens, including viruses and MP [108,109].
4.5. Unresolved issues in vaccines
Vaccines have been developed for the prevention of spreading of infectious diseases. As for viral diseases, inactivated or killed viruses, attenuated live viruses, viral proteins, and viral nucleic acids such as DNA and mRNA have been used for vaccine development [110]. The attenuated live vaccines against measles virus, mumps virus, rubella virus, and rotavirus are very effective and have reduced the incidence of the diseases markedly, although there are intermittent outbreaks after vaccination occur over time [111,112]. Clinical application of whole killed bacterial vaccines against diphtheria, tetanus, typhoid fever, and MP have failed because of severe systemic immune reactions owing to extensive immunogenic and/or pathogenic substances originating from these pathogens compared to viruses [113]. There are many viral infections that need more effective vaccines, including HIV and Zika virus [114]. In general, vaccines against RNA viruses, including SARS-CoV-2, influenza viruses, and HIV are less effective because the viruses can easily exchange genomic RNA within a host and possibly across species. Also, viruses have evolved many immunomodulatory strategies to counter host’s innate and adaptive immune responses to viral infection.
It is an interesting issue whether vaccines can induce milder clinical symptoms and/or prevent from severe pneumonia and other organ cell injury. Theoretically, an ideal vaccine for prevention the spread of disease and mitigation of symptoms should have two functions: blocking the entrance of viruses into cells and inhibiting replication processes in the host cells, since etiological or inflammation-inducing substances are produced in the focus in the PHS hypothesis. Killed virus vaccine induce only immune reactions against virus-origin substances, including virus specific antibodies and T cell clones against virus-origin proteins and peptides. Vaccines targeting viral capsid proteins, including viral protein vaccines or vaccines using viral mRNA targeting viral capsid proteins, can elicit virus specific antibodies. Those vaccines induce a virus-neutralization in vitro and could block the viral entry, but the antibodies are not involved in the replication process. Whereas immune systems of the host who experience a natural virus infection react against all virus-origin substances produced during replication process, including genomic and mRNAs, DNAs, and polyproteins, and other diverse host cell-origin substances. Thus, immune status of previously infected host could more effectively act on viral entry and control all toxic substances from infectious insults if reinfection occurs. Likewise, it is possible that attenuated viruses could replicate in certain cells even though not completely as natural viruses; attenuated virus infected cells produce less amounts of substances that can be controlled easily and memorized by immune cells. Also, it is possible that certain attenuated live strains could substitute pre-existing strains in microbiota over time. Thus, vaccinees could experience asymptomatic or milder clinical course when infected during natural virus epidemics. Immune status of the host against diverse substances in infected cells may be responsible for protection of the disease and attenuation of clinical symptoms.
4.6. Viruses and cancers
It is well known that viral and possibly intracellular bacterial infections could be associated with cancers. The oncogenic viruses include HBV and HCV (hepatocellular carcinoma), human papillomaviruses (cervix cancer), Epstein-Barr virus (head and neck cancer), and HIV (Kaposi's sarcoma) [115,116,117]. These viruses may have latently infected cells after initial infection, and repeated reactivation of the viruses induce target cell injury in same organs. The organ cell injury could be caused by cytopathic effect of viruses or by toxic substances derived from the latently infected cells. The injured organ cells should be replaced new ones by host’s repair system.
In the PHS hypothesis, a cancer cell is caused by a gene defect which codes an essential protein for cell survival. A cancer cell is made during process of the proteostasis coping with missing protein [8]. Established cancer cell could be regarded as a new biosystem and show uncontrollable proliferation and perform persistent activation of new genes for proteostasis [8]. All cells present in humans are continuously regenerating and are replaced to renew tissues and might have ability to become cancer cells. It is known that somatic cells may have limitations in the number of cell divisions, and that they can undergo together with telomere shortening [118]. Because cancers occur predominantly in older persons and show a proportional increase to increasing ages, “old” somatic cells that have undergone many division cycles might be fragile enough to a gene mutation and subsequently easy to become cancer cell. Also, the incidence of cancer differs according to each organ, and each organ cell may have different replacement cycle although the mechanisms and the sites of renewal in specific organs during steady state are unknown. Likewise, it is possible that somatic cells that are repeatedly or continuously affected by chronic infection state experience more renewal with shortened turn-over time. The more regenerated cells become vulnerable to a gene defect and a higher risk of transforming to cancer cells over time.
The epidemiological and clinical characteristics of cancers may be changed by environmental factors as well as environment-associated diseases such as obesity, type II diabetes, and inflammatory bowel disease. In Korea, inflammatory bowel disease in children and colorectal cancer in adults were rare diseases a generation ago, but the incidence of both diseases has been increased over time, probably owing to changed diets to westernized style [119]. Additionally, epidemiology of colorectal cancer in Western countries is changing as increased occurrence in young adults [120]. We have proposed that major environment factor for susceptibility of infection-related immune mediated disease, including KD, MIS-C, and JIA is the composition of strains in microbiota, which is affected by ages, sex, and ethnic groups and can be changed by diets as previously discussed. Also, the incidence of some organ cancers could differ by ages, sex, and ethnic groups, suggesting that some cancers might be associated with chronic pathogen infections, including unidentified viruses in microbiota.
Besides old age and infectious insults, there are other factors that can directly affect oncogenic gene defect, and they include genetic diseases, immunodeficiency, radiation such as ultraviolet rays and X-rays, chemicals, and drugs. It is possible that the cells chronically affected in these conditions may also be associated with rapid turn-over time of re-placement with higher risk of cancer transformation.
Cancer cells are dependent on nutrients from the host and use normal host cells such as vascular and stromal cells for their living and growth, thus it possible that they use host’s immune cells when they are infected with intracellular infections or physically stressed. Although components of immune system, such as cytotoxic T cells and antibodies for tumor antigens, NK cells, interferons, and other immune materials could attack cancer cells in vitro and in vivo, many unresolved issues also remain in tumor immunology. Cancer cells also contain various inflammation-inducing substances, and some of them are different to normal cells.
5. Treatment of virus infections
5.1. Antimicrobials
Because infectious diseases are caused by etiological agents such as bacteria, viruses, fungi, protozoa, and parasites, effective drugs against each infectious agent have been developed and widely used. Early antimicrobial therapy, including antibiotics, antifungals, and antivirals, for pathogen-associated systemic diseases is critical to reduce the number of pathogens and pathogen-originated substances, thereby inducing early recovery from the disease. Antiviral drugs are based on targeting the viral lifecycle and/or directing the host cellular factors, since virus replication is performed within cells and the interaction of host cell is essential for virus life. On the other hand, antibiotics are more effective on extracellular pathogen infections, in cases of pneumonia and localized skin infections caused by extracellular pathogens since they can directly act to pathogens.
Besides appearance of antimicrobial-resistance strains, the use of antimicrobials is not always successful in patients with the infectious diseases, especially intracellular pathogens. For example, antivirals cannot prevent from developing pneumonia and progression of pneumonia, and some patients are shown to be rapidly progressive to ARDS despite early antiviral treatment in influenza and possibly COVID-19 [121,122]. Some pneumonia patients with influenza, COVID-19, or other pathogens in developed countries, especially elderly patients with underlying diseases, experience treatment failure with a high mortality, despite early application of antimicrobials [123,124]. In chronic viral infections, including HIV and HBV infections, some patients can be affected with reactivation of the diseases during long-term antiviral therapy [125,126]. In bacterial infections, some patients with MP pneumonia show progressive pneumonia despite early treatment with adequate antibiotics [127]. Patients with septic conditions show higher a mortality despite early antibiotic treatment, and some patients show transient deterioration of clinical symptoms following antibiotic treatment. This may be caused by a cytokine storm, characterized by extensive immune cell activation against large amounts of substances produced during the process of bacterial death [128].
As for these findings, the PHS hypothesis suggest that toxic or inflammation-inducing substances involve not only pathogens and pathogen-originated substances, but also other substances derived from injured cells and/or from activated immune status. Also, intracellular organisms, such as some viruses and bacteria, can reside certain cell as latently infected cells, and reactivation of pathogen in the cells produce the toxic substances as like initial infection. Moreover, host’s immune status plays critical roles for controlling of these substances.
5.2. Immune modulators
Immune modulators include corticosteroids, intravenous immunoglobulin (IVIG), and drugs targeting cytokines or their receptors such as interleukin-1 (IL-1), IL-6, and tumor necrosis factor-α (TNF-α). In general, the drugs have used for autoimmune diseases such as RA, SLE, and autoantibody-related diseases in CNS and other organs, and postinfectious systemic inflammatory diseases such as acute rheumatic fever, KD, and MIS-C. Also, corticosteroids have been used in acute infectious diseases, including septic conditions, tuberculous meningitis, typhoid fever, and EB virus infection [129,130]. During the COVID-19 pandemic, a variety of treatment modalities including antivirals, convalescent plasma, biologics targeting IL-1, IL-6, and TNF-α, and other anti-inflammatory drugs has been tried, and corticosteroids such as dexamethasone and methylprednisolone were reported to be effective in reducing mortality of severely affected patients [131,132].
The mechanisms of each immune modulator are somewhat different, but immune modulators reduce inflammation through blocking various inflammation-associated pathways [133]. Although the entire mode of action of corticosteroids and IVIG is unknown, corticosteroids (as analogues of cortisol) and IVIG (as serum IgG) could be regarded as host-origin immune controllers in vivo. It is possible that a host immune system cannot produce them in adequate doses within the short duration of exposure to acute extensive toxic substances from infectious insults. In the PHS hypothesis, corticosteroids induce early establishment of hyperimmune reaction caused by initially acting nonspecific adaptive immune cells against larger amount of etiological or inflammation inducing substances [2]. Also, in rapidly progressive viral infections, there is a critical time to extensive target cell injury by the T cell-induced immunologic insults before reaching to the peak of inflammation. Thus, for patients with ARDS or other acute whole organ-specific diseases with severe lymphopenia, early, as soon as possible, systemic immune modulator treatment before the occurrence of diffuse organ-specific cell injury is critical, especially in previously immune-competent patients. Because the severity of viral pneumonia or other organs is dependent on the loads of inflammation-inducing substances in the early stage and the effect of corticosteroids is dose-dependent, the initial dose of corticosteroid and the schedule of treatment could be determined on an individual basis. As for severe COVID-19, we have recommended early preemptive immune modulators, especially corticosteroids and/or IVIG, and proper treatment modality, which is consist of early, enough dose of cortico-steroids based on severity of the disease, and short-term use with rapid tapering [3,4,134]. The rationale of this treatment modality is based on the PHS hypothesis and our experience with influenza pneumonia, acute bronchiolitis, and MP pneumonia [1,2,135,136].
In HIV, HBV, and HCV infections, there are chronic carriers who do not show any clinical or laboratory abnormal signs, and some patients show a reactivation of virus on occasion, despite antiviral treatment. The risk factor or number of reactivations remain to be elucidated but might be dependent on host’s immune status, including long-term use of immunosuppressants [137]. Clinically these reactivations can vary, ranging from a subclinical course to a severe acute organ exacerbation such as fulminant hepatitis and even death as like initial infection. Thus, it is possible that various inflammation-inducing substances that are derived from latently infected cells may be responsible for the immunopathogenesis of reactivated viral infections. Accordingly, short-term immune modulators for severely relapsed patients could be helpful to early recovery from the diseases. Historically, it has been reported that immune modulators, including corticosteroids and interferons, were effective in relapsed or exacerbated patients with HIV, HCV, and HBV [138,139,140].
Each immune components act on each toxic substance (Table 1), and all immune components may be communicated with networks, including cytokines and MHC class I in the PHS hypothesis [6]. Therefore, the effect of immune modulators differs depending on the disease entity and even in similar phenotype of the disease. Corticosteroids are effective the diseases of which pathophysiology are associated with activation of non-specific or immature B cells and T cells or innate immune cells such as eosinophils in RA, SLE, and eosinophilic granulomatosis, although rebound phenomena occur in chronic immune-mediated diseases [141]. On the other hand, the drugs might take a long time to be effective or be ineffective in the disease whose pathophysiology is associated with the innate immune system, such as the immune-protein system acting against idiopathic nephrotic syndrome, amyloidosis, and Alzheimer’s disease [9].
Although outcomes of viral infections dependent on host factors, an early and adequate use of immune modulators could mitigate rapid disease progression, and reduce morbidity, and possibly prevent irreversible total organ destruction especially in rapidly progressive cases even in chronic viral infections. In clinical fields, refractory or severe cases in viral or MP infections and in immune-mediated diseases including, KD, MIS-C, and systemic JIA, might be associated with the duration of disease-onset at presentation (lately presented cases) and/or the initiation time and dose of immune modulators especially in immune-competent cases. Severely affected patients may need early, as soon as possible, higher dose of corticosteroids rather than a guideline-recommended dose, and next treatment modality could be determined based on clinical signs and short-term follow-up inflammatory biomarkers, including lymphopenia, CRP, and albumin [142,143,144].
Although immune modulators induce stabilization of hyperimmune reactions in the early stage in acute insults from infectious disease, they cannot control etiological substances, thereby cannot cure disease. Also, prolonged use of corticosteroids causes serious systemic complications, and prolonged use of other immune modulators and antivirals can also cause rare adverse reactions, including higher risk of infection and various metabolic changes. Patients with autoimmune diseases or chronic inflammatory diseases after infection may have improper immune status for controlling initial exposure to an etiological substance to protect against target cell injury or for control of toxic substances produced by injured target self-cells.
6. Conclusions
The mechanism and pathophysiology of viral infections are complex. The host's defense responses to viral infections that lead to viral clearance may also be associated with injury and destruction of the target organ cells. In this paper, we discuss on the unresolved issues on pathophysiology of viral diseases and propose early immune modulators against acute severe insults from virus infections based on our clinical experiences and the PHS hypothesis. Although understanding of viral diseases and human interactions have advanced, further studies are needed to understand the viral-host response mechanisms to improved preventive methods, therapeutic interventions, and decrease target organ sequelae.
Author Contributions
conceptualization & methodology, K.Y.L; data curation, writing, and editing, K.Y.L, H.M.K.
Funding
None.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, K.-Y.; Rhim, J.-W.; Kang, J.-H. Hyperactive immune cells (T cells) may be responsible for acute lung injury in influenza virus infections: A need for early immune-modulators for severe cases. Med Hypotheses 2011, 76, 64–69. [Google Scholar] [CrossRef]
- Lee, K.-Y. Pneumonia, Acute Respiratory Distress Syndrome, and Early Immune-Modulator Therapy. Int. J. Mol. Sci. 2017, 18, 388. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Rhim, J.-W.; Kang, J.-H. Immunopathogenesis of COVID-19 and early immunomodulators. Clin. Exp. Pediatr. 2020, 63, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y. The solution on enigmas in COVID-19: the protein-homeostasis-system hypothesis. J. Korean Med Assoc. 2020, 63, 366–372. [Google Scholar] [CrossRef]
- Lee, K.-Y.; Rhim, J.-W.; Kang, J.-H. Kawasaki Disease: Laboratory Findings and an Immunopathogenesis on the Premise of a "Protein Homeostasis System". Yonsei Med J. 2012, 53, 262–275. [Google Scholar] [CrossRef]
- Lee, K.-Y. A Common Immunopathogenesis Mechanism for Infectious Diseases: The Protein-Homeostasis-System Hypothesis. Infect. Chemother. 2015, 47, 12–26. [Google Scholar] [CrossRef]
- Lee, K.-Y. A unified pathogenesis for kidney diseases, including genetic diseases and cancers, by the protein-homeostasis-system hypothesis. Kidney Res. Clin. Pr. 2017, 36, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.-W.; Kang, J.-H.; Lee, K.-Y. Etiological and pathophysiological enigmas of severe coronavirus disease 2019, multisystem inflammatory syndrome in children, and Kawasaki disease. Clin. Exp. Pediatr. 2022, 65, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y. Common immunopathogenesis of central nervous system diseases: the protein-homeostasis-system hypothesis. Cell Biosci. 2022, 12, 184. [Google Scholar] [CrossRef]
- Edwards, R.A.; Rohwer, F. Viral metagenomics. Nat. Rev. Microbiol. 2005, 3, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Starokadomskyy, P. Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question. Stud. Hist. Philos. Sci. Part C: Stud. Hist. Philos. Biol. Biomed. Sci. 2016, 59, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, N. On the historical significance of Beijerinck and his contagium vivum fluidum for modern virology. Hist. Philos. Life Sci. 2018, 40, 41. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Rohwer, F. Here a virus, there a virus, everywhere the same virus? Trends Microbiol. 2005, 13, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Virus Taxonomy: 2021 Release". talk.ictvonline.org. International Committee on Taxonomy of Viruses. Retrieved 18 June 2023.
- Chauhan, R.P.; Gordon, M.L. An overview of influenza A virus genes, protein functions, and replication cycle highlighting important updates. Virus Genes 2022, 58, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Pol, S.; Lagaye, S. The remarkable history of the hepatitis C virus. Microbes Infect. 2019, 21, 263–270. [Google Scholar] [CrossRef]
- Silvestri, G. AIDS pathogenesis: a tale of two monkeys. J. Med Primatol. 2008, 37 (Suppl. 2), 6–12. [Google Scholar] [CrossRef]
- Tompa, D.R.; Immanuel, A.; Srikanth, S.; Kadhirvel, S. Trends and strategies to combat viral infections: A review on FDA approved antiviral drugs. Int. J. Biol. Macromol. 2021, 172, 524–541. [Google Scholar] [CrossRef]
- Kausar, S.; Khan, F.S.; Rehman, M.I.M.U.; Akram, M.; Riaz, M.; Rasool, G.; Khan, A.H.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef] [PubMed]
- Moelling, K.; Broecker, F. Viroids and the Origin of Life. Int. J. Mol. Sci. 2021, 22, 3476. [Google Scholar] [CrossRef] [PubMed]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar]
- de Vries, R.D.; Mesman, A.W.; Geijtenbeek, T.B.; Duprex, W.P.; de Swart, R.L. The pathogenesis of measles. Curr. Opin. Virol. 2012, 2, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.-L.; McGinley, J.P.; Drysdale, S.B.; Pollard, A.J. Epidemiology and Immune Pathogenesis of Viral Sepsis. Front. Immunol. 2018, 9, 2147. [Google Scholar] [CrossRef] [PubMed]
- Bösmüller, H.; Matter, M.; Fend, F.; Tzankov, A. The pulmonary pathology of COVID-19. Virchows Arch. 2021, 478, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Delorey, T.M.; Ziegler, C.G.K.; Heimberg, G.; Normand, R.; Yang, Y.; Segerstolpe, Å.; Abbondanza, D.; Fleming, S.J.; Subramanian, A.; Montoro, D.T.; et al. COVID-19 tissue atlases reveal SARS-CoV-2 pathology and cellular targets. Nature 2021, 595, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Johansson, A.; Mohamed, M.S.; Moulin, T.C.; Schiöth, H.B. Neurological manifestations of COVID-19: A comprehensive literature review and discussion of mechanisms. J. Neuroimmunol. 2021, 358, 577658. [Google Scholar] [CrossRef]
- Weiss, R.A. Human endogenous retroviruses: friend or foe? APMIS 2016, 124, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Dsouza, J.M.; Mathew, J.L. Comparison of microbiota in the upper versus lower respiratory tract in children during health and respiratory disease: protocol for a systematic review. Syst. Rev. 2021, 10, 253. [Google Scholar] [CrossRef]
- Virgin, H.W. The Virome in Mammalian Physiology and Disease. Cell 2014, 157, 142–150. [Google Scholar] [CrossRef]
- Clinton, N.A.; Hameed, S.A.; Agyei, E.K.; Jacob, J.C.; Oyebanji, V.O.; Jabea, C.E. Crosstalk between the Intestinal Virome and Other Components of the Microbiota, and Its Effect on Intestinal Mucosal Response and Diseases. J. Immunol. Res. 2022, 2022, 7883945. [Google Scholar] [CrossRef] [PubMed]
- Citti, C.; Blanchard, A. Mycoplasmas and their host: emerging and re-emerging minimal pathogens. Trends Microbiol. 2013, 21, 196–203. [Google Scholar] [CrossRef]
- Ezzatpour, S.; Portocarrero, A.d.C.M.; Cardelle-Cobas, A.; Lamas, A.; López-Santamarina, A.; Miranda, J.M.; Aguilar, H.C. The Human Gut Virome and Its Relationship with Nontransmissible Chronic Diseases. Nutrients 2023, 15, 977. [Google Scholar] [CrossRef] [PubMed]
- Iorio, A.; Biazzo, M.; Gardini, S.; Muda, A.O.; Perno, C.F.; Dallapiccola, B.; Putignani, L. Cross-correlation of virome–bacteriome–host–metabolome to study respiratory health. Trends Microbiol. 2021, 30, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- O’donnell, M.P.; Fox, B.W.; Chao, P.-H.; Schroeder, F.C.; Sengupta, P. A neurotransmitter produced by gut bacteria modulates host sensory behaviour. Nature 2020, 583, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Borkenhagen, L.K.; Salman, M.D.; Ma, M.-J.; Gray, G.C. Animal influenza virus infections in humans: A commentary. Int. J. Infect. Dis. 2019, 88, 113–119. [Google Scholar] [CrossRef]
- Schwartz, D.A. An Analysis of 38 Pregnant Women With COVID-19, Their Newborn Infants, and Maternal-Fetal Transmission of SARS-CoV-2: Maternal Coronavirus Infections and Pregnancy Outcomes. Arch. Pathol. Lab. Med. 2020, 144, 799–805. [Google Scholar] [CrossRef]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.-S.; Iliopoulos, D.; et al. Origins of theE. coliStrain Causing an Outbreak of Hemolytic–Uremic Syndrome in Germany. N. Engl. J. Med. 2011, 365, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Burgner, D.; Lee, H.-S.; Hong, J.-H.; Lee, M.-H.; Kang, J.-H.; Lee, B.-C. The changing epidemiology of pediatric aseptic meningitis in Daejeon, Korea from 1987 to 2003. BMC Infect. Dis. 2005, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; Lee, K.-Y.; Kim, C.-H.; Sim, D. Changing hepatitis A epidemiology and the need for vaccination in Korea. Asian Pac. J. Allergy Immunol. 2004, 22, 237–242. [Google Scholar]
- El-Shabasy, R.M.; Nayel, M.A.; Taher, M.M.; Abdelmonem, R.; Shoueir, K.R.; Kenawy, E.R. Three waves changes, new variant strains, and vaccination effect against COVID-19 pandemic. Int. J. Biol. Macromol. 2022, 204, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y. New Insights for Febrile Urinary Tract Infection (Acute Pyelonephritis) in Children. Child. Kidney Dis. 2016, 20, 37–44. [Google Scholar] [CrossRef]
- Lee, H.; Yun, K.W.; Lee, H.J.; Choi, E.H. Antimicrobial therapy of macrolide-resistantMycoplasma pneumoniaepneumonia in children. Expert Rev. Anti-infective Ther. 2018, 16, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kil, H.-R.; Yu, J.-W.; Lee, S.-C.; Rhim, J.-W.; Lee, K.-Y. Changes in clinical and laboratory features of Kawasaki disease noted over time in Daejeon, Korea. Pediatr. Rheumatol. 2017, 15, 60. [Google Scholar] [CrossRef] [PubMed]
- Kuem, S.-W.; Hur, S.-M.; Youn, Y.-S.; Rhim, J.-W.; Suh, J.-S.; Lee, K.-Y. Changes in Acute Poststreptococcal Glomerulonephritis: An Observation Study at a Single Korean Hospital Over Two Decades. Child. Kidney Dis. 2015, 19, 112–117. [Google Scholar] [CrossRef]
- Rhim, J.-W.; Lee, Y.-T.; Kang, H.-M.; Suh, J.-S.; Lee, K.-Y. Changes in clinical features in Henoch-Schönlein purpura during three decades: an observational study at a single hospital in Korea. Clin. Rheumatol. 2019, 38, 2811–2818. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Hamada, H.; Arvonen, M. Editorial: Infection-Related Immune-Mediated Diseases and Microbiota. Front. Pediatr. 2020, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed]
- A Kennedy, E.; Holtz, L.R. Gut virome in early life: origins and implications. Curr. Opin. Virol. 2022, 55, 101233–101233. [Google Scholar] [CrossRef] [PubMed]
- Fallani, M.; Young, D.; Scott, J.; Norin, E.; Amarri, S.; Adam, R.; Aguilera, M.; Khanna, S.; Gil, A.; Edwards, C.A.A.; et al. Intestinal Microbiota of 6-week-old Infants Across Europe: Geographic Influence Beyond Delivery Mode, Breast-feeding, and Antibiotics. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Beyrer, C.; Baral, S.D.; van Griensven, F.; Goodreau, S.M.; Chariyalertsak, S.; Wirtz, A.L.; Brookmeyer, R. Global epidemiology of HIV infection in men who have sex with men. Lancet 2012, 380, 367–377. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Chaabna, K.; Abu-Raddad, L.J. The Epidemiology of Hepatitis C Virus in the Fertile Crescent: Systematic Review and Meta-Analysis. PLOS ONE 2015, 10, e0135281. [Google Scholar] [CrossRef]
- Yasumura, J.; Yashiro, M.; Okamoto, N.; Shabana, K.; Umebayashi, H.; Iwata, N.; Okura, Y.; Kubota, T.; Shimizu, M.; Tomiita, M.; et al. Clinical features and characteristics of uveitis associated with juvenile idiopathic arthritis in Japan: first report of the pediatric rheumatology association of Japan (PRAJ). Rheumatol. 2019, 17, 15. [Google Scholar] [CrossRef]
- Rhim, J.-W.; Kang, H.M.; Han, J.-W.; Lee, K.-Y. A Presumed Etiology of Kawasaki Disease Based on Epidemiological Comparison With Infectious or Immune-Mediated Diseases. Front. Pediatr. 2019, 7, 202. [Google Scholar] [CrossRef] [PubMed]
- Bouayad, A. Multifaceted roles of Fcγ receptors in COVID-19 and vaccine responses. Am. J. Transl. Res. 2023, 15, 3040–3059. [Google Scholar] [PubMed]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Katze, M.G.; He, Y.; Gale, M., Jr. Viruses and interferon: a fight for supremacy. Nat. Rev. Immunol. 2002, 2, 675–687. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-Hussaniy, H.A.; Al-Harcan, N.A.H.; Alexiou, A.; Batiha, G.E.-S. Neutrophil Extracellular Traps (NETs) and Covid-19: A new frontiers for therapeutic modality. Int. Immunopharmacol. 2022, 104, 108516. [Google Scholar] [CrossRef] [PubMed]
- Pires, B.G.; Calado, R.T. Hyper-inflammation and complement in COVID-19. Am. J. Hematol. 2023, 98 (Suppl. 4), S74–S81. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Quintin, J.; van der Meer, J.W. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Quintin, J. Innate immune memory in mammals. Semin. Immunol. 2016, 28, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Tso, F.Y.; Lidenge, S.J.; Poppe, L.K.; Peña, P.B.; Privatt, S.R.; Bennett, S.J.; Ngowi, J.R.; Mwaiselage, J.; Belshan, M.; Siedlik, J.A.; et al. Presence of antibody-dependent cellular cytotoxicity (ADCC) against SARS-CoV-2 in COVID-19 plasma. PLOS ONE 2021, 16, e0247640. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zuo, X.; Zhang, S.; Ouyang, Z.; Jiang, S.; Wang, F.; Wang, G. The Mechanism behind Influenza Virus Cytokine Storm. Viruses 2021, 13, 1362. [Google Scholar] [CrossRef]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Afzali, B.; Noris, M.; Lambrecht, B.N.; Kemper, C. The state of complement in COVID-19. Nat. Rev. Immunol. 2022, 22, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Potere, N.; Del Buono, M.G.; Caricchio, R.; Cremer, P.C.; Vecchié, A.; Porreca, E.; Gasperina, D.D.; Dentali, F.; Abbate, A.; Bonaventura, A. Interleukin-1 and the NLRP3 inflammasome in COVID-19: Pathogenetic and therapeutic implications. EBioMedicine 2022, 85, 104299. [Google Scholar] [CrossRef]
- Alfaro, E.; Díaz-García, E.; García-Tovar, S.; Zamarrón, E.; Mangas, A.; Galera, R.; López-Collazo, E.; García-Rio, F.; Cubillos-Zapata, C. Upregulated Proteasome Subunits in COVID-19 Patients: A Link with Hypoxemia, Lymphopenia and Inflammation. Biomolecules 2022, 12, 442. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Colley, L.; Agbaedeng, T.A.; Ellison-Hughes, G.M.; Ross, M.D. Vascular Manifestations of COVID-19 – Thromboembolism and Microvascular Dysfunction. Front. Cardiovasc. Med. 2020, 7, 598400. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodpoor, A.; Sanaie, S.; Roudbari, F.; Sabzevari, T.; Sohrabifar, N.; Kazeminasab, S. Understanding the role of telomere attrition and epigenetic signatures in COVID-19 severity. Gene 2022, 811, 146069. [Google Scholar] [CrossRef]
- Johnson, B.S.; Laloraya, M. A cytokine super cyclone in COVID-19 patients with risk factors: the therapeutic potential of BCG immunization. Cytokine Growth Factor Rev. 2020, 54, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Heimdal, I.; Moe, N.; Krokstad, S.; Christensen, A.; Skanke, L.H.; Nordbø, S.A.; Døllner, H. Human Coronavirus in Hospitalized Children With Respiratory Tract Infections: A 9-Year Population-Based Study From Norway. J. Infect. Dis. 2019, 219, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Robb, M.L.; Eller, L.A.; Kibuuka, H.; Rono, K.; Maganga, L.; Nitayaphan, S.; Kroon, E.; Sawe, F.K.; Sinei, S.; Sriplienchan, S.; et al. Prospective Study of Acute HIV-1 Infection in Adults in East Africa and Thailand. N. Engl. J. Med. 2016, 374, 2120–2130. [Google Scholar] [CrossRef]
- Indolfi, G.; Easterbrook, P.; Dusheiko, G.; Siberry, G.; Chang, M.-H.; Thorne, C.; Bulterys, M.; Chan, P.-L.; El-Sayed, M.H.; Giaquinto, C.; et al. Hepatitis B virus infection in children and adolescents. Lancet Gastroenterol. Hepatol. 2019, 4, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Indolfi, G.; Easterbrook, P.; Dusheiko, G.; El-Sayed, M.H.; Jonas, M.M.; Thorne, C.; Bulterys, M.; Siberry, G.; Walsh, N.; Chang, M.-H.; et al. Hepatitis C virus infection in children and adolescents. Lancet Gastroenterol. Hepatol. 2019, 4, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.Y.; Kim, J.H. The epidemiology of hepatitis B virus infection in Korea. Korean J. Intern. Med. 2019, 34, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Casey, M.J.; Stumpf, P.S.; MacArthur, B.D. Theory of cell fate. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1471. [Google Scholar] [CrossRef] [PubMed]
- Lyapina, I.; Ivanov, V.; Fesenko, I. Peptidome: Chaos or Inevitability. Int. J. Mol. Sci. 2021, 22, 13128. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Yang, C.; Castro, L.M.; Tashima, A.K.; Ferro, E.S.; Moir, R.D.; Willis, I.M.; Fricker, L.D. Analysis of the Yeast Peptidome and Comparison with the Human Peptidome. PLOS ONE 2016, 11, e0163312. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, G.G.; Hipp, M.S.; Hartl, F.U. Functional Modules of the Proteostasis Network. Cold Spring Harb. Perspect. Biol. 2020, 12, a033951. [Google Scholar] [CrossRef] [PubMed]
- Hommen, F.; Bilican, S.; Vilchez, D. Protein clearance strategies for disease intervention. J. Neural Transm. 2022, 129, 141–172. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhong, M.B.; Toro, C.A.; Zhang, L.; Cai, D. Endo-lysosomal pathway and ubiquitin-proteasome system dysfunction in Alzheimer's disease pathogenesis. Neurosci. Lett. 2019, 703, 68–78. [Google Scholar] [CrossRef]
- Morozov, A.V.; Karpov, V.L. Proteasomes and Several Aspects of Their Heterogeneity Relevant to Cancer. Front. Oncol. 2019, 9, 761. [Google Scholar] [CrossRef]
- Verbrugge, S.E.; Scheper, R.J.; Lems, W.F.; de Gruijl, T.D.; Jansen, G. Proteasome inhibitors as experimental therapeutics of autoimmune diseases. Arthritis Res. Ther. 2015, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Talnir, R.; Lavid, O.; Rubinstein, U.; Niven, M.; First, Y.; I Tsivion, A.J.; Schachter, Y. Transient lymphopenia and neutropenia: pediatric influenza A/H1N1 infection in a primary hospital in Israel. Isr. Med Assoc. J. : IMAJ.
- Yan, S.; Wu, G. Is lymphopenia different between SARS and COVID-19 patients? FASEB J. 2021, 35, e21245. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Kobune, F.; Sato, T.A.; Kohama, T.; Takeuchi, Y.; Abe, T.; Takayama, N.; Tsuchiya, T.; Tashiro, M. Extensive lymphopenia due to apoptosis of uninfected lymphocytes in acute measles patients. Arch. Virol. 2000, 145, 905–920. [Google Scholar] [CrossRef]
- Lee, K.-Y.; Lee, H.-S.; Hong, J.-H.; Lee, M.-H.; Lee, J.-S.; Burgner, D.; Lee, B.-C. Role of prednisolone treatment in severeMycoplasma pneumoniae pneumonia in children. Pediatr. Pulmonol. 2006, 41, 263–268. [Google Scholar] [CrossRef]
- Lerner, C.W.; Tapper, M.L. Opportunistic Infection Complicating Acquired Immune Deficiency Syndrome. Clinical features of 25 cases. Medicine 1984, 63, 155–164. [Google Scholar] [CrossRef]
- Lai, Y.W.; Tan, T.-C. Atypical presentation of Good syndrome: acute hepatitis from hepatitis B virus reactivation. Asia Pac. Allergy 2020, 10, e37. [Google Scholar] [CrossRef]
- Tumpey, T.M.; Lu, X.; Morken, T.; Zaki, S.R.; Katz, J.M. Depletion of Lymphocytes and Diminished Cytokine Production in Mice Infected with a Highly Virulent Influenza A (H5N1) Virus Isolated from Humans. J. Virol. 2000, 74, 6105–6116. [Google Scholar] [CrossRef] [PubMed]
- Douek, D.C.; Picker, L.J.; Koup, R.A. T Cell Dynamics in HIV-1 Infection. Annu. Rev. Immunol. 2003, 21, 265–304. [Google Scholar] [CrossRef] [PubMed]
- Lisco, A.; Ortega-Villa, A.M.; Mystakelis, H.; Anderson, M.V.; Mateja, A.; Laidlaw, E.; Manion, M.; Roby, G.; Higgins, J.; Kuriakose, S.; et al. Reappraisal of Idiopathic CD4 Lymphocytopenia at 30 Years. New Engl. J. Med. 2023, 388, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Lee, J.-S. Immunoglobulin G has a role for systemic protein modulation in vivo: A new concept of protein homeostasis. Med Hypotheses 2006, 67, 848–855. [Google Scholar] [CrossRef]
- Han, J.-W.; Oh, J.-H.; Rhim, J.-W.; Lee, K.-Y. Correlation between elevated platelet count and immunoglobulin levels in the early convalescent stage of Kawasaki disease. Medicine 2017, 96, e7583. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Pillat, M.M.; Tárnok, A. Dengue Fever,COVID-19 (SARS-CoV-2), andAntibody-DependentEnhancement (ADE): A Perspective. Cytom. Part A 2020, 97, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Müller, M.A.; Seok, H.; Park, G.E.; Lee, J.Y.; Cho, S.Y.; Ha, Y.E.; Baek, J.Y.; Kim, S.H.; Kang, J.-M.; et al. Serologic responses of 42 MERS-coronavirus-infected patients according to the disease severity. Diagn. Microbiol. Infect. Dis. 2017, 89, 106–111. [Google Scholar] [CrossRef]
- Zilla, M.; Wheeler, B.J.; Keetch, C.; Mitchell, G.; McBreen, J.; Wells, A.; Shurin, M.R.; Peck-Palmer, O.; Wheeler, S.E. Variable Performance in 6 Commercial SARS-CoV-2 Antibody Assays May Affect Convalescent Plasma and Seroprevalence Screening. Am. J. Clin. Pathol. 2021, 155, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Youn, Y.-S.; Lee, K.-Y.; Hwang, J.-Y.; Rhim, J.-W.; Kang, J.-H.; Lee, J.-S.; Kim, J.-C. Difference of clinical features in childhood Mycoplasma pneumoniae pneumonia. BMC Pediatr. 2010, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Bien, C.I. Autoimmune encephalitis in children and adolescents. Neurol. Res. Pr. 2020, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Sucher, E.; Sucher, R.; Gradistanac, T.; Brandacher, G.; Schneeberger, S.; Berg, T. Autoimmune Hepatitis—Immunologically Triggered Liver Pathogenesis—Diagnostic and Therapeutic Strategies. J. Immunol. Res. 2019, 2019, 9437043. [Google Scholar] [CrossRef]
- Arevalo-Rodriguez, I.; Buitrago-Garcia, D.; Simancas-Racines, D.; Zambrano-Achig, P.; Del Campo, R.; Ciapponi, A.; Sued, O.; Martinez-García, L.; Rutjes, A.W.; Low, N.; et al. False-negative results of initial RT-PCR assays for COVID-19: A systematic review. PLOS ONE 2020, 15, e0242958. [Google Scholar] [CrossRef] [PubMed]
- Winichakoon, P.; Chaiwarith, R.; Liwsrisakun, C.; Salee, P.; Goonna, A.; Limsukon, A.; Kaewpoowat, Q. Negative Nasopharyngeal and Oropharyngeal Swabs Do Not Rule Out COVID-19. J. Clin. Microbiol. 2020, 58, e00297–20. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, V.; Negro, A.; Pirotti, T.; Trenti, T. Estimate false-negative RT-PCR rates for SARS-CoV-2. A systematic review and meta-analysis. Eur. J. Clin. Investig. 2022, 52, e13706. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-C.; Youn, Y.-S.; Rhim, J.-W.; Kang, J.-H.; Lee, K.-Y. Early Serologic Diagnosis of Mycoplasma pneumoniae Pneumonia: An Observational Study on Changes in Titers of Specific-IgM Antibodies and Cold Agglutinins. Medicine 2016, 95, e3605. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.E.; Kang, H.M.; Yang, E.A.; Han, H.Y.; Han, S.B.; Rhim, J.W.; Lee, K.-Y. Early Confirmation of Mycoplasma pneumoniae Infection by Two Short-Term Serologic IgM Examination. Diagnostics 2021, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.; Denizer, G.; Friedland, L.R.; Krishnan, J.; Shapiro, M. Understanding modern-day vaccines: what you need to know. Ann. Med. 2018, 50, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Lee, H.-S.; Hur, J.-K.; Kang, J.-H.; Lee, B.-C. The changing epidemiology of hospitalized pediatric patients in three measles outbreaks. J. Infect. 2007, 54, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.-U.; Kim, E.-K.; Youn, Y.-S.; Rhim, J.-W.; Lee, K.-Y. Outbreaks of mumps: an observational study over two decades in a single hospital in Korea. Korean J. Pediatr. 2014, 57, 396–402. [Google Scholar] [CrossRef]
- Jiang, Z.; Li, S.; Zhu, C.; Zhou, R.; Leung, P.H.M. Mycoplasma pneumoniae Infections: Pathogenesis and Vaccine Development. Pathogens 2021, 10, 119. [Google Scholar] [CrossRef]
- Morabito, K.M.; Graham, B.S. Zika Virus Vaccine Development. J. Infect. Dis. 2017, 216 (Suppl. 10), S957–S963. [Google Scholar] [CrossRef] [PubMed]
- D'Souza, S.; Lau, K.C.; Coffin, C.S.; Patel, T.R. Molecular mechanisms of viral hepatitis induced hepatocellular carcinoma. World J. Gastroenterol. 2020, 26, 5759–5783. [Google Scholar] [CrossRef]
- Della Fera, A.N.; Warburton, A.; Coursey, T.L.; Khurana, S.; McBride, A.A. Persistent Human Papillomavirus Infection. Viruses 2021, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Safai, B.; Sarngadharan, M. G.; Koziner, B.; Godbold, J.; Myskowski, P. L.; Cunningham-Rundles, S.; et al. Spectrum of Kaposi's sarcoma in the epidemic of AIDS. Cancer Res. 1985, 45(9 Suppl), 4646s-4648s. 2: PMID, 2990. [Google Scholar]
- Rossiello, F.; Jurk, D.; Passos, J.F.; di Fagagna, F.D. Telomere dysfunction in ageing and age-related diseases. Nature 2022, 24, 135–147. [Google Scholar] [CrossRef]
- Lee, J.W.; Eun, C.S. Inflammatory bowel disease in Korea: epidemiology and pathophysiology. Korean J. Intern. Med. 2022, 37, 885–894. [Google Scholar] [CrossRef]
- Saraiva, M.R.; Rosa, I.; Claro, I. Early-onset colorectal cancer: A review of current knowledge. World J. Gastroenterol. 2023, 29, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, T.; Doshi, P. Multisystem failure: the story of anti-influenza drugs. BMJ 2014, 348, g2263. [Google Scholar] [CrossRef]
- Rhim, J.-W.; Lee, K.-Y.; Youn, Y.-S.; Kang, J.-H.; Kim, J.-C. Epidemiological and clinical characteristics of childhood pandemic 2009 H1N1 virus infection: an observational cohort study. BMC Infect. Dis. 2011, 11, 225. [Google Scholar] [CrossRef]
- Garcia-Vidal, C.; Carratalà, J. Early and Late Treatment Failure in Community-Acquired Pneumonia. Semin. Respir. Crit. Care Med. 2009, 30, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Menendez, R.; Torres, A.; Zalacaín, R.; Aspa, J.; Villasclaras, J.J.M.; Borderías, L.; Moya, J.M.B.; Ruiz-Manzano, J.; de Castro, F.R.; Blanquer, J.; et al. Risk factors of treatment failure in community acquired pneumonia: implications for disease outcome. Thorax 2004, 59, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Bang, K.B.; Kim, H.J. Management of antiviral drug resistance in chronic hepatitis B. World J. Gastroenterol. 2014, 20, 11641–11649. [Google Scholar] [CrossRef] [PubMed]
- Endalamaw, A.; Mekonnen, M.; Geremew, D.; Yehualashet, F.A.; Tesera, H.; Habtewold, T.D. HIV/AIDS treatment failure and associated factors in Ethiopia: meta-analysis. BMC Public Heal. 2020, 20, 82. [Google Scholar] [CrossRef] [PubMed]
- Youn, Y.-S.; Lee, K.-Y. Mycoplasma pneumoniaepneumonia in children. Korean J. Pediatr. 2012, 55, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ma, S. The cytokine storm and factors determining the sequence and severity of organ dysfunction in multiple organ dysfunction syndrome. Am. J. Emerg. Med. 2008, 26, 711–715. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, Y.; Tang, J.; Lunsford, L.D.; Li, T.; Tang, R.; He, J.; Xu, P.; Faramand, A.; Xu, J.; et al. Association of Corticosteroid Treatment With Outcomes in Adult Patients With Sepsis: A Systematic Review and Meta-analysis. JAMA Intern. Med. 2019, 179, 213–223. [Google Scholar] [CrossRef]
- Wallis, R.S.; O’garra, A.; Sher, A.; Wack, A. Host-directed immunotherapy of viral and bacterial infections: past, present and future. Nat. Rev. Immunol. 2023, 23, 121–133. [Google Scholar] [CrossRef]
- The RECOVERY Collaborative Group. ; Horby, P.; Lim, W. S.; Emberson, J. R.; Mafham, M.; Bell, J. L.; Linsell, L.; et al. Dexamethasone in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- The WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group; Sterne, J. A.C.; Murthy, S.; Diaz, J.V.; Slutsky, A.S.; Villar, J.; Angus, D.C.; Annane, D.; Azevedo, L.C.P.; Berwanger, O.; et al. Association Between Administration of Systemic Corticosteroids and Mortality Among Critically Ill Patients With COVID-19: A Meta-analysis. JAMA 2020, 324, 1330–1341. [Google Scholar] [CrossRef]
- Rizk, J.G.; Kalantar-Zadeh, K.; Mehra, M.R.; Lavie, C.J.; Rizk, Y.; Forthal, D.N. Pharmaco-Immunomodulatory Therapy in COVID-19. Drugs 2020, 80, 1267–1292. [Google Scholar] [CrossRef]
- Lee, K.-Y.; Rhim, J.-W.; Kang, J.-H. Early preemptive immunomodulators (corticosteroids) for severe pneumonia patients infected with SARS-CoV-2. Clin. Exp. Pediatr. 2020, 63, 117–118. [Google Scholar] [CrossRef]
- Kil, H.-R.; Lee, J.-H.; Lee, K.-Y.; Rhim, J.-W.; Youn, Y.-S.; Kang, J.-H. Early corticosteroid treatment for severe pneumonia caused by 2009 H1N1 influenza virus. Crit. Care 2011, 15, 413–413. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.-A.; Kang, H.-M.; Rhim, J.-W.; Kang, J.-H.; Lee, K.-Y. Early Corticosteroid Therapy for Mycoplasma pneumoniae Pneumonia Irrespective of Used Antibiotics in Children. J. Clin. Med. 2019, 8, 726. [Google Scholar] [CrossRef]
- López-Serrano, P. .; de la Fuente Briongos, E.; Alonso, E.C.; Pérez-Calle, J.L.; Rodríguez, C.F. Hepatitis B and immunosuppressive therapies for chronic inflammatory diseases: When and how to apply prophylaxis, with a special focus on corticosteroid therapy. World J. Hepatol. 2015, 7, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Ewald, H.; Raatz, H.; Boscacci, R.; Furrer, H.; Bucher, H.C.; Briel, M. Adjunctive corticosteroids for Pneumocystis jiroveci pneumonia in patients with HIV infection. Cochrane Database Syst. Rev. 2015, 2015, CD006150. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Zhang, H.-Y.; You, J.-P.; Zhang, X.-Q. Severe acute exacerbation of chronic hepatitis B during pegylated interferon treatment and early intervention with corticosteroid. Virol. J. 2012, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Perelson, A.S. Treatment of Hepatitis C Virus Infection With Interferon and Small Molecule Direct Antivirals: Viral Kinetics and Modeling. Crit. Rev. Immunol. 2010, 30, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Trivioli, G.; Terrier, B.; Vaglio, A. Eosinophilic granulomatosis with polyangiitis: understanding the disease and its management. Rheumatology 2020, 59 (Suppl. 3), iii84–iii94. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-Y.; Lee, K.-Y.; Rhim, J.-W.; Youn, Y.-S.; Oh, J.-H.; Han, J.-W.; Lee, J.-S.; Burgner, D. Assessment of intravenous immunoglobulin non-responders in Kawasaki disease. Arch. Dis. Child. 2011, 96, 1088–1090. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.-M.; Kang, H.-M.; Lee, S.-C.; Yu, J.-W.; Kil, H.-R.; Rhim, J.-W.; Han, J.-W.; Lee, K.-Y. Clinical implications in laboratory parameter values in acute Kawasaki disease for early diagnosis and proper treatment. Korean J. Pediatr. 2018, 61, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Han, H.Y.; Park, K.C.; Yang, E.-A.; Lee, K.-Y. Macrolide-Resistant and Macrolide-Sensitive Mycoplasma pneumoniae Pneumonia in Children Treated Using Early Corticosteroids. J. Clin. Med. 2021, 10, 1309. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



