
Submitted:
01 August 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
Urinary tract infections (UTIs) are the second most common bacterial infection with high recurrence rates. A major causative agent of UTIs is uropathogenic Escherichia coli (UPEC), which can form persistent biofilms on urothelial surfaces and/or urinary catheters. Biofilms are inherently tolerant to antimicrobials, making them difficult to eradicate. Ciprofloxacin hybrids functionalised with nitroxides have previously shown enhanced biofilm eradication activity in vitro. Here, we evaluate a ciprofloxacin di-nitroxide hybrid (CDN11), previously reported to have antibiofilm activity against UPEC, as a potential UTI therapeutic in various infection models. We report potent CDN11 activity against mature biofilms formed by cystitis strain UTI89 inside polyethylene catheters. In infected human bladder cell monolayers, CDN11 afforded a 3-log reduction in UPEC colony forming units (CFU) compared to controls, including intracellular bacteria. Infected mouse bladders carrying biofilm-like intracellular UPEC reservoirs had reduced bacterial burdens following treatment with CDN11 ex vivo. Activity trends for CDN11 were comparable throughout all models, indicating that our testing pipeline was robust. New antimicrobial development is arduous and requires large investments of time and money. The pipeline described here could be readily used in testing new compounds, fast tracking the development of novel UTI therapeutics.
Keywords:
bladder
; urinary tract infection
; UPEC
; bacterial biofilms
; catheters
; nitroxides
; antibiotic resistance
; fluoroquinolones
; therapeutics
1. Introduction
Urinary tract infections (UTIs) are one of the most common bacterial infections with a high rate of recurrence (~30%). UTI clinical syndromes include cystitis, pyelonephritis, urosepsis and asymptomatic bacteriuria (ASB) [1]. Hospital-acquired UTIs represent high-burden nosocomial infections, especially in patients undergoing long periods of urinary catheterisation, known as catheter-associated UTI (CAUTI) [2]. Uropathogenic Escherichia coli (UPEC) are the major causative agents of UTIs. UPEC are Gram-negative rod-shaped bacteria that attach to urinary epithelial cells and are capable of invading into bladder cells and persisting intracellularly to evade host responses [3]. UPEC persistence during UTI can be attributed to a number of virulence factors, such as adhesins, flagella, siderophores, and the ability to form biofilms [3]. Biofilms are sessile multicellular communities of bacteria embedded in a self-produced extracellular matrix, capable of forming on both biotic and abiotic surfaces. In UTI, UPEC forms intracellular bacterial communities (IBCs) inside bladder cells during early stages of the infection [4], or in the case of CAUTI, UPEC forms biofilms inside the lumen of indwelling urinary catheters [5,6]. Biofilms are inherently tolerant to environmental stress, antimicrobials, and host immune responses, making them difficult to eradicate and allowing bacteria to persist [7-10]. UPEC IBCs have similar molecular attributes to biofilms, and their establishment and maintenance in the bladder requires the production of type 1 pili, antigen 43, and a polysaccharide-rich extracellular matrix [4,11-13]. Following IBC formation during acute UTI, UPEC can persist in urinary tissues for longer periods in the form of intracellular quiescent reservoirs (QIRs) or in the case of CAUTI, as residual (persister) bacterial cells remaining attached on the catheter surface after antibiotic therapy; both forms share characteristics with biofilms and are intrinsically difficult to eradicate [14-16].
Nitric oxide (NO) has been reported to have broad-spectrum activity against biofilms (bacterial and fungal) and also in combination with antibiotics providing one of the most promising avenues for clinical therapy [17,18]. However, NO’s gaseous and highly reactive nature makes it difficult to work with, or to control the dose and timing of its delivery which can lead to NO-mediated cytotoxicity and off-target effects [17,19], Recently the evaluation of nitroxides as stable NO mimics has uncovered their potential as alternative biofilm dispersal agents [18]. Similar to NO, nitroxides were shown to reduce biofilm biomass of various Gram-negative species in vitro [20]. As nitroxides are stable (unlike NO) and thus more amenable to chemical manipulation, they have been successfully used in the synthesis of functionalised antibiotics and biocides [21-24], ensuring co-delivery with antimicrobials. Some nitroxide-antibiotic hybrids were reported to have improved biofilm eradication activity compared to the parent drug, with promising activity reported against several clinically important Gram-negative and Gram-positive bacteria in vitro [21,24]. Biofilm eradication by these hybrid drugs is likely achieved by simultaneous bacterial dispersal and killing (induced by the nitroxide and antibiotic/biocide moieties, respectively) making them highly suitable for clinical applications [25], as using biofilm dispersal agents alone (or even in combination with antibiotics, but unlinked) may pose a risk for the seeding of new infection sites by viable dispersed cells [26].
Here, we evaluate CDN11, a ciprofloxacin-dinitroxide hybrid previously reported to have antibiofilm activity against E. coli [24], as a potential UTI therapeutic. Ciprofloxacin is a commonly prescribed fluoroquinolone antibiotic for both uncomplicated and complicated UTIs [27], with diminishing efficacy over the past decade due to the rise of fluoroquinolone resistant UPEC lineages [28,29]. We report that CDN11 displays antibiofilm activity against ciprofloxacin sensitive UPEC in multiple relevant models of UTI, including in vitro infected catheters and human bladder cells. Activity was also observed against UPEC IBCs in the murine bladder following ex vivo treatment. Our findings showcase nitroxide-antibiotic hybrids as a promising drug co-administration approach targeting common biofilm infections. Moreover, the preclinical pipeline of in vitro and in vivo UTI models employed in this study, should serve as a robust platform for testing new UTI therapeutics in future studies.
2. Materials and Methods
2.1. Ethics Statement
All UTI mouse work was conducted under ethics approval number A1703-600M issued by the Animal Ethics Committee at the QIMR Berghofer Medical Research Institute.
2.2. Bacterial Strains and Culture Conditions
UPEC strain UTI89 [30] was used in this study as a reference cystitis strain, and EC958 [31] as a multi-drug resistant strain. Unless otherwise stated, all bacteria were routinely maintained and cultured in LB media at 37°C, 200 rpm (1% w/v tryptone (Oxoid, CM0087) 0.5% w/v yeast extract (Gibco, 212750), 0.5% w/v salt) from a single colony on a plate (1.5% w/v agar (BD, DF0140-01-0)), 37°C. For all antimicrobial activity assays, tested was conducted in MH medium (30% w/v dehydrated beef infusion, 1.75% w/v casein hydrolysate, 0.15% w/v starch, Oxoid, CM0405).
2.3. Antimicrobial Compounds
Ciprofloxacin (Sigma, 17850) stocks were routinely made from powder and dissolved in 0.1 M HCl (30 mM, 10 mg/mL). For MIC assays, ciprofloxacin was used at a concentration range of 3000 μM – 0.0023 μM. For MBEC assays, ciprofloxacin was used at a concentration range of 800 μM - 1.57 μM. For all other assays, ciprofloxacin was used at a concentration of 400 μM.
CDN11 was synthesised as previously described [24]. Briefly, EDC-promoted amide coupling of ciprofloxacin ethyl ester with Boc-Lys(Cbz)-OH gave the fully protected conjugate which was deprotected by hydrogenolysis at the α-amine and reacted in another EDC amide coupling reaction with Boc-Lys(Cbz)-OH. Deprotection of the two Cbz-protected amines and subsequent amidation with CTEMPO gave the nitroxide functionalised ciprofloxacin, which was deprotected by base mediated ester hydrolysis, followed by Boc-deprotection to give CDN11. Stocks were routinely made from powder and dissolved in 100% DMSO (10.5 mM, 10 mg/mL). For MIC assays, CDN11 was used at a concentration range of 800 μM – 2.63 μM. For MBEC assays, CDN11 was used at a concentration range of 800 μM - 1.57 μM. For all other assays, CDN11 was used at a concentration of 400 μM.
2.4. Minimum Inhibitory Concentration (MIC) Assay
MICs for nitroxides, antibiotics, and hybrid compounds were as described in Clinical and Laboratory Standards Institute (CLSI) [32]. Briefly, in a 96-well plate, twofold serial dilutions of each compound were prepared to a final volume of 100 μL in MH medium (CDN11: 800 μM - 2.63 μM, CIP: 3000 μM – 0.0023 μM), with a matched DMSO vehicle control for the highest concentration. Each well was inoculated with 100 μL bacteria suspension in MH containing 5 x 106 bacterial colony forming units (CFUs), prepared from overnight cultures in LB. The MIC for a compound was defined as the lowest concentration that prevented visible bacterial growth after 18 h of static incubation at 37 °C. MICs were performed a minimum of two times with three technical replicates per condition.
2.5. Minimum Biofilm Eradication Concentration (MBEC) Assay
MBEC assays were performed as per the manufacturer’s protocol with a Calgary biofilm device (CBD) (Innovotech Inc., Canada). The CBD consists of a 96-well plate and an associated lid consisting of small protrusions (pegs) that are immersed in each well when the lid is covered, facilitating individual biofilm growth on each peg. Briefly, UTI89 grown for 18h in LB were diluted to 1 x 107 CFU/mL in LB and used to inoculate the plate with 130 μL of culture per well. The CBD was incubated for 24 hours with shaking (150 rpm) at 37°C in 95% relative humidity. Following 24 h of growth, biofilms were washed once in PBS (130 µL) by incubating statically for 10 s to remove planktonic cells and then the peg lid was transferred to a new 96-well plate containing ciprofloxacin and CDN11 in MH, in concentrations ranging from 800 μM - 1.57 μM, with a matched HCl/DMSO vehicle control for the highest concentration, respectively. The plate was then incubated for a further 24 hours at 37°C with shaking (150 rpm), then washed once in PBS (200 µL, 10 s static incubation) and sonicated in a water bath (Elma Ultrasonic Cleaner S10H) for 20 min at 20°C to suspend the biomass in 200 µL of PBS. Each well was serially diluted 10-fold and 5 μL was spotted onto LB agar plates in triplicate to determine CFU recovered from each peg biofilm. Plates were incubated overnight at 37°C with colonies counted the following day to obtain log10(CFU/mL) values for each treatment condition. Biofilm experiments were performed a minimum of two times, with 3-8 pegs per group. MBEC values are reported as the concentration at which there are no recoverable biofilm CFU (complete eradication).
2.6. Catheter Biofilm Model
The in vitro catheter biofilm assays were performed similar to the MBEC assay. UTI89 grown for 18 h in LB were diluted to 1 x 107 CFU/ml in LB and 400 μL/well used to inoculate a 48-well plate. Each well contained catheters made from ~3 mm sections of UV sterilised polyethylene tubing (BD, 0.28 mm inner diameter, 0.61 mm outer diameter). Plates were incubated for 24 hours with shaking (150rpm) at 37°C in 95% relative humidity. Following 24 hours of growth, catheters were washed once in PBS (400 μL) to remove planktonic bacterial cells and then were transferred to a new plate containing CTEMPO, ciprofloxacin, and CDN11 in MH, at a concentration of 400 μM each, with matched vehicle controls. Catheters were then incubated for a further 24 hours at 37°C shaking (150 rpm), and then washed once in PBS (400 µL) and sonicated and processed as above to determine CFU.
2.7. Human Cell Infection Assays
Type 1 fimbriae (T1F) enriched, UTI89 was routinely prepared cultured from highly fimbriated stocks, which were incubated overnight statically at 37°C for 18 h. Bladder epithelial cells T24 (ATCC HTB-4), were maintained in Roswell Park Memorial Institute (RPMI) 1640 medium (Invitrogen) supplemented with 5% heat-inactivated foetal bovine serum (Invitrogen) at 37°C. Adhesion and invasion assays with UPEC strain UTI89 were performed as previously described [33]. Briefly, T24 cells were seeded at a density of 5 x 105 in 24-well plates, and then grown overnight to form confluent monolayers. Confluent T24 monolayers were infected with UTI89 for 1 hour at 37°C at a multiplicity of infection (MOI) of 10 (5 x 106 CFU/well), then washed with PBS. For adhesion assays, monolayers were then incubated with either RPMI only (untreated), DMSO/0.1M HCl (vehicle controls), 400 μM ciprofloxacin (antibiotic) or 400 μM CDN11 (dinitroxide-ciprofloxacin antibiotic hybrid) for 2 hours at 37°C. For invasion assays, infected monolayers as above were first incubated with 200 μg/mL gentamicin for 1 hour to eliminate extracellular bacteria, then washed three times with PBS. Monolayers were then treated with the same compound groups as the adhesion assays. Post treatment, both monolayers were washed three times with PBS followed by lysis with 500 μL 0.1% Triton X-100 in PBS for 10 min at 37°C. CFU recovered from each monolayer were determined by serial dilution plating onto LB agar plates. All compound working solutions were made up in RPMI, to a concentration of 400 μM (40x MIC of CDN11).
2.8. In Vivo UTI Mouse Model and Ex Vivo Bladder Antibiotic Treatment
6–7-week-old female C3H/HeJ mice were catheterised as previously described [34]. Briefly, mice were anaesthetised by isoflurane inhalation and catheterised with ~1-2 x 108 CFU in 30 μL of UTI89 in PBS. T1F enriched UTI89 was prepared as above. The prepared inoculum was deposited directly into the bladder using a sterile polyethylene catheter (same material as in vitro testing) followed by immediate removal [34]. C3H/HeJ mice were sourced from the ARC (Animal Resource Centre, Western Australia). Mice were sacrificed at 16 hours post inoculation and the bladders were extracted.
Bladders were bisected to expose the lumen and were washed in PBS for 5 min in a well of a 96-well plate (one mouse per well). Bladder halves were then transferred to fresh wells in another plate containing 200 μg/mL gentamicin in PBS and incubated for 1 hour 37°C to remove extracellular bacteria, then washed again in PBS. Each bladder half was then incubated with either PBS only (untreated), DMSO (vehicle control), 400 μM ciprofloxacin (antibiotic) or 400 μM CDN11 (nitroxide functionalised antibiotic) for 6 hours at 37°C. Following treatment, bladders were immediately transferred to tubes containing metal beads for homogenisation. Bladders were homogenised in 50 μL of PBS using a Mini Bead beater (BioSpec Products) before topping up to 1 mL with PBS and aliquoting 200 μL of each sample into a 96-well plate. Samples were serially diluted and 5 µL of each well for each sample was plated onto LB agar in quadruplicate and then incubated overnight at 37°C. The following day, colonies were enumerated, and bacterial CFU was expressed as log10(CFU/0.1g) of tissue.
2.9. Statistical Analysis
All statistical analysis was performed in GraphPad Prism Version 9 software (GraphPad Software). An ordinary one-way ANOVA with Dunn’s multiple comparisons correction was used to test for statistically significant differences between CDN11-treated groups and control group means. Statistical significance was set at p <0.05.
3. Results
3.1. CDN11 Eradicates UPEC Biofilms Formed on Urinary Catheters In Vitro
Dinitroxide-ciprofloxacin hybrid CDN11 was synthesised as previously described [24]. The minimum inhibitory concentration (MIC) and minimum biofilm eradication concentration (MBEC) of CDN11 against reference cystitis UPEC isolate UTI89 was confirmed as ≤10.5 μM and ≤400 μM, respectively, in agreement with previous reports [24]. To simulate UPEC biofilm formation in the context of CAUTI, mature biofilms of UTI89 were established at high cell density inside polyethylene catheters and then treated with antibiotics. After 24 hours of treatment with CDN11 at 400 μM (MBEC concentration), viable CFU remaining in the biofilm were reduced by >4 logs compared to vehicle treated (DMSO) and untreated biofilm controls (Figure 1). Thus, CDN11 antibiofilm activity in the catheter biofilm treatment model exceeded the 99.9% reduction in CFU cut-off used to define biofilm eradication in vitro and was significantly better than ciprofloxacin administered at the same concentration (p = <0.0001, Figure 1).
3.2. In Vitro Treatment of Infected Human Bladder Cells with CDN11 Reduces Extracellular and Intracellular UPEC Titers
We next tested if CDN11 could effectively control UPEC infection of human cells in vitro. Human T24 bladder epithelial cell monolayers infected with UPEC strain UTI89 for 1 hour at a multiplicity of infection (MOI) of 10 were treated for 2 hours with antibiotics. CDN11 at 400 μM significantly reduced the total CFU recovered from infected monolayers compared to untreated and vehicle treated controls (mean reduction of ≥3.5 logs, Figure 2A). Similarly, ciprofloxacin afforded a significant reduction of the total UPEC titer and cleared infection in one of eight treated monolayers (Figure 2A).
UPEC infection of T24 human bladder cells involves both attachment (extracellular bacteria) and invasion (intracellular bacteria) of bladder cells, resembling UPEC pathogenesis during bladder infection (cystitis) of mice and humans [35]. To specifically test CDN11 activity against intracellular UPEC reservoirs, we performed gentamicin protection assays [33], whereby the infected T24 cell monolayers were first treated with gentamicin to selectively eliminate extracellular UPEC followed by test antibiotics. Despite its large molecular size (>900 Da), CDN11 displayed activity against intracellular UPEC when administered at 400 μM, reducing the mean intracellular titer by ~40% compared to untreated and vehicle controls and clearing infection in one of five treated monolayers (Figure 2B). Ciprofloxacin-treated monolayers were completely cleared of intracellular UPEC, with no detectable CFU recovered from all monolayers examined (Figure 2B).
3.3. Ex Vivo Treatment of Infected Mouse Bladders with CDN11 Reduces Intracellular UPEC Reservoirs
To evaluate early preclinical efficacy for CDN11 as a UTI therapeutic, we next tested its activity against intracellular UPEC reservoirs that are established in the bladder of mice during experimental acute UTI. Infected bladders collected from female C3H/HeJ mice with acute cystitis established by UPEC strain UTI89 were treated ex vivo with gentamicin to eliminate extracellular (luminal) UPEC, followed by treatment with test antibiotics (Figure 3A). CDN11 treatment at 400 μM completely cleared UPEC from 25% of the infected bladder tissues, and significantly reduced the mean intracellular CFU of the group compared to untreated controls (Figure 3B). In comparison, ciprofloxacin treatment cleared intracellular UPEC from all bladder tissues examined (Figure 3B).
3.4. CDN11 Displays Activity against Biofilms by fluoroquinolone-Sensitive not Fluoroquinolone Resistant UPEC
UTIs are increasingly caused by multidrug resistant UPEC lineages [28]. In particular, E. coli sequence type 131 (ST131) is the most recent globally disseminated lineage of multidrug resistant (MDR) UPEC causing UTIs and bloodstream infections, with most ST131 strains being resistant to fluoroquinolones and beta-lactam antibiotics. To investigate whether CDN11 displays antibiofilm activity against MDR UPEC we compared its activity against UTI89 (ciprofloxacin sensitive) and EC958 biofilms, the latter being a representative ST131 MDR UPEC strain (ciprofloxacin resistant) [36]. While CDN11 displayed biofilm CFU reduction against UTI89 in a dose-dependent manner and at concentrations as low as 12.5 μM, it did not impact established EC958 biofilms even at the highest concentration tested (Figure 4). As EC958 contains both chromosomal and plasmid-encoded ciprofloxacin resistance genes, this finding suggests that CDN11 targets the same enzymes as ciprofloxacin (parent drug) in both planktonic and biofilm bacteria [37].
4. Discussion
This study evaluated a new hybrid antibiotic as a UTI therapeutic in various preclinical models that replicate clinically important aspects of the disease. Currently, the only therapeutics that treat the causative agents of UTI are antibiotics. Biofilms formed on patient catheters however are difficult to treat with antibiotics alone, and are often hard to eradicate due to high antibiotic tolerance [38]. During UTI, the formation of biofilms and intracellular bacterial reservoirs contribute to disease persistence and recurrence. Greater emphasis should be placed on targeting biofilms for treating high-burden infections like UTIs. CDN11 is an antibiotic conjugate designed to target biofilm bacteria through its nitroxide moieties that were previously demonstrated to display biofilm dispersal activity [20,39]. Here we report that administration of CDN11 can effectively control UPEC infection in relevant preclinical models, significantly reducing the overall bacterial burden associated with infection on both biotic and abiotic surfaces, and in some cases even clearing UPEC from infected cells and tissues. Our study is the first to showcase the potential of this hybrid antibiotic as a new UTI therapeutic.
The Calgary biofilm device (biofilms cultured on pegs) is widely used as a simple high-throughput assay to assess antibiofilm activity of new potential therapeutics [40]. Using this assay, we determined the MBEC value of CDN11 as the minimum concentration required to fully eradicate UPEC biofilms (i.e. kill 100% of cells), not reduce 99.9% of the cells compared to growth controls that is defined as the biofilm bactericidal concentration (BBC) and is often used interchangeably with MBEC [41]. This concentration guided our downstream testing in biofilm models of UPEC in the context of UTI. We could translate our findings from the peg model to the catheter model, which more closely replicates aspects of UTI disease, such as biofilm formation on patient catheters in CAUTI. Using concentrations and treatment durations derived from the peg model, we noted similar trends between in vitro models. While catheter biofilms were not completely eradicated by CDN11 treatment at 1 x MBEC, the observed reduction in biofilm CFU was significant (over 4-logs), bringing remaining CFU very close to the limit of detection for this assay, which is characteristically low (~35 CFU/peg). The in vitro catheter model simulates some clinically relevant factors of CAUTI, such as replicating bacterial titers seen on patient catheters and physicochemical interactions with catheter materials. The latter makes it an especially useful model for testing different catheter coatings that may contain antimicrobial compounds to limit biofilm formation or disrupt established biofilms [42]. However, the catheter model has limitations in representing important host factors during CAUTI, such as fibrinogen deposition and recruitment of inflammatory immune cells [16,43].
In order to replicate more host-pathogen interactions during in vitro testing, we included a human cell infection model. UPEC in vitro infection of human bladder cells is commonly employed in UPEC pathogenesis studies and has been shown to recapitulate the key initial steps in UPEC pathogenesis in the urinary tract: initial FimH-mediated adherence to epithelial cells followed by UPEC uptake into the cell and intracellular persistence [33,44,45]. Important host factors necessary for this initial adhesion step are the mannosylated uroplakins present on the surface of bladder epithelial cells [46], and proinflammatory cytokines IL-6 and IL-8, which are responsible for the recruitment of immune cells to the bladder [47,48], both of which can be recapitulated using the T24 bladder cell line [49,50]. During UTI, UPEC strains such as UTI89 are able to persist inside host cells, so by using the gentamicin protection assay model [33], we can test for compound activity specifically against intracellular bacteria. In these models, UPEC are mostly planktonic, but have been observed to form IBC-like structures [51]. While these have largely been observed in mouse UTI models and patient urine [4,52], several in vitro culture models have reported the presence of structures similar to IBCs [33,53]. CDN11 showed antimicrobial activity against both planktonic and biofilm-like UPEC in the human cell infection model, suggesting that it can penetrate mammalian cells to access intracellular bacteria, albeit less effectively than ciprofloxacin.
Cell culture models can also discern potential compound cytotoxicity, an important step before progressing to ex vivo and in vivo testing. In a previous study, CDN11 exhibited no cytotoxicity to T24 bladder cells even up to a concentration of 500 μM [24], which exceeds the concentration used here to treat infected T24 cells. We did however note some morphological changes to T24 monolayers (cell rounding) that appears to be due partly to the vehicle (DMSO) used, which has been previously observed in other studies using bladder and other cell lines [54,55]. While commonly used cytotoxicity assays like the lactate dehydrogenase (LDH) assay can inform about a compound’s mammalian cell toxicity, they are not indicative of alterations to cell-cell interactions. These are important in the context of many mucosal infections, including UTI, and have important implications for example in maintaining bladder tissue integrity during infection, which is important for limiting extensive UPEC invasion during UTI [56,57]. Investigating drug effects on cell-cell interactions in this model could screen out compounds that may disrupt the bladder architecture, before progressing them to in vivo testing.
In vivo infection models provide a powerful preclinical model for testing UTI therapeutics that replicate interactions between multiple tissues/cell types, UPEC (extracellular and intracellular reservoirs), the candidate compound, and host physiology. The mouse UTI model is widely used in the field to investigate host-pathogen interactions and potential drug candidates [34]. Here we treated infected mouse bladders for 6 hours with CDN11, to simulate the half-life of ciprofloxacin in the human body, while the dosage (1xMBEC; 400 μM) roughly equated to half the therapeutic ciprofloxacin dose commonly prescribed to adults for acute UTI [58,59]. The mouse UTI model can be adapted to study both acute and chronic UTI, and allows for many measurable outcomes, such as bacterial load in urine and urinary organs, immune responses, and tissue morphology changes. In the mouse UTI model, UPEC can be found as free-living (planktonic) extracellular bacteria, in an IBC (biofilm), or as filamentous cells upon exiting infected urothelial cells [4,60]. Using a model that incorporated filamentous UPEC, QIRs, recruited immune cells (which release nitric oxide), and the bladder’s tight cell layer architecture would provide insights into how the compound might interact with host responses and penetrate tissue layers, which would provide clinicians and researchers alike with more detailed information about a candidate compound’s activity [14,60,61]. We noted that throughout all our testing of CDN11, it retained antibiofilm activity in in vitro and ex vivo treatment models.
We observed that CDN11 is a potent antibiofilm agent in in vitro models, outperforming ciprofloxacin in both peg and catheter models, despite its lowered antibiotic activity against planktonic UTI89. We further probed CDN11’s antibiofilm activity by testing its activity against the ciprofloxacin-resistant strain EC958. Both planktonic and biofilm EC958 were resistant to high concentrations of CDN11 (>800 μM for MIC and MBEC), suggesting that CDN11’s bactericidal mode of action is the same against planktonic and biofilm bacteria. Loss of activity against ciprofloxacin resistant strains also suggests CDN11 still targets the same enzymes as ciprofloxacin. Given that nitroxides alone are not bactericidal [39], it is most likely the dual action of the nitroxide and antibiotic that is responsible for the enhanced activity seen against UTI89 biofilms, which are ciprofloxacin sensitive..
We observed less CDN11 activity in models where biofilms formed on biotic surfaces, and in particular intracellularly (human cell infection assays and infected mouse bladder ex vivo treatment). Comparing CDN11 to ciprofloxacin, CDN11 has a much higher molecular weight (952.20 g mol-1 vs. 331.34 g mol-1), suggesting that it may have reduced cell permeability of both bacterial and mammalian cells [62,63]. Other factors known to reduce cell permeability and tissue penetration include a higher polar surface area and bond flexibility [62], both of which are increased for CDN11 compared to ciprofloxacin. Ciprofloxacin’s bactericidal activity results from its binding and inhibition of DNA gyrase and topoisomerase IV [64], and so is most potent when inside bacterial cells. Despite a possible decrease in cell permeability, CDN11 was still able to reduce bacterial numbers in all models tested. Therefore, in chemically linking an antibiotic to nitroxide(s), there may be some loss in activity against planktonic bacteria but an enhancement of activity against biofilms formed on abiotic surfaces, as reported here. We also speculate that CDN11 might have reduced activity against UTI89 biofilm-like cells (IBC and filamentous) present in cell infection assays and murine bladders as a result of its reduced mammalian cell permeability.
A challenge in designing effective antimicrobials for UTI is catering to the different niches and forms of UPEC during pathogenesis. Drugs that can access intracellular UPEC would be desirable, as extracellular UPEC can be more readily accessed by antibiotics and immune cells, and also voided through urine. Additionally, drugs with antibiofilm activity would be beneficial for targeting catheter biofilms and IBCs. CDN11’s reduced activity in cellular infection models lends itself to being better purposed as a topical antibiofilm agent, as it had better activity in the catheter biofilm model. However, CDN11 could be further chemically modified to attempt to address these shortcomings, given its ability to reduce recoverable bacteria in all models. Alternatively, first-line antibiotics such as fosfomycin and nitrofurantoin [27] have yet to be hybridised with nitroxides and present an unexplored avenue for UTI antibiofilm agents. To date, several other antimicrobial agents have been hybridised to nitroxides, and have shown similar trends in antibiofilm activity against their target species [21,65,66]. Thus paired with our data, nitroxide antibiotic hybridisation is a promising strategy for eradicating biofilms, especially in infections that involve biofilms or biofilm-like structures. However in order to design and synthesise candidates with a high chance of therapeutic success, more fundamental investigations need to be conducted into the mode of action of nitroxides. Understanding how nitroxides impact biofilms will be key to designing better therapeutics that will reflect better efficacy in preclinical models, allowing for faster translation into human trials.
5. Conclusions
In conclusion, we present here a robust pipeline for preclinical testing of drugs targeting bacterial biofilms in urinary tract infections, using the nitroxide ciprofloxacin hybrid CDN11 as a candidate. New antibiotic development is arduous and requires large investments of time and money [67]. As new compounds and antibiotics become available, streamlining their testing through relevant preclinical pipelines that incorporate different aspects of the disease represents an efficient and time-saving approach to selecting lead candidates for clinical development.
Author Contributions
Conceptualisation, K.F.S. and M.T.; methodology, S.H. and J.Q.; investigation, S.H., J.Q., and S.W.; resources, S.W., K.F.S., and M.T.; writing—original draft preparation, S.H.; writing—review and editing, S.H., J.Q., and M.T.; visualization, S.H., J.Q.; supervision, J.Q. and M.T.; project administration, M.T.; funding acquisition, K.F.S. and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants by the Australian Research Council (DP210101317 to M.T. and K.F.S) and the National Health and Medical Research Council (GNT1144046 to M.T.). M.T. acknowledges support from a Georgina Sweet Award for Women in Quantitative Biomedical Science and S.H. is the recipient of an Australian Government Research Training Program (RTP) Scholarship.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Ethics Committee of the Queensland Institute for Medical Research (#A1703-600M., approved 16/06/2021).
Acknowledgments
The authors would like to thank the QIMR-B Animal Facility staff members Jonathan Mauclair and Agustin Mercau for housing and maintaining the mice for this project, and providing technical training.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tamadonfar, K.O.; Omattage, N.S.; Spaulding, C.N.; Hultgren, S.J. Reaching the End of the Line: Urinary Tract Infections. Microbiol Spectr 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.; Hreha, T.N.; Hunstad, D.A. Pathophysiology, Treatment, and Prevention of Catheter-Associated Urinary Tract Infection. Top Spinal Cord Inj Rehabil 2019, 25, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.D.; Hultgren, S.J. Urinary tract infections: microbial pathogenesis, host-pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 2020, 18, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and developmental pathways of uropathogenic Escherichia coli in urinary tract pathogenesis. Proc Natl Acad Sci USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef]
- Guiton, P.S.; Hung, C.S.; Hancock, L.E.; Caparon, M.G.; Hultgren, S.J. Enterococcal biofilm formation and virulence in an optimized murine model of foreign body-associated urinary tract infections. Infect. Immun. 2010, 78, 4166–4175. [Google Scholar] [CrossRef]
- García-Bonillo, C.; Texidó, R.; Reyes-Carmenaty, G.; Gilabert-Porres, J.; Borrós, S. Study of the Human Albumin Role in the Formation of a Bacterial Biofilm on Urinary Devices Using QCM-D. ACS Appl Bio Mater 2020, 3, 3354–3364. [Google Scholar] [CrossRef]
- Lewis, K. Multidrug tolerance of biofilms and persister cells. Curr. Top. Microbiol. Immunol. 2008, 322, 107–131. [Google Scholar] [CrossRef]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef]
- Gonzalez, M.J.; Da Cunda, P.; Notejane, M.; Zunino, P.; Scavone, P.; Robino, L. Fosfomycin tromethamine activity on biofilm and intracellular bacterial communities produced by uropathogenic Escherichia coli isolated from patients with urinary tract infection. Pathog Dis 2019, 77. [Google Scholar] [CrossRef]
- Hung, C.; Zhou, Y.; Pinkner, J.S.; Dodson, K.W.; Crowley, J.R.; Heuser, J.; Chapman, M.R.; Hadjifrangiskou, M.; Henderson, J.P.; Hultgren, S.J. Escherichia coli biofilms have an organized and complex extracellular matrix structure. mBio 2013, 4, e00645–00613. [Google Scholar] [CrossRef]
- Anderson, G.G.; Goller, C.C.; Justice, S.; Hultgren, S.J.; Seed, P.C. Polysaccharide capsule and sialic acid-mediated regulation promote biofilm-like intracellular bacterial communities during cystitis. Infect. Immun. 2010, 78, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.J.; Seed, P.C.; Hultgren, S.J. Development of intracellular bacterial communities of uropathogenic Escherichia coli depends on type 1 pili. Cell. Microbiol. 2007, 9, 2230–2241. [Google Scholar] [CrossRef] [PubMed]
- Totsika, M.; Kostakioti, M.; Hannan, T.J.; Upton, M.; Beatson, S.A.; Janetka, J.W.; Hultgren, S.J.; Schembri, M.A. A FimH inhibitor prevents acute bladder infection and treats chronic cystitis caused by multidrug-resistant uropathogenic Escherichia coli ST131. J Infect Dis 2013, 208, 921–928. [Google Scholar] [CrossRef]
- Mysorekar, I.U.; Hultgren, S.J. Mechanisms of uropathogenic Escherichia coli persistence and eradication from the urinary tract. Proc Natl Acad Sci U S A 2006, 103, 14170–14175. [Google Scholar] [CrossRef] [PubMed]
- Hannan, T.J.; Mysorekar, I.U.; Hung, C.S.; Isaacson-Schmid, M.L.; Hultgren, S.J. Early severe inflammatory responses to uropathogenic E. coli predispose to chronic and recurrent urinary tract infection. PLoS Pathog 2010, 6, e1001042. [Google Scholar] [CrossRef]
- Andersen, M.J.; Fong, C.; La Bella, A.A.; Molina, J.J.; Molesan, A.; Champion, M.M.; Howell, C.; Flores-Mireles, A.L. Inhibiting host-protein deposition on urinary catheters reduces associated urinary tract infections. Elife 2022, 11. [Google Scholar] [CrossRef]
- Barraud, N.; Kelso, M.J.; Rice, S.A.; Kjelleberg, S. Nitric oxide: a key mediator of biofilm dispersal with applications in infectious diseases. Curr. Pharm. Des. 2015, 21, 31–42. [Google Scholar] [CrossRef]
- Hawas, S.; Verderosa, A.D.; Totsika, M. Combination Therapies for Biofilm Inhibition and Eradication: A Comparative Review of Laboratory and Preclinical Studies. Frontiers in Cellular and Infection Microbiology 2022, 12. [Google Scholar] [CrossRef]
- Poh, W.H.; Rice, S.A. Recent Developments in Nitric Oxide Donors and Delivery for Antimicrobial and Anti-Biofilm Applications. In Molecules, 2022; Vol. 27. [CrossRef]
- Reffuveille, F.; Fuente-Nunez Cde, L.; Fairfull-Smith, K.E.; Hancock, R.E. Potentiation of ciprofloxacin action against Gram-negative bacterial biofilms by a nitroxide. Pathog Dis 2015, 73. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Dhouib, R.; Fairfull-Smith, K.E.; Totsika, M. Nitroxide Functionalized Antibiotics Are Promising Eradication Agents against Staphylococcus aureus Biofilms. Antimicrob. Agents Chemother. 2019, 64, e01685–01619. [Google Scholar] [CrossRef]
- Verderosa, A.D.; de la Fuente-Nunez, C.; Mansour, S.C.; Cao, J.; Lu, T.K.; Hancock, R.E.W.; Fairfull-Smith, K.E. Ciprofloxacin-nitroxide hybrids with potential for biofilm control. Eur J Med Chem 2017, 138, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Verderosa, A.D.; Mansour, S.C.; de la Fuente-Nunez, C.; Hancock, R.E.; Fairfull-Smith, K.E. Synthesis and Evaluation of Ciprofloxacin-Nitroxide Conjugates as Anti-Biofilm Agents. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Verderosa, A.D.; Harris, J.; Dhouib, R.; Totsika, M.; Fairfull-Smith, K.E. Eradicating uropathogenic Escherichia coli biofilms with a ciprofloxacin-dinitroxide conjugate. Medchemcomm 2019, 10, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Verderosa, A.D.; Dhouib, R.; Fairfull-Smith, K.E.; Totsika, M. Profluorescent Fluoroquinolone-Nitroxides for Investigating Antibiotic⁻Bacterial Interactions. Antibiotics (Basel) 2019, 8. [Google Scholar] [CrossRef]
- Fleming, D.; Rumbaugh, K. The Consequences of Biofilm Dispersal on the Host. Sci Rep 2018, 8, 10738. [Google Scholar] [CrossRef]
- Kot, B. Antibiotic Resistance Among Uropathogenic Escherichia coli. Pol J Microbiol 2019, 68, 403–415. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Totsika, M.; Moriel, D.G.; Idris, A.; Rogers, B.A.; Wurpel, D.J.; Phan, M.D.; Paterson, D.L.; Schembri, M.A. Uropathogenic Escherichia coli mediated urinary tract infection. Curr. Drug Targets 2012, 13, 1386–1399. [Google Scholar] [CrossRef]
- Hultgren, S.J.; Schwan, W.R.; Schaeffer, A.J.; Duncan, J.L. Regulation of production of type 1 pili among urinary tract isolates of Escherichia coli. Infect. Immun. 1986, 54, 613–620. [Google Scholar] [CrossRef]
- Forde, B.M.; Ben Zakour, N.L.; Stanton-Cook, M.; Phan, M.D.; Totsika, M.; Peters, K.M.; Chan, K.G.; Schembri, M.A.; Upton, M.; Beatson, S.A. The complete genome sequence of Escherichia coli EC958: a high quality reference sequence for the globally disseminated multidrug resistant E. coli O25b:H4-ST131 clone. PLoS One 2014, 9, e104400. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). In Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute, 950 West Valley Road, Suite 2500, Wayne, Pennsylvania 19087 USA, 2018.
- Berry, R.E.; Klumpp, D.J.; Schaeffer, A.J. Urothelial cultures support intracellular bacterial community formation by uropathogenic Escherichia coli. Infect. Immun. 2009, 77, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Hannan, T.J.; Hunstad, D.A. A Murine Model for Escherichia coli Urinary Tract Infection. Methods Mol Biol 2016, 1333, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.O.; Flores, C.; Williams, C.; Flusberg, D.A.; Marr, E.E.; Kwiatkowska, K.M.; Charest, J.L.; Isenberg, B.C.; Rohn, J.L. Recurrent Urinary Tract Infection: A Mystery in Search of Better Model Systems. Front Cell Infect Microbiol 2021, 11, 691210. [Google Scholar] [CrossRef] [PubMed]
- Totsika, M.; Beatson, S.A.; Sarkar, S.; Phan, M.D.; Petty, N.K.; Bachmann, N.; Szubert, M.; Sidjabat, H.E.; Paterson, D.L.; Upton, M. , et al. Insights into a multidrug resistant Escherichia coli pathogen of the globally disseminated ST131 lineage: genome analysis and virulence mechanisms. PLoS One 2011, 6, e26578. [Google Scholar] [CrossRef]
- Phan, M.D.; Peters, K.M.; Alvarez Fraga, L.; Wallis, S.C.; Hancock, S.J.; Nhu, N.T.K.; Forde, B.M.; Bauer, M.J.; Paterson, D.L.; Beatson, S.A. , et al. Plasmid-Mediated Ciprofloxacin Resistance Imparts a Selective Advantage on Escherichia coli ST131. Antimicrob. Agents Chemother. 2022, 66, e0214621. [Google Scholar] [CrossRef]
- Werneburg, G.T. Catheter-Associated Urinary Tract Infections: Current Challenges and Future Prospects. Res Rep Urol 2022, 14, 109–133. [Google Scholar] [CrossRef]
- de la Fuente-Nunez, C.; Reffuveille, F.; Fairfull-Smith, K.E.; Hancock, R.E. Effect of nitroxides on swarming motility and biofilm formation, multicellular behaviors in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 4877–4881. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: new technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef]
- Macia, M.D.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef]
- Majeed, A.; Sagar, F.; Latif, A.; Hassan, H.; Iftikhar, A.; Darouiche, R.O.; Mohajer, M.A. Does antimicrobial coating and impregnation of urinary catheters prevent catheter-associated urinary tract infection? A review of clinical and preclinical studies. Expert Rev Med Devices 2019, 16, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Talja, M.; Korpela, A.; Järvi, K. Comparison of urethral reaction to full silicone, hydrogen-coated and siliconised latex catheters. Br. J. Urol. 1990, 66, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Kerkez, I.; Tulkens, P.M.; Tenson, T.; Van Bambeke, F.; Putrinš, M. Uropathogenic Escherichia coli Shows Antibiotic Tolerance and Growth Heterogeneity in an In Vitro Model of Intracellular Infection. Antimicrob. Agents Chemother. 2021, 65, e0146821. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, A.; Ognenovska, S.; Paino, D.; Whiteley, G.; Glasbey, T.; Kriel, F.H.; Farrell, J.; Moore, K.H.; Manos, J.; Das, T. N-Acetylcysteine Protects Bladder Epithelial Cells from Bacterial Invasion and Displays Antibiofilm Activity against Urinary Tract Bacterial Pathogens. Antibiotics (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Zhou, G.; Mo, W.J.; Sebbel, P.; Min, G.; Neubert, T.A.; Glockshuber, R.; Wu, X.R.; Sun, T.T.; Kong, X.P. Uroplakin Ia is the urothelial receptor for uropathogenic Escherichia coli: evidence from in vitro FimH binding. J. Cell Sci. 2001, 114, 4095–4103. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.N.; Miao, Y. The nature of immune responses to urinary tract infections. Nat. Rev. Immunol. 2015, 15, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Ligon, M.M.; Mysorekar, I.U. Trans-mission control in the urinary tract: Local cytokine regulation of monocyte proliferation to combat infection. J Leukoc Biol 2018, 103, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Billips, B.K.; Forrestal, S.G.; Rycyk, M.T.; Johnson, J.R.; Klumpp, D.J.; Schaeffer, A.J. Modulation of host innate immune response in the bladder by uropathogenic Escherichia coli. Infect. Immun. 2007, 75, 5353–5360. [Google Scholar] [CrossRef]
- Schilling, J.D.; Mulvey, M.A.; Vincent, C.D.; Lorenz, R.G.; Hultgren, S.J. Bacterial invasion augments epithelial cytokine responses to Escherichia coli through a lipopolysaccharide-dependent mechanism. J. Immunol. 2001, 166, 1148–1155. [Google Scholar] [CrossRef]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef]
- Rosen, D.A.; Hooton, T.M.; Stamm, W.E.; Humphrey, P.A.; Hultgren, S.J. Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med 2007, 4, e329. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Dhar, N.; Thacker, V.V.; Simonet, T.M.; Signorino-Gelo, F.; Knott, G.W.; McKinney, J.D. Dynamic persistence of UPEC intracellular bacterial communities in a human bladder-chip model of urinary tract infection. Elife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Haghighitalab, A.; Matin, M.M.; Bahrami, A.R.; Iranshahi, M.; Saeinasab, M.; Haghighi, F. In vitro investigation of anticancer, cell-cycle-inhibitory, and apoptosis-inducing effects of diversin, a natural prenylated coumarin, on bladder carcinoma cells. Z Naturforsch C J Biosci 2014, 69, 99–109. [Google Scholar] [CrossRef]
- Lee, H.; Park, J.B. Evaluation of the effects of dimethylsulphoxide on morphology, cellular viability, mRNA, and protein expression of stem cells culture in growth media. Biomed Rep 2017, 7, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Miao, Y.; Abraham, S.N. The multiple antibacterial activities of the bladder epithelium. Ann Transl Med 2017, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, G.S.; Loudon, K.W.; Suchanek, O.; Clatworthy, M.R. Tissue Immunity in the Bladder. Annu. Rev. Immunol. 2022, 40, 499–523. [Google Scholar] [CrossRef] [PubMed]
- Karachalios, G.N.; Georgiopoulos, A.N.; Nasopoulou-Papadimitriou, D.D.; Adracta, D.J. Value of single-dose ciprofloxacin in the treatment of acute uncomplicated urinary tract infection in women. Drugs Exp. Clin. Res. 1991, 17, 521–524. [Google Scholar]
- Jakobsen, L.; Lundberg, C.V.; Frimodt-Møller, N. Ciprofloxacin Pharmacokinetics/Pharmacodynamics against Susceptible and Low-Level Resistant Escherichia coli Isolates in an Experimental Ascending Urinary Tract Infection Model in Mice. Antimicrob. Agents Chemother. 2020, 65. [Google Scholar] [CrossRef]
- Justice, S.S.; Hunstad, D.A.; Seed, P.C.; Hultgren, S.J. Filamentation by Escherichia coli subverts innate defenses during urinary tract infection. Proc Natl Acad Sci U S A 2006, 103, 19884–19889. [Google Scholar] [CrossRef]
- Engel, D.; Dobrindt, U.; Tittel, A.; Peters, P.; Maurer, J.; Gutgemann, I.; Kaissling, B.; Kuziel, W.; Jung, S.; Kurts, C. Tumor necrosis factor alpha- and inducible nitric oxide synthase-producing dendritic cells are rapidly recruited to the bladder in urinary tract infection but are dispensable for bacterial clearance. Infect. Immun. 2006, 74, 6100–6107. [Google Scholar] [CrossRef]
- Ross, A.G.; Benton, B.M.; Chin, D.; De Pascale, G.; Fuller, J.; Leeds, J.A.; Reck, F.; Richie, D.L.; Vo, J.; LaMarche, M.J. Synthesis of ciprofloxacin dimers for evaluation of bacterial permeability in atypical chemical space. Bioorganic & Medicinal Chemistry Letters 2015, 25, 3468–3475. [Google Scholar] [CrossRef]
- Zhang, G.F.; Liu, X.; Zhang, S.; Pan, B.; Liu, M.L. Ciprofloxacin derivatives and their antibacterial activities. Eur J Med Chem 2018, 146, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.T. The mode of action of 4-quinolones and possible mechanisms of resistance. J. Antimicrob. Chemother. 1986, 18 Suppl D, 21–29. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Hawas, S.; Harris, J.; Totsika, M.; Fairfull-Smith, K.E. Isothiazolone-Nitroxide Hybrids with Activity against Antibiotic-Resistant Staphylococcus aureus Biofilms. ACS Omega 2022, 7, 5300–5310. [Google Scholar] [CrossRef] [PubMed]
- Ndukwe, A.R.N.; Hawas, S.; Qin, J.; Wiedbrauk, S.; Totsika, M.; Boase, N.R.B.; Fairfull-Smith, K.E. Simple and Efficient Synthesis of 3-Aryl-2-oxazolidinone Scaffolds Enabling Increased Potency toward Biofilms. Mol. Pharm. 2023, 20, 3484–3493. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
Figure 1.
CDN11 has potent antibiofilm activity against UPEC biofilms formed inside urinary catheters in vitro. A) Schematic of the in vitro catheter biofilm model. ~3 mm sections of UV-sterilised (lightning symbol) polyethylene catheter were incubated with 4 x 106 CFU of UTI89 in LB media for 24 h followed by 24 h of treatment with 400 μM CDN11, 400 μM ciprofloxacin, 3.5% DMSO (vehicle), or media only (MH, Untreated), before disruption with mild sonication for CFU enumeration. B) Biofilm recovered bacteria from each treatment group were plotted as log10(CFU/mL) from 8 biological replicates, with group means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
Figure 1.
CDN11 has potent antibiofilm activity against UPEC biofilms formed inside urinary catheters in vitro. A) Schematic of the in vitro catheter biofilm model. ~3 mm sections of UV-sterilised (lightning symbol) polyethylene catheter were incubated with 4 x 106 CFU of UTI89 in LB media for 24 h followed by 24 h of treatment with 400 μM CDN11, 400 μM ciprofloxacin, 3.5% DMSO (vehicle), or media only (MH, Untreated), before disruption with mild sonication for CFU enumeration. B) Biofilm recovered bacteria from each treatment group were plotted as log10(CFU/mL) from 8 biological replicates, with group means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
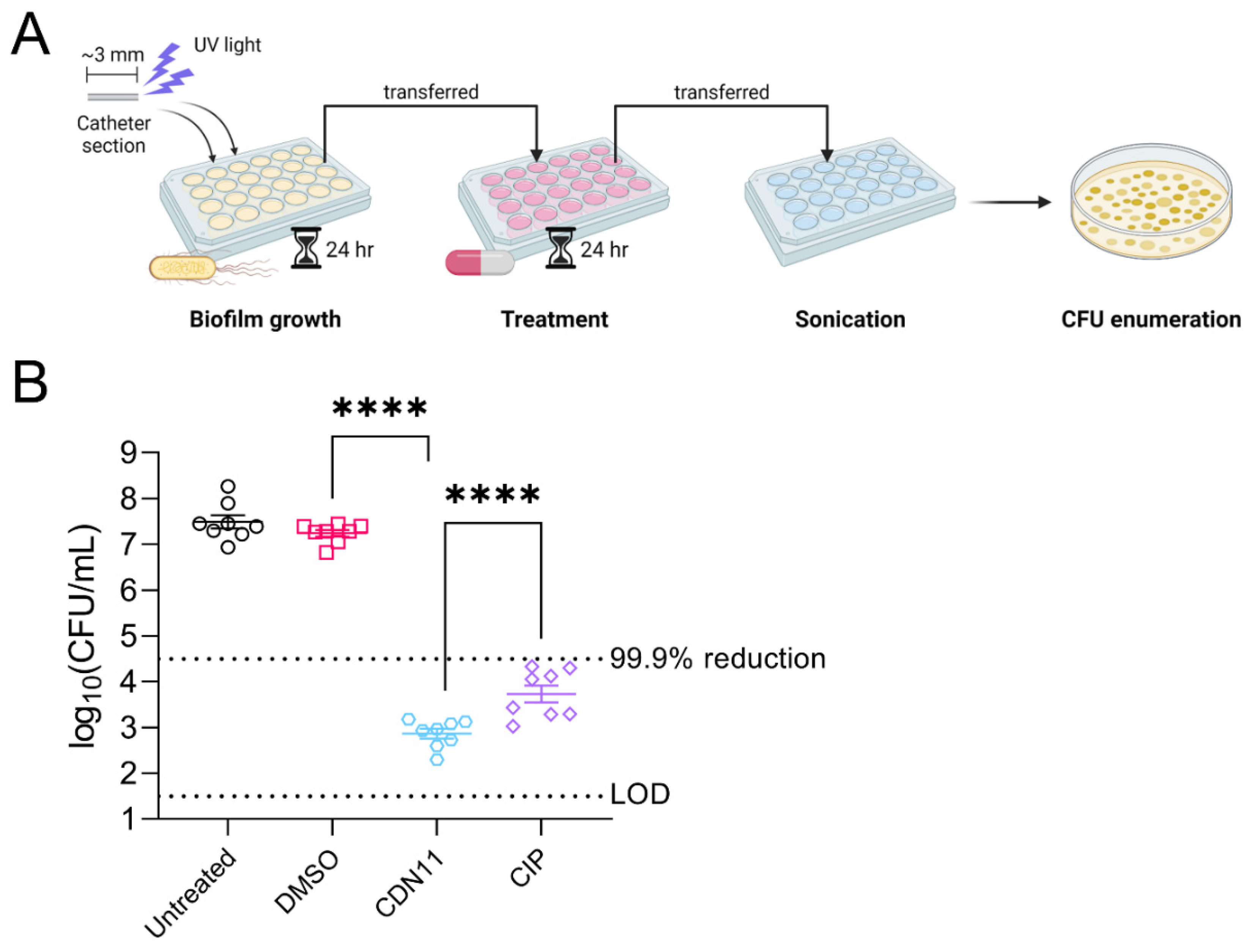
Figure 2.
CDN11 controls extracellular and intracellular UPEC during in vitro human bladder cell infection. Confluent monolayers of T24 bladder cells were incubated with UPEC strain UTI89 at an MOI of 10 for 1 h followed by A) 2 h of treatment with either 400 μM CDN11, 400 μM ciprofloxacin, an equivalent percentage of DMSO instead of CDN11 (vehicle), or media only (RPMI + 5% FCS, Untreated) or B) 1h of treatment with 200 μg/mL gentamicin to first eliminate extracellular bacteria, followed by 2 h of treatment with either 400 μM CDN11, 400 μM ciprofloxacin, an equivalent percentage of DMSO instead of CDN11 (vehicle), or media only (RPMI + 5% FCS). Bacteria recovered from infected monolayers were plotted per treatment group as A) log10(CFU/mL) or B) % intracellular bacteria compared to the media only control group. Data shown are from 5-6 biological repeats, with means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as ** p ≤ 0.01, *** p ≤ 0.001, and **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
Figure 2.
CDN11 controls extracellular and intracellular UPEC during in vitro human bladder cell infection. Confluent monolayers of T24 bladder cells were incubated with UPEC strain UTI89 at an MOI of 10 for 1 h followed by A) 2 h of treatment with either 400 μM CDN11, 400 μM ciprofloxacin, an equivalent percentage of DMSO instead of CDN11 (vehicle), or media only (RPMI + 5% FCS, Untreated) or B) 1h of treatment with 200 μg/mL gentamicin to first eliminate extracellular bacteria, followed by 2 h of treatment with either 400 μM CDN11, 400 μM ciprofloxacin, an equivalent percentage of DMSO instead of CDN11 (vehicle), or media only (RPMI + 5% FCS). Bacteria recovered from infected monolayers were plotted per treatment group as A) log10(CFU/mL) or B) % intracellular bacteria compared to the media only control group. Data shown are from 5-6 biological repeats, with means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as ** p ≤ 0.01, *** p ≤ 0.001, and **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
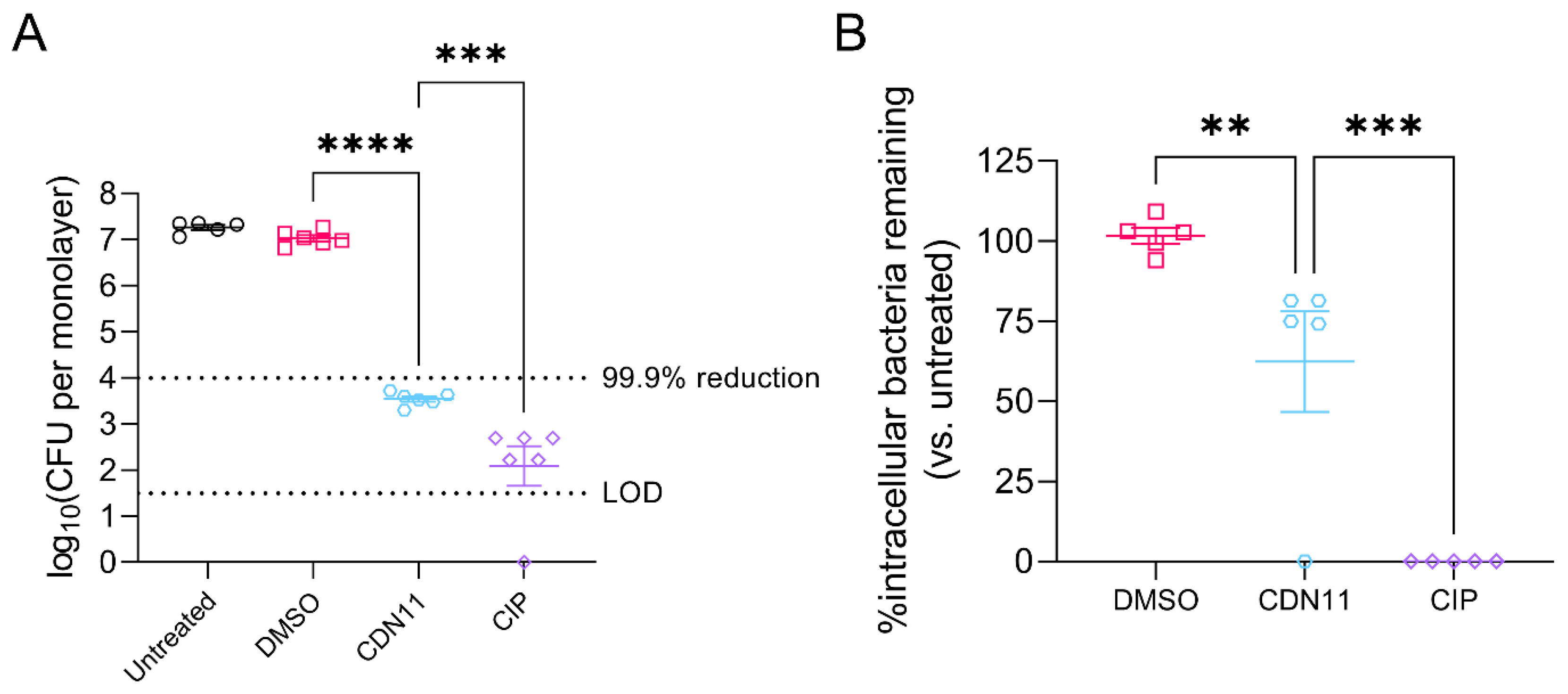
Figure 3.
CDN11 reduces intracellular UPEC reservoirs in infected mouse bladders after 6 hours of treatment ex vivo. A) Schematic overview of acute UTI mouse model ex vivo bladder treatment timeline. 6-week-old female C3H/HeJ mice were inoculated with 1 x 108 CFU of UPEC strain UTI89 and allowed to establish acute UTI for 16 h. Mice were then culled, bladders extracted and bisected, followed by 1 h of treatment with gentamicin, and then 6 h of treatment with either 400 μM CDN11, ciprofloxacin, or an equivalent percentage of DMSO instead of CDN11 (vehicle), or PBS only. Bladder halves were homogenised post treatment, serially diluted, and CFU enumerated. B) Bacteria recovered from mouse bladders were plotted per treatment group as log10(CFU/0.1g tissue). Data shown are from 8 mice per group with means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as ns not significant, ** p ≤ 0.01, **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
Figure 3.
CDN11 reduces intracellular UPEC reservoirs in infected mouse bladders after 6 hours of treatment ex vivo. A) Schematic overview of acute UTI mouse model ex vivo bladder treatment timeline. 6-week-old female C3H/HeJ mice were inoculated with 1 x 108 CFU of UPEC strain UTI89 and allowed to establish acute UTI for 16 h. Mice were then culled, bladders extracted and bisected, followed by 1 h of treatment with gentamicin, and then 6 h of treatment with either 400 μM CDN11, ciprofloxacin, or an equivalent percentage of DMSO instead of CDN11 (vehicle), or PBS only. Bladder halves were homogenised post treatment, serially diluted, and CFU enumerated. B) Bacteria recovered from mouse bladders were plotted per treatment group as log10(CFU/0.1g tissue). Data shown are from 8 mice per group with means and standard deviation shown by horizontal lines and error bars, respectively. Differences between group means were calculated using ordinary one-way ANOVA with Dunn’s correction, and significance was shown as ns not significant, ** p ≤ 0.01, **** p ≤ 0.0001. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
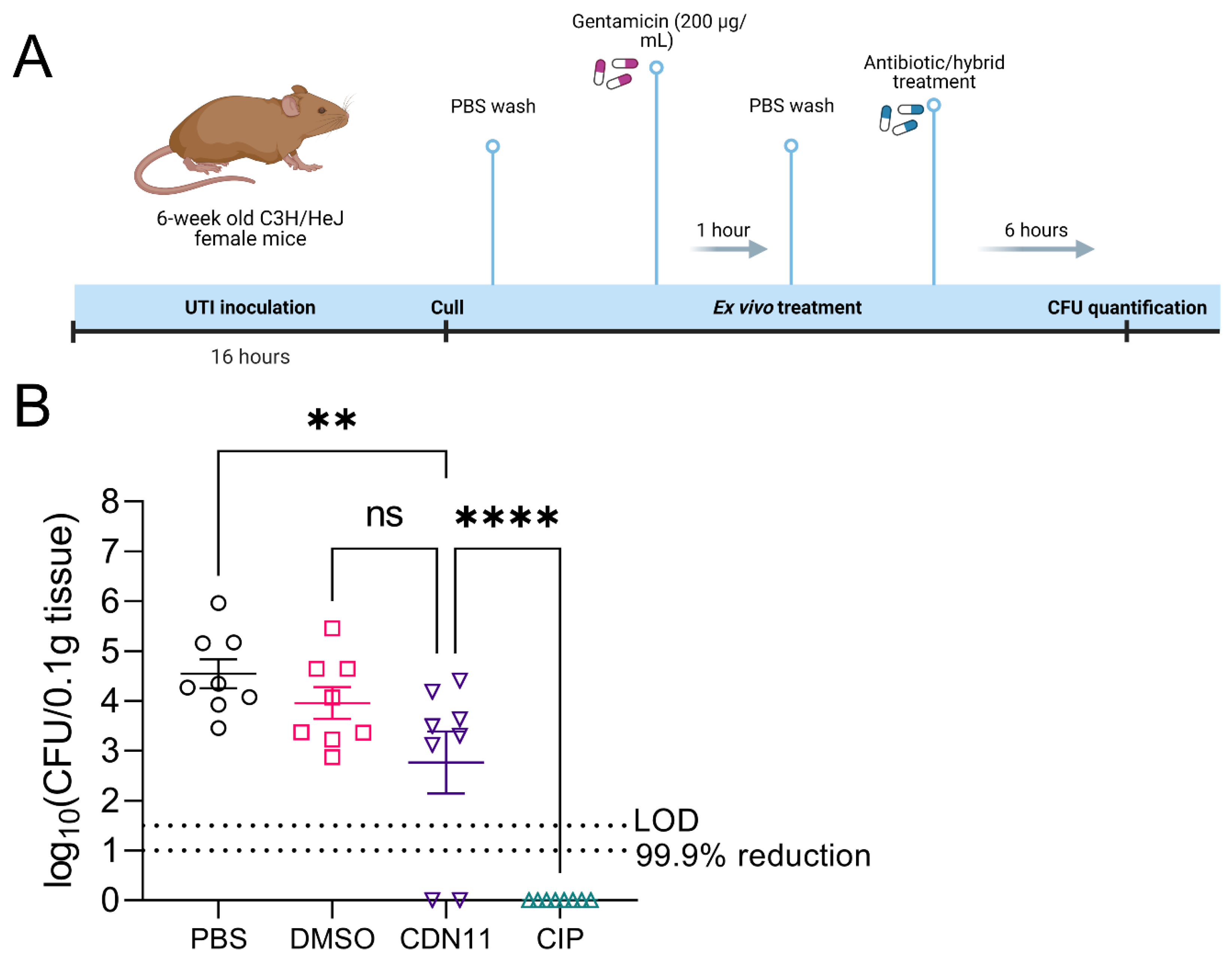
Figure 4.
EC958 planktonic and biofilm cells are resistant to CDN11. Minimum biofilm eradication concentration (MBEC) values are reported as complete biofilm eradication (concentration at which there is no recoverable viable biofilm CFU). UTI89 and EC958 biofilms were grown for 24 h using the Calgary Biofilm Device at 37°C with shaking, followed by treatment for 24 h in CDN11 at 37°C with shaking. Biofilms were disrupted with mild sonication for CFU enumeration. Enumerated bacterial CFU was plotted as log10(CFU/mL). Data represents n = 16 (UTI89) and n = 4 (EC958) biological repeats with means and standard deviation shown. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
Figure 4.
EC958 planktonic and biofilm cells are resistant to CDN11. Minimum biofilm eradication concentration (MBEC) values are reported as complete biofilm eradication (concentration at which there is no recoverable viable biofilm CFU). UTI89 and EC958 biofilms were grown for 24 h using the Calgary Biofilm Device at 37°C with shaking, followed by treatment for 24 h in CDN11 at 37°C with shaking. Biofilms were disrupted with mild sonication for CFU enumeration. Enumerated bacterial CFU was plotted as log10(CFU/mL). Data represents n = 16 (UTI89) and n = 4 (EC958) biological repeats with means and standard deviation shown. The technical limit of CFU detection (LOD) and the 99.9% CFU reduction cut-off (compared to vehicle control) are marked by dotted lines.
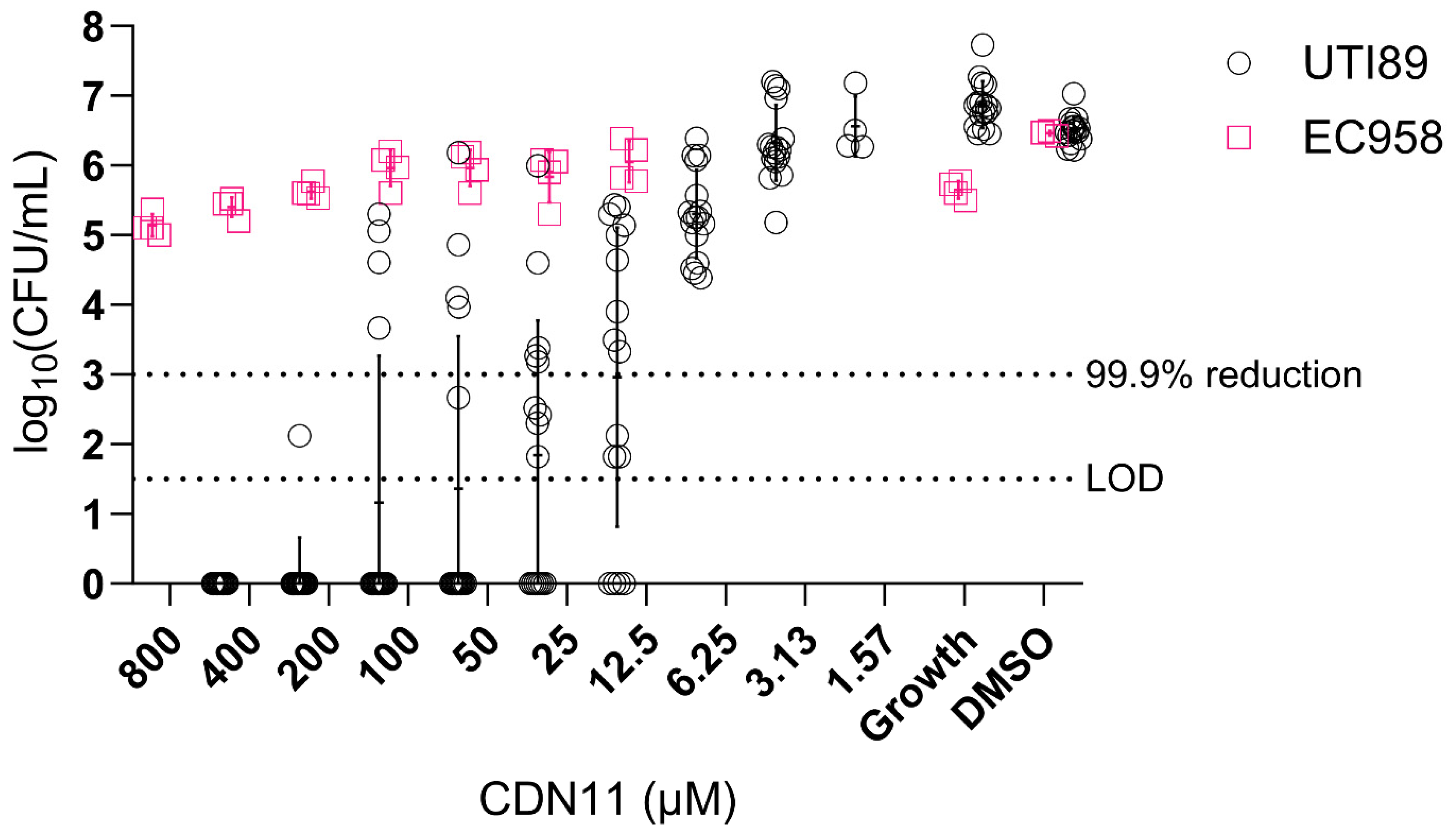
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



