
Submitted:
31 July 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
Fire is a common disturbance in the Brazilian savanna (Cerrado), wherein high-frequency fires drive the vegetation structure, composition, function, and dynamics of savanna ecosystems. Under climate change pressure, further understanding of fire-vegetation relationships and interactions can provide new approaches for establishing strategies for integrated fire management and promote savanna vegetation recovery post-fire. To understand how 15 years of yearly manipulated burning affects the vertical and horizontal structure of the vegetation, the species composition and diversity metrics (species richness, Shannon's diversity, and Pielou's evenness), and the aboveground carbon stocks, we surveyed all woody plant species with a diameter greater than three centimeters in 15 plots of a typical Brazilian savanna (cerrado stricto sensu) at an experimental research station in central Brazil (Cerrado biome). Fifteen plots (five per treatment) were differently affected by fire events over a decade, comprising three treatments: (i) annual fire, (ii) legacy fire (> 11 years since the last fire event), and control (not burned in the past 30 years). Non-metric multidimensional scaling (NMDS) analysis indicated a significant effect of fire on the species composition among treatments and that some species benefited from fire, such as Erythroxylum suberosum, whereas others propagated better without fire, such as Camisala montana and Dalbergia miscolobium. Over a decade of annual fire events led to decreases in Shannon's diversity, species richness, and stem density, which were significantly lower in the annual fire treatment than in the control and legacy fire treatments. Stem density by diameter and height size class (except for the 1-2 m class and above 8 m) was higher in the control than in the annual fire treatment, but the number of dead trees did not differ between the control and annual fire treatments. Our results also showed that fire was a factor in changes in the evaluated parameters, such as annual fire treatment, which reduced the amount of biomass and, therefore, the carbon stock. This study suggests that, if burned yearly, typical Brazilian savannas can become less biodiverse in terms of woody plant species and negatively affect their fire resilience. Therefore, fire management practices should focus on determining the frequency with which these ecosystems may benefit the most from fire disturbances.
Keywords:
Fire ecology
; cerrado stricto sensu
; savanna ecosystem
; woody vegetation
; plant ecology
1. Introduction
Understanding the natural and anthropic alterations that occur in savanna vegetation and biomass pools is essential for conservation science, especially as it can reflect on the elaboration of protocols for vegetation recovery, and on evaluations of the human-fire influence on the environment. Studies on fire disturbance effects on savanna vegetation provide evidence and insights on the resilience and dynamics of these ecosystems, in addition to monitoring the factors that govern changes in vegetation structure, composition, and productivity [1]. Tropical savanna biomes are particularly relevant, as there is a predominance of trees without a continuous canopy, a low density in the woody stratum, and a grass dominance [2]. They are among the most widespread terrestrial biomes [3,4], characterized by a biologically diverse flora [5], and potentially resistant and resilient to extreme drought conditions [6].
The Cerrado [Brazilian savanna biome], the second largest biome in Brazil, covers approximately 2.5 million square kilometers, or approximately 22% of the national territory [7,8], and is one of the world's biodiversity hotspots [9]. This biome is highly threatened because it is not entirely legally protected and has high rates of anthropic conversion to alternative land uses [4,10]. It is globally recognized for its exuberance and biodiversity and provides numerous ecosystem services [11,12], which stands out in the elaboration of ecosystem restoration protocols and potential for climate change mitigation [13]. Savanna ecosystems in the Cerrado are particularly influenced by the availability of water and nutrients, with fire as an agent maintaining the biomass gradient in this morphoclimatic domain [14,15], altering the vegetation composition and structure. These savanna ecosystems have been more recently investigated to estimate aboveground biomass because of the complexity of the expansion of anthropogenic activities that tend to affect their different formations [16,17] and predict information about their nutrient and carbon cycles [16].
In addition to intrinsic regional characteristics, such as acidic soils, poor nutrient content, and a superficial water table [18,19], fire is an agent that modifies the structure and composition of vegetation in savannas [20], thereby contributing to the establishment of different environments. Although fire is a prevailing disturbance in Cerrado's savanna ecosystems, it is challenging for ecosystem managers and scholars to use preventive fire as a method to avoid highly impactful fire events, which, according to current legislation, do not cover the specificities of each ecosystem type across the biome, and does not consider the ecological importance of biodiversity maintenance. Therefore, information regarding the need and usefulness of fire management is essential for the development and perpetuation of some plant species in the Cerrado [21]. Considering the importance of savanna fire management, there are records of the presence of fires for at least 32 thousand years [22], with fire being considered responsible for the antagonism of the physiognomies found in the cerrado [23]. Fire management legislation needs to be adapted to deal with fire issues, with the establishment of goals anchored in sustainability and effective and attainable long-term commitments to reduce negative fire effects in these environments.
Considering this pressing challenge, one method to minimize immeasurable damage caused by ecosystem fires is the use of integrated fire management, which has excellent applicability, occurring through prescribed, planned, and authorized burnings, considering each type of ecosystem. This approach results in different proposals for decision-making, including fire as a modeling agent [24], and considers the demands of local populations in the area where it is applied, their cultural traditions, and local socioeconomic conditions [25]. Therefore, there is an improvement in the understanding of fire-human-vegetation relationships in different Brazilian ecosystems, especially those in the Cerrado, considering their ecology [26] and acting as agents of the dynamic balance of savannas, forests, and the regulation of the advance and retreat of vegetation [27].
The overall objective of this study was to quantify the effects of fire on the structure, composition, diversity, and aboveground carbon stocks of the shrub-tree component of savanna ecosystems differently exposed to fire in central Brazil. The study was carried out in three savanna treatments, two of which were subjected to constant fire regimes (with and without planned annual fire during 15 years). Specifically, we quantified the effects of fire on [i] the vertical and horizontal structure of the vegetation, [ii] diversity metrics [richness, diversity, and evenness], and [iii] the aboveground biomass carbon stocks of shrub-tree savanna vegetation. We aimed to answer the following questions: i] Are 15 years of annual fire sufficient to change the floristic composition and diversity of savanna ecosystems? ii] What is the effect of annual fires on the vertical and horizontal structures of savannah vegetation? iii] Do aboveground biomass and carbon stocks change after 15 years of annual fire relative to the control areas? We tested the following hypotheses: H1] Annual burning in the cerrado alters the composition and diversity of the vegetation; H2] the incidence of fire diversifies the structure and modifies the distribution of species among size classes; H3] phytosociology, horizontal structure, and vegetation loss are parameters for evaluating fire effects on a savanna plant community; H4] burning alters vertical and horizontal parameters, compared to communities where there is no regular fire; and H5] fire influences species richness, diversity, and evenness, negatively altering richness in regularly burned areas.
2. Materials and Methods
2.1. Study Site
This study was conducted in savanna ecosystems in the Federal District, Brazilian Cerrado. The Federal District presents a District Law number 742 July 28, 1994, which deals with the protection of its natural vegetation, with a history of native vegetation unchanged until the 1960s. With the onset of the construction of Brasilia, the vegetation in this area was converted into agriculture and urbanization, through fires. From that period until now, high rates of deforestation and degradation have been witnessed, especially through the use of fire, which raises questions about the effects of fire on the composition, richness of plant species, and structure of vegetation, motivating these and other studies [21] to answer some questions about the recurrence and importance of these anthropic disturbances.
The study was conducted at Fazenda Água Limpa (FAL), the experimental farm of the University of Brasília [UnB], in a cerrado stricto sensu [15° 56' - 15° 59' S and 47° 55' – 47° 58' W; between 1,048 m and 1,150 m elevation], the predominant phytophysiognomy that covers approximately 1,480 ha [28,29]. With about 4,340 hectares of total area, the farm is part of the Cerrado Biosphere Reserve [30], which includes the Environmental Protection Areas (APA) Gama and Cabeça-de-Veado, the Botanical Garden of Brasília (JBB) and the Ecological Reserve of the Brazilian Institute of Geography and Statistics (IBGE). In FAL, there are preservation areas, such as the Relevant Area of Ecological Interest [Capetinga-Taquara stream], covering 2,100 hectares, with a history that exceeds 30 years of teaching, research dedicated to protection and conservation, and environmental sustainability [31]. The study area has an Aw-type climate [32], with dry seasons between May and October, rainy seasons from November to April, average annual rainfall of 1,600 mm, and monthly averages between 9 mm and 249 mm [33]. Air temperature ranges from 12 to 28.5°C and relative humidity from 15 to 70%. These soils are largely characterized as dystrophic red-yellow latosols, with high acidity and low nutrient availability [34,35], influencing the type of local vegetation. The area is composed of approximately 45% cerrado stricto sensu [36], distributed in flat terrain. For this research, the areas (treatments) selected at FAL were: an area with a history of annual burning [area 1, or annual fire], area with 15 years after the last fire event (area two, or legacy fire); and an area with no record of fire occurrence for 30 years (area three, or control) (Figure 1).
The three cerrado treatments chosen for the study were also designated for their high species richness and biodiversity, and for contemplating environments with easy access by the population, as they border a highway, and have a history of manipulated fire interventions. Also, approximately 38% of FAL's area are composed of savanna ecosystems [37].
2.2. Fire Experiment
Sampling plots were allocated within a cerrado stricto sensu vegetation area covering approximately 7.50 km. Fifteen plots (20 m x 50 m) were randomly allocated, three in each treatment, according to a well established protocol [38], totaling 0.5 ha in each sampled treatment, and 1.5 ha in the entire experimental area. In each treatment (Figure 1), the fire treatments were defined as Annual Fire, with a history of recurrent annual burning for at least 15 years, with the use of black firebreaks in the months of June or July, as a reduction in the load of combustible material in order to prevent larger fires; Legacy fire, with a history of fire occurrence, where approximately 15 years ago, an accidental fire accidently occurred, reaching approximately 30% of the FAL area; and Control, with no record of burning for over 30 years, according to the institution's documents.
The black firebreak of the annual fire treatment has the support of brigade members from the farm, a team from the Federal District military fire department, the Brazilian army, the Institute of the Environment and Renewable Natural Resources (IBAMA), the Brasília Environmental Institute (IBRAM), and volunteers in the vegetation that borders the highway between the months of June and July, the beginning of the fire risk season in the region. Activity commonly used as a way to reduce deposited plant material, thus avoiding possible outbreaks of fire or even large fires, falls within the regulations established by integrated fire management legislation.
2.3. Field Vegetation Survey
To estimate aboveground carbon stocks, we characterized the structure and floristic composition of the woody vegetation through a forest inventory of the tree-shrub component. Between June 2021 and ending in December 2021, each of the five plots (20 m x 50 m) of each treatment, systematically distributed in the area, was delimited by iron stakes, subdivided into 10 subplots of 100 m², and, all living and dead woody individuals were surveyed. The forest inventory was carried out in the dry season, before the black firebreak was carried out in the area with annual burning, followed by areas with a history of fire for 15 years, and areas with no record of burning for at least 30 years.
For sampling, plots of 0.1 ha were used in each treatment, with a sampling intensity of 0.5 ha, totaling 1.5 ha (3 areas x 0.5 ha) combining all three treatments. All individuals with a base diameter (Db) 0.30 cm from the ground, with a height equal to or greater than 5 cm [38] were measured and recorded. Diametric measurements were performed using uma suta and total height with a hypsometric ruler [39]. Bifurcated individuals were measured separately to obtain basal area and stem biomass. All trees falling within the established criteria were botanically identified at the family, genus, and species levels using the APG IV botanical classification system [40] and updated according to the nomenclature of the Flora do Brasil list [41]. This procedure aimed to quantify the parameters of Wealth, Shannon Diversity (H'), and Pielou Equability (J), according to [42], and phytosociological parameters of the horizontal structure: absolute and relative density, absolute and relative dominance, absolute and relative frequency, and importance value index [43,44,45].
2.4. Aboveground Carbon Stock Estimation
Fire affects ecosystem carbon fluxes [46,47] because trees and shrubs store carbon both above and belowground [48,49]. Thus, in the management of fire in ecosystems with high richness and phytosociological diversity of species, such as in the cerrado stricto sensu [50,51,52], a factor of great importance in indirect measurements is the accuracy of productivity estimates, which depends on the application of specific allometric equations generated for these environments [53,54]. In this study, aboveground biomass was estimated using models adjusted by [157]. The carbon stock was estimated as the product of biomass values by a factor of 0.47 [55]. Tree and shrub aboveground carbon stocks across treatments were estimated as: EC = 0,24564 + 0,01456 * Db2 * Ht (R² = 98,29% and Syx = 25,79%), where EC = carbon stock per tree, in kg; Db = Diameter taken at the base (30 cm above ground level) in cm; Ht = Total Height of the Tree or Shrub, in m; R² = coefficient of determination; Syx = standard error of the estimate [53].
2.5. Statistical Analysis
Floristic Composition and Structure
The analysis of the floristic composition and structure of the vegetation at the study site was conducted using the inventory data, using quantitative parameters established by [28,44]: stem density, dominance expressed by the basal area, absolute frequency expressed in absolute and relative values, by species, aiming to characterize the local tree-shrub cover. The assessment of diversity among the three treatments was performed using the Shannon Weaver (H') index [57,58,156] and Pielou's evenness [59].
To investigate the composition patterns of existing species in the areas, a non-metric multidimensional scaling (NMDS) analysis was carried out in the three areas, depending on the structural and floristic characteristics, for richness, abundance, and density of the species with the R software, to evaluate the similarities and data dissimilarities across functions. ANOVA was performed to detect differences among treatments in the evaluated parameters.
Carbon Stocks
The determination of aboveground carbon stocks in natural woody ecosystems has been largely conducted through allometric models that involve tree attribute data such as diameter, wood density and height that are collected from individuals in the studied plant community [60]. Thus, in this research, from inventory data, the number of trees, species, and diameters were obtained, which were used in the determination of aboveground carbon. Understanding how the above-ground biomass varies in the Cerrado, the carbon stock will bring environmental responses to this parameter, providing information on estimates of carbon stocks, and assisting in conservation policies [61]. The results of this evaluation of the analyzed parameters were made using boxplots, and other analyzes were performed in the R software (versão 3.6.2; [62].
3. Results
3.1. Species Composition across Treatments
The NMDS analysis indicated that the species composition differed significantly among treatments based on the species abundance matrix, suggesting that fire influenced the floristic composition of the studied cerrado ecosystems (Figure 1). NMDS showed that the floristic composition of the control treatment was more similar to that of the legacy fire treatment than to that of the annual fire treatment, reflecting the effect of fire on the composition of species in the studied savanna ecosystems. The ordination demonstrated the separation between the species in the treatments, with the control area and a fire record for 15 years, distinct from the group with annual fire occurrence, which resulted in changes in the composition of the species. In this case, there is evidence for the existence of a considerable number of species that are sensitive to repeated fires.
Figure 1.
Distribution of species by treatment based on non-metric multidimensional scaling (NMDS) analysis for the three treatments, including species with vectors significant at the 99% significance level (Appendix Table A1). Circles represent the annual fire treatment, triangles represent the control (30 years without fire), and squares represent the legacy fire treatment (i.e., where the fire occurred 15 years prior to the survey).
Figure 1.
Distribution of species by treatment based on non-metric multidimensional scaling (NMDS) analysis for the three treatments, including species with vectors significant at the 99% significance level (Appendix Table A1). Circles represent the annual fire treatment, triangles represent the control (30 years without fire), and squares represent the legacy fire treatment (i.e., where the fire occurred 15 years prior to the survey).
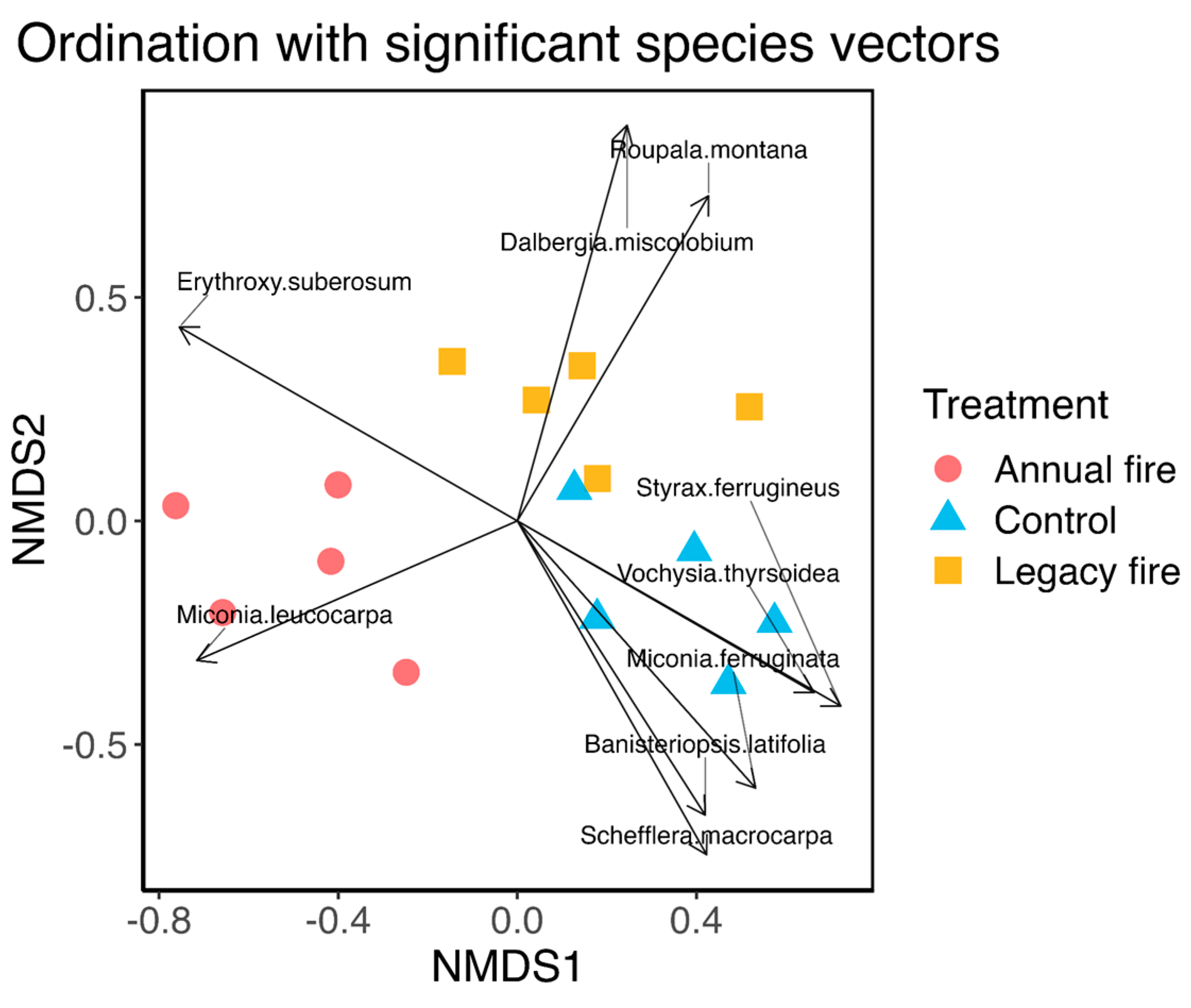
Our results indicated that the absence of fire for relatively longer periods contributed to the increase in species density. Among the three treatments, 2,734 individuals were identified, where the control presented 38.30% of the identified species, followed by the legacy fire treatment, with 37.6% of the species, and in the annual fire treatment the percentage of taxa was 24.1%. Some species, such as the species Miconia leucocarpa (Figure 1), were strongly associated with the occurrence of fire, however, species such as Camisala montana, showed a greater association in the legacy fire treatment, where greater intervals of fire incidence occurred. However, species such as Miconia ferruginata had a higher occurrence in the fire extinguishing treatment. These different behaviors may be associated with the strategies of each species, the architecture of individuals [63].
3.2. Diversity Metrics across Treatments
Floristic diversity in a community can be assessed by the number of existing species combined with their abundance, or by the combination of these two components [58]. At the research sites, species diversity was determined using the Shannon-Wiener index (H') (Figure 2a), and the Pielou evenness index (J) was also calculated (Figure 2c), which evaluates the relative abundance of species within the same studied sample [64]. From the obtained data, it was possible to observe differences in Shannon diversity and Pielou uniformity or evenness based on the evaluated area and fire.
Shannon's diversity varied significantly among the three treatments, wherein diversity was highest in the control treatment, with values ranging between 2.9 and 3.1, and lowest in the annual fire treatment (Figure 2a). The control treatment did not differ significantly from the legacy fire treatment. Species richness also varied between treatments, with values in the annual fire treatment lower than the other two treatments (Figure 2b). Periodic fire influences the characteristics of this biome, benefiting the ecosystem by enriching the soil through the action of burning ash, increasing organic matter [14], and nutrients produced and accumulated, thereby improving primary productivity in this biome, and forest nutrition processes [65]. [66] highlighted the frequency and time of burning, which is essential for the reduction of biomass, in biennial and four-year periods, helping to reduce grasses and the risk of severe fires.
Pielou evenness did not differ among treatments, with values ranging from 0.80 to 0.86. The value found for the legacy fire treatment, 0.85, was the highest average among the three treatments, suggesting that such treatment had the greatest uniformity (Figure 2c). The number of woody individuals per treatment was significantly different. Figure 2d shows the effects of fire on the number of individuals in each treatment, with the greatest number in the control plots, followed by the legacy fire and annual fire treatments.
3.3. Structural Differences across Treatments
Figure 3a shows the number of dead trees in each treatment. In the control, we identified 19 dead individuals, similar in quantity for the annual fire treatment, contrasting with the legacy fire, with 10 dead individuals. Among the diameter distributions obtained in each area (Figure 3b), the legacy fire treatment had the highest number of individuals [286] in the 1 – 5 cm classes; the control treatment stood out with greater quantity for diameter classes 5 -10 cm had individuals and 10 – 15 cm had 165 individuals. However, in the following diameter, 15 – 20 cm, the annual fire treatment stood out with 42 individuals. Evaluations of the number of individuals among the established diameters demonstrated their uniformity in the following diameter classes for the established treatments. Among the height classes (Figure 3c), there was variation from a few centimeters (0.4 m) to more than 9 m. In this analysis, the legacy fire treatment had the highest number of individuals, 606, with heights of up to 3 m. For the following height classes, 3-4m to 7 m, the control treatment group comprised more individuals, 435. However, in the 7 – 8m height class, the legacy fire treatment stood out with 11 specimens and 10 in the 8 – 9m class. This is in contrast to the three subjects for the control treatment and four for the annual fire. Above 9 m, the two treatments (control and legacy fire) had four individuals each, with no record of this height class for the annual fire treatment.
3.4. Aboveground Carbon Stocks across Treatments
Figure 4 shows the variation in live shrub-tree aboveground carbon stocks among the three treatments, indicating substantial variation between treatments. Our data indicate that aboveground carbon was highest in the control treatment (11.3 MgC/ha), which was followed by the legacy fire and annual fire treatments (7.2 MgC/ha). This analysis highlights the main consequences of fire on the vegetation of the Brazilian savanna in terms of carbon stock. Carbon stock values are good predictors of the structural conditions of a forest [67], such that forests in natural succession with smaller trees have less dispersed distributions of aboveground carbon [68]. In addition to areas with less interference, the intrinsic characteristics of the site influence carbon stocks, composition, and diversity [69].
Our aboveground carbon data support discussions on the changes imposed by fire frequency in the Brazilian Cerrado. Less frequent fires (e.g., control treatment) contribute to the differentiation of the levels of carbon stock in a region, influences the height of trees, the type of existing tree cover, and maximizes aboveground woody biomass [70]. However, savannas affected by fire sporadically, as in the legacy fire treatment, have increased structural complexity, changes in habitat, increase and diversity of biomass, and consequently greater availability of nutrients in the soil. Thus, in this study, the high rates of burning were not favorable for the carbon stock, as the seasonality of fire events considerably reduced the input of grasses, branches, leaves, and small shrubs, and therefore the stocks of carbon.
4. Discussion
4.1. Relationship between Fire and Cerrado Species Composition
Our findings support the hypothesis of the influence of annual burning on changing the composition and diversity of species in the evaluated savanna ecosystem. We expected that, in general, the fire differences would negatively affect the composition and diversity of the vegetation among the three the treatments in the Brazilian Cerrado. Fire is an intrinsic factor in altering vegetation in this environment [71], conferring to the evolution, resistance, and resilience of some species [72]. Several studies have shown that in Brazil, for the management of areas for agricultural and livestock purposes, the use of uncontrolled fire has caused high rates of fires every year, modifying the interactions of humans with the vegetation, changing the climate, the microorganisms existing in soils, being the main agent causing environmental problems, destroying and threatening biodiversity [73]. Due to the incidence of fire exposure, we observed that there are losses in composition and diversity, which contributes to the threat of the vegetation and its inclusion in the list of global biodiversity hotspots [9].
Our results demonstrated that species affected by fire recurrence, such as Miconia leucocarpa from the Melastomaceae family, the one with the highest representation in the Brazilian Cerrado [74], and Davilla elliptica, family Dilleniaceae, whose annual burning changes the ability for regrowth and development [75]. Fire frequency matters for vegetation because of the time needed for the recovery of nutrients needed for the regrowth of the aboveground component, which in constant regimes becomes unfeasible and the plant does not reach the minimum size for resistance and resilience to fire [21]. However, our research showed that annual, biennial, or four-year burnings did not occur, but with annual fires, the ability to overcome, regrow, and the development of species with medium heights and diameters was significant, such as Dalbergia miscolobium and Camisala montana, developing well with the inclusion of fire inclusion in its growth cycle. The findings of previous studies with prescribed fires are in agreement with ours, showing that these individuals have a greater height after a prescribed fire [111,116].
Fire has detrimental consequences on the radial growth of some plant species and can negatively affect sexual reproduction [77]. The lack of structures in the diaspores of monocotyledonous plants, such as hard integuments or more heat-resistant coverings, also contributes to seed loss during fire exposure. All layers of Cerrado vegetation are affected by fire, although in different ways, depending on the phenological characteristics of the species. Fire can negatively affect the sexual reproduction of some species, while favoring others. Some species inhabiting fire-prone environments have developed mechanisms that allow survival. According to [78], the bark is considered the most important mechanism by which trees protect themselves against forest fires because of its excellent thermal insulation capacity. The presence of fire can influence several aspects of plants, such as flowering [30], fruit and seed production, clonal reproduction [79], and plant architecture.[63]. [80] conclude that increasing the thickness of the shell promotes a higher level of exchange rate protection.
We tested a second hypothesis regarding the effects of fire on the vegetation composition and diversity by looking at the differences in the distribution and abundance of species across treatments. Our findings corroborate previous studies describing fire in the Cerrado as a conditioner of some species, where higher density and frequency occurred for individuals in the Araliaceae (Scheflera macrocarpa) and Styracaceae (Styrax ferrugineus) families. In the control and legacy fire treatments, other families followed this trend. This can be explained by the sensitivity of some individuals to environmental changes [2], which reported that in a savanna ecosystem, some species are more tolerant to fire than in other environments [77]. However, it can also depend on the peculiarities of each family. Fire studies in Brazilian savannas suggest that changes are promoted by fire, but to understand which species and families are most fire resistant, several data are needed over relatively longer periods of time [81], unlike the objective of this work.
To this end, the NMDS analysis confirmed the distinction between the species compositions across treatments. Although the results reinforce the interference of recurrent burnings on the cerrado ecosystems, it is still not possible and desirable to completely suppress fire because it is a natural disturbance that is historically needed and the vegetation is adapted [14,82,83]. Furthermore, completely suppressing fires is not a viable alternative [23], as it produces accumulation of organic matter and influences the intensity of fires, contributing to catastrophic events such as increased carbon emissions and loss of biodiversity [85,86,87,88]. Therefore, fires are required to eliminate excess shrubs and grassy materials [89]. These results support decision-making for operations using fire, such as the firebreak or black line that Fazenda Água Limpa that are performed annually.
4.2. Fire Effect on Vegetation Diversity
A total of 473 live species were registered in all treatments in this study, with 110 individuals in the annual fire treatment, 170 in the legacy fire treatment, and 193 in the control. The difference in floristic diversity was strongly correlated with the number of fires in the last 15 years, suggesting that fire is a determining factor in the structure of communities in this savanna, indicating the species are resistant to this fire gradient. Our study demonstrated that the plots excluding fire resulted in a more closed phytophysiognomic formation, dominated by trees such as Tachigali subvelutina and Caryocar brasiliense and shrubs (Erythroxylum suberosum and Erythroxylum tortuosum) sensitive to fire. In general, in places where fire is excluded, there is a possibility of invasion of the forest onto the savanna, reducing the diversity of shrub and herbaceous vegetation, which we call the process of "woody encroachment" [90]. Our results make sense, since changes in the herbaceous layer can demonstrate a change in the structure of the community, which is influenced by fire, changing the number of species between areas with regular fires and those not burned [81], indicating that periodic fire events gradually reduce species richness and diversity [91].
In this study, it was evident that the average diversity of the Shannon index and species richness in all plots differed significantly between treatments, being larger for control and legacy fire treatments. Our findings suggest that constant fire changes the richness and diversity in the communities where it occurs. Ratifying studies by [92 and 93] showed that fire alters the structure, composition, and density of the vegetation, affecting nutrient fluxes and input. Different studies have shown that the absence of fire allows the establishment of sensitive species and the possibility of conversion to different phytophysiognomies [30,94,95,96], being positive for resistant species and negative for species sensitive to high temperatures [94,97,98,99].
The differences in Pielou uniformity for all plots across treatments outlined in this study emphasized the interference of fire on the post-fire dynamics of the savanna vegetation. We found differences in the evenness of distributions between the control and legacy fire treatments and that in the annual fire treatment, which can be explained by the fact that areas with high rates of burning have more open tree cover due to a high mortality of adult trees [100]. These results confirm our hypothesis because regular burning simplified the vertical and horizontal parameters of the plant community. Certainly, plant species that are sensitive to fires are lost in areas with intense fires [101]. However, previous studies on the effects of the fire regime in Cerrado communities [21] suggested that fire is essential for the Cerrado, acting as a natural element for the biome and contributing to the evolution and diversity of species in this region. physiognomy.
As expected, the plots treated with annual fire had fewer woody individuals, with a total of 147 individuals, in contrast with the legacy fire treatment with 200 species, and the control with the greatest number of woody species, which in turn affected the development of vegetation. A study in the same type of savanna biome [21] evaluated the incidence of fire for a few years, observing a reduction in the shrub-tree vegetation, emphasizing that recurrent burning causes the mortality of woody plants [117,156,173], increases the dominance of grasses [21], alters the landscape, changes the floristic composition [111,128], impoverishes the complex cerrado system, and modifies the environmental structure [28].
4.3. Fire Effects on Vegetation Structure
We found a lower tree mortality in the legacy fire treatment, with 10 dead trees, in contrast to the 18 dead trees found in the two other treatments (Figure 3a). This perishing of plant species in Brazilian savannas is remarkable because of the flammability of individuals, favoring selective processes of plants with greater and lesser adaptation [105]. In places with constant burning events, the number of dead trees and species is derived from those with thinner bark and less developed roots, making them more susceptible [9,142,188]. Another factor that contributes to high mortality rates, in this case, for an area with an annual burning record, is the amount of fine fuel, grasses, leaves, trunks, and fine branches deposited in the soil [108]. However, we also found that non-occurrence of fire does not mean a reduction in mortality, explaining the need for fire for the growth of species in the Brazilian Cerrado. Therefore, mortality does not show major differences between forests where sporadic and interspersed fires occur [107].
Diametric values (Figure 3b) were strongly influenced by the minimum adopted criterion of 5 cm, according to a study underestimating individuals of shrub and subshrub species [109]. Legacy fire treatment stands out for this initial diameter class, indicating that burning, in addition to poor soils and rainfall seasonality, are determinants of the vegetation composition of the Cerrado [21]. These results indicate non-generalized diametric patterns in the Brazilian Cerrado, supporting research where the zero fire policy does not conserve, much less protect, the existing biodiversity [110]. Different fire regimes, determined by frequency, associated with global change factors and fire vectors, influence and drive the diverse characteristics of the biome [111]. As shown in the legacy fire treatment, certain periods of fire benefit species growth, and although fire positively influences treatment, for the evaluated metric, its effects differed strongly in each treatment [88]. According to [20], the sporadic nature of fire events can be a renewing factor for plants, with evidence that some species need fire to develop, which may have driven individuals of smaller diameter classes to stand out because of the inclusion of fire in their life cycle.
However, in the evaluation of the diametric classes, the plants measured between 5 and 10 cm, the burnings were not favorable, pointing to a greater diametric growth; for the control, a greater number of plants are within the mentioned class and regular burning reduces the number of individuals. For the following diameter class, 10 to 15 cm, there was a considerable difference in the number of individuals for the three treatments, indicating the main characteristics of the effects of fire on the savanna vegetation, reinforcing that it is an environment with smaller diameter trees and, consequently, thinner bark, and shrub species, which suffer more injuries during burning [111]. The following diameter class values corroborate the suggestion by [75] that species with diameters greater than 5 cm survive frequent fires, especially because of the complex dynamics of the savanna ecosystems, reflecting species adaptations to different climate variations, temperature fluctuations, edaphic particularities, anthropic interventions, and post-burn damage, and the interaction among all these variables is reflected in the basal area of savanna ecosystems.
Tree-shrub height ranged from 0.4 m to over 9 m across treatments (Figure 3c), with differences in the number of individuals within each size class among treatments. We noted that the legacy fire treatment stood out with the highest number of individuals (606), with heights of up to three meters. Among the following height classes, from 0.4 to 1 m, the treatment that presented the most specimens was the treatment with the occurrence of fire, indicating that for plant growth, fire is essential for development. In the height classes 3 - 4 m to 7 m the control treatment grouped most individuals (435). However, in the 7-8m size, the legacy fire treatment stood out with 11 specimens and 10 in the 8 - 9m class. Above 9 meters, the two treatments, control and legacy fire, had 4 individuals each, with no record of this height class for the annual fire treatment. In terms of vertical structure of the studied ecosystem, the treatments showed significant differences for the smallest sizes, where the treatments with fire occurrence had most individuals, supporting the role of fire in shaping the structure of savanna ecosystems [112,113,114].
The legacy fire treatment had the highest stem density within a height class of 1–3 m. In this height class, Dalbergia miscolobium, Pouteria ramiflora, Camisala montana, and Miconia burchellii occurred with maximum heights, benefiting from an episode of fire to maximize their development and thus stand out compared with other species. However, for heights above 3 m to 7 m, as the trees reached larger sizes, in addition to a reduction in the number of species, the treatments also differed. Between 8 m, we identified two species in the legacy fire treatment and one for the three treatments up to 9 m in height. We also identified a species of Bowdichia virgilioides and Caryocar brasiliense above 9 m in height in the control treatment. Our findings indicate that the control treatment had more individuals in the higher size classes, and there was no occurrence of individuals with great heights. Also, the studied species may have benefited from the indirect effects of fire on the composition and functioning of the ecosystem, increasing the availability of nutrients, light, and water. The direct and indirect effects of fire on the development of tropical vegetation are the result of competition, rapid release of nutrients into the soil, seasonality, and the characteristics of fire, intensity, frequency, and duration [115].
4.4. Fire Effects on Aboveground Carbon Stocks across Treatments
Areas with different values of carbon estimates in their natural habitat may be related to peculiarities, such as variations in height and diameter, age, type of anthropic interference, number of individuals [116,117], soil type and fertility, topographies [118], and individual density [117]. In our study, aboveground carbon stocks varied as a function of the direct effects of fire on biomass. The disturbance regime induced the treatment with annual fire and the reduction of the main constituents of the vegetation, influencing the dynamics of the carbon stocks. We suggest that the reduction of tree components where fire occurred regularly increased the availability of light and modified the microclimate causing carbon stocks to change, also due to exposure of the vegetation to high temperatures during fire events. The carbon stock estimates obtained in this research of 7.2 Mg/ha in the annual fire, 8.6 Mg/ha in the legacy fire, and 11.3 Mg/ha in the control were lower than those found by [119], who quantified the carbon stock at Fazenda Água Limpa and estimated stocks at 8.60 Mg/ha, [28], who found values around 13.27 Mg/ha. These differences may be related to the fact may have been caused because of the occurrences of fire in the treatments, but in general, we argue that the suppression of fires for this evaluation, contributed to the increase of the carbon stock.
Even if the values in this analysis were not relatively high for treatments with fire, because collections were carried out in a shorter period than most previous studies, measuring the impact of fire on carbon balance is extremely important because of its complexity and performance in controlling species composition, influencing ecosystem development, and impacts on ecosystem services. Thus, for our study site, the frequency of fire occurrence in this vegetation influences aboveground carbon stocks. Furthermore, vegetation responses to fire vary according to the duration, frequency, and intensity [77]. Therefore, the long-term effects of fires can modify the patterns, fluxes, and carbon stocks of Brazilian savannas. Thus, distinguishing the consequences of fire, in the short and long term, in the distribution, species composition, and carbon stocks in these places is a challenge, due to the diverse characteristics of each phytophysiognomy, in addition to the fact that the dissemination of harmful effects overlaps the current debates, where fire management in tropical savannas should be considered in decision-making processes. The loss of biomass by fire events contributes to the exposure of the tree stratum and the loss of woody individuals, altering the climate and plant communities [121]. Our results substantiate claims that fire should not be completely excluded from this ecosystem because of the accumulation of dry biomass, which intensifies the possibility of large and intense fires [121,122,148].
5. Conclusions
This study explored the effects of fire on the vegetation of a Brazilian Neotropical savanna, suggesting interference of annual fires on the diversity, composition, richness, and vertical and horizontal distribution of individuals. Our hypotheses related to the composition and structure of the vegetation were supported, since annual fires altered the evaluated vegetation parameters, diversifying the distribution and imposing divergent patterns when compared to treatments without regular fire. Contrary to our expectations, tree mortality was determined by both fire and its absence. We deduced that the absence of fire did not characterize the exemption of tree deaths. In general, aboveground carbon stocks are reduced with annual fire events. In the future, the assessment of biomass over a longer period of time will be able to predict these estimates to consider seasonality and determine the levels of anthropogenic and natural interference, as well as to establish fire disturbance parameters for restoration and conservation of carbon stocks in these savanna ecosystems.
There are many challenges to overcome when estimating carbon stocks in Cerrado ecosystems due to environmental and structural aspects, different types of vegetation, and the application of tools and methodologies that consider the variability and specificity of each location. Additional studies, which take longer to evaluate in the field, are essential to deepen the understanding of the main effects of fire on the entire composition and structure of savanna ecosystems, whether for the evaluation of behavioral patterns in the mediation of every component of the biomass, the consequences on nutrient cycling, the development and mortality of woody individuals, and the increment that fire events impose and influence on carbon stocks and on the ecosystem dynamics and regeneration post-fire.
Author Contributions
Conceptualization, S.C.M.N., R.S.P., P.V.S; methodology, S.C.M.N., R.S.P, P.V.S.; formal analysis, R.S.P.; UNB., CAPES.; data curation, S.C.M.N., R.S.P; writing—original draft preparation, S.C.M.N., B.B., R.S.P.; writing—review and editing, S.C.M.N., B.B., R.S.P. P.V., A.F.T.; supervision, B.B and R.S.P.; All authors have read and agreed to the published version of the manuscript.
Funding
Graduate Program at the University of Brasilia - UNB; Coordination for the Improvement of Higher Education Personnel - CAPES
Data Availability Statement
The data can be made available upon request to the first author.
Acknowledgments
The authors thank the Fazenda Água Limpa research station and its employees Sebastião, Geraldo, Rodrigo, Augusto, Alcides, Augustinho, Luciano, Mauro, Miron, Zico, Queen, Alexandre Palermo, Lícia, teacher Ana Maria for all the support to develop this study, Pâmela Vírgilio for field and laboratory work support,; André for his support in carrying out the forest inventory; to the employees of Fazenda Água Limpa,, the Forest Sciences graduate program at the University of Brasilia, and CAPES for the doctoral scholarship and financial incentives for survival and carrying out the analyses.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Table A1.
Results of the Non-Metric Multidimensional Scaling (NMDS) analysis. R2 is the coefficient of determination and the asteristics refer to the significance level, where those with three asteristics are the vectors included in the NMDS Figure 2.
Table A1.
Results of the Non-Metric Multidimensional Scaling (NMDS) analysis. R2 is the coefficient of determination and the asteristics refer to the significance level, where those with three asteristics are the vectors included in the NMDS Figure 2.
| Species | NMDS1 | NMDS2 | R2 | P-value |
|---|---|---|---|---|
| Agonandra brasiliensis | 0.54628 | 0.83760 | 0.2012 | 0.321 |
| Aspidosperm .tomentosum | 0.90325 | 0.42912 | 0.1018 | 0.511 |
| Banisteriopsis latifolia | 0.53772 | -0.84312 | 0.6093 | 0.001 *** |
| Blepharocalyx salicifolius | -0.15560 | 0.98782 | 0.0870 | 0.598 |
| Byrsonima.pachyphylla | -0.86522 | -0.50139 | 0.1873 | 0.295 |
| Byrsonima.verbascifolia | -0.91416 | -0.40534 | 0.0769 | 0.654 |
| Bowdichia.virgilioides | 0.29841 | 0.95444 | 0.1670 | 0.328 |
| Casearia.sylvestris | -0.91062 | -0.41324 | 0.3848 | 0.052 . |
| Caryocar.brasiliense | 0.88254 | -0.47024 | 0.4325 | 0.014 * |
| Connarus.suberosus | -0.93377 | 0.35786 | 0.0402 | 0.779 |
| Dalbergia.miscolobium | 0.26686 | 0.96374 | 0.8432 | 0.001 *** |
| Dalbergia.elliptica | 0.63236 | -0.77468 | 0.2087 | 0.275 |
| Davilla.elliptica | -0.80026 | -0.59965 | 0.3734 | 0.035 * |
| Diospyros.burchellii | -0.57408 | 0.81880 | 0.0911 | 0.676 |
| Diospyros.lasiocalyx | 0.20319 | 0.97914 | 0.5188 | 0.014 * |
| Dimorphandra.mollis | 0.38512 | -0.92287 | 0.2701 | 0.056 . |
| Enterolobium.gummiferum | 0.81475 | -0.57982 | 0.0684 | 0.691 |
| Eremanthus.glomerulatus | -0.43466 | -0.90059 | 0.2094 | 0.241 |
| Eriotheca.pubescens | 0.99989 | 0.01483 | 0.0304 | 0.823 |
| Erythroxylum.deciduum | -0.22797 | -0.97367 | 0.0002 | 0.999 |
| Erythroxy.suberosum | -0.86718 | 0.49800 | 0.7576 | 0.001 *** |
| Erythroxylum.tortuosum | 0.76754 | -0.64100 | 0.2761 | 0.125 |
| Guapira.noxia | 0.77366 | -0.63360 | 0.4021 | 0.047 * |
| Handroanthus.ochraceus | 0.84702 | 0.53156 | 0.1538 | 0.370 |
| Hancornia.speciosa | -0.22933 | -0.97335 | 0.1786 | 0.412 |
| Hymenaea.stigonocarpa | 0.32592 | -0.94540 | 0.4944 | 0.026 * |
| Heteropterys.byrsonimiifolia | 0.45763 | -0.88914 | 0.0534 | 0.768 |
| Kielmeyera.coriacea | 0.05631 | 0.99841 | 0.1677 | 0.411 |
| Kielmeyera.speciosa | 0.25950 | 0.96574 | 0.4671 | 0.022 * |
| Lafoensia.pacari | 0.88586 | -0.46396 | 0.0725 | 0.861 |
| Leptolobium.dasycarpum | -0.49854 | -0.86686 | 0.1652 | 0.359 |
| Machaerium.opacum | 0.73150 | -0.68184 | 0.4537 | 0.013 * |
| Miconia.albicans | 0.60432 | 0.79674 | 0.2459 | 0.162 |
| Miconia.leucocarpa | -0.91636 | -0.40035 | 0.6099 | 0.002 ** |
| Miconia.burchelli | 0.16307 | 0.98661 | 0.3891 | 0.046 * |
| Miconia.speciosa | 0.13303 | 0.99111 | 0.1687 | 0.519 |
| Miconia.ferruginata | 0.66501 | -0.74684 | 0.6406 | 0.004 ** |
| Mimosa.claussenii | 0.76611 | -0.64271 | 0.1956 | 0.265 |
| Myrsine.guianensis | -0.04177 | 0.99913 | 0.1592 | 0.425 |
| Neea.theifera | -0.90570 | -0.42392 | 0.0731 | 0.640 |
| Ouratea.hexasperma | 0.10205 | 0.99478 | 0.0456 | 0.739 |
| Palicourea.rigida | 0.21823 | 0.97590 | 0.3388 | 0.061 . |
| Pouteria.ramiflora | 0.60027 | -0.79979 | 0.4059 | 0.027 * |
| Piptocarpha.rotundifolia | -0.84519 | -0.53447 | 0.2268 | 0.209 |
| Psidium.laruotteanum | 0.19464 | 0.98087 | 0.0218 | 0.913 |
| Plenckia.populnea | -0.71841 | -0.69562 | 0.2406 | 0.224 |
| Pterodon.emarginatus | 0.79357 | -0.60848 | 0.3462 | 0.072 . |
| Qualea.grandiflora | 0.42728 | -0.90412 | 0.3066 | 0.129 |
| Qualea.multiflora | -0.03812 | 0.99927 | 0.1074 | 0.524 |
| Qualea.parviflora | 0.86802 | -0.49653 | 0.2768 | 0.123 |
| Roupala.montana | 0.50624 | 0.86239 | 0.7116 | 0.001 *** |
| Rourea.induta | 0.98941 | 0.14514 | 0.0767 | 0.674 |
| Salacia.crassifolia | 0.63236 | -0.77468 | 0.2087 | 0.275 |
| Schefflera.macrocarpa | 0.49271 | -0.87019 | 0.7362 | 0.001 *** |
| Symplocos.rhamnifolia | 0.53729 | 0.84340 | 0.2863 | 0.147 |
| Strychnos.pseudoquina | 0.83442 | -0.55114 | 0.1168 | 0.541 |
| Syagrus.comosa | 0.05081 | 0.99871 | 0.0976 | 0.609 |
| Styrax.ferrugineus | 0.86754 | -0.49737 | 0.6936 | 0.001 *** |
| Stryphnodendron.adstringens | 0.77755 | -0.62883 | 0.1788 | 0.291 |
| Solanum.lycocarpum | 0.00000 | 0.00000 | 0.0000 | 1.000 |
| Tachigali.subvelutina | -0.49209 | -0.87054 | 0.1239 | 0.422 |
| Vatairea.macrocarpa | 0.35803 | -0.93371 | 0.0222 | 0.900 |
| Vochysia.thyrsoidea | 0.86539 | -0.50109 | 0.5867 | 0.002 ** |
| Vochysia.elliptica | 0.67429 | 0.73847 | 0.4986 | 0.012 * |
| --- | ||||
| Signif. codes: 0 ‘***’ 0.00 | 1 ‘**’ 0.0 | 1 ‘*’ 0. | 05 ‘.’ 0 | .1 ‘ ’ 1 |
References
- Wang, J.; Han, W.; Ge, C.; Guan, H.; Yang, H.; Zhang, X. Form-stable oxalic acid dihydrate/glycolic acid-based composite PCMs for thermal energy storage. Renew. Energy 2019, 136, 657–663. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Walter, B.M.T. As principais ftofsionomias do bioma Cerrado. In Cerrado: Ecologia e Fora; Sano, S.M., Almeida, S.P., Ribeiro, J.F., Eds.; . Embrapa Informação Tecnológica: Brasília, Brazil, 2008; pp. 151–212. [Google Scholar]
- Terra, M.d.C.N.S.; Prado-Júnior, J.A.D.; de Souza, C.R.; Pinto, L.O.R.; Silveira, E.M.d.O.; Cordeiro, N.G.; Cirne-Silva, T.M.; Mantovani, V.A.; Scolforo, J.R.S.; de Mello, J.M. Tree species dominance in neotropical savanna aboveground biomass and productivity. For. Ecol. Manag. 2021, 496, 119430. [Google Scholar] [CrossRef]
- Sano, E.E.; Rodrigues, A.A.; Martins, E.S.; Bettiol, G.M.; Bustamante, M.M.C.; Bezerra, A.S.; Couto, A.F., Jr.; Vasconcelos, V.; Schüler, J.; Bolfe, E.L. Cerrado ecoregions: A spatial framework to assess and prioritize Brazilian savanna environmental diversity for conservation. J. Environ. Manag. 2019, 232, 818–828. [Google Scholar] [CrossRef]
- Rocha, G.F.; Ferreira, L.G.; Ferreira, N.C.; Ferreira, M.E. Detecção de desmatamento no bioma Cerrado entre 2002 e 2009: Padrões, Tendência e Impactos. Rev. Bras. Cartogr. 2011, 63, 341–349. [Google Scholar] [CrossRef]
- Swann, A.L.S. Plants and Drought in a Changing Climate. Curr. Clim. Chang. Rep. 2018, 4, 192–201. [Google Scholar] [CrossRef]
- Strassburg, B.B.N.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R.; Loyola, R.; Latawiec, A.E.; Filho, F.J.B.O.; Scaramuzza, C.A.D.M.; Scarano, F.R.; et al. Moment of truth for the Cerrado hotspot. Nat. Ecol. Evol. 2017, 1, 99. [Google Scholar] [CrossRef]
- BrasiL. Ministério do Meio Ambiente. Cadastro Nacional de Unidades de Conservação. Brasília: MMA. 2016. Available online: http://www.mma.gov.br/images/arquivo/80112/CNUC_Agosto%20-%20Biomas%201.pdf (accessed on 20 July 2021).
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Silveira, E.M.O.; Silva, S.H.G.; Acerbi-Junior, F.W.; Carvalho, M.C.; Carvalho, L.M.T.; Scolforo, J.R.S.; Wulder, M.A. Object-based random forest modelling of aboveground forest biomass outperforms a pixel-based approach in a heterogeneous and mountain tropical environment. Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 175–188. [Google Scholar] [CrossRef]
- Gomes, L.; Miranda, H.S.; Maria, M. Forest Ecology and Management How can we advance the knowledge on the behavior and e ff ects of fi re in the Cerrado biome? Forest Ecol. Manag. 2018, 417, 281–290. [Google Scholar] [CrossRef]
- Strassburg, B.B.N.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R.; Loyola, R.; Latawiec, A.E.; Filho, F.J.B.O.; Scaramuzza, C.A.D.M.; Scarano, F.R.; et al. Moment of truth for the Cerrado hotspot. Nat. Ecol. Evol. 2017, 1, 99. [Google Scholar] [CrossRef]
- Strassburg, B.B.N.; Iribarrem, A.; Beyer, H.L.; Cordeiro, C.L.; Crouzeilles, R.; Jakovac, C.C.; Junqueira, A.B.; Lacerda, E.; Latawiec, A.E.; Balmford, A.; et al. Global priority areas for ecosystem restoration. Nature 2020, 586, 724–729. [Google Scholar] [CrossRef]
- Coutinho, L.M. Fire in the Ecology of the Brazilian Cerrado. In Fire in the Tropical Biota: Ecosystem Processes and Global Challenges; Springer: Berlin, Germany, 1990; pp. 82–105. [Google Scholar]
- Mistry, J. World Savannas: Ecology and Human Use; Great Britain: Pearson Education Limited; Prentice Hall: Hoboken, NJ, USA, 2000. [Google Scholar]
- Pereira, K.M.G.; Cordeiro, N.G.; Terra, M.d.C.N.S.; Pyles, M.V.; Cabacinha, C.D.; de Mello, J.M.; Berg, E.v.D. Protection status as determinant of carbon stock drivers in Cerrado sensu stricto. J. Plant Ecol. 2020, 13, 361–368. [Google Scholar] [CrossRef]
- Rocha, G.F.; Ferreira, L.G.; Ferreira, N.C.; Ferreira, M.E. Detecção de desmatamento no bioma Cerrado entre 2002 e 2009: Padrões, Tendência e Impactos. Rev. Bras. Cartogr. 2011, 63, 341–349. [Google Scholar] [CrossRef]
- Haridasan, M. Nutrição mineral de plantas nativas do cerrado. R Bras Fisiol Veg 2000, 12, 54–64. [Google Scholar]
- Eiten, G. Cerrado: Caracterização, ocupação e perspectivas. UnB: SEMATEC; 1994; pp. 1–73.
- Medeiros, M.B.; Miranda, H.S. Mortalidade pós-fogo em espécies lenhosas de campo sujo submetido a três queimadas prescritas anuais. Acta Bot. Bras. 2005, 19, 493–500. [Google Scholar] [CrossRef]
- Miranda, H.S. Efeitos do regime de fogo sobre a estrutura de comunidades de Cerrado: Projeto Fogo; IBAMA: Brasília, Brazil, 2010; 144p. [Google Scholar]
- Ferraz-Vicentini, K.R.C. História do Fogo no Cerrado. Tese (Doutorado)—Departamento de Ecologia; Universidade de Brasília, Brasília, Brazil, 1999.
- Solbrig, O.T.; Young, M.D. Economic and ecological driving forces affecting tropical savannas. The world’s savannas: Economic driving forces, ecological constraints and policy options for sustainable land use. Unesco, Paris: The Parthenon Publishing Group, 1993; pp. 3–8.
- Pivello, V.R.; Norton, G.A. Firetool: An expert system for the use of prescribed fires in cerrado (Brazlian savanna) conservation areas. J. Appl. Ecol. 1996, 33, 348–356. [Google Scholar] [CrossRef]
- Eloy, E.; da Silva, D.A.; Caron, B.O.; Elli, E.F.; Schwerz, F. Effect of age and spacing on biomass production in forest plantations. 2018, 42. 42. [CrossRef]
- Bilbao, B.; Mistry, J.; Millán, A.; Berardi, A. Sharing Multiple Perspectives on Burning: Towards a Participatory and Intercultural Fire Management Policy in Venezuela, Brazil, and Guyana. Fire 2019, 2, 39. [Google Scholar] [CrossRef]
- Hopkins, B. Ecological processes at the forest-savanna boundary. Nature and Dynamics of Forest-Savanna Boundaries; Chapman & Hall: London, UK, 1992; pp. 21–33. [Google Scholar]
- Rezende, A.V.; Vale, A.T.; Sanquetta, C.R.; Figueiredo Filho, A.; Felfili, J.M. Comparação de modelos matemáticos para estimativa do volume, biomassa e estoque de carbono da vegetação lenhosa de um cerrado sensu stricto em Brasília, DF. Sci.For. 2005, 71, 65–76. [Google Scholar]
- Sanquetta, C.R. Métodos de determinação de biomassa florestal. As florestas e o carbono. Curitiba.
- Felfili, J.M.; Carvalho, F.A.; Haidar, R.F. Manual para o monitoramento de parcelas permanentes nos biomas Cerrado e Pantanal. Brasília, 2005.
- Granado, L.M.A. Estimativa de biomassa e combustível em diferentes fitofisionomias do Cerrado / Larissa Moreira Alves Granado -- Brasília - DF, 2019. 71 p. Dissertação (Mestrado) -- Faculdade de Tecnologia -Universidade de Brasília - UnB.
- Koppen, W. Climatologia: Con un estudio de los climas de la tierra; Fondo de Cultura Econômica: México, 1948; 479p. [Google Scholar]
- Nimer, E. Climatologia do Brasil; IBGE: Rio de Janeiro, Brazil, 1989; 422p. [Google Scholar]
- Haridasan, M. Nutritional adaptation of native plnts of the cerrado biome in acid soils. Braz. J. Plant Physiol. 2008, 20, 183–195. [Google Scholar] [CrossRef]
- dos Santos, G.L.; Pereira, M.G.; Delgado, R.C.; Torres, J.L.R. Natural regeneration in anthropogenic environments due to agricultural use in the cerrado, Uberaba, MG, Brazil. Biosci. J. 2017, 33, 169–176. [Google Scholar] [CrossRef]
- Felfili, J.M.; Rezende, A.V.; Júnior, M.C.D.S.; Silva, M.A. Changes in the floristic composition of cerrado sensu stricto in Brazil over a nine-year period. J. Trop. Ecol. 2000, 16, 579–590. [Google Scholar] [CrossRef]
- Azevedo, G.B. Amostragem e modelagem da biomassa de raízes em um Cerrado sentido restrito no Distrito Federal. Dissertação (Mestrado em Ciências Florestais). Universidade de Brasília, 2014.
- Felfili, J.M. Composição florística e fitossociológica de um cerrado sensu stricto em Água Boa - MT. Acta Bot. Bras. 2002, 16, 103–112. [Google Scholar] [CrossRef]
- Durigan, G. Estrutura e diversidade de florestas tropicais. In: S.V. Martins (org.). Ecologia de Florestas Tropicais do Brasil. Editora UFV, Viçosa. 2009; pp. 185–215.
- Angiosperm Phylogeny Group (APG IV). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Forzza, R.C.; Baumgratz, J.F.A.; Bicudo, C.E.M.; Canhos, D.A.L.; Carvalho, A.A.; Coelho, M.A.N.; Costa, A.F.; Costa, D.P.; Hopkins, M.G.; Leitman, P.M.; et al. New Brazilian Floristic List Highlights Conservation Challenges. BioScience 2012, 62, 39–45. [Google Scholar] [CrossRef]
- Felfili, J.M.; Rezende, R.P. Conceitos e métodos em fitossociologia. Brasília: Universidade de Brasília, Departamento de Engenharia Florestal. Comun. Técnicas Florestais 2003, 5, 1–68. [Google Scholar]
- Lamprecht, H. Silvicultura nos trópicos: Ecossistemas florestais e respectivas espécies arbóreas – possibilidades e métodos de aproveitamento sustentado. rossdorf: República Federal da Alemanha: Dt. Ges. Für Techn. Zusammenarbeit; 1990.
- Mueller-Dombois, D.; Ellenberg, H. Aims and methods of vegetation ecology. The Blackburn Press: New Jersey, 2002; 547p.
- Kent, M.; Coker, P. Vegetation Description: A Practical Approach; Belhaven Press: London, UK, 1992; 363p. [Google Scholar]
- Lal, R. Carbon sequestration. Philos. Trans. R. Soc. B 2008, 363, 815–830. [Google Scholar] [CrossRef]
- Grace, J.; Jose, J.S.; Meir, P.; Miranda, H.S.; Montes, R.A. Productivity and carbon fluxes of tropical savannas. J. Biogeogr. 2006, 33, 387–400. [Google Scholar] [CrossRef]
- Bustamante, M.M.C.; Oliveira, E.L. Impacto das atividades agrícolas, florestais e pecuárias nos recursos naturais. Savanas: Desafios e estratégias para o equilíbrio entre sociedade, agronegócio e recursos naturais. Planaltina: Embrapa Cerrados, 2008.
- Houghton, R.A.; Hall, F.; Goetz, S.J. Importance of biomass in the global carbon cycle. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Felfili, J.M.; Eisenlohr, P.V.; Melo, M.M.R.F.; Andrade, L.A.; Meira Neto, J.A.A. Fitossociologia no Brasil – Volume 1: Métodos e estudos de casos. UFV, 2011.
- Felfili, J.M.; Fagg, C.W. Floristic composition, diversity and structure of the “cerrado” sensu stricto on rocky soils in northern Goiás and southern Tocantins, Brazil. Rev. Bras. Bot. 2007, 30, 375–385. [Google Scholar] [CrossRef]
- Mendonça, R.C.; Felfili, J.M.; Walter, B.M.T.; Silva Júnior, M.C.; Rezende, A.V.; Filgueiras, T.S.; Nogueira, P.E. Flora vascular do cerrado. Cerrado: Ambiente e flora. Embrapa- CPAC. Planaltina, DF. 1998.
- Rezende, A.V.; Vale, A.T.; Sanquetta, C.R.; Figueiredo Filho, A.; Felfli, J.M. Comparação de modelos matemáticos para estimativa do volume, biomassa e estoque de carbono da vegetação lenhosa de um cerrado sensu stricto em Brasília, DF. Sci. For. 2006, 71, 65–76. [Google Scholar]
- Sanquetta, C.R. Análise da estrutura vertical de florestas através do diagrama h-M. Ciência Florest. 1995, 5, 55–68. [Google Scholar] [CrossRef]
- IPCC. National Greenhouse Gas Inventories Programme; IGES: Japan, 2006. [Google Scholar]
- Martins, F.R.; Santos, F.A.M. Técnicas usuais de estimativa da biodiversidade. Revista Holos 1999, 1, 236–267. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; Urbana University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University: Princeton, NJ, USA, 1988; 192p. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley Interscience: New York, NY, USA, 1975. [Google Scholar]
- de Souza, C.R.; de Souza, F.C.; Maia, V.A.; de Aguiar-Campos, N.; Coelho, P.A.; Farrapo, C.L.; Santos, A.B.M.; Araújo, F.C.; Gianasi, F.M.; Paula, G.G.P.; et al. Tropical forests structure and diversity: A comparison of methodological choices. Methods Ecol. Evol. 2021, 12, 2017–2027. [Google Scholar] [CrossRef]
- Morandi, P.S.; Marimon, B.S.; Marimon-Junior, B.H.; Ratter, J.A.; Feldpausch, T.R.; Colli, G.R.; Munhoz, C.B.R.; Júnior, M.C.d.S.; Lima, E.d.S.; Haidar, R.F.; et al. Tree diversity and above-ground biomass in the South America Cerrado biome and their conservation implications. Biodivers. Conserv. 2018, 29, 1519–1536. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 3.4.2); R Foundation for Statistical Computing: Vienna, Austria. 2017. [Google Scholar]
- Klink, C.A.; Machado, R.B. Conservation of the Brazilian Cerrado. Conserv. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science Ltd: Oxford, UK, 2004. [Google Scholar]
- Inkotte, J.; Martins, R.C.C.; Scardua, F.P.; Pereira, R.S. Métodos de avaliação da ciclagem de nutrientes no bioma Cerrado: Uma revisão sistemática. 2019, 29, 988–1003. [CrossRef]
- Pivello, V.R.; Oliveras, I.; Miranda, H.S.; Haridasan, M.; Sato, M.N.; Meirelles, S.T. Effect of fires on soil nutrient availability in an open savanna in Central Brazil. Plant Soil 2010, 337, 111–123. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and Status and trends of amphibian declines and extinction worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Piponiot, C.; Derroire, G.; Descroix, L.; Mazzei, L.; Rutishauser, E.; Sist, P.; Hérault, B. Assessing timber volume recovery after disturbance in tropical forests – A new modelling framework. Ecol. Model. 2018, 384, 353–369. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Zhou, Y.; Singh, J.; Butnor, J.R.; Coetsee, C.; Boucher, P.B.; Case, M.F.; Hockridge, E.G.; Davies, A.B.; Staver, A.C. Limited increases in savanna carbon stocks over decades of fire suppression. Nature 2022, 603, 445–449. [Google Scholar] [CrossRef]
- Simon, M.F.; Grether, R.; de Queiroz, L.P.; Skema, C.; Pennington, R.T.; Hughes, C.E. Recent assembly of the Cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proc. Natl. Acad. Sci. USA 2009, 106, 20359–20364. [Google Scholar] [CrossRef]
- Dantas, J.S. Relação solo-paisagem e predição da erodibilidade de solos coesos dos Tabuleiros Costeiros no Estado do Maranhão. Tese (Programa de Pós Graduação em Agronomia – Ciência do Solo) – Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Jaboticabal, SP. 2013.
- Lopes, E.R.N.; Silva, A.P.P.S. ; Peruchi,, J.F. Zoneamento de risco de incêndio e queimadas no município de Sorocaba - São Paulo. Revista do Departamento de Geografia, Universidade de São Paulo - SP. 2018.
- Goldenberg, R.; Baumgratz, J.F.A.; Souza, M.L.D.E.R. Taxonomia de Melastomataceae no Brasil: Retrospectiva, perspectivas e chave de identificação para os gêneros. Rodriguésia 2012, 63, 145–161. [Google Scholar] [CrossRef]
- Medeiros, M.B. Efeitos do fogo nos padrões de rebrotamento em plantas lenhosas, em campo sujo, após queimadas prescritas. Tese, Universidade de Brasília. 2002.
- Medeiros, M.B.; Miranda, H.S. Post-fire resprouting and mortality in cerrado woody plant species over A three-year period. Edinb. J. Bot. 2008, 65, 53–68. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Moreira, A. The role of fire in population dynamics of woody plants. In Ecology and Natural History of a Neotropical Savanna: The cerrados of Brazil; The University of Columbia Press: New York, NY, USA, 2002; pp. 159–177. [Google Scholar]
- Soares, R.V.; Batista, A.C.; Tetto, A.F. Incêndios florestais: Controle, efeitos e uso do fogo. Curitiba, 2ª edição revisada, 2017; 255p.
- Hoffman, W.A. The effects of fire and cover on seedling establishment in a neotropical savana. J. Ecol. 1996, 84, 383–393. [Google Scholar] [CrossRef]
- Vale, A.T.; Elias, P.S. Nível de proteção térmica da casca de quatro espéciees lenhosas e a relação da arquitetura da casca com a transferência de calor. Ciência Florestal. Santa Maria 2014, 24, 979–987. [Google Scholar]
- Lima, J.M.; Castro, A.B.; Lima, A.P.; Magnusson, W.E.; Landeiro, V.L.; Fadini, R.F. Influência do regime de queimadas sobre a riqueza e composição florística de uma savana isolada na amazônia - peld oeste do pará. Oecologia Aust. 2020, 24. [Google Scholar] [CrossRef]
- Miranda, H.S.; Sato, M.N.; Andrade, S.M.; Haridasan, M.; Moraes, H.C. Queimadas de Cerrado: Caracterização e impactos. In Aguiar, L.M.S.; Camargo, A.J.A. (Ed.). Cerrado: Ecologia e caracterização. Brasília: Embrapa Cerrados, 2004; pp. 69–123. [Google Scholar]
- Pivello, V.R. Fire management for biological conservation in the Brazilian cerrado. In Savannas and Dry Forests; Routledge: Abingdon-on-Thames, UK, 2017; pp. 141–166. [Google Scholar]
- Bowman, D.M.; Murphy, B.P.; Boer, M.M.; A Bradstock, R.; Cary, G.J.; A Cochrane, M.; Fensham, R.J.; A Krawchuk, M.; Price, O.F.; Williams, R.J. Forest fire management, climate change, and the risk of catastrophic carbon losses. Front. Ecol. Environ. 2013, 11, 66–67. [Google Scholar] [CrossRef]
- Bond, W.J.; Archibald, S. Confronting complexity: Fire policy choices in South African savanna parks. Int. J. Wildland Fire 2003, 12, 381–389. [Google Scholar] [CrossRef]
- Pivello, V.R. The Use of Fire in the Cerrado and Amazonian Rainforests of Brazil: Past and Present. Fire Ecol. 2011, 7, 24–39. [Google Scholar] [CrossRef]
- Wang, J.; Han, W.; Ge, C.; Guan, H.; Yang, H.; Zhang, X. Form-stable oxalic acid dihydrate/glycolic acid-based composite PCMs for thermal energy storage. Renew. Energy 2019, 136, 657–663. [Google Scholar] [CrossRef]
- Wang, B.; Nolan, R.; Marshall, H. COVID-19 Immunisation, Willingness to Be Vaccinated and Vaccination Strategies to Improve Vaccine Uptake in Australia. Vaccines 2021, 9, 1467. [Google Scholar] [CrossRef] [PubMed]
- Durigan. Zero-fire: Not possible nor desirable in the Cerrado of Brazil. Flora, 2020; 268.
- Abreu, R.C.R.; Hoffmann, W.A.; Vasconcelos, H.L.; Pilon, N.A.; Rossatto, D.R.; Durigan, G. The biodiversity cost of carbon sequestration in tropical savanna. Sci. Adv. 2017, 3, e1701284–1701284. [Google Scholar] [CrossRef]
- Durigan, G.; Leitão Filho, H.F.; Rodrigues, R.R. Phytosociology and stricture of a frequently burnt cerrado vegetation in SE- Brazil. Flora 1994, 189, 153–160. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Sanford, R.L.; Cummings, D.L.; Salcedo, I.H.; Sampaio, E.V.S.B. Biomass and Nutrient Dynamics Associated with Slash Fires in Neotropical Dry Forests. Ecology 1993, 74, 140–151. [Google Scholar] [CrossRef]
- Koch, A.; Brierley, C.; Maslin, M.M.; Lewis, S.L. Earth system impacts of the European arrival and Great Dying in the Americas after 1492. Quat. Sci. Rev. 2019, 207, 13–36. [Google Scholar] [CrossRef]
- Almeida, A.Q.; Mello, A.A.; Neto, A.L.D.; Ferraz, R.C. Relações empíricas entre características dendrométricas da Caatinga brasileira e dados TM Landsat 5. Pesqui. Agropecuária Bras. 2014, 49, 306–315. [Google Scholar] [CrossRef]
- Lopes, J.F.B. Deposição e decomposição de serapilheira em área da Caatinga. Revista Agro@mbiente On-line Boa Vista 2009, 3, 72–79. [Google Scholar] [CrossRef]
- Moreira, A.G. Effects of fire protection on savanna structure in Central Brazil. J. Biogeogr. 2000, 27, 1021–1029. [Google Scholar] [CrossRef]
- Lapsley, J.; Hayes, G.M.; Janvier, V. Influence of locoregional lymph node aspiration cytology vs sentinel lymph node mapping and biopsy on disease stage assignment in dogs with integumentary mast cell tumors. Vet. Surg. 2020, 1–9. [Google Scholar] [CrossRef]
- Líbano, A.M.; Felfili, J.M. Mudanças temporais na composição florística e na diversidade de um cerrado sensu stricto do Brasil Central em um período de 18 anos (1985–2003). Acta Bot. Bras. 2007, 20, 927–936. [Google Scholar] [CrossRef]
- Roitman, I.; Bustamante, M.M.C.; Haidar, R.F.; Shimbo, J.Z.; Abdala, G.C.; Eiten, G.; Fagg, C.W.; Felfili, M.C.; Felfili, J.M.; Jacobson, T.K.B.; et al. Optimizing biomass estimates of savanna woodland at different spatial scales in the Brazilian Cerrado: Re-evaluating allometric equations and environmental influences. PLoS ONE 2018, 13, e0196742. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt-Römermann, M.; Baeten, L.; Craven, D.; De Frenne, P.; Hédl, R.; Lenoir, J.; Bert, D.; Brunet, J.; Chudomelová, M.; Decocq, G.; et al. Drivers of temporal changes in temperate forest plant diversity vary across spatial scales. Glob. Chang. Biol. 2015, 21, 3726–3737. [Google Scholar] [CrossRef]
- Altomare, M.; Vasconcelos, H.L.; Raymundo, D.; Lopes, S.; Vale, V.; Prado-Junior, J. Assessing the fire resilience of the savanna tree component through a functional approach. Acta Oecologica 2021, 111, 103728. [Google Scholar] [CrossRef]
- Sato, M.N.; Garda, A.A.; Miranda, H.S. Fire effects in the mortality rate of woody vegetation in Central Brazil. Proceedings of the 3rd International Conference on Forest Fires Research Coimbra. 1998.
- Ramos, P.C.M. Sistema nacional de prevenção e combate aos incêndios florestais. In Fórum Nacional Sobre Incêndios Florestais; IPEF: 1995; pp. 29–58.
- Miranda, S.C. Variação espacial e temporal da biomassa vegetal em áreas de Cerrado. Tese (Doutorado em Ecologia). Universidade de Brasília, 2013.
- Bond, W.J.; Keeley, J.E. 2005. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Poorter, L. Bark thickness and fire regime. Funct. Ecol. 2015, 29, 315–327. [Google Scholar] [CrossRef]
- Araújo, F.D.; Tng, D.Y.; Apgaua, D.M.; Coelho, P.A.; Pereira, D.G.; Santos, R.M. Post-fire plant regeneration across a closed forest-savanna vegetation transition. For. Ecol. Manag. 2017, 400, 77–84. [Google Scholar] [CrossRef]
- Luke, R.; Mcarthur, A. Bushfire in Australia. Australian Government Publishing Service, Canberra. 1978.
- Maracahipes, L.; Marimon, B.S.; Lenza, E.; Marimon-Junior, B.H.; de Oliveira, E.A.; Mews, H.A.; Gomes, L.; Feldpausch, T.R. Post-fire dynamics of woody vegetation in seasonally flooded forests (impucas) in the Cerrado-Amazonian Forest transition zone. Flora 2014, 209, 260–270. [Google Scholar] [CrossRef]
- Silveira, F.A.O.; Ordóñez-Parra, C.A.; Moura, L.C.; Schmidt, I.B.; Andersen, A.N.; Bond, W.; Buisson, E.; Durigan, G.; Fidelis, A.; Oliveira, R.S.; et al. Biome Awareness Disparity is BAD for tropical ecosystem conservation and restoration. J. Appl. Ecol. 2021, 59, 1967–1975. [Google Scholar] [CrossRef]
- Pausas, J.G.; E Keeley, J. Wildfires and global change. Front. Ecol. Environ. 2021, 19, 387–395. [Google Scholar] [CrossRef]
- Hagmann, R.K.; Hessburg PFHagmann, P.F.; Hessburg, R.B.; Salter, A.G.; Merschel, M.J. Contemporary wildfires further degrade resistance and resilience of fire-excluded forests. For. Ecol. Manag. 2022, 506, 119975. [Google Scholar] [CrossRef]
- Mattsson, M.O.; Simkó, M. Grouping of experimental conditions as an approach to evaluate effects of extremely low-frequency magnetic fields on oxidative response in in vitro studies. Front. Public Health 2014, 2, 132. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Spies, T.A.; Perry, D.A.; Skinner, C.N.; Taylor, A.H.; Brown, P.M.; Stephens, S.L.; Larson, A.J.; Churchill, D.J.; Povak, N.A.; et al. Tamm Review: Management of mixed-severity fire regime forests in Oregon, Washington, and Northern California. For. Ecol. Manag. 2016, 366, 221–250. [Google Scholar] [CrossRef]
- Aka, G.M.; Elogne, C.P.; Irie, C.; Zo-bi Bienvenu, H.K.; Amani, V.V.D.M. , Bruno, H. Life after fire - Long-term responses of 20 timber species in semi-deciduous forests of West Africa. For. Ecol. Manag 2023, 538, 120977. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Nogueira, E.M.; Nelson, B.W.; Fearnside, P.M.; França, M.B.; de Oliveira, C.A. Tree height in Brazil's ‘arc of deforestation’: Shorter trees in south and southwest Amazonia imply lower biomass. For. Ecol. Manag. 2008, 255, 2963–2972. [Google Scholar] [CrossRef]
- Alves, R.J.V.; Kolbek, J. Can campo rupestre vegetation be floristically delimited based on vascular plant genera? Plant Ecol. 2009, 207, 67–79. [Google Scholar] [CrossRef]
- Paiva, A.O.; Rezende, A.V.; Pereira, R.S. Estoque de carbono em cerrado sensu stricto do Distrito Federal. 2011, 35, 527–538. [CrossRef]
- Felfili, J.M.; Rezende, R.P. Conceitos e métodos em fitossociologia. Brasília: Universidade de Brasília, Departamento de Engenharia Florestal. Comunicações Técnicas Florestais 2003, 5, 1–68. [Google Scholar]
- Cianciaruso, M.V.; da Silva, I.A.; Batalha, M.A. Aboveground biomass of functional groups in the ground layer of savannas under different fire frequencies. Aust. J. Bot. 2010, 58, 169–174. [Google Scholar] [CrossRef]
- Ramos-Neto, M.B.; Pivello, V.R. Lightning Fires in a Brazilian Savanna National Park: Rethinking Management Strategies. Environ. Manag. 2000, 26, 675–684. [Google Scholar] [CrossRef]
Figure 1.
Study area location: Água Limpa Farm (FAL). The studied treatments (plots) are delimited in the google earth images (2021), as following: Area 1 (in pink) = annual fire; Area 2 (in blue) = legacy fire; Area 3 (in yellow) = control (fire-free area for the past 30 years).
Figure 1.
Study area location: Água Limpa Farm (FAL). The studied treatments (plots) are delimited in the google earth images (2021), as following: Area 1 (in pink) = annual fire; Area 2 (in blue) = legacy fire; Area 3 (in yellow) = control (fire-free area for the past 30 years).
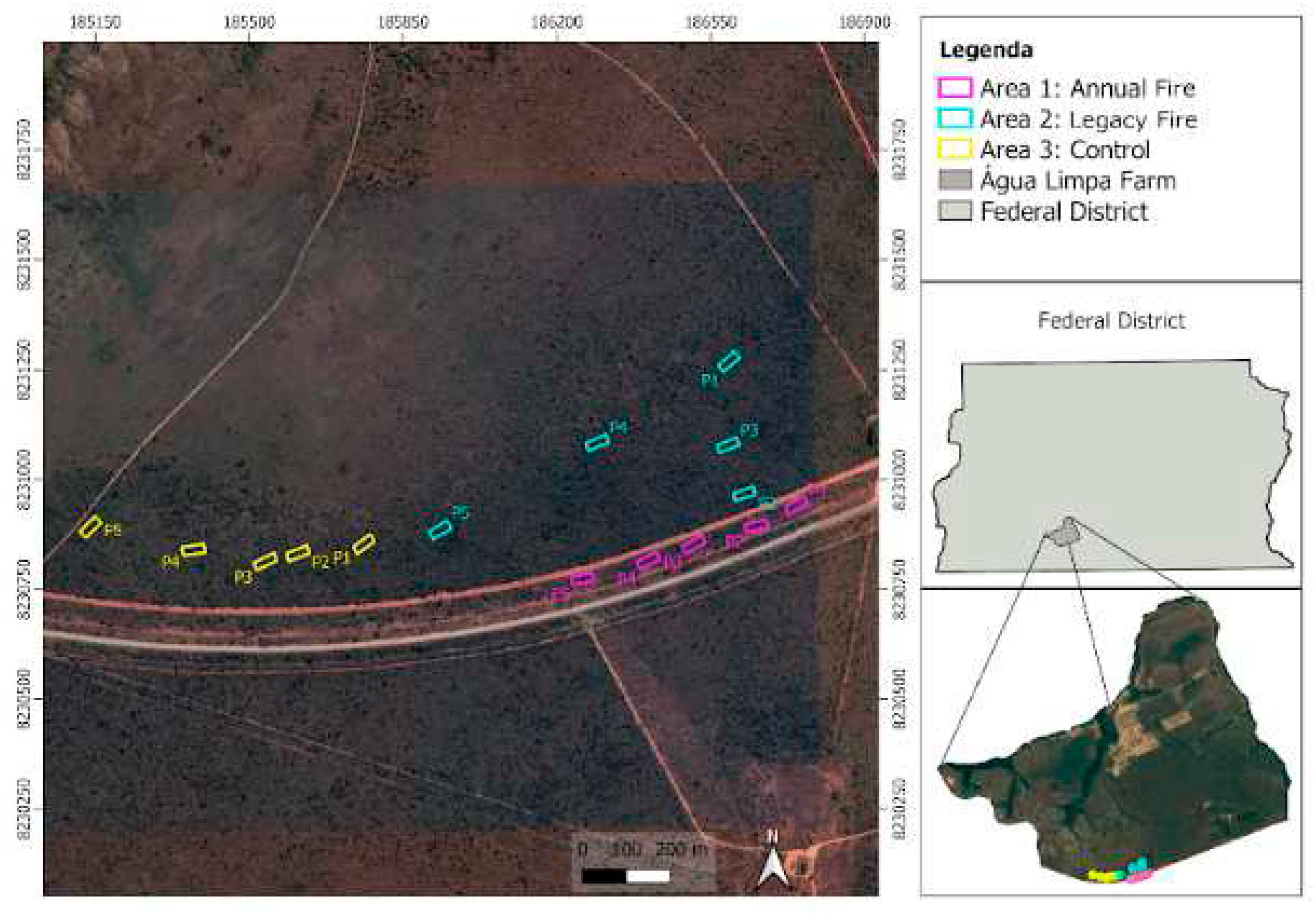
Figure 2.
Structure, richness, diversity (Shannon-Wiener), and evenness of the tree community in cerrado stricto sensu at Fazenda Água Limpa, DF. (a) Shannon diversity, (b) species richness, (c) Pielou evenness, and (d) number of woody individuals per plot in the Brazilian Cerrado. The boxplots show the median bands representing the median values of the variables, the upper and lower indicate the first and third quartiles. The bars indicate the minimum and maximum values. Dots denote outliers.
Figure 2.
Structure, richness, diversity (Shannon-Wiener), and evenness of the tree community in cerrado stricto sensu at Fazenda Água Limpa, DF. (a) Shannon diversity, (b) species richness, (c) Pielou evenness, and (d) number of woody individuals per plot in the Brazilian Cerrado. The boxplots show the median bands representing the median values of the variables, the upper and lower indicate the first and third quartiles. The bars indicate the minimum and maximum values. Dots denote outliers.
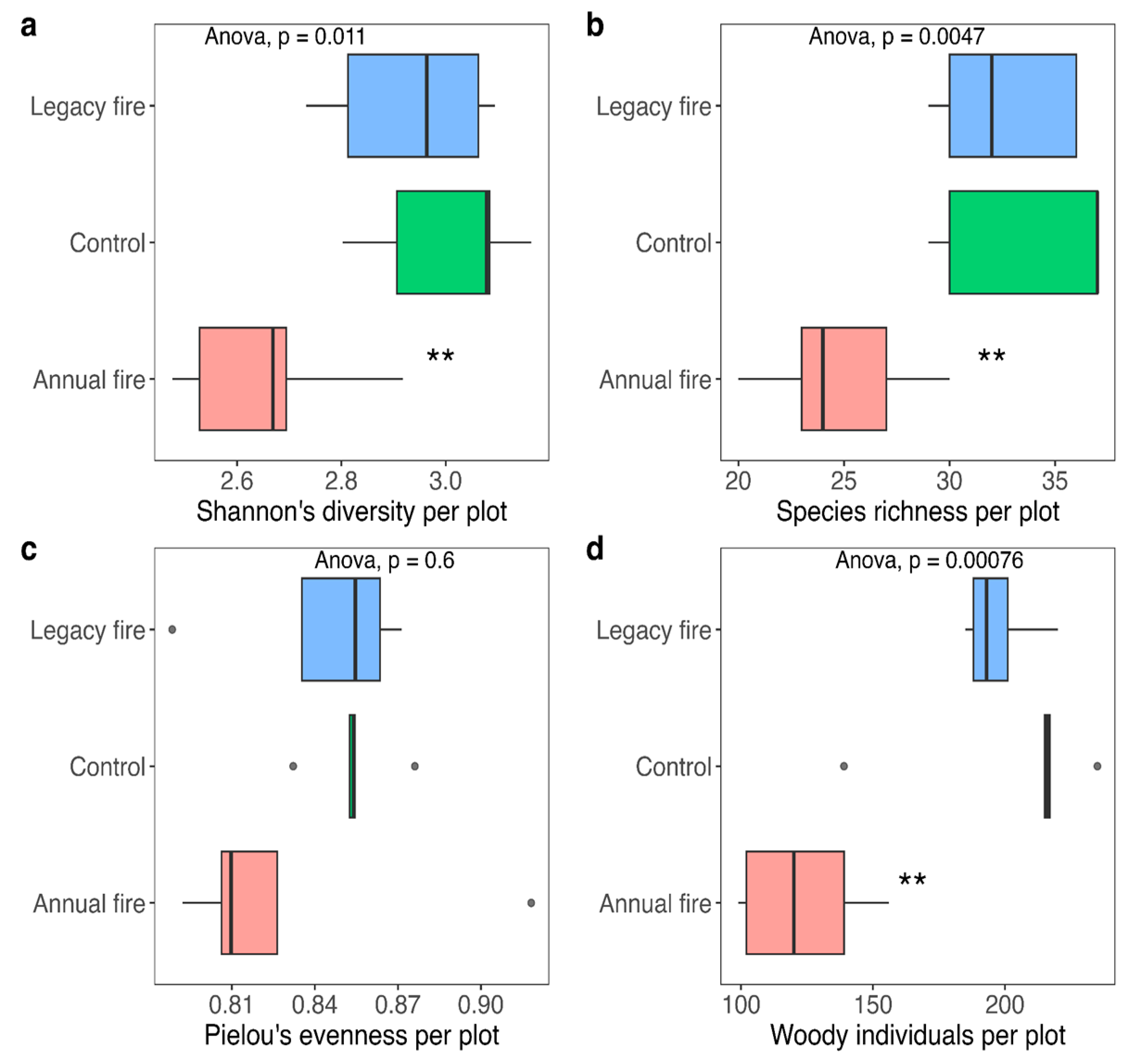
Figure 3.
(a) Structural differences in vegetation between treatments under the effects of fire, number of dead individuals, (b) diameter classes, and (c) height classes in the three evaluated treatments.
Figure 3.
(a) Structural differences in vegetation between treatments under the effects of fire, number of dead individuals, (b) diameter classes, and (c) height classes in the three evaluated treatments.
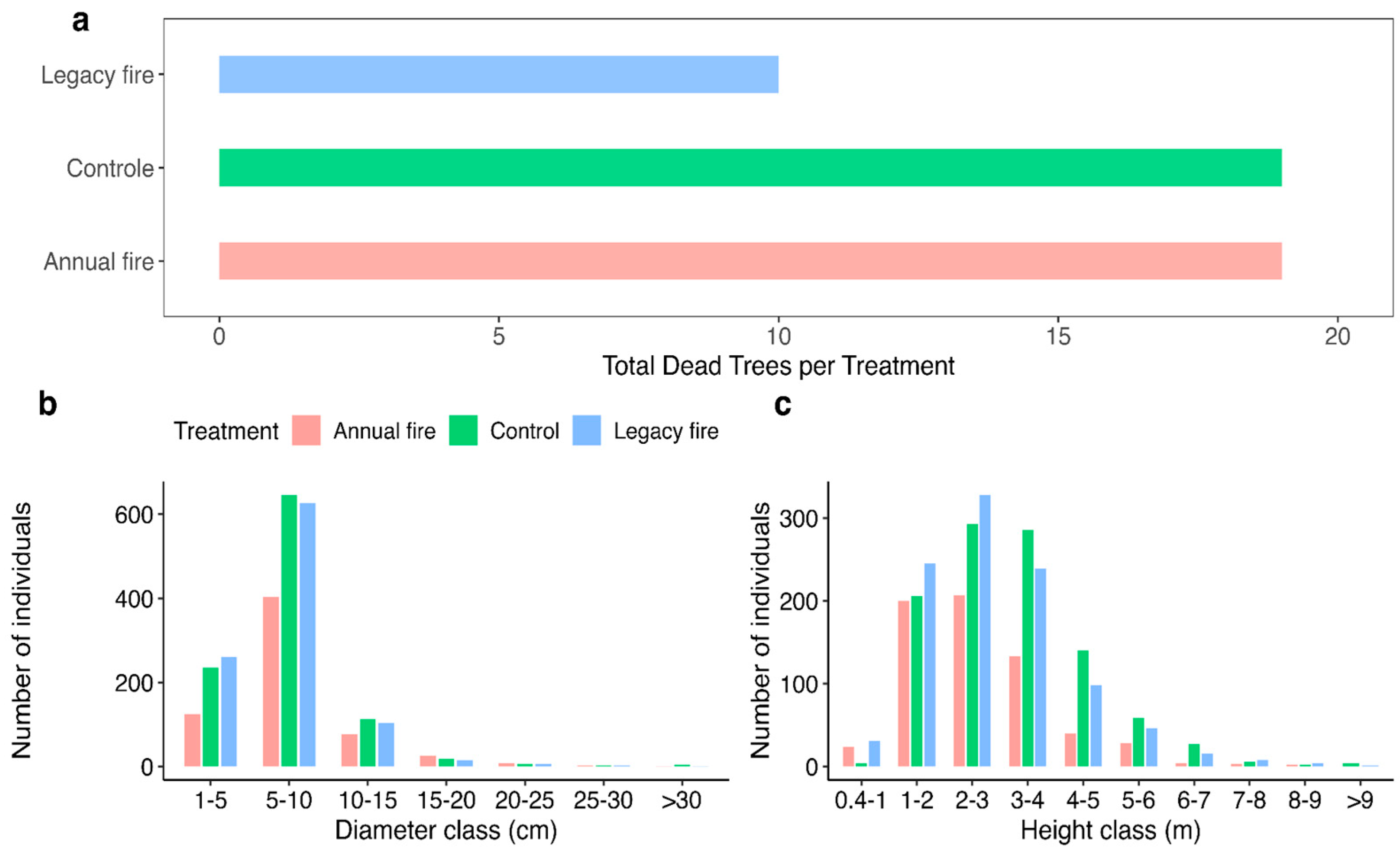
Figure 4.
Estimated live shrub-tree aboveground carbon stocks (in MgC/ha) per treatment.
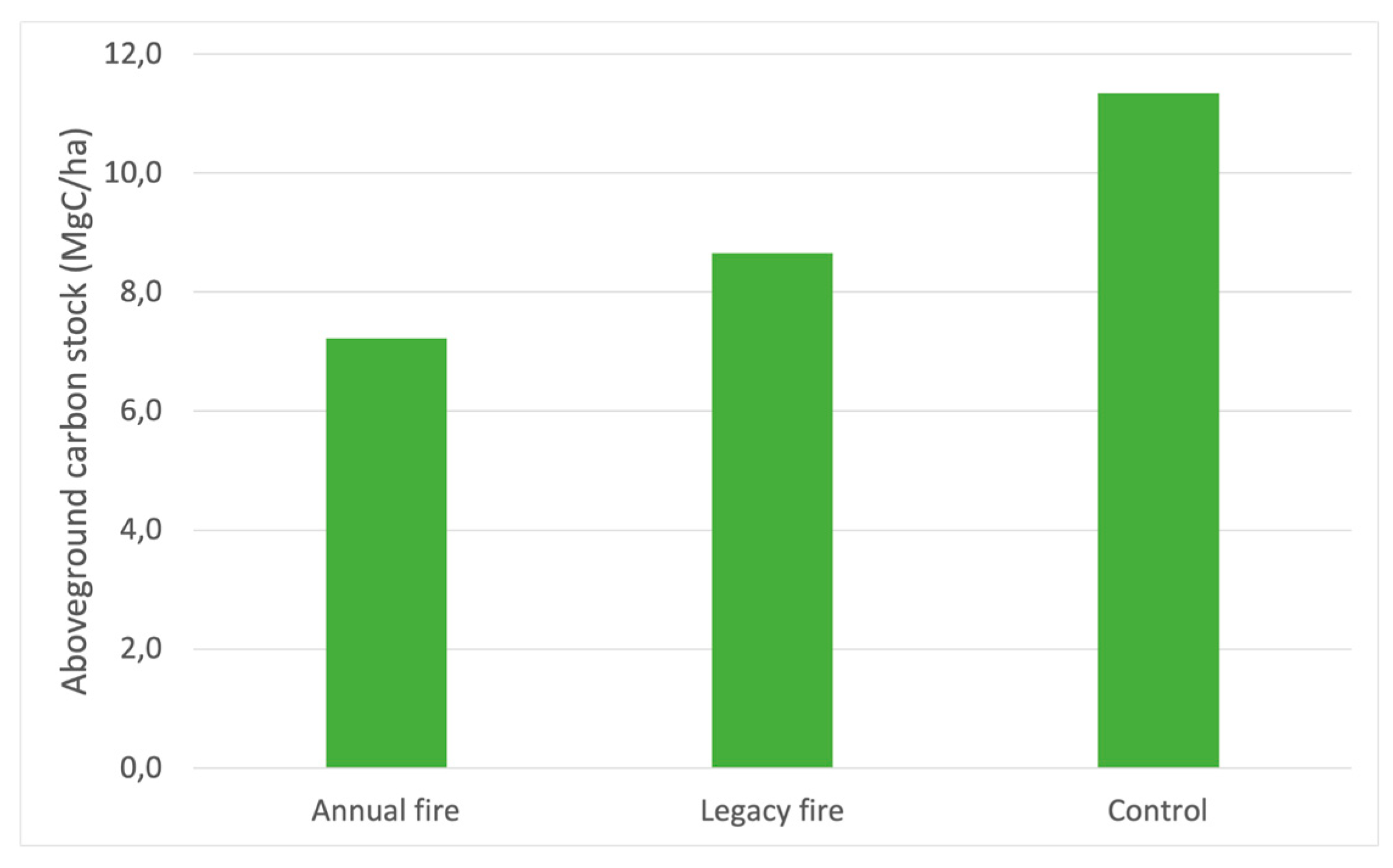
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



