
Submitted:
01 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
Skin cancer is a condition characterized by the abnormal growth of skin cells, primarily caused by exposure to ultraviolet (UV) radiation from the sun or artificial sources like tanning beds. Different types of skin cancer include melanoma, basal cell carcinoma, and squamous cell carcinoma. Despite the advancements in targeted therapies, there is still a need for a safer, highly efficient approach to preventing and treating cutaneous malignancies. Spices have a rich history dating back thousands of years and are renowned for their ability to enhance the flavor, taste, and color of food. Derived from various plant parts like seeds, fruits, bark, roots, or flowers, spices are important culinary ingredients. However, their value extends beyond the culinary realm. Some spices contain bioactive compounds, including phenolic compounds, which are known for their significant biological effects. These compounds have attracted attention in scientific research due to their potential health benefits, including their possible role in disease prevention and treatment, such as cancer. This review focuses on examining the potential of spice-derived phenolic compounds as preventive or therapeutic agents for managing skin cancers. By compiling and analyzing the available knowledge, this review aims to provide insights that can guide future research in identifying new anticancer phytochemicals and uncovering additional mechanisms for combating skin cancer.
Keywords:
melanoma
; non-melanoma skin cancer
; spice
; phenolic compounds
; anticancer effect
1. Introduction
Spices are defined as aromatic vegetable substances derived from various parts of plants, including seeds, bark, roots, fruits, and leaves. They have been used globally for centuries as a food ingredient to enhance flavor and extend the shelf life of food [1]. Besides their culinary uses, spices are also recognized for their medicinal properties and have been traditionally employed for treating various diseases and ailments. In recent years, there has been a notable increase in research exploring the potential health benefits of spices. Studies suggest that spices may provide protection against conditions such as cardiovascular disease, neurodegenerative disorders, chronic inflammation, obesity, or type 2 diabetes [2,3,4]. The wide range of health benefits associated with spices can be credited to their various active components, with phenolic compounds being the predominant phytochemicals responsible for these effects [5].
In the last decades, phenolic compounds have been intensively studied as potential anticancer agents against various types of cancers including breast [6,7,8,9,10,11]. Moreover, based on scientific literature, there is strong evidence supporting the potential of phenolic compounds as promising agents in the battle against both melanoma and non-melanoma skin cancers.
This review article provides an overview of the research on spice-derived phenolic compounds and their effects on melanoma and non-melanoma skin cancers, encompassing in vitro and in vivo studies.
2. Skin cancers
Skin cancers encompass a range of malignancies that can affect individuals worldwide. Skin cancer incidence and mortality rates vary depending on several factors, including geographical location, sun exposure, skin type, and preventive measures. Skin cancers are among the most commonly diagnosed cancers globally. Non-melanoma skin cancers (NMSC), including basal cell carcinoma (BCC) and squamous cell carcinoma (SCC), are more prevalent than melanoma [12]. On the other hand, melanoma, although less common than NMSC, is more aggressive and has a higher potential for metastasis. Over the past few decades, there has been a notable rise in the incidence of melanoma, particularly among fair-skinned populations with significant sun exposure [13]. However, studies indicate that there is a projected decline in melanoma rates in the future [14].
Basal cell carcinoma (BCC) is derived from immature pluripotent cells of the lower layers of the epidermis. The etiology is multifactorial. Exposure to UV radiation is the most important cause, as BCC occurs predominantly in places exposed to the sun. Previous exposure to chemical carcinogens, ionizing radiation or phototherapy for skin diseases, chronic inflammatory changes, or trauma can also contribute to BCC. A higher incidence of BCC is observed in immunocompromised patients. Multipotent differentiation potential is reflected in great clinical diversity and morphology. Nodular – solid and cystic BCC, ulcerative, superficial, morphea-like sclerosing, keratotic and pigment variant. The ulcerative BCC can lead to extremely extensive lesions. Metastases are extremely rare, and the morbidity associated with BCC is related to local tissue invasion and destruction [15,16,17].
Actinic keratosis (AK) is the proliferation of cytologically atypical keratinocytes. Nowadays AK is considered an early in situ squamous cell carcinoma. Age, the cumulative dose of UV radiation, outdoor activities, male gender, solar lentigines, and patient´s immunological condition are considered risk factors. Scaly or hyperkeratotic brown-red, brown-yellow macules or papules with an erythematous base, usually less than 1 cm in diameter, in sun-exposed parts of the body. AKs can either rare spontaneously regress, they can remain stable without significant change, or progress to invasive squamous cell carcinoma [18,19,20].
Squamous cell carcinoma (SCC) represents the most aggressive type of non-melanoma skin cancer. Up to 60% of invasive SCC arise from previous AK. Actinic keratosis, radiation (arsenic) keratosis, Bowen disease, Bowenoid papulosis, actinic cheilitis, and Queyrat erythroplasia are considered to be its in situ variants. The etiology is multifactorial and includes mainly chronic exposure to UV radiation, light phototype, long-term previous phototherapy for other dermatoses, professional exposure to X-ray radiation or radiation for internal malignancies, chemical carcinogens, chronic inflammatory skin changes, immunosuppression, infection with HPV viruses. It grows de novo or from a previous precancer. Initially, indurated, painless papules or macules of grayish, brownish-yellow, and reddish skin color may progress to nodular lesions with crusts and ulceration on the surface. The tumor grows quickly, disintegrates, and can outgrow soft tissues, cartilage, and bone. It can metastasize to regional lymph nodes, and later also to other organs. The prognosis depends on the location, size, and degree of tumor differentiation [20,21,22].
Early and correct diagnosis of NMSCs is a basic prerequisite for their successful treatment. The gold standard for the diagnosis of BCC, SCC, and AK is histopathological examination. The choice of individual therapeutic strategies depends on the anatomical location, the thickness of the tumor, the affected area of the skin surface, the histological type of the tumor, patient comorbidity, and the availability of the method. Surgical excision is still the predominant therapeutic strategy. Other treatment possibilities include curettage, electrodesiccation, cryotherapy, and radiotherapy. Only a few drugs meet the criteria for field treatment: 5% fluorouracil, 3% diclofenac in a gel with hyaluronic acid, photodynamic therapy, chemical peeling (destruction is achieved with 35% trichloroacetic acid, alpha-hydroxy acids, zinc chloride, and phenolic acid), retinoids (topical or systemic) and 5% imiquimod in cream [20,21,22,23,24,25].
Melanoma is a malignant skin tumor that arises from the malignant transformation of melanocytes anywhere in the human body. It is mainly localized in the skin, but it also often appears in the eye or mucous membranes. It belongs to the most insidious tumors due to its ability to quickly form metastases. One-third of melanomas arise from various pigment nevi, and two-thirds of melanomas arise from so-called "de novo" - without a pre-existing pigmented lesion on clinically normal skin. Melanoma is considered to be the human tumor with the greatest immunogenic response. The incidence of melanoma is increasing more than the incidence of any other malignancy [26,27].
The causes of the malignant transformation of a melanocyte into a melanoma cell are still unclear. The genetic predisposition of an individual with a positive family history of melanoma in combination with the influences of the external environment plays an important role in the development of melanoma. UV radiation is one of the best-documented risk factors for the development of melanomas - whether from existing dysplastic nevi or arising de novo. Other risk factors include burning the skin at a young age, phototherapy, repeated use of tanning beds, immunosuppressive therapy, acquired and congenital disorders of the immune system, and oncological diseases treated with aggressive chemotherapy or immunotherapy [28,29].
Clinical diagnosis is also based on the ABCDEF criteria, which in many cases will help to diagnose developing melanoma. These criteria include asymmetry of the lesion, borders irregularity, color variety, diameter larger than 5mm, and firm (nodular) or funny-looking lesion or ugly duckling sign (different from the others) [26]. Cutaneous melanoma occurs as an irregular oval or polycyclic lesion with a typical variety of colors (brown, gray, blue-black, whitish, pink). In the beginning, horizontal growth of the tumor prevails, the progression will show vertical growth and the formation of the nodular lesion, with a tendency to ulcerate and bleed after minimal trauma which signals the growth of the tumor in depth and thus a worse prognosis of the disease. A rare amelanotic or a hypomelanotic subtype of melanoma due to lack of pigmentation can mimic other benign and malignant conditions [30,31]. Melanoma can occur anywhere on the skin, on the mucous membranes, in the iris of the eye, in the soft coverings of the brain, in the heart, in the urogenital tract, and in the lymph node [28].
Early surgical extirpation of the primary tumor with a sufficient safety margin with the extirpation of the adjacent subcutaneous tissue is currently the only curative treatment for this cancer. After a precise histological examination, the TNM staging, and the further course of treatment are determined. If a complete examination (using sonography, CT, or PET/CT) does not detect metastatic spread of the disease, patients remain in follow-up with their oncologist/oncodermatologist. In case of evidence of metastases, adequate surgical treatment, radiotherapy, chemotherapy, or biological treatment is chosen. For unresectable or distant metastatic disease, a combination therapy with nivolumab/ipilimumab has been a recommended option for first-line or second- or subsequent-line systemic therapy. Patients diagnosed with melanoma should be regularly examined [32,33,34].
3. Phenolic compounds
Phenolic compounds are widely distributed throughout the plant kingdom and represent a significant class of secondary metabolites in plants. They can be found in various plant tissues, including fruits, nuts, grains, legumes, and spices. These compounds play essential roles in numerous physiological processes, such as improving plant quality, adding to the color and flavor profiles, and enhancing stress resistance. Phenolic compounds share a common chemical structure consisting of an aromatic ring accompanied by one or more hydroxyl substituents. These compounds can be further categorized into several classes, with the primary groups of phenolic compounds encompassing phenolic acids, flavonoids, tannins, stilbenes, coumarins, and lignans (Figure 1) [35].
Phenolic acids, which belong to the group of non-flavonoid phenolic compounds, are extensively found in plants in various forms, including free, conjugated-soluble, and insoluble-bound forms. They are commonly found in fruits, vegetables, whole grains, coffee, and spices. Phenolic acids in plants are associated with a wide range of functions, although much remains to be understood about their specific roles. These functions include facilitating nutrient uptake, promoting protein synthesis, enhancing enzyme activity, supporting photosynthesis, and contributing to structural components. Phenolic acids are derivatives of benzoic (e.g. 4-hydroxybenzoic acid, protocatechuic acid, vanillic acid, gallic acid, and syringic acid) and cinnamic (e.g. p-coumaric acid, caffeic acid, chlorogenic acid, ferulic acid, and sinapic acid) acids [36,37].
Flavonoids are a large class of phenolic compounds that include subclasses such as flavones, flavonols, flavanones, flavanols (catechins), anthocyanins, and isoflavones. They are widely distributed in fruits, vegetables, grains, and beverages like tea and wine. Flavonoids in plants serve multiple roles, including the regulation of cell growth, the attraction of pollinating insects, the protection of enzymes and vitamins, and providing protection against both biotic and abiotic stresses [38,39].
Tannins are a group of phenolic compounds that are commonly found in plant-based foods such as tea, coffee, cocoa, and fruits like grapes and pomegranates. They contribute to the astringent taste in these foods [40].
Stilbenes are a class of phenolic compounds that are characterized by the presence of a stilbene backbone. Several plants produce natural stilbenes as a defense mechanism against various pathological conditions, including excessive ultraviolet (UV) radiation, exposure to high temperatures, attacks by insects, as well as fungal or bacterial infections. Resveratrol is the most well-known stilbene and is found in grapes, berries, and red wine [41].
Coumarins are phenolic compounds with a benzopyrone structure. They can be found in various plants such as tonka beans, cinnamon, and sweet clover. They participate in the antimicrobial plant’s chemical defense strategy, in adaptive plant responses to iron deficiency, or in the interaction between plant roots and beneficial microbes [42].
Lignans are phenolic compounds found in plant sources such as flaxseeds, sesame seeds, whole grains, and berries. They are known for their potential health benefits, including antioxidant and hormone-balancing effects. Lignans in plants serve as defensive compounds, safeguarding them against potential threats from insects, microorganisms, and even neighboring plants [43].
Apart from their natural role, phenolic compounds exhibit notable pharmacological properties that encompass anti-inflammatory [44], immunosuppressive [45], cardioprotective [46], antioxidant [47], neuroprotective [48] and antimicrobial activities [49]. Moreover, several phenolic compounds have shown anti-cancer activity by inhibiting the growth of cancer cells [50], promoting cell death such as apoptosis [51], autophagy [52], necroptosis [53] or targeting numerous signaling pathways [54,55,56]. In addition, the formation of new blood vessels that support tumor growth has also been reported as a possible target of phenolic compounds [57,58,59].
4. Overview of selected spices
Allspice
Allspice (Pimenta dioica), is an evergreen tree belonging to the myrtle family (Myrtaceae). It is commonly referred to as Jamaican pepper or pimento. Originating from the West Indies and Central America, the allspice plant derives its name from the distinctive taste of its dried berries, which is reminiscent of a blend of cloves, cinnamon, and nutmeg In folk medicine, Allspice berries have been used for the treatment of colds, dysmenorrhea, dyspepsia, diabetes, myalgia and sore joints [60]. Furthermore, Allspice has been found to have different pharmacological activities such as antimicrobial, antioxidant anti-inflammatory, or anticancer effects [61,62,63,64] due to the presence of numerous phytochemicals among them eugenol is a main bioactive constituent [65]. Other phenolic compounds such as methyl eugenol, isoeugenol, chavicol, flavonoids (e.g. quercetin, quercitrin, kaempferol, catechin, naringenin), tannins (e.g. vascalaginone, grandininol) or phenolic acids (e.g. syringic acid, caffeic acid, coumaric acid, cinnamic acid) have also been reported as active constituents of Allspice [66,67,68].
Alpinia galanga
Alpinia galanga (L.) Willd, also known as galangal, is a plant species in the ginger family (Zingiberaceae). It is native to Southeast Asia and is widely cultivated in Indonesia, Malaysia, Thailand, and other tropical countries [69]. Galangal rhizome is a widely utilized spice available in fresh, frozen, dried, or powdered forms [70]. In traditional medicine, it is used to treat a variety of ailments such as digestive problems, respiratory infections, rheumatic pain, fever, and skin diseases. It is also believed to have anti-inflammatory, antioxidant, and anticancer properties [71]. The analysis of the phytochemical constituents of A. galanga indicated the presence of numerous bioactive compounds such as alkaloids, tannins, terpenoids, saponins, and phenolics [72]. Regarding phenolic compounds, several flavonoids and non-flavonoid phenolic compounds are present in A. galanga. Flavonoids, such as galangin, galangin-3-methylether, kaempferol, quercetin, isorhamnetin, apigenin, kumatakenin, pinocembrin have been isolated from the rhizomes and seeds of A. galanga [73]. In addition, numerous other, non-flavonoid phenolic compounds such as phenolic acids (e.g. ferulic acid, gallic acid), lignans (galanganal, galanganol A-C), chalcones (galanganones A-C), and other phenolic compounds such as ellagic acid, 1′-acetoxychavicol acetate, methyleugenol and many other have also been identified in A. galanga [74,75,76].
Black cumin
Nigella sativa (L.), commonly known as black cumin or black seed, is an annual plant belonging to the Ranunculaceae family. It is cultivated for its highly aromatic seeds that are utilized as a spice and in traditional medicine. This plant, also referred to as black caraway, and is is indigenous to South and Southwest Asia. Furthermore, it is prevalent in regions such as Northern Africa, the Middle East, and Southern Europe [80]. The seeds and oil of this plant are widely used in traditional medicine for managing various ailments such as rheumatoid arthritis, asthma, inflammatory conditions, diabetes, and digestive disorders [81]. Nigella sativa is known to contain a range of active compounds, with thymoquinone being the most significant. Among others, several phenolic compounds have been isolated from black cumin seeds including phenolic acids (caftaric acid, p-hydroxybenzoic acid, syringic acid, protocatechuic acid, and chlorogenic acid) flavonoids (kaempferol, kaempferol-3-glucoside, quercetin, quercitrin, and diosmin) or other phenolic compounds (p-cymene, thymohydroquinone, thymol, carvacrol, t-anethole) [81,82,83].
Black pepper
Black pepper (Piper nigrum, L.) is a plant belonging to the Piperaceae family. The dried fruits of this plant are utilized as a spice across the globe. It originates from the Malabar Coast of India and it is one of the earliest known spices. It has a pungent, spicy taste and a distinct aroma, making it a popular seasoning and spice in cuisines all over the world. Black pepper is rich in bioactive compounds including alkaloids (piperine), sesquiterpenes (e.g. β-caryophyllene), monoterpenes (e.g. limonene, β-pinene, α-pinene, sabinene, camphene), oxygenated terpenes (e.g. linalool, terpinene-4-ol, eugenol). Moreover, several phenolic acids (e.g. hydroxybenzoic acid, gallic acid, caffeic acid, and hydroxycinnamic acids) and flavonoids (e.g. quercetin, catechin, epicatechin, myricetin kaempferol, isoquercetin, isorhamnetin) have been extracted from black pepper [84,85,86]. Black pepper is also used for medicinal purposes and has various health benefits including antioxidant, antimicrobial, anti-inflammatory, or gastroprotective [87]. Moreover, some bioactive compounds isolated from black pepper have also anticancer effects [88,89,90].
Cinnamon
Cinnamon, scientifically known as Cinnamomum verum, and commonly referred to as Ceylon cinnamon or true cinnamon, belongs to the Lauraceae family. This tree is native to Sri Lanka (formerly Ceylon), the neighboring Malabar Coast of India, and Myanmar. The spice is derived from the inner bark of the cinnamon tree. Other notable species within the same family include C. cassia, C. burmannii, C. loureiroi, and C. citriodorum. It is widely recognized as one of the most essential spices and is embraced and utilized by people across the globe in their daily lives [91]. Although cinnamon is preferably used as a spice, it also possesses several biological effects such as anti-tumor [92], anti-diabetic [93], anti-inflammatory [94], anti-microbial [95] and anti-oxidant activities [96]. The phytochemical content of cinnamon has shown the presence of numerous active constituents extracted from the leaf and bark oils with eugenol and cinnamaldehyde as the major constituents [97]. Among bioactive constituents, several phenolic compounds have been detected including above mentioned eugenol and cinnamaldehyde, phenolic acid (e.g. caffeic acid, chlorogenic acid, gallic, rosmarinic acid, p-coumaric acid, protocatechuic acid, p-hydroxybenzoic acid, and trans-vanillic acid), flavonoids (e.g. rutin, apigenin, catechin, and epicatechin) [98,99]. Other phenolic compounds such as cinnamyl alcohol, α-terpineol, cinnamyl acetate, p-cymene, coumarin, and pyrogallol have also been found in cinnamon extracts [100].
The phenol commonly investigated in relation to skin cancers is eugenol, which is found abundantly in this spice.
Coriander
Coriander (Coriandrum sativum L), a plant from Apiaceae family, is one of the earliest spices used by humans. It grows wild over a wide area of Western Asia and Southern Europe but today is cultivated in The Netherlands, Central and Eastern Europe, China, India, Bangladesh, and North Africa [101]. Traditional medicines, it has been used for digestive and gastric troubles, fever, cough, dysentery, and many other complaints [102]. Analyzes of the bioactive constituents of coriander showed numerous phytochemicals responsible for the therapeutic and nutritional effects of this plant including fatty acids, sterols, tocols, essential oils, and water-soluble compounds [103]. Moreover, many phenolic compounds have been isolated either from coriander seeds including phenolic acids (caffeic acid, chlorogenic acid, ferulic acid, gallic acid, o-coumaric acid, trans-hydroxycinnamic acid, p-coumaric acid, rosmarinic acid, salicylic acid, trans-cinnamic acid, vanillic acid), flavonoids (e.g. rutin, luteolin quercetin, kaempferol naringin, apigenin) [104]. The primary active constituents discovered in the leaves of coriander were quercetin derivatives such as quercetin-3-O-rutinoside quercetin 3-O-glucuronide, quercetin-3-O-glucoside and kaempferol-3-O-rutinoside [105]. In addition, numerous other phenolic compounds have been detected in the aerial parts of the coriander plant including esculin, esculetin, apigenin, luteolin, diosmin, catechin, orientin, vanillic acid, p-coumaric acid, cis- and trans-ferulic acid, gallic acid, maleic acid and others [106,107]. However, it is important to mention that the constituents of coriander can differ depending on factors such as the variety, growth stage, planting location, and season of cultivation.
Several reports documented the antiproliferative effect of either coriander extract or its pure phytochemicals [108,109,110,111]. In addition, coriander extract significantly suppressed the migration and invasion of murine melanoma cells as well as the number of metastatic regions in mice bearing B16F10 melanoma cells [112]. Furthermore, some of the coriander’s bioactive components showed also the potential to modulate the activity of skin cancer cells.
Fenugreek
Fenugreek (Trigonella foenum-graecum) is a herbaceous plant that belongs to the Leguminosae family. It is commonly used as a spice and medicinal herb, and its seeds are utilized in various culinary preparations, herbal supplements, and traditional medicines [113,114,115]. Fenugreek is native to the Mediterranean region, southern Europe, and western Asia and is widely cultivated in many parts of the world for its culinary and medicinal properties [116].
In traditional medicine, fenugreek is used for its potential benefits in addressing various health conditions, including but not limited to cancer, hypercholesterolemia, diabetes, cold cough, splenomegaly, hepatitis, backache, and inflammation [117]. Phytochemical analyses showed that fenugreek contains a diverse array of secondary metabolites, which include saponins, steroids, alkaloids, flavonoids, terpenes, phenolic acid derivatives, amino acids, as well as fatty acids and their derivatives [118,119]. Related to phenolic compounds, several flavonoids (e.g. quercetin, luteolin, vitexin, isovitexin, kaempferol, tricin, and naringenin) and phenolic acid (e.g. p-coumaric acid, caffeic acid, chlorogenic acid) have been isolated either from whole plants or seeds [120,121]. In the past decade, there has been extensive research on fenugreek and its active compounds as potential agents in the fight against cancer [122,123,124,125]. Regarding skin cancers, several phenolic fenugreek constituents have been reported to suppress the viability and growth of skin cancer cells either in vitro or in vivo.
Ginger
Ginger is the rhizome of Zingiber officinale a plant belonging to the ginger family (Zingiberaceae). The same family also includes other plants used as spices, namely turmeric, cardamom, and galangal. It is native to southeastern Asia and was later introduced to other parts of the world. Ancient civilizations, including the Chinese, Indians, Greeks, and Romans, valued ginger for its medicinal benefits [126]. Ginger has a pungent, spicy flavor and a warm aroma, which is why it is a popular ingredient in various cuisines worldwide. In traditional medicine, ginger was commonly used to treat digestive disorders, including nausea, indigestion, and flatulence. It was also used as a warming herb to improve circulation and alleviate symptoms of colds, coughs, and respiratory ailments [127]. The chemical composition of ginger reveals the presence of more than 400 distinct compounds. Ginger is rich in terpenes such as cineole, citral, limonene, α/β pinenes, β-elemene, farnesene zerumbone, zingiberene, and phenolic compounds including gingerols (e.g. 6-gingerol, 8-gingerol, 10-gingerol), shogalols (e.g. 6-shogaol, 8-shogaol, 10-shogaol). In addition, several other phenolic compounds have been identified in ginger including quercetin, gingerenone-A, zingerone, 8-paradol, or 6-dehydrogingerdione [128,129,130]. Ginger extract or its constituents were reported to have a broad spectrum of pharmacological activities such as antioxidant, anti-inflammatory, antimicrobial, neuroprotective, cardioprotective, and gastroprotective [128,131]. Moreover, there are numerous studies focusing on the anticancer effect of ginger [132,133].
Oregano
Origanum vulgare L., also known as oregano, is a Mediterranean plant species belonging to the Lamiaceae family which, nowadays, represents one of the most used culinary herbs. However, the application of oregano in several ethnobotanical practices, including folk medicine, dates to ancient times. Regarding its phytotherapeutic effect, various investigations have been performed, documenting that oregano essential oil possesses antimicrobial, antiviral, antifungal, antioxidant, anti-inflammatory, digestive, expectorant, and neuroprotective effects. In addition, antiproliferative and anticancer effects of oregano have been reported [134,135]. Although the chemical composition of oregano strongly depends on the vegetative periods of the growing season, generally carvacrol and thymol are the two main components of this spice. In addition, terpenes (e.g. α-thujene, α-pinene, camphene, sabinene, β-pinene, β-myrcene, α-phellandrene, α-cubebene, β–elemene, trans-caryophyllene, eucalyptol, linalool), flavonoids (e.g. rutin, naringin, hesperetin, naringenin, apigenin, luteolin, acacetin, vitexin) or phenolic acids (caffeinic acid, rosmarinic acid) has also been identified in oregano extract or essential oils [136,137].
Nutmeg
Nutmeg is a spice derived from the seed of the Myristica fragrans, which is native to Indonesia. Now, for commercial purposes, is widely cultivated in several countries, including India, Thailand, Japan, China, and South Africa. The spice is obtained by grinding the seed into a powder or grating it into a fine texture. As spice is commonly used in both sweet and savory dishes, as well as beverages. Additionally, nutmeg has been used in traditional medicine to treat anxiety, gastrointestinal discomfort, insomnia rheumatism, and also as an aphrodisiac [138,139]. Moreover, several pharmacological actions such as antidiabetic [85], anti-inflammatory and analgetic [140,141], antimicrobial [142,143], hepatoprotective [144] or antioxidant [145]. The effects of nutmeg are thought to be mediated by its bioactive constituents, which include lignans, neolignans, diphenyl alkanes, phenylpropanoids, terpenoids, alkanes, fatty acids, fatty acid esters, as well as a few minor components like steroids, saponins, triterpenoids, and flavonoids. Phenolic compounds found in nutmeg encompass various categories such as phenolic acids (e.g. protocatechuic acid, caffeic acid, vanillic acid, p-coumaric acid, ferulic acid, and sinapic acid), flavonoids (e.g. catechin, epicatechin, rutin, quercitrin, isoquercitrin, quercetin, and kaempferol), as well as other phenolic compounds like ellagic acid, myristicin, and elemicin [146,147].
Red chili
Red chili (Capsicum annuum L) known also as hot pepper, chili pepper or red pepper is one of the most widely cultivated spices from the genus Capsicum. This plant probably originated from central-east Mexico [148]. Archaeological evidence suggests that chili peppers have been cultivated and consumed in this region for at least 6,000 years. It has been utilized for centuries in different countries and civilizations. In ancient times, the Maya civilization employed pepper as a remedy for conditions such as asthma, coughs, and sore throats. Similarly, the Aztecs used it to alleviate toothaches. Chemical analyses of red chili showed a broad spectrum of active secondary metabolites. The spiciness of red chili peppers is primarily caused by capsaicinoids, alkaloids that are exclusively found in the Capsicum genus. Capsaicin and dihydrocapsaicin, among all capsaicinoids, account for approximately 77-98% of the pungency found in chili peppers. Other capsaicinoids include nordihydrocapsaicin, homodihydrocapsaicin, and homocapsaicin [149]. Furthermore, carotenoids responsible for the red color of chili peppers (capsanthin and capsorubin) or yellow-orange color (e.g. violaxanthin, zeaxanthin, β-cryptoxanthin, lutein, and β-carotene) have also been identified in a red chili fructi [150]. Among flavonoids, quercetin and luteolin have been described as the main flavonoids in Capsicum [151]. However, besides these two, several other flavonoids, including kaempferol, myricetin, apigenin, naringenin, catechin, epigallocatechin, and their derivatives, have also been identified in this spice [152]. Moreover, a number of phenolic acids such as gallic acid, protocatechuic acid, vanillic acid, hydroxyl cinnamic acids, caffeic acid, ferulic acid, chlorogenic acid, and cinnamic acid have been detected in red chili extract [153]. Capsaicin, a main active compound, exhibits a wide range of pharmaceutical applications due to its analgetic anti-arthritic and anti-inflammatory properties, as well as its effectiveness against bacterial infections, Moreover, it was also reported to suppress the growth of cancer cells [154,155].
Rosemary
Salvia rosmarinus Spenn. (syn. Rosmarinus officinalis), commonly known as rosemary, is a medicinal aromatic plant of the Lamiaceae family, native to the Mediterranean region. It is usually a small shrub with evergreen small needle-like leaves of light grey-green color. It is used with popularity as a spice and flavoring. Although it is most preferred as a spice in food, this herb is one of the most popular medicinal herbs in the world. Rosemary is used in folk medicine for wound healing, in treating inflammatory diseases and mycoses, and to relieve renal colic, muscle spasms, dysmenorrhea, and headaches. It also has antiviral, antibacterial, antithrombotic, antidepressant, and antioxidant effects [156,157]. Chemical analyses showed a broad spectrum of bioactive constituents. Numerous compounds were identified in the essential oil of rosemary. Among them, the predominant constituents were 1,8-cineol, camphor, α-pinene, limonene, camphene, and linalool, respectively [158]. Furthermore, several phenolic compounds have been identified in rosemary extract including phenolic acids (salvianic acid, caffeic acid, rosmarinic acid, and salvianic acid A), flavonoids (luteolin, luteolin−7-O-rutinoxide, luteolin-7-glucoronide, hesperidin, apigenin, cirsimaritin, genkwanin, and salvigenin) and phenolic diterpenes including rosmadial, 7-methylrosmanol, carnosol, carnosic acid and 12-methylcarnosic acid [159]. In addition to above mentioned biological effects, several studies have specifically investigated the mechanisms underlying the anticancer effects of rosemary or its active constituents [160,161].
Saffron
Crocus sativus, commonly known as saffron, is a flowering plant in the Iridaceae family. It is cultivated for its highly prized saffron spice, which is derived from the dried stigmas of the flowers. Saffron is known for its vibrant red color, distinct aroma, and unique flavor. This plant is primarily cultivated in regions with a Mediterranean climate, such as Iran, Spain, India, and Greece [162]. Apart from its culinary uses, saffron has also been traditionally valued for its medicinal properties. In traditional medicine, it has been used for antioxidant, anti-inflammatory, anti-vomiting, and mood-enhancing effects [163]. Analyzes of chemical composition showed that saffron contains numerous active constituents belonging to carotenoids such as crocin, crocetin, and
lycopene, monoterpenes including picrocrocin, crocusatins, isophorone, and amino acids and alkaloids (e.g. 5-methyluracil, pyridin-3-ylmethanol, uracil, harman, tribulusterine). Moreover, saffron also contains numerous phenolic compound including phenolic acids (e.g.protocatechuic acid, 4-hydroxybenzoic acid, vanillic acid benzoic acid, p-coumaric acid) or flavonoids and flavonoid glycosides (e.g. kaempferol, kaempferide, kaempferol-3-O-sophoroside-7-O-glucoside, kaempferol-3,7,4′-triglucoside, kaempferol 7-O-β-D-glucopyranoside, isorhamnetin-3,4′-diglucoside, isorhamnetin-3-O-glucoside, astragalin, sophoraflavonoloside, helichrysoside) [164].
Sichuan pepper
Plants belonging to the genus Zanthoxylum L. (the fruits of this genus are known as Sichuan pepper) form the largest group within the Rutaceae family and encompass approximately 250 species, including Z. bungeanum, Z. zanthoxyloides, Z. ovalifolium, Z. armatum, and many others [165]. These are deciduous, heat-loving, evergreen plants found in subtropical and warm climates worldwide, particularly in Asia, Africa, and America. Traditionally, they have been used as food and for their positive effects on the human body in the prevention and treatment of various diseases in countries like China, Japan, India, and Vietnam [166]. As a spice, not only the fruits but also the seeds, leaves, roots, and bark are utilized. It has a pungent aroma, and citrus-floral taste, and causes a tingling sensation in the mouth. Despite being commonly referred to as "pepper", it is not related to peppercorn from the Piperaceae family [167]. Current research pays significant attention to this genus. Many of its effects have been demonstrated, including anti-tumor, neuroprotective, antiparasitic, antimicrobial, antiviral, antioxidant, anti-inflammatory, antidiabetic, hepatoprotective, anti-obesity, and other properties [166,167,168]. These effects are mediated by bioactive compounds, among which volatile odor-bearing alcohols, ethers, terpenoids, and related compounds, as well as non-volatile alkyl amides, polyphenols, and their glycosides. Polyphenols include isovitexin, vitexin, hyperoside, isoquercitrin, rutin, foeniculin, trifolin, quercitrin, astragalin, and afzelin [169].
Star anise
Illicium verum Hook f., also known as badian, is an aromatic evergreen tree belonging to the Illiciaceae family [170]. It is named for its typically star-shaped fruit. It is naturally found almost exclusively in southern China and Vietnam but is also commercially grown in other Asian and European countries [171]. Star anise has a long history in China as both a spice and medicinal plant, listed in the Compendium of Materia Medica (Bencaogangmu) during the Ming Dynasty and in the Chinese Pharmacopoeia [170,172]. It is used in various forms, such as raw, dried, powdered, or as an essential oil. Traditionally, it has been used as a carminative, digestive, and spasmolytic for abdominal pain, colic, and vomiting, as a sedative for nervousness and insomnia, as an analgesic and anti-inflammatory for lower back pain, joint pain, and rheumatism, and also for symptoms of cold [170]. Its pleiotropic effects, including antimicrobial, antifungal, antiviral, antioxidant, anti-inflammatory, gastroprotective, and others are due to phytochemicals naturally occurring in star anise fruits [173,174,175,176]. Chemical analyses have shown that fruits contain significant amounts of terpenes, alkaloids, essential oils, and tannins. The most abundant phenolic compounds are phenylpropanoids cis- and trans-anethole (85-95%), estragole, anisylacetone, ρ-anisaldehyde, foeniculin, and others [177]. Additionally, it is a valuable source of shikimic acid which is an essential intermediate for the synthesis of the antiviral drug oseltamivir (Tamiflu®) [172]. In addition to the aforementioned health benefits, an anticancer effect of Illicium verum has been demonstrated in various in vitro and in vivo tumor models [178,179,180,181,182,183].
Sumac
Sumac, scientifically referred to as Rhus coriaria L., is a plant belonging to the Anacardiaceae family and is native to southern Europe. Its dried fruits are widely employed as a spice. Traditionally, sumac has been commonly utilized for addressing conditions such as gout, obesity, bleeding disorders, and diabetes. Its rich history of traditional usage aligns with its medicinal applications, making it a popular choice for these ailments [184]. Moreover, Rhus coriaria's phytochemicals have been reported to have several pharmacological effects including antioxidant, anti-inflammatory, antimicrobial antiemetic, hypolipidemic, or analgesic [185,186]. In addition, the anticancer effect of sumac against various types of cancers has also been reported [187,188,189]. The chemical analyses showed that sumac contains numerous active compounds including essential oils, steroids, terpenoids, and organic acids. However, the dominant groups of bioactive compounds found in sumac are phenolic compounds, mainly hydrolyzable tanins (e.g. galloylhexose, O-galloylmorbergenin, O-galloyl arbutin), flavonoids (e.g. quercetin, myrecetin, apigenin, kaempferol, quercetin 3-glucoside), phenolic acids (gallic acid, ellagic acid, protocatechuic acid) and anthocyanins (e.g. delphidin-3-glucoside, cyanidin 3-(2″-galloyl)galactoside, cyanidin-3-glucoside) [184,190,191,192].
Thyme
Thymus vulgaris L. known as thyme (the Greek word ‘thymos’ means strength or courage) belongs to the Lamiaceae family. This plant grows in dry and hot climates in southern Europe, North Africa, and several parts of Asia. The genus Thymus includes around 300 species distributed worldwide. Thyme has been widely used as a spice. Its essential oils are used in traditional medicine for their antiseptic, anthelmintic, expectorant, antispasmodic, antimicrobial, antiviral, antifungal, antioxidant, sedative, as well as antirheumatic, antihypertensive, and calming effects. Therefore, it is used to treat a variety of ailments (respiratory diseases, prevention of atherosclerosis, urinary tract infections, dyspepsia, toothache, and many others). Its aromatic extracts are also used in cosmetics [193,194]. In addition, several authors reported the potential of either thyme extract or thyme bioactive compounds to suppress the growth and viability of different cancer cells [195,196,197]. Analyzes of chemical composition revealed that thyme contains abundant secondary metabolites of which thymol and carvacrol are the major constituents. Other including terpenoids (e.g. β-pinene, α-fellandrene, p-cymene, m-cymene, eucalyptol, trans-caryophyllene), phenolic acids (e.g. caffeic acid, quinic acid, p-coumaric acid, ferulic acid, cinnamic acid, rosmarinic acid) and flavonoids (e.g. apigenin, naringnin, luteolin-7-o-rutinose) [198,199].
Turmeric
The ginger family (Zingiberaceae) includes the perennial herbaceous plant known as turmeric (Curcuma longa). Since antiquity, several uses have been made for its underground stems, known as tuberous rhizomes. They are used as a common condiment, a natural textile dye, and have therapeutic benefits as an aromatic stimulant. In addition to being a spice, turmeric has also been used historically as a perfume. It comes from southern India and Indonesia. Due to its flavor and color, turmeric is a common spice used in the creation of curries in India and other Asian nations [200]. Turmeric has been used for treating cough, diabetic wounds, hepatic diseases, cardiovascular illness, dyspepsia, and epigastric discomfort for ages in Chinese and Ayurvedic medicine. A study focusing on turmeric composition led to the extraction of more than one hundred and fifty secondary metabolites. These compounds comprised a diverse range of chemical classes including monoterpenoids, sesquiterpenoids, diterpenoids, and triterpenoids. Furthermore, the phytochemical analysis of turmeric also revealed the presence of saccharides, steroids, fatty acids, and alkaloids among its constituents. Moreover, turmeric is a rich source of phenolic compounds including curcuminoids (e.g. curcumin, demethoxycurcumin, bisdemethoxycurcumin) phenolic acids (e.g. gallic acid, protocatechuic acid, chlorogenic acid, cinnamic acid, ferulic acid), flavonoids (e.g. rutin, myricetin, genistein, quercetin, catechin, epicatechin) and other phenylpropanoids (e.g. coumarin) [201,202].
In addition to its various biological effects, the potential of turmeric and its active constituents as anticancer agents has been explored in recent research studies [203,204]. Furthermore, several of the aforementioned phenolic compounds have been investigated for their potential as agents against skin cancers [205,206,207,208].
5. Effect of spices-derived phenolic compounds against melanoma and non-melanoma skin cancer
5.1. Phenolic acids
Caffeic acid (CA) is widely distributed in nature and can be found in various plants, including coffee beans, fruits, vegetables, herbs, and spices. Caffeic acid has been studied for its potential in the prevention and treatment of skin cancer due to its antioxidant and anti-inflammatory properties. In the study conducted by Yang et al. (2014), CA in human A431 skin cancer cells suppressed colony formation and in non-cancer HaCaT cells prevented EGF-induced neoplastic transformation. Moreover, in animal experiments, it significantly decreased tumor incidence and volume in a solar UV (SUV)-induced skin carcinogenesis [209]. Furthermore, Western blot analyses demonstrated that CA effectively suppressed ERK1/2 activities both in vitro and in vivo. This indicates that CA may exert chemopreventive effects against SUV-induced skin carcinogenesis by specifically targeting ERK1/2 signaling pathways.
In JB6 P+ mouse skin epidermal cells, a separate study revealed that CA demonstrated the capability to inhibit UVB-induced COX-2 expression. This inhibition was achieved by suppressing the activity of Fyn kinase, which is a crucial player in skin carcinogenesis. Moreover, caffeic acid also blocked Fyn kinase downstream MAPKs contributing to its potential as a preventive agent against skin carcinogenesis caused by UVB exposure [210]. Additionally, it has been shown that CA effectively inhibits UVB-induced DNA damage, reduces the generation of ROS, and decreases the frequency of apoptotic cell death in human dermal fibroblasts. Moreover, it has been found to prevent the UVB-induced expression of PI3K and AKT kinases by activating PTEN, a tumor suppressor protein. This suggests that CA may possess protective properties against UVB-induced skin damage and potentially contribute to skin health and anti-cancer effects through these mechanisms [211]. Other authors documented the antiproliferative effect of CA in melanoma cells. Decreased cell viability, increased number of dead cells, and inhibited colony formation have been found in CA-treated human SK-Mel-28 melanoma cells. In addition, it increased the percentage of apoptotic cells and increased the number of cells at the G0–G1 phase of the cell cycle [212].
In another study, CA and chlorogenic acid (ChA) induced apoptosis in amelanotic melanoma C32 cells. This effect was potentiated with static magnetic field (SMF) exposition. Caffeic acid also significantly upregulated mRNA expression of the Casp3 and caspase-3 activity in C32 cells [213]. Later, they reported that both phenolic acids in the same melanoma cell line in combination with SMF decreased TGFβ protein levels [214]. Moreover, ChA in C32 cells suppressed the viability of melanoma cells and expression of genes of antioxidant enzymes superoxide dismutase (SOD1, SOD2) and glutathione peroxidase (GPX1), as well as activity of SOD, GPx, and catalase (CAT). Surprisingly, in combination with SMF, the activity of antioxidant enzymes was increased [215].
p-Coumaric acid (p-CA), another member of phenolic acids, has a broad spectrum of biological activities including anti-inflammatory, antioxidant, antidiabetic and hepatoprotective [216,217,218,219]. Morover, anticancer effect of p-CA has also been reported [220,221]. Regarding skin cancer, this phenolic acid decreased both human and murine melanoma cell viability and proliferation. The antiproliferative effect of p-CA has been associated with the cell-dependent arrest of the cell cycle – at the S phase in human A375 cells and at the G0–G1 phase in murine B16 cells. In human melanoma cells downregulation of cyclin A and CDK2 while in murine cells cyclin E and CDK2 have been observed. In addition, p-CA induced apoptosis in both melanoma cell lines [222]. In a recent study conducted by Gastaldello et al. (2021), gavage administration of p-CA to Balb/C mice bearing B16F10 melanoma cells resulted in a significant decrease in tumor volume and a reduction of a number of mitoses in the tumor region [223]. Moreover, p-CA modulated also some factors in the tumor microenvironment such as angiogenesis and inflammation.
Ferulic acid (FA) has been evaluated as a perspective compound for skin cancer prevention. Ultraviolet B (UVB) radiation can cause chronic skin inflammation which can result in the development of non-melanoma skin cancer, particularly basal cell carcinoma (BCC) and squamous cell carcinoma (SCC) [224]. In the context of UVB-induced skin inflammation, reducing inflammation can help minimize the risk of cancer formation by mitigating the inflammatory response and subsequent DNA mutations. In the study by Stanifort and co-workers (2012), topically applied FA attenuated UVB-induced changes in the skin of experimental animals [225]. Simultaneously, FA caused the degradation of matrix metalloproteinase-2 and -9 (MMP-2 and MMP-9) overexpressed after UVB irradiation. Later, it was documented that the protective effect of FA in UVB skin carcinogenesis was associated with the modulation of angiogenesis, inflammation, and apoptosis. UVB irradiation increased the expression of several angiogenic and inflammatory markers such as iNOS, IL-6, TNF-α, or VEGF. Moreover, the expression of anti-apoptotic Bcl-2 protein was upregulated while anti-apoptotic Bax was downregulated. The administration of FA before the UVB exposition significantly reverted all of these events [226].
Gallic acid (GA), a phenolic compound broadly distributed in the plant kingdom, has been referred to possesses numerous biological activities [227]. Regarding skin cancers, GA has been referred to induce apoptosis in A375.S2 human melanoma cells. Molecular analyses showed that GA induced both caspase-dependent and caspase-independent pathways of apoptosis [228]. Subsequently, these researchers found that GA can hinder the invasiveness of melanoma cells by inhibiting MMP-2 through its involvement in the Ras and ERK signaling pathways [229]. A great proteomic analysis performed in the study by Liu et al. (2014) revealed the co-occurrence of apoptosis and modulation of glycolysis in the GA-treated B16F10 melanoma cells [230]. It was shown that gallic acid stimulated the mitochondrial apoptosis pathway and simultaneously upregulated several proteins involved in glycolysis including glucokinase, pyruvate kinase, glyceraldehyde-3-phosphate dehydrogenase α-enolase, and aldolase.
Protocatechuic acid (PA), a natural phenolic compound, is commonly found in various plant-based foods, such as fruits, vegetables, certain medicinal plants, and spices. In the study by Tseng and co-workers (1998), PA significantly inhibited the incidence of chemically-induced skin tumors and simultaneously inhibited the activity of epidermal ornithine decarboxylase and myeloperoxidase as well as the formation of hydrogen peroxide in the mouse skin [231]. On the other hand, Nakamura et al. (2000) found that PA at a low dose (16 nM) decreased the number of tumors, and, surprisingly, at a higher dose (1,600 nM) it increased the number of tumors [232]. The authors later verified that the effect of PA was dose-dependent. Application of this phenolic acid at a dose of 16 nM significantly prevented inflammation and oxidative stress. However, at a higher dose of 1,600 nM, it had the opposite effect and actually intensified both inflammation and oxidative stress [233]. In addition, a study conducted by Lin et al. (2010) revealed the anti-metastatic potential of PA [234]. It significantly decreased the number of liver metastasis in mice injected with B16-F10 melanoma cells. Furthermore, levels of MMP-2, as well as the expression of Ras and phosphorylated Akt, were downregulated in PA-treated animals.
Rosmarinic acid (RA), another phenolic acid found in several spices, decreased viability and suppressed migration of metastatic SK-MEL-28 melanoma cells. Rosmarinic acid-induced apoptosis was linked to increased caspase-8 and caspase-3 gene expression together with increased activity of caspase-3. Moreover, decreased gene expression of NLRP3 inflammasome has also been observed [235]. In another study, RA showed an inhibitory effect on melanoma cells via suppression of the ADAM17/EGFR/AKT/GSK3β axis resulting in the inhibition of melanoma cell proliferation, invasion, and migration. In addition, RA also enhanced the sensitivity of melanoma cells to cisplatin [236]. Besides, the radiosensitizing effect of RA in melanoma B16F10 cells was documented and this effect can be associated with decreased GSH-dependent protection against oxidative stress [237,238].
Syringic acid (SA), a phenolic compound found in numerous plants, has been reported to exhibit a wide range of biological activities such as antioxidant, anti-inflammatory, antimicrobial, and anticancer [239,240,241]. Regarding skin cancers, SA remarkably inhibited UVB-induced tumor incidence. Molecular analyses in human epidermal keratinocytes exposed to UVB irradiation showed that SA significantly downregulated the expression of COX-2, MMP-1, and prostaglandin E2. Additionally, this phenolic acid inhibited MAPK and EGFR phosphorylation induced by UVB [242].
The chemical structures of phenolic acids discussed in this review are shown in Figure 2.
5.2. Flavonoids
Apigenin, similar to many other flavonoids, has been found in different fruits and vegetables and a number of studies documented its broad spectrum of biological activities [243] including anticancer effects [244]. In addition, a number of studies were focused on its anticancer effect against both melanoma and non-melanoma skin cancers. In the study conducted by Zhao and co-workers (2017), apigenin decreased viability, migration, and invasion in A375 and C8161 human melanoma cells [245]. Moreover, cell cycle arrest at the G2/M phase and apoptosis induction has been promoted in apigenin-treated cells. In addition, apigenin decreased the activation of several proteins including ERK 1/2, AKT, and mTOR. Additionally, the apigenin-induced anti-metastatic effect was associated with the modulation of STAT3 activity. In murine melanoma B16F10 cells, apigenin decreased STAT3 phosphorylation and STAT3 nuclear localization resulting in suppression of STAT3 transcriptional activity. Furthermore, apigenin was observed to decrease the expression of genes targeted by STAT3, including MMP-2, MMP-9, VEGF, and Twist1, which play a role in cell invasion and migration [246]. In another study, it was shown that apigenin induced apoptosis in the A375SM human melanoma cells via upregulation of phosphorylated p38 with simultaneous downregulation of ERK, JNK, and Akt, the kinases involved in the proliferation and growth of melanoma cells [247]. Besides, it was reported that the antiproliferative and anticancer effects of apigenin can be associated with immune response modulation. Xu et al. (2018) found, that apigenin suppressed PD-L1 expression in melanoma cells melanoma cells that have become more sensitive to cytotoxic T cells [248]. In addition, the same effect was observed in apigenin-treated host dendritic cells resulting in an augmented host immune response.
In experimental animals, apigenin has been found to prevent UVB-induced skin inflammation and carcinogenesis via different mechanisms. In the study by Mirzoeva and co-workers (2018), apigenin suppressed carcinogenesis in mice exposed to UVB irradiation due to maintaining of expression of thrombospondin-1, which serves as a natural inhibitor of angiogenesis and tumorigenesis [249]. Furthermore, other targets such as mTOR, Src kinase, and COX-2 are also suggested [250,251,252]. In addition, in the study conducted by Caltagirone et al. (2000), apigenin was found to delay tumor growth in mice bearing B16-BL6 melanoma cell xenograft [253]. Moreover, apigenin also significantly decreased the number of colonies in the lung which suggest the anti-invasive and anti-metastatic potential of this flavonoid.
Astragalin (kaempferol-3-glucoside) is a significant component of plant extracts that have demonstrated antioxidant, anti-inflammatory, antimicrobial, antidiabetic, antidepressant, and anticancer effects [254]. The cytotoxic effect of astragalin on A375P and SK-MEL-2 melanoma cells was evidenced by an increase in the sub-G1 population and TUNEL-positive cell population, indicating the presence of DNA fragmentation in astragalin-treated cells. Cell death was accompanied by a decrease in cyclin D1 expression and the anti-apoptotic Bcl-2 protein Mcl-1, upregulation of Bax, and activation of caspase-3. A specific pro-apoptotic effect of astragalin includes a significant reduction in SOX10 expression in non-transfected cells as well as in cells overexpressing SOX10 [255]. SOX10 (Sry-related HMg-Box Gene 10) is a transcription factor that facilitates neural crest cell development and is also involved in melanoma cell growth through the regulation of immune checkpoint protein expression [256].
Catechin and its related molecules, such as epigallocatechin-3-gallate (EGCG) and epicatechin-3-gallate (ECG) have been found to possess antiproliferative and anticancer properties, including in skin tumors [257,258,259]. The main mechanism of their action is the induction of apoptosis and cell cycle arrest. For example, in A375 cells, in addition to mitochondrial damage, changes in the level of the Bcl-2 protein, and activation of caspase -3, catechin-induced modulation of autophagy has also been observed. Increased PI3K, Akt, and mTOR phosphorylation activated this signaling pathway that suppresses autophagy. Accordingly, reduced levels of other autophagic proteins such as Beclin-1, LC3, and phospho-AMPK were observed [260]. Immunotherapy is also a significant treatment approach for melanomas, and research on natural substances with anticancer effects in this area is continuously advancing. A study by Ravindran Menon et al. (2021) investigated the influence of EGCG on immune responses and the tumor microenvironment in human metastatic melanoma cell lines 1205Lu, HS294T, and A375 [261]. It was found that EGCG inhibited IFN-γ-induced PD-L1/PD-L2 expression. This effect depended on the suppression of JAK/STAT signaling, which mediates the interaction between cytokine receptors and the cell nucleus. Similar to in vitro experiments, in vivo inhibition of the JAK/STAT pathway led to reduced tumor growth in mice, decreased Ki-67-positive cells, and increased expression of granzyme in CD8+ cells in the tumor microenvironment. CD8+ T cells were found to be essential for suppressing tumor growth, and treatment with EGCG reactivated the immune response against the tumor even more effectively than anti-PD-1 antibody treatment.
Chrysin has also been referred to suppress melanoma cell proliferation and invasion. In human A375.S2 melanoma cells, chrysin inhibited cell migration and invasion via matrix metalloproteinase-2 (MMP-2) inhibition, and due to inhibition of expression or activation of several signaling pathways including phosphoinositide 3-kinase (PI3K), protein kinase B (AKT), and protein kinase C (PKC), focal adhesion kinase (FAK), Ras homolog family member A (RhoA), p-c-Jun. Moreover, the downregulation of N-cadherin and upregulation of E-cadherin indicated that chrysin can also affects epithelial-mesenchymal transition [262]. Another study documented the antiproliferative and anticancer effects of chrysin in vitro and in vivo [263]. In murine B16F10 melanoma cells, exposition to chrysin led to the induction of cell cycle arrest at the G2/M phase associated with apoptosis induction. Moreover, in BALB/c mice bearing melanoma xenograft chrysin decreased the size and weight of tumors after 21 days of treatment. In addition, the cytotoxic activity of NK cells, cytotoxic T lymphocytes, and macrophages has been found in chrysin-treated animals. Results of another study showed that chrysin induced apoptosis in human and murine melanoma cells through caspase activation and modulation of the activity of MAPK members including extracellular signal-regulated kinase (ERK) 1/2 and p38 MAPK [264].
Diosmin, a naturally occurring flavonoid identified in some spices, has been found to act synergistically with IFN-α in the treatment of metastatic pulmonary melanoma [265]. Diosmin is also able to induce apoptosis in non-melanoma skin cancer A431 cells associated with increased ROS generation, downregulation of Bcl-2, MMP- and MMP-9 as well as upregulation of p53, caspase-3, and caspase-9 [266].
In the above-mentioned study [267], diosmin significantly reduced the number of lung metastasis as well as growth and invasion index in animals bearing B16F10 cell xenograft. In another study, these authors compared the ability of diosmin, grape seed extract, and red wine to suppress the growth of lung metastasis in B16F10 melanoma cells inoculated experimental animals. Ahong studied agents, diosmin showed the greatest reduction in pulmonary metastases [268].
Galangin has been documented to suppress the proliferation of the murine melanoma B16F10 cells in a dose-dependent manner. Moreover, galangin significantly reduced the ability of melanoma cells to form colonies indicating its antimetastatic potential as well. This effect has been associated with decreased cell adhesion to fibronectin and decreased cell motility and migration. In addition, galangin inhibited lung metastasis in mice bearing B16F10 xenografts [269]. Later, Benguedouar and co-workers (2016) presented that galangin-induced suppression of melanoma cell growth was associated with the induction of autophagy and apoptosis [270]. In addition, it was shown that galangin rescued human keratinocytes from undergoing apoptosis triggered by UVB-induced oxidative stress by restoring mitochondrial function and reducing the levels of apoptotic proteins [271]. In another study, they found that galangin in human keratinocytes increased the expression of GSH-synthesizing enzymes via activation of ERK/AKT-Nrf2 (nuclear factor erythroid 2-related factor 2) signaling [272]. These results indicate the possible preventive effect of galangin against UVB-induced skin carcinogenesis.
Genistein (4′, 5, 7-trihydroxy isoflavone) is a natural isoflavone and phytoestrogen mostly found in soybeans and also in other foods - alfalfa and clover sprouts, barley meal, broccoli, cauliflower, and sunflower, caraway, and clover seeds [273].
It has a variety of pharmacological activities including anti-inflammatory, antioxidant, and anticancer. Furthermore, the anticancer effect of this isoflavone has also been intensively studied [274,275]. Regarding melanoma cells, genistein was referred to inhibit human and animal melanoma cell growth and proliferation. These effects were associated with cell cycle arrest at the G2/M phase and cell differentiation promotion [276]. As reported by Darbon and co-workers (2000), G2/M arrest in human melanoma cells was associated with impaired dephosphorylation of Cdk1 and checkpoint kinase Chk2 activation [277]. Through detailed molecular analyzes, the actions of genistein have been found to operate through a multifactorial mechanism. It is known that activation of the FAK/paxillin pathway and mitogen-activated protein kinases (MAPKs) signaling may serve as a possible indicator of melanoma metastasis. Genistein in B16F10 cells inhibited proliferation, invasion, and cell migration and induced apoptosis in a dose-dependent manner. Moreover, genistein significantly suppressed the expression of numerous proteins including a phosphorylated form of FAK, paxillin, p38, ERK, and JNK as well as the expression of tensin-2, vinculin, and α-actinin. Additionally, real-time PCR showed decreased gene expression of FAK and paxillin. Interestingly, it has been observed that low concentrations of genistein can activate the FAK/paxillin and MAPK signaling pathways, leading to enhanced invasion and migration of melanoma cells [278]. Another study reported the ability of genistein to inhibit the growth of A375SM melanoma cells via modulation of p21, cyclin E, and/or cyclin B genes expression and stimulation of ROS production followed by activation of p53 and p38 MAPK and activation of the ER stress-mediated apoptotic pathway [279]. Another author showed that genistein inhibited basal and PGE2-induced proliferation of melanoma cells and this effect was associated with suppression of IL-8 and EP3 receptor expression [207]. Moreover, genistein has been shown to increase the sensitivity of melanoma cells to cisplatin, as their combination has significantly induced apoptosis while suppressing the expression of antiapoptotic proteins [280]. In addition to melanoma cells, genistein has also been reported to restrict cell growth in the HN4 squamous cell carcinoma due to blocking of the cell cycle at the S/G2-M phases and apoptosis induction in a time- and dose-dependent manner. Molecular analyzes showed the down-regulation of Cdc25C which plays an important role in the activation of the cyclinB1-Cdk1 complex [281].
In addition to in vitro studies, several in vivo experiments confirmed the anticancer effect of genistein in the xenograft model of carcinogenesis, chemically induced skin carcinogenesis, or UV-induced skin cancer. Genistein significantly prevented the growth of tumors in the female C57BL6J mice subcutaneously injected with B16 melanoma cells. Tumor volume was significantly reduced (by 50%) in animals feeding a diet with genistein compared to the untreated group [282]. Furthermore, genistein suppressed the growth of lung metastasis in C57BL/6 mice inoculated B16F-10 melanoma cells. Moreover, genistein significantly prolongs the survival of tumor-bearing animals [283]. In addition, in the study conducted by Farina and co-workers (2006), it was found that genistein in experimental animals injected with B16F0 melanoma cells significantly inhibited the formation of new blood vessels by tumor implants [284]. In another study, Wei et al. (1998) reported that genistein significantly decreased the incidence and multiplicity of skin tumors in two-stages tumor models when dimethylbenz[a]anthracene /DMBA) was used as an initiator and 12-O-tetradecanoyl phorbol-13-acetate (TPA) as a promotor of skin carcinogenesis [285]. Genistein significantly reduced the DMBA-induced formation of DNA adduct as well as reduced TPA-induced H2O2 formation and inflammatory reaction. Furthermore, the preventive effect of genistein on UV-induced skin carcinogenesis has also been documented. Later, they reported a protective effect of genistein in UVB-induced skin carcinogenesis in mice. Genistein was applicated either orally or topically. They found that genistein inhibited tumor incidence and multiplicity although topically applied was more effective. Moreover, a strong protective effect of genistein was found when UVB irradiation was combined either with DMBA or TPA [286]. Because UV irradiation induces ROS production, it is believed that the protective effect of genistein can be related to its antioxidant effect [287].
Isoquercitrin or quercetin-3-O-β-D-glucopyranoside, found similarly to other flavonoids in fruits, vegetables, and other plant-based foods and beverages, has been studied on several skin tumor cell lines (SK-MEL-2, SK-MEL-28, B16) and non-tumor cell lines (HaCaT) [288]. At a concentration of 25 µmol/L, it significantly inhibited the viability and clonogenicity of SK-MEL-2 cells and induced their death, but this effect was not observed in healthy cells. Apoptotic cell death was preceded by DNA fragmentation and cell cycle arrest in the G1/S phase of the cell cycle. This led to the activation of the mitochondrial apoptotic pathway, which was accompanied by a decrease in the anti-apoptotic protein Bcl-2, and an increase in pro-apoptotic proteins Bax, AIF, Endo G, and cleaved PARP. Caspases contribute to PARP cleavage. After isoquercitrin treatment, there was a reduction in inactive forms of caspases, pro-caspases -8 and -9. Another important mechanism is the modulation of the signaling pathways. The PI3K/Akt/mTOR transduction pathway has a significant impact on cell proliferation, survival, growth, cell cycle regulation, and cell death [289]. Isoquercitrin significantly reduced the levels of phosphorylated forms of all its members, indicating that the antiproliferative effect of IQ is also mediated by the suppression of the PI3K/Akt/mTOR signaling pathway [290].
Isorhamnetin, a derivative of quercetin (quercetin-3-methyl ether), has been shown to have a wide range of biological effects including anticancer [291]. In melanoma B16F10 cell isorhamnetin dose-dependently inhibited proliferation and migration. Moreover, the exposition of cells to isorhamnetin resulted in apoptosis induction associated with the dysregulation of Bcl-2 family proteins and caspase-3 activation. Moreover, it inhibited the phosphorylation of Akt and suppressed the nuclear accumulation of NF-κB. In addition, animal experiments showed that isorhamnetin significantly decreased the volume of tumors initiated by B16F10 cell injection in C57BL/6 mice [292]. Another study showed the anticancer effect of isorhamnetin in non-melanoma cancer cells [293]. In vitro, isorhamnetin prevented EGF-induced neoplastic transformation of JB6 epidermal cells. Moreover, isorhamnetin inhibited the growth of A431 squamous carcinoma cells as well as the expression of COX-2, an important inflammatory mediator associated with skin carcinoma. In vivo, significant suppression of tumor volume and tumor weight in isorhamnetin-treated cells has been observed. Subsequent molecular analyses showed aberrant activation/deactivation of several signaling pathways such as ERK, Akt, PI3, and MEK in isorhamnetin-treated cells. Furthermore, Li and co-workers (2012) reported that quercetin-3-methyl ether suppressed the malignant transformation of mouse skin epidermal JB6 cells exposed to 12-O-tetradecanoylphorbol-13-acetate [294]. In addition, this chalcone inhibited the phosphorylation of ERK2 and induced cell cycle arrest at the G2/M phase.
Kaempferol, a flavonoid found in several spices, is also a potent inhibitor of melanoma cell proliferation and survival. In the study by Qiang et al. (2021), kaempferol induced cell cycle arrest at the G2/M phase in murine melanoma cells followed by induction of apoptosis. Moreover, kaempferol also decreased the volumes and weights of tumors in mice injected with melanoma cells [295]. Other authors showed that kaempferol-induced apoptosis was associated with the downregulation of the m-TOR/PI3K/AKT pathway in human melanoma A375 cells [296]. Furthermore, kaempferol has been recently reported to inhibit melanoma metastasis due to the suppression of aerobic glycolysis of both human and mouse melanoma cells [297]. In another study, this flavonoid has been found to suppress UV-induced skin cancer. In experimental animals, local administration of kaempferol reduced tumor volume and incidence by 68% and 91%, respectively. As showed detailed analyses, this effect has been associated with the inhibition of p90 ribosomal S6 kinase (RSK) and mitogen and stress-activated protein kinase (MSK), downstream molecules of the MAPK cascade [298]. In addition, the protective effect of kaempferol in UV-induced skin cancer can also be associated with the inhibition of Src kinase activity, which is an upstream MAPK regulator [299].
Luteolin, a flavonoid found in numerous fruits and vegetables, has been reported to possess a multi-targeted mechanism of anti-melanoma action. The research conducted by Schomberg et al. (2020) revealed that the growth inhibition of melanoma cells induced by luteolin was linked to the alteration of numerous genes involved in various pathways, including the extracellular matrix (ECM) pathway, oncogenic pathway, and immune response signaling [300]. Moreover, luteolin also showed a significant anticancer effect in nude mice injected with melanoma cells. In another study, luteolin suppressed melanoma cell proliferation and induced apoptosis via the inhibition of phosphorylation of AKT1 and PI3K, two pathways involved in the control of cell growth and survival. In addition, downregulated expression of MMP-2 and MMP-9 with concomitant increased expression of tissue inhibitors of metalloproteinases TIMP-1 and TIMP-2 has also been observed in the luteolin-treated A375 melanoma cells [301].
Epithelial-mesenchymal transition (EMT) plays an important role in cancer progression and metastasis. During cancer metastasis, tumor cells can undergo EMT, which allows them to detach from the primary tumor, invade surrounding tissues, enter the bloodstream or lymphatic system, and establish secondary tumors in distant organs. EMT is associated with downregulated expression of epithelial markers such as E-cadherin, while the expression of mesenchymal markers, such as N-cadherin, vimentin, and fibronectin, is upregulated [302]. The study realized by Li et al. (2019) indicated that the antiproliferative effect of luteolin in melanoma cells can be associated with decreased expression of proteins involved in hypoxia-inducible factor-1α/vascular endothelial growth factor (HIF-1α/VEGF) signaling pathway including HIF-1α, VEGF-A, and vascular endothelial growth factor receptor (VEGFR-2) [303]. In addition, decreased expression of MMP-2 and MMP-9 as well as p-Akt has also been found. Moreover, luteolin reversed EMT via N-cadherin and vimentin downregulation and E-cadherin upregulation.
Additionally, another mechanism leading to suppression of proliferation and induction of melanoma cell death such as induction of ER stress, generation of reactive oxygen species (ROS), cell cycle arrest, inhibition of β3 integrin or signal transducers and activators of transcription 3 (STAT3) signal pathways has also been reported [304,305,306,307].
Myricetin is a naturally occurring flavonoid with wide distribution in various food sources. As a flavonol, it is predominantly found in onions, red wine, tea, berries, fruits, vegetables, medicinal herbs, and spices. In berries, vegetables, and fruits, myricetin is commonly present in the form of glycosides rather than free aglycones. Notably, the skins of several fruits, like red grapes, contain notably high levels of myricetin [308]. Myricetin exhibits strong anticancer activity and demonstrates promising therapeutic potential by targeting and regulating the expression of several molecular targets involved in critical processes such as cell proliferation, invasion, and metastasis as well as inflammation, apoptosis, and angiogenesis [309]. Jung et al. (2008) show that myricetin reduced UVB-induced COX-2 expression in mouse skin epidermal cells and this effect has been associated with the inhibition of Fyn kinase activity which plays a crucial role in the development of skin cancers [310]. Moreover, myricetin also significantly suppressed skin tumor incidence in animals exposed to UVB in a dose-dependent manner. In addition, the antiproliferative activity of myricetin against A431 human skin cancer cell lines was also evaluated. Myricetin suppressed A431 cell proliferation and decreased their colony-forming capacity. Moreover, it induced apoptosis associated with changes in the Bcl2/Bax ratio increased production of ROS, and by reducing mitochondrial membrane potential. In addition, myricetin significantly inhibited the migration and invasion of skin cancer cells [311]. Furthermore, it was found that myricetin inhibited TPA-induced COX-2 expression in JB6 P+ cells by modulation of NF-кB activation [312]. In another study, these authors found that myricetin inhibited TPA- or EGF-induced transformation of JB6 P+ mouse epidermal cells together with inhibition of mitogen-activated protein kinase kinase (MEK), a kinase involved in tumor development. This effect was associated with the inhibition of ERK and RSK phosphorylation [313]. Later, Kumamoto et al. (2009) showed that EGF-induced JB6 P+ transformation can be efficiently inhibited by myricetin via inhibition of Janus Kinase (JAK) 1 phosphorylation and, to a lesser extent, via STAT3 inhibition [314]. In addition, myricetin has been documented that in vivo, in mouse skin, myricetin exerts a suppressive effect on UVB-induced angiogenesis by modulating the activity of PI-3 kinase [315].
Naringenin, another flavonoid present in several spices, has been reported to possess an antiproliferative effect in a melanoma model. In a recent study, naringenin in SK-MEL-28 human and B16F10 murine melanoma cells induced apoptosis associated with activation of caspase-3 and PARP cleavage. Furthermore, in naringenin-treated melanoma cells suppression of migration with a concomitant decrease of ERK1/2 and c-Jun N-terminal kinase (JNK) phosphorylation was observed. In addition, naringenin also inhibited angiogenesis as documented by suppression of human umbilical vein endothelial cells (HUVECs) migration, inhibition of tube formation of HUVECs as well as microvessel sprouting from rat aorta [316]. Another study showed that the antiangiogenic effect of naringenin can be associated with the inhibition of Two-Pore Channel 2 activity, an intracellular Ca2+ channel involved in several pathophysiological processes such as cancer cell proliferation, metastasis or angiogenesis [317,318]. In the study by Lentiny and co-workers (2007), the anti-metastatic potential of naringenin has been evaluated [319]. Oral administration of naringenin to C57BL6/N mice bearing B16F10 cells significantly decreased the number of lung metastases by about 69% compared to untreated animals. Furthermore, in the study by Ahamad et al. (2014), naringenin has been reported to inhibit proliferation and induce apoptosis also in non-melanoma human epidermoid carcinoma A431 cells [320]. DNA fragmentation, ROS generation, mitochondrial depolarization, and caspase activation were associated with apoptosis in naringenin-treated cells.
Quercetin, a natural flavonoid found abundantly in vegetables and fruits, is believed to have a variety of health benefits, including anti-oxidative, anti-inflammatory, antimicrobial, boosting the immune system, cardioprotective, antidiabetic or anticancer [321]. Regarding skin cancers, quercetin's anti-cancer effects can be attributed to several potential mechanisms. Quercetin has been found to inhibit proliferation and induce apoptosis in both, melanoma and non-melanoma cells. It exerts these effects by modulating various mechanisms such as cell cycle arrest, downregulation of the anti-apoptotic proteins, and upregulation of the pro-apoptotic protein expression or caspase activation [322,323,324]. In addition to these effects, quercetin has been reported to modulate the activity of several signaling pathways associated with cell survival, growth, migration, malignant transformation, or cell death. Cao et al. (2014) demonstrated that the anticancer effect of quercetin can be associated with the inhibition of STAT3 phosphorylation and subsequent suppression of STAT3 nuclear localization [325]. Moreover, quercetin also decreased STAT3 downstream gene expression including MMP-, MMP-9, and VEGF. According to Kim et al. (2019), the mechanism of the antiproliferative effect of quercetin in human A375SM melanoma cells may include the regulation of the MAPK signaling pathway [326]. Moreover, significantly increased JNK and p38 phosphorylation has been observed in the tumors from mice transplanted with A375SM melanoma cells. In another study, the exposition of murine melanoma B16-BL6 cells to quercetin resulted in significant inhibition of the cell invasion and migration associated with the suppressed gelatinolytic activity of MMP-9 and PKC signaling [327]. Some other studies showed that quercetin is capable to decreased the metastatic potential of melanoma cells also via suppression of EMT. In the study conducted by Lin et al. (2011), it was observed that treating the highly invasive human epidermal carcinoma A431-III cells with quercetin caused a transformation of their mesenchymal-like morphology into an epithelial-like morphology [328]. Additionally, the treatment with quercetin resulted in the downregulation of mesenchymal markers such as N-cadherin, vimentin, fibronectin, Twist, and Snail while simultaneously upregulating the expression of the epithelial marker E-cadherin. Later, Patel and Sharma (2016) reported that quercetin in SK-MEL-28 human melanoma reversed collagen I-induced EMT via suppression of the expression of mesenchymal markers including vimentin, N-cadherin, Twist, and SNAIL [329]. Conversely, quercetin also increased the expression of epithelial markers such as VCAM1, ICAM1, and E-cadherin. Overall, these findings indicated that quercetin had the ability to reverse the EMT process and promote the restoration of an epithelial phenotype in melanoma cells.
In addition, the association between the antiproliferative effect of quercetin and the modulation of signaling pathways such as Src/Stat3/S100A7 signaling [330], hepatocyte growth factor/c-Met signaling [331], epidermal growth factor signaling [332], phosphoinositide 3-kinase signaling [333] or retinoic acid-inducible gene I signaling [334].
Rutin, a flavonoid found in many plants, has a broad spectrum of biological activities such as anti-inflammatory, anti-oxidative, neuroprotective, anti-nociceptive, anticancer and many others [335]. In human melanoma RPMI-7951 and SK-MEL-28 cell lines, rutin decreased viability, caused loss of adherence, and initiated changes in cell morphology. Moreover, rutin induced nuclear fragmentation, and chromatin condensation indicating apoptosis induction. In addition, rutin has been found to induce senescence in melanoma cells as confirmed by increased expression of beta-galactosidase, a protein associated with cell senescence [336]. Furthermore, in the study by Martínez Conesa et al. (2005) anti-metastatic potential of rutin has been evaluated [267]. Results of the study showed the reduction of lung metastasis in rutin-treated animals injected with B16F10 melanoma cells. The ability of rutin to inhibit the lung tumor nodule induced by B16F10 melanoma cells injection in experimental animals has also been documented by Menon et al. (1995) [337]. Another study showed the potential of rutin to inhibit UVB-induced inflammation of hairless mouse skin. Topical application of rutin before UVB radiation significantly reduced UVB-induced expression of several pro-inflammatory mediators such as COX-2, and iNOS as well as prevented activation of signaling pathways including STAT3, p38 MAPK, and JNK [338].
Vitexin, a flavonoid found in various plants, particularly in fruits, vegetables, and certain medicinal herbs and spices. It is often recognized for its potential health benefits due to its antioxidant and anti-inflammatory properties. Regarding skin cancers, in both extract and pure substance forms, vitexin has demonstrated the remarkable ability to effectively inhibit the growth, proliferation, and ability to form colonies of various melanoma cell lines, including mouse melanoma cell B16F10 and human malignant melanoma cell lines A375, and vemurafenib-resistant melanoma cells (A375, Sk-Mel-5, and Sk-Mel-28). In addition, vitexin induces a block at the G2/M phase of the cell cycle and apoptosis. In vitexin-treated cells, down-regulation of the anti-apoptotic protein Bcl-2 and up-regulation of the pro-apoptotic protein Bax have been observed followed by cleavage of the DNA repair enzyme PARP. The antiproliferative activity of vitexin was associated with DNA damage induced by increased levels of ROS and oxidative stress. At the protein level, phosphorylated ATM and ATR, protein kinases activated in response to DNA damage, were significantly upregulated. In addition, checkpoint kinase Chk2 was activated, levels of tumor suppressor proteins p21 and p53 were increased, and accumulation of γH2AX, which interacts with kinases such as ATM and ATR, occurred. These results are consistent with in vivo experiments on xenografted mice, where vitexin at doses of 40 and 80 mg/kg resulted in a significant reduction in tumor growth [339]. As reported by Zhang et al. (2020), vitexin in human melanoma A375 and C8161 cells inhibited the expression of MMP-2 and MMP-9 as well as the expression of vimentin, Slug and Twist a transcription factor involved in EMT [340]. In addition, vitexin also decreased the phosphorylation of Src, Janus Kinase (JAK)1, and JAK2 kinases followed by the deactivation of STAT3 signaling.
The chemical structures of flavonoids discussed in this review are shown in Figure 3.
5.3. Other phenolic compounds
1’-acetoxychavicol acetate (ACA), derived from A. galanga, has been identified as the primary compound with a range of biological activities [71]. Among others, ACA has been evaluated as a chemopreventive agent in experimental skin cancers. Murakami et al. (1996) found that ACA significantly reduced 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced skin cancers in female JCR mice [341]. In addition, ACA was found to be an inhibitor of reactive oxygen species (ROS) production and a suppressor of lipid hydroperoxide formation. Another study confirmed the chemopreventive effect of ACA in TPA-induced skin carcinogenesis and this activity was associated with the inhibition of NF-κB activation [342].
trans-Anethole is classified as a phenylpropene and is found in various plants, particularly in spices and herbs. It has been demonstrated that trans-anethole has excellent antibacterial, antifungal, antioxidant, and anti-inflammatory effects and it is „generally recognized as safe“ and is, therefore, a valuable substance for use in medicine and skincare [171]. Its antiproliferative activity has been demonstrated on CRL-6475 tumor cells, where it exhibited significant selectivity compared to healthy human epidermal melanocytes (HEMa-LP) [343]. In concentrations of 3 - 12 µmol/L, trans-anethole inhibited the proliferation and clonogenicity of tumor cells. Apoptosis was detected using double staining with AO/EB and Annexin V/PI, and there was an increased population of dead cells. Furthermore, this phenylpropanoid significantly upregulated the expression of miR-498. When the cells were exposed to the miR-498 inhibitor, the antiproliferative effect of anethole was attenuated, demonstrating its critical role in cell proliferation, invasiveness, and metastasis [344,345]. The authors assume that this effect is caused by the modulation of the miR-498/STAT4 axis. The pro-apoptotic effects of alcoholic star anise extract, rich in trans-anethole, were demonstrated in several tumor cell lines, including the human melanoma cell line M14WM. They found a dose-dependent inhibition of proliferation (10, 25, and 50 μg/mL) and morphological changes typical of apoptosis [346].
Arbutin, a hydroquinone glucoside detected in many plants, is capable of inhibiting melanin production by inhibiting tyrosinase. In addition to this effect, arbutin has been proven to be an active anticancer agent against various types of cancer, such as bladder, breast, brain, cervical, colon, stomach, and other types of cancer [347]. The proapoptotic effect of arbutin and its acetyl derivative was studied in B16 melanoma cells. After cell cycle arrest in the G1 phase, apoptosis occurred, which was associated with the induction of mitochondrial dysfunction. Mitochondrial damage induced by arbutin and its acetyl derivative led to the loss of MMP and changes in the expression of Bcl-2 proteins, where Bcl-2 and Bcl-xL were downregulated [348]. Its antiproliferative effects were also confirmed by proteomic analysis in A375 cells (human malignant melanoma cells). Upregulated proteins included p53, VDAC-1, and 14-3-3G, while others were downregulated, such as vimentin, HSP90, and others [349].
Capsaicin (8-methyl-N-vanillyl-6-nonenamide), the major active component of chili peppers is one of the most commonly used spices in the world. It is responsible for the pungent and spicy flavor of chili peppers. It is known to activate the transient receptor potential vanilloid 1 (TRPV1) receptor, which is involved in the perception of pain and heat, so when capsaicin comes into contact with the skin or mucous membranes, it can cause a burning or stinging sensation [350]. Apart from its sensory effects, capsaicin has been studied for its potential health benefits. It has shown anti-inflammatory properties and may have analgesic (pain-relieving) effects. Some studies suggest that capsaicin may also have antioxidant and anticancer properties [351]. Regarding skin cancers, some studies have suggested that capsaicin may have anti-cancer effects in melanoma. Capsaicin inhibited cell growth and induced apoptosis in human melanoma A375 cells. These effects have been associated with increased generation of nitric oxide (NO•) leading to p53 activation and subsequent activation of both mitochondrial as well as extrinsic apoptotic pathways [352]. Other authors reported that capsaicin in highly metastatic B16-F10 mouse melanoma cells suppressed migration via inhibition of the PI3-K/Akt/Rac1 signal pathway. A detailed study showed that capsaicin significantly decreased p85 phosphorylation, a regulatory subunit of PI3-K [353]. Another study showed that capsaicin in this type of melanoma cells induced apoptosis due to modulation of Bcl-2 family proteins expression, PARP cleavage, and caspase-3 activation [354]. A recent study by Chu et al. (2019) confirmed the significant antiproliferative effect of capsaicin on melanoma cells [355]. Using A375 and C8161 human melanoma cell lines showed the pro-apoptotic effect of this compound associated with PARP cleavage and caspase-3 activation. Additionally, the study revealed that capsaicin also triggered autophagy, as evidenced by the formation of autophagosomes and the accumulation of autophagy markers such as beclin1 and LC3 proteins. However, the inhibition of autophagy by 3-MA significantly enhanced the antiproliferative effect of capsaicin. This suggests that capsaicin-induced autophagy acts as a pro-survival process in these melanoma cells. Furthermore, a recent study revealed that the anti-melanoma effect of capsaicin is also associated with the modulation of the tumor-associated NADH oxidase (tNOX). Capsaicin directly inhibited tNOX activity and reduced tNOX expression on both protein and genetic levels. In addition, capsaicin in melanoma cells induced ROS generation followed by autophagy but not apoptosis induction. Moreover, capsaicin in female C57BL/6 mice bearing B16F10 melanoma cells significantly reduced tumor volume and tumor weight. In addition, protein analysis of the tumor tissue showed that capsaicin decreased tNOX expression also in vivo [356].
Carnosic acid (CrA) is classified as a phenolic diterpene compound and is predominantly present in plants of the Lamiaceae family, which encompasses notable herbs like rosemary, sage, and thyme. Rosemary, in particular, is widely recognized as a prominent natural source of carnosic acid It is known for its biological activities including antioxidants, anti-inflammatory, and neuroprotective. In addition, the anticancer effect of CrA has been intensively studied in both solid cancers and hematological malignities [357]. Regarding skin cancers, CrA has been reported to inhibit the growth, migration, and proliferation of B16F10 melanoma cells. Moreover, it blocked cell cycle arrest at the G0/G1 phase, up-regulated p21 and down-regulated p27. In addition, a combination study showed that CrA potentiated the effect of “classical” chemotherapeutic agents lomustine and carmustine. Finally, CrA decreased tumor growth in mice injected with B16F10 melanoma cells [358]. In another study, carnosic acid inhibited the migration and adhesion of B16F10 cells. Moreover, it inhibited the secretion of MMP-9, urokinase plasminogen activator (uPA), and vascular cell adhesion molecule (VCAM)-1. In addition, CrA was observed to promote the expression of E-cadherin while reducing the expression of vimentin and N-cadherin, which are markers associated with a mesenchymal phenotype. Based on these results, it is suggested that the inhibition of EMT might be a key factor contributing to the reduced cell migration in B16F10 cells treated with CrA [359]. In a recent study by Alcaraz et al. (2022), CrA was identified as a potent radiosensitizing agent [360]. The research demonstrated that when B16F10 melanoma cells were exposed to X-ray irradiation, CrA significantly reduced their cell survival, indicating its potential to enhance the effectiveness of radiotherapy in treating melanoma. Furthermore, another study showed the potential of CrA to enhance the cytotoxicity of β-lapachone in melanoma cells. The mechanism of β-lapachone cytotoxicity is due to the production of ROS via NAD(P)H quinone oxidoreductase 1 (NQO1) activity. Carnosic acid was able to induce the activity of NQO1 and enhanced the cytotoxicity of β-lapachone in melanoma cells [361].
Carnosol, a phenolic diterpene, found mostly in rosemary, gained attention due to its potential health benefits and therapeutic properties. It exhibits antioxidant, anti-inflammatory, and anticancer properties, among others. Regarding anticancer effects, studies have shown that carnosol may exhibit anticancer effects by interfering with various processes involved in cancer development and progression. It has been found to inhibit the growth of cancer cells, induce apoptosis, and suppress the formation of blood vessels that support tumor growth [362,363]. Furthermore, several studies reported also the preventive effect of carnosol against skin cancers. In the study by Huang et al. (2005), carnosol was found to inhibit the invasivity of highly metastatic mouse melanoma B16/F10 cells [364]. Treatments of cells with carnosol resulted in the inhibition of B16/F10 cell migration and invasivity. This effect was associated with decreased activity and expression of MMP-9. In addition, carnosol inhibited the activation of several signaling pathways, including ERK 1/2, AKT, p38, and JNK as well as inhibited NF-kappaB activity. Another author reported that the anti-proliferative and pro-apoptotic effect of carnosol in human melanoma G361 cells is associated with a marked elevation of ROS level. Detailed analyses showed that carnosol modulated the expression of several Bcl-2 family proteins, increased the cellular level of p53, and, on the other hand, prevented activation of Src and STAT3. Subsequently, the expression of STAT3-dependent genes such as D-cyclins and survivin has also been down-regulated [365]. Another study showed that carnosol prevented UVB-induced ROS production in human keratinocyte HaCat cells resulting in a DNA damage decrease as documented by suppressed phosphorylation of γH2AX and Chk1, two markers of DNA damage. Moreover, carnosol prevented UVB-induced cell transformation into cancerous cells [366]. Another study showed that topical application of carnosol decreased the number of chemically induced (TPA/DMBA model) skin cancers in experimental animals. In addition, carnosol inhibited TPA-induced skin inflammation and activity of ornithine decarboxylase [367]. Recently, Yeo et al (2019) reported the anti-inflammatory effect of carnosol in UVB-irradiated animals [368]. Topical application of carnosol inhibited erythema, epidermal thickness, and decreased inflammatory markers, iNOS, and COX-2. Furthermore, carnosol prevented STAT3 activation as well as decreased expression of proinflammatory cytokines including TNF-α and IL-1β.
Carvacrol, a phenolic monoterpenoid that can be isolated from various plants including oregano or thyme, exhibits a diverse array of bioactivities that have proven valuable in clinical applications. These include antimicrobial properties, anti-inflammatory, and antioxidant effects as well as anticancer activity [369,370,371,372]. Regarding melanoma, in carvacrol-treated human melanoma A375 cancer cells significant suppression of cell growth and induction of apoptosis has been reported. Flow cytometric analyzes showed cell cycle arrest at the G2/M phase in carvacrol-treated cells and an increased number of cells with sub-G0 DNA content, which is considered a marker of apoptosis. Western blot analyzes revealed a decrease in Bcl-2 protein expression and PARP cleavage [373]. Another study showed that carvacrol entrapped in chitosan nanoparticles was significantly more potent against A-375 melanoma cells compared to carvacrol only [374]. Recently, Nanni et al. (2020) reported strong anti-melanoma oregano extract [375]. Oregano extract in murine and human melanoma cells induced ROS production and inhibited melanoma cell proliferation. Moreover, it triggered apoptosis and necroptosis via DNA and mitochondria damage. On the other hand, no cytotoxicity of oregano extract was observed in non-cancer cells.
Cinnamaldehyde is an organic compound that gives cinnamon its characteristic flavor and aroma and it is widely used as a flavoring agent in food and beverages due to its sweet, spicy, and warm taste. Apart from its culinary applications, cinnamaldehyde has gained attention for its potential health benefits. Studies have suggested that it may have antioxidant, anti-inflammatory, and anti-cancer properties [376,377,378].
Regarding melanoma, cinnamaldehyde has been reported to impair melanoma cell viability, proliferation, invasiveness, as well as tumor growth. In the study by Cabello et al. (2009), the exposition of human melanoma A375, G361, and LOX cells to cinnamaldehyde led to decreased melanoma cell proliferation associated with cell cycle arrest at the G1 phase [379]. Further analyses showed that cinnamaldehyde induced apoptosis as confirmed by annexin-V/propidium iodide staining and activation of caspase-3. At sub-apoptotic concentrations, cinnamaldehyde inhibited cell migration. Furthermore, the application of cinnamaldehyde on A375 human melanoma cells resulted in an increase in the generation of ROS, along with the activation of genes involved in the cellular response to oxidative stress, including heme oxygenase-1, sulfiredoxin 1 homolog, and thioredoxin reductase 1. Moreover, daily application of cinnamaldehyde to mice bearing human A375 melanoma xenograft led to a decrease in tumor volume and weight when compared to untreated controls. Another study showed that the application of cinnamaldehyde to mice injected with melanoma cells led to significant suppression of tumor growth and decreased density of new blood vessels in the tumor. Western blot analyses showed downregulation of HIF-α and VEGF expression indicating that cinnamaldehyde-induced inhibition of angiogenesis can contribute to its anti-melanoma effect [380]. A study conducted by Patra et al. (2019) uncovered that the antiangiogenic effect of cinnamaldehyde is linked to the PI3/Akt/mTOR pathway, which is involved in the regulation of the transcription and translation of HIF-1α [381]. Moreover, at the molecular level, cinnamaldehyde decreased VEGF secretion, phosphorylation of VEGF receptor, and activity of MMP-2 and MMP-9. In vivo experiments showed that cinnamaldehyde suppressed invasion and metastasis in mice bearing B16F10 melanoma cells xenograft.
Curcumin, a diarylheptanoid, belonging to the group of curcuminoids, is the primary bioactive compound found in turmeric, giving it its characteristic bright yellow-orange color. Vogel and Pelletier originally extracted curcumin from the rhizomes of C. longa in 1815 [200]. Curcumin's impact on melanoma cell survival and proliferation was examined in a number of in vitro experiments that documented its pleiotropic mechanism of action. Zhang et al. (2015) studied the antiproliferative effect of curcumin using A375 human melanoma cells [382]. They found that curcumin decreased melanoma cell proliferation, migration, and invasion. Moreover, western blot analyzes showed decreased expression of MMP-2 and MMP-9. Curcumin also decreased phosphorylation of JAK and STAT3 proteins followed by induction of apoptosis. Another study showed that the pro-apoptotic effect of curcumin is associated with the suppression of the transcriptional activity of NF-кB. In murine melanoma cells exposed to curcumin, Marín et al. (2007) documented inhibition of melanoma cell proliferation, viability, and apoptosis induction [383]. Further study showed inhibition of NF-кB activity and decreased expression of NF-кB-targeted genes including COX-2 and cyclin D1. The association between apoptosis induction and modulation of NF-кB activity in curcumin-treated melanoma cells has also been referred also by Siwak et al. (2005) and Zheng et al. (2004) [384,385]. Furthermore, in the study conducted by Jiang et al. (2014), curcumin has been reported to initiate the mitochondrial pathway of apoptosis in human melanoma cell lines [386]. They found that curcumin stimulated the expression of pro-apoptotic Bax protein and vice versa inhibited the expression of antiapoptotic Mcl-1 and Bcl-2 proteins. Apoptosis has also been associated with decreased levels of NF-κB p65 protein resulting in inhibition of NF-кB signaling, modulation of p38 and p53 activity as well as with DNA damage. Interestingly, curcumin also activated caspase-8 and caspase-3 indicating, that also extrinsic apoptosis pathway may be activated in curcumin-treated melanoma cells. The same results i.e. activation of caspase-8 and caspase-3 were reported by Bush et al. (2001) who also found that curcumin caused aggregation of Fas receptor and these effects were p53 and Bcl-2 proteins independent. In another study, curcumin-induced apoptosis has been associated with curcumin-induced oxidative stress in human melanoma A357 cells [387]. Curcumin increased the generation of ROS and, on the other hand, decreased intracellular GSH levels resulting in apoptosis initiation [388]. ROS-induced cell death in curcumin-treated mouse melanoma cells has also been reported in the study conducted by Kocyigit and Guler (2017) [389]. Curcumin in B16-F10 melanoma cells increased ROS levels, caused DNA damage, and decreased mitochondrial membrane potential (MMP) with subsequent apoptosis induction. In addition, numerous studies reported different mechanisms of curcumin-induced growth or survival suppression such as induction of ER stress [390], autophagy induction [391], or modulation of signaling pathways [205,382,392,393].
Additionally, several studies also confirmed an anticancer effect of curcumin in vivo. In mice bearing B16F10 melanoma cells xenograft, orally administered curcumin significantly decreased the number of lung metastasis and also prolonged survival of experimental animals [337]. The ability of curcumin to prevent lung metastasis in mice injected with B16F10 melanoma cells was confirmed recently. Pulmonary administration (for better bioavailability) of curcumin-protected melanoma lung metastasis in a dose-dependent manner. Histological analysis showed a decrease in cell proliferation marker Ki-67 in curcumin-treated animals [394]. Furthermore, curcumin has been reported to inhibit skin tumors also in mice bearing SCC cell line (SRB12-p9) xenograft. Curcumin applied topically or by oral gavage, significantly reduced tumor growth and volume. Further analyses revealed that this effect was associated with the inhibition of AKT, pS6, 4EBP1, STAT3, and ERK1/2 phosphorylation [395]. In addition, curcumin also prevented skin cancer formation in UVB-irradiated animals. Both, oral and topical applications of curcumin to UVB-irradiated SKH-1 hairless mice led to slower tumor onset and decreased tumor multiplicity when compared to untreated animals [396]. The protective effect of curcumin on UVB-induced skin tumorigenesis has also been reported by Tsai et al. (2012) [397]. Topically applied curcumin significantly decreased the number of tumors and tumor volume. Furthermore, curcumin significantly suppressed UVB-induced apoptosis and thymine dimers, a marker of UVB-induced DNA damage.
Ellagic acid (EA), is a naturally occurring polyphenolic compound found in various plant-based foods, particularly in fruits such as strawberries, raspberries, blackberries, pomegranates, and walnuts. It belongs to the class of hydrolyzable tannins and has been the subject of scientific research due to its potential health benefits. Some studies have investigated the effects of EA on melanoma. Human metastatic melanoma cell lines 1205Lu, WM852c, and A375 were incubated with different concentrations of EA for 72 h [398]. According to the study, EA was found to induce apoptosis and cause cell cycle arrest at the G1 phase, followed by apoptosis induction. Additionally, the study revealed that EA's antiproliferative effects may be attributed to its ability to inhibit the NF-κB pathway. Other authors investigated the anti-cancer effects of EA on melanoma cells in vitro and in vivo [399]. The findings from their study demonstrated that EA notably suppressed the proliferation, migration, and invasion of WM115 and A375 melanoma cells, and these effects were achieved by targeting the EGFR signaling pathway. As the authors reported, incubation of WM115 and A375 melanoma cells with EA led to a reduction in EGFR expression. Furthermore, there are several reports documenting the protective effect of EA against UVB-induced damage to human keratinocytes. Hseu et al. (2012) reported that EA significantly decrease UVB-induced ROS generation and prolonged HaCaT viability with concomitant inhibition of UVB-induced apoptosis [400]. The preventive effect of EA correlated with increased expression of heme oxygenase 1 (HO-1) and SOD. This was followed by the downregulation of Keap1 and increased activation of Nrf2. Another author found that EA modulated the activity of cytokines in UVB-irradiated HaCaT cells. It decreased the expression of proinflammatory IL-6 and up-regulated expression of antiinflammatory IL-10. However, other cytokines such as IL-6, IL-8, MCP-1 (monocyte chemoattractant protein), and TNF-α did not show significant changes in response to EA treatment [401].
Eugenol (4-allyl-2-methoxyphenol), a phenolic compound highly abundant in various spices such as cinnamon, basil, bay leaves, cloves, and nutmeg, demonstrates antioxidant, antibacterial, antiviral, anti-inflammatory, and anti-proliferative properties through diverse mechanisms [402,403,404]. Mechanisms of antiproliferative and apoptosis-inducing mechanisms have been reviewed in detail by Jaganthan and Supriyanto (2012) [405]. Regarding melanoma cells, eugenol has been reported to induce apoptosis in human melanoma G361 cells. Apoptosis was associated with the activation of caspase-3, PARP-cleavage, and DNA fragmentation [406]. Another study showed that eugenol inhibited different melanoma cells in a time-dependent manner. Moreover, it blocked the cell cycle at the G1 phase and induced apoptosis associated with blebbing of membranes, chromatin condensation, and cytoplasm shrinking. In addition, it was also found that eugenol modulated the expression of the members of the E2F family of transcription factors and it is suggested that via this effect eugenol can inhibit cell proliferation and cell cycle and induce apoptosis. Furthermore, eugenol significantly decreased tumor size and decreased the development of metastasis in B16F10 xenograft mice [407]. The ability of eugenol to cause cell cycle arrest at the G1 phase has also been reported by Ra Choi et al. (2011) [408]. G1 arrest in G361 melanoma cells exposed to eugenol has been associated with a decrease in the expression of cycle-related molecules including cyclins A, D3, and E, cyclin-dependent kinases cdk2, cdk4, and cdc2. On the other hand, in the study conducted by Júnior et al (2016) eugenol-induced apoptosis in SK-Mel-28 and A2058 melanoma was accompanied by cell cycle arrest at the G2/m phase indicating that the effect of eugenol can be cell type-dependent [409]. Moreover, the study conducted by Valizahed et al. (2021) indicated that the nanoformulation of eugenol exhibit improved chemotherapeutic effectiveness [410]. Chitosan nanoparticles loaded with eugenol demonstrated a significant effect against human melanoma cell lines (A-375) compared to eugenol in its non-formulated state.
In addition to melanomas, eugenol has also been reported to restrict experimentally induced skin cancers. In the study by Pal et al. (2010), skin cancers have been induced by the application of DMBA as an initiator and croton oil as a promoter [411]. In the untreated control animals, the tumor incidence was 100% while in eugenol-treated cells the number of mice with skin tumors was significantly reduced (42%). Furthermore, the size of tumors in the eugenol group was significantly smaller and the overall survival in eugenol-treated animals was significantly increased. In addition, eugenol significantly decreased cellular proliferation in skin tumors and this effect was associated with the suppression of expression of two oncogenes (c-Myc and H-ras) associated with cell proliferation. A similar effect of eugenol in the prevention of DMBA/croton oil-induced skin carcinogenesis reported also Sukumaran and co-workers (1994) [412]. A strong preventive effect of eugenol has also been observed in chemically induced skin cancer where 12-O-tetradecanoylphorbol-13-acetate (TPA) has been used as a tumor promoter instead of croton oil. Eugenol applicated before TPA significantly decreased cell proliferation, ornithine decarboxylase (ODC) activity, expression of COX-2 and iNOS as well as secretion of proinflammatory cytokines including IL-6, TNF-α, and PGE2. Moreover, in eugenol-treated animals suppression of NF-κB/p65 translocation has been detected [413].
In addition, an in vitro study in non-melanoma A431 cells (human epidermoid carcinoma) showed that eugenol induced block of the cell cycle at the G0/G1 phase. Furthermore, it also induced the translocation of the aryl hydrocarbon receptor (AhR) to the nucleus, and this effect can be associated with the modulation of cell cycle regulatory proteins involved in the transition from the G0/G1 phase to the S phase [414].
6-Gingerol, together with other gingerols, is responsible for the pungent taste. Gingerols are the predominant compounds in fresh ginger rhizomes. Ginger contains several gingerols with different chain lengths, ranging from n6 to n10, with 6-gingerol being the most abundant among them [415]. 6-gingerol has been reported to possess several pharmacological activities such as antioxidant, anti-inflammation, anti-aggregatory or antimicrobial [416]. In addition, it has been studied for its potential anticancer properties. Studies conducted in cell cultures and animal models have shown that gingerol can inhibit the growth and proliferation of cancer cells. It has been found to induce apoptosis and suppress angiogenesis. Gingerol has also demonstrated the ability to modulate various signaling pathways involved in cancer development and progression [417,418,419]. Furthermore, 6-gingerol has also been studied as a potential anticancer agent against skin cancers. In the study by Nigam et al (2009), 6-gingerol demonstrated significant cytotoxic effects against non-melanoma human epidermoid carcinoma A431 cells [420]. This effect was associated with the generation of ROS followed by a decrease in MMP and apoptosis induction. The findings also indicated that the perturbation in the mitochondrial membrane was associated with the disruption of the Bax/Bcl-2 ratio, both at the gene and protein levels. Another study showed that topically applied 6-gingerol inhibited COX-2 expression in mouse skin stimulated by the TPA. Furthermore, 6-gingerol suppressed the activity of NF-кB. In addition, it prevented TPA-induced activation of the p38 MAP kinase that regulates COX-2 expression in mouse skin [421]. Later they reported that 6-gingerol in vitro reduced UVB-induced ROS production and caspase activation in human keratinocyte HaCaT cells. Moreover, it inhibited UVB-induced activation of NF-кB. In addition, local application of 6-gingerol prior to UVB irradiation of hairless mice significantly suppressed UVB-induced COX-2 expression and NF-кB activation in mouse skin indicating that 6-gingerol can be effective against UVB-induced skin damage [422]. In the study by Ju et al. (2011) 6-gingerol decreased tumor incidence, growth, and prolonged survival of mice inoculated with B16F1 melanoma cells [423]. Additionally, treatment with 6-gingerol resulted in a significant influx of tumor-infiltrating lymphocytes such as CD4 and CD8 T-cells, as well as B220+ B-cells, into the tumor microenvironment and this effect can play a role in the anticancer effect of 6-gingerol. Another study demonstrated that 6-gingerol exhibits antiangiogenic effects. In vitro experiments revealed that 6-gingerol inhibited the proliferation of HUVECs stimulated by VEGF and bFGF. It also suppressed the formation of capillary-like tubes induced by VEGF in different experimental models. Furthermore, in an in vivo study using mice implanted with B16F10 melanoma cells, treatment with 6-gingerol led to a reduction in the number of lung metastases. These findings suggest that 6-gingerol may have therapeutic potential in targeting angiogenesis and metastasis in cancer [419]. In a study by Park et al. (1998), topical application of 6-gingerol prevented TPA-induced inflammation in experimental animals and decreased the number of skin papillomas in the DMBA/TPA model of chemical skin carcinogenesis [424]. Additionally, it inhibited TPA-induced increased activity of epidermal ornithine decarboxylase, which plays an important role in skin carcinogenesis.
6-shogaol is another bioactive compound found in ginger. It is derived from gingerol during the drying or cooking process of ginger. It possesses unique properties and potential health benefits including anti-inflammatory, antioxidant, anticancer, and neuroprotective effects [129,425]. In the context of skin cancers, a study reported that 6-shogaol exhibited greater efficacy than 6-gingerol or curcumin in inhibiting tumor promotion induced by TPA in experimental animals. Topically applied to mouse skin, 6-shogaol significantly suppressed TPA-stimulated transcription of iNOS and COX-2 mRNA expression. Additionally, it was observed that 6-shogaol reduced the nuclear translocation of NF-кB, a protein involved in inflammatory responses and cell survival. Moreover, 6-shogaol demonstrated significant inhibition of signaling pathways associated with cell proliferation, survival, and inflammation including Erk1/2, p38 MAPK, PI3/Akt, and JNK1/2. Furthermore, in a TPA/DMBA skin tumor model, 6-shogaol significantly reduced the occurrence of multiple skin tumors [426]. In a study conducted by Chen et al. (2019), it was found that 6-shogaol exhibited a protective effect against UVB-induced oxidative stress in HaCaT cells [427]. When HaCaT cells were exposed to UVB radiation without any treatment, it resulted in elevated levels of ROS and increased activation of p38, ERK, and JNK signaling. However, when HaCaT cells were treated with 6-shogaol prior to UVB irradiation, it significantly reduced ROS levels and prevented the activation of p38, ERK, and JNK. This suggests that 6-shogaol can mitigate the oxidative stress induced by UVB radiation by inhibiting the activation of the MAPK signaling pathway. Additionally, the study revealed that 6-shogaol prevented the depletion of the Nrf2 protein levels induced by UVB irradiation. Nrf2 plays a crucial role in cellular defense against oxidative stress. Therefore, the protective effect of 6-shogaol against oxidative stress can be partially attributed to its ability to prevent the depletion of Nrf2 protein levels.
Thymol, a phenolic monoterpene, is a major component of essential oils derived from thyme. It shares the same chemical formula as carvacrol, another compound found in thyme and oregano, but they differ in the position of the hydroxyl group on the benzene ring. While carvacrol has the hydroxyl group in the ortho position, thymol has it in the para position. Both thymol and carvacrol are aromatic compounds present in various plants belonging to the Lamiaceae and Verbenaceae families [428]. Thymol exhibits a wide range of biological activities, such as antioxidative, antimicrobial, antiviral, antihypertensive, immunomodulatory, and anticancer properties [429]. However, the research on the impact of thymol specifically on skin cancers is limited, with only a few studies conducted in this area. Thymol in B16-F10 melanoma cells exhibited moderate cytotoxicity and this effect was associated with induction of oxidative stress. In addition, radical scavengers such as GSH, vitamin E, butylated hydroxyanisole, or butylated hydroxytoluene suppressed thymol-induced cytotoxicity in melanoma cells [430]. Another study showed that dry extract from T. vulgaris L. and thymol, its main active compound, prevented UVA- and UVB-induced damage of keratinocytes cell line NCTC 2544. Both treatments decreased UV-induced ROS production as well as DNA damage [431]. Later these authors documented the protective effect of T. vulgaris extract and thymol in UVA- and UVB-irradiated HaCaT (keratinocytes) cell line. Both compounds prevented UVA- and UVB-induced genotoxicity as well as the generation of ROS and apoptosis induction [432].
Another author reported that thymol decreased UVB-induced genotoxicity in ex vivo human skin models. Moreover, both compounds prevented UVB-induced cell toxicity as documented by decreased levels of lactate dehydrogenase (LDH) [433].
The chemical structures of other selected phenolic compounds discussed in this review are shown in Figure 4.
6. Conclusion
Throughout history, spices have been utilized for both culinary and medicinal purposes. Their ability to enhance the taste, smell, and appearance of food and beverages is well-known, but they also offer protection against various acute and chronic diseases. Recent research has highlighted the health properties of spices, particularly their bioactive constituents, with a focus on phenolic compounds.
Phenolic compounds have gained attention for their potential role in preventing and managing cancer. While they may not be as potent as conventional chemotherapeutic drugs, their capacity for cancer prevention is evident. In the case of skin cancer, utilizing phenolic compounds is particularly appealing due to their widespread availability, cost-effectiveness, and good tolerance. The skin's accessibility makes it an advantageous area for both observation and topical treatment, encouraging research into the effectiveness of these natural compounds.
Studies have demonstrated that phenolic compounds exhibit anticancer effects through various mechanisms, such as antioxidant, proapoptotic, anti-inflammatory, anti-metastatic, and antiangiogenic actions, and their modulation of signaling pathways related to cell life and death. Additionally, they may counteract the damaging effects of solar UV radiation and environmental carcinogens.
However, it is crucial to acknowledge that much of the current evidence comes from laboratory and animal studies. To establish phenolic compounds' true effectiveness and safety in preventing or treating skin cancers, further clinical trials involving human subjects are essential. By conducting more research, we can better understand the potential benefits and limitations of using these natural compounds in the fight against skin cancer.
Author Contributions
Conceptualization, J.B., and J.M.; investigation, J.B, R.M., Z.B., G.M., Z.F., and J.M.; writing—original draft preparation J.B, R.M., Z.B., G.M., Z.F., and J.M.; writing—review and editing, J.M.; visualization, R.M.; supervision, J.M.; funding acquisition, G.M. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the Grant Agency of the Ministry of the Education, Science, Research and Sport of the Slovak Republic (VEGA 1/0539/21, VEGA 1/0513/21) and the Slovak Research and Development Agency under contract No. APVV-16–0446. Moreover, this publication is the result of the project implementation: “Open scientific community for modern interdisciplinary research in medicine (OPENMED)”, ITMS2014+: 313011V455, supported by the Operational Programme Integrated Infrastructure, funded by the ERDF. This publication was also supported by the Operational program Integrated Infra-structure within the project: Demand-driven research for the sustainable and innovative food, Drive4SIFood 313011V336, co-financed by the European Regional Development Fund.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shabir Ahmad, R., Imran, M., Kamran Khan, M., Haseeb Ahmad, M., Sajid Arshad, M., Ateeq, H.,Abdul Rahim, M., Introductory Chapter: Herbs and Spices - An Overview. In Herbs and Spices - New Processing Technologies; R. Shabir Ahmad; IntechOpen Limited: London, UK, 2021; Vol. 1, p. 276. [CrossRef]
- Bi, X., Lim, J.,Henry, C.J. Spices in the management of diabetes mellitus. Food Chem 2017, 217: p. 281-293. [CrossRef]
- Tapsell, L.C., Hemphill, I., Cobiac, L., Patch, C.S., Sullivan, D.R., Fenech, M., Roodenrys, S., Keogh, J.B., Clifton, P.M., Williams, P.G., Fazio, V.A.,Inge, K.E. Health benefits of herbs and spices: the past, the present, the future. Med J Aust 2006, 185(S4): p. S1-S24. [CrossRef]
- Essa, M.M., Akbar, M.,Guillemin, G., In The Benefits of Natural Products for Neurodegenerative Diseases; M.M. Essa, M. Akbar, and G. Guillemin; Springer Cham: Switzerland, 2016; Vol. 12, p. 496. [CrossRef]
- Singh, N.,Yadav, S.S. A review on health benefits of phenolics derived from dietary spices. Curr Res Food Sci 2022, 5: p. 1508-1523. [CrossRef]
- Avila-Galvez, M.A., Gimenez-Bastida, J.A., Espin, J.C.,Gonzalez-Sarrias, A. Dietary Phenolics against Breast Cancer. A Critical Evidence-Based Review and Future Perspectives. Int J Mol Sci 2020, 21(16). [CrossRef]
- Muller, A.G., Sarker, S.D., Saleem, I.Y.,Hutcheon, G.A. Delivery of natural phenolic compounds for the potential treatment of lung cancer. Daru 2019, 27(1): p. 433-449. [CrossRef]
- De, S., Paul, S., Manna, A., Majumder, C., Pal, K., Casarcia, N., Mondal, A., Banerjee, S., Nelson, V.K., Ghosh, S., Hazra, J., Bhattacharjee, A., Mandal, S.C., Pal, M.,Bishayee, A. Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies. Cancers (Basel) 2023, 15(3). [CrossRef]
- Kountouri, A.M., Kaliora, A.C., Koumbi, L.,Andrikopoulos, N.K. In-vitro gastric cancer prevention by a polyphenol-rich extract from olives through induction of apoptosis. Eur J Cancer Prev 2009, 18(1): p. 33-9. [CrossRef]
- Spagnuolo, C., Flores, G., Russo, G.L.,Ruiz Del Castillo, M.L. A Phenolic Extract Obtained from Methyl Jasmonate-Treated Strawberries Enhances Apoptosis in a Human Cervical Cancer Cell Line. Nutr Cancer 2016, 68(7): p. 1140-50. [CrossRef]
- Ji, X., Usman, A., Razalli, N.H., Sambanthamurthi, R.,Gupta, S.V. Oil palm phenolics (OPP) inhibit pancreatic cancer cell proliferation via suppression of NF-kappaB pathway. Anticancer Res 2015, 35(1): p. 97-106.
- Leiter, U., Keim, U.,Garbe, C. Epidemiology of Skin Cancer: Update 2019. Adv Exp Med Biol 2020, 1268: p. 123-139. [CrossRef]
- O'Neill, C.H.,Scoggins, C.R. Melanoma. J Surg Oncol 2019, 120(5): p. 873-881. [CrossRef]
- Garbe, C., Keim, U., Gandini, S., Amaral, T., Katalinic, A., Hollezcek, B., Martus, P., Flatz, L., Leiter, U.,Whiteman, D. Epidemiology of cutaneous melanoma and keratinocyte cancer in white populations 1943-2036. Eur J Cancer 2021, 152: p. 18-25. [CrossRef]
- Krynitz, B., Olsson, H., Lundh Rozell, B., Lindelof, B., Edgren, G.,Smedby, K.E. Risk of basal cell carcinoma in Swedish organ transplant recipients: a population-based study. Br J Dermatol 2016, 174(1): p. 95-103. [CrossRef]
- Peris, K., Fargnoli, M.C., Garbe, C., Kaufmann, R., Bastholt, L., Seguin, N.B., Bataille, V., Marmol, V.D., Dummer, R., Harwood, C.A., Hauschild, A., Holler, C., Haedersdal, M., Malvehy, J., Middleton, M.R., Morton, C.A., Nagore, E., Stratigos, A.J., Szeimies, R.M., Tagliaferri, L., Trakatelli, M., Zalaudek, I., Eggermont, A., Grob, J.J., European Dermatology Forum, t.E.A.o.D.-O., the European Organization for, R.,Treatment of, C. Diagnosis and treatment of basal cell carcinoma: European consensus-based interdisciplinary guidelines. Eur J Cancer 2019, 118: p. 10-34. [CrossRef]
- Tanese, K. Diagnosis and Management of Basal Cell Carcinoma. Curr Treat Options Oncol 2019, 20(2): p. 13. [CrossRef]
- Roewert-Huber, J., Stockfleth, E.,Kerl, H. Pathology and pathobiology of actinic (solar) keratosis - an update. Br J Dermatol 2007, 157 Suppl 2: p. 18-20. [CrossRef]
- Eisen, D.B., Asgari, M.M., Bennett, D.D., Connolly, S.M., Dellavalle, R.P., Freeman, E.E., Goldenberg, G., Leffell, D.J., Peschin, S., Sligh, J.E., Wu, P.A., Frazer-Green, L., Malik, S.,Schlesinger, T.E. Guidelines of care for the management of actinic keratosis. J Am Acad Dermatol 2021, 85(4): p. e209-e233. [CrossRef]
- Kallini, J.R., Hamed, N.,Khachemoune, A. Squamous cell carcinoma of the skin: epidemiology, classification, management, and novel trends. Int J Dermatol 2015, 54(2): p. 130-40. [CrossRef]
- Mogensen, M.,Jemec, G.B. Diagnosis of nonmelanoma skin cancer/keratinocyte carcinoma: a review of diagnostic accuracy of nonmelanoma skin cancer diagnostic tests and technologies. Dermatol Surg 2007, 33(10): p. 1158-74. [CrossRef]
- Waldman, A.,Schmults, C. Cutaneous Squamous Cell Carcinoma. Hematol Oncol Clin North Am 2019, 33(1): p. 1-12. [CrossRef]
- Neale, H., Michelon, M., Jacob, S., Pinkston, M., Ukaegbu, R., Zamor, W., Morrison, E., Deng, A.,Levin, N.A. Topical 5% 5-fluorouracil versus procedural modalities for squamous cell carcinoma in situ and superficial basal cell carcinoma: A retrospective cohort analysis. J Am Acad Dermatol 2022, 87(2): p. 423-425. [CrossRef]
- Dirschka, T., Bierhoff, E., Pflugfelder, A.,Garbe, C. Topical 3.0% diclofenac in 2.5% hyaluronic acid gel induces regression of cancerous transformation in actinic keratoses. J Eur Acad Dermatol Venereol 2010, 24(3): p. 258-63. [CrossRef]
- Piquero-Casals, J., Morgado-Carrasco, D., Gilaberte, Y., Del Rio, R., Macaya-Pascual, A., Granger, C.,Lopez-Estebaranz, J.L. Management Pearls on the Treatment of Actinic Keratoses and Field Cancerization. Dermatol Ther (Heidelb) 2020, 10(5): p. 903-915. [CrossRef]
- Garbe, C., Amaral, T., Peris, K., Hauschild, A., Arenberger, P., Basset-Seguin, N., Bastholt, L., Bataille, V., Del Marmol, V., Dreno, B., Fargnoli, M.C., Forsea, A.M., Grob, J.J., Holler, C., Kaufmann, R., Kelleners-Smeets, N., Lallas, A., Lebbe, C., Lytvynenko, B., Malvehy, J., Moreno-Ramirez, D., Nathan, P., Pellacani, G., Saiag, P., Stratigos, A.J., Van Akkooi, A.C.J., Vieira, R., Zalaudek, I., Lorigan, P., European Dermatology Forum, t.E.A.o.D.-O., the European Organization for, R.,Treatment of, C. European consensus-based interdisciplinary guideline for melanoma. Part 1: Diagnostics: Update 2022. Eur J Cancer 2022, 170: p. 236-255. [CrossRef]
- Erdei, E.,Torres, S.M. A new understanding in the epidemiology of melanoma. Expert Rev Anticancer Ther 2010, 10(11): p. 1811-23. [CrossRef]
- Rastrelli, M., Tropea, S., Rossi, C.R.,Alaibac, M. Melanoma: epidemiology, risk factors, pathogenesis, diagnosis and classification. In Vivo 2014, 28(6): p. 1005-11.
- Leonardi, G.C., Falzone, L., Salemi, R., Zanghi, A., Spandidos, D.A., McCubrey, J.A., Candido, S.,Libra, M. Cutaneous melanoma: From pathogenesis to therapy (Review). Int J Oncol 2018, 52(4): p. 1071-1080. [CrossRef]
- Rashid, S., Shaughnessy, M.,Tsao, H. Melanoma classification and management in the era of molecular medicine. Dermatol Clin 2023, 41(1): p. 49-63. [CrossRef]
- Gong, H.Z., Zheng, H.Y.,Li, J. Amelanotic melanoma. Melanoma Res 2019, 29(3): p. 221-230. [CrossRef]
- Garbe, C., Peris, K., Hauschild, A., Saiag, P., Middleton, M., Bastholt, L., Grob, J.J., Malvehy, J., Newton-Bishop, J., Stratigos, A.J., Pehamberger, H.,Eggermont, A.M. Diagnosis and treatment of melanoma. European consensus-based interdisciplinary guideline - Update 2016. Eur J Cancer 2016, 63: p. 201-17. [CrossRef]
- Dummer, R., Hauschild, A., Lindenblatt, N., Pentheroudakis, G.,Keilholz, U. Cutaneous melanoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol 2015, 26 Suppl 5: p. v126-32. [CrossRef]
- Swetter, S.M., Thompson, J.A., Albertini, M.R., Barker, C.A., Baumgartner, J., Boland, G., Chmielowski, B., DiMaio, D., Durham, A., Fields, R.C., Fleming, M.D., Galan, A., Gastman, B., Grossmann, K., Guild, S., Holder, A., Johnson, D., Joseph, R.W., Karakousis, G., Kendra, K., Lange, J.R., Lanning, R., Margolin, K., Olszanski, A.J., Ott, P.A., Ross, M.I., Salama, A.K., Sharma, R., Skitzki, J., Sosman, J., Wuthrick, E., McMillian, N.R.,Engh, A.M. NCCN Guidelines(R) Insights: Melanoma: Cutaneous, Version 2.2021. J Natl Compr Canc Netw 2021, 19(4): p. 364-376. [CrossRef]
- Alu'datt, M.H., Rababah, T., Alhamad, M.N., Al-Mahasneh, M.A., Almajwal, A., Gammoh, S., Ereifej, K., Johargy, A.,Alli, I. A review of phenolic compounds in oil-bearing plants: Distribution, identification and occurrence of phenolic compounds. Food Chem 2017, 218: p. 99-106. [CrossRef]
- Robbins, R.J. Phenolic acids in foods: an overview of analytical methodology. J Agric Food Chem 2003, 51(10): p. 2866-87. [CrossRef]
- Zhang, Y., Cai, P., Cheng, G.,Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Natural Product Communications 2022, 17(1): p. 1934578X211069721. [CrossRef]
- Rodriguez De Luna, S.L., Ramirez-Garza, R.E.,Serna Saldivar, S.O. Environmentally Friendly Methods for Flavonoid Extraction from Plant Material: Impact of Their Operating Conditions on Yield and Antioxidant Properties. ScientificWorldJournal 2020, 2020: p. 6792069. [CrossRef]
- Dias, M.C., Pinto, D.,Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26(17). [CrossRef]
- Serrano, J., Puupponen-Pimia, R., Dauer, A., Aura, A.M.,Saura-Calixto, F. Tannins: current knowledge of food sources, intake, bioavailability and biological effects. Mol Nutr Food Res 2009, 53 Suppl 2: p. S310-29. [CrossRef]
- Pecyna, P., Wargula, J., Murias, M.,Kucinska, M. More Than Resveratrol: New Insights into Stilbene-Based Compounds. Biomolecules 2020, 10(8). [CrossRef]
- Stringlis, I.A., de Jonge, R.,Pieterse, C.M.J. The Age of Coumarins in Plant-Microbe Interactions. Plant Cell Physiol 2019, 60(7): p. 1405-1419. [CrossRef]
- Saleem, M., Kim, H.J., Ali, M.S.,Lee, Y.S. An update on bioactive plant lignans. Nat Prod Rep 2005, 22(6): p. 696-716. [CrossRef]
- Ginwala, R., Bhavsar, R., Chigbu, D.I., Jain, P.,Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants (Basel) 2019, 8(2). [CrossRef]
- Song, B., Guan, S., Lu, J., Chen, Z., Huang, G., Li, G., Xiong, Y., Zhang, S., Yue, Z.,Deng, X. Suppressive effects of fisetin on mice T lymphocytes in vitro and in vivo. J Surg Res 2013, 185(1): p. 399-409. [CrossRef]
- Kamisah, Y., Jalil, J., Yunos, N.M.,Zainalabidin, S. Cardioprotective Properties of Kaempferol: A Review. Plants (Basel) 2023, 12(11). [CrossRef]
- Shen, N., Wang, T., Gan, Q., Liu, S., Wang, L.,Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem 2022, 383: p. 132531. [CrossRef]
- Dourado, N.S., Souza, C.D.S., de Almeida, M.M.A., Bispo da Silva, A., Dos Santos, B.L., Silva, V.D.A., De Assis, A.M., da Silva, J.S., Souza, D.O., Costa, M.F.D., Butt, A.M.,Costa, S.L. Neuroimmunomodulatory and Neuroprotective Effects of the Flavonoid Apigenin in in vitro Models of Neuroinflammation Associated With Alzheimer's Disease. Front Aging Neurosci 2020, 12: p. 119. [CrossRef]
- Górniak, I., Bartoszewski, R.,Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem Rev 2019, 18(1): p. 241-272. [CrossRef]
- Fernandez, J., Silvan, B., Entrialgo-Cadierno, R., Villar, C.J., Capasso, R., Uranga, J.A., Lombo, F.,Abalo, R. Antiproliferative and palliative activity of flavonoids in colorectal cancer. Biomed Pharmacother 2021, 143: p. 112241. [CrossRef]
- Abdullah, H., Ismail, I., Suppian, R.,Zakaria, N.M. Natural Gallic Acid and Methyl Gallate Induces Apoptosis in Hela Cells through Regulation of Intrinsic and Extrinsic Protein Expression. Int J Mol Sci 2023, 24(10). [CrossRef]
- Zhang, L.C., Liu, Y.N., La, X.Q., Li, S.T., Wen, L.N., Liu, T., Li, H.Q., Li, A.P., Wu, H., Wu, C.X.,Li, Z.Y. The bound polyphenols of foxtail millet (Setaria italica) inner shell inhibit breast cancer by promoting lipid accumulation-induced autophagic death. Food Chem Toxicol 2023, 177: p. 113855. [CrossRef]
- Lin, W.,Tongyi, S. Role of Bax/Bcl-2 family members in green tea polyphenol induced necroptosis of p53-deficient Hep3B cells. Tumour Biol 2014, 35(8): p. 8065-75. [CrossRef]
- Michalkova, R., Kello, M., Kudlickova, Z., Gazdova, M., Mirossay, L., Mojzisova, G.,Mojzis, J. Programmed Cell Death Alterations Mediated by Synthetic Indole Chalcone Resulted in Cell Cycle Arrest, DNA Damage, Apoptosis and Signaling Pathway Modulations in Breast Cancer Model. Pharmaceutics 2022, 14(3). [CrossRef]
- Barreca, M.M., Alessandro, R.,Corrado, C. Effects of Flavonoids on Cancer, Cardiovascular and Neurodegenerative Diseases: Role of NF-kappaB Signaling Pathway. Int J Mol Sci 2023, 24(11). [CrossRef]
- Russo, M., Moccia, S., Luongo, D.,Russo, G.L. Senolytic Flavonoids Enhance Type-I and Type-II Cell Death in Human Radioresistant Colon Cancer Cells through AMPK/MAPK Pathway. Cancers (Basel) 2023, 15(9). [CrossRef]
- Mirossay, L., Varinska, L.,Mojzis, J. Antiangiogenic Effect of Flavonoids and Chalcones: An Update. Int J Mol Sci 2017, 19(1). [CrossRef]
- Varinska, L., van Wijhe, M., Belleri, M., Mitola, S., Perjesi, P., Presta, M., Koolwijk, P., Ivanova, L.,Mojzis, J. Anti-angiogenic activity of the flavonoid precursor 4-hydroxychalcone. Eur J Pharmacol 2012, 691(1-3): p. 125-33. [CrossRef]
- Khater, M., Greco, F.,Osborn, H.M.I. Antiangiogenic Activity of Flavonoids: A Systematic Review and Meta-Analysis. Molecules 2020, 25(20). [CrossRef]
- Zhang, L.,Lokeshwar, B.L. Medicinal properties of the Jamaican pepper plant Pimenta dioica and Allspice. Curr Drug Targets 2012, 13(14): p. 1900-6. [CrossRef]
- Al-Rehaily, A., Al-Said, M., Al-Yahya, M., Jaber, S.,Rafatullah, S. Ethnopharmacological Studies on Allspice (Pimenta dioica) in Laboratory Animals. Pharm Biol 2002, 40: p. 200-205. [CrossRef]
- Miyajima, Y., Kikuzaki, H., Hisamoto, M.,Nikatani, N. Antioxidative polyphenols from berries of Pimenta dioica. Biofactors 2004, 21(1-4): p. 301-3. [CrossRef]
- El Gizawy, H.A., Boshra, S.A., Mostafa, A., Mahmoud, S.H., Ismail, M.I., Alsfouk, A.A., Taher, A.T.,Al-Karmalawy, A.A. Pimenta dioica (L.) Merr. Bioactive Constituents Exert Anti-SARS-CoV-2 and Anti-Inflammatory Activities: Molecular Docking and Dynamics, In Vitro, and In Vivo Studies. Molecules 2021, 26(19). [CrossRef]
- Marzouk, M.S., Moharram, F.A., Mohamed, M.A., Gamal-Eldeen, A.M.,Aboutabl, E.A. Anticancer and antioxidant tannins from Pimenta dioica leaves. Z Naturforsch C J Biosci 2007, 62(7-8): p. 526-36. [CrossRef]
- Milenkovic, A., Stanojević, J., Stojanović-Radić, Z., Pejčić, M., Cvetkovic, D., Zvezdanović, J.,Stanojević, L. Chemical composition, antioxidative and antimicrobial activity of allspice (Pimenta dioica (L.) Merr.) essential oil and extract. Advanced Technologies 2020, 9: p. 27-36. [CrossRef]
- Rao, P.S., Navinchandra, S.,Jayaveera, K.N. An important spice, Pimenta dioica (Linn.) Merill: A Review. Int Curr Pharm J 2012, 1(8): p. 221-225. [CrossRef]
- Rema, J.,Krishnamoorthy, B., Allspice. 2nd Edition ed. Handbook of Herbs and Spices. 2012: Woodhead Publishing. 166-192.
- Mandal, D., Sarkar, T.,Chakraborty, R. Critical Review on Nutritional, Bioactive, and Medicinal Potential of Spices and Herbs and Their Application in Food Fortification and Nanotechnology. Appl Biochem Biotechnol 2023, 195(2): p. 1319-1513. [CrossRef]
- Chudiwal, A.K., Jain, D.P.,Rahul Somani, R. Alpinia galanga Willd.- An overview on phyto-pharmacological properties. Indian J Nat Prod Resour 2010, 1(2): p. 143-149.
- Eram, S., Mujahid, M., Bagga, P., Ansari, V., Ahmad, M., Kumar, A., Ahsan, F.,Akhter, M. A Review on Phytopharmacological Activity of Alpinia Galanga. Int J Pharm Pharm Sci 2019: p. 6-11. [CrossRef]
- Chouni, A.,Paul, S. A Review on Phytochemical and Pharmacological Potential of Alpinia galanga. Pharmacogn J 2018, 10(1): p. 09-15. [CrossRef]
- Namdeo, A.G.,Kale, V.M. Comparative pharmacognostic and phytochemical investigation of two Alpinia species from Zingiberaceae Family. World J Pharm Res 2015, 4(5): p. 1417-32.
- Tungmunnithum, D., Tanaka, N., Uehara, A.,Iwashina, T. Flavonoids Profile, Taxonomic Data, History of Cosmetic Uses, Anti-Oxidant and Anti-Aging Potential of Alpinia galanga (L.) Willd. Cosmetics 2020, 7(4): p. 89. [CrossRef]
- Ramanunny, A.K., Wadhwa, S., Gulati, M., Vishwas, S., Khursheed, R., Paudel, K.R., Gupta, S., Porwal, O., Alshahrani, S.M., Jha, N.K., Chellappan, D.K., Prasher, P., Gupta, G., Adams, J., Dua, K., Tewari, D.,Singh, S.K. Journey of Alpinia galanga from kitchen spice to nutraceutical to folk medicine to nanomedicine. J Ethnopharmacol 2022, 291: p. 115144. [CrossRef]
- Zhou, Y.-Q., Liu, H., He, M.-X., Wang, R., Zeng, Q.-Q., Wang, Y., Ye, W.-C.,Zhang, Q.-W., Chapter 11 - A Review of the Botany, Phytochemical, and Pharmacological Properties of Galangal. In Natural and Artificial Flavoring Agents and Food Dyes; A.M. Grumezescu and A.M. Holban; Academic Press, 2018, p. 351-396. [CrossRef]
- Nampoothiri, S.V., Esakkidurai, T.,Pitchumani, K. Identification and Quantification of Phenolic Compounds in Alpinia galanga and Alpinia calcarata and its Relation to Free Radical Quenching Properties: A Comparative Study. J Herbs Spices Med Plants 2015, 21(2): p. 140-147. [CrossRef]
- Suja, S.,Chinnaswamy, P. Inhibition of in vitro cytotoxic effect evoked by Alpinia galanga and Alpinia officinarum on PC - 3 cell line. Anc Sci Life 2008, 27(4): p. 33-40.
- Choi, J.Y., Lee, N.K., Wang, Y.Y., Hong, J.P., Son, S.R., Gu, D.H., Jang, D.S.,Choi, J.H. 1'-Acetoxyeugenol Acetate Isolated from Thai Ginger Induces Apoptosis in Human Ovarian Cancer Cells by ROS Production via NADPH Oxidase. Antioxidants (Basel) 2022, 11(2). [CrossRef]
- Suciati, A.,Maryati. Systematic Review: Anticancer Potential of Active Compounds from Galangal (Alpinia galanga). In Proceedings of the 4th International Conference Current Breakthrough in Pharmacy (ICB-Pharma 2022), Sukoharjo, Indonesia, 14-15 January 2022. [CrossRef]
- Ali, B.H.,Blunden, G. Pharmacological and toxicological properties of Nigella sativa. Phytother Res 2003, 17(4): p. 299-305. [CrossRef]
- Kooti, W., Hasanzadeh-Noohi, Z., Sharafi-Ahvazi, N., Asadi-Samani, M.,Ashtary-Larky, D. Phytochemistry, pharmacology, and therapeutic uses of black seed (Nigella sativa). Chin J Nat Med 2016, 14(10): p. 732-745. [CrossRef]
- Ismail, N., Abd Ghafar, S.A.,Abu Bakar, M.Z., Chapter Fifteen - Antioxidant activity and phenolic content of black cumin seeds. In Biochemistry, Nutrition, and Therapeutics of Black Cumin Seed; A.A. Mariod; Academic Press, 2023, p. 169-188. [CrossRef]
- Srinivasan, K. Cumin (Cuminum cyminum) and black cumin (Nigella sativa) seeds: traditional uses, chemical constituents, and nutraceutical effects. Food Qual Saf 2018, 2(1): p. 1-16. [CrossRef]
- Milenkovic, A.,Stanojević, L. Black pepper: Chemical composition and biological activities. Advanced Technologies 2021, 10: p. 40-50. [CrossRef]
- Al-Khayri, J.M., Upadhya, V., Pai, S.R., Naik, P.M., Al-Mssallem, M.Q.,Alessa, F.M. Comparative Quantification of the Phenolic Compounds, Piperine Content, and Total Polyphenols along with the Antioxidant Activities in the Piper trichostachyon and P. nigrum. Molecules 2022, 27(18). [CrossRef]
- Ashokkumar, K., Murugan, M., Dhanya, M.K., Pandian, A.,Warkentin, T.D. Phytochemistry and therapeutic potential of black pepper [Piper nigrum (L.)] essential oil and piperine: a review. Clin Phytoscience 2021, 7(1): p. 52. [CrossRef]
- Butt, M.S., Pasha, I., Sultan, M.T., Randhawa, M.A., Saeed, F.,Ahmed, W. Black pepper and health claims: a comprehensive treatise. Crit Rev Food Sci Nutr 2013, 53(9): p. 875-86. [CrossRef]
- Zhang, M., Qiu, B., Sun, M., Wang, Y., Wei, M., Gong, Y.,Yan, M. Preparation of Black pepper (Piper nigrum L.) essential oil nanoparticles and its antitumor activity on triple negative breast cancer in vitro. J Food Biochem 2022, 46(12): p. e14406. [CrossRef]
- Cardoso, L.P., de Sousa, S.O., Gusson-Zanetoni, J.P., de Melo Moreira Silva, L.L., Frigieri, B.M., Henrique, T., Tajara, E.H., Oliani, S.M.,Rodrigues-Lisoni, F.C. Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway. Pharmaceuticals (Basel) 2023, 16(1). [CrossRef]
- Manayi, A., Nabavi, S.M., Setzer, W.N.,Jafari, S. Piperine as a Potential Anti-cancer Agent: A Review on Preclinical Studies. Curr Med Chem 2018, 25(37): p. 4918-4928. [CrossRef]
- Rao, P.V.,Gan, S.H. Cinnamon: a multifaceted medicinal plant. Evid Based Complement Alternat Med 2014, 2014: p. 642942. [CrossRef]
- Sadeghi, S., Davoodvandi, A., Pourhanifeh, M.H., Sharifi, N., ArefNezhad, R., Sahebnasagh, R., Moghadam, S.A., Sahebkar, A.,Mirzaei, H. Anti-cancer effects of cinnamon: Insights into its apoptosis effects. Eur J Med Chem 2019, 178: p. 131-140. [CrossRef]
- Sahib, A.S. Anti-diabetic and antioxidant effect of cinnamon in poorly controlled type-2 diabetic Iraqi patients: A randomized, placebo-controlled clinical trial. J Intercult Ethnopharmacol 2016, 5(2): p. 108-13. [CrossRef]
- Schink, A., Naumoska, K., Kitanovski, Z., Kampf, C.J., Frohlich-Nowoisky, J., Thines, E., Poschl, U., Schuppan, D.,Lucas, K. Anti-inflammatory effects of cinnamon extract and identification of active compounds influencing the TLR2 and TLR4 signaling pathways. Food Funct 2018, 9(11): p. 5950-5964. [CrossRef]
- Raeisi, M., Tajik, H., Yarahmadi, A.,Sanginabadi, S. Antimicrobial Effect of Cinnamon Essential Oil Against Escherichia Coli and Staphylococcus aureus. Health Scope 2015, 4(4): p. e21808. [CrossRef]
- Pagliari, S., Forcella, M., Lonati, E., Sacco, G., Romaniello, F., Rovellini, P., Fusi, P., Palestini, P., Campone, L., Labra, M., Bulbarelli, A.,Bruni, I. Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum verum J. Presl) Bark Extract after In Vitro Digestion Simulation. Foods 2023, 12(3): p. 452. [CrossRef]
- Shahidi, F.,Hossain, A. Bioactives in spices, and spice oleoresins: Phytochemicals and their beneficial effects in food preservation and health promotion. J Food Bioact 2018, 3: p. 8-75. [CrossRef]
- Cardoso-Ugarte, G.A., López-Malo, A.,Sosa-Morales, M.E., Chapter 38 - Cinnamon (Cinnamomum zeylanicum) Essential Oils. In Essential Oils in Food Preservation, Flavor and Safety; V.R. Preedy; Academic Press: San Diego, 2016, p. 339-347. [CrossRef]
- Vallverdu-Queralt, A., Regueiro, J., Martinez-Huelamo, M., Rinaldi Alvarenga, J.F., Leal, L.N.,Lamuela-Raventos, R.M. A comprehensive study on the phenolic profile of widely used culinary herbs and spices: rosemary, thyme, oregano, cinnamon, cumin and bay. Food Chem 2014, 154: p. 299-307. [CrossRef]
- Singh, N., Rao, A.S., Nandal, A., Kumar, S., Yadav, S.S., Ganaie, S.A.,Narasimhan, B. Phytochemical and pharmacological review of Cinnamomum verum J. Presl-a versatile spice used in food and nutrition. Food Chem 2021, 338: p. 127773. [CrossRef]
- Baliga, M.S., Mane, P.P., Timothy Nallemgera, J., Thilakchand, K.R.,Kalekhan, F., Chapter 5 - Dietary Spices in the Prevention of Rheumatoid Arthritis: Past, Present, and Future. In Foods and Dietary Supplements in the Prevention and Treatment of Disease in Older Adults; R.R. Watson; Academic Press: San Diego, 2015, p. 41-49. [CrossRef]
- Kumar, V., Marković, T., Mila, E.,Dey, A., Herbs: Composition and Dietary Importance. In Encyclopedia of Food and Health; B. Caballero, P.M. Finglas, and F. Toldrá; Elsevier: Kidlington, Oxford, 2015; Vol. 3, p. 332-337. [CrossRef]
- Laribi, B., Kouki, K., M'Hamdi, M.,Bettaieb, T. Coriander (Coriandrum sativum L.) and its bioactive constituents. Fitoterapia 2015, 103: p. 9-26. [CrossRef]
- Msaada, K., Jemia, M.B., Salem, N., Bachrouch, O., Sriti, J., Tammar, S., Bettaieb, I., Jabri, I., Kefi, S., Limam, F.,Marzouk, B. Antioxidant activity of methanolic extracts from three coriander (Coriandrum sativum L.) fruit varieties. Arab J Chem 2017, 10: p. S3176-S3183. [CrossRef]
- Barros, L., Dueñas, M., Dias, M.I., Sousa, M.J., Santos-Buelga, C.,Ferreira, I.C.F.R. Phenolic profiles of in vivo and in vitro grown Coriandrum sativum L. Food Chem 2012, 132(2): p. 841-848. [CrossRef]
- Oganesyan, E.T., Nersesyan, Z.M.,Parkhomenko, A.Y. Chemical composition of the above-ground part of Coriandrum sativum. Pharm Chem J 2007, 41(3): p. 149-153. [CrossRef]
- Nambiar, V., Daniel, M.,Guin, P. Characterization of polyphenols from coriander leaves (Coriandrum sativum), red amaranthus (A paniculatus) and green amaranthus (A frumentaceus) using paper chromatography and their health implications. J Herb Med Toxicol 2010, 4: p. 173-177.
- Mechchate, H., Costa de Oliveira, R., Es-Safi, I., Vasconcelos Mourao, E.M., Bouhrim, M., Kyrylchuk, A., Soares Pontes, G., Bousta, D.,Grafov, A. Antileukemic Activity and Molecular Docking Study of a Polyphenolic Extract from Coriander Seeds. Pharmaceuticals (Basel) 2021, 14(8). [CrossRef]
- Elmas, L., Secme, M., Mammadov, R., Fahrioglu, U.,Dodurga, Y. The determination of the potential anticancer effects of Coriandrum sativum in PC-3 and LNCaP prostate cancer cell lines. J Cell Biochem 2019, 120(3): p. 3506-3513. [CrossRef]
- Eroglu, C., Secme, M., Bagci, G.,Dodurga, Y. Assessment of the anticancer mechanism of ferulic acid via cell cycle and apoptotic pathways in human prostate cancer cell lines. Tumour Biol 2015, 36(12): p. 9437-46. [CrossRef]
- Kim, S.J., Pham, T.H., Bak, Y., Ryu, H.W., Oh, S.R.,Yoon, D.Y. Orientin inhibits invasion by suppressing MMP-9 and IL-8 expression via the PKCalpha/ ERK/AP-1/STAT3-mediated signaling pathways in TPA-treated MCF-7 breast cancer cells. Phytomedicine 2018, 50: p. 35-42. [CrossRef]
- Huang, H., Nakamura, T., Yasuzawa, T.,Ueshima, S. Effects of Coriandrum sativum on Migration and Invasion Abilities of Cancer Cells. J Nutr Sci Vitaminol (Tokyo) 2020, 66(5): p. 468-477. [CrossRef]
- Wani, S.A.,Kumar, P. Fenugreek: A review on its nutraceutical properties and utilization in various food products. J Saudi Soc Agric Sci 2018, 17(2): p. 97-106. [CrossRef]
- Sun, W., Shahrajabian, M.H.,Cheng, Q. Fenugreek Cultivation with Emphasis on Historical Aspects and its uses in Traditional Medicine and Modern Pharmaceutical Science. Mini Rev Med Chem 2021, 21(6): p. 724-730. [CrossRef]
- Visuvanathan, T., Than, L.T.L., Stanslas, J., Chew, S.Y.,Vellasamy, S. Revisiting Trigonella foenum-graecum L.: Pharmacology and Therapeutic Potentialities. Plants (Basel) 2022, 11(11). [CrossRef]
- Bahmani, M., Shirzad, H., Mirhosseini, M., Mesripour, A.,Rafieian-Kopaei, M. A Review on Ethnobotanical and Therapeutic Uses of Fenugreek (Trigonella foenum-graceum L). J Evid Based Complementary Altern Med 2016, 21(1): p. 53-62. [CrossRef]
- Nagulapalli Venkata, K.C., Swaroop, A., Bagchi, D.,Bishayee, A. A small plant with big benefits: Fenugreek (Trigonella foenum-graecum Linn.) for disease prevention and health promotion. Mol Nutr Food Res 2017, 61(6). [CrossRef]
- Skaltsa, H., Fenugreek - The Genus Trigonella. 1st Edition ed. 2002, London: CRC Press. 226. [CrossRef]
- Benayad, Z., Gomez-Cordoves, C.,Es-Safi, N.E. Characterization of flavonoid glycosides from fenugreek (Trigonella foenum-graecum) crude seeds by HPLC-DAD-ESI/MS analysis. Int J Mol Sci 2014, 15(11): p. 20668-85. [CrossRef]
- Niknam, R., Kiani, H., Mousavi, Z.,Mousavi, M., Extraction, Detection, and Characterization of Various Chemical Components of Trigonella foenum-graecum L. (Fenugreek) Known as a Valuable Seed in Agriculture. 2021, p. 189-217. [CrossRef]
- Shawky, E., Sobhy, A.A., Ghareeb, D.A., Shams Eldin, S.M.,Selim, D.A. Comparative metabolomics analysis of bioactive constituents of the leaves of different Trigonella species: Correlation study to α-amylase and α-glycosidase inhibitory effects. Ind Crops Prod 2022, 182: p. 114947. [CrossRef]
- Al-Timimi, L.A.N. Antibacterial and Anticancer Activities of Fenugreek Seed Extract. Asian Pac J Cancer Prev 2019, 20(12): p. 3771-3776. [CrossRef]
- Alsemari, A., Alkhodairy, F., Aldakan, A., Al-Mohanna, M., Bahoush, E., Shinwari, Z.,Alaiya, A. The selective cytotoxic anti-cancer properties and proteomic analysis of Trigonella Foenum-Graecum. BMC Complement Altern Med 2014, 14: p. 114. [CrossRef]
- Ajmal, S., Shafqat, M., Ajmal, L., Younas, H., Tasadduq, R.,Mahamood, N. Evaluation of Anti-cancer and Anti-proliferative Activity of Medicinal Plant Extracts (Saffron, Green Tea, Clove, Fenugreek) on Toll Like Receptors Pathway. Nat Prod Sci 2022, 28(3): p. 121-129. [CrossRef]
- Alrumaihi, F.A., Khan, M.A., Allemailem, K.S., Alsahli, M.A., Almatroudi, A., Younus, H., Alsuhaibani, S.A., Algahtani, M.,Khan, A. Methanolic Fenugreek Seed Extract Induces p53-Dependent Mitotic Catastrophe in Breast Cancer Cells, Leading to Apoptosis. J Inflamm Res 2021, 14: p. 1511-1535. [CrossRef]
- Singletary, K. Ginger: An Overview of Health Benefits. Nutr Today 2010, 45: p. 171-183. [CrossRef]
- Khodaie, L.,Sadeghpoor, O. Ginger from ancient times to the new outlook. Jundishapur J Nat Pharm Prod 2015, 10(1): p. e18402. [CrossRef]
- Kiyama, R. Nutritional implications of ginger: chemistry, biological activities and signaling pathways. J Nutr Biochem 2020, 86: p. 108486. [CrossRef]
- Semwal, R.B., Semwal, D.K., Combrinck, S.,Viljoen, A.M. Gingerols and shogaols: Important nutraceutical principles from ginger. Phytochemistry 2015, 117: p. 554-568. [CrossRef]
- Schadich, E., Hlavac, J., Volna, T., Varanasi, L., Hajduch, M.,Dzubak, P. Effects of Ginger Phenylpropanoids and Quercetin on Nrf2-ARE Pathway in Human BJ Fibroblasts and HaCaT Keratinocytes. Biomed Res Int 2016, 2016: p. 2173275. [CrossRef]
- Ozkur, M., Benlier, N., Takan, I., Vasileiou, C., Georgakilas, A.G., Pavlopoulou, A., Cetin, Z.,Saygili, E.I. Ginger for Healthy Ageing: A Systematic Review on Current Evidence of Its Antioxidant, Anti-Inflammatory, and Anticancer Properties. Oxid Med Cell Longev 2022, 2022: p. 4748447. [CrossRef]
- Kundu, J.K., Na, H.K.,Surh, Y.J. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr 2009, 61: p. 182-192. [CrossRef]
- Bae, W.Y., Choi, J.S., Kim, J.E., Park, C.,Jeong, J.W. Zingerone suppresses angiogenesis via inhibition of matrix metalloproteinases during tumor development. Oncotarget 2016, 7(30): p. 47232-47241. [CrossRef]
- Gird, C.E., Costea, T.,Mitran, V. Evaluation of cytotoxic activity and anticancer potential of indigenous Rosemary (Rosmarinus officinalis L.) and Oregano (Origanum vulgare L.) dry extracts on MG-63 bone osteosarcoma human cell line. Rom J Morphol Embryol 2021, 62(2): p. 525-535. [CrossRef]
- Bouyahya, A., Chamkhi, I., Benali, T., Guaouguaou, F.E., Balahbib, A., El Omari, N., Taha, D., Belmehdi, O., Ghokhan, Z.,El Menyiy, N. Traditional use, phytochemistry, toxicology, and pharmacology of Origanum majorana L. J Ethnopharmacol 2021, 265: p. 113318. [CrossRef]
- Ozkan, G., Baydar, H.,Erbas, S. The influence of harvest time on essential oil composition, phenolic constituents and antioxidant properties of Turkish oregano (Origanum onites L.). J Sci Food Agric 2010, 90(2): p. 205-9. [CrossRef]
- Teixeira, B., Marques, A., Ramos, C., Serrano, C., Matos, O., Neng, N.R., Nogueira, J.M., Saraiva, J.A.,Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare) extracts and essential oil. J Sci Food Agric 2013, 93(11): p. 2707-14. [CrossRef]
- Geetha, P.P.S.R.P.G.L.P.G.S.S.R.S.B.S. Pharmacology and chemistry of Myristica fragrans Houtt. - a review. J Spices Aromat 2005, 14(2): p. 94-101.
- Kuete, V., Chapter 23 - Myristica fragrans: A Review. In Medicinal Spices and Vegetables from Africa; V. Kuete; Academic Press, 2017, p. 497-512. [CrossRef]
- Zhang, W.K., Tao, S.S., Li, T.T., Li, Y.S., Li, X.J., Tang, H.B., Cong, R.H., Ma, F.L.,Wan, C.J. Nutmeg oil alleviates chronic inflammatory pain through inhibition of COX-2 expression and substance P release in vivo. Food Nutr Res 2016, 60: p. 30849. [CrossRef]
- Hayfaa, A.A., Sahar, A.M.,Awatif, M.A. Evaluation of analgesic activity and toxicity of alkaloids in Myristica fragrans seeds in mice. J Pain Res 2013, 6: p. 611-5. [CrossRef]
- Shafiei, Z., Shuhairi, N.N., Md Fazly Shah Yap, N., Harry Sibungkil, C.A.,Latip, J. Antibacterial Activity of Myristica fragrans against Oral Pathogens. Evid Based Complement Alternat Med 2012, 2012: p. 825362. [CrossRef]
- Narasimhan, B.,Dhake, A.S. Antibacterial principles from Myristica fragrans seeds. J Med Food 2006, 9(3): p. 395-9. [CrossRef]
- Morita, T., Jinno, K., Kawagishi, H., Arimoto, Y., Suganuma, H., Inakuma, T.,Sugiyama, K. Hepatoprotective effect of myristicin from nutmeg (Myristica fragrans) on lipopolysaccharide/d-galactosamine-induced liver injury. J Agric Food Chem 2003, 51(6): p. 1560-5. [CrossRef]
- Rezende, D., Oliveira, C.D., Batista, L.R., Ferreira, V.R.F., Brandao, R.M., Caetano, A.R.S., Alves, M.V.P.,Cardoso, M.G. Bactericidal and antioxidant effects of essential oils from Satureja montana L., Myristica fragrans H. and Cymbopogon flexuosus. Lett Appl Microbiol 2022, 74(5): p. 741-751. [CrossRef]
- Bandyopadhyay, P.S.V.C. Estimation of phenolic acids in cinnamon, clove, cardamom, nutmeg and mace by high performance liquid chromatography. J Spices Aromat 1995, 4(2): p. 129-134.
- Odubanjo, O., Olasehinde, T., Oyeleye, I., Oboh, G.,Boligon, A. Seed extracts from Myristica fragrans (Nutmeg) and Moringa oleifera (Drumstick tree) inhibits enzymes relevant to erectile dysfunction and metal-induced oxidative damage in rats' penile tissues. J Food Biochem 2017, 42: p. e12452. [CrossRef]
- Kraft, K.H., Brown, C.H., Nabhan, G.P., Luedeling, E., Luna Ruiz Jde, J., Coppens d'Eeckenbrugge, G., Hijmans, R.J.,Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc Natl Acad Sci U S A 2014, 111(17): p. 6165-70. [CrossRef]
- Reyes-Escogido Mde, L., Gonzalez-Mondragon, E.G.,Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16(2): p. 1253-70. [CrossRef]
- Pugliese, A., O'Callaghan, Y., Tundis, R., Galvin, K., Menichini, F., O'Brien, N.,Loizzo, M.R. In vitro investigation of the bioaccessibility of carotenoids from raw, frozen and boiled red chili peppers (Capsicum annuum). Eur J Nutr 2014, 53(2): p. 501-10. [CrossRef]
- Antonio, A.S., Wiedemann, L.S.M.,Veiga Junior, V.F. The genus Capsicum: a phytochemical review of bioactive secondary metabolites. Rsc Adv 2018, 8(45): p. 25767-25784. [CrossRef]
- Zhang, D., Sun, X., Battino, M., Wei, X., Shi, J., Zhao, L., Liu, S., Xiao, J., Shi, B.,Zou, X. A comparative overview on chili pepper (capsicum genus) and sichuan pepper (zanthoxylum genus): From pungent spices to pharma-foods. Trends Food Sci 2021, 117: p. 148-162. [CrossRef]
- Ayob, O., Hussain, P.R., Suradkar, P., Naqash, F., Rather, S.A., Joshi, S.,Ahmad Azad, Z.R.A. Evaluation of chemical composition and antioxidant activity of Himalayan Red chilli varieties. LWT 2021, 146: p. 111413. [CrossRef]
- Batiha, G.E., Alqahtani, A., Ojo, O.A., Shaheen, H.M., Wasef, L., Elzeiny, M., Ismail, M., Shalaby, M., Murata, T., Zaragoza-Bastida, A., Rivero-Perez, N., Magdy Beshbishy, A., Kasozi, K.I., Jeandet, P.,Hetta, H.F. Biological Properties, Bioactive Constituents, and Pharmacokinetics of Some Capsicum spp. and Capsaicinoids. Int J Mol Sci 2020, 21(15). [CrossRef]
- Clark, R.,Lee, S.H. Anticancer Properties of Capsaicin Against Human Cancer. Anticancer Res 2016, 36(3): p. 837-43.
- Heinrich, M., Kufer, J., Leonti, M.,Pardo-de-Santayana, M. Ethnobotany and ethnopharmacology--interdisciplinary links with the historical sciences. J Ethnopharmacol 2006, 107(2): p. 157-60. [CrossRef]
- de Oliveira, J.R., Camargo, S.E.A.,de Oliveira, L.D. Rosmarinus officinalis L. (rosemary) as therapeutic and prophylactic agent. J Biomed Sci 2019, 26(1): p. 5. [CrossRef]
- Hussain, A.I., Anwar, F., Chatha, S.A., Jabbar, A., Mahboob, S.,Nigam, P.S. Rosmarinus officinalis essential oil: antiproliferative, antioxidant and antibacterial activities. Braz J Microbiol 2010, 41(4): p. 1070-8. [CrossRef]
- Hcini, K., Abidi, M., Quílez, M., Jordán Maria, J.,Sadok, B., Total Phenolic Content and Polyphenolic Profile of Tunisian Rosemary (<em>Rosmarinus officinalis</em> L.) Residues. In Natural Drugs from Plants; A.E.-S. Hany; IntechOpen: Rijeka, 2021, p. Ch. 9. [CrossRef]
- Moore, J., Yousef, M.,Tsiani, E. Anticancer Effects of Rosemary (Rosmarinus officinalis L.) Extract and Rosemary Extract Polyphenols. Nutrients 2016, 8(11). [CrossRef]
- Gonzalez-Vallinas, M., Reglero, G.,Ramirez de Molina, A. Rosemary (Rosmarinus officinalis L.) Extract as a Potential Complementary Agent in Anticancer Therapy. Nutr Cancer 2015, 67(8): p. 1221-9. [CrossRef]
- Saxena, R.B. Botany, Taxonomy and Cytology of Crocus sativus series. Ayu 2010, 31(3): p. 374-81. [CrossRef]
- Melnyk, J.P., Wang, S.,Marcone, M.F. Chemical and biological properties of the world's most expensive spice: Saffron. Food Res Int 2010, 43(8): p. 1981-1989. [CrossRef]
- Xing, B., Li, S., Yang, J., Lin, D., Feng, Y., Lu, J.,Shao, Q. Phytochemistry, pharmacology, and potential clinical applications of saffron: A review. J Ethnopharmacol 2021, 281: p. 114555. [CrossRef]
- Groppo, M.,Pirani, J.R. A new species of Zanthoxylum (Rutaceae) with a key to the species from Northeastern Brazil. Phytotaxa 2017, 314: p. 259. [CrossRef]
- Okagu, I.U., Ndefo, J.C., Aham, E.C.,Udenigwe, C.C. Zanthoxylum Species: A Comprehensive Review of Traditional Uses, Phytochemistry, Pharmacological and Nutraceutical Applications. Molecules 2021, 26(13). [CrossRef]
- Ekka, G., Jadhav, S.,Quraishi, A., An Overview of Genus Zanthoxylum with Special Reference to Its Herbal Significance and Application. 2020, p. [CrossRef]
- Tanoh, E.A., Boue, G.B., Nea, F., Genva, M., Wognin, E.L., Ledoux, A., Martin, H., Tonzibo, Z.F., Frederich, M.,Fauconnier, M.L. Seasonal Effect on the Chemical Composition, Insecticidal Properties and Other Biological Activities of Zanthoxylum leprieurii Guill. & Perr. Essential oils. Foods 2020, 9(5). [CrossRef]
- Ji, Y., Li, S.,Ho, C.-T. Chemical composition, sensory properties and application of Sichuan pepper (Zanthoxylum genus). Food Sci Hum Wellness 2019, 8(2): p. 115-125. [CrossRef]
- Wang, G.W., Hu, W.T., Huang, B.K.,Qin, L.P. Illicium verum: a review on its botany, traditional use, chemistry and pharmacology. J Ethnopharmacol 2011, 136(1): p. 10-20. [CrossRef]
- Sharafan, M., Jafernik, K., Ekiert, H., Kubica, P., Kocjan, R., Blicharska, E.,Szopa, A. Illicium verum (Star Anise) and Trans-Anethole as Valuable Raw Materials for Medicinal and Cosmetic Applications. Molecules 2022, 27(3). [CrossRef]
- Ghosh, S., Chisti, Y.,Banerjee, U.C. Production of shikimic acid. Biotechnol Adv 2012, 30(6): p. 1425-31. [CrossRef]
- Li, W., Wu, Z., Xia, Y., Tan, J., Zhao, H., Chen, S., Li, Y., Tang, H., Wang, G.,Zhang, Y. Antiviral and Antioxidant Components from the Fruits of Illicium verum Hook.f. (Chinese Star Anise). J Agric Food Chem 2022, 70(12): p. 3697-3707. [CrossRef]
- De, M., De, A.K., Sen, P.,Banerjee, A.B. Antimicrobial properties of star anise (Illicium verum Hook f). Phytother Res 2002, 16(1): p. 94-5. [CrossRef]
- Majali, I.S. Antioxidant and Anti-Inflammatory Activity of Star Anise (Illicium Verum) in Murine Model. Biomed Pharmacol J 2022, 15(2): p. 1097-1108. [CrossRef]
- Ibrahim, F., Ibrahim, A., E, E.L., Hussein, M.,Ahmed, K. Illicium verum Extracts Anti-Gastro Ulcerogenic Potential on Experimentally Rat Models. Int J PharmTech Res 2016, 9: p. 65-80.
- Patra, J.K., Das, G., Bose, S., Banerjee, S., Vishnuprasad, C.N., Del Pilar Rodriguez-Torres, M.,Shin, H.S. Star anise (Illicium verum): Chemical compounds, antiviral properties, and clinical relevance. Phytother Res 2020, 34(6): p. 1248-1267. [CrossRef]
- Asif, M., Yehya, A.H.S., Al-Mansoub, M.A., Revadigar, V., Ezzat, M.O., Khadeer Ahamed, M.B., Oon, C.E., Murugaiyah, V., Abdul Majid, A.S.,Abdul Majid, A.M.S. Anticancer attributes of Illicium verum essential oils against colon cancer. S Afr J Bot 2016, 103: p. 156-161. [CrossRef]
- Pahore, A.K., Khan, S.,Karim, N. Anticancer effect of Illicium verum (star anise fruit) against human breast cancer MCF-7 cell line. Pak J Med Sci 2023, 39(1): p. 70-74. [CrossRef]
- Chen, C.H.,deGraffenried, L.A. Anethole suppressed cell survival and induced apoptosis in human breast cancer cells independent of estrogen receptor status. Phytomedicine 2012, 19(8-9): p. 763-7. [CrossRef]
- Khan, M.,Shamim, S. Anisi Stellati Fructus, a Significant Traditional Chinese Medicine (TCM) Herb and Its Bioactivity against Gastric Cancer. Evid Based Complement Alternat Med 2022, 2022: p. 4071489. [CrossRef]
- Najar, B., Shortrede, J.E., Pistelli, L.,Buhagiar, J. Chemical Composition and in Vitro Cytotoxic Screening of Sixteen Commercial Essential Oils on Five Cancer Cell Lines. Chem Biodivers 2020, 17(1): p. e1900478. [CrossRef]
- Kim, A., Im, M.,Ma, J.Y. Anisi stellati fructus extract attenuates the in vitro and in vivo metastatic and angiogenic potential of malignant cancer cells by downregulating proteolytic activity and pro-angiogenic factors. Int J Oncol 2014, 45(5): p. 1937-48. [CrossRef]
- Batiha, G.E., Ogunyemi, O.M., Shaheen, H.M., Kutu, F.R., Olaiya, C.O., Sabatier, J.M.,De Waard, M. Rhus coriaria L. (Sumac), a Versatile and Resourceful Food Spice with Cornucopia of Polyphenols. Molecules 2022, 27(16). [CrossRef]
- Alsamri, H., Athamneh, K., Pintus, G., Eid, A.H.,Iratni, R. Pharmacological and Antioxidant Activities of Rhus coriaria L. (Sumac). Antioxidants (Basel) 2021, 10(1).
- Sakhr, K.,El Khatib, S. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac - Rhus coriaria): A review. Heliyon 2020, 6(1): p. e03207. [CrossRef]
- Kubatka, P., Kello, M., Kajo, K., Samec, M., Liskova, A., Jasek, K., Koklesova, L., Kuruc, T., Adamkov, M., Smejkal, K., Svajdlenka, E., Solar, P., Pec, M., Busselberg, D., Sadlonova, V.,Mojzis, J. Rhus coriaria L. (Sumac) Demonstrates Oncostatic Activity in the Therapeutic and Preventive Model of Breast Carcinoma. Int J Mol Sci 2020, 22(1). [CrossRef]
- Athamneh, K., Hasasna, H.E., Samri, H.A., Attoub, S., Arafat, K., Benhalilou, N., Rashedi, A.A., Dhaheri, Y.A., AbuQamar, S., Eid, A.,Iratni, R. Rhus coriaria increases protein ubiquitination, proteasomal degradation and triggers non-canonical Beclin-1-independent autophagy and apoptotic cell death in colon cancer cells. Scientific Reports 2017, 7(1): p. 11633. [CrossRef]
- Gabr, S.A.,Alghadir, H.A. Potential anticancer activities of Rhus coriaria (sumac) extract against human cancer cell lines. Biosci Rep 2021, 41(5). [CrossRef]
- Romeo, F.V., Ballistreri, G., Fabroni, S., Pangallo, S., Nicosia, M.G., Schena, L.,Rapisarda, P. Chemical Characterization of Different Sumac and Pomegranate Extracts Effective against Botrytis cinerea Rots. Molecules 2015, 20(7): p. 11941-58. [CrossRef]
- Bozan, B., Koşar, M., Z.Tunalier, Öztürk, N.,Baser, K.H.C. Antioxidant and Free Radical Scavenging Activities of Rhus coriaria and Cinnamomum cassia Extracts. Acta Alimentaria 2003, 32: p. 53-61. [CrossRef]
- Abu-Reidah, I.M., Ali-Shtayeh, M.S., Jamous, R.M., Arraez-Roman, D.,Segura-Carretero, A. HPLC-DAD-ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem 2015, 166: p. 179-191. [CrossRef]
- Patil, S.M., Ramu, R., Shirahatti, P.S., Shivamallu, C.,Amachawadi, R.G. A systematic review on ethnopharmacology, phytochemistry and pharmacological aspects of Thymus vulgaris Linn. Heliyon 2021, 7(5): p. e07054. [CrossRef]
- Hammoudi Halat, D., Krayem, M., Khaled, S.,Younes, S. A Focused Insight into Thyme: Biological, Chemical, and Therapeutic Properties of an Indigenous Mediterranean Herb. Nutrients 2022, 14(10). [CrossRef]
- Ahmad, A., Saeed, M.,Ansari, I.A. Molecular insights on chemopreventive and anticancer potential of carvacrol: Implications from solid carcinomas. J Food Biochem 2021, 45(12): p. e14010. [CrossRef]
- Niksic, H., Becic, F., Koric, E., Gusic, I., Omeragic, E., Muratovic, S., Miladinovic, B.,Duric, K. Cytotoxicity screening of Thymus vulgaris L. essential oil in brine shrimp nauplii and cancer cell lines. Sci Rep 2021, 11(1): p. 13178. [CrossRef]
- Kowalczyk, A., Przychodna, M., Sopata, S., Bodalska, A.,Fecka, I. Thymol and Thyme Essential Oil-New Insights into Selected Therapeutic Applications. Molecules 2020, 25(18). [CrossRef]
- Roby, M.H.H., Sarhan, M.A., Selim, K.A.-H.,Khalel, K.I. Evaluation of antioxidant activity, total phenols and phenolic compounds in thyme (Thymus vulgaris L.), sage (Salvia officinalis L.), and marjoram (Origanum majorana L.) extracts. Ind Crops Prod 2013, 43: p. 827-831. [CrossRef]
- Teixeira, B., Marques, A., Ramos, C., Neng, N.R., Nogueira, J.M.F., Saraiva, J.A.,Nunes, M.L. Chemical composition and antibacterial and antioxidant properties of commercial essential oils. Ind Crops Prod 2013, 43: p. 587-595. [CrossRef]
- Kotha, R.R.,Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24(16). [CrossRef]
- Meng, F.-C., Zhou, Y.-Q., Ren, D., Wang, R., Wang, C., Lin, L.-G., Zhang, X.-Q., Ye, W.-C.,Zhang, Q.-W., Chapter 10 - Turmeric: A Review of Its Chemical Composition, Quality Control, Bioactivity, and Pharmaceutical Application. In Natural and Artificial Flavoring Agents and Food Dyes; A.M. Grumezescu and A.M. Holban; Academic Press, 2018, p. 299-350. [CrossRef]
- Qiong-Qiong, Y. Phenolic profiles, antioxidant, and antiproliferative activities of turmeric (Curcuma longa). Ind Crops Prod 2020, v. 152: p. 112561. [CrossRef]
- Fan, Y., Zhang, X., Tong, Y., Chen, S.,Liang, J. Curcumin against gastrointestinal cancer: A review of the pharmacological mechanisms underlying its antitumor activity. Front Pharmacol 2022, 13: p. 990475. [CrossRef]
- Shahrajabian, M.H.,Sun, W. The Golden Spice for Life: Turmeric with the Pharmacological Benefits of Curcuminoids Components, including Curcumin, Bisdemethoxycurcumin, and Demethoxycurcumin. Curr Org Synth 2023. [CrossRef]
- Tang, Y.,Cao, Y. Curcumin Inhibits the Growth and Metastasis of Melanoma via miR-222-3p/SOX10/Notch Axis. Dis Markers 2022, 2022: p. 3129781. [CrossRef]
- Rocha Silva, L., Alves Nunes, J., Zhan, P., K, Z.L., Helena Cardoso, S.,Ferreira da Silva-Junior, E. Natural Coumarin Derivatives Targeting Melanoma. Curr Med Chem 2023. [CrossRef]
- Venza, I., Visalli, M., Oteri, R., Beninati, C., Teti, D.,Venza, M. Genistein reduces proliferation of EP3-expressing melanoma cells through inhibition of PGE2-induced IL-8 expression. Int Immunopharmacol 2018, 62: p. 86-95. [CrossRef]
- Danciu, C., Borcan, F., Bojin, F., Zupko, I.,Dehelean, C. Effect of the isoflavone genistein on tumor size, metastasis potential and melanization in a B16 mouse model of murine melanoma. Nat Prod Commun 2013, 8(3): p. 343-6. [CrossRef]
- Yang, G., Fu, Y., Malakhova, M., Kurinov, I., Zhu, F., Yao, K., Li, H., Chen, H., Li, W., Lim, D.Y., Sheng, Y., Bode, A.M., Dong, Z.,Dong, Z. Caffeic acid directly targets ERK1/2 to attenuate solar UV-induced skin carcinogenesis. Cancer Prev Res (Phila) 2014, 7(10): p. 1056-66. [CrossRef]
- Kang, N.J., Lee, K.W., Shin, B.J., Jung, S.K., Hwang, M.K., Bode, A.M., Heo, Y.S., Lee, H.J.,Dong, Z. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis 2009, 30(2): p. 321-30. [CrossRef]
- Balupillai, A., Nagarajan, R.P., Ramasamy, K., Govindasamy, K.,Muthusamy, G. Caffeic acid prevents UVB radiation induced photocarcinogenesis through regulation of PTEN signaling in human dermal fibroblasts and mouse skin. Toxicol Appl Pharmacol 2018, 352: p. 87-96. [CrossRef]
- Pelinson, L.P., Assmann, C.E., Palma, T.V., da Cruz, I.B.M., Pillat, M.M., Mânica, A., Stefanello, N., Weis, G.C.C., de Oliveira Alves, A., de Andrade, C.M., Ulrich, H., Morsch, V.M.M., Schetinger, M.R.C.,Bagatini, M.D. Antiproliferative and apoptotic effects of caffeic acid on SK-Mel-28 human melanoma cancer cells. Mol Biol Rep 2019, 46(2): p. 2085-2092. [CrossRef]
- Kimsa-Dudek, M., Synowiec-Wojtarowicz, A., Krawczyk, A., Kosowska, A., Kimsa-Furdzik, M.,Francuz, T. The Apoptotic Effect of Caffeic or Chlorogenic Acid on the C32 Cells That Have Simultaneously Been Exposed to a Static Magnetic Field. Int J Mol Sci 2022, 23(7). [CrossRef]
- Kimsa-Dudek, M., Synowiec-Wojtarowicz, A.,Krawczyk, A. Phenolic acids and a static magnetic field change the expression of transforming growth factor beta isoforms in amelanotic melanoma cells. Mol Biol Rep 2023, 50(5): p. 4207-4216. [CrossRef]
- Kimsa-Dudek, M., Krawczyk, A., Synowiec-Wojtarowicz, A., Dudek, S.,Pawlowska-Goral, K. The impact of the co-exposure of melanoma cells to chlorogenic acid and a moderate-strength static magnetic field. J Food Biochem 2020, 44(12): p. e13512. [CrossRef]
- Cui, K., Zhang, L., La, X., Wu, H., Yang, R., Li, H.,Li, Z. Ferulic Acid and P-Coumaric Acid Synergistically Attenuate Non-Alcoholic Fatty Liver Disease through HDAC1/PPARG-Mediated Free Fatty Acid Uptake. Int J Mol Sci 2022, 23(23). [CrossRef]
- Ghaderi, S., Gholipour, P., Komaki, A., Salehi, I., Rashidi, K., Esmaeil Khoshnam, S.,Rashno, M. p-Coumaric acid ameliorates cognitive and non-cognitive disturbances in a rat model of Alzheimer's disease: The role of oxidative stress and inflammation. Int Immunopharmacol 2022, 112: p. 109295. [CrossRef]
- Moradi, M., Farbood, Y., Mard, S.A., Dianat, M., Goudarzi, G., Khorsandi, L.,Seyedian, S.S. p-Coumaric acid has pure anti-inflammatory characteristics against hepatopathy caused by ischemia-reperfusion in the liver and dust exposure. Iran J Basic Med Sci 2023, 26(2): p. 164-175. [CrossRef]
- Venkatesan, A., Roy, A., Kulandaivel, S., Natesan, V.,Kim, S.J. p-Coumaric Acid Nanoparticles Ameliorate Diabetic Nephropathy via Regulating mRNA Expression of KIM-1 and GLUT-2 in Streptozotocin-Induced Diabetic Rats. Metabolites 2022, 12(12). [CrossRef]
- Tehami, W., Nani, A., Khan, N.A.,Hichami, A. New Insights Into the Anticancer Effects of p-Coumaric Acid: Focus on Colorectal Cancer. Dose Response 2023, 21(1): p. 15593258221150704. [CrossRef]
- Saremi, S., Kolahi, M., Tabandeh, M.R.,Hashemitabar, M. Induction of apoptosis and suppression of Ras gene expression in MCF human breast cancer cells. J Cancer Res Ther 2022, 18(4): p. 1052-1060. [CrossRef]
- Hu, X., Yang, Z., Liu, W., Pan, Z., Zhang, X., Li, M., Liu, X., Zheng, Q.,Li, D. The Anti-tumor Effects of p-Coumaric Acid on Melanoma A375 and B16 Cells. Front Oncol 2020, 10: p. 558414. [CrossRef]
- Gastaldello, G.H., Cazeloto, A.C.V., Ferreira, J.C., Rodrigues, D.M., Bastos, J.K., Campo, V.L., Zoccal, K.F.,Tefe-Silva, C. Green Propolis Compounds (Baccarin and p-Coumaric Acid) Show Beneficial Effects in Mice for Melanoma Induced by B16f10. Medicines (Basel) 2021, 8(5). [CrossRef]
- Ciazynska, M., Olejniczak-Staruch, I., Sobolewska-Sztychny, D., Narbutt, J., Skibinska, M.,Lesiak, A. Ultraviolet Radiation and Chronic Inflammation-Molecules and Mechanisms Involved in Skin Carcinogenesis: A Narrative Review. Life (Basel) 2021, 11(4). [CrossRef]
- Staniforth, V., Huang, W.C., Aravindaram, K.,Yang, N.S. Ferulic acid, a phenolic phytochemical, inhibits UVB-induced matrix metalloproteinases in mouse skin via posttranslational mechanisms. J Nutr Biochem 2012, 23(5): p. 443-51. [CrossRef]
- Ambothi, K., Prasad, N.R.,Balupillai, A. Ferulic acid inhibits UVB-radiation induced photocarcinogenesis through modulating inflammatory and apoptotic signaling in Swiss albino mice. Food Chem Toxicol 2015, 82: p. 72-8. [CrossRef]
- Wianowska, D.,Olszowy-Tomczyk, M. A Concise Profile of Gallic Acid-From Its Natural Sources through Biological Properties and Chemical Methods of Determination. Molecules 2023, 28(3). [CrossRef]
- Lo, C., Lai, T.Y., Yang, J.H., Yang, J.S., Ma, Y.S., Weng, S.W., Chen, Y.Y., Lin, J.G.,Chung, J.G. Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int J Oncol 2010, 37(2): p. 377-85. [CrossRef]
- Lo, C., Lai, T.Y., Yang, J.S., Yang, J.H., Ma, Y.S., Weng, S.W., Lin, H.Y., Chen, H.Y., Lin, J.G.,Chung, J.G. Gallic acid inhibits the migration and invasion of A375.S2 human melanoma cells through the inhibition of matrix metalloproteinase-2 and Ras. Melanoma Res 2011, 21(4): p. 267-73. [CrossRef]
- Liu, C., Lin, J.J., Yang, Z.Y., Tsai, C.C., Hsu, J.L.,Wu, Y.J. Proteomic study reveals a co-occurrence of gallic acid-induced apoptosis and glycolysis in B16F10 melanoma cells. J Agric Food Chem 2014, 62(48): p. 11672-80. [CrossRef]
- Tseng, T.H., Hsu, J.D., Lo, M.H., Chu, C.Y., Chou, F.P., Huang, C.L.,Wang, C.J. Inhibitory effect of Hibiscus protocatechuic acid on tumor promotion in mouse skin. Cancer Lett 1998, 126(2): p. 199-207. [CrossRef]
- Nakamura, Y., Torikai, K., Ohto, Y., Murakami, A., Tanaka, T.,Ohigashi, H. A simple phenolic antioxidant protocatechuic acid enhances tumor promotion and oxidative stress in female ICR mouse skin: dose-and timing-dependent enhancement and involvement of bioactivation by tyrosinase. Carcinogenesis 2000, 21(10): p. 1899-907. [CrossRef]
- Nakamura, Y., Torikai, K.,Ohigashi, H. A catechol antioxidant protocatechuic acid potentiates inflammatory leukocyte-derived oxidative stress in mouse skin via a tyrosinase bioactivation pathway. Free Radic Biol Med 2001, 30(9): p. 967-78. [CrossRef]
- Lin, H.H., Chen, J.H., Chou, F.P.,Wang, C.J. Protocatechuic acid inhibits cancer cell metastasis involving the down-regulation of Ras/Akt/NF-kappaB pathway and MMP-2 production by targeting RhoB activation. Br J Pharmacol 2011, 162(1): p. 237-54. [CrossRef]
- da Silva, G.B., Manica, D., da Silva, A.P., Marafon, F., Moreno, M.,Bagatini, M.D. Rosmarinic acid decreases viability, inhibits migration and modulates expression of apoptosis-related CASP8/CASP3/NLRP3 genes in human metastatic melanoma cells. Chem Biol Interact 2023, 375: p. 110427. [CrossRef]
- Huang, L., Chen, J., Quan, J.,Xiang, D. Rosmarinic acid inhibits proliferation and migration, promotes apoptosis and enhances cisplatin sensitivity of melanoma cells through inhibiting ADAM17/EGFR/AKT/GSK3beta axis. Bioengineered 2021, 12(1): p. 3065-3076. [CrossRef]
- Olivares, A., Alcaraz-Saura, M., Achel, D.G.,Alcaraz, M. Effect of Rosmarinic Acid and Ionizing Radiation on Glutathione in Melanoma B16F10 Cells: A Translational Opportunity. Antioxidants (Basel) 2020, 9(12). [CrossRef]
- Alcaraz, M., Alcaraz-Saura, M., Achel, D.G., Olivares, A., López-Morata, J.A.,Castillo, J. Radiosensitizing effect of rosmarinic acid in metastatic melanoma B16F10 cells. Anticancer Res 2014, 34(4): p. 1913-21.
- Srinivasulu, C., Ramgopal, M., Ramanjaneyulu, G., Anuradha, C.M.,Suresh Kumar, C. Syringic acid (SA) ‒ A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed Pharmacother 2018, 108: p. 547-557. [CrossRef]
- Cikman, O., Soylemez, O., Ozkan, O.F., Kiraz, H.A., Sayar, I., Ademoglu, S., Taysi, S.,Karaayvaz, M. Antioxidant Activity of Syringic Acid Prevents Oxidative Stress in l-arginine-Induced Acute Pancreatitis: An Experimental Study on Rats. Int Surg 2015, 100(5): p. 891-6. [CrossRef]
- Mihanfar, A., Darband, S.G., Sadighparvar, S., Kaviani, M., Mirza-Aghazadeh-Attari, M., Yousefi, B.,Majidinia, M. In vitro and in vivo anticancer effects of syringic acid on colorectal cancer: Possible mechanistic view. Chem Biol Interact 2021, 337: p. 109337. [CrossRef]
- Ha, S.J., Lee, J., Park, J., Kim, Y.H., Lee, N.H., Kim, Y.E., Song, K.M., Chang, P.S., Jeong, C.H.,Jung, S.K. Syringic acid prevents skin carcinogenesis via regulation of NoX and EGFR signaling. Biochem Pharmacol 2018, 154: p. 435-445. [CrossRef]
- Ghitu, A., Schwiebs, A., Radeke, H.H., Avram, S., Zupko, I., Bor, A., Pavel, I.Z., Dehelean, C.A., Oprean, C., Bojin, F., Farcas, C., Soica, C., Duicu, O.,Danciu, C. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients 2019, 11(4). [CrossRef]
- Madunic, J., Madunic, I.V., Gajski, G., Popic, J.,Garaj-Vrhovac, V. Apigenin: A dietary flavonoid with diverse anticancer properties. Cancer Lett 2018, 413: p. 11-22. [CrossRef]
- Zhao, G., Han, X., Cheng, W., Ni, J., Zhang, Y., Lin, J.,Song, Z. Apigenin inhibits proliferation and invasion, and induces apoptosis and cell cycle arrest in human melanoma cells. Oncol Rep 2017, 37(4): p. 2277-2285. [CrossRef]
- Cao, H.H., Chu, J.H., Kwan, H.Y., Su, T., Yu, H., Cheng, C.Y., Fu, X.Q., Guo, H., Li, T., Tse, A.K., Chou, G.X., Mo, H.B.,Yu, Z.L. Inhibition of the STAT3 signaling pathway contributes to apigenin-mediated anti-metastatic effect in melanoma. Sci Rep 2016, 6: p. 21731. [CrossRef]
- Woo, J.S., Choo, G.S., Yoo, E.S., Kim, S.H., Lee, J.H., Han, S.H., Kim, H.J., Jung, S.H., Park, Y.S., Kim, B.S., Kim, S.K., Park, B.K., Cho, S.D., Nam, J.S., Choi, C.S., Che, J.H.,Jung, J.Y. Apigenin induces apoptosis by regulating Akt and MAPK pathways in human melanoma cell A375SM. Mol Med Rep 2020, 22(6): p. 4877-4889. [CrossRef]
- Xu, L., Zhang, Y., Tian, K., Chen, X., Zhang, R., Mu, X., Wu, Y., Wang, D., Wang, S., Liu, F., Wang, T., Zhang, J., Liu, S., Zhang, Y., Tu, C.,Liu, H. Apigenin suppresses PD-L1 expression in melanoma and host dendritic cells to elicit synergistic therapeutic effects. J Exp Clin Cancer Res 2018, 37(1): p. 261. [CrossRef]
- Mirzoeva, S., Tong, X., Bridgeman, B.B., Plebanek, M.P.,Volpert, O.V. Apigenin Inhibits UVB-Induced Skin Carcinogenesis: The Role of Thrombospondin-1 as an Anti-Inflammatory Factor. Neoplasia 2018, 20(9): p. 930-942. [CrossRef]
- Bridgeman, B.B., Wang, P., Ye, B., Pelling, J.C., Volpert, O.V.,Tong, X. Inhibition of mTOR by apigenin in UVB-irradiated keratinocytes: A new implication of skin cancer prevention. Cell Signal 2016, 28(5): p. 460-468. [CrossRef]
- Byun, S., Park, J., Lee, E., Lim, S., Yu, J.G., Lee, S.J., Chen, H., Dong, Z., Lee, K.W.,Lee, H.J. Src kinase is a direct target of apigenin against UVB-induced skin inflammation. Carcinogenesis 2013, 34(2): p. 397-405. [CrossRef]
- Van Dross, R.T., Hong, X., Essengue, S., Fischer, S.M.,Pelling, J.C. Modulation of UVB-induced and basal cyclooxygenase-2 (COX-2) expression by apigenin in mouse keratinocytes: role of USF transcription factors. Mol Carcinog 2007, 46(4): p. 303-14. [CrossRef]
- Caltagirone, S., Rossi, C., Poggi, A., Ranelletti, F.O., Natali, P.G., Brunetti, M., Aiello, F.B.,Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int J Cancer 2000, 87(4): p. 595-600. [CrossRef]
- Riaz, A., Rasul, A., Hussain, G., Zahoor, M.K., Jabeen, F., Subhani, Z., Younis, T., Ali, M., Sarfraz, I.,Selamoglu, Z. Astragalin: A Bioactive Phytochemical with Potential Therapeutic Activities. Adv Pharmacol Sci 2018, 2018: p. 9794625. [CrossRef]
- You, O.H., Shin, E.A., Lee, H., Kim, J.H., Sim, D.Y., Kim, J.H., Kim, Y., Khil, J.H., Baek, N.I.,Kim, S.H. Apoptotic Effect of Astragalin in Melanoma Skin Cancers via Activation of Caspases and Inhibition of Sry-related HMg-Box Gene 10. Phytother Res 2017, 31(10): p. 1614-1620. [CrossRef]
- Rosenbaum, S.R., Tiago, M., Caksa, S., Capparelli, C., Purwin, T.J., Kumar, G., Glasheen, M., Pomante, D., Kotas, D., Chervoneva, I.,Aplin, A.E. SOX10 requirement for melanoma tumor growth is due, in part, to immune-mediated effects. Cell Rep 2021, 37(10): p. 110085. [CrossRef]
- Min, K.J.,Kwon, T.K. Anticancer effects and molecular mechanisms of epigallocatechin-3-gallate. Integr Med Res 2014, 3(1): p. 16-24. [CrossRef]
- Ravindranath, M.H., Ramasamy, V., Moon, S., Ruiz, C.,Muthugounder, S. Differential growth suppression of human melanoma cells by tea (Camellia sinensis) epicatechins (ECG, EGC and EGCG). Evid Based Complement Alternat Med 2009, 6(4): p. 523-30. [CrossRef]
- Sudha, T., Salaheldin, T.A., Darwish, N.H.,Mousa, S.A. Antitumor/anti-angiogenesis efficacy of epigallocatechin gallate nanoformulated with antioxidant in melanoma. Nanomedicine (Lond) 2022, 17(15): p. 1039-1053. [CrossRef]
- Du, B.X., Lin, P.,Lin, J. EGCG and ECG induce apoptosis and decrease autophagy via the AMPK/mTOR and PI3K/AKT/mTOR pathway in human melanoma cells. Chin J Nat Med 2022, 20(4): p. 290-300. [CrossRef]
- Ravindran Menon, D., Li, Y., Yamauchi, T., Osborne, D.G., Vaddi, P.K., Wempe, M.F., Zhai, Z.,Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals (Basel) 2021, 14(11). [CrossRef]
- Chen, H.Y., Jiang, Y.W., Kuo, C.L., Way, T.D., Chou, Y.C., Chang, Y.S.,Chung, J.G. Chrysin inhibit human melanoma A375.S2 cell migration and invasion via affecting MAPK signaling and NF-kappaB signaling pathway in vitro. Environ Toxicol 2019, 34(4): p. 434-442. [CrossRef]
- Sassi, A., Maatouk, M., El Gueder, D., Bzeouich, I.M., Abdelkefi-Ben Hatira, S., Jemni-Yacoub, S., Ghedira, K.,Chekir-Ghedira, L. Chrysin, a natural and biologically active flavonoid suppresses tumor growth of mouse B16F10 melanoma cells: In vitro and In vivo study. Chem Biol Interact 2018, 283: p. 10-19. [CrossRef]
- Pichichero, E., Cicconi, R., Mattei, M.,Canini, A. Chrysin-induced apoptosis is mediated through p38 and Bax activation in B16-F1 and A375 melanoma cells. Int J Oncol 2011, 38(2): p. 473-83. [CrossRef]
- Alvarez, N., Vicente, V.,Martínez, C. Synergistic effect of diosmin and interferon-alpha on metastatic pulmonary melanoma. Cancer Biother Radiopharm 2009, 24(3): p. 347-52. [CrossRef]
- Buddhan, R.,Manoharan, S. Diosmin reduces cell viability of A431 skin cancer cells through apoptotic induction. J Cancer Res Ther 2017, 13(3): p. 471-476. [CrossRef]
- Martínez Conesa, C., Vicente Ortega, V., Yáñez Gascón, M.J., Alcaraz Baños, M., Canteras Jordana, M., Benavente-García, O.,Castillo, J. Treatment of metastatic melanoma B16F10 by the flavonoids tangeretin, rutin, and diosmin. J Agric Food Chem 2005, 53(17): p. 6791-7. [CrossRef]
- Martinez, C., Vicente, V., Yanez, J., Alcaraz, M., Castells, M.T., Canteras, M., Benavente-Garcia, O.,Castillo, J. The effect of the flavonoid diosmin, grape seed extract and red wine on the pulmonary metastatic B16F10 melanoma. Histol Histopathol 2005, 20(4): p. 1121-9. [CrossRef]
- Zhang, W., Tang, B., Huang, Q.,Hua, Z. Galangin inhibits tumor growth and metastasis of B16F10 melanoma. J Cell Biochem 2013, 114(1): p. 152-61. [CrossRef]
- Benguedouar, L., Lahouel, M., Gangloff, S.C., Durlach, A., Grange, F., Bernard, P.,Antonicelli, F. Ethanolic Extract of Algerian Propolis and Galangin Decreased Murine Melanoma Tumor Progression in Mice. Anticancer Agents Med Chem 2016, 16(9): p. 1172-83. [CrossRef]
- Madduma Hewage, S.R., Piao, M.J., Kim, K.C., Cha, J.W., Han, X., Choi, Y.H., Chae, S.,Hyun, J.W. Galangin (3,5,7-trihydroxyflavone) shields human keratinocytes from ultraviolet B-induced oxidative stress. Biomol Ther (Seoul) 2015, 23(2): p. 165-73. [CrossRef]
- Madduma Hewage, S.R.K., Piao, M.J., Kang, K.A., Ryu, Y.S., Fernando, P., Oh, M.C., Park, J.E., Shilnikova, K., Moon, Y.J., Shin, D.O.,Hyun, J.W. Galangin Activates the ERK/AKT-Driven Nrf2 Signaling Pathway to Increase the Level of Reduced Glutathione in Human Keratinocytes. Biomol Ther (Seoul) 2017, 25(4): p. 427-433. [CrossRef]
- Jaiswal, N., Akhtar, J., Singh, S.P., Badruddeen,Ahsan, F. An Overview on Genistein and its Various Formulations. Drug Res (Stuttg) 2019, 69(6): p. 305-313. [CrossRef]
- Hou, S. Genistein: Therapeutic and Preventive Effects, Mechanisms, and Clinical Application in Digestive Tract Tumor. Evid Based Complement Alternat Med 2022, 2022: p. 5957378. [CrossRef]
- Bhat, S.S., Prasad, S.K., Shivamallu, C., Prasad, K.S., Syed, A., Reddy, P., Cull, C.A.,Amachawadi, R.G. Genistein: A Potent Anti-Breast Cancer Agent. Curr Issues Mol Biol 2021, 43(3): p. 1502-1517. [CrossRef]
- Wang, H.Z., Zhang, Y., Xie, L.P., Yu, X.Y.,Zhang, R.Q. Effects of genistein and daidzein on the cell growth, cell cycle, and differentiation of human and murine melanoma cells(1). J Nutr Biochem 2002, 13(7): p. 421-426. [CrossRef]
- Darbon, J.M., Penary, M., Escalas, N., Casagrande, F., Goubin-Gramatica, F., Baudouin, C.,Ducommun, B. Distinct Chk2 activation pathways are triggered by genistein and DNA-damaging agents in human melanoma cells. J Biol Chem 2000, 275(20): p. 15363-9. [CrossRef]
- Cui, S., Wang, J., Wu, Q., Qian, J., Yang, C.,Bo, P. Genistein inhibits the growth and regulates the migration and invasion abilities of melanoma cells via the FAK/paxillin and MAPK pathways. Oncotarget 2017, 8(13): p. 21674-21691. [CrossRef]
- Heo, J.R., Lee, G.A., Kim, G.S., Hwang, K.A.,Choi, K.C. Phytochemical-induced reactive oxygen species and endoplasmic reticulum stress-mediated apoptosis and differentiation in malignant melanoma cells. Phytomedicine 2018, 39: p. 100-110. [CrossRef]
- Tamura, S., Bito, T., Ichihashi, M.,Ueda, M. Genistein enhances the cisplatin-induced inhibition of cell growth and apoptosis in human malignant melanoma cells. Pigment Cell Res 2003, 16(5): p. 470-6. [CrossRef]
- Alhasan, S.A., Ensley, J.F.,Sarkar, F.H. Genistein induced molecular changes in a squamous cell carcinoma of the head and neck cell line. Int J Oncol 2000, 16(2): p. 333-341. [CrossRef]
- Record, I.R., Broadbent, J.L., King, R.A., Dreosti, I.E., Head, R.J.,Tonkin, A.L. Genistein inhibits growth of B16 melanoma cells in vivo and in vitro and promotes differentiation in vitro. Int J Cancer 1997, 72(5): p. 860-864. [CrossRef]
- Menon, L.G., Kuttan, R., Nair, M.G., Chang, Y.C.,Kuttan, G. Effect of isoflavones genistein and daidzein in the inhibition of lung metastasis in mice induced by B16F-10 melanoma cells. Nutr Cancer 1998, 30(1): p. 74-7. [CrossRef]
- Farina, H.G., Pomies, M., Alonso, D.F.,Gomez, D.E. Antitumor and antiangiogenic activity of soy isoflavone genistein in mouse models of melanoma and breast cancer. Oncol Rep 2006, 16(4): p. 885-91. [CrossRef]
- Wei, H., Bowen, R., Zhang, X.,Lebwohl, M. Isoflavone genistein inhibits the initiation and promotion of two-stage skin carcinogenesis in mice. Carcinogenesis 1998, 19(8): p. 1509-14. [CrossRef]
- Wei, H., Saladi, R., Lu, Y., Wang, Y., Palep, S.R., Moore, J., Phelps, R., Shyong, E.,Lebwohl, M.G. Isoflavone genistein: photoprotection and clinical implications in dermatology. J Nutr 2003, 133(11 Suppl 1): p. 3811S-3819S. [CrossRef]
- Wei, H., Zhang, X., Wang, Y.,Lebwohl, M. Inhibition of ultraviolet light-induced oxidative events in the skin and internal organs of hairless mice by isoflavone genistein. Cancer Lett 2002, 185(1): p. 21-9. [CrossRef]
- Valentova, K., Vrba, J., Bancirova, M., Ulrichova, J.,Kren, V. Isoquercitrin: pharmacology, toxicology, and metabolism. Food Chem Toxicol 2014, 68: p. 267-82. [CrossRef]
- Wullschleger, S., Loewith, R.,Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124(3): p. 471-84. [CrossRef]
- Won, Y.S., Kim, J.H., Lizardo, R.C.M., Min, H.J., Cho, H.D., Hong, S.M.,Seo, K.I. The Flavonol Isoquercitrin Promotes Mitochondrial-Dependent Apoptosis in SK-Mel-2 Melanoma Cell via the PI3K/AKT/mTOR Pathway. Nutrients 2020, 12(12). [CrossRef]
- Gong, G., Guan, Y.Y., Zhang, Z.L., Rahman, K., Wang, S.J., Zhou, S., Luan, X.,Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomed Pharmacother 2020, 128: p. 110301. [CrossRef]
- Duan, R., Liang, X., Chai, B., Zhou, Y., Du, H., Suo, Y., Chen, Z., Li, Q.,Huang, X. Isorhamnetin Induces Melanoma Cell Apoptosis via the PI3K/Akt and NF-kappaB Pathways. Biomed Res Int 2020, 2020: p. 1057943. [CrossRef]
- Kim, J.E., Lee, D.E., Lee, K.W., Son, J.E., Seo, S.K., Li, J., Jung, S.K., Heo, Y.S., Mottamal, M., Bode, A.M., Dong, Z.,Lee, H.J. Isorhamnetin suppresses skin cancer through direct inhibition of MEK1 and PI3-K. Cancer Prev Res (Phila) 2011, 4(4): p. 582-91. [CrossRef]
- Li, J., Mottamal, M., Li, H., Liu, K., Zhu, F., Cho, Y.Y., Sosa, C.P., Zhou, K., Bowden, G.T., Bode, A.M.,Dong, Z. Quercetin-3-methyl ether suppresses proliferation of mouse epidermal JB6 P+ cells by targeting ERKs. Carcinogenesis 2012, 33(2): p. 459-65. [CrossRef]
- Qiang, D., Ci, C., Liu, W., Wang, J., He, C., Ji, B.,Shao, X. Inhibitory effect of kaempferol on mouse melanoma cell line B16 in vivo and in vitro. Postepy Dermatol Alergol 2021, 38(3): p. 498-504. [CrossRef]
- Yang, J., Xiao, P., Sun, J.,Guo, L. Anticancer effects of kaempferol in A375 human malignant melanoma cells are mediated via induction of apoptosis, cell cycle arrest, inhibition of cell migration and downregulation of m-TOR/PI3K/AKT pathway. J buon 2018, 23(1): p. 218-223. [PubMed]
- Zheng, X., Pan, Y., Yang, G., Liu, Y., Zou, J., Zhao, H., Yin, G., Wu, Y., Li, X., Wei, Z., Yu, S., Zhao, Y., Wang, A., Chen, W.,Lu, Y. Kaempferol impairs aerobic glycolysis against melanoma metastasis via inhibiting the mitochondrial binding of HK2 and VDAC1. Eur J Pharmacol 2022, 931: p. 175226. [CrossRef]
- Yao, K., Chen, H., Liu, K., Langfald, A., Yang, G., Zhang, Y., Yu, D.H., Kim, M.O., Lee, M.H., Li, H., Bae, K.B., Kim, H.G., Ma, W.Y., Bode, A.M., Dong, Z.,Dong, Z. Kaempferol targets RSK2 and MSK1 to suppress UV radiation-induced skin cancer. Cancer Prev Res (Phila) 2014, 7(9): p. 958-967. [CrossRef]
- Lee, K.M., Lee, K.W., Jung, S.K., Lee, E.J., Heo, Y.S., Bode, A.M., Lubet, R.A., Lee, H.J.,Dong, Z. Kaempferol inhibits UVB-induced COX-2 expression by suppressing Src kinase activity. Biochem Pharmacol 2010, 80(12): p. 2042-9. [CrossRef]
- Schomberg, J., Wang, Z., Farhat, A., Guo, K.L., Xie, J., Zhou, Z., Liu, J., Kovacs, B.,Liu-Smith, F. Luteolin inhibits melanoma growth in vitro and in vivo via regulating ECM and oncogenic pathways but not ROS. Biochem Pharmacol 2020, 177: p. 114025. [CrossRef]
- Yao, X., Jiang, W., Yu, D.,Yan, Z. Luteolin inhibits proliferation and induces apoptosis of human melanoma cells in vivo and in vitro by suppressing MMP-2 and MMP-9 through the PI3K/AKT pathway. Food Funct 2019, 10(2): p. 703-712. [CrossRef]
- Lamouille, S., Xu, J.,Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 2014, 15(3): p. 178-96. [CrossRef]
- Li, C., Wang, Q., Shen, S., Wei, X.,Li, G. HIF-1α/VEGF signaling-mediated epithelial-mesenchymal transition and angiogenesis is critically involved in anti-metastasis effect of luteolin in melanoma cells. Phytother Res 2019, 33(3): p. 798-807. [CrossRef]
- Ruan, J.S., Liu, Y.P., Zhang, L., Yan, L.G., Fan, F.T., Shen, C.S., Wang, A.Y., Zheng, S.Z., Wang, S.M.,Lu, Y. Luteolin reduces the invasive potential of malignant melanoma cells by targeting beta3 integrin and the epithelial-mesenchymal transition. Acta Pharmacol Sin 2012, 33(10): p. 1325-31. [CrossRef]
- Kim, J.K., Kang, K.A., Ryu, Y.S., Piao, M.J., Han, X., Oh, M.C., Boo, S.J., Jeong, S.U., Jeong, Y.J., Chae, S., Na, S.Y.,Hyun, J.W. Induction of Endoplasmic Reticulum Stress via Reactive Oxygen Species Mediated by Luteolin in Melanoma Cells. Anticancer Res 2016, 36(5): p. 2281-9.
- Li, T., Fu, X., Liu, B., Wang, X., Li, J., Zhu, P., Niu, X., Bai, J., Liu, Y., Lu, X.,Yu, Z.L. Luteolin binds Src, promotes STAT3 protein ubiquitination and exerts anti-melanoma effects in cell and mouse models. Biochem Pharmacol 2022, 200: p. 115044. [CrossRef]
- George, V.C., Naveen Kumar, D.R., Suresh, P.K., Kumar, S.,Kumar, R.A. Comparative studies to evaluate relative in vitro potency of luteolin in inducing cell cycle arrest and apoptosis in HaCaT and A375 cells. Asian Pac J Cancer Prev 2013, 14(2): p. 631-7. [CrossRef]
- Kang, N.J., Jung, S.K., Lee, K.W.,Lee, H.J. Myricetin is a potent chemopreventive phytochemical in skin carcinogenesis. Ann N Y Acad Sci 2011, 1229: p. 124-32. [CrossRef]
- Afroze, N., Pramodh, S., Hussain, A., Waleed, M.,Vakharia, K. A review on myricetin as a potential therapeutic candidate for cancer prevention. 3 Biotech 2020, 10(5): p. 211. [CrossRef]
- Jung, S.K., Lee, K.W., Byun, S., Kang, N.J., Lim, S.H., Heo, Y.S., Bode, A.M., Bowden, G.T., Lee, H.J.,Dong, Z. Myricetin suppresses UVB-induced skin cancer by targeting Fyn. Cancer Res 2008, 68(14): p. 6021-9. [CrossRef]
- Sun, W., Tao, Y., Yu, D., Zhao, T., Wu, L., Yu, W.,Han, W. Myricetin exerts potent anticancer effects on human skin tumor cells. Trop J Pharm 2018, 17: p. 1067-1072. [CrossRef]
- Lee, K.M., Kang, N.J., Han, J.H., Lee, K.W.,Lee, H.J. Myricetin down-regulates phorbol ester-induced cyclooxygenase-2 expression in mouse epidermal cells by blocking activation of nuclear factor kappa B. J Agric Food Chem 2007, 55(23): p. 9678-84. [CrossRef]
- Lee, K.W., Kang, N.J., Rogozin, E.A., Kim, H.G., Cho, Y.Y., Bode, A.M., Lee, H.J., Surh, Y.J., Bowden, G.T.,Dong, Z. Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1. Carcinogenesis 2007, 28(9): p. 1918-27. [CrossRef]
- Kumamoto, T., Fujii, M.,Hou, D.X. Myricetin directly targets JAK1 to inhibit cell transformation. Cancer Lett 2009, 275(1): p. 17-26. [CrossRef]
- Jung, S.K., Lee, K.W., Byun, S., Lee, E.J., Kim, J.E., Bode, A.M., Dong, Z.,Lee, H.J. Myricetin inhibits UVB-induced angiogenesis by regulating PI-3 kinase in vivo. Carcinogenesis 2010, 31(5): p. 911-7. [CrossRef]
- Choi, J., Lee, D.H., Jang, H., Park, S.Y.,Seol, J.W. Naringenin exerts anticancer effects by inducing tumor cell death and inhibiting angiogenesis in malignant melanoma. Int J Med Sci 2020, 17(18): p. 3049-3057. [CrossRef]
- Pafumi, I., Festa, M., Papacci, F., Lagostena, L., Giunta, C., Gutla, V., Cornara, L., Favia, A., Palombi, F., Gambale, F., Filippini, A.,Carpaneto, A. Naringenin Impairs Two-Pore Channel 2 Activity And Inhibits VEGF-Induced Angiogenesis. Sci Rep 2017, 7(1): p. 5121. [CrossRef]
- Skelding, K.A., Barry, D.L., Theron, D.Z.,Lincz, L.F. Targeting the two-pore channel 2 in cancer progression and metastasis. Explor Target Antitumor Ther 2022, 3(1): p. 62-89. [CrossRef]
- Lentini, A., Forni, C., Provenzano, B.,Beninati, S. Enhancement of transglutaminase activity and polyamine depletion in B16-F10 melanoma cells by flavonoids naringenin and hesperitin correlate to reduction of the in vivo metastatic potential. Amino Acids 2007, 32(1): p. 95-100. [CrossRef]
- Ahamad, M.S., Siddiqui, S., Jafri, A., Ahmad, S., Afzal, M.,Arshad, M. Induction of apoptosis and antiproliferative activity of naringenin in human epidermoid carcinoma cell through ROS generation and cell cycle arrest. PLoS One 2014, 9(10): p. e110003. [CrossRef]
- Anand David, A.V., Arulmoli, R.,Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn Rev 2016, 10(20): p. 84-89. [CrossRef]
- Soll, F., Ternent, C., Berry, I.M., Kumari, D.,Moore, T.C. Quercetin Inhibits Proliferation and Induces Apoptosis of B16 Melanoma Cells In Vitro. Assay Drug Dev Technol 2020, 18(6): p. 261-268. [CrossRef]
- Zhang, X.M., Chen, J., Xia, Y.G.,Xu, Q. Apoptosis of murine melanoma B16-BL6 cells induced by quercetin targeting mitochondria, inhibiting expression of PKC-alpha and translocating PKC-delta. Cancer Chemother Pharmacol 2005, 55(3): p. 251-62. [CrossRef]
- Zhang, X., Xu, Q.,Saiki, I. Quercetin inhibits the invasion and mobility of murine melanoma B16-BL6 cells through inducing apoptosis via decreasing Bcl-2 expression. Clin Exp Metastasis 2000, 18(5): p. 415-21. [CrossRef]
- Cao, H.H., Tse, A.K., Kwan, H.Y., Yu, H., Cheng, C.Y., Su, T., Fong, W.F.,Yu, Z.L. Quercetin exerts anti-melanoma activities and inhibits STAT3 signaling. Biochem Pharmacol 2014, 87(3): p. 424-34. [CrossRef]
- Kim, S.H., Yoo, E.S., Woo, J.S., Han, S.H., Lee, J.H., Jung, S.H., Kim, H.J.,Jung, J.Y. Antitumor and apoptotic effects of quercetin on human melanoma cells involving JNK/P38 MAPK signaling activation. Eur J Pharmacol 2019, 860: p. 172568. [CrossRef]
- Zhang, X.M., Huang, S.P.,Xu, Q. Quercetin inhibits the invasion of murine melanoma B16-BL6 cells by decreasing pro-MMP-9 via the PKC pathway. Cancer Chemother Pharmacol 2004, 53(1): p. 82-8. [CrossRef]
- Lin, Y.S., Tsai, P.H., Kandaswami, C.C., Cheng, C.H., Ke, F.C., Lee, P.P., Hwang, J.J.,Lee, M.T. Effects of dietary flavonoids, luteolin, and quercetin on the reversal of epithelial-mesenchymal transition in A431 epidermal cancer cells. Cancer Sci 2011, 102(10): p. 1829-39. [CrossRef]
- Patel, D.H.,Sharma, N. Inhibitory effect of quercetin on epithelial to mesenchymal transition in SK-MEL-28 human melanoma cells defined by in vitro analysis on 3D collagen gels. Onco Targets Ther 2016, 9: p. 6445-6459. [CrossRef]
- Fan, J.J., Hsu, W.H., Lee, K.H., Chen, K.C., Lin, C.W., Lee, Y.A., Ko, T.P., Lee, L.T., Lee, M.T., Chang, M.S.,Cheng, C.H. Dietary Flavonoids Luteolin and Quercetin Inhibit Migration and Invasion of Squamous Carcinoma through Reduction of Src/Stat3/S100A7 Signaling. Antioxidants (Basel) 2019, 8(11). [CrossRef]
- Cao, H.H., Cheng, C.Y., Su, T., Fu, X.Q., Guo, H., Li, T., Tse, A.K., Kwan, H.Y., Yu, H.,Yu, Z.L. Quercetin inhibits HGF/c-Met signaling and HGF-stimulated melanoma cell migration and invasion. Mol Cancer 2015, 14: p. 103. [CrossRef]
- Huang, Y.T., Hwang, J.J., Lee, P.P., Ke, F.C., Huang, J.H., Huang, C.J., Kandaswami, C., Middleton, E., Jr.,Lee, M.T. Effects of luteolin and quercetin, inhibitors of tyrosine kinase, on cell growth and metastasis-associated properties in A431 cells overexpressing epidermal growth factor receptor. Br J Pharmacol 1999, 128(5): p. 999-1010. [CrossRef]
- Rafiq, R.A., Quadri, A., Nazir, L.A., Peerzada, K., Ganai, B.A.,Tasduq, S.A. A Potent Inhibitor of Phosphoinositide 3-Kinase (PI3K) and Mitogen Activated Protein (MAP) Kinase Signalling, Quercetin (3, 3', 4', 5, 7-Pentahydroxyflavone) Promotes Cell Death in Ultraviolet (UV)-B-Irradiated B16F10 Melanoma Cells. PLoS One 2015, 10(7): p. e0131253. [CrossRef]
- Peng, D., Chen, L., Sun, Y., Sun, L., Yin, Q., Deng, S., Niu, L., Lou, F., Wang, Z., Xu, Z., Wang, C., Fan, L., Wang, H.,Wang, H. Melanoma suppression by quercein is correlated with RIG-I and type I interferon signaling. Biomed Pharmacother 2020, 125: p. 109984. [CrossRef]
- Ganeshpurkar, A.,Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm J 2017, 25(2): p. 149-164. [CrossRef]
- Pinzaru, I., Chioibas, R., Marcovici, I., Coricovac, D., Susan, R., Predut, D., Georgescu, D.,Dehelean, C. Rutin Exerts Cytotoxic and Senescence-Inducing Properties in Human Melanoma Cells. Toxics 2021, 9(9). [CrossRef]
- Menon, L.G., Kuttan, R.,Kuttan, G. Inhibition of lung metastasis in mice induced by B16F10 melanoma cells by polyphenolic compounds. Cancer Letters 1995, 95(1): p. 221-225. [CrossRef]
- Choi, K.S., Kundu, J.K., Chun, K.S., Na, H.K.,Surh, Y.J. Rutin inhibits UVB radiation-induced expression of COX-2 and iNOS in hairless mouse skin: p38 MAP kinase and JNK as potential targets. Arch Biochem Biophys 2014, 559: p. 38-45. [CrossRef]
- Liu, N., Wang, K.S., Qi, M., Zhou, Y.J., Zeng, G.Y., Tao, J., Zhou, J.D., Zhang, J.L., Chen, X.,Peng, C. Vitexin compound 1, a novel extraction from a Chinese herb, suppresses melanoma cell growth through DNA damage by increasing ROS levels. J Exp Clin Cancer Res 2018, 37(1): p. 269. [CrossRef]
- Zhang, W., LiPing, Z.,Guo, L. Vitexin inhibited the invasion, metastasis, and progression of human melanoma cells by targeting STAT3 signaling pathway. bioRxiv 2020: p. 2020.09.24.311233. [CrossRef]
- Murakami, A., Ohura, S., Nakamura, Y., Koshimizu, K.,Ohigashi, H. 1'-Acetoxychavicol acetate, a superoxide anion generation inhibitor, potently inhibits tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in ICR mouse skin. Oncology 1996, 53(5): p. 386-91. [CrossRef]
- Batra, V., Syed, Z., Gill, J.N., Coburn, M.A., Adegboyega, P., DiGiovanni, J., Mathis, J.M., Shi, R., Clifford, J.L.,Kleiner-Hancock, H.E. Effects of the tropical ginger compound,1'-acetoxychavicol acetate, against tumor promotion in K5.Stat3C transgenic mice. J Exp Clin Cancer Res 2012, 31(1): p. 57. [CrossRef]
- Li, J., Mao, Y.,Li, W. Anethole suppresses the growth of human skin cancer cells by targeting microRNA498/STAT4 axis. Trop J Pharm 2022, 21(11): p. 2391-2396. [CrossRef]
- Wang, T., Ma, L., Li, W., Ding, L.,Gao, H. MicroRNA-498 reduces the proliferation and invasion of colorectal cancer cells via targeting Bcl-2. FEBS Open Bio 2020, 10(1): p. 168-175. [CrossRef]
- Zhang, X., Xu, X., Ge, G., Zang, X., Shao, M., Zou, S., Zhang, Y., Mao, Z., Zhang, J., Mao, F., Qian, H.,Xu, W. miR-498 inhibits the growth and metastasis of liver cancer by targeting ZEB2. Oncol Rep 2019, 41(3): p. 1638-1648. [CrossRef]
- Lombardi, V.R., Carrera, I.,Cacabelos, R. In Vitro Screening for Cytotoxic Activity of Herbal Extracts. Evid Based Complement Alternat Med 2017, 2017: p. 2675631. [CrossRef]
- Nahar, L., Al-Groshi, A., Kumar, A.,Sarker, S.D. Arbutin: Occurrence in Plants, and Its Potential as an Anticancer Agent. Molecules 2022, 27(24). [CrossRef]
- Jiang, L., Wang, D., Zhang, Y., Li, J., Wu, Z., Wang, Z.,Wang, D. Investigation of the pro-apoptotic effects of arbutin and its acetylated derivative on murine melanoma cells. Int J Mol Med 2018, 41(2): p. 1048-1054. [CrossRef]
- Nawarak, J., Huang-Liu, R., Kao, S.H., Liao, H.H., Sinchaikul, S., Chen, S.T.,Cheng, S.L. Proteomics analysis of A375 human malignant melanoma cells in response to arbutin treatment. Biochim Biophys Acta 2009, 1794(2): p. 159-67. [CrossRef]
- Caterina, M.J., Schumacher, M.A., Tominaga, M., Rosen, T.A., Levine, J.D.,Julius, D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 1997, 389(6653): p. 816-24. [CrossRef]
- Chen, Z.Y., Huang, H.H., Li, Q.C., Zhan, F.B., Wang, L.B., He, T., Yang, C.H., Wang, Y., Zhang, Y.,Quan, Z.X. Capsaicin Reduces Cancer Stemness and Inhibits Metastasis by Downregulating SOX2 and EZH2 in Osteosarcoma. Am J Chin Med 2023, 51(4): p. 1041-1066. [CrossRef]
- Kim, M.Y. Nitric oxide triggers apoptosis in A375 human melanoma cells treated with capsaicin and resveratrol. Mol Med Rep 2012, 5(2): p. 585-91. [CrossRef]
- Shin, D.H., Kim, O.H., Jun, H.S.,Kang, M.K. Inhibitory effect of capsaicin on B16-F10 melanoma cell migration via the phosphatidylinositol 3-kinase/Akt/Rac1 signal pathway. Exp Mol Med 2008, 40(5): p. 486-94. [CrossRef]
- Jun, H.S., Park, T., Lee, C.K., Kang, M.K., Park, M.S., Kang, H.I., Surh, Y.J.,Kim, O.H. Capsaicin induced apoptosis of B16-F10 melanoma cells through down-regulation of Bcl-2. Food Chem Toxicol 2007, 45(5): p. 708-15. [CrossRef]
- Chu, H., Li, M.,Wang, X. Capsaicin induces apoptosis and autophagy in human melanoma cells. Oncol Lett 2019, 17(6): p. 4827-4834. [CrossRef]
- Islam, A., Hsieh, P.F., Liu, P.F., Chou, J.C., Liao, J.W., Hsieh, M.K.,Chueh, P.J. Capsaicin exerts therapeutic effects by targeting tNOX-SIRT1 axis and augmenting ROS-dependent autophagy in melanoma cancer cells. Am J Cancer Res 2021, 11(9): p. 4199-4219. [PubMed]
- Bahri, S., Jameleddine, S.,Shlyonsky, V. Relevance of carnosic acid to the treatment of several health disorders: Molecular targets and mechanisms. Biomed Pharmacother 2016, 84: p. 569-582. [CrossRef]
- Lin, K.I., Lin, C.C., Kuo, S.M., Lai, J.C., Wang, Y.Q., You, H.L., Hsu, M.L., Chen, C.H.,Shiu, L.Y. Carnosic acid impedes cell growth and enhances anticancer effects of carmustine and lomustine in melanoma. Biosci Rep 2018, 38(4). [CrossRef]
- Park, S.Y., Song, H., Sung, M.K., Kang, Y.H., Lee, K.W.,Park, J.H. Carnosic acid inhibits the epithelial-mesenchymal transition in B16F10 melanoma cells: a possible mechanism for the inhibition of cell migration. Int J Mol Sci 2014, 15(7): p. 12698-713. [CrossRef]
- Alcaraz, M., Olivares, A., Andreu-Galvez, M., Achel, D.G., Mercado, A.M.,Alcaraz-Saura, M. Paradoxical Radiosensitizing Effect of Carnosic Acid on B16F10 Metastatic Melanoma Cells: A New Treatment Strategy. Antioxidants (Basel) 2022, 11(11). [CrossRef]
- Arakawa, N., Okubo, A., Yasuhira, S., Takahashi, K., Amano, H., Akasaka, T., Masuda, T., Shibazaki, M.,Maesawa, C. Carnosic acid, an inducer of NAD(P)H quinone oxidoreductase 1, enhances the cytotoxicity of beta-lapachone in melanoma cell lines. Oncol Lett 2018, 15(2): p. 2393-2400. [CrossRef]
- Johnson, J.J. Carnosol: a promising anti-cancer and anti-inflammatory agent. Cancer Lett 2011, 305(1): p. 1-7. [CrossRef]
- O'Neill, E.J., Hartogh, D.J.D., Azizi, K.,Tsiani, E. Anticancer Properties of Carnosol: A Summary of in Vitro and In Vivo Evidence. Antioxidants (Basel) 2020, 9(10). [CrossRef]
- Huang, S.C., Ho, C.T., Lin-Shiau, S.Y.,Lin, J.K. Carnosol inhibits the invasion of B16/F10 mouse melanoma cells by suppressing metalloproteinase-9 through down-regulating nuclear factor-kappa B and c-Jun. Biochem Pharmacol 2005, 69(2): p. 221-32. [CrossRef]
- Choi, S.M., Kim, D.-H., Chun, K.-S.,Choi, J.-S. Carnosol induces apoptotic cell death through ROS-dependent inactivation of STAT3 in human melanoma G361 cells. Appl Biol Chem 2019, 62(1): p. 55. [CrossRef]
- Tong, L.,Wu, S. The Mechanisms of Carnosol in Chemoprevention of Ultraviolet B-Light-Induced Non-Melanoma Skin Cancer Formation. Sci Rep 2018, 8(1): p. 3574. [CrossRef]
- Huang, M.T., Ho, C.T., Wang, Z.Y., Ferraro, T., Lou, Y.R., Stauber, K., Ma, W., Georgiadis, C., Laskin, J.D.,Conney, A.H. Inhibition of skin tumorigenesis by rosemary and its constituents carnosol and ursolic acid. Cancer Res 1994, 54(3): p. 701-8. [PubMed]
- Yeo, I.J., Park, J.H., Jang, J.S., Lee, D.Y., Park, J.E., Choi, Y.E., Joo, J.H., Song, J.K., Jeon, H.O.,Hong, J.T. Inhibitory effect of Carnosol on UVB-induced inflammation via inhibition of STAT3. Arch Pharm Res 2019, 42(3): p. 274-283. [CrossRef]
- Maczka, W., Twardawska, M., Grabarczyk, M.,Winska, K. Carvacrol-A Natural Phenolic Compound with Antimicrobial Properties. Antibiotics (Basel) 2023, 12(5). [CrossRef]
- Joshi, S., Kundu, S., Priya, V.V., Kulhari, U., Mugale, M.N.,Sahu, B.D. Anti-inflammatory activity of carvacrol protects the heart from lipopolysaccharide-induced cardiac dysfunction by inhibiting pyroptosis via NLRP3/Caspase1/Gasdermin D signaling axis. Life Sci 2023, 324: p. 121743. [CrossRef]
- Cerrah, S., Ozcicek, F., Gundogdu, B., Cicek, B., Coban, T.A., Suleyman, B., Altuner, D., Bulut, S.,Suleyman, H. Carvacrol prevents acrylamide-induced oxidative and inflammatory liver damage and dysfunction in rats. Front Pharmacol 2023, 14: p. 1161448. [CrossRef]
- Singh, J., Luqman, S.,Meena, A. Carvacrol as a Prospective Regulator of Cancer Targets/Signalling Pathways. Curr Mol Pharmacol 2023, 16(5): p. 542-558. [CrossRef]
- Govindaraju, S.,Arulselvi, P.I. Characterization of Coleus aromaticus essential oil and its major constituent carvacrol for in vitro antidiabetic and antiproliferative activities. J Herbs Spices Med 2018, 24(1): p. 37-51. [CrossRef]
- Osanloo, M., Alipanah, H., Farjam, M., Taheri, A.,Zarenezhad, E. Anticancer Activity of Chitosan Nanoparticles Containing Satureja khuzistanica Essential Oil, and Carvacrol against Human Melanoma and Breast Cancer. Russ J Bioorg 2023, 49(3): p. 594-601. [CrossRef]
- Nanni, V., Di Marco, G., Sacchetti, G., Canini, A.,Gismondi, A. Oregano Phytocomplex Induces Programmed Cell Death in Melanoma Lines via Mitochondria and DNA Damage. Foods 2020, 9(10). [CrossRef]
- de Oliveira, I.C.V., Galvao-Moreira, L.V., Vilela, J.L., Duarte-Silva, M., Aguiar-da-Silva, L.D., Pereira, C.A.A., Pereira, D.M.S., Pinheiro, A., Lima-Neto, L.G., Fernandes, E.S., Cardoso, C.R.B.,Branco-de-Almeida, L.S. Cinnamaldehyde modulates host immunoinflammatory responses in rat ligature-induced periodontitis and peripheral blood mononuclear cell models. Int Immunopharmacol 2023, 115: p. 109669. [CrossRef]
- Qureshi, K.A., Mohammed, S.A.A., Khan, O., Ali, H.M., El-Readi, M.Z.,Mohammed, H.A. Cinnamaldehyde-Based Self-Nanoemulsion (CA-SNEDDS) Accelerates Wound Healing and Exerts Antimicrobial, Antioxidant, and Anti-Inflammatory Effects in Rats' Skin Burn Model. Molecules 2022, 27(16). [CrossRef]
- Zhang, G., Li, T., Liu, J., Wu, X.,Yi, H. Cinnamaldehyde-Contained Polymers and Their Biomedical Applications. Polymers (Basel) 2023, 15(6). [CrossRef]
- Cabello, C.M., Bair, W.B., 3rd, Lamore, S.D., Ley, S., Bause, A.S., Azimian, S.,Wondrak, G.T. The cinnamon-derived Michael acceptor cinnamic aldehyde impairs melanoma cell proliferation, invasiveness, and tumor growth. Free Radic Biol Med 2009, 46(2): p. 220-31. [CrossRef]
- Zhou, L., Lu, Y., Yang, G.,Wu, J. Research on tumorigenicity of cinnamaldehyde in melanoma cell lines and its mechanism. Tumour Biol 2014, 35(6): p. 5717-22. [CrossRef]
- Patra, K., Jana, S., Sarkar, A., Mandal, D.P.,Bhattacharjee, S. The inhibition of hypoxia-induced angiogenesis and metastasis by cinnamaldehyde is mediated by decreasing HIF-1alpha protein synthesis via PI3K/Akt pathway. Biofactors 2019, 45(3): p. 401-415. [CrossRef]
- Zhang, Y.P., Li, Y.Q., Lv, Y.T.,Wang, J.M. Effect of curcumin on the proliferation, apoptosis, migration, and invasion of human melanoma A375 cells. Genet Mol Res 2015, 14(1): p. 1056-67. [CrossRef]
- Marin, Y.E., Wall, B.A., Wang, S., Namkoong, J., Martino, J.J., Suh, J., Lee, H.J., Rabson, A.B., Yang, C.S., Chen, S.,Ryu, J.H. Curcumin downregulates the constitutive activity of NF-kappaB and induces apoptosis in novel mouse melanoma cells. Melanoma Res 2007, 17(5): p. 274-83. [CrossRef]
- Siwak, D.R., Shishodia, S., Aggarwal, B.B.,Kurzrock, R. Curcumin-induced antiproliferative and proapoptotic effects in melanoma cells are associated with suppression of IkappaB kinase and nuclear factor kappaB activity and are independent of the B-Raf/mitogen-activated/extracellular signal-regulated protein kinase pathway and the Akt pathway. Cancer 2005, 104(4): p. 879-90. [CrossRef]
- Zheng, M., Ekmekcioglu, S., Walch, E.T., Tang, C.H.,Grimm, E.A. Inhibition of nuclear factor-kappaB and nitric oxide by curcumin induces G2/M cell cycle arrest and apoptosis in human melanoma cells. Melanoma Res 2004, 14(3): p. 165-71. [CrossRef]
- Jiang, A.J., Jiang, G., Li, L.T.,Zheng, J.N. Curcumin induces apoptosis through mitochondrial pathway and caspases activation in human melanoma cells. Mol Biol Rep 2015, 42(1): p. 267-75. [CrossRef]
- Bush, J.A., Cheung, K.J., Jr.,Li, G. Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp Cell Res 2001, 271(2): p. 305-14. [CrossRef]
- Liao, W., Xiang, W., Wang, F.F., Wang, R.,Ding, Y. Curcumin inhibited growth of human melanoma A375 cells via inciting oxidative stress. Biomed Pharmacother 2017, 95: p. 1177-1186. [CrossRef]
- Kocyigit, A.,Guler, E.M. Curcumin induce DNA damage and apoptosis through generation of reactive oxygen species and reducing mitochondrial membrane potential in melanoma cancer cells. Cell Mol Biol (Noisy-le-grand) 2017, 63(11): p. 97-105. [CrossRef]
- Bakhshi, J., Weinstein, L., Poksay, K.S., Nishinaga, B., Bredesen, D.E.,Rao, R.V. Coupling endoplasmic reticulum stress to the cell death program in mouse melanoma cells: effect of curcumin. Apoptosis 2008, 13(7): p. 904-14. [CrossRef]
- Zhao, G., Han, X., Zheng, S., Li, Z., Sha, Y., Ni, J., Sun, Z., Qiao, S.,Song, Z. Curcumin induces autophagy, inhibits proliferation and invasion by downregulating AKT/mTOR signaling pathway in human melanoma cells. Oncol Rep 2016, 35(2): p. 1065-74. [CrossRef]
- Chiu, Y.J., Yang, J.S., Tsai, F.J., Chiu, H.Y., Juan, Y.N., Lo, Y.H.,Chiang, J.H. Curcumin suppresses cell proliferation and triggers apoptosis in vemurafenib-resistant melanoma cells by downregulating the EGFR signaling pathway. Environ Toxicol 2022, 37(4): p. 868-879. [CrossRef]
- Yu, T., Ji, J.,Guo, Y.L. MST1 activation by curcumin mediates JNK activation, Foxo3a nuclear translocation and apoptosis in melanoma cells. Biochem Biophys Res Commun 2013, 441(1): p. 53-8. [CrossRef]
- Shimada, K., Ushijima, K., Suzuki, C., Horiguchi, M., Ando, H., Akita, T., Shimamura, M., Fujii, J., Yamashita, C.,Fujimura, A. Pulmonary administration of curcumin inhibits B16F10 melanoma lung metastasis and invasion in mice. Cancer Chemother Pharmacol 2018, 82(2): p. 265-273. [CrossRef]
- Sonavane, K., Phillips, J., Ekshyyan, O., Moore-Medlin, T., Roberts Gill, J., Rong, X., Lakshmaiah, R.R., Abreo, F., Boudreaux, D., Clifford, J.L.,Nathan, C.A. Topical curcumin-based cream is equivalent to dietary curcumin in a skin cancer model. J Skin Cancer 2012, 2012: p. 147863. [CrossRef]
- Phillips, J., Moore-Medlin, T., Sonavane, K., Ekshyyan, O., McLarty, J.,Nathan, C.A. Curcumin inhibits UV radiation-induced skin cancer in SKH-1 mice. Otolaryngol Head Neck Surg 2013, 148(5): p. 797-803. [CrossRef]
- Tsai, K.D., Lin, J.C., Yang, S.M., Tseng, M.J., Hsu, J.D., Lee, Y.J.,Cherng, J.M. Curcumin Protects against UVB-Induced Skin Cancers in SKH-1 Hairless Mouse: Analysis of Early Molecular Markers in Carcinogenesis. Evid Based Complement Alternat Med 2012, 2012: p. 593952. [CrossRef]
- Jensen, J.D., Dunn, J.H., Luo, Y., Liu, W., Fujita, M.,Dellavalle, R.P. Ellagic acid inhibits melanoma growth in vitro. Dermatol Reports 2011, 3(3): p. e36. [CrossRef]
- Wang, F., Chen, J., Xiang, D., Lian, X., Wu, C.,Quan, J. Ellagic acid inhibits cell proliferation, migration, and invasion in melanoma via EGFR pathway. Am J Transl Res 2020, 12(5): p. 2295-2304.
- Hseu, Y.C., Chou, C.W., Senthil Kumar, K.J., Fu, K.T., Wang, H.M., Hsu, L.S., Kuo, Y.H., Wu, C.R., Chen, S.C.,Yang, H.L. Ellagic acid protects human keratinocyte (HaCaT) cells against UVA-induced oxidative stress and apoptosis through the upregulation of the HO-1 and Nrf-2 antioxidant genes. Food Chem Toxicol 2012, 50(5): p. 1245-55. [CrossRef]
- Lembo, S., Balato, A., Di Caprio, R., Cirillo, T., Giannini, V., Gasparri, F.,Monfrecola, G. The modulatory effect of ellagic acid and rosmarinic acid on ultraviolet-B-induced cytokine/chemokine gene expression in skin keratinocyte (HaCaT) cells. Biomed Res Int 2014, 2014: p. 346793. [CrossRef]
- Islam, S.U., Ahmed, M.B., Ahsan, H., Islam, M., Shehzad, A., Sonn, J.K.,Lee, Y.S. An Update on the Role of Dietary Phytochemicals in Human Skin Cancer: New Insights into Molecular Mechanisms. Antioxidants (Basel) 2020, 9(10). [CrossRef]
- Zari, A.T., Zari, T.A.,Hakeem, K.R. Anticancer Properties of Eugenol: A Review. Molecules 2021, 26(23). [CrossRef]
- Padhy, I., Paul, P., Sharma, T., Banerjee, S.,Mondal, A. Molecular Mechanisms of Action of Eugenol in Cancer: Recent Trends and Advancement. Life (Basel) 2022, 12(11). [CrossRef]
- Jaganathan, S.K.,Supriyanto, E. Antiproliferative and molecular mechanism of eugenol-induced apoptosis in cancer cells. Molecules 2012, 17(6): p. 6290-304. [CrossRef]
- Kim, G.-C., Choe, D.S., Im, J.-S., Jeong, H., Kim, I., Lee, M.-H.,Park, B.-S. Caspases-dependent Apoptosis in Human Melanoma Cell by Eugenol. Anat Cell Biol 2006, 39: p. 245-253.
- Ghosh, R., Nadiminty, N., Fitzpatrick, J.E., Alworth, W.L., Slaga, T.J.,Kumar, A.P. Eugenol causes melanoma growth suppression through inhibition of E2F1 transcriptional activity. J Biol Chem 2005, 280(7): p. 5812-9. [CrossRef]
- Ra Choi, B.-B., Shin, S.-H., Kim, U.-K., Hong, J.-W.,Kim, G.-C. S Phase Cell Cycle Arrest and Apoptosis is Induced by Eugenol in G361 Human Melanoma Cells. Int J Oral Biol 2011, 36(3): p. 129-134.
- Júnior, P.L., Câmara, D.A., Costa, A.S., Ruiz, J.L., Levy, D., Azevedo, R.A., Pasqualoto, K.F., de Oliveira, C.F., de Melo, T.C., Pessoa, N.D., Fonseca, P.M., Pereira, A., Araldi, R.P.,Ferreira, A.K. Apoptotic effect of eugenol envolves G2/M phase abrogation accompanied by mitochondrial damage and clastogenic effect on cancer cell in vitro. Phytomedicine 2016, 23(7): p. 725-35. [CrossRef]
- Valizadeh, A., Khaleghi, A.A., Alipanah, H., Zarenezhad, E.,Osanloo, M. Anticarcinogenic Effect of Chitosan Nanoparticles Containing Syzygium aromaticum Essential Oil or Eugenol Toward Breast and Skin Cancer Cell Lines. BioNanoScience 2021, 11(3): p. 678-686. [CrossRef]
- Pal, D., Banerjee, S., Mukherjee, S., Roy, A., Panda, C.K.,Das, S. Eugenol restricts DMBA croton oil induced skin carcinogenesis in mice: downregulation of c-Myc and H-ras, and activation of p53 dependent apoptotic pathway. J Dermatol Sci 2010, 59(1): p. 31-9. [CrossRef]
- Sukumaran, K., Unnikrishnan, M.C.,Kuttan, R. Inhibition of tumour promotion in mice by eugenol. Indian J Physiol Pharmacol 1994, 38(4): p. 306-8.
- Kaur, G., Athar, M.,Alam, M.S. Eugenol precludes cutaneous chemical carcinogenesis in mouse by preventing oxidative stress and inflammation and by inducing apoptosis. Mol Carcinog 2010, 49(3): p. 290-301. [CrossRef]
- Kalmes, M., Hennen, J.,Blömeke, B. Eugenol and isoeugenol as antiproliferative agents in skin cancer cells. Toxicol Lett 2009, 189. [CrossRef]
- Chrubasik, S., Pittler, M.H.,Roufogalis, B.D. Zingiberis rhizoma: a comprehensive review on the ginger effect and efficacy profiles. Phytomedicine 2005, 12(9): p. 684-701. [CrossRef]
- Ganjikunta, V.S., Maddula, R.R., Bhasha, S., Sahukari, R., Kondeti Ramudu, S., Chenji, V., Kesireddy, S.R., Zheng, Z.,Korivi, M. Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats. Molecules 2022, 27(23). [CrossRef]
- de Lima, R.M.T., Dos Reis, A.C., de Menezes, A.P.M., Santos, J.V.O., Filho, J., Ferreira, J.R.O., de Alencar, M., da Mata, A., Khan, I.N., Islam, A., Uddin, S.J., Ali, E.S., Islam, M.T., Tripathi, S., Mishra, S.K., Mubarak, M.S.,Melo-Cavalcante, A.A.C. Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phytother Res 2018, 32(10): p. 1885-1907. [CrossRef]
- Aloliqi, A.A. Therapeutic Potential of 6-Gingerol in Prevention of Colon Cancer Induced by Azoxymethane through the Modulation of Antioxidant Potential and Inflammation. Curr Issues Mol Biol 2022, 44(12): p. 6218-6228. [CrossRef]
- Kim, E.C., Min, J.K., Kim, T.Y., Lee, S.J., Yang, H.O., Han, S., Kim, Y.M.,Kwon, Y.G. [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochem Biophys Res Commun 2005, 335(2): p. 300-8. [CrossRef]
- Nigam, N., Bhui, K., Prasad, S., George, J.,Shukla, Y. [6]-Gingerol induces reactive oxygen species regulated mitochondrial cell death pathway in human epidermoid carcinoma A431 cells. Chem Biol Interact 2009, 181(1): p. 77-84. [CrossRef]
- Kim, S.O., Kundu, J.K., Shin, Y.K., Park, J.H., Cho, M.H., Kim, T.Y.,Surh, Y.J. [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene 2005, 24(15): p. 2558-67. [CrossRef]
- Kim, J.K., Kim, Y., Na, K.M., Surh, Y.J.,Kim, T.Y. [6]-Gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radic Res 2007, 41(5): p. 603-14. [CrossRef]
- Ju, S.A., Park, S.M., Lee, Y.S., Bae, J.H., Yu, R., An, W.G., Suh, J.H.,Kim, B.S. Administration of 6-gingerol greatly enhances the number of tumor-infiltrating lymphocytes in murine tumors. Int J Cancer 2012, 130(11): p. 2618-28. [CrossRef]
- Park, K.K., Chun, K.S., Lee, J.M., Lee, S.S.,Surh, Y.J. Inhibitory effects of [6]-gingerol, a major pungent principle of ginger, on phorbol ester-induced inflammation, epidermal ornithine decarboxylase activity and skin tumor promotion in ICR mice. Cancer Lett 1998, 129(2): p. 139-44. [CrossRef]
- Bischoff-Kont, I.,Furst, R. Benefits of Ginger and Its Constituent 6-Shogaol in Inhibiting Inflammatory Processes. Pharmaceuticals (Basel) 2021, 14(6). [CrossRef]
- Wu, H., Hsieh, M.C., Lo, C.Y., Liu, C.B., Sang, S., Ho, C.T.,Pan, M.H. 6-Shogaol is more effective than 6-gingerol and curcumin in inhibiting 12-O-tetradecanoylphorbol 13-acetate-induced tumor promotion in mice. Mol Nutr Food Res 2010, 54(9): p. 1296-306. [CrossRef]
- Chen, F., Tang, Y., Sun, Y., Veeraraghavan, V.P., Mohan, S.K.,Cui, C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J Photochem Photobiol B 2019, 197: p. 111518. [CrossRef]
- Krause, S.T., Liao, P., Crocoll, C., Boachon, B., Forster, C., Leidecker, F., Wiese, N., Zhao, D., Wood, J.C., Buell, C.R., Gershenzon, J., Dudareva, N.,Degenhardt, J. The biosynthesis of thymol, carvacrol, and thymohydroquinone in Lamiaceae proceeds via cytochrome P450s and a short-chain dehydrogenase. Proc Natl Acad Sci U S A 2021, 118(52). [CrossRef]
- Rathod, N.B., Kulawik, P., Ozogul, F., Regenstein, J.M.,Ozogul, Y. Biological activity of plant-based carvacrol and thymol and their impact on human health and food quality. Trends Food Sci 2021, 116: p. 733-748. [CrossRef]
- Satooka, H.,Kubo, I. Effects of thymol on B16-F10 melanoma cells. J Agric Food Chem 2012, 60(10): p. 2746-52. [CrossRef]
- Calo, R., Visone, C.M.,Marabini, L. Thymol and Thymus Vulgaris L. activity against UVA- and UVB-induced damage in NCTC 2544 cell line. Mutat Res Genet Toxicol Environ Mutagen 2015, 791: p. 30-7. [CrossRef]
- Mapelli, M., Calò, R.,Marabini, L. Thymol and Thymus vulgaris extract protects human keratinocyte cell line (HaCaT) from UVA and UVB damage. Oxid Antioxid Med Sci 2016, 5: p. 39. [CrossRef]
- Cornaghi, L., Arnaboldi, F., Calo, R., Landoni, F., Baruffaldi Preis, W.F., Marabini, L.,Donetti, E. Effects of UV Rays and Thymol/Thymus vulgaris L. Extract in an ex vivo Human Skin Model: Morphological and Genotoxicological Assessment. Cells Tissues Organs 2016, 201(3): p. 180-92. [CrossRef]
Figure 1.
Classification of phenolic compounds.
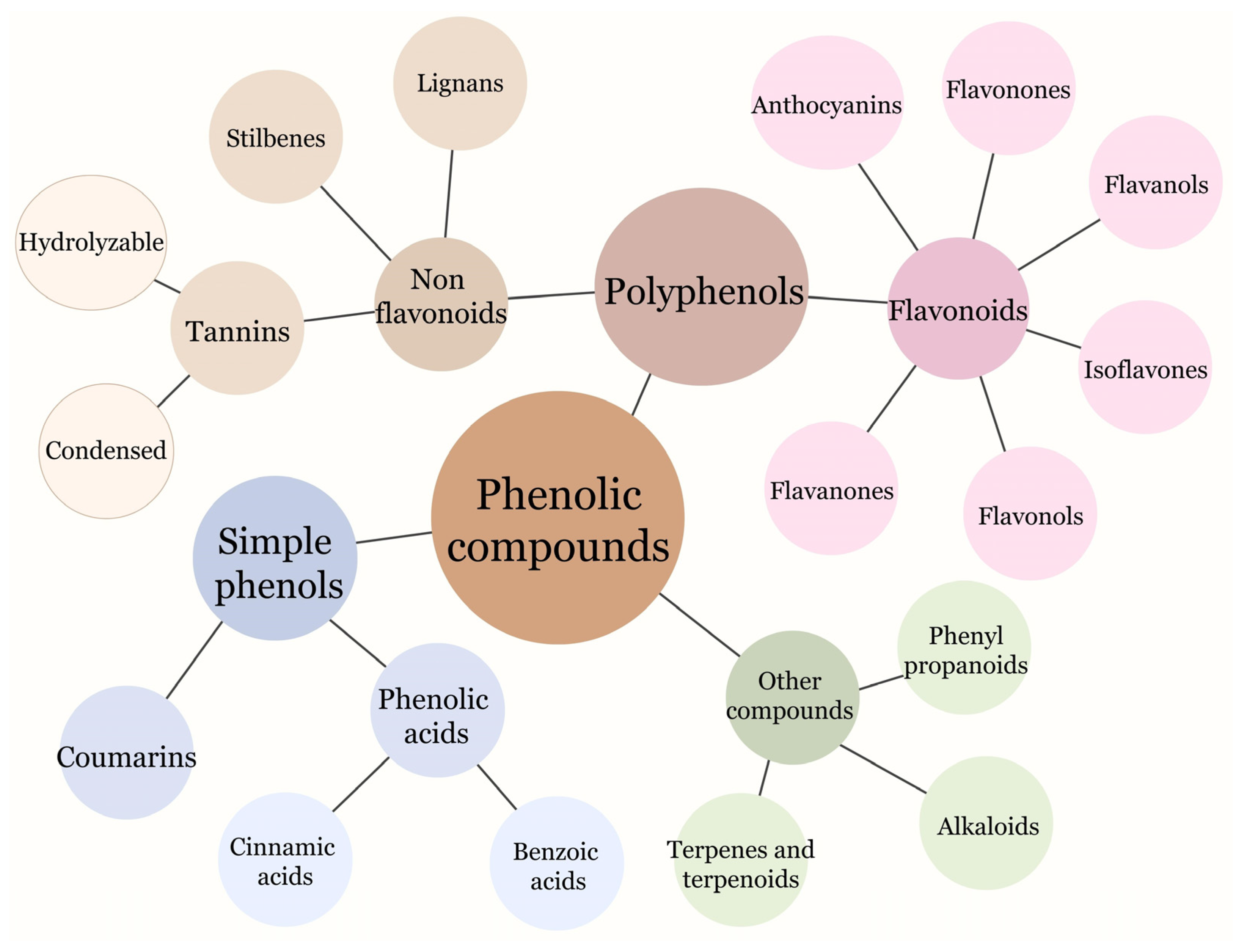
Figure 2.
Structures of phenolic acids discussed in this review. A - hydroxycinnamic acid derivatives, B - hydroxybenzoic acid derivatives.
Figure 2.
Structures of phenolic acids discussed in this review. A - hydroxycinnamic acid derivatives, B - hydroxybenzoic acid derivatives.
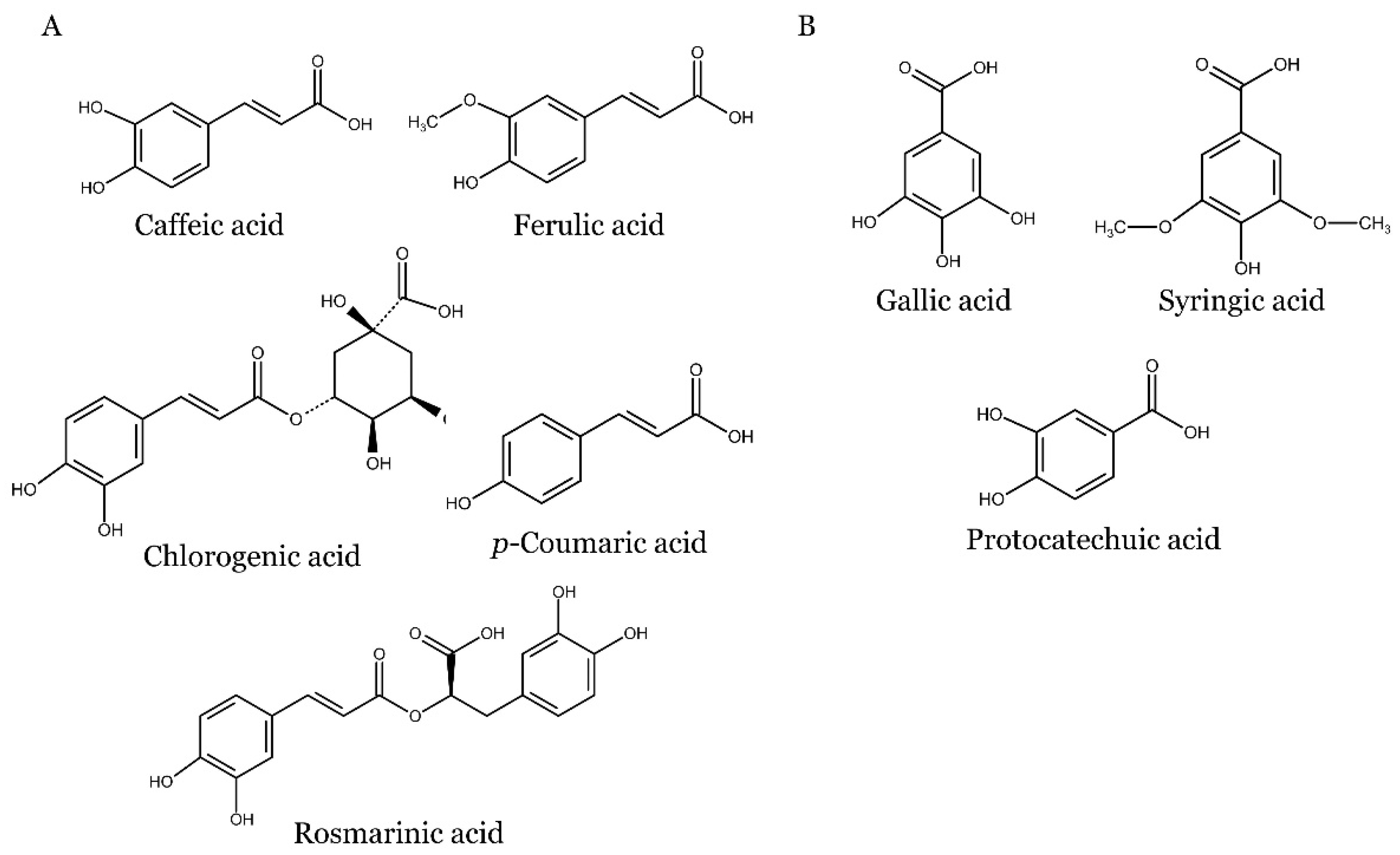
Figure 3.
Structures of selected flavonoids discussed in this review.
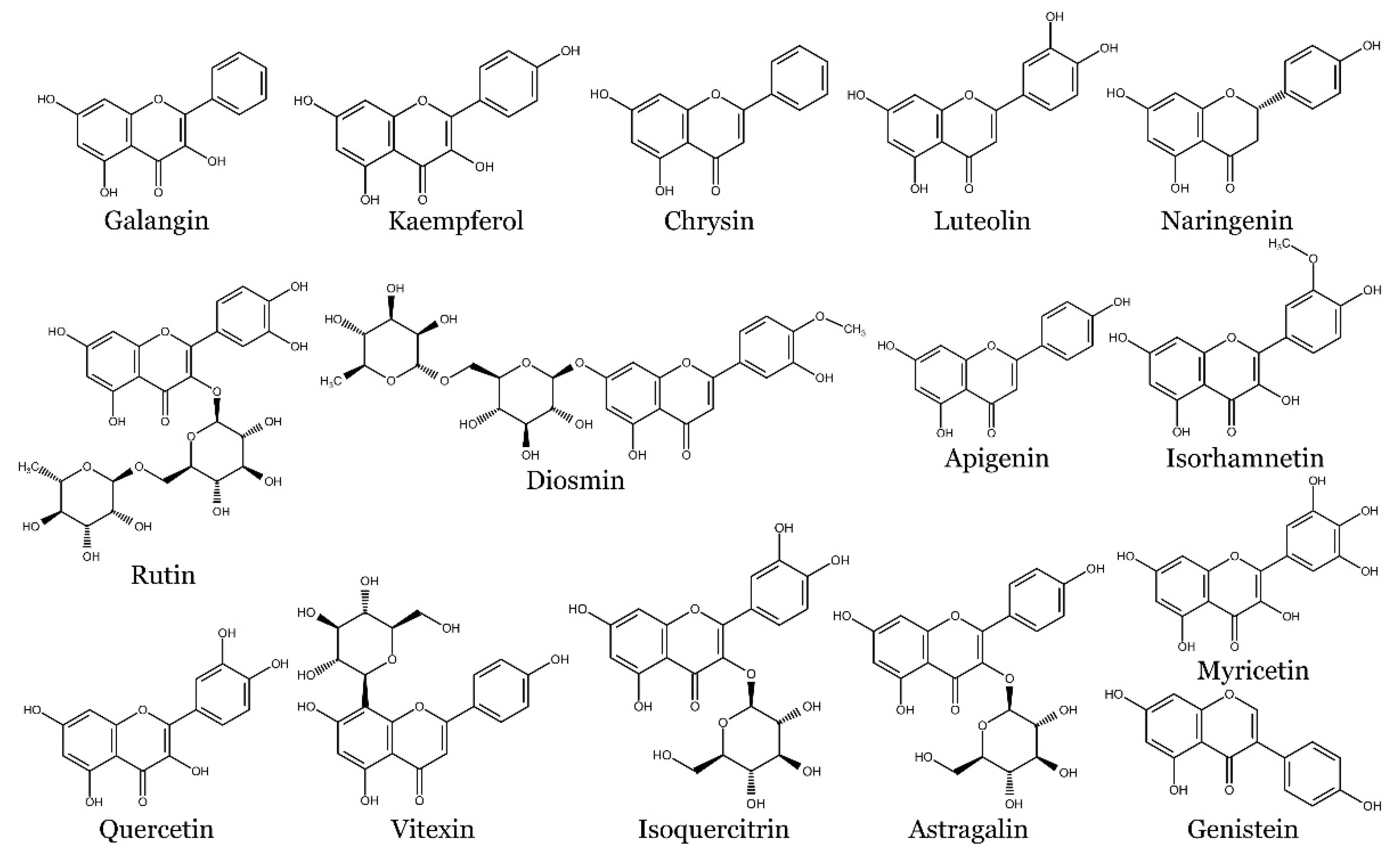
Figure 4.
Structures of selected phenolic compounds discussed in this review.
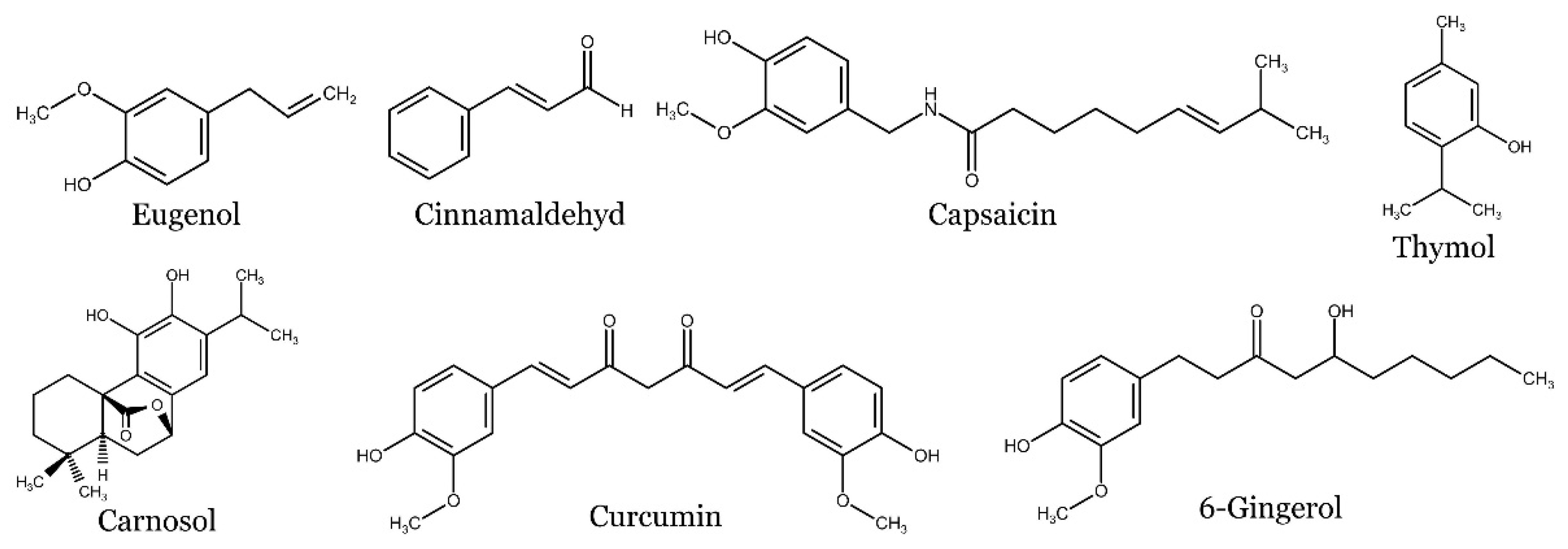
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



