
Submitted:
01 August 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
nutrition, macro and micronutrients, bone health, exercise, exogenous and endogenous microRNA, epigenetics
Keywords:
nutrition
; macro and micronutrients
; bone health
; exercise
; exogenous and endogenous microRNA
; epigenetics
1. Introduction
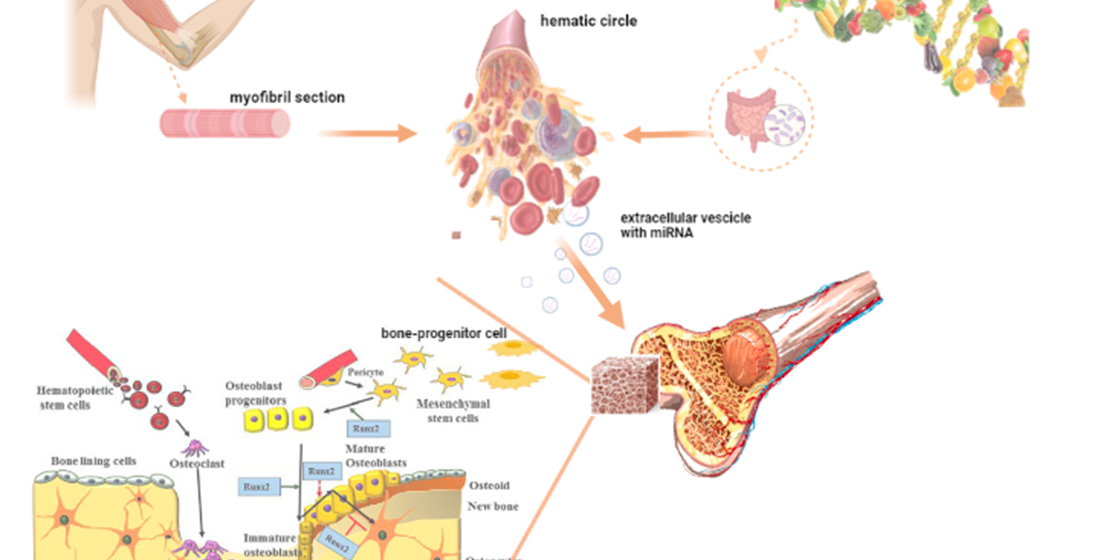
Epigenetic mechanisms may influence gene activity at the transcriptional and post-transcriptional levels [1,2]. Among the most studied epigenetic modifications, there are changes in the expression profile of microRNAs (miRNAs or miRs) [3]. Furthermore, it is already known that diet along with exercise, can modulate the expression of endogenous miRNAs and prevent or delay the development of some metabolic diseases such as bone disorders confirming the importance of epigenetics in bone regeneration [4,5,6,7,8,9,10,11]. In addition, some foods contain miRNAs that after ingestion can influence the various biological processes [12,13]. The bone tissue is metabolically active and it is constantly remodelling in response to different stimuli [14,15,16,17]. MiRNAs expression together with specific transcription factors control the differentiation of the mesenchymal cells from which the osteogenic line cells originate [9]. They can therefore regulate multiple processes crucial for homeostasis, such as cell proliferation, differentiation, survival, and apoptosis and critically influence the regulation of different cellular processes and post-transcriptional gene expression allowing cells to adapt to the environment [1,15]. Recent findings show that both exogenous and endogenous miRNAs may be able to withstand several critical conditions of the extracellular environment and circulate in body fluids because they are protected from degradation by ribonucleases by "packing" into lipid nanovesicles called exosomes [12,18,19,20,21] or associated with argonaute protein 2 (Ago2) that is the key components of the RNA induced silencing complex (RISC) [1,12,18,21]. In humans, Ago2 possess the activity of miRNA-guided mRNA cleavage or translational inhibition; it also has been demonstrated that it interacts with the mRNA-binding protein GW182, a mRNA-binding protein weighing 182 kDa, which is a marker for cytoplasmic foci referred to as GW bodies (GWBs) proposed to be cytoplasmic sites for mRNA storage and degradation [22]. Alterations in the expression of specific miRNAs, which can be collected from peripheral blood and used as biomarkers to study the organic responses, are associated with an increased risk of some medical conditions such as fractures and bone disorders [15]. Several miRNAs are involved in the osteogenic differentiation of progenitor cells and influence some cellular processes in response to exercise and nutrition as we will describe in the following sections.
2. Exercise and Osteogenic miRNAs Expression
Bone tissue is a tissue that is sensitive to mechanical stimuli. Mechanical loads, including compression and deformation, are the stimuli that play essential roles in the differentiation and mineralization of osteoblasts, as well as maintaining high bone mass and density decreasing the risk of osteoporosis [23]. Some studies show that high-impact exercise increases bone mineral density (BMD) [9,24]. The load applied to the bone must always be gradual and administered considering the age of the subject and eventually, the pathologies that are in place. However, in vitro experiment performed on osteoclast, indicates that mechanical stimulus increases the expression of some miRNAs; these mechanisms could be considered important therapeutic candidates for the prevention and treatment of bone diseases, in particular for osteoporosis [25,26]. Recently it has been shown that the half marathon increases the expression of miR-21-5p that promotes the proliferation of mesenchymal stem cells by targeting the two antagonists of the Transforming growth factor beta (TGFb) pathway: Phosphatase and tensin homologue (PTEN) and Small mother against decapentaplegic 7 (Smad7) by reducing their expression [9]. PTEN antagonizes the phosphatidylinositol 3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) pathway via lipid phosphatase activity and controls different cellular processes including survival, proliferation, energy metabolism, and cellular architecture [27]. TGFb is a pleiotropic cytokine that regulates many processes such as cell growth, differentiation, apoptosis, migration, and immune response. Smad7 is a TGF-β/ Smad signal antagonist and is a negative regulator of Runt-related transcription factor 2 (RUNX2); it reduces PTEN expression, accelerates osteoblast differentiation and increases cell survival through PI3K/Akt signalling [28,29]. In fact, in vitro, PTEN-deficient osteoblasts proliferate faster by reducing apoptosis and increasing the cell size; this is consistent with the effects of activation of Akt and mTOR pathways [30]. Reduced expression of PTEN and SMAD7 induced an increase in the AKT/pAKT and SMAD4 protein levels, and this overregulates RUNX2 gene expression [9].
In addition, it has been seen that the proliferative potential of mesenchymal stem cells decreases with age due to the telomere shortening. During cellular division, it is possible to lose more telomeres losing consequently genetic information, an ageing process that leads to cell death. Telomere length is therefore important to counteract the senescence of mesenchymal stem cells and telomerase plays an important role in this regard and in promoting the differentiation process [31]. Telomerase is an RNA-dependent DNA polymerase that synthesizes telomeric DNA sequences and provides the molecular basis for unlimited proliferative potential [32]. Oxidative stress in response to exercise increases the expression of the human telomerase reverse transcriptase gene (hTERT) in human marrow stromal cells (hMSC), increasing telomerase activity that promotes osteoblast differentiation [33].
Hua-Yu Zhu et al. have shown that the same miR-21 can regulate hTERT via PTEN in different processes [34]. Telomerase has also been reported to regulate autophagy with an important role in the differentiation and regulation of stem cells of various cell types, including mesenchymal stem cells [35]. The increased expression of telomerase may be due to oxidative stress resulting from physical activity. Autophagy would appear to be involved in the regulation of the cellular redox state induced by physical activity through the degradation of iron-binding proteins such as ferritin [35].
Furthermore, is already known that resistance exercise induces an increase in iron levels, probably related to tissue damage. Excess iron is considered toxic precisely because of its ability to accept and donate electrons and to be involved in the reaction called “The Fenton reaction” in which reacting with hydrogen peroxide generates both hydroxyl radicals and higher oxidation states of the iron evolving into free radicals [36,37].
The increase in osteogenesis-related genes expression following physical exercises such as RUNX2, muscle segment homeobox 1 (MSX1) and secreted phosphoprotein (SPP1) act to increase the expression of hTERT and telomere repeat binding factor 1 (TERF1), indicates the activity of mesenchymal stem cells and their increased capability to differentiate [33].
3. Endogenous miRNAs and Bone Metabolism
As we already said the miRNAs are a group of small non-coding RNAs that regulate gene expression at the post-transcriptional level and cellular processes [38]. Notwithstanding most miRNAs are found inside cells, there is a large portion that migrates outside in the body fluids such as blood, urine, saliva, seminal fluid, breast milk and other biological specimens in response to tissue damage, apoptosis and necrosis through active transport into exosomes or bonded to a protein as previously described [39]. Early studies have shown that about 10% of circulating miRNAs are secreted into exosomes, while the remaining 90% form complexes with proteins such as Ago2 [40,41].
These systems protect them from the degradation induced by ribonucleases and ensure their stability in body fluids; maybe this is the reason why they could play an essential role as biomarkers, because of their high specificity for the type of tissue or cell of the origin [1]. Recently, has increased the interest to point out the miRNAs directly involved in osteogenic processes in response to exercise and nutrition. A study performed by Vimalraj et al. performed on mice has identified in RUNX2 gene expression the predominant factors required for osteoblast differentiation [14]. In particular, the isoform Type II of this gene has been found in undifferentiated mesenchymal cells, pre-osteoblasts and chondrocyte precursors e, in general, it is necessary for the differentiation of pluripotent mesenchymal cells to osteoblasts [42]. The osteoblasts differentiation is enhanced by the transcription factor Osterix (Osx), a downstream effector of RUNX2. Up to the present, several regulators such as signal transducer and activator of transcription 1 (Stat1), Twist Family BHLH Transcription Factor 1 (Twist) and Hes-related family bHLH transcription factor with YRPW motif 1 (Hey1) form a complex with RUNX2 resulting in osteoblast differentiation inhibitions. This is a fairly complex phenomenon as a critical balance between the differentiation of osteoblasts and the activity of osteoclasts to maintain bone homeostasis [14]. One of the most studied miRNAs is miR-21; the studies confirm the main role to promote stem cell osteogenesis through the Smad7-Smad1/5/8-RUNX2 and Akt pathways [15,29]. The over-expression of miR-21 acts as a promoter of osteogenesis by improving matrix mineralization and fracture healing [14]; Chen et al., highlighted the reduction in miR-21 levels in bone tissue and serum in osteoporosis patients [43]. The miR-21 inhibitions determine a reduction of the osteogenic capacity of human bone marrow mesenchymal stem cells (BMSCs) and calcium deposition by inducing osteoporosis [15]. However, the investigations on the miRNAs that can influence the RUNX2 expressions are constantly evolving [14]. In addition, as we will discuss below, the expression of osteogenic miRNAs is influenced by physical exercise but also by nutrition.
4. Micronutrients Intake and Osteogenic miRNAs Expression
Nutrients can interact directly with the genome and indirectly through modulation of mechanisms, including DNA methylation, histone modification and non-coding RNA expression, in particular the miRNAs. Vitamins and minerals can induce the expression of miRNAs through the activation of transcription factors, which regulate the gene expression by induction of messenger RNA (mRNA) degradation or inhibit their translation. They can also modify the expression of DNA methyltransferases (DNMT) and different enzymes such as histone deacetylase and histone acetyltransferase that are involved in several processes such as transcription activation, gene silencing, DNA repair and cell cycle progression [44]. DNMT would appear to prevent demethylation in postmitotic neurons, which together with DNA methylation provides an epigenetic mechanism of gene regulation in neural development, function and disorders [45]. The modulation of the activity of these enzymes leads to changes in the methylation state of DNA as well as the histones, which in turn modulate the expression of some genes, including the miRNAs themselves [6]. Among the micronutrients, vitamin D plays a key role in promoting bone mineralization [5], facilitating the intestinal absorption of phosphorus and calcium intake with the diet that is involved in bone calcification. Vitamin D is a member of the steroid hormones families with nuclear steroid receptors (NR) signalling function and is involved in the biogenesis and regulation of miRNAs expression [5,6]. It is already known that estrogens have fundamental anticatabolic and anabolic effects on bones; therefore, the lack of estrogen plays a central role in the development of osteoporosis [46,47]. The active form, 1,25 dihydroxy vitamin D (1,25(OH)2 D), can regulate the expression of osteoblastic mineralization factors by affecting the expression of some specific miRNAs like miR-637 and miR-1228. The miR-637 would seem to act by degrading the mRNA of Collagen Type IV Alpha 1 Chain (COL4A1), whose expression, during osteoblastic differentiation, inhibits matrix mineralization; while miR-1228, is a mirtron, an alternative precursor for microRNA biogenesis that was recently described in invertebrates, that uses a different mechanism of action than classical miRNAs, as they bypass the cleavage of enzyme DROSHA, is exported out of the nucleus, split by endoribonuclease Dicer and incorporated into the RISC [48]. The miR-1228 reduces the expression of the Bone Morphogenetic Protein 2 (BMP2K) inducible kinase, a protein potentially implicated in cellular endocytosis and differentiation, but its molecular functions have remained unknown; it seems to be a potent inducer of bone formation through its stimulation of osteoblast differentiation [5]. Given the importance of vitamin D, it is therefore very important to guarantee high levels which can be achieved through diet, such as cholecalciferol, thanks to animal foods intake such as salmon and blue fish or mackerel or cod liver oil consumed mainly as a supplement. Another source of vitamin D is vitamin D2 or ergocalciferol which is a bit less active but of which plant-based food is rich, for example, mushrooms. In any case, 80% of the vitamin D needed is guaranteed by sun exposition. Therefore the best practice to increase vitamin D levels is to carry out physical exercise outdoors to guarantee good production and positive action also on bone metabolism [9,24]. Even vitamin C plays a positive role in bone health; in fact, it is a cofactor in multiple biological processes such as collagen synthesis and antioxidant capability, regulating stem cell differentiation and improving osteoblast activity [6,8,49] Clinical studies performed in humans and animals have shown that a deficiency of vitamin C leads to musculoskeletal alterations; noteworthy, the results showed that 100 mM of vitamin C effectively activates genes related to the musculoskeletal system in BMSCs, whilst lower doses, 25 mM, didn’t induce any effect. Similar results have also been observed for the regulation of vitamin C-dependent miRNAs production [50,51]. There is a possibility that vitamin C treatments regulate miR-29b-1 and miR-589-5p expression by promoting Octamer-binding transcription factor ¾ (Oct3/4), Nanog, sex-determining region Y-box 2 (SOX2) and Mitogen-Activated Protein Kinase Kinase Kinase 8 (MAP3K8) expressions in BMSCs contributing to cell proliferation and differentiation. In addition, it increases the expression of miR-371b-5p, miR-181a and miR-215 in BMSCs. The study hypothesis would be that these miRNAs respectively promote cell proliferation and differentiation of these cells [51,52,53]. Among the various micronutrients that contribute to bone health, is also orthosilicic acid (OSA) which stimulates osteoblastic differentiation. Studies on ovariectomized rats orthosilicic acid fed compared to other deprived rats, showed a higher Bone Mineral Density (BMD) and trabecular thickness. Recently it has been discovered that miR-130b plays a role in cell proliferation, differentiation, and apoptosis; its expression increased during the osteogenic differentiation of multipotent mesenchymal stem cells of the human bone marrow. The study provided by Yunhao You et al. found an increase in his level during osteogenesis in response to 20 mM of orthosilicic acid and suggested that overexpression of miR-130b promoted osteogenic differentiation. However, the mechanisms of action which promote his transcription, have yet to be explored. Nutrition can contribute to the intake of OSA. Indeed its absorption is more effective if taken from the liquid phase in which it is dissolved and easily assimilated not having to undergo major changes [11]. Regular mineral water contains about 6.8 mg per litre but there are some water whose silicon content can range from 14.4 mg up to 60 mg per litre. In food it is mainly contained in the leathery parts of vegetables; for this reason, it is important to consume vegetables and fruits with peel and legumes and prefer foods that have undergone few industrial processes to preserve their structure and content. Recent studies suggest that other micronutrients, such as natural phenolic acids, usually found in plants that are commonly intake by diet, may have an important bone anti-resorption activity [4,54,55]. One of these compounds is syringic acid, 3,5-dimethoxy-4-hydroxybenzoic acid (SA), a phenolic acid acts on mouse mesenchymal stem cells (mMSC) cells inducing the differentiation of osteoblasts. It increases miR-21 expression which reduces Smad7 activity by targeting the TGF-b/BMP signalling pathway, resulting in increased RUNX2 expression, thus leading to the expression of osteoblast differentiation markers genes such as alkaline phosphatase (ALP), Collagen Type I Alpha 1 (Col-I) and osteocalcin (OCN) in BMSCs [56]. Also, isoflavones i.e. syringe, genistein, laminarin, hesperetin and sulphurin, promote osteoblast differentiation through activation of the BMP2/SMAD5/Akt/RUNX2 pathway [57]. Deep SA has several positive effects on bone health due to its strong antioxidant activity and also through antihypertensive, antiproliferative, antiendotoxic, antitumor, hepatoprotective and antihyperglycemic effects. It is mainly present in olives, walnuts and dates but also in blue-coloured fruits and berries where it is formed by the decomposition of lenin, an anthocyanin, and its aglycone, malvidin [58]. This justifies its presence in red wine (0.27 mg/100 mL) and red vinegar (0.30 mg/100 mL) whilst in dried fruits such as walnuts or peanuts as well as in olives, cocoa pulp, pumpkin, durum wheat and in smaller quantities in peas and cauliflower, the concentration can reach 33mg/100g [11].
5. Macronutrients Intake and Osteogenic miRNAs Expression
Macronutrients influence important signalling pathways that regulate human metabolism. However, it's not effortless to discriminate whether a nutrition-gene interaction is the result of a direct or indirect effect due to the involvement of several bioactive components. In other words, nutrients can induce epigenetic changes, either through methylation of DNA or changes in some miRNAs expression. Carbohydrates or Carbs (CHO), proteins and fats are broken down during digestion into the composition monomers respectively monosaccharides, amino acids and fatty acids [10]. Few studies have been done on the role of carbs and lipids on bone health. Currently, Kang Gan et al. investigated the roles of miR-221-3p and miR-222-3p, in regulating the osteogenic differentiation of BMSCs under high blood glucose conditions. The results showed increased expression of these two miRNAs in the bone tissue of diabetic mice, inhibiting osteogenic differentiation via the IGF-1/ERK pathway activation [59]. Further studies showed that high blood glucose levels, together with increased free fatty acids (FFA), increased the expression of miR-449, which inhibits osteogenic differentiation of BMSCs through suppression of the Sirt1/Fra-1 (Fos-related antigen) pathway [60]. MiR-449 directly targets sirtuin 1 (Sirt1) of the SIRT family by binding the 3’-UTR sequence. Sirt1 belongs to the NAD+-dependent enzymes classes that catalyze the deacylation of acyl-lysine residues that regulate the life span of mammals, cellular energy metabolism and the balance between osteoclastic and osteoblastic activity through different signalling pathways [61]. Shakibaei M. et al. reported that Sirt-1 activated by resveratrol (a flavonoid) inhibits osteoclastogenesis; also 1 Fra-1, a protein belonging to the activator protein 1 (AP-1) family of transcription factors, plays an essential role in osteogenesis. Increased expression of miR-449, also significantly reduced mRNA and protein expression levels of osteogenic differentiation-related marker genes, including RUNX2, bone sialoprotein (BSP), collagen I, and OCN. The latter regulates bone mineralization and bone turnover in the late stages of osteoblast differentiation, whilst BSP is involved in the mineralization [62].
Some studies suggest that a diet containing low amounts of methionine (belonging to the sulfur amino acids, SAAs) increases the expression of the miRNAs that alter RUNX2 expression by altering bone structure in mice. This is important because the methionine cycle generates S-adenosyl methionine (SAM), a coenzyme used by DNA methyltransferases to methylate the histones and regulate the gene expression. Particularly these studies demonstrated that mice fed low-methionine food, compared to control mice, had high expression of miR-204 in the bone marrow; this miRNA regulates Osterix and RUNX2 in bone, inhibiting the amount and function of osteoblasts and by inducing bone fragility. Maternal bone mass decreases during lactation since skeletal calcium is released into breast milk. Although renal calcium excretion is reduced with increased tubular reabsorption and this is not sufficient to prevent bone loss. During lactation there is evidence of increased parathormone-related peptide (PTHrP) homologs with the N-terminal fragment of parathyroid hormone (PTH) produced by the mammary glands, which plays a key role in increasing blood calcium concentration and, in combination with low estradiol levels, leads to high rates of bone resorption. However, weaning triggers skeletal recovery that occurs very rapidly after the end of lactation [63].
Regarding the effects of the proteins, a study carried out on maternal nutrition has shown that a low-protein diet negatively regulates mother and child bone mass [64]; however, there are no studies investigating the epigenetic effect of a high-protein diet.
Ioannis Kanakis et al. showed that there was a correlation between bone mineralization and the level of protein dietary intake during lactation in mice; the expression of RUNX2, as well as Alp and Col1a1, were all decreased mostly in mice with a low-protein intake diet compared to the control mice. This indicates a direct correlation resulting in decreased osteoblastic differentiation and activity, particularly in miR-26a, 34a and 125b expression. In deep, miR-125b normally regulates the osteogenic differentiation of human MSCs whilst miR-26a reverses the bone regeneration deficit of MSCs and miR-34a inhibits osteoclastogenesis. The main pathways concerned appear to be the Wnt and IL-6 signalling pathways [65]. Therefore, protein malnutrition increases bone loss, and slows down and delays bone recovery.
Fully understanding the mechanisms could lead to draft nutritional guidelines for improving bone health. However, the effects of dietary proteins on bone health must be considered according to age, health, the diet habits of the population and exercise practice [66]. Certainly, is well known that a high protein intake increases urinary excretion of Calcium (Ca) and on average is estimated that at least 1 mg of Ca is excreted for each additional gram of protein consumed [67]. This relationship is mainly attributable to the metabolism of sulfur amino acids contained especially in animal proteins and some plant proteins, resulting in increased acidity buffered by organic calcium release from the skeleton. The effects of proteins on bones can also depend on the intake of foods rich in calcium and alkalis, such as fruits and vegetables. A low protein intake reduces insulin-like growth factor production, which in turn hinders calcium and phosphate metabolism, bone formation, and the promotion of satellite cell activation [68]. These effects probably depend on the amount and type of protein and influence bone health through epigenetic mechanisms [69].
6. Exogenous miRNA and Bone Metabolism
So far, we have focused on the interaction between macronutrients and endogenous microRNA production and how this can induce epigenetic changes that affect bone health. New scientific evidence suggests that microRNA content in foods can survive the processes of digestion, absorption, and transport in the biological fluids by regulating the expression of some genes correlated with different biological processes [18,70]. Food-derived microRNAs could act as a new functional component of foods, such as vitamins and other nutrients, that can influence human health [6,12]. It is important to consider the typical specificity of miRNAs depending on the kingdom of belonging. Often miRNA genes remain the same between species and kingdoms throughout evolution; however, it’s possible that their nucleotide sequences may change and which makes interspecies influence difficult. However, recent studies show that many human miRNAs share identical sequences with miRNAs from different species, demonstrating a possible influence on humans [18,71,72]. Nevertheless, a plant-based miR-168a has been detected in human serum and regulated the expression of the mammalian gene low-density lipoprotein receptor adaptor protein 1 (LDLRAP1) involved in the LDL-cholesterol metabolism in liver [13,73]. Another example is miR-29b found in cow's milk, which promotes bone health by negatively regulating osteoclast differentiation and positively regulating osteoblast differentiation in humans by increasing RUNX2 activity [17]. In human the food-derived miRNAs, to influence gene expression, must remain unmodified until it reaches their final destination. Studies of plant-derived foods have shown that miR-168 found in soybean and miR-166 found in rice, remained stable after processing and cooking [74,75]. Their stability depends on several factors one in particular is that may be secreted encapsulated in exosomes, which gives resistance to degradation [70]. The exosomes have special membrane characteristics that can withstand harsh conditions in the extracellular environment such as hypoxia, hypermetabolic, enzymatic and acidic environments such as that found in the stomach. This characteristic is probably due to the lipid composition of the membrane, which can change in composition based on pH modification to determine an asymmetric distribution of phosphatidylethanolamine that allows a change in lipid composition between the two membrane layers [76]. These characteristics could allow for instance vegetable miRNAs to resist the digestive phase. The exosomal miRNAs from soy, rice and milk, demonstrated their resistance to saliva, gastric, biliary and pancreatic juice digestion, thus being able to cross even the intestinal barrier [70,77]. This is impressive since the transferred miRNAs can influence target cells by actively participating in intracellular events [21]. As regards the intestinal permeability and its role in the absorption of exogenous miRNAs, there are many evidences to support the hypothesis that the microbiota plays a key role in this process and, in turn, the intestine modulates the bioavailability of nutrients [19]. Is already known that there is variability among individuals in response to physiological and pathological conditions based on age, sex, environmental factors, genetics, diet and so on. Some studies argue that there may be an interaction between miRNAs carried by extracellular microvesicles released for instance by plants and the gut microbiota and there are several supporting hypotheses that try to explain what happens. The first one point out that plant miRNAs might regulate intestinal permeability by regulating the microbiota gene expressions as a result of interaction with different proteins of bacteria; another one, highlights there are different bacterial mRNA targets, that facilitate their absorption from intestine [78,79]. A study performed on the plant-derived microRNA absorptions, found a peak within 3 to 6 hours in serum and tissues, indicating that the gastrointestinal environment is responsible for their absorption. In addition, it seems they can survive for about 36 hours in the tissues and be subsequently metabolized in a similar way to endogenous miRNAs [12,80]. Exogenous miRNAs can regulate the functionality of specific bacteria in the microbiome and have an important impact on an individual's health; it seems that, for instance, corn miRNAs can decrease the concentration of Firmicutes (an intestinal bacterium), which is correlated with an increased risk of obesity and metabolic diseases or the amount of A. Muciniphila, which increases the risk of cardiovascular disease and diabetes [20]. Interesting results emerge from studies by Chen et al, suggesting that plant exosome-like nanoparticles from the ginger rhizome, could be taken up by immune system cells (particularly macrophages) and exert immunomodulatory effects by inhibiting inflammasome activation whose activity would be associated with increased intestinal permeability and progression of diseases such as obesity [81]. Recently, a study by Cao et al. [82] also showed that ginseng-derived nanoparticles led to a reduction in melanoma tumour growth in mice by targeting macrophages and suppressing the switching of the pro-inflammatory M1 phenotype to anti-inflammatory M2 [83].
Zhao et al. suggested the reason that would hinder the detection of plant-derived miRNAs in animals would be the sequence of the miRNAs. Not all miRNAs seem to be able to be taken up, but that selective uptake would occur based on their sequence; determining factors would be, for example, high GC content, a short length and the "CAG" motif, which could promote the uptake and delivery of these exogenous miRNAs [84].
7. Conclusions
Gene activity is influenced by various external stimuli through complex mechanisms that depend on specific transcription factors and the expression of non-coding RNAs whose expression is influenced by exercise and nutrition [1,6,9,10,24,33]. In recent years, a study has also focused on understanding their influence on osteogenic processes. Factors required for osteoblast differentiation have been identified, in particular the gene expression of Runt-related transcription factor 2 (RUNX2) [14]. In particular, the miRNA whose activated pathway has been described is miR-21, which appears to play a role in promoting stem cell osteogenesis via the Smad7-Smad1/5/8-RUNX2 and Akt pathway [15,29]. Exercise as a mechanical load, plays an essential role in both the differentiation and mineralisation of osteoblasts and bone density, reducing the risk of osteoporosis [23]. Among different types of physical activity, half marathons have been shown to increase the expression of many miRNAs including miR-21-5p osteoblast differentiation and proliferation [27]. Among the micronutrients, one of the compounds that have been found to have positive effects is syringic acid (SA), a phenolic acid that on mouse mesenchymal stem cells has been shown to induce osteoblast differentiation [62]. In particular, it would appear to perform this function through increased expression of miR-21, which reduces Smad7 activity by inducing increased expression of RUNX2, with a positive effect on osteoblast differentiation [56]. It is advisable to engage in outdoor physical activity to promote vitamin D production, the intake of which is recommended along with vitamin C and silicic acid as they target the same osteogenic markers. Regarding the effect of macronutrients, Kang Gan et al. analysed the effect of high glucose levels on the regulation of osteogenic differentiation of bone marrow mesenchymal stem cells (MSCs). The results showed that increased expression of miR-221-3p and miR-222-3p on the bone tissue of diabetic mice inhibited osteogenic differentiation through activation of the IGF-1/ERK pathway [59]. Regarding the effects of protein, there are not many studies but it would appear that a low-protein diet negatively regulates bone mass [64]; however, there are no studies investigating the epigenetic effect of a high-protein diet. Therefore, exogenous microRNAs belonging to different species may influence human cellular processes through mechanisms that need further investigation to be better understood [12,13].
In conclusion, a correct lifestyle that includes moderate physical activity and a balanced diet, avoiding obesity conditions, are essential for prevention of bone metabolism-related diseases because they act against abnormal epigenetic regulation. What seems clear to optimise the regenerative potential of BMSCs is to avoid low protein intake, except sulphur amino acids, and high fatty acids and carbohydrates which negatively affect bone mass. To improve bone health, further studies looking at modulation of bone epigenetic biomarkers due to protocols of prevention or effective alternative therapies are necessary.
Author Contributions
Conceptualization and writing, P.P., C.P. and C.R.; original draft preparation, P.P., and C.P.; review and editing, A.A. (Alessandra Amato), A.A. (Anna Alioto), S.B and S.K. All authors have contributed to the article. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledges
No acknowledges
Conflicts of Interest
authors declare no conflict of interest.
References
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9. [Google Scholar] [CrossRef]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions--beyond repression of gene expression. Nature reviews. Genetics 2014, 15, 599–612. [Google Scholar] [CrossRef]
- Schiera, G.; Contrò, V.; Sacco, A.; Macchiarella, A.; Cieszczyk, P.; Proia, P. From epigenetics to anti-doping application: a new tool of detection. Human Movement 2017, 18, 3–10. [Google Scholar] [CrossRef]
- Folwarczna, J.; Zych, M.; Burczyk, J.; Trzeciak, H.; Trzeciak, H.I. Effects of natural phenolic acids on the skeletal system of ovariectomized rats. Planta medica 2009, 75, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Lisse, T.S.; Adams, J.S.; Hewison, M. Vitamin D and microRNAs in bone. Critical reviews in eukaryotic gene expression 2013, 23, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Beckett, E.L.; Yates, Z.; Veysey, M.; Duesing, K.; Lucock, M. The role of vitamins and minerals in modulating the expression of microRNA. Nutrition research reviews 2014, 27, 94–106. [Google Scholar] [CrossRef]
- Arumugam, B.; Balagangadharan, K.; Selvamurugan, N. Syringic acid, a phenolic acid, promotes osteoblast differentiation by stimulation of Runx2 expression and targeting of Smad7 by miR-21 in mouse mesenchymal stem cells. Journal of cell communication and signaling 2018, 12, 561–573. [Google Scholar] [CrossRef]
- Kolhe, R.; Mondal, A.K.; Pundkar, C.; Periyasamy-Thandavan, S.; Mendhe, B.; Hunter, M.; Isales, C.M.; Hill, W.D.; Hamrick, M.W.; Fulzele, S. Modulation of miRNAs by Vitamin C in Human Bone Marrow Stromal Cells. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical Exercise Modulates miR-21-5p, miR-129-5p, miR-378-5p, and miR-188-5p Expression in Progenitor Cells Promoting Osteogenesis. Cells 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.M.H. Macronutrient modulation of mRNA and microRNA function in animals: A review. Animal Nutrition 2020, 6, 258–268. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Ma, W.; Jiao, G.; Zhang, L.; Zhou, H.; Wu, W.; Wang, H.; Chen, Y. Ortho-silicic acid enhances osteogenesis of osteoblasts through the upregulation of miR-130b which directly targets PTEN. Life Sciences 2021, 264, 118680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, T.; Yin, Y.; Zhang, C.Y.; Zhang, Y.L. Dietary microRNA-A Novel Functional Component of Food. Advances in nutrition (Bethesda, Md.) 2019, 10, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Research 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Vimalraj, S.; Arumugam, B.; Miranda, P.J.; Selvamurugan, N. Runx2: Structure, function, and phosphorylation in osteoblast differentiation. International journal of biological macromolecules 2015, 78, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Li, X.; Zhang, F.; Tewari, N.; Ma, R.; Zhong, L.; Makishima, M.; Liu, Y.; Bhawal, U.K. MicroRNA-21 facilitates osteoblast activity. Biochemistry and biophysics reports 2021, 25, 100894. [Google Scholar] [CrossRef] [PubMed]
- Smieszek, A.; Marcinkowska, K.; Pielok, A.; Sikora, M.; Valihrach, L.; Marycz, K. The Role of miR-21 in Osteoblasts-Osteoclasts Coupling In Vitro. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hassan, M.Q.; Jafferji, M.; Aqeilan, R.I.; Garzon, R.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Biological functions of miR-29b contribute to positive regulation of osteoblast differentiation. The Journal of biological chemistry 2009, 284, 15676–15684. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Chiang, K.; Zempleni, J.; Cui, J. Computational Characterization of Exogenous MicroRNAs that Can Be Transferred into Human Circulation. PloS one 2015, 10, e0140587. [Google Scholar] [CrossRef] [PubMed]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Frontiers in nutrition 2021, 8, 586564. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Pham, Q.; Davis, C.D.; Yu, L.; Wang, T.T.Y. Delineating effect of corn microRNAs and matrix, ingested as whole food, on gut microbiota in a rodent model. Food science & nutrition 2020, 8, 4066–4077. [Google Scholar] [CrossRef]
- Di Liegro, C.M.; Schiera, G.; Di Liegro, I. Extracellular vesicle-associated RNA as a carrier of epigenetic information. Genes 2017, 8, 240. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Satoh, M.; Pauley, K.M.; Fritzler, M.J.; Reeves, W.H.; Chan, E.K. Detection of the argonaute protein Ago2 and microRNAs in the RNA induced silencing complex (RISC) using a monoclonal antibody. Journal of immunological methods 2006, 317, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.D.F.; Oliveira, M.L.d.; Lirani-Galvão, A.P.; Marin-Mio, R.V.; Santos, R.N.d.; Lazaretti-Castro, M. Physical exercise and osteoporosis: effects of different types of exercises on bone and physical function of postmenopausal women. Arquivos Brasileiros de Endocrinologia & Metabologia 2014, 58, 514–522. [Google Scholar] [CrossRef]
- Amato, A.; Baldassano, S.; Cortis, C.; Cooper, J.; Proia, P.J.H.M. Physical activity, nutrition, and bone health. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Rossi, M.; Pitari, M.R.; Amodio, N.; Di Martino, M.T.; Conforti, F.; Leone, E.; Botta, C.; Paolino, F.M.; Del Giudice, T.; Iuliano, E.; et al. miR-29b negatively regulates human osteoclastic cell differentiation and function: implications for the treatment of multiple myeloma-related bone disease. Journal of cellular physiology 2013, 228, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Chen, X.; Zhang, S.; Huang, M.; Shen, X.; Xu, J.; Zou, J. The Effect of Exercise on the Prevention of Osteoporosis and Bone Angiogenesis. BioMed research international 2019, 2019, 8171897. [Google Scholar] [CrossRef] [PubMed]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nature reviews Molecular cell biology 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, Z.; Chen, Y. Regulation of TGF-beta signaling by Smad7. Acta biochimica et biophysica Sinica 2009, 41, 263–272. [Google Scholar] [CrossRef]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical exercise modulates miR-21-5p, miR-129-5p, miR-378-5p, and miR-188-5p expression in progenitor cells promoting osteogenesis. Cells 2019, 8, 742. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bruxvoort, K.J.; Zylstra, C.R.; Liu, J.; Cichowski, R.; Faugere, M.C.; Bouxsein, M.L.; Wan, C.; Williams, B.O.; Clemens, T.L. Lifelong accumulation of bone in mice lacking Pten in osteoblasts. Proceedings of the National Academy of Sciences of the United States of America 2007, 104, 2259–2264. [Google Scholar] [CrossRef] [PubMed]
- Weng, N.-p.; Granger, L.; Hodes, R.J. Telomere lengthening and telomerase activation during human B cell differentiation. Proceedings of the National Academy of Sciences 1997, 94, 10827–10832. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.-S.; Wright, W.E.; Shay, J.W. Human telomerase and its regulation. Microbiology and molecular biology reviews 2002, 66, 407–425. [Google Scholar] [CrossRef] [PubMed]
- Dalle Carbonare, L.; Mottes, M.; Cheri, S.; Deiana, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Salvagno, G.L.; Lippi, G.; Valenti, M.T. Increased Gene Expression of RUNX2 and SOX9 in Mesenchymal Circulating Progenitors Is Associated with Autophagy during Physical Activity. Oxidative medicine and cellular longevity 2019, 2019, 8426259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.Y.; Li, C.; Bai, W.D.; Su, L.L.; Liu, J.Q.; Li, Y.; Shi, J.H.; Cai, W.X.; Bai, X.Z.; Jia, Y.H.; et al. MicroRNA-21 regulates hTERT via PTEN in hypertrophic scar fibroblasts. PloS one 2014, 9, e97114. [Google Scholar] [CrossRef]
- Ahmed, S.; Passos, J.F.; Birket, M.J.; Beckmann, T.; Brings, S.; Peters, H.; Birch-Machin, M.A.; von Zglinicki, T.; Saretzki, G. Telomerase does not counteract telomere shortening but protects mitochondrial function under oxidative stress. Journal of cell science 2008, 121, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M. Biological origin of free radicals, and mechanisms of antioxidant protection. Chemico-biological interactions 1994, 91, 133–140. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: the Fenton reaction. Toxicology letters 1995, 82, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Rossbach, M. Small non-coding RNAs as novel therapeutics. Current molecular medicine 2010, 10, 361–368. [Google Scholar] [CrossRef]
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.-M.; Xu, Y.-M.; Huang, L.-F.; Wang, X.-Z. Exosomes: novel biomarkers for clinical diagnosis. The scientific world journal 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proceedings of the National Academy of Sciences 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic acids research 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Xiao, Z.S. Advances in Runx2 regulation and its isoforms. Medical hypotheses 2007, 68, 169–175. [Google Scholar] [CrossRef]
- Cheng, V.K.F.; Au, P.C.M.; Tan, K.C.; Cheung, C.L. MicroRNA and human bone health. JBMR plus 2019, 3, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Carrozza, M.J.; Utley, R.T.; Workman, J.L.; Cote, J. The diverse functions of histone acetyltransferase complexes. TRENDS in Genetics 2003, 19, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Fan, G. The role of DNA methylation in the central nervous system and neuropsychiatric disorders. International review of neurobiology 2009, 89, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Raisz, L.G. Pathogenesis of osteoporosis: concepts, conflicts, and prospects. The Journal of clinical investigation 2005, 115, 3318–3325. [Google Scholar] [CrossRef]
- Nilsson, S.; Makela, S.; Treuter, E.; Tujague, M.; Thomsen, J.; Andersson, G.r.; Enmark, E.; Pettersson, K.; Warner, M.; Gustafsson, J.-Å. Mechanisms of estrogen action. Physiological reviews 2001, 81, 1535–1565. [Google Scholar] [CrossRef]
- Finnegan, E.F.; Pasquinelli, A.E. MicroRNA biogenesis: regulating the regulators. Critical reviews in biochemistry and molecular biology 2013, 48, 51–68. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Nutrition, N.F.; Allergens, F.; Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Knutsen, H.K.; Maciuk, A.; Mangelsdorf, I.; et al. Guidance for establishing and applying tolerable upper intake levels for vitamins and essential minerals. 2022, 20, e200102. [Google Scholar] [CrossRef]
- Wu, J.; Liang, J.; Li, M.; Lin, M.; Mai, L.; Huang, X.; Liang, J.; Hu, Y.; Huang, Y. Modulation of miRNAs by vitamin C in H2O2-exposed human umbilical vein endothelial cells. International Journal of Molecular Medicine 2020, 46, 2150–2160. [Google Scholar] [CrossRef] [PubMed]
- Kolhe, R.; Mondal, A.K.; Pundkar, C.; Periyasamy-Thandavan, S.; Mendhe, B.; Hunter, M.; Isales, C.M.; Hill, W.D.; Hamrick, M.W.; Fulzele, S. Modulation of miRNAs by vitamin C in human bone marrow stromal cells. Nutrients 2018, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Abak, A.; Shoorei, H.; Mohaqiq, M.; Majidpoor, J.; Sayad, A.; Taheri, M. Regulatory role of microRNAs on PTEN signaling. Biomedicine & Pharmacotherapy 2021, 133, 110986. [Google Scholar] [CrossRef]
- Yang, L.; Li, C.; Liang, F.; Fan, Y.; Zhang, S. MiRNA-155 promotes proliferation by targeting caudal-type homeobox 1 (CDX1) in glioma cells. Biomedicine & Pharmacotherapy 2017, 95, 1759–1764. [Google Scholar] [CrossRef]
- Lai, Y.L.; Yamaguchi, M. Phytocomponent p-hydroxycinnamic acid stimulates bone formation and inhibits bone resorption in rat femoral tissues in vitro. Molecular and cellular biochemistry 2006, 292, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.L.; Yamaguchi, M. Oral administration of phytocomponent p-hydroxycinnamic acid has anabolic effects on bone calcification in femoral tissues of rats in vivo. Journal of health science 2006, 52, 308–312. [Google Scholar] [CrossRef]
- Raut, N.; Wicks, S.M.; Lawal, T.O.; Mahady, G.B. Epigenetic regulation of bone remodeling by natural compounds. Pharmacological research 2019, 147, 104350. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Liang, H.L.; Hung, C.H.; Kuo, P.L. Syringetin, a flavonoid derivative in grape and wine, induces human osteoblast differentiation through bone morphogenetic protein-2/extracellular signal-regulated kinase 1/2 pathway. Molecular nutrition & food research 2009, 53, 1452–1461. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.; Kumar, C.S. Syringic acid (SA)‒a review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomedicine & Pharmacotherapy 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Gan, K.; Dong, G.H.; Wang, N.; Zhu, J.F. miR-221-3p and miR-222-3p downregulation promoted osteogenic differentiation of bone marrow mesenchyme stem cells through IGF-1/ERK pathway under high glucose condition. Diabetes research and clinical practice 2020, 167, 108121. [Google Scholar] [CrossRef] [PubMed]
- Qu, B.; Gong, K.; Yang, H.-S.; Li, Y.-G.; Jiang, T.; Zeng, Z.-M.; Cao, Z.-R.; Pan, X.-M. MiR-449 overexpression inhibits osteogenic differentiation of bone marrow mesenchymal stem cells via suppressing Sirt1/Fra-1 pathway in high glucose and free fatty acids microenvironment. Biochemical and biophysical research communications 2018, 496, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yang, C.; Tan, J.; Dou, C.; Chen, Y. Modulation of SIRT6 activity acts as an emerging therapeutic implication for pathological disorders in the skeletal system. Genes & Diseases 2022. [Google Scholar] [CrossRef]
- Qu, B.; Gong, K.; Yang, H.S.; Li, Y.G.; Jiang, T.; Zeng, Z.M.; Cao, Z.R.; Pan, X.M. MiR-449 overexpression inhibits osteogenic differentiation of bone marrow mesenchymal stem cells via suppressing Sirt1/Fra-1 pathway in high glucose and free fatty acids microenvironment. Biochemical and biophysical research communications 2018, 496, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Role of Neonatal Dietary Ca in Bone Development and Characteristics of Porcine Mesenchymal Stem Cells; North Carolina State University: 2014.
- Ellur, G.; Sukhdeo, S.V.; Khan, M.T.; Sharan, K. Maternal high protein-diet programs impairment of offspring's bone mass through miR-24-1-5p mediated targeting of SMAD5 in osteoblasts. Cellular and molecular life sciences : CMLS 2021, 78, 1729–1744. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, I.; Alameddine, M.; Scalabrin, M.; van 't Hof, R.J.; Liloglou, T.; Ozanne, S.E.; Goljanek-Whysall, K.; Vasilaki, A. Low protein intake during reproduction compromises the recovery of lactation-induced bone loss in female mouse dams without affecting skeletal muscles. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020, 34, 11844–11859. [Google Scholar] [CrossRef] [PubMed]
- Proia, P.; Amato, A.; Drid, P.; Korovljev, D.; Vasto, S.; Baldassano, S. The impact of diet and physical activity on bone health in children and adolescents. Frontiers in Endocrinology 2021, 12, 704647. [Google Scholar] [CrossRef]
- Remer, T.; Manz, F. Estimation of the renal net acid excretion by adults consuming diets containing variable amounts of protein. The American journal of clinical nutrition 1994, 59, 1356–1361. [Google Scholar] [CrossRef]
- Baldassano, S.; Alioto, A.; Amato, A.; Rossi, C.; Messina, G.; Bruno, M.R.; Stallone, R.; Proia, P. Fighting the Consequences of the COVID-19 Pandemic: Mindfulness, Exercise, and Nutrition Practices to Reduce Eating Disorders and Promote Sustainability. Sustainability 2023, 15, 2120. [Google Scholar] [CrossRef]
- Conigrave, A.D.; Brown, E.M.; Rizzoli, R. Dietary protein and bone health: roles of amino acid-sensing receptors in the control of calcium metabolism and bone homeostasis. Annual review of nutrition 2008, 28, 131–155. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.; Ferro, V.A.; Tate, R.J. Determination of the potential bioavailability of plant microRNAs using a simulated human digestion process. Molecular nutrition & food research 2015, 59, 1962–1972. [Google Scholar] [CrossRef]
- Saunders, M.A.; Liang, H.; Li, W.-H. Human polymorphism at microRNAs and microRNA target sites. Proceedings of the National Academy of Sciences 2007, 104, 3300–3305. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Pal Bhadra, M.; Girschick, H.J.; Bhadra, U. MicroRNAs–micro in size but macro in function. The FEBS journal 2008, 275, 4929–4944. [Google Scholar] [CrossRef] [PubMed]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. The Journal of nutrition 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.; Ferro, V.A.; Tate, R.J. Determination of the potential bioavailability of plant microRNAs using a simulated human digestion process. Molecular nutrition & food research 2015, 59, 1962–1972. [Google Scholar] [CrossRef]
- Xie, W.; Melzig, M.F. The stability of medicinal plant microRNAs in the herb preparation process. Molecules 2018, 23, 919. [Google Scholar] [CrossRef] [PubMed]
- Askenase, P.W. Exosomes provide unappreciated carrier effects that assist transfers of their miRNAs to targeted cells; I. They are 'The Elephant in the Room'. RNA biology 2021, 18, 2038–2053. [Google Scholar] [CrossRef]
- Rani, P.; Vashisht, M.; Golla, N.; Shandilya, S.; Onteru, S.K.; Singh, D. Milk miRNAs encapsulated in exosomes are stable to human digestion and permeable to intestinal barrier in vitro. Journal of Functional Foods 2017, 34, 431–439. [Google Scholar] [CrossRef]
- Horne, R.; St. Pierre, J.; Odeh, S.; Surette, M.; Foster, J.A. Microbe and host interaction in gastrointestinal homeostasis. Psychopharmacology 2019, 236, 1623–1640. [CrossRef] [PubMed]
- Peck, B.C.; Mah, A.T.; Pitman, W.A.; Ding, S.; Lund, P.K.; Sethupathy, P. Functional transcriptomics in diverse intestinal epithelial cell types reveals robust microRNA sensitivity in intestinal stem cells to microbial status. Journal of Biological Chemistry 2017, 292, 2586–2600. [Google Scholar] [CrossRef]
- Liang, G.; Zhu, Y.; Sun, B.; Shao, Y.; Jing, A.; Wang, J.; Xiao, Z. Assessing the survival of exogenous plant microRNA in mice. Food science & nutrition 2014, 2, 380–388. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Y.; Yu, J. Exosome-like nanoparticles from ginger rhizomes inhibited NLRP3 inflammasome activation. Molecular pharmaceutics 2019, 16, 2690–2699. [Google Scholar] [CrossRef]
- Cao, M.; Yan, H.; Han, X.; Weng, L.; Wei, Q.; Sun, X.; Lu, W.; Wei, Q.; Ye, J.; Cai, X. Ginseng-derived nanoparticles alter macrophage polarization to inhibit melanoma growth. Journal for immunotherapy of cancer 2019, 7, 1–18. [Google Scholar] [CrossRef]
- Kim, H.; Wang, S.Y.; Kwak, G.; Yang, Y.; Kwon, I.C.; Kim, S.H. Exosome-guided phenotypic switch of M1 to M2 macrophages for cutaneous wound healing. Advanced science 2019, 6, 1900513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Mao, Q.; Zhao, Z.; Dou, T.; Wang, Z.; Cui, X.; Liu, Y.; Fan, X. Prediction of plant-derived xenomiRs from plant miRNA sequences using random forest and one-dimensional convolutional neural network models. BMC genomics 2018, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



