
Submitted:
01 August 2023
Posted:
02 August 2023
You are already at the latest version
Abstract
The green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae), is an economically important pest of crops worldwide. A M. persicae (SEF-R) population from a cabbage field in southeastern China was tested for susceptibilities to 13 insecticides. Compared with the susceptible clone (FFJ-S), extremely high and high-resistance to beta-cypermethrin (324-fold) and imidacloprid (106.9-fold) were detected in SEF-R. More importantly, this is the first report of resistance in field M. persicae population to sulfoxaflor, flupyradifurone, pymetrozine, spirotetramat, flonicamid, and broflanilide in China. The resistance factor decayed to a low level to sulfoxaflor and pymetrozine after 15 generations without any selection pressure. The resistance-related mutations (R81T and kdr) were detected in SEF, conferring target-site resistance to neonicotinoids and pyrethroids, respectively. Biochemistry bioassays confirmed the involvement of monooxygenase enzyme, carboxylesterase, superoxide dismutase and peroxidase in multi-insecticide resistance mechanism. The overexpression of P450s, esterases, and a UDP-glycosyltransferase, might be responsible for the multi-insecticide resistance in SEF-R. The knockdown of CYP6CY3 in SEF-R increased its susceptibility to imidacloprid, thiacloprid, and thiamethoxam, which verified that P450s play vital roles in neonicotinoid metabolism. This study will be helpful for chemical control, as well as for resistance monitoring and management of GPA, in China.
Keywords:
Myzus persicae
; insecticides
; resistance monitoring
; detoxification enzymes
; protective enzymes
; metabolic mechanism
1. Introduction
The green peach aphid (GPA) Myzus persicae (Sulzer, 1776) is an enormously polyphagous pest worldwide that has been reported to breed on more than 400 kinds of plants belong to 50 families, including crop, vegetable, fruit, and tobacco. Infestations of GPA reduced plant growth rates and caused reductions in crop quantity and quality [1]. In addition, the aphid vectors over 100 plant viruses, accounting for 67.7% of the aphid–vector viruses, which can lead to large yield reduction [2]. As a highly variable species, aphids evolve different strains and biotypes, distinguished by biology, host-plant preference, and insecticide resistance [3], which favor spread and rapid colonization. The cabbage Brassica oleracea var. capitata Linnaeus is the primary vegetable, having the largest vegetable planting area in China of 0.4-million hectares, which accounts for 25%–30% of Chinese vegetables. GPA is a destructive pest of B. oleracea throughout the country. Effective control is crucial to protect B. oleracea from aphid infection and prevent viral transmission.
Although some other Integrated Pest Management measures, such as natural enemies, cultural control, and mechanical control, have been used to control the GPA, chemical control is still the main way to suppress the pest in practice. Based on IRAC, there are 16 groups of effective insecticides that kill the GPA by acting on nerve and muscle (10 groups), growth and development (four groups) and respiration targets (two groups) targets [4]. In China, the systemic use of insecticides has played a vital role in controlling this pest since the introduction of organophosphates and carbamates insecticides in the 1950s [5]. To date, more than 600 pesticide products have been registered to manage the cabbage GPA in China. Among these products, 336 products, accounting for 55.17%, contain neonicotinoids, and 158 products, accounting for 25.94%, contain pyrethroids [6], which indicates that the two kinds of insecticides are sill the major insecticides used to eliminate the GPA of cabbage, and they have been unitized for several decades. Overreliance on these two kinds of insecticides has led to the occurrence and expansion of insecticide resistance in the GPA and threatens their continued efficacy. Insecticide-resistance monitoring data in China indicates that GPA field populations have high resistance to organophosphate insecticides, such as dimethoate and malathion, carbamate insecticides, such as pirimicarb and methomyl, pyrethroids, such as fenvalerate and cypermethrin, and neonicotinoids, such as imidacloprid [5,7,8].
The evolution and spread of resistance in GPA represent major threats to its sustainable control [9]. Understanding resistance mechanisms is critical for the development of rational strategies to prolong the lives of current insecticides. To date, seven independent mechanisms of resistance have been found in this species, including alterations in target site, metabolic mechanisms, and physical resistance mechanisms [9]. The enhanced expression of insecticide detoxifying enzymes is the most common mechanism in pests and often presents the greatest challenge. The overproduction of carboxylesterases (E4 and EF4) causes resistance of M. persicae to organophosphates and carbamates [10,11]. The amplification of the cytochrome P450 gene CYP6CY3 is associated with low level resistance to neonicotinoids [12,13,14], whereas the upregulation of glutathione S transferase (GST) and UDP-glycosyltransferases (UGTs) have been shown to affect insecticide resistance [15,16]. In addition, the protective antioxidant enzymes, superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT), which play important roles in immunity, help maintain a state of equilibrium in insects after insecticide exposure [17,18,19,20]. Field populations are exposed to different insecticides under variable selection pressures, which favors the evolution of multiple resistance through resistance mechanisms. Understanding these mechanisms will help us estimate the prevalence of resistant phenotypes when bioassays cannot be used to detect resistance [21,22].
Brassica species are cultivated year-round in Fujian Province, southeast China and are vulnerable to GPA and other notorious pests, such as Plutella xylostella, Phyllotreta striolata (Fabricius), Spodoptera litura Fabricius, and Pieris rapae. Various insecticides are frequently applied by farmers to control the damage from these pests. Presently, some active ingredients, such as flonicamid, cyantraniliprole, spirotetramat, afidopyropen, sulfoxaflor, and flupyradifurone, having excellent aphicidal activities, have been registered for GPA management in China [23]. Furthermore, other insecticides, such as broflanilide, have also entered the Chinese market and showed high potential in aphid control, although they have not been registered for aphid management [24]. These compounds belong to different groups of insecticides and may be alternated with pyrethroids or neonicotinoids in the field. Resistance to some insecticides, such as spirotetramat [25], flupyradifurone [26], and sulfoxaflor [16], has been detected in other countries, but the resistance status of GPA is unavailable in China, including Fujian Province. To warn, delay, or prevent the development of resistance in GPA to these compounds and better guide management practices, resistance monitoring in a GPA field population collected from Fujian Province was conducted. To further investigate the resistance mechanisms, biochemical assays and molecular methods were used to determine the activities and expression levels of detoxifying and protective enzymes, as well as related candidate genes, that might help monitor insecticide resistance.
2. Materials and methods
2.1. Insects
The laboratory susceptible strain of the GPA (FFJ-S) was continuously reared without insecticides exposure in the Fujian Institute of Plant Protection, Fujian Academy of Agricultural Science, Fuzhou, China, since it was collected from Arabidopsis thaliana in Jianxin District of Fuzhou, China in 2008. The resistant field population (SEF-R) was collected from cabbage in the Jin’an District of Fuzhou (26°12′84′′N, 119°33′59′′E) in 2022, where heavy insecticide spraying was carried out to control the aphid and other pests. Both GPA strains were reared on seedlings of Brassica campestris ssp. Chinensis (L.) at 21 ± 2°C and 65% ± 5% relative humidity, under a photoperiod of 16:8 h (light: dark). The SEF-R was reared one generation before being subject to a bioassay.
2.2. Pesticides, synergists, and other chemicals
The insecticides used for bioassays included flupyradifurone (96% purity; Bayer AG, Germany), thiacloprid (97.5% purity; Bayer AG), imidacloprid (97% purity; Bayer AG), esfenvalerate (90% purity; Shandong Huayang Technology Co., Ltd., China), beta-cypermethrin (95% purity; Nanjing Red Sun Co., LTD, China), flonicamid (96% purity; Ishihara Sangyo Kaisha, Ltd., Japan), cyantraniliprole (FMC Corporation, USA), spirotetramat (96% purity; Bayer AG), afidopyropen (92.5% purity; BASF SE, Germany), broflanilide (98% purity; Mitsui Chemical lnc., Japan), thiamethoxam (98% purity; Syngenta Group, Switzerland), Pymetrozine (95% purity; Syngenta Group, Switzerland), and sulfoxaflor (95.9% purity; Corteva Agriscience, USA). Sulfoxaflor
Synergists piperonyl butoxide (PBO; reagent grade), S,S,S-tributyl phosphorotrithioate (DEF; reagent grade), and diethyl maleate (DEM; reagent grade) were purchased from Sinopharm Group (Holding) Co. Ltd, China. Ethylenediaminetetraacetic acid, albumin bovine (BSA), and sodium dodecyl sulfate were purchased from Shanghai Aladdin Bio-Chem Technology Co., LTD (Shanghai, China). Eserine, α-naphthyl acetate, fast blue B salt, p-nitroanisole, n-phenylthiourea, Coomassie Brilliant Blue G250, DL-dithiothreitol, phenylmethylsulfonyl fluoride, Triton X-100, and other chemicals were purchased from Sigma-Aldrich, USA.
2.3. Bioassay methods
To assess the susceptibility of GPA to selected insecticides, the leaf-dip method recommended by IRAC [4] was used under laboratory conditions. All the tested insecticides were dissolved in 0.1% Triton X-100 to prepare five to seven concentrations. Clean leaf-discs of B. oleracea L. were dipped in insecticide solutions for 10 s. After being air dried at room, the leaf-discs were placed onto 1% agar plate (20-mm depth) in a petri dish (30-mm diameter and 40-mm depth). Then 20 2-d-old apterous aphids (for spirotetramat and pymetrozine test) or apterous adults (for other insecticides) were each transferred onto a treated leaf-disc using a paint brush, and each unit was covered with a close-fitting, ventilated lid. Mortalities were observed 4 d later to determine the impact of spirotetramat and pymetrozine and 3 d later to determine other insecticide effects. Aphids that could not right themselves within 10 s once turned on their backs were considered as dead.
2.4. Resistance recession
To test the stability of resistance, the SEF-R strain was reared for 15 generations (equivalent of approximately 1 year in the field) without insecticide exposure to investigate whether resistance to imidacloprid, beta-cypermethrin, sulfoxaflor, and pymetrozine was stable. GPA of the 1st (G1), 4th (G4), 7th (G7), 10th (G10), 12th (G12), and 15th (G15) generations were subjected to a bioassay. The average rate of change in response per generation (R) was estimated in accordance with Tabashnik [27], as follows:
where n represents the number of generations. An increase or decrease in resistance is indicated by positive and negative R values, respectively.
R = log10[final LC50] - log10[initial LC50]/n,
2.5. Synergism experiment
The effects of three synergists (PBO, DEF, and DEM) each in combination with a thiacloprid or cypermethrin mixture against GPD were evaluated using the bioassay method. The highest doses of PBO, DEF, and DEM on the susceptible strain that led to zero mortality were 0.08, 0.05, and 0.06 g L−1, respectively, using the bioassay method. Apterous adult aphids were exposed to leaf discs that were treated with PBO, DEF, or DEM combined with the thiacloprid and cypermethrin mixture. Four replicates were performed for each concentration in the bioassay.
2.6. Detoxification enzyme activity assays
The esterase activity was measured using a microplate reader in accordance with Byrne and Devonshire [28]. Individual aphids were homogenized in 60 μL of ice-cold phosphate buffered saline (PBS) (0.02 M, pH 6.5) and centrifuged at 4°C and 10,000 × g for 10 min. The supernatant was used as the esterase source. After incubating for 5 min at room, 100 μL substrate α-naphthyl acetate (107 mM) and 100 μL of eserine (107 mM) were mixed with the color development reagent (6 mg fast blue RR salt dissolved in 10 mL of 0.02 M PBS, pH 6.5) in the dark. Then, 150 μL of this reaction liquid was added to a 96-well microplate (Corning Life Sciences, USA), followed by the addition of 50 μL of supernatant fluid and 50 μL of PBS per well. The absorbance at 450 nm was measured for 5 min continuously using the kinetic model in a microplate reader (SPECTRA max PLUS384, Molecular Devices, USA). The esterase activity was reported as mOD450/min/aphid. For each population, approximately 100 individuals were measured.
The GST activity was determined in accordance with a published method involving 1-chloro-2,4-dinitrobenzene, with slight modifications [29]. Briefly, 60 adult aphids were homogenized in 2.0 mL ice-cold PBS (0.04 mol L−1, pH 7.5), and the supernatant solution was used after being centrifuged at 10,000 × g for 10 min at 4°C. Briefly, a 300-μL reaction mixture containing 100 μL diluted enzyme solution, 100 μL 1-chloro-2,4-dinitrobenzene (1.2 mM) substrate solution, and 100 μL glutathione ((6 mM) was prepared, after which the absorbance was measured at 340 nm for 10 min using the kinetic model. The results were determined based on the protein concentration of an enzyme source, and the specific activity was converted from an OD value.
The monooxygenase enzyme (MFO) activity was measured in accordance to Shang’s method [30]. Briefly, 60 adult aphids were homogenized in 2.0 mL ice-cold PBS (0.04 mol L−1, pH 7.8). The supernatant obtained by centrifuging 10,000 × g for 10 min at 4°C was added to a reaction unit containing NADPH and nitroanisole (0.05 mol L−1 in acetone) as a substrate. Hydrochloric acid (1 mol L−1) was added to terminate the reaction after incubation for 30 min at 37°C. Then, the reaction unit was extracted by a sodium hydroxide (NaOH) and chloroform solution. Finally, the optical density (OD) of the enzyme source was recorded at 400 nm using a microplate reader. The specific activity was obtained using a nitrophenol standard curve and the protein concentration of the enzyme source.
2.7. Determination of protective enzyme activities in M. persicae
The activities of the protective enzymes CAT (catalog no. A007-2), POD (catalog no. A084-1), and SOD (catalog no. A001-3) were determined using commercial assay kits in accordance with the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, China). A spectrophotometric method based on the ultraviolet absorption of peroxide released from the activity of CAT on hydrogen peroxide was used to determine the CAT level, and the nitroblue tetrazolium reduction method and the guaiacol method were used to determine the SOD and POD activities, respectively.
The protein contents of the enzyme solutions were determined using the Bradford method [31]. Serial dilutions of a BSA solution, and the samples were measured together. The protein contents of the samples were calculated using a standard curve based on the BSA solution. The diluted enzyme solutions (50 μL) were mixed with Coomassie Brilliant Blue (200 μL). After incubating at 25°C for 10 min, the absorbance at 595 nm was measured. All the experiments were repeated three times, and the average values were obtained from the triple-replicated data.
2.8. Target-site resistance (Detection of R81T, L1014F and M918T mutations)
Genetic analyses of GPA from SEF-R were conducted for known targe site resistance. This included the nicotinic acetylcholine receptor R81T mutation and the voltage-gated sodium channel knockdown resistance mutations L1014F (kdr) and M918L (Super-kdr), responsible for neonicotinoids and pyrethroids resistance in GPA, respectively. The real-time TaqMan assays were used for the mutation identification of genome DNA iin a MasterCycler ep RealPlex4 machine (Eppendorf, Hamburg, GER) according to the methods described in other studies [32,33].
2.9. Gene expression analysis
The mRNA levels of different enzyme-encoding genes in the FFJ-S and SEF-R strains were measured using RT-qPCR with SYBR® Green Supermix (ThermoFisher, USA) in a qTOWER 2.2 real-time quantitative PCR system (Analytikjena, Germany). Total RNA was extracted as described above and quantified using a ScanDrop 100 spectrophotometer (Analytikjena) in accordance with the manufacturer’s protocols. The RNA concentration was diluted to 0.8 μg μL–1 with diethyl pyrocarbonate-treated H2O, and 0.8 μg of RNA was then reverse transcribed in a 20-μL reaction volume using a TUREscript 1st Stand cDNA Synthesis Kit (Aidlab, Beijing, China) with the actin gene as an internal control (NCBI gene ID: 836110). In total, 15 genes, CYP6CY3 (Gene ID: KF218356), CYP6K1 (Gene ID: XM_022323152), CYP6CY23 (Gene ID: MF155662), CYP380C40 (Gene ID: OM677847), CYP6CY9 (Gene ID: XM_022316140), CYP6CY56 (Gene ID: MT472683), CYP4G15 (Gene ID: XM_022323310), CYP3CY06 (Gene ID: XM_022312311), E4 (Gene ID: X74554), THEM (Gene ID: XM_022326659), GST (Gene ID: 111036096), UGT344P2 (Gene ID: OM677846), POD (Gene ID: 112683716), SOD (Gene ID: 111035379), and CAT (Gene ID: 111041019), were quantified. The primers for these genes were designed using the Primer Express 3.0 software based on the sequences of target genes in NCBI or according other research, and they are listed in Supplementary Table 1.
Each RT-qPCR was conducted in a 20-μL mixture containing 1 μL of sample cDNA, 1 μL of each primer (200 nM), 6 μL of diethyl pyrocarbonate-treated H2O, and 10 μL of 2×SYBR® Green Supermix. The qPCR cycling parameters were as follows: 95°C for 3 min, followed by 39 cycles of 95°C for 10 s and 58°C for 30 s. A plate reader was used for the analysis. Melting curve generation was performed from 60 to 95°C (+1℃ (+1ti, holding time 4 s). After adding all the components, samples were centrifuged at 6,000 ×g for 1 min to keep all the components in the tube bottom. To check the reproducibility of the assay results, the qPCR for each sample was performed using three technical replicates and three biological replicates. The relative gene expression was calculated automatically using qPCRsoft3.2 software. The comparative 2−ΔΔCT method [34] was used for the relative quantification calculation.
2.10. In vivo RNAi of CYP6CY3 and bioassays
Fragments of CYP6CY3 were cloned into pGEM-T (Promega, USA) and used as templates for dsRNA-CYP6CY3 synthesis using a T7 RiboMAX™ Express RNAi System (Promega) in accordance with Peng et al. [35]. The dsRNA-CYP6CY3 (100 ng mL–1 in diet) was added to the artificial diet [36], and DEPC water and dsGFP (dsRNA of GFP) were also added to the artificial diet as the control. The dsRNA-CYP6CY3-treated artificial diet containing imidacloprid (100 mg L–1), thiacloprid (40 mg L–1), thiamethoxam (30 mg L–1), sulfoxaflor (50 mg L–1), or flupyradifurone (10 mg L–1) was used in the toxicity assessment, and the diet supplemented with DEPC water or dsGFP, but without insecticides, was used as the control. The experiments were performed in triplicate. To determine the efficiency of the dsRNA knockdowns of CYP6CY3, the aphids were collected after they fed on the artificial diet for 48 h, and the samples were used for qRT-PCR analysis. The mortality of the aphids was also calculated for 48 h. Each treatment was replicated in triplicate.
2.11. Data analysis
The enzymatic activities and the gene expression data between SEF-R and FJJ-S were compared with Student’s t-tests, and the mortalities of the GPA that were RNAi plus insecticide-treated versus mortalities of the control were compared with a one-way analysis of variance in combination with a Fisher’s least significant difference multiple comparison tests. These experiments were analyzed using DPS Data Processing System (Hangzhou Ruifeng Technology LTD, China) [37]. All the data are presented as the means ± standard errors (SEs). Differences were considered significant at P-value < 0.05 or 0.01. Median lethal concentrations (LC50) of each insecticide for adult apterous aphids were obtained by DPS Data Processing System. The LC50 values without overlap among 95% confidence intervals (CIs) were considered significantly different. The Resistance Factor (RF) was calculated as the ratio of the LC50 value of the field population to the LC50 value of the FFJ-S strain. Resistance levels were classified in accordance with the standard reported by World Health Organization [38]: susceptibility (RF < 3-fold), decreased susceptibility (RF = 3 to 5-fold), low resistance (RF = 5 to 10-fold), moderate resistance (RF = 10 to 40-fold), high resistance (RF = 40 to 160-fold), and extremely high resistance (RR > 160-fold).
3. Results
3.1. Insecticide resistance
The bioassays of the field and susceptible populations indicated that the field population had a reduced susceptibility to the 13 tested insecticides according to the WHO standard (Table 1). The LC50 of the SEF-R strain showed a reduced susceptibility to five insecticides of Group 4 in IRAC, imidacloprid (116.48 mg L–1), thiacloprid (54.86 mg L–1), thiamethoxam (33.2 mg L–1), sulfoxaflor (51.51 mg L–1), and flupyradifurone (12.67 mg L–1). When compared with the FFJ-S strain, the SEF-R exhibited a 106.9-fold resistance to imidacloprid, 29-fold resistance to thiacloprid, 12.9-fold resistance to thiamethoxam, 32.4-fold resistance to sulfoxaflor, and 9.5-fold resistance to flupyradifurone.
The SEF-R strain showed a high resistance to esfenvalerate (RF = 79.1-fold; LC50 = 764.4 mg L–1) and extremely high resistance to beta-cypermethrin (RF = 324-fold; LC50 = 2,371.54 mg L–1). The SEF-R strain also develop high resistance to pymetrozine (RF = 34.8-fold; LC50 = 29.89 mg L–1) and broflanilide (RF = 15.5-fold; LC50 = 15.23 mg L–1). For other three insecticides, low resistance to spirotetramat (RF = 8.1-fold; LC50 = 8.45 mg L–1) and flonicamid (RF = 5.8-fold; LC50 = 7.85 mg L–1) was detected in SEF-R. The LC50 values for afidopyropen and cyantraniliprole in the SEF-R were 4.0 and 3.4 times higher than those in FFJ-S, respectively (Table 1), which indicated a decreased susceptibility in the SEF-R to the two insecticides.
3.2. Stability of resistance to selected insecticides
The field population of GPA, SEF-R, was raised under laboratory conditions for 15 generations without insecticide exposure. Susceptibility to imidacloprid, beta-cypermethrin, sulfoxaflor, and pymetrozine in GPA were evaluated in G1, G4, G7, G10, G12, and G15, respectively. The GPA became much more susceptible to the selected four insecticides in the absence of insecticide selection. The RF of GPA against imidacloprid decreased from 106.9-fold in the G1 to 18.7-fold in the G15 (Figure 1A) relative to susceptible strain FFJ-S. The resistance level of GPA to imidacloprid dropped slowly from G1 (106.9-fold) to G4 (101.7-fold), reduced sharply from G4 to G12, and then maintained a stable level from G12 to G15.
The resistance level of GPA to beta-cypermethrin quickly declined from 324-fold in the G1 to 109.9-fold in the G10) and then remained steady from G10 to G15 (Figure 1B). The resistance level of GPA to sulfoxaflor decreased slowly from G1 to G4, and then, it quickly dropped from G4 to G10 (Figure 1C). Meanwhile, the RF of the GPA to pymetrozine sharply dropped from 34.8-fold (G1) to 15.2-fold (G4) and trended to be stable from G4 to G15 (Figure 1D). The decline rates of resistance against imidacloprid, beta-cypermethrin, sulfoxaflor, and pymetrozine were -0.05, -0.04, -0.04, and -0.04 in G15, respectively.
3.3. Synergist assessments and enzyme activity levels
The synergistic effects of PBO, DEF, and DEM on the toxicity of imidacloprid and beta-cypermethrin to the resistance field strain SEF-R were evaluated. The synergism ratios of PBO, DEF, and DEM on imidacloprid to SEF-R were 4.16-, 2.15- and 3.2-fold (Table 2), respectively, and those on beta-cypermethrin to SEF-R were 6.43-, 3.84- and 1.91-fold (Table 3), respectively. These three synergists had little synergistic effects on imidacloprid or beta-cypermethrin to susceptive strain FFJ-S (Table 2 and Table 3).
The SEF-R strain possessed significantly higher P450 (3.06-fold), carboxylesterase (3.04-fold), GST (1.5-fold), SOD (1.2-fold), and POD (3.4-fold) activities compared with the FFJ-S strain (Figure 2). The CAT activity was reduced significantly in the SEF-R.
3.4. R81T, kdr and super-kdr resistance
The R81T and kdr mutations were detected in the SEF-R, but super-kdr mutation was not found. In the SEF-R, the predominant genotype was the heterozygote for the R81T and kdr mutations. Its percentage of heterozygote was 64.29% for R81T and 79.35% for kdr, while the percentage of RR was 10.2% for R81T, respectively (Figure 3).
3.5. Enzyme genes expression.
Based on the qPCR analysis, the expression differences in 15 metabolic and protective enzyme-encoding genes between SEF-R and FFJ-S were assessed (Figure 4). Among the eight selected P450 genes, six, CYP6CY3 (3.63-fold), CYP6CY23 (3.13-fold), CYP380C40 (2.51-fold), CYP6CY9 (2.46-fold), CYP4G15 (5.24-fold), and CYP3CY06 (1.71-fold), were significantly upgraded, but the CYP6K1 and CYP6CY56 genes were downgraded in the SEF-R population (Figure 4). A significant overexpression of all three esterase genes, E4 (7.11-fold), and THEM (11.47-fold), were observed in the SEF-R population. The expression of UGT344P2 was estimated to have increased 18-fold in SEF-R, which made it the most highly overexpressed gene tested. The GST showed no significant difference in expression level between the SEF-R and FFJ-F. For the three protective enzyme genes, POD (2.04-fold) was significantly upregulated and SOD showed no significant difference, but the CAT level (0.57-fold) was significantly downregulated in the SEF-R compared with in the FFJ-S.
3.6. Knockdown of P450 genes increases the sensitivity of GPA to sulfoxaflor.
To evaluate the functional role of CYP6CY3 sulfoxaflor resistance in GPA, the expression levels of these two P450 genes were decreased by RNA interference in the SEF-R population, and the toxicity levels of imidacloprid, thiacloprid, thiamethoxam, sulfoxaflor, and flupyradifurone were tested after RNAi exposure. After GPA were fed a dsRNA-incorporated diet for 48 h, the transcript levels of CYP6CY3 were reduced to 0.62-fold compared with that of the control, which contained dsGFP (Figure 5A). The mortality levels after the dsCYP6CY13 treatment, imidacloprid exposure, thiacloprid exposure and thiamethoxam exposure were 78.33%, 66.67%, and 68.33%, respectively, which were significantly higher than that of the control at the diagnostic dose of sulfoxaflor (Figure 5B). However, sulfoxaflor, and flupyradifurone exposure did not affect the mortality rate.
4. Discussion
The evolution and spread of insecticide resistance in the GPA are considered major challenges in the management and elimination of the pest and its transmitted virus. To date, the GPA has developed resistance to almost all insecticides used worldwide. An aphid monitoring survey suggested that the GPA in a cabbage field located in southeastern China, Fujian Province, was not susceptible to the 13 tested insecticides categorized to six MoA groups [4]. Multi-insecticide resistance might drastically reduce the number of insecticides that can be used to control the insect in this district. To delay resistance development and prolong the product’s effectiveness, implementing insecticide resistance management strategies, including rational applications of insecticides, is urgently needed according to monitoring data.
Similar to most other countries, neonicotinoids (Group 4A) are one of the most important chemical classes of insecticides used against the GPA in China [39]. More than 55% of all registered products in China contain neonicotinoids, which have been a key instrument in GPA management [6]. However, the GPA have the second-greatest number of cases of neonicotinoid resistance, behind only those of Bemisia tabaci, according to the Arthropod Pesticide Resistance Database (APRD), and this represents a serious threat to the effective control of GPA by neonicotinoids [40]. The high level of imidacloprid-resistance detected in SEF-R reflected its status in cabbage fields, which is consistent with other monitoring data. Since susceptibility in the GPA was shown to decrease in 2005 [41], elevated levels of resistance to imidacloprid have been observed in most of the provinces of China, such as Beijing [42], Hunan [43], Jiangsu, and Shanghai [44], in which GPA have been tested. According to published documents, confirmed resistance to imidacloprid is much higher in vegetable aphid populations (RF = 32–350.9-fold) than in tobacco aphid populations (RF = 6.03–22-fold) in China [8]. Additionally, GPA collected from cabbage field in Longyao, Heibei Province exhibited the highest resistance (2,853-fold) [44]. We speculated that selection pressures rather than host plants shaped the evolution of imidacloprid resistance because GPA collected from tobacco fields in Europe evolved moderate to high resistance to imidacloprid (RF >15 detected in 86.7% populations in 2015) [45]. Less insecticide needed to be used against GPA because of the short growing season of tobacco in China, which also indicated that the limited application of imidacloprid delayed resistance development. The present study revealed moderate level resistance in the SEF-R clone to the other two neonicotinoids, thiacloprid and thiamethoxam, which were registered in 2015 to kill aphids on cabbage. Zhang [46] found that GPA collected from cabbage fields in Guangdong and Hunan Provinces, in southern and southeastern China, respectively, are still susceptible to thiamethoxam. There are few products of thiamethoxam (eight products) and thiacloprid (four products) allowed in cabbage aphid management, which may reduce the number of their spray applications in fields [6]. We inferred that the moderate resistance to the two nicotinoids in SEF-R was caused by cross-resistance between thiacloprid or thiamethoxam with imidacloprid verified in other studies [47,48].
As nAChR competitive modulators, sulfoxaflor (Subgroup 4C) and flupyradifurone (Subgroup 4D) belong to the novel classes sulfoximines and butenolides, respectively. Previous studies showed that the two compounds are effective against some GPA populations exhibiting neonicotinoid resistance [49,50], but later researches confirmed the cross-resistance between sulfoxaflor or flupyradifurone with imidacloprid in some field populations [48,51]. SEF-R exhibited resistance to the two compounds. Today, sulfoxaflor (Tefuli®) and flupyradifurone (Jinxian®) are only registered by Corteva Agriscience and Bayer AG, respectively, which make us easily know their application history. Growers did not apply the two compounds in the field in the recent years. However, field populations of GPA in other countries have developed high resistance to the two compounds. In total, 25% of the field-collected GPA in Greece had an RF˃119.2 for flupyradifurone [26], and field populations collected in Western Australia and Spain exhibited 4–23-fold and 0.6-61-fold resistance to sulfoxaflor, respectively [16] (Mezei et al. 2022). Our study first reported the resistance of GPA to sulfoxaflor and flupyradifurone in China, which might cause by high resistance to imidacloprid. The loss of sensitivity of GPA to sulfoxaflor and flupyradifurone threatened the applications of the two insecticides as alternative tools for integrated pest and resistance management. The resistance of SEF-R to the five Group 4 insecticides makes it difficult select related products. Stability experiments showed that the RFs of imidacloprid and sulfoxaflor dropped to 18.7- and 9-fold, respectively, after 15 generations without selection (Figure 1A). The study highlighted that the cessation of selective pressure could help restore the susceptibility of GPA to imidacloprid and sulfoxaflor, but they should be prohibited for at least one year. For Group 4 compounds, thiamethoxam and flupyradifurone should be recommended as alternative insecticides to manage GPA in the field, but avoid repeated applications.
Synthetic pyrethroids (Group 4A) form another common insecticide group that have been used for several decades for controlling the GPA in China. Since GPA collected from Beijing and Hebei exhibited extremely high resistance levels to fenvalerate (RF = 675) and deltamethrin (RF = 1,468) in 1990 [52], pyrethroid resistance became widespread across China by the late 1990s. Surveys from 2011 to 2014 revealed that more than 85.5% of 62 collected field populations had developed a high resistance to beta-cypermethrin [44] in China. The SEF-R population exhibited extremely high resistance to beta-cypermethrin (RF = 324-fold) and high resistance to esfenvalerate (RF = 79.1-fold). The recommended doses for GPA control ranged from 25 to 90 mg L–1, far below the LC50 values of esfenvalerate (764.4 mg L–1) and beta-cypermethrin (2,371.54 mg L–1) (Table 1), which led to control failures of GPA after applications of the two pyrethroids at recommended concentrations. After the SEF-R was reared for 15 generations without exposure to any insecticides, the GPA maintained a 91.3-fold resistance to beta-cypermethrin (LC50= 667.97 mg L–1) (Figure 1B). The two pyrethroids were not advised for the control of the GPA in this field.
IRAC classified pymetrozine [53] and flonicamid [54] to Group 9 “Selective homopteran feeding blockers”, with pymetrozine in Subgroup 9B and flonicamid in Subgroup 9C. The two compounds target the chordotonal organ, resulting in an inhibitory effect on aphid feeding, and they offer attractive alternatives to conventional insecticides due to a lack of cross-resistance. Pymetrozine has entered the Chinese market for about two decades and flonicamid has also been used for ten years. According to the Arthropod Pesticide Resistance Database [55], there have been no reports of field resistance to flonicamid in GPA worldwide. In Greece, RFs to pymetrozine ranged from 1.1 to 2.2) in 2018 [56], even lower than those found in 2010 (1.0–3.7 and 1.0–6.0 in clones from peach and tobacco, respectively) [57] although ~10-year passed. Astonishingly, the SEF-R evolved 5.8- and 29.89-fold resistance to flonicamid and pymetrozine, respectively. However, 205.9-fold resistance to pymetrozine has been documented in A. gossypii collected from Cheongju, South Korea [58], and 1,100-fold resistance to pymetrozine has also been detected in B. tabaci [59]. These reports suggest, possible resistance development in GAP to the two compounds. Furthermore, resistance to pymetrozine decreased quickly during the first four generation and remained stable in the subsequent nine generations. Owing to the quick increase in susceptibility due to the suspended application of pymetrozine and the relative susceptibility of the SEF-R to flonicamid, they could be used as alternative to other insecticides. The insecticides spirotetramat (Group 23), afidopyropen (Group 9D), and cyantraniliprole (Group 28), were recently used as aphicides against GPA in China. Spirotetramat has a new mode of action and is from a class of chemicals called tetramic acids, which act as Lipid Biosynthesis Inhibitors. Afidopyropen is a derivative of pyripyropene A that has a new mode of action as a chordotonal organ Transient Receptor Potential Vanilloid-type channel modulator, and cyantraniliprole is a member of the bisamides class of insecticides with an excellent aphicidal activity [23]. Except for the detection of high levels of resistance to spirotetramat in two populations of GPA collected from Queensland (170.7-fold resistance in Alloway171 and 105.6-fold resistance in Osborne171) [25], there has been little evidence of variation in susceptibility to afidopyropen, or cyantraniliprole. No obvious resistance has been observed to either afidopyropen or cyantraniliprole, and only low resistance to spirotetramat (RF=8.1) was identified in SEF-R. The three insecticides should be applied as alternatives to insecticides provoking high resistance levels.
Broflanilide, a new meta-diamide pesticide that acts on the γ-aminobutyric acid receptor in insect pests through a novel mechanism of action, is highly active against a broad range of pest, including lepidoptera, coleoptera, and homoptera [60]. Resistance to broflanilide in the GPA has not been documented in previous studies. To date, broflanilide is not permitted for the control of GPA, but it has been registered for the management of Plutella xylostella and Phyllotreta striolata (Fabricius) on cruciferous vegetables in China. We confirmed that growers used broflanilide spray more than 5 times per year. Selection pressures owing to the extensive use of broflanilide to control P. xylostella and P. striolata might lead to the evolvement of resistance in the GPA in the field. In the future, risk assessment of resistance in GPA is necessary prior to the registration of broflanilide use on cabbage.
Resistance monitoring data indicated that SEF-R developed multifold resistance to different insecticides through multiple mechanisms. The study of resistance in GPA has showed the importance of target-site mechanisms in developing high levels of resistance by affecting insecticide binding to receptors [9]. The R81T mutation in the nAChR gene have been proved responsible for high resistance to neonicotinoids and also found conferred cross-resistance to other insecticide classes function as agonists on the nAChR, such as sulfoxaflor [39,49,51] and butenolides [48] since it was first detected in France in GPA [61]. This resistance mutation has been detected in SEF-R, which has also been observed in Chinese populations of GPA [62]. We speculated the R81T mutation conferred the resistance in SEF-R to all five tested Group 4 compounds, and sulfoxaflor and butenolides are not used as alternative to rotate with neonicotinoids. Otherwise, the SEF-R possessed kdr mutation (L1014F) in sodium channel with 79.35% genotypic frequencies of RS heterozygotes, but super-kdr mutation was not found. Similaryly, Tang [44] found that the L1014F kdr mutation was detected in 35 of 45 field popoulations with 2.7-100% genotypic frequencies of RS heterozygotes in China. The kdr mutation also meant it difficult to select other pyrethroid compounds, such as deltamethrin, bifenthrin and fenpropathrin to control the aphid in the field.Enhanced detoxification by pesticide detoxification enzymes is an important mechanism for reducing the susceptibility to insecticides. PBO and DEF had useful synergistic effects with imidacloprid and beta-cypermethrin, but DEM had only useful effects with cypermethrin. Synergism of PBO with neonicotinoids or permethrins has been comprehensively documented in field-collected resistant GPA and shows usefulness with imidacloprid [8,63] and cyhalothrin [63], which is consistent with our results. Increases associated with MFO, CarE, and GST were also observed. The results suggested the possible involvement of the metabolic resistance mechanisms of the three detoxification enzymes in SEF-R. This agreed with reports on field resistant populations collected from Hunan and Chongqing Provinces [46,60] and Korea [64].
To further investigate the functions of the MFOs that were related to multi-resistance in SEF-R, we chose eight P450 genes for an expression analysis. Compared with in FFJ-S, seven P450 genes were more than 2-fold upregulated in the SEF-R, with CYP6CY3 (3.63-fold), CYP6CY23 (3.13-fold), CYP380C40 (2.51-fold), CYP6CY9 (2.46-fold), and CYP4G15 (5.24-fold) being significantly overexpressed. The overexpression of CYP6CY3 (10–75-fold) is responsible for neonicotinoid resistance in the GPA, which confers a moderate level of resistance to neonicotinoids [9,12]. Additionally, two GPA clones with different copy numbers of CYP6CY3 display low levels (3–20-fold) of resistance to imidacloprid and thiamethoxam, but the two clones survive and reproduce well on Brassica napus seedlings grown from seed treated with commercial levels of neonicotinoids [14] in a large-scale semi-field trial. The knockdown of CYP6CY3 in SEF-R increased the susceptibility of SEF-R to imidacloprid, thiacloprid, and thiamethoxam, but it did not affect the susceptibility to sulfoxaflor and flupyradifurone, which is consistent with the results of Nakao et al. [13]. The CYP6CY23 also ungraduated greatly in SEF-R, but it showed no capacity to metabolize nicotine [65], its function in GPA is needed to study.
The reduced penetration of insecticide is an additional mechanism of resistance in the imidacloprid-resistant 5191A clone of GPA [9,12]. Additionally, CYP4G15 was significantly overexpressed in SEF-R. The CYP4 clan in insects is involved in the last step of cuticular hydrocarbon biosynthesis [5,66]. Although there was no evidence of the CYP4 clan’s involvement in insecticides resistance in the GPA before our study, the overexpression levels of CYP4EM10 [67] and CYP4PR1 [69] in Triatoma infectants and CYP4G19 in Blattella germanica [70] are associated with pyrethroid resistance, and the increased expression levels of CYP4C68 and CYP4G70 are associated with imidacloprid resistance in Diaphorina citri Kuwayama [71]. Further studies need to confirm the function of CYP4G15 in the GPA. The overexpression of CYP380C40 (21–76-fold) and UDP-glucuronosyltransferase UGT344P2 (6–33-fold) has been identified in field populations of the GPA in Australia, and this proved partially responsible for sulfoxaflor resistance in the aphid [16]. The expression levels of CYP380C40 and UGT344P2 were estimated to be 2.51- and 18-fold, respectively, and the overexpression might be responsible for the 32.4-fold increase in resistance to sulfoxaflor in SEF-R compared with in FFJ-S. Although the function of some P450s in GPA has been well identified, co-up-regulation of several P450 genes are often observed and how these genes work together is unclear.
Additionally, we evaluated the differences in the expression levels of two esterase genes E4, and THEM between SEF-R and FFJ-S and found that they all had increased expression levels in SEF-R. The carboxylesterase, E4, mainly confer resistance to carbamates and organophosphates [11]. Resistance to these two kinds of insecticides in the GPA are widely distributed throughout China, although resistance levels have not been investigated in SEF-R. THEM is a type of thioesterase that catalyzes the cleavage of thioester bonds present in a wide range of glutathione, acyl-carrier proteins, fatty acyl-coenzyme A substrates, and other cellular molecules [72]. Thioester-containing proteins may be an outcome of different environments and selective pressures that trigger the innate immune response. [73,74]. The GST gene was not upregulated in SEF-R, which indicated that it was not associated with insecticide resistance. Antioxidant mechanisms were also investigated. A biochemical analysis revealed that the activities of SOD and POD were higher, and the CAT activity was lower, in SEF-R than in FFJ-S. Higher SOD and POD activities have also been observed in another type of field resistance, which is consistent with our results, but the CAT activity also increased in that resistant strain, which is contrary to our result [64]. Quantitative PCR showed increased expressions of SOD and POD, and a decreased expression of CAT, in SEF-R, consistent with their activity levels. The protective enzymes often work as part of a basic immune response.
Previous resistance monitoring surveys in China put emphasis on conventional insecticides, such as neonicotinoids and pyrethroids. Some new ingredients with aphicidal activities are often not included in monitoring program. SEF-R exhibited resistance to some new introduced or unregistered insecticides, such as broflanilide, which meant careful monitoring of susceptibility to these compounds is necessary. The risk of insecticide resistance including establishment of susceptibility baseline needs to be assessed when a new ingredient will be registered for new target pest insects in China. However, other target pest insects are not considered in the risk assessment. Based on our results, neonicotinoids and pyrethroids are not suitable to control SEF-R, but afidopyropen, and cyantraniliprole are good alternative to the two kinds of insecticdes effectively control SEF-R. Flonicamid and spirotetramat are also recommended in SEF-R management in order to maintain diversity of modes of action and implement resistance management in practice, which is critical in preventing or delaying resistance development. Other control tactics, such as biological control, should integrate with insecticides for enhanced protection from aphid damage and reduced insecticide application.
5. Conclusion
Our study demonstrated that a field population of M. persicae (SEF-R) exhibited an extremely high level of resistance to beta-cypermethrin, high levels of resistance to imidacloprid and esfenvalerate, moderate levels of resistance to thiacloprid, thiamethoxam, sulfoxaflor, pymetrozine, and broflanilide, and low levels of resistance to flupyradifurone, flonicamid, and spirotetramat. Afidopyropen an cyantraniliprole were recommended as the primary insecticides and flonicamid and spirotetramat were exploited as alternative to control SEF-R. Molecular analysis showed a high frequency of R81T mutation and kdr mutation, conferring target-site resistance to neonicotinoids and pyrethroids in SEF-R. Exposure to the PBO synergist restored the susceptibility to imidacloprid and beta-cypermethrin in SEF-R, suggesting the role of MFO in neonicotinoids and pyrethroids. The activities of MFO, CarE, and GST in SEF-R were significantly higher than those in FFJ-S, confirming that a metabolic detoxification mechanism is involved in multi-resistance to selected insecticides. RNA-seq results revealed that the overexpression of P450s (CYP6CY3, CYP6CY23, CYP380C40, CYP6CY9, and CYP4G15), esterases (E4, EF4, and THEM), and a UGT (UGT344P2) might be responsible for the multi-resistance of SEF-R. A knockdown of CYP6CY3 in SEF-R increased the susceptibility of SEF-R to imidacloprid, thiacloprid, and thiamethoxam, which verified that the P450 plays a vital role in neonicotinoid metabolism. The increased activities of POD and SOD enzymes, and the upregulation of the two encoding genes, showed that the two protective enzymes play some role in the resistance of SEF-R to various insecticides. However, more work is required, in order to determine the exact detoxification mechanism conferring multi-resistance in SEF-R.
Author Contributions
Hu J.: Conceptualization; investigation; validation; writing—review and editing; funding acquisition; Wang J.: methodology; investigation; validation; Yu Y.: formal analysis; investigation; Chen F.: formal analysis; writing—review and editing; Rao W.: investigation; writing—review and editing; funding acquisition; Lin L.: methodology; investigation; Fan G.: Conceptualization; funding acquisition;
Funding
This work was supported by Fujian Natural Science Foundation (2022J01461), the program of Fujian Provincial Department of Science & Technology (2020R10240011), program of Science and Technology Innovation Foundation of FAAS Supported by Financial Department of Fujian Government (CXTD2021002-1, XTCXGC2021017).
Data Availability Statement
Data are available from the corresponding author.
Acknowledgments
We wish to thank Fei Shuhua for her support with rearing the aphids.
Conflicts of Interest
The authors declare no conflict of interest
References
- Sharma, S.; Sood, A.K.; Ghongade, D.S. Assessment of losses inflicted by the aphid, Myzus persicae (Sulzer) to sweet pepper under protected environment in north western Indian Himalayan region. Phytoparasitica 2022, 50, 51–62. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Bosque-Pérez, N.A.; Davis, T.S. Insect-Borne Plant Pathogens and Their Vectors: Ecology, Evolution, and Complex Interactions. Annu. Rev. Entomol. 2018, 63, 169–191. [Google Scholar] [CrossRef] [PubMed]
- van Emden, H.F. Plant resistance to Myzus persicae induced by a plant regulator and measured by aphid relative growth rate. Entomologia Experimentalis et Applicata 1969, 12, 125–131. [Google Scholar] [CrossRef]
- IRAC. 2023. Available online: https://irac-online.org/mode-of-action/Accession:2023.
- Gao, X.W.; Zheng, B.Z.; Cao, B.J. Resistance in Myzus persicae to organophosphorus and carbamate insecticides in China. Acta Phytophy Sinica 1992, 19, 365–371. [Google Scholar]
- ICAMA. Available online: http://www.chinapesticide.org.cn/zgnyxxw/kgls/indexAccession data: 2023.
- Tang, Q.-L.; Ma, K.-S.; Hou, Y.-M.; Gao, X.-W. Monitoring insecticide resistance and diagnostics of resistance mechanisms in the green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae) in China. Pestic. Biochem. Physiol. 2017, 143, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, Z.; Shi, L.; Shen, G.; He, L. Insecticide resistance monitoring and metabolic mechanism study of the green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae), in Chongqing, China. Pestic. Biochem. Physiol. 2016, 132, 21–28. [Google Scholar] [CrossRef]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Field, L.M.; Foster, S.P.; Moores, G.D.; Williamson, M.S.; Blackman, R.L. The evolution of insecticide resistance in the peach–potato aphid, Myzus persicaePhil. Trans. R. Soc. Lond. B 1998, 353, 1677–1684. [Google Scholar] [CrossRef]
- Lan, W.-S.; Cong, J.; Jiang, H.; Jiang, S.-R.; Qiao, C.-L. Expression and Characterization of Carboxylesterase E4 Gene from Peach–Potato Aphid (Myzus persicae) for Degradation of Carbaryl and Malathion. Biotechnol. Lett. 2005, 27, 1141–1146. [Google Scholar] [CrossRef]
- Puinean, A.M.; Foster, S.P.; Oliphant, L.; Denholm, I.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Bass, C. Amplification of a Cytochrome P450 Gene Is Associated with Resistance to Neonicotinoid Insecticides in the Aphid Myzus persicae. PLOS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef]
- Nakao, T.; Kawashima, M.; Banba, S. Differential metabolism of neonicotinoids by Myzus persicae CYP6CY3 stably expressed in Drosophila S2 cells. J. Pestic. Sci. 2019, 44, 177–180. [Google Scholar] [CrossRef]
- Kirkland, L.S.; Chirgwin, E.; E Ward, S.; Congdon, B.S.; van Rooyen, A.; A Umina, P. P450-mediated resistance in Myzus persicae (Sulzer) (Hemiptera: Aphididae) reduces the efficacy of neonicotinoid seed treatments in Brassica napus. Pest Manag. Sci. 2023, 79, 1851–1859. [Google Scholar] [CrossRef]
- Pan, Y.; Xu, P.; Zeng, X.; Liu, X.; Shang, Q. Characterization of UDP-Glucuronosyltransferases and the Potential Contribution to Nicotine Tolerance in Myzus persicae. Int. J. Mol. Sci. 2019, 20, 3637. [Google Scholar] [CrossRef] [PubMed]
- Pym, A.; Umina, P.A.; Reidy-Crofts, J.; Troczka, B.J.; Matthews, A.; Gardner, J.; Hunt, B.J.; van Rooyen, A.R.; Edwards, O.R.; Bass, C. Overexpression of UDP-glucuronosyltransferase and cytochrome P450 enzymes confers resistance to sulfoxaflor in field populations of the aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2022, 143, 103743. [Google Scholar] [CrossRef] [PubMed]
- Kayser, H.; Palivan, C.G. Stable free radicals in insect cuticles: Electron spin resonance spectroscopy reveals differences between melanization and sclerotization. Arch. Biochem. Biophys. 2006, 453, 179–187. [Google Scholar] [CrossRef]
- Qin, D.; Liu, B.; Zhang, P.; Zheng, Q.; Luo, P.; Ye, C.; Zhao, W.; Zhang, Z. Treating green pea aphids, Myzus persicae, with azadirachtin affects the predatory ability and protective enzyme activity of harlequin ladybirds, Harmonia axyridis. Ecotoxicol. Environ. Saf. 2021, 212, 111984. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, H.; Wang, Z.; Long, G.-Y.; Jin, D.-C. Protective and Detoxifying Enzyme Activity and ABCG Subfamily Gene Expression in Sogatella furcifera Under Insecticide Stress. Front. Physiol. 2019, 9, 1890. [Google Scholar] [CrossRef]
- Wang, H.; Xin, T.; Wang, J.; Zou, Z.; Zhong, L.; Xia, B. Sublethal effects of bifenazate on biological traits and enzymatic properties in the Panonychus citri (Acari: Tetranychidae). Sci. Rep. 2021, 11, 20934. [Google Scholar] [CrossRef] [PubMed]
- Balmert, N.J.; Rund, S.S.; Ghazi, J.P.; Zhou, P.; Duffield, G.E. Time-of-day specific changes in metabolic detoxification and insecticide resistance in the malaria mosquito Anopheles gambiae. J. Insect Physiol. 2014, 64, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Brogdon, W.G. Chapter 5: Insecticide Resistance Monitoring; microplate enzyme activity assays. M.Q. Benedict (Ed.), Methods in anopheles Research (fourth ed.), Centers for Disease Control and Prevention, Atlanta, USA (2014), pp. 240-247.
- de Little, S.C.; Umina, P.A. Susceptibility of Australian Myzus persicae (Hemiptera: Aphididae) to three recently registered insecticides: Spirotetramat, Cyantraniliprole, and Sulfoxaflor. J. Econ. Entomol. 2017, 110, 1764–1769. [Google Scholar] [CrossRef]
- Nakao, T.; Banba, S. Broflanilide: A meta-diamide insecticide with a novel mode of action. Bioorganic Med. Chem. 2016, 24, 372–377. [Google Scholar] [CrossRef]
- Umina, P.A.; Bass, C.; van Rooyen, A.; Chirgwin, E.; Arthur, A.L.; Pym, A.; Mackisack, J.; Mathews, A.; Kirkland, L. Spirotetramat resistance in Myzus persicae (Sulzer) (Hemiptera: Aphididae) and its association with the presence of the A2666V mutation. Pest Manag. Sci. 2022, 78, 4822–4831. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, F.; Folia, M.; Ilias, A.; Papapetrou, P.; Roditakis, E.; Bass, C.; Vontas, J.; Margaritopoulos, J.T. Flupyradifurone resistance in Myzus persicae populations from peach and tobacco in Greece. Pest Manag. Sci. 2021, 78, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E. Evolution of resistance to Bacillus thuringiensis. Annual Review of Entomology 1994, 39, 47–79. [Google Scholar] [CrossRef]
- Byrne, F.; Devonshire, A. Insensitive Acetylcholinesterase and Esterase Polymorphism in Susceptible and Resistant Populations of the Tobacco Whitefly Bemisia tabaci (Genn.). Pestic. Biochem. Physiol. 1993, 45, 34–42. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Shang, C.-C.; Soderlund, D.M. Monooxygenase activity of tobacco budworm (Heliothis virescens F.) larvae: tissue distribution and optimal assay conditions for the gut activity. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1984, 79, 407–411. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Puinean, A.M.; Elias, J.; Slater, R.; Warren, A.; Field, L.M.; Williamson, M.S.; Bass, C. Development of a high-throughput real-time PCR assay for the detection of the R81T mutation in the nicotinic acetylcholine receptor of neonicotinoid-resistant Myzus persicae. Pest Manag. Sci. 2013, 69, 195–199. [Google Scholar] [CrossRef]
- A Anstead, J.; Williamson, M.S.; Eleftherianos, I.; Denholm, I. High-throughput detection of knockdown resistance in Myzus persicae using allelic discriminating quantitative PCR. Insect Biochem. Mol. Biol. 2004, 34, 871–877. [Google Scholar] [CrossRef]
- Kenneth, J.L.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Peng, T.; Pan, Y.; Gao, X.; Xi, J.; Zhang, L.; Ma, K.; Wu, Y.; Zhang, J.; Shang, Q. Reduced abundance of the CYP6CY3-targeting let-7 and miR-100 miRNAs accounts for host adaptation of Myzus persicae nicotianae. Insect Biochem. Mol. Biol. 2016, 75, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.-H.; Yu, X.-R.; Shang, Q.-L.; Shi, X.-Y.; Gao, X.-W. Oral Delivery Mediated RNA Interference of a Carboxylesterase Gene Results in Reduced Resistance to Organophosphorus Insecticides in the Cotton Aphid, Aphis gossypii Glover. PLOS ONE 2014, 9, e102823. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.-Y.; Zhang, C.-X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2012, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]
- WHO. Status of resistance in houseflies, Musca domestica, Document VBC/EC/80.7, World Health Organization, Geneva, Switzerland, 1980.
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- Michigan State University, Arthropod pesticide resistance database. 2014 (accessed 2023.05.15).
- Gu, C.; Wang, G.; Wang, K.Y.; Ma, H.; Guo, Q. Studies on the resistance level of Myzus persicae (Sulzer) in main tobacco regions of southwest. J. Plant Protection. 2006, 33, 77–80. [Google Scholar]
- Gong, Y. .; Wang, Z.; Shi, B.; Kand, Z.; Zhu, L.; Guo, X.; Liu, J.; Wei, S. Resistance status of Myzus persicae (Sulzer) (Hemiptera: Aphididae) populations to pesticide in Beijing. Scientia Agricultura Sinica, 2011, 44, 4385–4394. [Google Scholar]
- Bo, J.; Zhou, X.; Fang, Y. Studies on monitoring of insecticide resistance of Myzus persicae. Agrochemicals Research & Application 2009, 13, 27–29. [Google Scholar]
- Tang, Q. Insecticide resistance and genetic variation of field populations of Myzus pericae (Sulzer), in China. Ph.D. Dissertation, China Agricultural University. 2015.
- Voudouris, C.C.; Williamson, M.S.; e, P.J. , Kati, A. N.; Sahinoglou, A.J.; Margaritopoulos, J.T. Evolution of imidacloprid resistance in Myzus persicae in Greece and susceptibility data for spirotetramat. Pest Manag. Sci. 2017, 73, 1804–1812. [Google Scholar]
- Zhang, P.Y. Resistance monitoring and biochemistry mechanism of thiamethoxam in Myzus persicae. Master Dissertation, Hunan Agricultural University. 2014.
- Foster, S.P.; Cox, D.; Oliphant, L.; Mitchinson, S.; Denholm, I. Correlated responses to neonicotinoid insecticides in clones of the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae). Pest Manag. Sci. 2008, 64, 1111–1114. [Google Scholar] [CrossRef]
- Cutler, P.; Slater, R.; Edmunds, A.J.; Maienfisch, P.; Hall, R.G.; Earley, F.G.; Pitterna, T.; Pal, S.; Paul, V.-L.; Goodchild, J.; et al. Investigating the mode of action of sulfoxaflor: a fourth-generation neonicotinoid. Pest Manag. Sci. 2013, 69, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Watson, G.B.; Loso, M.R.; Geng, C.; Babcock, J.M.; Thomas, J.D. Sulfoxaflor and the sulfoximine insecticides: Chemistry, mode of action and basis for efficacy on resistant insects. Pestic. Biochem. Physiol. 2013, 107, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Jeschke, P.; Velten, R.; E Beck, M.; Ebbinghaus-Kintscher, U.; Thielert, W.; Wölfel, K.; Haas, M.; Kunz, K.; Raupach, G. Flupyradifurone: a brief profile of a new butenolide insecticide. Pest Manag. Sci. 2015, 71, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Mezei, I.; Valverde-Garcia, P.; Siebert, M.W.; Gomez, L.E.; Torne, M.; Watson, G.B.; Raquel, A.M.; Fereres, A.; Sparks, T.C. Impact of the nicotinic acetylcholine receptor mutation R81T on the response of European Myzus persicae populations to imidacloprid and sulfoxaflor in laboratory and in the field. Pestic. Biochem. Physiol. 2022, 187, 105187. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zheng, B.; Cao, B. Resistance levels of Myzus persicae to pyrethroids in Beijing and Langfang of Hebei. Pesticides 1993, 32, 14. [Google Scholar]
- Flückiger, C.R.; Kristinsson, H.; Senn, R. ; Rindlisbacher, A, Buholzer, H.; Voss, G. CGA 215–944—a novel agent to control aphids and whiteflies. Proc 1992 Brighton Crop Prot Conf—Pests and diseases, vol 1, pp 43–50.
- Morita, M.; Ueda, T.; Yoneda, T.; Koyanagi, T.; Haga, T. Flonicamid, a novel insecticide with a rapid inhibitory effect on aphid feeding. Pest Manag. Sci. 2007, 63, 969–973. [Google Scholar] [CrossRef]
- Mota-Sanchez, D.; Wise, J.C. The Arthropod Pesticide Resistance Database. Michigan State University. 2023. Available online: http://www.pesticideresistance.orgAccession data: 2023.
- Margaritopoulos, J.T.; Kati, A.; Voudouris, C.; Skouras, P.; Tsitsipis, J. Long-term studies on the evolution of resistance of Myzus persicae (Hemiptera: Aphididae) to insecticides in Greece. Bull. Entomol. Res. 2020, 111, 1–16. [Google Scholar] [CrossRef]
- Margaritopoulos, J.T.; Tsamandani, K.; Kanavaki, O.M.; Katis, N.I; Tsitsipis, J.A. Efficacy of pymetrozine against Myzus persicae and in reducing potato virus Y transmission on tobacco plants. J. Appl. Entomo. 2010, 134, 323–332. [Google Scholar] [CrossRef]
- Koo, H.-N.; An, J.-J.; Park, S.-E.; Kim, J.-I.; Kim, G.-H. Regional susceptibilities to 12 insecticides of melon and cotton aphid, Aphis gossypii (Hemiptera: Aphididae) and a point mutation associated with imidacloprid resistance. Crop. Prot. 2014, 55, 91–97. [Google Scholar] [CrossRef]
- Gorman, K.; Slater, R.; Blande, J.D.; Clarke, A.; Wren, J.; McCaffery, A.; Denholm, I. Cross-resistance relationships between neonicotinoids and pymetrozine in Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2010, 66, 1186–1190. [Google Scholar] [CrossRef]
- Li, R.; Cheng, S.; Chen, Z.; Guo, T.; Liang, P.; Zhen, C.; Wang, J.; Zhang, L.; Liang, P.; Gao, X. Establishment of Toxicity and Susceptibility Baseline of Broflanilide for Aphis gossypii Glove. Insects 2022, 13, 1033. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, M.; Yokota, M.; Ihara, M.; Akamatsu, M.; Sattelle, D.B.; Matsuda, K. Role in the Selectivity of Neonicotinoids of Insect-Specific Basic Residues in Loop D of the Nicotinic Acetylcholine Receptor Agonist Binding Site. Mol. Pharmacol. 2006, 70, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ding, Q.; Wang, X.; Wang, R.; Ullah, F.; Gao, X.; Song, D. V101I and R81T mutations in the nicotinic acetylcholine receptor β1 subunit are associated with neonicotinoid resistance in Myzus persicae. Pest Manag. Sci. 2022, 78, 1500–1507. [Google Scholar] [CrossRef] [PubMed]
- Panini, M.; Dradi, D.; Marani, G.; Butturini, A.; Mazzoni, E. Detecting the presence of target-site resistance to neonicotinoids and pyrethroids in Italian populations of Myzus persicae. Pest Manag. Sci. 2013, 70, 931–938. [Google Scholar] [CrossRef]
- Choi, B.R.; Lee, S.W.; Yoo, J.K. Resistance mechanisms of green peach aphid, Myzus persicae (Homoptera: Aphididae), to imidacloprid. Korean J. Appl. Entomol. 2001, 40, 265–271, [in Korean]. [Google Scholar]
- Singh, K.S.; Troczka, B.J.; Duarte, A.; Balabanidou, V.; Trissi, N.; Carabajal Paladino, L.Z.C.; Nguyen, P.; Zimmer, C.T.; Papapostolou, K.M.; Randall, E.; et al. The genetic architecture of a host shift: An adaptive walk protected an aphid and its endosymbiont from plant chemical defenses. Sci. Adv. 2020, 6, eaba1070. [Google Scholar] [CrossRef]
- Qiu, Y.; Tittiger, C.; Wicker-Thomas, C.; LE Goff, G.; Young, S.; Wajnberg, E.; Fricaux, T.; Taquet, N.; Blomquist, G.J.; Feyereisen, R. An insect-specific P450 oxidative decarbonylase for cuticular hydrocarbon biosynthesis. Proc. Natl. Acad. Sci. USA 2012, 109, 14858–14863. [Google Scholar] [CrossRef]
- Feyereisen, R. Origin and evolution of the CYP4G subfamily in insects, cytochrome P450 enzymes involved in cuticular hydrocarbon synthesis. Mol. Phylogenetics Evol. 2020, 143, 106695. [Google Scholar] [CrossRef]
- Dulbecco, A.B.; Moriconi, D.E.; Calderón-Fernández, G.M.; Lynn, S.; McCarthy, A.; Roca-Acevedo, G.; Salamanca-Moreno, J.A.; Juárez, M.P.; Pedrini, N. Integument CYP genes of the largest genome-wide cytochrome P450 expansions in triatomines participate in detoxification in deltamethrin-resistant Triatoma infestans. Sci. Rep. 2018, 8, 2–13. [Google Scholar] [CrossRef]
- Dulbecco, A.B.; Moriconi, D.E.; Pedrini, N. Knockdown of CYP4PR1, a cytochrome P450 gene highly expressed in the integument tissue of Triatoma infestans, increases susceptibility to deltamethrin in pyrethroid-resistant insects. Pestic. Biochem. Physiol. 2021, 173, 104781. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Zhang, L.; Liu, N. Overexpression of CYP4G19 associated with a pyrethroid-resistant strain of the German cockroach, Blattella germanica (L.). Gene 2003, 314, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Mo, X.; Rizvi, S.A.H.; Li, C.; Zeng, X. Detection and biochemical characterization of insecticide resistance in field populations of Asian citrus psyllid in Guangdong of China. Sci. Rep. 2018, 8, 12587. [Google Scholar] [CrossRef]
- Swarbrick, C.M.; Nanson, J.D.; Patterson, E.I.; Forwood, J.K. Structure, function, and regulation of thioesterases. Prog. Lipid Res. 2020, 79, 101036. [Google Scholar] [CrossRef] [PubMed]
- Upasana, S.; Ioannis, E. Evolution and function of thioester-containing proteins and the complement system in the innate immune response. Front. Immunol. 2017, 29, 8. [Google Scholar]
Figure 1.
Change in LC50 values (95% CI) of imidacloprid (A), beta-cypermethrin (B), sulfoxaflor (C), and pymetrozine (D) to M. persicae collected from the field and reared in the laboratory without exposure to any insecticides. Numbers on the tops of the graph bars represent resistance ratios compared with those of a susceptible strain Pila.
Figure 1.
Change in LC50 values (95% CI) of imidacloprid (A), beta-cypermethrin (B), sulfoxaflor (C), and pymetrozine (D) to M. persicae collected from the field and reared in the laboratory without exposure to any insecticides. Numbers on the tops of the graph bars represent resistance ratios compared with those of a susceptible strain Pila.
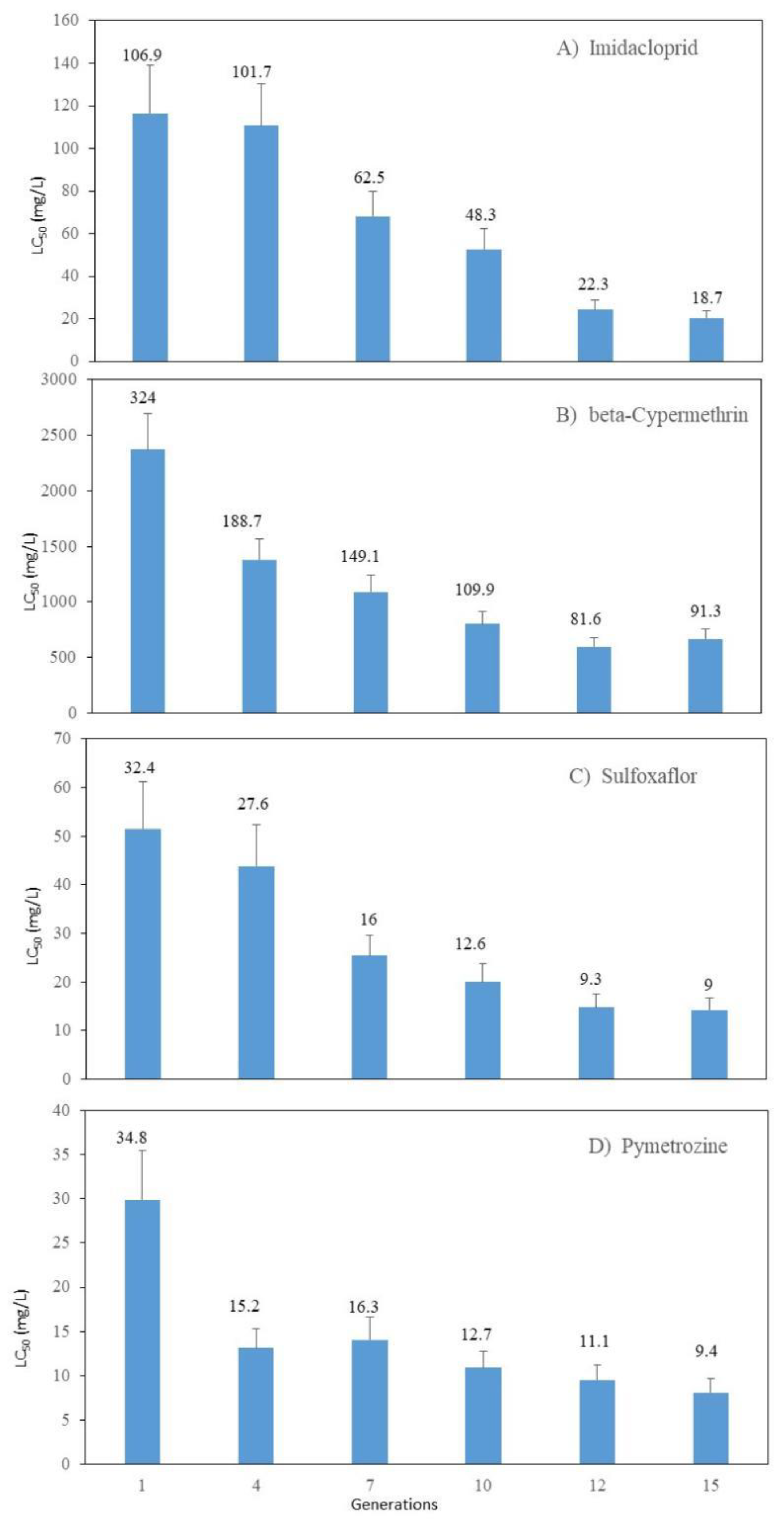
Figure 2.
Activities of detoxifying enzymes and protective enzymes in FFJ-S and SEF-R. The asterisk ** in the figureure indicates significant differences as determined by Student’s t-test (P < 0.01).
Figure 2.
Activities of detoxifying enzymes and protective enzymes in FFJ-S and SEF-R. The asterisk ** in the figureure indicates significant differences as determined by Student’s t-test (P < 0.01).
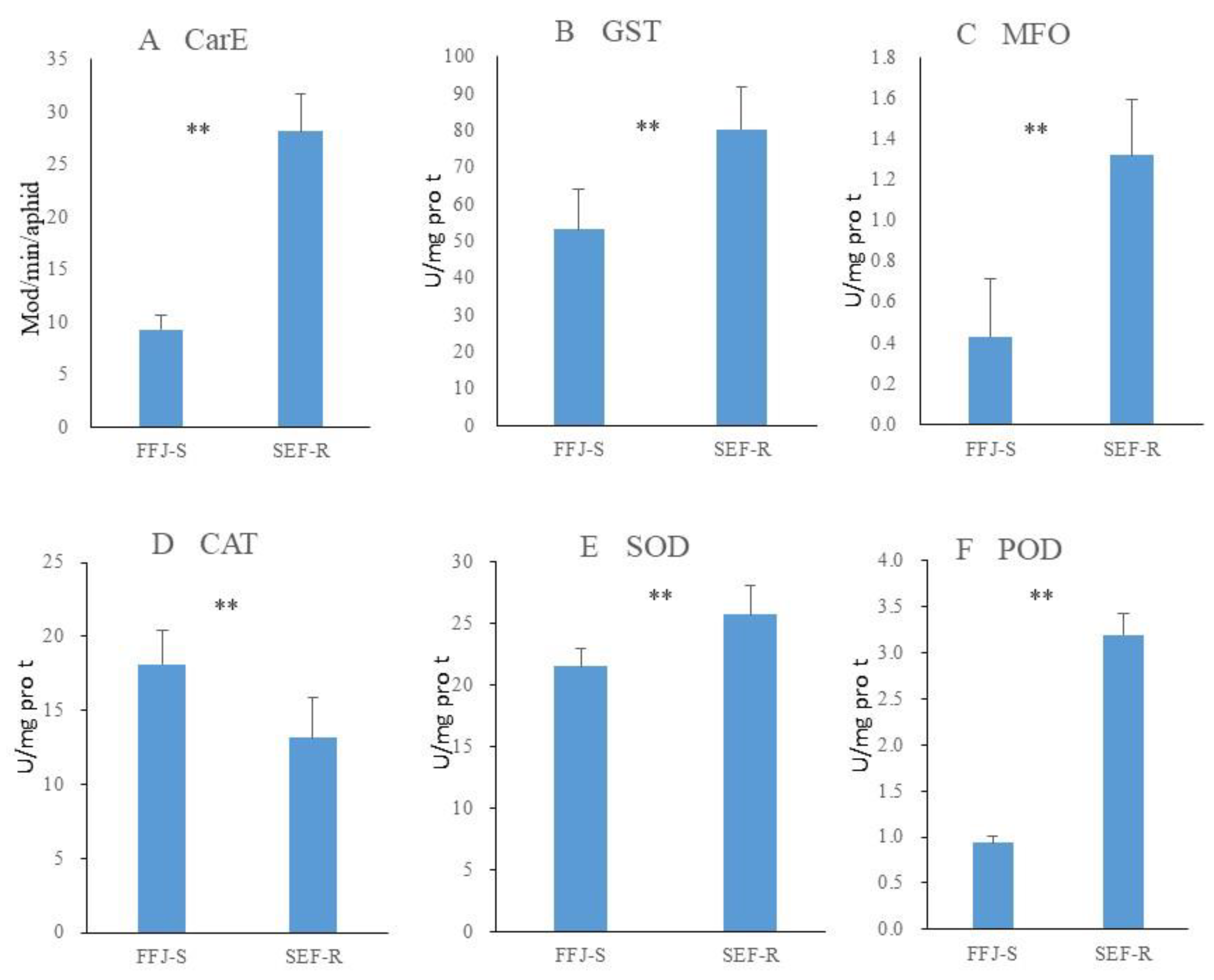
Figure 3.
Frequency of insecticide resistance genotypes (R81T, L1014F and M918T mutations) using the Taqman PCR assay. One hundred individual aphids were tested per genotype test. SS: wild type; RS: heterozygous resistant; RR: homozygous resistant.
Figure 3.
Frequency of insecticide resistance genotypes (R81T, L1014F and M918T mutations) using the Taqman PCR assay. One hundred individual aphids were tested per genotype test. SS: wild type; RS: heterozygous resistant; RR: homozygous resistant.
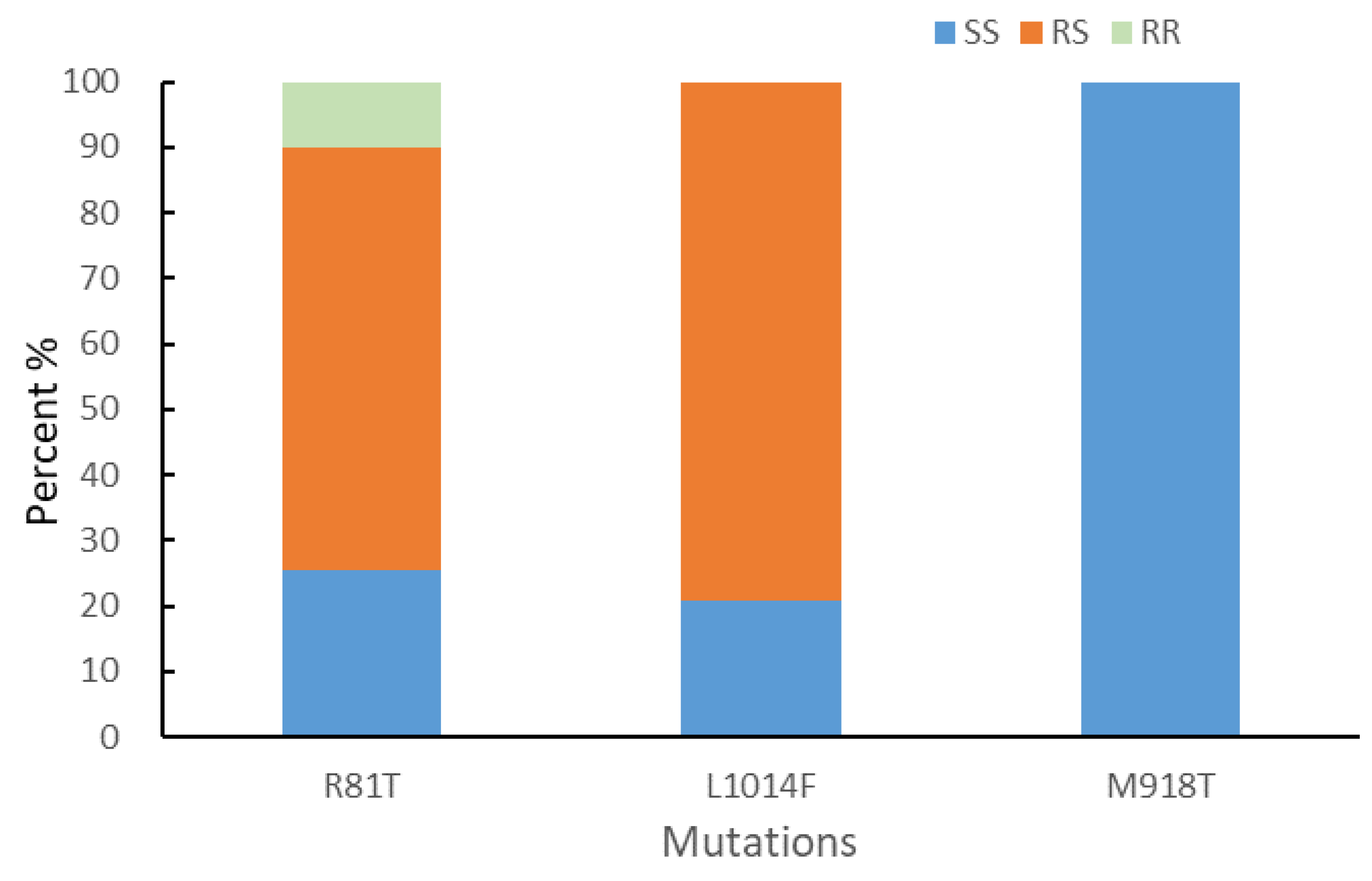
Figure 4.
Quantitative real-time PCR validation of the expression levels of differentially expressed P450 genes identified using RNA-sequencing. The expression levels were normalized to GAPDH and β-actin genes. The asterisk * in the Figure indicates significant differences as determined by Student’s t-test (P < 0.05). The asterisk ** in the Figure indicates significant differences as determined by Student’s t-test (P < 0.01).
Figure 4.
Quantitative real-time PCR validation of the expression levels of differentially expressed P450 genes identified using RNA-sequencing. The expression levels were normalized to GAPDH and β-actin genes. The asterisk * in the Figure indicates significant differences as determined by Student’s t-test (P < 0.05). The asterisk ** in the Figure indicates significant differences as determined by Student’s t-test (P < 0.01).
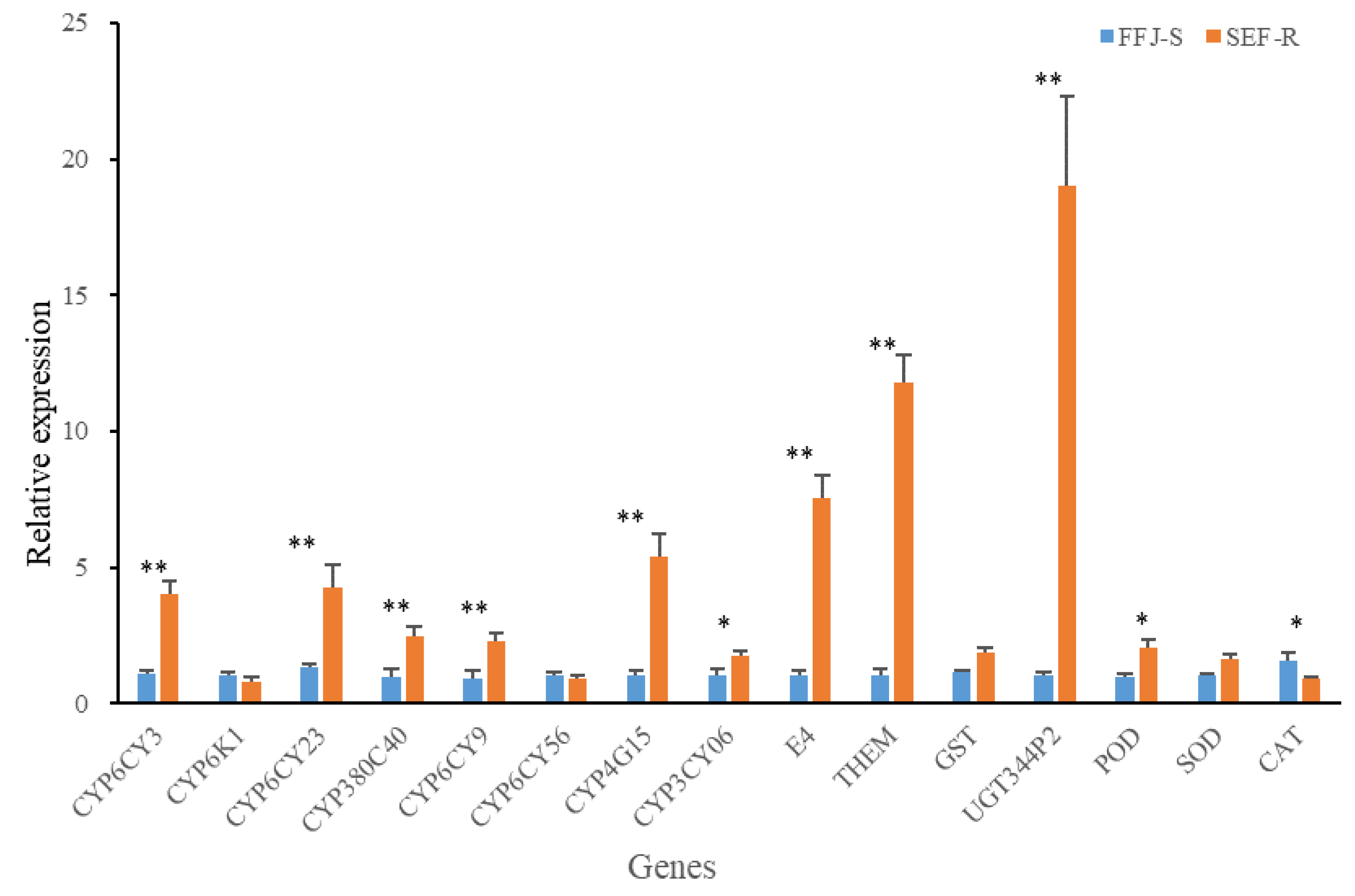
Figure 5.
Knockdown of CYP6CY3 increased the sensitivity of resistant aphids to imidacloprid, thiacloprid, thiamethoxam, sulfoxaflor, and flupyradifurone. (A) Relative expression levels of CYP6CY3, (B) (%) mortality at 48 h of dsRNA-fed adult GPA after treatment with imidacloprid (100 mg L−1), thiacloprid (40 mg L−1), thiamethoxam (30 mg L−1), sulfoxaflor (50 mg L−1) and flupyradifurone (10 mg L−1). The bars with lowercase letters (a, b) on top are significantly different according to the one-way analysis of variance followed by Tukey’s multiple comparison test (P < 0.05).
Figure 5.
Knockdown of CYP6CY3 increased the sensitivity of resistant aphids to imidacloprid, thiacloprid, thiamethoxam, sulfoxaflor, and flupyradifurone. (A) Relative expression levels of CYP6CY3, (B) (%) mortality at 48 h of dsRNA-fed adult GPA after treatment with imidacloprid (100 mg L−1), thiacloprid (40 mg L−1), thiamethoxam (30 mg L−1), sulfoxaflor (50 mg L−1) and flupyradifurone (10 mg L−1). The bars with lowercase letters (a, b) on top are significantly different according to the one-way analysis of variance followed by Tukey’s multiple comparison test (P < 0.05).
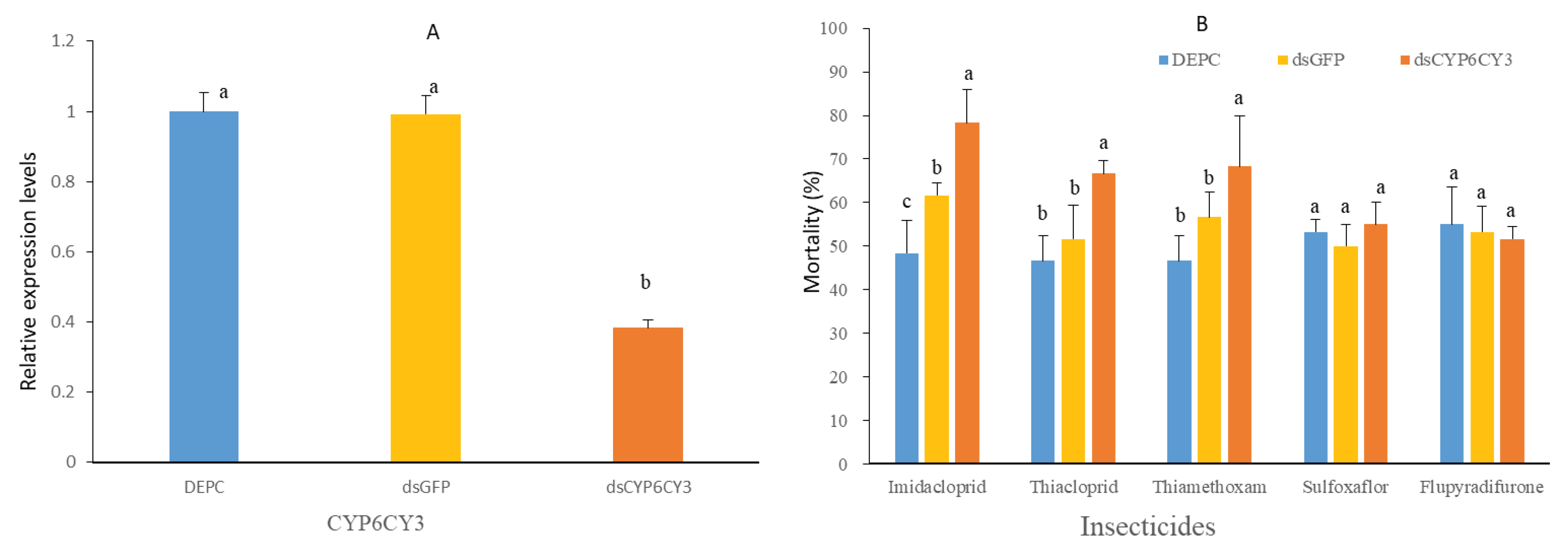

Table 1.
Cross-resistance of FFJ-S and SEF-R strains of GPA to 13 insecticides.
| Insecticides. | Strains | No. | Slope (SE) | LC50 (95%CI) mg·L-1 |
χ2 | RFa |
|---|---|---|---|---|---|---|
| Thiacloprid | FFJ-S | 480 | 4 (0.33) | 1.89 (1.74-2.06) | 3.32 (df=3) | |
| SEF-R | 560 | 1.57 (0.14) | 54.86 (45.64-66.52) | 3.04 (df=4) | 29 | |
| Imidacloprid | FFJ-S | 560 | 2.68 (0.21) | 1.09 (0.97-1.22) | 2.12 (df=4) | |
| SEF-R | 480 | 1.83 (0.18) | 116.48 (97.95-138.88) | 2.45 (df=3) | 106.9 | |
| Thiamethoxam | FFJ-S | 560 | 4.71 (0.36) | 2.57 (2.4-3.74) | 5.5 (df=4) | |
| SEF-R | 480 | 1.64 (0.18) | 33.2(27.37-40.49) | 2.61 (df=3) | 12.9 | |
| Sulfoxaflor | FFJ-S | 480 | 2.79 (0.25) | 1.59 (1.41-1.79) | 4.25 (df=3) | |
| SEF-R | 480 | 2.31 (0.26) | 51.51 (42.81-61.19) | 1.6 (df=3) | 32.4 | |
| Flupyradifurone | FFJ-S | 480 | 2.68 (0.24) | 1.34 (1.17-1.51) | 2.61 (df=3) | |
| SEF-R | 480 | 1.99 (0.19) | 12.67 (10.78-14.92) | 2.94 (df=3) | 9.5 | |
| Esfenvalerate | FFJ-S | 480 | 2.04 (0.19) | 9.66 (8.24-11.33) | 3.76 (df=3) | |
| SEF-R | 480 | 1.77 (0.17) | 764.4 (639.34-915.07) | 3.09 (df=3) | 79.1 | |
| beta-Cypermethrin | FFJ-S | 560 | 1.8 (0.15) | 7.32 (6.17-8.69) | 3.15 (df=4) | |
| SEF-R | 480 | 1.9 (0.18) | 2371.54 (2006.7-2823.87) | 0.64 (df=3) | 324 | |
| Flonicamid | FFJ-S | 560 | 2.02 (0.16) | 1.35 (1.15-1.57) | 2.47 (df=4) | |
| SEF-R | 480 | 1.79 (0.18) | 7.85 (6.55-9.42) | 2.53 (df=3) | 5.8 | |
| Pymetrozine | FFJ-S | 560 | 1.84 (0.15) | 0.86 (0.73-1.01) | 3.54 (df=4) | |
| SEF-R | 480 | 1.99 (0.19) | 29.89 (25.45-35.4) | 3.88 (df=3) | 34.8 | |
| Afidopyropen | FFJ-S | 480 | 2.03 (0.19) | 0.74 (0.62-0.87) | 2.38 (df=3) | |
| SEF-R | 480 | 2.1 (0.2) | 2.92 (2.49-3.44) | 2.81 (df=3) | 4 | |
| Spirotetramat | FFJ-S | 480 | 2.09 (0.19) | 1.05 (0.9-1.23) | 2.51 (df=3) | |
| SEF-R | 480 | 2.05 (0.19) | 8.45 (7.22-9.95) | 4.92 (df=3) | 8.1 | |
| Cyantraniliprole | FFJ-S | 480 | 1.95 (0.19) | 8.38 (7.07-9.87) | 3.55 (df=3) | |
| SEF-R | 480 | 1.81 (0.18) | 28.81 (24.21-34.55) | 0.94 (df=3) | 3.4 | |
| Broflanilide | FFJ-S | 560 | 1.78 (0.15) | 0.98 (0.83-1.17) | 4.88 (df=4) | |
| SEF-R | 480 | 1.82 (0.18) | 15.23 (12.81-18.27) | 0.43 (df=3) | 15.5 |
RF (resistance factor) = LC50 value of a field population/LC50 value of the Lab-SS strain.
Table 2.
Synergism of three enzyme inhibitors on imidacloprid toxicity to GPA.
| Treatment | No. | Slope±SE | LC50 (95%CI) | χ2 | SRb | |
|---|---|---|---|---|---|---|
| FFJ-S | IMDPa | 560 | 2.68 (0.21) | 1.09 (0.97-1.22) | 2.12 (df=4) | |
| IMDP+PBO | 480 | 3.18 (0.28) | 1.1 (.099-1.22) | 4.07 (df=3) | 0.99 | |
| IMDP+DEF | 480 | 3.3 (0.28) | 1.13 (1.02-1.25) | 5.08 (df=3) | 0.96 | |
| IMDP+DEM | 480 | 2.7 (0.26) | 1.01 (0.9-1.14) | 5.31 (df=3) | 1.08 | |
| SEF-R | IMDP | 480 | 1.83 (0.18) | 116.48 (97.95-138.88) | 2.45 (df=3) | |
| IMDP+PBO | 560 | 1.56 (0.14) | 27.99 (23.23-33.81) | 6.37 (df=4) | 4.16 | |
| 酯酶 | IMDP+DEF | 560 | 1.63 (0.16) | 54.13 (45.19-64.94) | 6.42 (df=4) | 2.15 |
| IMDP+DEM | 560 | 1.55 (0.14) | 36.36 (30.14-43.91) | 5.43 (df=4) | 3.2 |
a, IMDP: imidacloprid; b, SR (synergism ratio) = LC50 of imidacloprid/LC50 of imidacloprid with synergist.
Table 3.
Synergism of three enzyme inhibitors on beta-cypermethrin toxicity to GPA.
| Insecticides | Treatment | No. | Slope±SE | LC50 ((95%CI)) | χ2 | SRb |
|---|---|---|---|---|---|---|
| FFJ-S | CYPEa | 560 | 1.8 (0.15) | 7.32 (6.17-8.69) | 3.15 (df=4) | |
| CYPE+PBO | 560 | 1.75 (0.14) | 7.09 (5.98-8.42) | 3.26 (df=4) | 1.03 | |
| CYPE+DEF | 560 | 1.62 (0.14) | 7.1 (5.93-8.52) | 2.35 (df=4) | 1.03 | |
| CYPE+DEM | 560 | 1.82 (0.15) | 7.61 (6.43-9.03) | 3.74 (df=4) | 0.96 | |
| SEF-R | CYPE | 480 | 1.9 (0.18) | 2371.54 (2006.7-2823.87) | 0.64 (df=3) | |
| CYPE+PBO | 480 | 1.81 (0.18) | (309.07-439.99) | 3.6 (df=3) | 6.43 | |
| CYPE+DEF | 480 | 1.98 (0.18) | 617.18 (524.91-732.39) | 2.95 (df=3) | 3.84 | |
| CYPE+DEM | 480 | 2.1 (0.21) | 1241.92 (1054.77-1468.27) | 1.2 (df=3) | 1.91 |
a, CYPE: beta-cypermethrin; b, SR (synergism ratio) = LC50 of beta-cypermethrin/LC50 of beta-cypermethrin with synergist.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



