
Submitted:
02 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
Anthracnose and root rot are major foliar and root diseases of strawberry, respectively that cause most yield and quality losses. Anthracnose is caused by multiple species belonging to Colletotrichum spp complex. This disease alone can causeup to 70% yield loss in North America. Colletotrichum spp. cause several disease symptoms on strawberries, including root, fruit, and crown rot, lesions on petioles and runners, and irregular black spots on the leaf. In many cases, a lower level of infection on foliage remains non-symptomatic (quiescent), posing a challenge to growers as these plants can be a significant source of inoculum for the fruiting field. Reliable detection methods for quiescent infection should play an important role in preventing infected plants' entry into the production system or guiding growers to take appropriate preventative measures to control the disease. The selection of highly effective fungicides and including them in a schedule to prevent the development of fungicide resistance in fungal populations will remain an area of continued research. Disease management methods should entail sensitive molecular testing of suspected fungal isolates for resistance, rotation of products, and monitoring of quiescent infections in transplant materials. Testing and the inclusion of non-chemical methods such as biologicals and biorational treatments especially for soilborne pathogens will also be necessary to make disease management economically feasible and sustainable.
Keywords:
Anthracnose fruit rot
; Colletotrichum acutatum
; Diagnostics
; crop protection
; sustainable management techniques
; strawberry
1. Introduction
Strawberry (Fragaria× ananassa Duchesne), a major small fruit across the world, has an attractive flavor, taste and high content of essential nutrients that benefit human health [1]. The Food and Agriculture Organization of the United Nations (FAO) reports that strawberries were planted on 389,665 hectares and produced a total of 917,5384.43 metric tons globally in 2021 [2]. The United States of America is among the countries with the highest strawberry production and plants approximately 58,070 acres of strawberries, with a value of US$2.4 billion [3]. In addition, the south-Atlantic region of the United States produces 2,342 acres of strawberries, with an average yield of 13,979 lb/acre and a total farm gate value of $47,158,000 [4]. Strawberry consumption in the United States has grown tremendously over the last two decades, from 2 lb per capita in 1980 to 8 lbin 2013 [5]. However, diseases and pests significantly lower the quality and production of strawberry fruit incurring significant financial losses to growers.
The mid-Atlantic region of the United States, including the Commonwealth of Virginia, ranks third in the production of fresh market strawberries after California and Florida, and most growers are using the annual hill plasticulture (AHP) production system [4] likely due to the disease risk from the build-up of inoculum in a perennial system. Field preparation begins with debris removal, disking and tilling the soil followed by bedding, plastic covering, drip tape installation, and overhead watering in September. The raised soil beds in most cases are fumigated to control nematodes, fungi, and weeds. However, due to new regulations, fumigation with synthetic chemicals is becoming difficult for small growers and farms close to public places. Then, the transplants are planted and overhead watered for establishment between mid-September and early October, when temperatures typically reach above 30 degrees Celsius [6,7]. Strawberry fruit is -harvested two to three times each week on average when the berries are fully ripened from mid-April until late June. The strawberry plant can be infected by different organisms, including many arthropods, nematodes, fungi, bacteria, virus, and other pests [8]. Further, the strawberry plant is highly susceptible to a large variety of soilborne pathogens, including genera of Verticillium, Phytophthora, and Colletotrichum spp., which are considered the most damaging pathogens on this crop in the United States strawberry production system [8,9]. Anthracnose in general caused by fungal pathogens belonging to multiple species under C. acutatum and C. gloeosporioides species complex. All of these pathogens can cause infection on any parts of strawberry, however, C. acutatum species complex tend to be more destructive as fruit rot pathogens, and C. gloeosporioides species complex is more damaging as crown rot pathogens. Considering the overall importance and frequency of occurrence on the U.S. strawberries, this review highlights C. acutatum species complex and mentioned as C. acutatum.
In the strawberry industry, strawberry transplants are grown mainly by using starter plant material i.e. runner tips that come from mother plants located in northern latitudes, such as Canada, or mother plants that are located in higher altitudes in the U.S., such as in the mountain region of North Carolina and California, which have relatively cooler temperatures. Low temperatures may slow disease progression but do not eliminate the pathogen from strawberry transplants [10]. These tips are then rooted in propagation houses in July through September to generate strawberry plug plants at various locations in the U.S. Pathogens such as those under C. acutatum species complex may be transferred to freshly formed plants since transplants are propagated vegetatively [11]. In the propagation house, the warm temperatures and humid environment make a favorable environment for the moderate to high temperature loving pathogen to thrive. When the infected transplants arrive in strawberry-producing fields, plants are still subjected to ideal environmental conditions for anthracnose disease development, such as extended wetness periods and temperatures between 20 and 30°C [12]. Therefore, new technology for detecting C. acutatum in asymptomatic nursery plants would be appreciable in the North American Strawberry Nursery System to minimize the potential of selling transplants latently infected with Colletotrichum to fruit growers [13]. The aim of this review is to highlight the cutting edge technologies forthe detecting the latent presence of Colletotrichum on strawberry and its role in the management of the diseases. It also includes exciting new sustainable management techniques for root rot especially in organic strawberry system.
2. Anthracnose diseases in strawberry
Anthracnose, described as a disease that shows as black, sunken lesions on stems, runners or fruit and is caused by fungi that generate asexual spores in acervuli, is derived from the Greek roots "anthrak-" (coal) and "-nosos" (disease). Anthracnose fruit rot (AFR) disease on strawberries is caused by multiple strains belonging to Colletotrichum spp. including, C. acutatum species complex (including 31 species of Colletotrichum) [14,15,16]. In United States, C. fragariae was believed to be the primary agent of strawberry anthracnose until Smith and Black (1986) reported C. acutatum on strawberries [17]. These diseases are among the most dangerous, causing significant crop losses of up to 70% in commercial production fields planted with susceptible strawberry cultivars [12,18]. Colletotrichum spp. have a wide host range and cause several disease symptomsin strawberry, including root, fruits, and crown rot, lesions on petioles and runner, and irregular spots on the leaf [19]. C. acutatum can infect all parts of the strawberry plant (leaves, petioles, flowers, crowns, and roots) along with fruit, and lesions may expand and entirely cover the surface of the fruit under favorable conditions (high temperature and humidity), especially on susceptible varieties [20,21].
3. Taxonomy of Colletotrichum acutatum
The genus of Colletotrichum spp. consists of more than 189 species ordered into at least 11 major phylogenetic lineages, also known as species complexes [22,23]. The fungus Colletotrichum acutatum, taxonomically belongs to: Fungi, Ascomycota, Pezizomycotina, Sordariomycetes, Hypocreomycetidae, Glomerellales, Glomerellaceae, Colletotrichum, acutatum as reported by Simmonds (1965) and described it as a distinct species in Queensland, Australia, in a pathogen survey of fruit rot [24]. The pathogen was previously identified as a species of Gloeosporium that affected strawberries in Australia, causing mature fruit rot and lesions on the stolon, petiole, and peduncle [25]. Using rDNA-ITS and β-tubulin 2 (TUB2) sequence analyses, there were 34 species cluster in five clades that are accepted under the C. acutatum species complex [26,27,28] (Figure 1). According to Ulrike Damm (2012), many taxa that von Arx (1957) had thought were synonyms of C. gloeosporioides belong to the C. acutatum species complex, including C. mahoniae, C. godetiae, Gloeosporium phormii, Gloeosporium lycopersici , and Gloeosporium [15] (Fig. 1). In recent years, seven species of the C. acutatum complex including C. acutatum s.s., C. fioriniae, C. godetiae, C. miaoliense, C. nymphaeae, C. salicis, and C. simmondsii have been accepted as strawberry pathogens [29].
4. Epidemiology and lifestyle of Colletotrichum acutatum
C. acutatum inoculum in annual strawberry fields is derived from symptomless infected transplants from nurseries where overhead irrigation might have resulted in conidial dissemination [30]. Under warm temperatures and high humidity, this pathogen rapidly produces conidia, which can spread to flowers and fruit by splashing water, contaminated soil on farming equipment, harvesting operations, and consequently show the symptom from the infections by the pathogen. Several studies showed that the conidia dispersal occur within a 25 cm radius of the inoculum source and may vary depending on rainfall intensity and ground cover [31]. This fungus is considered to be a hemibiotrophic where the fungus initially enters a biotrophic phase and then switches to a necrotrophic phase [32]. The pathogen penetrates the cuticle via a specialized cell called an appressorium and grows within the cuticle and cell walls of epidermal, subepidermal, and subtending cells. Then, the fungus produces acervuli as a stroma immediately under the outer periclinal epidermal walls when the cortical tissue is substantially disturbed, and the conidia are released from these acervuli. Although C. acutatum may produce quiescent infections on strawberry plants, according to several studies; however, the epidemiological significance of latent infection before the colonization of fruits and senescing foliage has not been investigated [33].
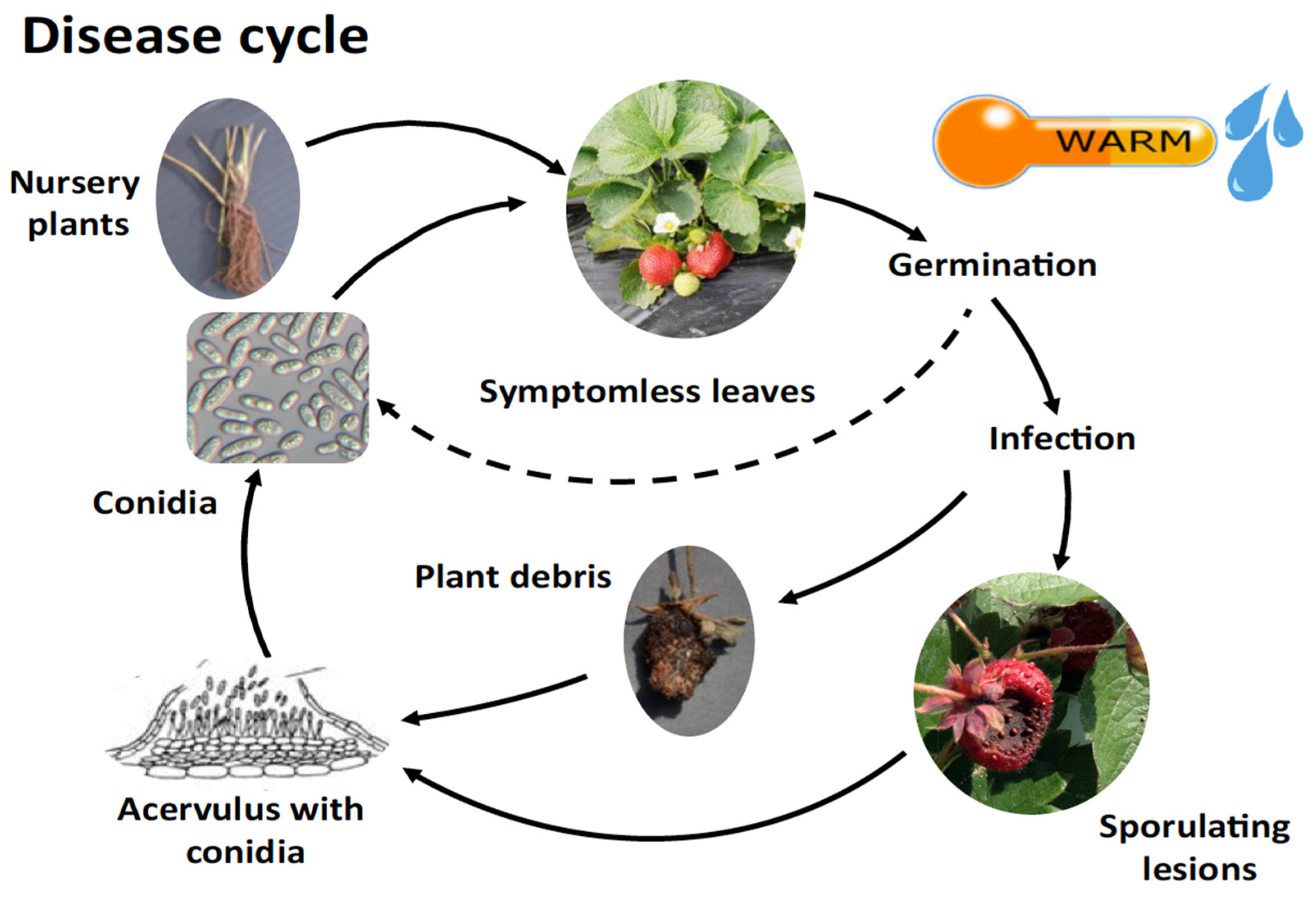
The quiescent stage (the latent period) is considered to be the time between fungal infection of the host and the first symptom appearance [34]. The latent period depends on temperature and is between 2-3 days at 25 ºC and 6-17 days at 5 ºC [35]. The germination of C. acutatum conidia, development of appressoria, latency on non-targeted vegetative organs, including leaves, serving as a source of overwintering inoculum is shown in Figure 2. In comparison to C. fragariae or C. gloeosporioides, C. acutatum can produce more conidia at lower temperatures and show the shortest latent period among the three species at 5 and 10 °C. The concentration of inoculum present as latent infections on strawberry plants determines the beginning of disease symptoms. Knowing the minimum inoculum concentration for the start of a disease is informative , particularly in the case of strawberry AFR as the disease begins as infected transplants from nurseries in commercial fields [36]; therefore, the development of new tools for detection of the latent infection at transplants stage in the nurseries is a key factor to control AFR diseases on strawberries in fruiting fields.
The primary source of inoculum for fruit infections may be C. acutatum appressoria and secondary conidia, which are formed on symptomless leaves and help keep inoculum available throughout the growing season. Wetness for more than 4 h is required for the secondary conidia production and appressoria formation [32]. In general, Colletotrichum spp. conidia are transmitted from plant to plant in the field primarily by rain splash. Conidia of C. acutatum are disseminated over short distances on low-growing crops such as strawberries, and using straw as a ground cover can drastically reduce conidia dispersal [37,38]. Further, the conidia of the three Colletotrichum spp. were evaluated with water splash, and it was discovered that conidia of C. fragariae dispersed over the shortest distance while those of C. acutatum spread over the longest distance due to high production of spores [39]. Although C. acutatum can survive in soil and on inanimate surfaces for various lengths duration depending on the conditions, it appears to compete poorly as a saprophyte [40]. The fungus may survive in the soil for at least two winters with temperatures below 0 °C, causing anthracnose to grow in the following years [41]. Therefore, the application of appropriate disease treatment is required not just for the current year's crop, but also for subsequent seasons. Additionally, it has been proven that weeds host the strawberry infection caused by C. acutatum. Nevertheless, a multicrop study conducted in Florida found that C. acutatum isolates can be host-specific and offer little threat to other crops [42].
5. Detection-morphological, molecular, and remote sensing
Transplants infected with Colletotrichum spp. can spread pathogen from the nursery to the field and the best management option to avoid the disease is, to begin with disease-free planting materials (Han et al., 2016). However, there is no reliable diagnostic method-based protocol to detect latent infection of anthracnose diseases in the early stages of production cycle in large production area. Currently, the production of the disease-free transplants in the nurseries relies on scouting for symptomatic plants and the identification of the diseases using colony and spore morphology, which is time-consuming, error-prone, and sometimes inaccurate [41,43]. Molecular techniques such as polymerase chain reaction (PCR), real-time PCR, and ELISA using DNA have become a robust detection and diagnostic tool for plant pathogens (Aljawasim & Vincelli, 2015; Freeman & Rodriguez, 1995; Martinez-Culebras et al., 2003). However, the polymerase chain reaction (PCR) has some challenges such as presence of PCR inhibitors in plant tissues, failure to amplify low DNA concentration, and detection of dead pathogens, which can give false positive. Further, all molecular techniques for diagnosis of plant pathogens are costly, time-consuming, and need the use of highly skilled human resources (Fazari et al., 2021).
Visual inspection, along with field sampling of plant material is the conventional way of detecting the infections in the field, but it is tedious and requires specialized skills [44,45]. The traditional methods can not detect the latent infection in early stage of infection. Other laboratory analyses, such as microscopy, molecular, biochemical, and microbiological methods have been applied for detection of crop diseases; however, these techniques have disadvantages, as the sampling process is destructive and offers limited diagnostic points, and it is not field-scalable or may not represent accurate field variability [44,46]. Therefore, precise, high-throughput, non-invasive, and field-scalable approaches are required [44,47]. As an alternative in recent years, non-destructive methods such as spectral vegetation indexing, multispectral imaging (MSI) or hyperspectral imaging (HSI) based on ground, aerial, and satellite platforms have emerged that are capable of crop disease diagnostics at high accuracy and high spatial scales (from leaf to plant to field). Additionally, MSI and HSI could also offer early detection even before the visual symptoms develop. Such detection can help with proactive management of anthracnose, thereby improving productivity [48].
5.1. Remote Sensing of Anthracnose
Remote sensing (RS) with MSI and HSI systems has demonstrated its ability for spatiotemporal vegetation monitoring including detection of crop diseases in early stages [49]. During the disease infection process, MSI captures light reflected from the surface of the target object such as leaf, which is dependent on both physiological and biological status, which is prone to variation during the disease infection process, e.g., alternations in plant pigmentations such as chlorophyll and carotenoids [48]. Since anthracnose causes physiological, morphological, and plant pigmentation variations, the MSI and HSI techniques could be useful in estimating its incidence. [48]. Remote sensing such as hyperspectral imaging, was used to detect anthracnose diseases on tea plants with a detection accuracy of 98% for identifying the disease at the leaf level and 94% at the pixel level where they identified disease-sensitive bands at 542, 686, and 754 nm, which were used to create two disease indices, including the Tea Anthracnose Ratio Index (TARI) and the Tea Anthracnose Normalized Index (TANI) [50].
MSI imaging corresponds to imaging within 3-10 bands of the electromagnetic spectrum in the optical range of 350–1000 nm. Each pixel in the image is represented by a vector, referred to as the spectral signature or fingerprint region of the spectrum [48]. Each fingerprint region of spectrum has complex absorption sequences due to various bending vibrations within molecules of the plant tissue, and a slight alteration in a compound's molecular structure will result in a significant change in spectral absorption [51]. The spectral data is helpful, although it may be redundant for adjacent wavelengths. To minimize data size and enhance data utilization efficiency, only significant wavelengths with essential information should be selected for the application of the multispectral imaging (MSI) to reduce expenses and increase speed of plant disease detection [46]. In the field, the sheath blight (ShB) disease on rice, caused by Rhizoctonia solani, has been detected with high efficiency using multispectral imagining (MSI) where five vegetation indices were then calculated from the multispectral images, including Normalized Difference Vegetation Index (NDVI), Ration Vegetation Index (RVI), Difference Vegetation Index (DVI), Normalized Difference Water Index (NDWI) and Red Edge (RE) [52]. The MSI technique was used to detect the light leaf spot infection with 92% accuracy on oilseed rape (Brassica napus) within 13 days prior to the beginning of visible symptoms, and they used false color mapping of spectral vegetation indices to quantify disease severity and its distribution within the plant canopy in the field [48]. A non-destructive model for evaluation of the firmness, total soluble solids (TSS) content and ripeness stage in strawberry fruit was established with 100% accuracy using multispectral imaging technique [53]. Hyperspectral Imaging (HIS) has been used to detect anthracnose on strawberries and the spectra of disease in symptomless and symptomatic sections of leaves varies significantly at wavelengths ranging from 540-570 nm to 750-900 nm in the laboratory [54]. Several studies used ultraviolet fluorescence (440–740 nm), multispectral (Green [540 nm], red [660 nm], and near-Infrared [800 nm]), RGB, and hyperspectral (900–1700 nm) imaging techniques for detection of crop diseases such as powdery mildew (Erysiphales) on grapevine (Vitis Vinifera) [55].
5.2. Unmanned Aerial Vehicle (UAV) platform
Unmanned aerial vehicles (UAV), typically referred to as drones, have seen extensive applications in the past decade for crop production management. Crop health monitoring is a particularly promising use of UAV since it may enhance conventional crop monitoring methods, including visual observation to assist in rapid detection, which has a significant positive effect on crop yield and quality [56]. UAVs are used with MSI or HSI systems to provide high spatial resolution images at flexible flight schedules and short data-acquisition timeframes [49]. Further, the unmanned aerial vehicle (UAV) based imagery data technology has been used successfully in different applications such as rapid evaluation of crop vigor and soil characteristics, crop water requirements, disease infestation, and yield prediction [57]. Three platforms, including an unmanned aerial vehicle (UAV) and PlanetScope and Sentinel-2 satellite platforms with multispectral (MS) imagery systems, were evaluated based on the analysis of the spatial resolution using soil adjusted vegetation index (SAVI) to monitor onion crops in the field, and the best result was achieved with the images provided by the UAV platform, which could give more detailed images at critical moments in the crop cycle [58]. Small unmanned aerial systems (UAS) equipped with high-resolution visible (red-green-blue [RGB]) and multispectral imaging technique was used to detect the powdery mildew (PM) in apple orchards with 77% accuracy [59]. Using a mobile platform, three algorithms, including the stepwise discriminant analysis (SDA), Fisher discriminant analysis (FDA), and KNN methods with 32 spectral vegetation indices, were applied to train the model to detect anthracnose diseases at different infection stages on strawberries in both indoor and field trails, and the three models' classification accuracies were 71.3%, 70.5%, and 73.6%, respectively [45]. To our knowledge, there has been no research on the detection of latent infection of anthracnose diseases on strawberries in the field by using multispectral imaging technology with UAV.
6. Management: Chemical, biological and biorational
Programs of Integrated Pest Management (IPM) can help producers in combining techniques that focus on the long-term disease and pest management including, usage of the pesticides when required, excellent sanitation practices, the planting of disease-free plants, and cultural controls [60]. Several cultural control methods were used to reduce the C. acutatum infection, such as removing the bloom clusters as they appear on both mother and daughter plants. Another cultural practice is the reduction of standing water irrigation by using drip irrigation instead of overhead irrigation, which help to reduce the movement of the conidia from one plant to another through water splash. Further, hot water therapy has been used for a long time to eradicate pest and disease issues, including cyclamen mites (Phytonemus pallidus ssp. fragariae Zimmerman) and endoparasitic nematodes, in dormant strawberry stock. From mother plants that had been inoculated with C. acutatum, runner cuttings were collected, and they were immersed for 7 minutes at 35°C, followed by 2 or 3 minutes at 50°C. Cuttings infected with C. acutatum were successfully reduced by both treatments from over 80% in the controls to between 6% and 17% [61].
Chemical methods such the usage of fungicides is a common method among growers to control AFR and other strawberry diseases, and frequently relies on a calendar schedule of weekly application [62]. Early in the season, between November and December, inoculum levels are low and typically unfavorable conditions for C. acutatum, therefore infected plants do not exhibit symptoms. During this time, the first step of chemical management includes the use of low label rates of broad-spectrum protectant fungicides like Captan. Then, the inoculum levels increase, and the environment will reach the ideal condition for AFR development from January to March; therefore, higher label rates of broad-spectrum fungicides must be applied weekly depending on the detection of latent infections [62]. In the past, mancozeb, carbendazim, prochloraz, and Tecto 60 have all been used as synthetic fungicides to control the Colletotrichum spp. that causes anthracnose in fruits including strawberry[63,64].
In the last update of 2023 southeast regional strawberry integrated pest management guide focused on plasticulture production, many fungicides are labeled as "Exellent" for controlling AFR of strawberry such as merivon (7+11), pristine (7+11), luna sensation (7+11), quadris top (3+11), quilt xcel (3+11), cabrio (11), abound (11), flint extra (11), and miravis prime (12+7). On other hand, other common fungicides such as captan and thiram were rated a "good" and "fair" respectively [65]. However, to avoid the development of resistance strains of C. acutatum, all fungicides must be used carefully and in rotation using different active chemicals ingredients.
Many biological controls and biofungicides have been evaluated on fruit crops against Colletotrichum spp., but none have consistently demonstrated field efficacy. Several bacterial biocontrol agents such as Bacillus spp. were summarized and evaluated against Colletotrichum spp. such as C. acutatum, C. gloeosporioides, and C. truncatum clades, and they exhibit some inhibitory activities due to the production of antifungal activity via secretion of antifungal metabolites and enzymes or via induction of disease resistance in fruits. [66]. B. subtilis, P. polymyxa, and B. amyloliquefaciens have generally been the best agents for managing C. acutatum. According to reports, Paenibacillus polymyxa secretes antifungal enzymes that have the ability to break down chitin, amylase, cellulose, and proteins [67]. The conidial germination of C. acutatum was decreased by more than 60% by using Bacillus spp. that isolated from the apple phylloplane due to the production of fixed and volatile compounds [68]. Prestop (Gliocladium catenulatum) and PlantShield (Trichoderma harzianum), two commercial fungal biocontrol agents, significantly decreased anthracnose development by up to 45% when sprayed three times onto plants between blooming and fruit ripening [69]. Six isolates of yeast (Saccharomyces cerevisiae) were successfully controlled C. acutatum on citrus plants during preharvest due to several actions including the production antifungal compounds, competition for nutrients, inhibition pathogen germination, and production of killer activity and hydrolytic enzymes when in contact with the fungus wall [70].
7. Anthracnose Diseases Management Challenges
Commercial strawberry growers depend on several management strategies, such as disease-free plants, proper irrigation, mulching, good sanitation practices, pesticides, crop rotation, and diseases resistant cultivars, but none of these have achieved effective control. To control different phytopathogenic fungi, including Colletotrichum species, growers rely on the use of the expensive fungicide input in strawberry production systems in the Northeast and Mid-Atlantic; however, usage of agrochemicals in the management of Anthracnose fruit rot (AFR) and crown rot (ACR) diseases on strawberry faces many challenges including: (i) fumigant methyl bromide (MeBr) is banned in many countries including United States because of its ozone-depleting properties since 2005 (ii) in some cases, fungicide applications failed to control anthracnose diseases epidemics due to several reasons including fungicides resistant fungi [10]. (iii) In the fresh strawberry fruit market, pesticide usage is less desirable by consumers and disease-free transplants in the field is a good starting point to achieve that [71]. (v) few number of fungicides were effective against diseases caused by Colletotrichum spp. on strawberry due to the variability on fungicides sensitivity [72]. In addition, AFR is difficult to control since symptomless infected and infested plants avoid examination and the pathogen may buildup to large levels in the field without being detected, creating the perfect environment for severe epidemics on ripening fruit under disease-favorable weather conditions. All these considerations highlight the important need for viable nonfumigated options in strawberry production systems for the management of soilborne diseases and pests.
8.3. Alternative and Sustainable Integrated Pest Management Strategies for soilborne diseases
Sustainable integrated pest management strategies are needed to meet the global demand [73]. Many alternative soil fumigation methods with synthetic chemicals such as glucosinolate-containing Brassica spp. is known to release volatile isothiocyanates (ITCs) which is lethal to different soilborne plants pathogens [74]. Several studies reported that bio-fumigation with ITC-producing plants is effective against some soilborne plant pathogens including, Rhizoctonia, Verticillium, Fusarium, Pythium, and Phytophthora spp. [75]. However, this fumigation method is not consistent due to the variable concentration of ITCs in different mustard varieties. From the grower's perspective, the efficacy of bio-fumigation was investigated on different plants and the level of adoption was low. Low efficacy of the treatment has been ascribed to many factors, including variation in soil texture, moisture, temperature, soil microbial community, and pH [76].
8.4. Overview of Anaerobic Soil Disinfestation (ASD)
Anaerobic soil disinfestation (ASD), a preplant soil disinfestation strategy, is another alternative promising soil bio-rational method that shows promise to control a wide range of soilborne pathogens and plant-parasitic nematodes. This strategy has not been experimented well in the Northeast U.S. [77,78]. The ASD process depends on adding carbon (C) source to stimulate microbially driven anaerobic soil conditions in moist soils covered with polyethylene mulch which is supposed to convert organic material into other organic compounds that should be lethal to soilborne pathogens [79]. This technique causes changes in soil physical and chemical characteristics, such as the formation of volatile fatty acids, a decrease in soil pH, a rise in soil moisture, and changes in soil nutrients as a result of organic matter addition [80]. Another mechanism of anaerobic soil disinfestation (ASD) against soilborne pathogens is lowering the redox potential below the critical redox potential (about +200 mV), which is able to reduce the survival stage of soilborne pathogens [81]. In soils treated with anaerobic soil disinfestation (ASD), the inoculum of soilborne pathogens such as V. dahlia was reduced by 80–100% compared with nontreated control and produced marketable fruit yields equivalent to fumigation [82]. In addition, root rot diseases complexes on tomato plants, caused by some pathogens including Colletotrichum spp., Verticillium dahliae, and Meloidogyne spp., was significantly reduced in ASD-treated soils in high tunnels compared with plants grown in control soils [83]. However, this technique needs to be optimized in terms of engaging beneficial microorganisms to control different soilborne pathogens and to enhance plant vigor and productivity in the same time [84]. The mechanisms of ASD are not fully understood; it may be due to the toxic by-products of anaerobic decomposition, volatile compounds, biocontrol by anaerobic soil microorganisms, or oxygen deficiency [77].
8.5. Optimizing Anaerobic soil disinfestation (ASD) with endophytic bacteria.
In general, several factors influence ASD effect application, such as carbon source, addition of beneficial microorganisms, and environmental conditions including soil types, pH, and temperature. Based on our previous work, brewer's spent grain (BSG) was used effectively as carbon sources to support soil microbial growth in ASD application in filed trails [85]. Engaging beneficial microorganisms such as endophytic bacteria that are used as biofertilizers or bio-stimulants with an anaerobic soil disinfestation (ASD) technique could generate a powerful tool to control soilborne pathogens and improve the growth and yield of strawberries, which may play a crucial role in sustainable crop production in the future [86]. Beneficial microorganisms can improve plant nutrition and support plant development under natural or stressed conditions, and increase yield and quality of many important crops [87]. In the interaction between the beneficial microorganisms and plants, these organisms act as nutrients supplier, phytohormones producer, plant growth enhancer, biocontrol of phytopathogens, and improver of soil structure [88]. Root dipping of seedlings (plug plants), followed by spray treatments of both probiotic bacteria including Bacillus amylolequefaciens (BChi1) and Paraburkholderia fungorum (BRRh-4) on leaves in the field, dramatically enhanced the fruit yield of strawberry by 48%, compared to non-treated controls [89]. In greenhouse, three strains of Bacillus velezensis, an endophyte bacterium, significantly suppressed strawberry pathogen growth (Colletotrichum gloeosporioides), and increased the marketable fruit yields in the field [90]. The gray mold disease in strawberry, caused by Botrytis cinerea, was controlled by five different isolates of Bacillus spp. via the production of diffusible and volatile antifungal chemicals [91]. The severity of Rhizoctonia root rot disease on Viburnum plants (Viburnum odoratissimum) was reduced on both greenhouse and field trials using TerraGrow product which is a complex of five Bacillus strains, including B. Licheniformis, B. Subtilis, B. Pumilus, B. Amylolquefiens, and B. Megaterium [92]. In a perennial strawberry production system, the combination of beneficial microbes and ASD enhanced plant vigor and fruit yield and suppressed the weed population and pathogenic microbes compared with untreated plants [93].
Future perspectives
The strawberry production has increased around the world in the past few years due to rising demand. Cutting edge research programs are ongoingto solve problems that threaten strawberry production while also enhancing fruit quality to meet customer demands. There is an urgent need for the adoption of sustainable alternative disease management measures that pose little threat to human health and the ecological system[93]. Integrated pest management (IPM) systems, which combine biological, cultural, and chemical tools with other supporting technologies, are the greatest way to achieve pest management that is effective, efficient, and sustainable. The combination of Anaerobic soil disinfestation (ASD) with beneficial microbes is being introduced lately in agricultural practices in place of fungicide and soil fumigants due to their economically viable and environment friendly which is supported by other studies recommendation [93]. It appears that ASD with different C sources is a viable approach to disease control, yield increase, and soil improvement specially in limited sources, organic, as well as smallholder farming. Furthermore, integrating ASD with beneficial microbes could reduce the initial investment for ASD treatments alone and create even powerful tool for pest management including fungi, bacteria, nematodes, and weeds in strawberry production system. Future research should concentrate on understanding how to incorporate the suitable beneficial microbe to control specific pathogen, as well as better understanding which mechanism(s) are responsible for disease control under different situations.
A precise detection method for pathogens that cause a crop disease is a requirement for the application of the appropriated disease control techniques. The current review may also highlight the need for rapid, non-destructive, and accurate method for anthracnose fruits rot (AFR) the early stage of the infection (latent period). Further, strawberry growers benefit from early incubation stage identification because it allows them to immediately remove contaminated plants before the disease spreads and causes further damage. In recent years, a combination of small unmanned aerial systems (UAS) equipped with multispectral imagining (MSI) sensors, which integrates spectral and image data, has demonstrated considerable benefits for non-destructive inspections, plant disease identification, and the safety of agricultural products. We believe this review of using remote sensing to diagnosis anthracnose fruit rot (AFR) will provide novel thoughts and encourage the development of appropriate theories, methods, and tools to monitor strawberry transplants in the nurseries which consider the main source of inoculum to the production farms.
References
- Giampieri, F. , et al., The strawberry: Composition, nutritional quality, and impact on human health. Nutrition, 2012. 28(1): p. 9-19.
- FAO STAT, C.a.L.P. 2023, FAO.
- Wu, F., Z. Guan, and A.J. Whidden, An Overview of the US and Mexico Strawberry Industries. EDIS, 2020. 2016(1): p. 4-4.
- Samtani, J.B. , et al., The status and future of the strawberry industry in the United States. HortTechnology, 2019. 29(1): p. 11-24.
- (USDA), U.S.D.o.A. U.S. strawberry consumption continues to grow. 2014 [cited 2022 05-20-2022]; Available from: https://www.ers.usda.gov/dataproducts/chart/gallery/gallery/chartdetail/?chartId=77884.
- Brown, M. , Florida strawberry production and marketing. The Strawberry: A Book for Growers. NF Childers, ed. Dr. Norman N. Childers Publications, Gainesville, FL, 2003: p. 31-42.
- Christman, J. and J.B. Samtani, A survey of strawberry production practices in Virginia. 2019.
- Garrido, C. , et al., New insights in the study of strawberry fungal pathogens. Genes Genomes Genomics, 2011. 5(1): p. 24-39.
- Amil-Ruiz, F. , et al., Partial activation of SA-and JA-defensive pathways in strawberry upon Colletotrichum acutatum interaction. Frontiers in plant science, 2016. 7: p. 1036.
- Forcelini, B.B. and N.A. Peres, Widespread resistance to QoI fungicides of Colletotrichum acutatum from strawberry nurseries and production fields. Plant Health Progress, 2018. 19(4): p. 338-341.
- Dale, A., B. R. Hughes, and D. Donnelly, The role of micropropagation in producing specific pathogen-tested plants. HortScience, 2008. 43(1): p. 74-77.
- Forcelini, B.B., F. P. Gonçalves, and N.A. Peres, Effect of inoculum concentration and interrupted wetness duration on the development of anthracnose fruit rot of strawberry. Plant disease, 2017. 101(2): p. 372-377.
- Poling, E.B. , Anthracnose on strawberry: Its etiology, epidemiology, and pathology, together with management strategies for strawberry nurseries: Introduction to the workshop. HortScience, 2008. 43(1): p. 59-65.
- Miller-Butler, M.A. , et al., Evaluation of detached strawberry leaves for anthracnose disease severity using image analysis and visual ratings. HortScience, 2019. 54(12): p. 2111-2117.
- Damm, U. , et al., The Colletotrichum acutatum species complex. Studies in mycology, 2012. 73: p. 37-113.
- Weir, B., P. Johnston, and U. Damm, The Colletotrichum gloeosporioides species complex. Studies in mycology, 2012. 73: p. 115-180.
- Smith, B. , First Report ofColletotrichum acutatumon Strawberry in the United States. Plant Disease, 1986. 70(11): p. 1074e.
- Agusti, L. , et al., Biocontrol of root rot of strawberry caused by Phytophthora cactorum with a combination of two Pseudomonas fluorescens strains. Journal of Plant Pathology, 2011. 93(1): p. 363-372.
- Denoyes-Rothan, B. , et al., Genetic diversity and pathogenic variability among isolates of Colletotrichum species from strawberry. Phytopathology, 2003. 93(2): p. 219-228.
- Mertely, J. and N. Peres, Root necrosis of strawberries caused by Colletotrichum acutatum. 2008, Online. Electronic Data Info. Source PP211. Univ. of Florida-IFAS ….
- Seijo, T.E. , et al. Resistance of strawberry cultivars and advanced selections to anthracnose and Botrytis fruit rots. in Proceedings of the Florida State Horticultural Society. 2008.
- Baroncelli, R. , et al., The Colletotrichum acutatum species complex as a model system to study evolution and host specialization in plant pathogens. Frontiers in Microbiology, 2017: p. 2001.
- Talhinhas, P. and R. Baroncelli, Colletotrichum species and complexes: geographic distribution, host range and conservation status. Fungal Diversity, 2021. 110(1): p. 109-198.
- Simmonds, J. , A study of the species of Colletotrichum causing ripe fruit rots in Queensland. Queensland Journal of Agricultural and Animal Sciences, 1966. 22(4): p. 437-459.
- Von Arx, J.A. , A revision of the fungi classified as Gloeosporium. A revision of the fungi classified as Gloeosporium., 1970.
- Bragança, C.A. , et al., Species of the Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal biology, 2016. 120(4): p. 547-561.
- Jayawardena, R. , et al., Notes on currently accepted species of Colletotrichum. Mycosphere, 2016. 7(8): p. 1192-1260.
- De Silva, D. , et al., Colletotrichum species associated with chili anthracnose in Australia. Plant Pathology, 2017. 66(2): p. 254-267.
- dos Santos Vieira, W.A. , et al., Optimal markers for the identification of Colletotrichum species. Molecular phylogenetics and evolution, 2020. 143: p. 106694.
- Strand, L.L. , Integrated pest management for strawberries. Vol. 3351. 2008: UCANR Publications.
- Yang, X. , et al., Effects of surface topography and rain intensity on splash dispersal of Colletotrichum acutatum. Phytopathology, 1990. 80(10): p. 1115-1120.
- Smith, B.J. , Epidemiology and pathology of strawberry anthracnose: a North American perspective. HortScience, 2008. 43(1): p. 69-73.
- Leandro, L. , et al., Germination and sporulation of Colletotrichum acutatum on symptomless strawberry leaves. Phytopathology, 2001. 91(7): p. 659-664.
- Prusky, D. , Pathogen quiescence in postharvest diseases. Annual review of Phytopathology, 1996. 34(1): p. 413-434.
- King, W. , et al., Effects of temperature on sporulation and latent period of Colletotrichum spp. infecting strawberry fruit. Plant Disease, 1997. 81(1): p. 77-84.
- Howard, C.M. , Anthracnose of strawberry caused by the Colleiotrichum complex in Florida. Plant Dis, 1992. 76: p. 976-981.
- Madden, L., L. Wilson, and M. Ellis, Field spread of anthracnose fruit rot of strawberry in relation to ground cover and ambient weather conditions. Plant Disease, 1993. 77(9): p. 861-866.
- Madden, L., X. Yang, and L. Wilson, Effects of rain intensity on splash dispersal of Colletotrichum acutatum. Phytopathology, 1996. 86(8): p. 864-874.
- Ntahimpera, N. , et al., Comparison of rain effects on splash dispersal of three Colletotrichum species infecting strawberry. Phytopathology, 1999. 89(7): p. 555-563.
- Agostini, J. and L. Timmer, Population dynamics and survival of strains of Colletotrichum gloeosporioides on citrus in Florida. Phytopathology, 1994. 84(4): p. 420-425.
- Lilja, A.T. , et al., Phytophthora cactorum and Colletotrichum acutatum: Survival and Detection. Agriculturae conspectus scientificus, 2006. 71(4): p. 121-128.
- MacKenzie, S. , et al., Host range and genetic relatedness of Colletotrichum acutatum isolates from fruit crops and leatherleaf fern in Florida. Phytopathology, 2009. 99(5): p. 620-631.
- Malarczyk, D., J. Panek, and M. Frąc, Alternative molecular-based diagnostic methods of plant pathogenic fungi affecting berry crops—A Review. Molecules, 2019. 24(7): p. 1200.
- Fazari, A. , et al., Application of deep convolutional neural networks for the detection of anthracnose in olives using VIS/NIR hyperspectral images. Computers and Electronics in Agriculture, 2021. 187: p. 106252.
- Lu, J. , et al., Field detection of anthracnose crown rot in strawberry using spectroscopy technology. computers and electronics in agriculture, 2017. 135: p. 289-299.
- Yeh, Y.-H. , et al., Strawberry foliar anthracnose assessment by hyperspectral imaging. Computers and Electronics in Agriculture, 2016. 122: p. 1-9.
- Alijani, Z. , et al., Biocontrol of strawberry anthracnose disease caused by Colletotrichum nymphaeae using Bacillus atrophaeus strain DM6120 with multiple mechanisms. Tropical Plant Pathology, 2022: p. 1-15.
- Veys, C. , et al., Multispectral imaging for presymptomatic analysis of light leaf spot in oilseed rape. Plant methods, 2019. 15(1): p. 1-12.
- Khaliq, A. , et al., Comparison of satellite and UAV-based multispectral imagery for vineyard variability assessment. Remote Sensing, 2019. 11(4): p. 436.
- Yuan, L. , et al., Detection of anthracnose in tea plants based on hyperspectral imaging. Computers and Electronics in Agriculture, 2019. 167: p. 105039.
- Canteri, M.H. , et al., ATR-FTIR spectroscopy to determine cell wall composition: Application on a large diversity of fruits and vegetables. Carbohydrate Polymers, 2019. 212: p. 186-196.
- Zhang, D. , et al., Detection of rice sheath blight using an unmanned aerial system with high-resolution color and multispectral imaging. PloS one, 2018. 13(5): p. e0187470.
- Liu, C. , et al., Application of multispectral imaging to determine quality attributes and ripeness stage in strawberry fruit. PloS one, 2014. 9(2): p. e87818.
- Wang, C. , et al. Strawberry anthracnose disease assessment using hyperspectral imaging. in Proc. 6th Int. Symp. Machinery and Mechatronics Agric. and Biosyst. Eng. 2012.
- Pérez-Roncal, C. , et al., Hyperspectral imaging to assess the presence of powdery mildew (Erysiphe necator) in cv. Carignan noir grapevine bunches. Agronomy, 2020. 10(1): p. 88.
- Pham, H. , et al. A novel bistatic lidar system for early-detection of plant diseases from unmanned aircraft. in Proceedings of the 31th Congress of the International Council of the Aeronautical Sciences (ICAS 2018), Belo Horizonte, Brazil. 2018.
- Zhang, C. and J.M. Kovacs, The application of small unmanned aerial systems for precision agriculture: a review. Precision agriculture, 2012. 13(6): p. 693-712.
- Messina, G. , et al., A comparison of UAV and satellites multispectral imagery in monitoring onion crop. An application in the 'Cipolla Rossa di Tropea' (Italy). Remote Sensing, 2020. 12(20): p. 3424.
- Chandel, A.K., L. R. Khot, and B. Sallato, Apple powdery mildew infestation detection and mapping using high-resolution visible and multispectral aerial imaging technique. Scientia Horticulturae, 2021. 287: p. 110228.
- Miller-Butler, M.A. , Screening strawberry clones for anthracnose disease resistance using traditional techniques and molecular markers. 2016: The University of Southern Mississippi.
- Simpson, D., A. Berrie, and A. Johnson. Hot Water Treatment to Eliminate Colletotrichum acutatum from Strawberry Runner Cuttings. in V International Strawberry Symposium 708. 2004.
- Mertely, J.C. and N.A. Peres, Anthracnose Fruit Rot of Strawberry: PP-207/PP130, rev. 9/2012. EDIS, 2012. 2012(9).
- Chechi, A. , et al., Diversity in species composition and fungicide resistance profiles in Colletotrichum isolates from apples. Pesticide biochemistry and physiology, 2019. 158: p. 18-24.
- Sengupta, P. , et al., Postharvest diseases of Indian gooseberry and their management: a review. International Journal of Fruit Science, 2020. 20(2): p. 178-190.
- Melanson, R.A. , et al., 2020 Southeast Regional Strawberry Integrated Pest Management Guide For Plasticulture Production. 2020.
- Shi, X.-C. , et al., Biocontrol strategies for the management of Colletotrichum species in postharvest fruits. Crop Protection, 2021. 141: p. 105454.
- Kim, Y.S., K. Balaraju, and Y. Jeon, Biological control of apple anthracnose by Paenibacillus polymyxa APEC128, an antagonistic rhizobacterium. The plant pathology journal, 2016. 32(3): p. 251.
- Moreira, R.R., C. N. Nesi, and L.L.M. De Mio, Bacillus spp. and Pseudomonas putida as inhibitors of the Colletotrichum acutatum group and potential to control Glomerella leaf spot. Biological control, 2014. 72: p. 30-37.
- Verma, N., L. MacDonald, and Z. Punja, Inoculum prevalence, host infection and biological control of Colletotrichum acutatum: causal agent of blueberry anthracnose in British Columbia. Plant Pathology, 2006. 55(3): p. 442-450.
- Lopes, M.R. , et al., Saccharomyces cerevisiae: a novel and efficient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiological research, 2015. 175: p. 93-99.
- McInnes, T., L. Black, and J. Gatti Jr, Disease-free plants for management of strawberry anthracnose crown rot. Plant Disease, 1992. 76(3): p. 260-264.
- Dowling, M. , et al., Managing Colletotrichum on fruit crops: A "complex" challenge. Plant Disease, 2020. 104(9): p. 2301-2316.
- Oldfield, T.L. , et al., A life cycle assessment of biosolarization as a valorization pathway for tomato pomace utilization in California. Journal of cleaner production, 2017. 141: p. 146-156.
- Matthiessen, J.N. and J.A. Kirkegaard, Biofumigation and enhanced biodegradation: opportunity and challenge in soilborne pest and disease management. Critical reviews in plant sciences, 2006. 25(3): p. 235-265.
- Hansen, Z. and A. Keinath, Increased pepper yields following incorporation of biofumigation cover crops and the effects on soilborne pathogen populations and pepper diseases. Applied soil ecology, 2013. 63: p. 67-77.
- Morra, M. and J. Kirkegaard, Isothiocyanate release from soil-incorporated Brassica tissues. Soil Biology and Biochemistry, 2002. 34(11): p. 1683-1690.
- Butler, D.M. , et al., Impact of anaerobic soil disinfestation combined with soil solarization on plant–parasitic nematodes and introduced inoculum of soilborne plant pathogens in raised-bed vegetable production. Crop Protection, 2012. 39: p. 33-40.
- Shrestha, U., R. M. Augé, and D.M. Butler, A meta-analysis of the impact of anaerobic soil disinfestation on pest suppression and yield of horticultural crops. Frontiers in plant science, 2016. 7: p. 1254.
- Molendijk, L. , et al. Perspectives of anaerobic soil disinfestation. in VII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation 883. 2009.
- Shrestha, U. , et al., Soil inoculation with Trichoderma asperellum, T. harzianum or Streptomyces griseoviridis prior to anaerobic soil disinfestation (ASD) does not increase ASD efficacy against Sclerotium rolfsii germination. Applied Soil Ecology, 2020. 147: p. 103383.
- Butler, D.M. , et al., Exploring warm-season cover crops as carbon sources for anaerobic soil disinfestation (ASD). Plant and Soil, 2012. 355(1): p. 149-165.
- Shennan, C. , et al., Anaerobic soil disinfestation is an alternative to soil fumigation for control of some soilborne pathogens in strawberry production. Plant pathology, 2018. 67(1): p. 51-66.
- Testen, A.L. , et al., On-farm evaluations of anaerobic soil disinfestation and grafting for management of a widespread soilborne disease complex in protected culture tomato production. Phytopathology, 2020(ja).
- Lamers, J. , et al. Anaerobic soil disinfestation for soil borne disease control in strawberry and vegetable systems: current knowledge and future directions. in VIII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation 1044. 2014.
- Liu, D. , Evaluation of anaerobic soil disinfestation using brewers spent grain and yeast inoculation on weed control in annual hill plasticulture strawberry production, in School of Plant and Environmental Sciences. 2021, Virginia Polytechnic Institute and State University. p. 115.
- Momma, N. , et al., Development of biological soil disinfestations in Japan. Applied microbiology and biotechnology, 2013. 97(9): p. 3801-3809.
- Kundan, R. , et al., Plant growth promoting rhizobacteria: mechanism and current prospective. J Fertil Pestic, 2015. 6(2): p. 9.
- Jayaprakashvel, M., C. Chitra, and N. Mathivanan, Metabolites of plant growth-promoting rhizobacteria for the management of soilborne pathogenic fungi in crops, in Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms. 2019, Springer. p. 293-315.
- Rahman, M. , et al., Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Scientific reports, 2018. 8(1): p. 1-11.
- Mei, C. , et al., A Potential Application of Endophytic Bacteria in Strawberry Production. Horticulturae, 2021. 7(11): p. 504.
- de Melo Pereira, G.V. , et al., A multiphasic approach for the identification of endophytic bacterial in strawberry fruit and their potential for plant growth promotion. Microbial ecology, 2012. 63(2): p. 405-417.
- Baysal, F. , Comparative performance of fungicides and biocontrol products in suppression of Rhizoctonia root rot in viburnum. J. Plant Pathol. Microbiol, 2018. 9(9).
- Rahman, M. , et al., Probiotic Bacteria, Anaerobic Soil Disinfestation and Mustard Cover Crop Biofumigation Suppress Soilborne Disease and Increase Yield of Strawberry in a Perennial Organic Production System. Plant Disease, 2023(ja).
- Zhang, X. , Detection and management of Colletotrichum acutatum sensu lato on strawberry. 2015, Iowa State University.
Figure 1.
Phylogenetic analysis of the 42 Colletotrichum acutatum species complex strains based on a multilocus concatenated alignment of the ITS, GAPDH, CHS-1, HIS3, ACT, and TUB2 genes. Accepted species by Jayawardena (2016) are highlighted in black while isolates not fitting with those are reported in gray. Colletotrichum orchidophilum was used as an outgroup.[15,27].
Figure 1.
Phylogenetic analysis of the 42 Colletotrichum acutatum species complex strains based on a multilocus concatenated alignment of the ITS, GAPDH, CHS-1, HIS3, ACT, and TUB2 genes. Accepted species by Jayawardena (2016) are highlighted in black while isolates not fitting with those are reported in gray. Colletotrichum orchidophilum was used as an outgroup.[15,27].
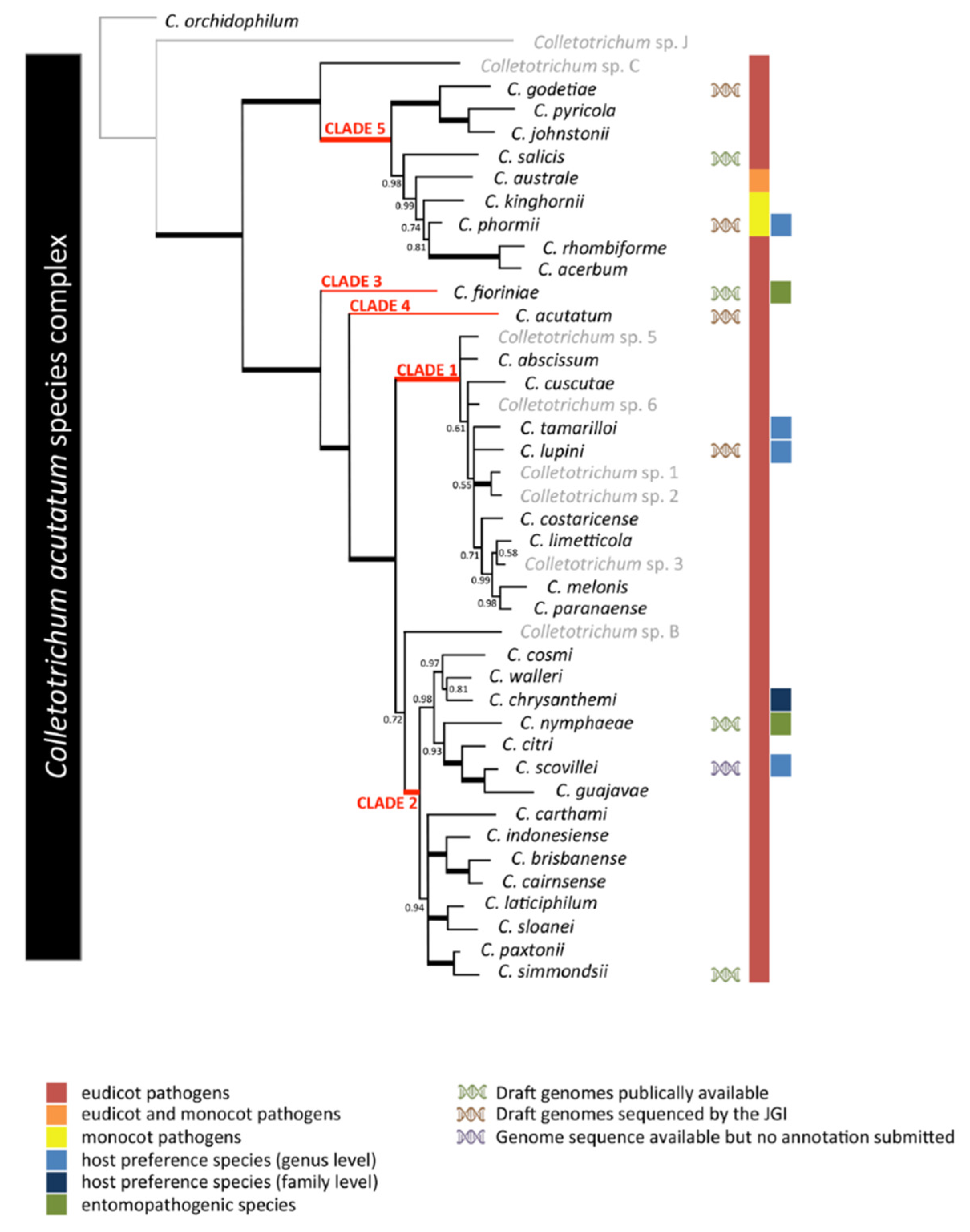
Figure 2.
The disease cycle of anthracnose fruit rot caused (AFR) by C. acutatum [94].
Figure 2.
The disease cycle of anthracnose fruit rot caused (AFR) by C. acutatum [94].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



