
Submitted:
02 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
Studies evaluating candidate tick-derived proteins as anti-tick vaccines in natural hosts have been limited due to high costs. To overcome this problem, animal models are used in immunization tests. The aim of this article was to review the use of rabbits as an experimental model for the evaluation of tick-derived proteins as vaccines. A total of 57 tick proteins have been tested for their immunogenic potential using rabbit as model for vaccination. The most commonly used rabbit breeds were New Zealand (73.8%), Japanese white (19%), Californians (4.8%) and Flemish lop-eared rabbit (2.4%). Anti-tick vaccines efficacy resulted in up to 99.9%. Haemaphysalis longicornis (17.9%) and Ornithodoros moubata (12.8%) were the most common tick model in vaccination trials. Experiments in rabbits have revealed that some proteins (CoAQP, OeAQP, OeAQP1, Bm86, GST-Hl, 64TRP, serpins and voraxin) can induce immune responses against various tick species. In addition, in some cases it was possible to determine that the vaccine efficacy in rabbits was similar to experiments performed in natural hosts (e.g. Bm86, IrFER2, RmFER2, serpins and serine protease inhibitor). In conclusion, results have shown that prior to performing anti-tick vaccination trials using natural hosts, rabbits can be used as suitable experimental models for these studies
Keywords:
rabbit
; immunization
; humoral and adaptive response
; tick
; antigen
1. Introduction
Ticks are obligate blood-sucking ectoparasites that parasitize a large number of terrestrial and semi-terrestrial vertebrates including humans [1,2,3]. Although they have been considered cosmopolitan parasites, most tick species are restricted to specific habitats, especially in tropical and subtropical regions [4,5]. Ticks transmit a wide variety of pathogens, being the second most important vectors of pathogens affecting humans, and the main vector in domestic and wild animals [6,7].
Traditional methods to control these arthropods are mainly based on the use of synthetic acaricides [8,9,10]. However, the application of these products has disadvantages, including the selection of resistant tick populations, environmental contamination, and residues in products of animal origin, such as milk and meat [11].
These issues raise the need to develop alternative control methods, including the selection of parasite-resistant breeds [12,13], biological control using entomopathogenic fungi (Metarhizium spp., Beauveria spp.) [14,15], entomopathogenic nematodes (Heterorhabditidae and Steinernematidae) [16,17], regulator ants (Solenopsis germinata, S. saevissima, Camponotus rengira, and Ectatomma quadridens) [18,19], and pesticides [20,21], as well as immunological control through the application of anti-tick vaccines [22,23,24].
Several proteins have been studied to date as candidates for the development of tick vaccines [25,26]. The immune response against target tick-derived proteins can affect tick’s biological functions like feeding, blood digestion, protein regulation, water transport, reproduction, embryogenesis, immune response and tick-pathogen interactions [27,28,29,30]. The first commercial anti-tick vaccine was obtained from the Bm86 protein [31]. The antigen hindered feeding and reproductive ability of the Rhipicephalus microplus [31,32], and was used in two leading tick vaccines, TickGARD® and GAVAC® [33]. This landmark result obtained by Willadsen et al. (1989) [31] paved the way for the investigation of new antigens and the development of vaccines that reduced infestations by R. microplus as well as other tick species.
The evaluation of tick vaccines in natural hosts has limitations mainly due to the high costs of maintaining and using farm or wild animals in experiments. For this reason, animal models such as hamsters, guinea pigs and rabbits are commonly used [34,35,36]. These animals have been used as models for basic and applied research, not only to test immune responses generated by anti-tick vaccines, but also to study resistance to chemical acaricides and tick-borne pathogens infection under laboratory conditions [37,38,39,40].
The use of hamsters, guinea pigs and rabbits in the tick vaccination experiments comes with generally low maintenance costs, little space requirements, short reproductive cycles and large numbers of pups produced per year compared to some natural hosts [41,42,43]. However, there are distinct benefits and disadvantages in each of these models. For instance, the use of hamsters is limited by low blood volume compared to guinea pigs and rabbits [44,45]. On the other hand, guinea pigs have thick skin, which makes blood collection relatively difficult, sometimes even requiring anesthetic techniques to collect small volumes, in contrast to rabbits, which do not require anesthetic techniques for blood collection [46].
Another limitation in experimental animal models is the number of ticks that can be used when performing the infestation. Studies in rabbits have reported these animals can supports a higher burden of adult ticks [23,47], compared to mice, hamsters or guinea pigs [48,49]. Interestingly, the rabbit model was the first animal model used in several immunological studies and was crucial, for example, for the development of Louis Pasteur's rabies vaccine in 1881 [50]. In 1976, the World Health Organization (WHO) [51] highlighted rabbit as one of the most important laboratory animals for the study of different diseases [51,52,53,54]. The most common breeds of laboratory rabbits are derived from the European rabbit (Oryctolagus cuniculus) [55]. The American Rabbit Breeders Association (ARBA) enlisted 30 rabbit breeds that are used for experiments [56], among which the most used is the New Zealand white rabbit [30,42,57].
Laboratory rabbits have proven to be the most suitable and accessible hosts for all life stages of various tick species during infestation and vaccination experiments [41,58]. This is because it has several advantages over the use of laboratory mice and rats, such as: (i) longer life span than mice and rats [59], (ii) larger body size (up to four times larger than rats), (iii) more blood volume, cell and tissue samples [60], (iv) production of copious antiserum [51,56], and (v) easy maintenance and breeding [56].
Historically, the evaluation of the tick-stimulated immune responses in rabbits began by studying the skin reactions caused by tick bites. A study by Trager observed that a single infestation of rabbits with Dermacentor variabilis larvae induced immunity that prevented subsequent larval infestations [61]. This work served as the basis for the subsequent use of rabbits as a model host for the development of anti-tick vaccines in the 1970s [62]. The main objective of this article reviews the current literature underscoring the importance of laboratory rabbits as an experimental model for the development of anti-tick vaccines, comparing the immune responses developed in rabbits and the natural host, and evaluating the vaccine efficacy against potential anti-tick antigens.
2. Vaccination in Rabbits
Rabbits are currently used as a model organism in anti-tick vaccines assays against ticks of the genera Amblyomma, Dermacentor, Hyalomma, Haemaphysalis, Ixodes, Ornithodoros, and Rhipicephalus (Figure 1) [23,35,63,64,65,66]. The following paragraphs will discuss the main trials carried out on rabbits for anti-tick vaccine development.
2.1. Haemaphysalis spp.
The tick Haemaphysalis longicornis tick is native to East Asia, and with sparse distribution in Australia, New Zealand and the USA [67,68]. It has a three-host life cycle, infesting cattle and wild animals such as ungulates, lagomorphs, carnivores and birds [69,70]. Immunological studies have shown different immunogenic proteins with potential to develop a vaccine against H. longicornis from China and Japan. Japanese white rabbit and New Zealand breeds were mostly used in the infestation experiments.
Eight proteins were evaluated for the purpose of vaccine development against H. longicornis using rabbit as an animal model (Figure 2). Wang et al. used New Zealand white rabbits to test the immune response against the lipocalin homologous protein of H. longicornis, obtaining a 60% reduction in the blood-feeding period of ticks, which would affect egg number, oviposition and hatching rate [24]. In an experiments Japanese white rabbits were immunized with protein 34 from H. longicornis, obtaining a partial reduction of this tick infestation [64,71].
2.2. Ornithodoros spp.
Ornithodoros erraticus and Ornithodoros moubata are nidicolous and endophilic argasid ticks that are widely distributed in different regions [72,73,74,75], and can intermittently feed on various vertebrates, such as birds and canines [76,77]. Eight tick-derived proteins have been evaluated for the development of vaccines against O. erraticus and O. moubata using rabbit as an animal model. Oleaga et al. tested the O. moubata ferritin 2 orthologues in New Zealand white rabbits, obtaining 71% efficacy for OmFer2, corresponding to decreased egg hatching rate and subsequent number of emerging O. moubata larvae [78]. On the other hand, Pérez-Sánchez´s research group tested the immune response against aquaporin, showing moderate vaccine efficacy against O. erraticus [35].
A study carried out by Manzano-Román used the New Zealand rabbits breed to test the protective effect induced by the recombinant subolesin proteins against O. erraticus and O. moubata. The results showed a higher reduction in O. erraticus oviposition when compared with O. moubata [79]. Manzano-Román et al. used the same rabbits breed to verify the vaccine efficiency of the synthetic peptides of subolesin/akirin, obtaining 83.1% vaccine efficacies against O. erraticus (Figure 3) [80]. These results show that rabbits presented anti-O. erraticus antibodies that recognized both subolesin proteins, and protected against both argasid ticks with comparable efficacy.
2.3. Rhipicephalus spp.
Rhipicephalus appendiculatus, Rhipicephalus microplus, and Rhipicephalus sanguineus s.l. are medically important ixodid ticks of the genus Rhipicephalus [81]. Rhipicephalus appendiculatus is distributed in central, eastern and southeastern Africa [82,83]. Rhipicephalus microplus and R. sanguineus s.l. are cosmopolitan ticks, distributed in the tropical and subtropical regions of the globe [6,84]. They present monoxene (R. microplus) and hetorexone (R. sanguineus s.l. and R. appendiculatus) life cycles, preferring domestic hosts such as bovines, canines and some wild animals. They feed on humans as incidental hosts [84,85].
New Zealand white, Californians, Japanese white and Flemish lop-eared rabbit breeds have been studied as model hosts for R. appendiculatus, R. microplus, and R. sanguineus s.l. in experiments carried out in Brazil, Cuba, Kenya, Japan and Mexico, proving to be successfully used in evaluating anti-tick proteins. Regarding the evaluation of proteins for the development of vaccines against Rhipicephalus species using rabbit as an animal model. To date, a total of 15 molecules have been evaluated in rabbits for vaccine development against Rhipicephalus species, being the most frequent tick genus in this kind of study.
Tthe voraxin α homologue of the R. appendiculatus tick was used to immunize the Japanese white rabbits, which resulted in a reduction in the weight of ticks, followed by a 50% reduction in egg mass [86]. On the other hand, a different study determined the vaccinal efficacy of rGST in New Zealand white rabbits, showing that rGST caused a reduction in the number of female R. sanguineus s.l. infestation [28].
Parizi et al. using the New Zealand white rabbit for immunization with the R. microplus cystatin 2c, reported a reduction in the number of fully engorged adult female ticks, causing damage to R. appendiculatus tissues such as intestine, salivary glands and ovary [23]. A study used Californian breed rabbits, immunized with the P0 protein, and demonstrated a 90% reduction efficacy against R. sanguineus s.l., observing a decrease in nymphs and larvae fed on vaccinated rabbits [57]. In tum, Jittapalapong et al. determined the vaccine efficacy of recombinant R. microplus salivary gland serpin in New Zealand rabbits, obtaining an 83% reduction in adult R. microplus engorgement compared to the control [88]. These results indicated that this tick serpin is immunogenic for rabbits and suggested that this vaccine candidate antigen may confer protective immunity against the cattle ticks in this experimental model.
Also for the cattle tick, Lagunes-Quintanilla et al. initially studied the recombinant peptide derived from the serpin RmS-17 protein as a vaccine in the New Zealand rabbit breed, to later vaccinate cattle. The results showed that the experimental vaccination reached 79% efficacy, limiting the number of infested adult ticks, oviposition and fertility of the eggs [47]. Rabbits developed a strong humoral immune response expressed by high levels of anti-RmS-17 IgG. This was the first study evaluating the efficacy of the RmS-17 peptide against infestation by R. microplus ticks and demonstrating that it is immunogenic and protective in a rabbit model. A study used New Zealand rabbits to verify the vaccinal efficacy of the 50-kDa protein from Rhipicephalus haemaphysaloides, reporting a 74.7% protection in feeding ticks just 24 hours after infestation [89]. The protein stimulated a specific protective immune response in tick-infested rabbits, demonstrating the success of rabbits as an animal model for these tick experiments.
2.4. Ixodes spp.
Ixodes ricinus and Ixodes scapularis are ixodid ticks that are characterized by presenting a heteroxenous life cycle, infesting cattle, deer, dogs, and a wide variety of vertebrates, including humans [90,91,92]. The nymphal stage is the most frequently responsible for transmitting pathogens to humans [93,94]. Of the 265 species of Ixodes, 55 are distributed in the Neotropical region of the planet [5], however, I. ricinus and I. scapularis can be found only in the northern hemisphere [90]. Vaccination studies against I. ricinus and I. scapularis using the New Zealand rabbit breed have been reported in the USA, Spain and the Netherlands (Figure 4) [48,95,96].
Five proteins were tested for the development of vaccines against I. ricinus and I. scapularis using rabbits as an animal model. For instance, Contreras and de la Fuente evaluated the efficacy of CoAQP proteins against I. ricinus infesting New Zealand white rabbits, obtaining an efficiency of 80% [96]. Meanwhile, Schuijt et al. evaluated P8, P19 and P23 proteins of I. scapularis in New Zealand white rabbits, demonstrating a reduction of feeding by infested nymphs in rabbits immunized with the cocktail antigens [95].
Contreras and de la Fuente used the New Zealand rabbits as a vaccination model to describe the effect of the Q38 chimeric protein that conserved protective epitopes from I. ricinus. The vaccine had an efficacy of 99.9% in the reduction of I. ricinus larvae of, with a cumulative effect in the reduction of the tick survival and molting to the next life stage [39]. Also, vaccination with I. ricinus recombinant protein ferritin 2 significantly reduced the number, weight and fertility of ticks in vaccinated rabbits infested with I. ricinus, with overall 98% vaccine efficacy [97]. These results demonstrated the feasibility of using ferritin 2 to develop vaccines to control tick infestations. A study showed that recombinant forms of the tick cement antigen R. appendiculatus 64P, act as a "double-acting" tick vaccine for I. ricinus, providing cross-protection for this ixodid tick, possibly by attacking antigens in the midgut and salivary glands of adults and nymphs. The tick mortality rate was 60%, and the results indicated the potential of 64TRPs as a broad-spectrum tick vaccine [48].
2.5. Dermacentor spp.
Dermacentor marginatus is an ixodid tick that has a heteroxenous life cycle and a variety of hosts including canines, horses and humans [98,99]. It is a tick with a cosmopolitan distribution, present mainly in the Nearctic, Palearctic and Netotropic ecozones of the planet [100,101,102]. In the search for proteins for the development of a vaccine against D. marginatus, the New Zealand white rabbit has been used as an animal model in infestations and vaccination experiments. A study infested New Zealand breed rabbits with D. marginatus after administering the last dose of the immunogen of GST, recording moderate vaccine efficacy against D. marginatus (Figure 5 and Table 1) [30].
3. Discussion
To date, 57 tick-derived proteins have been evaluated as potential anti-tick vaccines by studying the immunogenic responses generated using rabbits as an experimental model. Rabbit models for anti-tick vaccination trials have allowed a better understanding of the physiological mechanisms of ticks infesting mammal hosts. For example, the study of the serpins HLS1, rHLS2, rSerpin, and RmS-17 in rabbits stimulated an immune response that affected the prolonged duration of feeding, increased mortality and reduced oviposition in ticks like H. longicornis and R. microplus [47,64,88,103].
Globally, the use of rabbits has provided novel evidence on a vaccine based on salivary glycine-rich proteins in various medically important tick species. According to the findings obtained by Zhou et al. using rabbits immunized with the glycine-rich protein RH50, this protein was only expressed in salivary glands of partially fed ticks, not in salivary glands of unfed ticks or in the midgut, fat body, or ovary of partially fed ticks, in contrast to what is reported for p29 and Bm86 proteins [63,89,120].
Rabbits have been used as an immunization model to evaluate immunological response to a given antigen (Q38, Bm86, GST, serpins and voraxin) against different tick species. For example, high vaccine efficacy against both I. ricinus and D. reticulatus has been obtained with the chimeric protein Q38 containing subolesin/akirin [39].
Similarly, experiments in rabbits using voraxin α, a protein derived from the male tick and transferred to the female through copulation to stimulate female blood feeding [105], have yielded vaccine efficiency by reducing feeding time in Amblyomma hebraeum. There is amino acid sequence similarity between the voraxin α of A. hebraeum (85%) and that of D. variabilis (92%) and R. appendiculatus (85%) [86]. The immunization results could therefore potentially be similar, making this protein a good multispecies vaccine candidate. By reducing feeding time of ticks, it would also reduce salivation and consequently pathogen transmission, in addition to impairing oocyte development [105].
The use of rabbits as animal models in the discovery of anti-tick molecules has been fundamental in enabling testing of these molecules before inoculation into the natural hosts. It has been verified that rabbits present an immune response similar to that obtained in the natural hosts. For example, the use of the ferritin 2 protein to immunize rabbits infested with I. ricinus (IrFER2) yielded an efficiency of 98%, while the efficiency of the same protein used in bovines infested with R. microplus and R. annulatus (RmFER2) was 64% and 72%, respectively [97]. Also, recombinant peptides derived from serpins have shown efficacies against R. microplus between 67% and 79% in rabbits [47,88], while in bovines, this protein has offered an efficacy of 67% against R. appendiculatus [121].
Studies carried out with the Bm86 antigen in rabbits and cattle have shown that rabbits and cattle have a very similar reduction efficacy against R. microplus, obtaining a 62% reduction in rabbits [47], and 60% in cattle [97]. These results further indicate that rabbits are an excellent experimental model for initial vaccination experiments with anti-tick antigens, prior to the application of these in the natural hosts such as bovines. Another benefit that has been obtained by using rabbits as an animal model in research is the high recovery rate of fully engorged individuals of different tick species when carrying out infestations, compared to non-definitive domestic hosts. An experiment using R. microplus obtained a recovery of 33% in rabbits [122], compared to 3.7% in goats [123], 0.4% in dogs [124], and 1.8% in horses [125].
On the other hand, in a vaccination experiment in rabbits, Canales et al. reported for the first time that the recombinant bacterial membrane fraction containing the BM95-MSP1a chimera was effective for the control of R. microplus infestations. The BM95-MSP1a vaccine reduced oviposition and fertility of R. microplus similarly to the commercial vaccine Bm86, having a significantly greater immune response in vaccinated rabbits compared to controls [107]. The results obtained in this experiment demonstrated that rabbit is an excellent animal model to continue exploring new techniques or novel anti-tick antigens.
The infestation time of the R. microplus nymphal to adult stages on rabbits takes on average of 30 days to complete [126], while the time it takes to complete these two life stages in bovines, it is between 14 and 20 days on average [127]. These data indicate that the infestation of some tick species such as R. microplus in rabbits could be a valuable alternative animal model for evaluation of candidate vaccines and new molecules with acaricidal activity against this ectoparasite. A possible analysis of why the life cycle takes longer in the nymphal to adult life stages of R. microplus in rabbits could be due to the inflammatory cellular response caused by tick bites, at the beginning of feeding, preventing them from accessing the blood source, which can lead to increased mortality and feeding time [128]. Another aspect could be the strong competition between ticks due to the little physical body space for feeding provided by rabbits as a feeding model, which can cause the death of some ticks by not being able to adhere to the skin at the beginning of the infestation [129,130].
Rabbits immunized with 64TRP and infestated by R. sanguineus s.l. or I. ricinus developed local inflammatory immune responses, involving leukocytes, basophils, eosinophils, lymphocytes, mast cells, and macrophages. In turn, bovines immunized with 64TRP and challenged with R. appendiculatus showed dermal migration of dendritic cells, actively degranulating mast cells, basophils, and eosinophils [48]. These authors also found that the GST protein has generated very similar inflammatory responses (mainly caused by eosinophils or mast cells) in hamsters, rabbits and bovines [48]. Furthermore, infestation-only studies revealed a similar immune response against R. appendiculatus, with infiltration of neutrophils, macrophages, eosinophils and basophils, in both rabbits and cattle [54].
The immune responses generated by the different proteins studied in rabbits could vary depending on the challenges being made with ticks in immature or mature life stages. For example, the response generated by the p29 and HL34 proteins in the life stages of larvae, nymphs and adults of H. longicornis fed on immunized rabbits suggest that these proteins may be involved in mediating key physiological functions in the tick [63,71]. Although mature and immature ticks commonly express native p29, their sensitivities to rabbit immune response against rp29 appear to be different [63], while the native HL34 is expressed in both immature (larvae and nymphs) and adult ticks. It is thus likely that immunity against rHL34 is directed against immature and mature ticks [71].
This result can be supported by Kemp et al. who recorded that R. microplus in immature and mature states have different sensitivities to acquire resistance against anti-tick molecules. While there was severe intestinal damage in adult females and males feeding on cattle infested with R. microplus-derived antigens, there was no effect on tick larvae feeding on the same protected cattle [120]. Therefore, we can suggest that the different vaccine effects between immature and mature H. longicornis ticks fed on rp29-vaccinated rabbits could be consistent as well in natural hosts.
Additionally, studies in rabbits have allowed us to broaden our knowledge about "exposed" and “hidden” antigens of anti-tick proteins. For example, it has been reported that HLS1 acts on the expression of hidden antigens, inhibiting the secretion of rHLS1 in rabbits during feeding [64]. Also, 64TRP isoforms have been characterized as "dual-acting" anti-tick proteins against R. sanguineus s.l. and I. ricinus: they target both "exposed" and "hidden" antigens, preventing attachment and feeding by affecting the feeding site, as well as cross-reacting with 'hidden' midgut antigens, resulting in the death of engorged ticks [48].
Only a few studies focus on identifying molecules that affect male feeding or reproduction. One of the proteins that has been identified and tested in rabbits is voraxin [105]. Preliminary vaccination of rabbits with rvoraxin α has demonstrated humoral immunity and conferred protective immunity against female R. appendiculatus ticks, resulting in reduced feeding weight [86]. This may indicate that the antibodies against voraxin α effect female ticks of the same species. This same result has been observed in the female A. hebraeum, which has been studied in infestations in rabbits immunized with voraxin α, obtaining a reduction in feeding of up to 72% compared to engorged ticks from control rabbits [105]. These results suggest that rabbits may be a good model not only for the study of anti-tick molecules that act on females, but also for those molecules that act on males.
Results obtained from the study of the tick saliva proteome have shown a variety of proteins that protect ticks against host immune responses and antihemostatic mechanisms [131,132,133,134,135,136]. This is because during hematophagy, tick salivary glands undergo remarkable growth and differentiation, accompanied by a significant increase in the synthesis of different proteins [137]. Tirloni et al. identified 187 tick and 68 bovine proteins in the saliva proteome of R. microplus, demonstrating that R. microplus saliva is rich in hemolipoproteins, lipocalins, peptidase inhibitors, antimicrobial peptides, glycine, and maintenance proteins [133]. These proteins, together with pharmacological bioactive lipids, can counteract the host's defenses and hemostatic mechanisms [138,139], while the host physiological systems can trigger changes in the feeding activity of ticks [140] by stimulating proteins to limit the host defense mechanisms [141].
Another study by Tirloni et al. looked at the saliva proteome of non-fed adult ticks of I. scapularis and A. americanum stimulated in different hosts, including rabbits, dog and human, identifying a total of 276 proteins in I. scapularis and 340 proteins in A. americanum. Among these proteins, 55 (I. scapularis) and 67 (A. americanum) belonged to the same functional classes [142]. These data suggest that A. americanum and I. scapularis used a core set of functionally similar proteins that regulated key host defense pathways to successfully feed. I. scapularis saliva had a high abundance of proteins related to heme/iron metabolism, followed by extracellular matrix/cell adhesion, oxidative metabolism/detoxification, cytoskeletal metabolism, proteasome machinery, nuclear regulation, conserved protein with unknown function, modification proteins, protein synthesis machinery proteins, and transport/storage. In turn, A. americanum saliva had a high abundance of extracellular matrix/cell adhesion proteins and proteinase inhibitors, followed by immune-related heme/iron metabolism, energy protein metabolism, cytoskeletal, protein synthesis machinery and proteasome machinery [142].
The above results indicated that these two tick species could inject the same protein at different levels into different hosts, and that the protein composition in the saliva of different tick species feeding on the same host is likely to be different. Furthermore, they suggest that ticks of the same species differentially express tick salivary proteins when stimulated to start feeding on different hosts, expressing unique protein profiles in their saliva. There is evidence that ticks differentially express specific sets of genes when stimulated to start feeding [143,144]. For example, A. americanum saliva proteins contain a diversity of protease inhibitors (PI), expressing a total of 155 PI proteins belonging to eight families. Approximately 74% of these PI (115/155) were secreted into saliva within the first 120 h of feeding, indicating that the functions of the PI are associated with the regulation of the early stages of feeding in A. americanum which could also include the transmission of TBD agents by A. americanum [145].
On the other hand, Tirloni et al. identified differences in the expression of proteins in the development stages of nymphs and adult females of H. longicornis, obtaining 30 proteins in the saliva of nymphs, 74 proteins in the saliva of fully engorged adult females, and 31 proteins that were detected at both stages [134]. Proteins expressed in adult saliva may be related to exposure to different vertebrate hosts and the different stages of development, leading to changes in the dynamics of salivary transcription [132,146]. Taken together, those studies demonstrate that, even though the protein profile of tick saliva is strongly influenced by the host they infest, rabbits can be used as an alternative non-natural host to continue exploring and describing proteins that serve as candidates for tick vaccines
4. Conclusions
The current review summarized the evaluation of 57 antigens as anti-tick vaccines in different rabbit breeds. These breeds include New Zealand, Japanese white, Flemish lop-eared, and California rabbits. The most widely used is the New Zealand breed, in countries located in Africa, Asia, America, Oceania, and Europe. Rabbits are not the natural host of most tick species, however this has not been a limitation to obtain vaccination results very similar to those on natural hosts for different tick species. Likewise, the use of rabbits has provided valuable insight on the immunological responses generated by novel antigens prior to vaccination trials in natural hosts. Rabbits stand out among other animal models used in vaccination experiments because they are suitable and commercially accessible alternative hosts for challenging the larval, nymphal, and adult life stages of various tick species.
Author Contributions
Conception and literature search, writing, review and editing, A.R. and L.F.; Conceptualization, A.R and I.D..; writing, review and editing, A.R., L.F, S.U., A.A., and I.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grants from the Instituto Nacional de Ciência e Tecnologia em Entomologia Molecular of the Brazilian Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, National Council for Scientific and Technological Development; grant no. 465678/2014-9), the Brazilian National Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Office for the Advancement of Higher Education; grant no. 88881.068421/2014-01), the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ, Carlos Chagas Filho Foundation for the Support of Research in the State of Rio de Janeiro; grant no. E-26/210.012/2018; SEI-260003/001743/2023), and the Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS, Foundation for the Support of Research in the State of Rio Grande do Sul; grant no. 21/2551-0002221-3) and Ministerio de Ciencia Tecnología e Innovación de Colombia number 885/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brisola, C. Mites (ticks and others). In Medical and veterinary entomology; Publishing Athens, 2011; pp. 263–315. [Google Scholar]
- Alcantara, E.; Ferreira da Silva, C.; Ávila, R.; Pacheco, R.; Muñoz, L.; Honorio, D. Ticks (Acari: Argasidae and Ixodidae) infesting amphibians and reptiles in northeastern Brazil. Syst. Appl. Acarol. 2018, 23, 1497. [Google Scholar] [CrossRef]
- Santos, M.; Bahiense, T.; Silva, A.; Onofrio, V.; Barral, T.; Souza, B.; Lira-da-Silva, R.; Biondi, I.; Meyer, R.; Portela, R. Ticks and associated pathogens from rescued wild animals in rainforest fragments of northeastern Brazil. Front. Vet. Sci. 2020, 7, 177. [Google Scholar] [CrossRef]
- Cortés-Vecino, J. Changes in the distribution and abundance of ticks and their relationship with global warming. J. Vet. Med. Zoot. 2010, 57, 65–75. [Google Scholar]
- Guglielmone, A.; Nava, S.; Robbins, R. Geographic distribution of the hard ticks (Acari: Ixodida: Ixodidae) of the world by countries and territories. Zootaxa 2023, 5251, 1–274. [Google Scholar] [CrossRef]
- Jongejan, F.; Uilenberg, G. The global importance of tick. Parasitology 2004, 129, 3–14. [Google Scholar] [CrossRef]
- de la Fuente, J.; Estrada-Pena, A.; Venzal, J.; Kocan, K.; Sonenshine, D. Overview: ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2007, 13, 6938–6946. [Google Scholar] [CrossRef]
- Abbas, R.; Zaman, M.; Colwell, D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: the state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. Expert consultation on the sustainable management of parasites in livestock challenged by the global emergence of resistance - Part 1: Current status and management of acaricide resistance in livestock ticks. FAO Animal Production and Health Report No. 17; Rome, 2022; pp. 9–10. [Google Scholar] [CrossRef]
- Obaid, M.; Islam, N.; Alouffi, A.; Zeb, A.; da Silva Vaz, I.; Tetsuya, T.; Abid, A. Acaricides resistance in ticks: selection, diagnosis, mechanisms, and mitigation. Front. Cell. Infect. Microbiol. 2022, 12, 941831. [Google Scholar] [CrossRef]
- Willadsen, P. Tick control: thoughts on a research agenda. Vet. Parasitol. 2006, 138, 161–168. [Google Scholar] [CrossRef]
- Mapholi, N.; Maiwashe, A.; Matika, O.; Riggio, V.; Banga, C.; MacNeil, M.; Dzama, K. Genetic parameters for tick counts across months for different tick species and anatomical locations in South African Nguni cattle. Trop. Anim. Health Prod. 2017, 49, 1201–1210. [Google Scholar] [CrossRef]
- Porto-Neto, L.; Reverter, A.; Prayaga, K.; Barendse, W. The genetic architecture of climatic adaptation of tropical cattle. PloS One 2014, 9, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Chi, M.; Rodriguez-Vivas, R.; Galindo-Velasco, E.; Lezama-Gutierrez, R. Laboratory and field evaluation of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) for the control of Rhipicephalus microplus (Acari: Ixodidae) in the Mexican tropics. Vet. Parasitol. 2010, 170, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, L.; Mesquita, E.; Almeida, T.; de Oliveira, R.; Oliveira, J.; Fernandes, F.; Guedes, M.; Pinheiro, E. Encapsulation of entomopathogenic fungal conidia: evaluation of stability and control potential of Rhipicephalus microplus. Ticks Tick Borne Dis. 2023, 14, 102184. [Google Scholar] [CrossRef]
- Freitas-Ribeiro, G.; Furlong, J.; Vasconcelos, V.; Dolinski, C.; Ribeiro, A. Analysis of biological parameters of Boophilus microplus Canestrini, 1887 exposed to entomopathogenic nematodes Steinernema carpocapsae Santa Rosa and ALL strains (Steinernema: Rhabditidae). Braz. Arch. Biol. Technol. 2005, 48, 911–919. [Google Scholar] [CrossRef]
- de Oliveira, C.; da Silva Matos, R.; Xavier, L.; de Souza, W.; Rita, V.; Pinheiro, E.; Dolinski, C.; de Azevedo, C. First report of pathogenicity of entomopathogenic nematodes of the genus Heterorhabditis on partially engorged females of Dermacentor nitens (Acari: Ixodidae). Biol. Control 2014, 69, 78–81. [Google Scholar] [CrossRef]
- Zingg, S.; Dolle, P.; Voordouw, M.; Kern, M. The negative effect of wood ant presence on tick abundance. Parasit. Vectors 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Platts-Mills, T.; Retterer, M.; Workman, L.; Wilson, J. A consistent “shortage” of cases of the alpha-gal syndrome (AS) on the gulf coast: possible relevance of fire ants as a predator of lone star ticks. J. Allergy Clin. Immunol. 2019, 143, AB278. [Google Scholar] [CrossRef]
- Adenubi, O.; Ahmed, A.; Fasina, F.; McGaw, L.; Eloff, J.; Naidoo, V. Pesticidal plants as a possible alternative to synthetic acaricides in tick control: A systematic review and meta-analysis. Ind. Crops. Prod. 2018, 123, 779–806. [Google Scholar] [CrossRef]
- Charlie-Silva, I.; Giglioti, R.; Magalhaes, P.; Sousa, I.; AnnFoglio, M.; Oliveira, M.; Chagas, A. Lack of impact of dietary inclusión of dried Artemisia annua leaves for cattle on infestation by Rhipicephalus (Boophilus) microplus tick. Ticks Tick Borne Dis. 2018, 9, 1115–1119. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M. Tick vaccines: current status and future directions. Expert Rev. Vaccines 2015, 14, 1367–1376. [Google Scholar] [CrossRef]
- Parizi, L.; Konrdörfer, C.; Alves, G.; Fagundes, B.; Kiio, I.; Amaral, M.; da Silva, R.; Camargo-Mathias, M.; Seixas, A.; Konnai, S.; Ohashi, K.; Wangombe, N.; da Silva Vas, I. Rhipicephalus microplus cystatin as a potential cross-protective tick vaccine against Rhipicephalus appendiculatus. Ticks Tick Borne Dis. 2020, 11, 101378. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lihong, L.; Pinxing, W.; Hongmeng, D.; Shuwen, X.; Jingze, L.; Yonghong, H. Gene cloning, analysis and effect of a new lipocalin homologue from Haemaphysalis longicornis as a protective antigen for an anti-tick vaccine. Vet. Parasitol. 2021, 290, 109–358. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ahmad, S.; de Albuquerque, P.; Kamil, A.; Alshammari, F.; Alouffi, A.; da Silva Vaz, I. Prediction of novel drug targets and vaccine candidates against human lice (Insecta), Acari (Arachnida), and their associated pathogens. Vaccines 2022, 10, 8. [Google Scholar] [CrossRef]
- Yadav, N.; Upadhyay, R. European journal of biological research tick saliva antigen-based vaccines, disease protection and prophylaxis. Eur. J. Biol. Res. 2020, 12, 77–101. [Google Scholar] [CrossRef]
- de la Fuente, J.; Kopáček, P.; Lew-Tabor, A.; Maritz -Olivier, C. Strategies for new and improved vaccines against ticks and tick-borne diseases. Parasite Immunol. 2016, 38, 754–769. [Google Scholar] [CrossRef] [PubMed]
- Ndawula, C.; Alves, G.; Parizi, L.; da Silva Vaz, I. Constituting a glutathione S-transferase-cocktail vaccine against tick infestation. Vaccine 2019, 37, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, B.; Han, Q. Understanding tick biology and its implications in anti-tick and transmission blocking vaccines against tick-borne pathogens. Front. Vet. Sci. 2020, 7, 319–456. [Google Scholar] [CrossRef] [PubMed]
- Huercha, R.; Min, L.; Xinli, F.; Zhengxiang, H.; Lijiang, W.; Yongchang, L.; Wei, Z.; Yang, Z.; Yuhui, M.; Chahan, B. Characterization of glutathione S-transferase of Dermacantor marginatus and effect of the recombinant antigen as a potential anti tick vaccine. Vet. Parasitol. 2020, 279, 109043. [Google Scholar] [CrossRef]
- Willadsen, P.; Riding, G.; McKenna, R.; Kemp, D.; Tellam, R.; Nielsen, J.; Gough, J. Immunologic control of a parasitic arthropod. Identification of a protective antigen from Boophilus microplus. J. Immunol. 1989, 143, 1346–1351. [Google Scholar] [CrossRef]
- Johnston, L.; Kemp, D.; Pearson, R. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Effects of induced immunity on tick populations. Int. J. Parasitol. 1986, 16, 27–34. [Google Scholar] [CrossRef]
- Willadsen, P.; Smith, D.; Cobon, G.; McKenna, R. Comparative vaccination of cattle against Boophilus microplus with recombinant antigen Bm86 alone or in combination with recombinant Bm91. Parasite Immunol. 1999, 18, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Gomes, H.; Moraes, J.; Githaka, N.; Martins, R.; Isezaki, M.; da Silva Vaz, I.; Logullo, C.; Konnai, C.; Ohashi, K. Vaccination with cyclin-dependent kinase tick antigen confers protection against Ixodes infestation. Vet. Parasitol. 2015, 211, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, R.; Manzano-Román, R.; Obolo-Mvoulouga, P.; Oleaga, A. Function-guided selection of midgut antigens from Ornithodoros erraticus ticks and an evaluation of their protective efficacy in rabbits. Vet. Parasitol. 2019, 272, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lynn, G.; Černý, L.; Kurokawa, C.; Diktaş, H.; Matias, J.; Sajid, A.; Arora, G.; DePonte, K.; Narasimhan, S.; Fikrig, E. Immunization of guinea pigs with cement extract induces resistance against Ixodes scapularis ticks. Ticks Tick Borne Dis. 2022, 13, 10201. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Chen, Z.; Liu, A.; Ren, Q.; Liu, J.; Liu, Z.; Li, Y.; Yin, H.; Guan, G.; Luo, J. Biological parameters of Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) fed on rabbits, sheep, and cattle. Korean. J. Parasitol. 2016, 54, 301–305. [Google Scholar] [CrossRef]
- Colby, L.; Quenee, L.; Zitzow, L. Considerations for infectious disease research studies using animals. Comp Medi. 2017, 67, 222–231. [Google Scholar]
- Contreras, M.; de la Fuente, J. Control of Ixodes ricinus and Dermacentor reticulatus tick infestations in rabbits vaccinated with the Q38 subolesin/akirin chimera. Vaccine 2016, 34, 3010–3013. [Google Scholar] [CrossRef]
- Stokes, J.; Walker, D.; Varela-Stokes, A. The guinea pig model for tick-borne spotted fever rickettsioses: A second look. Ticks Tick Borne Dis 2020, 11, 101538. [Google Scholar] [CrossRef]
- Burkholder, T.; Linton, G.; Hoyt, J.; Young, R. The laboratory rabbit, guinea pig, hamster, and other rodents. In The rabbit as an experimental model; Suckow, M., Stevens, K., Wilson, R., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 529–560. [Google Scholar]
- Esteves, P.; Abrantes, J.; Baldauf, H.; BenMohamed, L.; Chen, Y.; Christensen, N.; González-Gallego, J.; Giacani, L.; Hu, J.; Kaplan, G.; Keppler, O.; Knight, K.; Kong, X.; Lanning, D.; Pendu, J.; Lemos, A.; Liu, J.; Liu, S.; Lopes, A.; Lu, S.; Lukehart, S.; Manabe, Y.; Neves, F.; McFadden, G.; Mage, R. The wide utility of rabbits as models of human diseases. Exp. Mol. Med. 2018, 50, 1–10. [Google Scholar] [CrossRef]
- Soares, J.; Pinheiro, A.; Esteves, P. The rabbit as an animal model to study innate immunity genes: Is it better than mice? Front. Immunol. 2022, 13, 981815. [Google Scholar] [CrossRef]
- Valentine, H.; Daugherity, E.; Singh, B.; Maurer, K. The experimental use of Syrian hamsters. In American College of Laboratory Animal Medicine. The laboratory rabbit, guinea pig, hamster, and other rodents; Suckow, M., Stevens, K., Wilson, R., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 875–906. [Google Scholar] [CrossRef]
- Miedel, E.; Hankenson, F. Biology and diseases of hámsters. In American College of Laboratory Animal Medicine. Laboratory Animal Medicine, 3rd ed.; Fox, J., Anderson, L., Otto, G., Pritchett-Corning, K., Whary, M., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 209–245. [Google Scholar] [CrossRef]
- Shomer, N.; Holcombe, H.; Harkness, J. Biology and diseases of guinea pigs. In American College of Laboratory Animal Medicine. Laboratory Animal Medicine, 3rd ed.; Fox, J., Anderson, L., Otto, G., Pritchett-Corning, K., Whary, M., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 247–283. [Google Scholar] [CrossRef]
- Lagunes-Quintanilla, R.; Valdez-Espinoza, U.; Hernández-Ortiz, R.; Castro-Saines, E.; Merino, O.; Mendoza-Martínez, N. Experimental vaccination in rabbits using the peptide RmS-17 antigen reduces the performance of a Mexican Rhipicephalus microplus tick strain. Ticks Tick Borne Dis. 2022, 13, 102044. [Google Scholar] [CrossRef] [PubMed]
- Trimnell, A.; Davies, G.; Lissina, O.; Hails, R.; Nuttall, P. A cross-reactive tick cement antigen is a candidate broad-spectrum tick vaccine. Vaccine 2005, 23, 4329–4341. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Narasimhan, S.; Vidyarthi, A.; Sameet, C.; Meister, L.; Diktas, H.; Strank, N.; Lynn, G.; DePonte, K.; Craft, J.; Fikrig, E. Repeat tick exposure elicits distinct immune responses in guinea pigs and mice. Ticks Tick Borne Dis. 2020, 11, 101529. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Santos-Burgoa, C. Treatment against human rabies: a bit of its history. Public Health. 1994, 28, 454–63. [Google Scholar]
- Russell, R.; Schilling, P. Selected topics on laboratory medicine: The rabbit. In Series of scientific and technical monographs. WHO 1976, 4, 12–86. [Google Scholar]
- Kirkland, W. Ultrastructural changes in the nymphal salivary glands of the rabbit tick, Haemaphysalis leporispalustris, during feeding. J. Insect Physio. 1971, 17, 1933–1946. [Google Scholar] [CrossRef]
- McGowan, M.; Homer, T.; Odell, G.; McNew, R.; Barker, R. Performance of ticks fed on rabbits inoculated with extracts derived from homogenized tick Amblyomma maculatum Koch (Acari: Ixodidae). J. Parasitol. 1980, 66, 42–48. [Google Scholar] [CrossRef]
- Walker, A.; Fletcher, J. Histological study of the attachment sites of adult Rhipicephalus appendiculatus on rabbits and cattle. Int. J. Parasitol. 1986, 16, 399–413. [Google Scholar] [CrossRef]
- Fox, R. The biology of the laboratory rabbit. In Taxonomy and genetics; Weisbroth, S., Kraus, A., Eds.; Academic Press: New York, 1974; p. 22. [Google Scholar]
- ARBA. American Rabbit Breeders Association. Available online: https://www.arba.net/breeds.htm (accessed on 30 October 2018).
- Rodríguez-Mallon, A.; Fernández, E.; Encinosa, P.; Bello, Y.; Méndez-Pérez, L.; Cepero, L.; Pérez, D.; González, M.; Garay, H.; Reyes, O.; Méndez, L.; Estrada, M. A novel tick antigen shows high vaccine efficacy against the dog tick, Rhipicephalus sanguineus. Vaccine 2012, 30, 1782–1789. [Google Scholar] [CrossRef]
- Graur, D.; Duret, L.; Gouy, M. Phylogenetic position of the order Lagomorpha (rabbits, hares and allies). Nature 1996, 379, 333–335. [Google Scholar] [CrossRef]
- Bryda, E. The mighty mouse: the impact of rodents on advances in biomedical research. Mo Med. 2013, 110, 207–211. [Google Scholar]
- Mullane, K.; Williams, M. Animal models of asthma: reprise or reboot? Biochem Pharmacol. 2014, 87, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Trager, W. Acquired immunity to ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Suckow, M.; Stevens, K.; Wilson, R. The laboratory rabbit, guinea pig, hamster, and other rodents; Suckow, M., Stevens, K., Wilson, R., Eds.; A volume in American College of Laboratory Animal Medicine, 2012; pp. 572–689. [Google Scholar] [CrossRef]
- Mulenga, A.; Sugimoto, Y.; Sako, K.; Musoke, A.; Mozaria, S.; Onuma, M. Molecular characterisation of a Haemaphysalis longicornis tick salivary gland-associated 29-kilodalton protein and its effect as a vaccine against tick infestation in rabbits. Infect. Immun. 1999, 40, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Sugino, M.; Imamura, S.; Mulenga, A.; Nakajima, M.; Tsuda, A.; Ohashi, K.; Onuma, M. A serine proteinase inhibitor (serpin) from ixodid tick Haemaphysalis longicornis; cloning and preliminary assessment of its suitability as a candidate for a tick vaccine. Vaccine 2003, 21, 2844–2851. [Google Scholar] [CrossRef]
- Galay, R.; Umemiya-Shirafuji, R.; Bacolod, E.; Maeda, H.; Kusakisako, K.; Koyama, J. Two kinds of ferritin protect ixodid ticks from iron overload and consequent oxidative stress. PLoS ONE 2014, 9, e90661. [Google Scholar] [CrossRef]
- Liang, N.; Hong-Meng, D.; Xiang-Yuan, F.; Ya-Xue, W.; Feng, Y.; Xiao-Ya, L.; Yong-Hong, H. Characterization and evaluation of a new triosephosphate isomerase homologue from Haemaphysalis longicornis as a candidate vaccine against tick infection. Ticks Tick Borne Dis. 2022, 13, 101–968. [Google Scholar] [CrossRef]
- Egizi, A.; Bulaga-Seraphin, L.; Alt, E.; Bajwa, W.; Bernick, J.; Bickerton, M.; Fonseca, D. First glimpse into the origin and spread of the Asian longhorned tick Haemaphysalis longicornis, in the United States. Zoonoses Public Health 2020, 67, 637–650. [Google Scholar] [CrossRef]
- Zhao, L.; Li, J.; Cui, X.; Jia, N.; Wei, J.; Xia, L.; Wang, H.; Zhou, Y.; Wang, Q.; Liu, X.; Yin, C.; Pan, Y.; Wen, H.; Wang, Q.; Fuzhong, X.; Sun, Y.; Jiang, J.; Li, S.; Cao, W. Distribution of Haemaphysalis longicornis and associated pathogens: analysis of pooled data from a China field survey and global published data. Lancet Planet. Health 2020, 4, e320–e329. [Google Scholar] [CrossRef]
- Luo, L.; Zhao, L.; Wen, H.; Zhang, Z.; Liu, J.; Fang, L.; Yu, X. Haemaphysalis longicornis ticks as reservoir and vector of severe fever with thrombocytopenia syndrome virus in China. J. Emerg. Infect. Dis. 2015, 21, 1770. [Google Scholar] [CrossRef]
- Tufts, D.; Sameroff, S.; Tagliafierro, T.; Jain, K.; Oleynik, A.; VanAcker, M.; Tokarz, R. A metagenomic examination of the pathobiome of the invasive tick species, Haemaphysalis longicornis, collected from a New York City borough, USA. Ticks Tick Borne Dis. 2020, 11, 101516. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, A.; Mulenga, A.; Sugimoto, C.; Nakajima, M.; Ohashi, K.; Onuma, M. cDNA cloning, characterization and vaccine effect analysis of Haemaphysalis longicornis tick saliva proteins. Vaccine 2001, 19, 4287–4296. [Google Scholar] [CrossRef] [PubMed]
- Pospelova-Shtrom, M. On the system of classification of ticks of the family Argasidae CAN. Acarologia 1969, 11, 1–22. [Google Scholar] [PubMed]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R. African swine fever: how can global spread be prevented? Philos Trans R Soc Lond B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef]
- ECDC. European Centre for Disease Prevention and Control and European Food Safety Authority. Available online: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data (accessed on 12 June 2022).
- Baizheng, W.; Xin, L.; Jingze, L.; Rong, B. Predicting the potential habitat for Ornithodoros tick species in China. Vet. Parasitol. 2022, 311, 109793. [Google Scholar] [CrossRef]
- De Morais, J.; Lopes, I.; Nuncio, M. Spanish-African recurrent fever in Portugal: historical and clinical-epidemic escorco. Int. Med. 2007, 14, 170–178. [Google Scholar]
- Assous, M.; Wilamowski, A. Relapsing fever borreliosis in Eurasia-forgotten, but certainly not gone! Clin. Microbiol. Infect. 2009, 15, 407–414. [Google Scholar] [CrossRef]
- Oleaga, A.; González-Pérez, S.; Pérez-Sánchez, R. First molecular and functional characterisation of ferritin 2 proteins from Ornithodoros argasid ticks. Vet. Parasitol. 2022, 304, 109–684. [Google Scholar] [CrossRef]
- Manzano-Román, R.; Díaz-Martín, V.; Oleaga, A.; Siles-Lucas, M.; Pérez-Sánchez, R. Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet. Parasitol. 2012, 185, 248–259. [Google Scholar] [CrossRef]
- Manzano-Román, R.; Díaz-Martín, A.; Pérez-Sánchez, R. Identification of protective linear B-cell epitopes on the subolesin/akirin orthologues of Ornithodoros spp. soft ticks. Vaccine 2015, 33, 1046–1055. [Google Scholar] [CrossRef]
- Guglielmone, A.; Petney, T.; Robbins, R. Ixodidae (Acari: Ixodoidea): descriptions and redescriptions of all known species from 1758 to December 31, 2019. Zootaxa 2020, 4871, 1–322. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.; Keirans, J.; Horak, I. The genus Rhipicephalus (Acari, Ixodidae): a guide to the brown ticks of the world, 1st ed.; Cambridge University Press: Cambridge, 2000; pp. 79–104. [Google Scholar]
- Makwarela, T.; Nyangiwe, N.; Masebe, T.; Mbizeni, S.; Nesengani, L.; Djikeng, A.; Mapholi, N. Tick diversity and distribution of hard (Ixodidae) cattle ticks in South Africa. Microbiol. Res. 2023, 14, 42–59. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasit. Vectors 2010, 3, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Brophy, M.; Riehle, M.; Mastrud, N.; Ravenscraft, A.; Adamson, J.; Walker, K. Genetic variation in Rhipicephalus sanguineus s.l. ticks across Arizona. Int. J. Environ. Res. Public Health 2022, 19, 4223. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Konnai, S.; Imamura, S.; Ito, T.; Onuma, M.; Ohashi, K. Cloning and characterization of Rhipicephalus appendiculatus voraxinα and its effect as anti-tick vaccine. Vaccine 2009, 27, 5989–5997. [Google Scholar] [CrossRef]
- Leal, B.; Alzugaray, M.; Seixas, A.; da Silva Vaz, I.; Ferreira, C. Characterization of a glycine-rich protein from Rhipicephalus microplus: Tissue expression, gene silencing and immune recognition. Parasitolgy 2018, 145, 927–938. [Google Scholar] [CrossRef]
- Jittapalapong, S.; Kaewhom, P.; Pumhom, P.; Canales, M.; de la Fuente, J.; Stich, R. Immunization of rabbits with recombinant serine protease inhibitor reduces the performance of adult female Rhipicephalus microplus. Transbound. Emerg. Dis. 2010, 57, 103–106. [Google Scholar] [CrossRef]
- Zhou, J.; Gong, H.; Zhou, Y.; Xuan, X.; Fujisaki, K. Identification of a glycine-rich protein from the tick Rhipicephalus haemaphysaloides and evaluation of its vaccine potential against tick feeding. Parasitol. Res. 2006, 100, 77–84. [Google Scholar] [CrossRef]
- Gray, J. The ecology of ticks transmitting Lyme borreliosis. Exp. Appl. Acarol. 1998, 22, 249–258. [Google Scholar] [CrossRef]
- Gilbert, L.; Maffey, G.; Ramsay, S.; Hester, A. The effect of deer management on the abundance of Ixodes ricinus in Scotland. Ecol Appl 2012, 22, 658–667. [Google Scholar] [CrossRef]
- Hofmeester, T.; Sprong, H.; Jansen, P.; Prins, H.; Van, S. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in dutch forests. Parasit. Vectors 2017, 10, 1–8. [Google Scholar] [CrossRef]
- Ostfeld, R.; Canham, C.; Oggenfuss, K.; Winchcombe, R.; Keesing, F. Climate, deer, rodents, and acorns as determinants of variation in lyme-disease risk. PLoS ONE 2006, 4, e145. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.; Palli, S. Mapping distributions of the Lyme disease vector, Ixodes scapularis, and spirochete, Borrelia burgdorferi, in Kentucky using passive and active surveillance. Ticks Tick Borne Dis. 2022, 13, 101885. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.; Narasimhan, S.; Daffre, S.; de Ponte, K.; Hovius, J.; Veer, V. Identification and characterization of Ixodes scapularis antigens that elicit immunity to ticks by visualizing the yeast surface. PLoS ONE 2011, 6, e15926. [Google Scholar] [CrossRef] [PubMed]
- Contreras, M.; de la Fuente, J. Control of infestations by Ixodes ricinus tick larvae in rabbits vaccinated with aquaporin recombinant antigens. Vaccine 2017, 35, 1323–1328. [Google Scholar] [CrossRef]
- Hajdusek, O.; Almazán, C.; Loosova, G.; Villar, M.; Canales, M.; Grubhoffer, L.; Kopacek, P.; de la Fuente, J. Characterization of ferritin 2 for the control of tick infestations. Vaccine 2010, 28, 2993–2998. [Google Scholar] [CrossRef]
- Yunker, C.; Keirans, J.; Cliffornd, C.; Easton, E. Dermacentor ticks (Acari: Ixodoidea: Ixodidae) of the new world: a scanning electron microscope atlas. Proc. Entomol. Soc. Wash. 1986, 88, 609–627. [Google Scholar]
- Eisen, R.; Kugeler, K.; Eisen, J.; Beard, C.; Paddock, C. Tick-borne zoonoses in the United States: persistent and emerging threats to human health. ILAR J 2017, 58, 319–335. [Google Scholar] [CrossRef]
- Dergousoff, S.; Galloway, T.; Lindsay, L.; Curry, P.; Chilton, N. Range expansion of Dermacentor variabilis and Dermacentor andersoni (Acari: Ixodidae) near their northern distributional limits. J. Med. Entomol. 2013, 50, 510–520. [Google Scholar] [CrossRef]
- Zhang, G.; Zheng, D.; Tian, Y.; Li, S. A dataset of distribution and diversity of ticks in China. Sci. Data 2019, 6, 105. [Google Scholar] [CrossRef]
- Martin, J.; Fischhoff, I.; Castellanos, A.; Han, B. Ecological predictors of zoonotic vector status among Dermacentor ticks (Acari: Ixodidae): A trait-based approach. J. Med. Entomol. 2022, 59, 2158–2166. [Google Scholar] [CrossRef] [PubMed]
- Imamura, S.; da Silva Vaz, I.; Sugino, M.; Ohashi, K.; Onuma, M. A serine protease inhibitor (Serpin) from Haemaphysalis longicornis as an anti-tick vaccine. Vaccine 2005, 23, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Chinzei, Y.; Minoura, H. Reduced oviposition in Ornithodoros moubata (Acari: Argasidae) fed on tick-sensitized and vitellin-immunized rabbits. J. Med. Entomol. 1988, 25, 26–31. [Google Scholar] [CrossRef]
- Weiss, B.; Kaufman, W. Two feeding-induced proteins from the male gonad trigger engorgement of the female tick, Amblyomma hebraeum. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 5874–5879. [Google Scholar] [CrossRef] [PubMed]
- Shahein, Y. Molecular cloning and expression of a larval immunogenic protein from the cattle tick Boophilus annulatus. Vet. Immunol. Immunopathol. 2008, 121, 281–289. [Google Scholar] [CrossRef]
- Canales, C.; Labruna, M.; Soares, J.; Prudencio, C.; de la Fuente, J. Protective efficacy of bacterial membranes containing surface-exposed BM95 antigenic peptides for the control of cattle tick infestations. Vaccine 2009, 27, 7244–7248. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Martín, V.; Manzano-Román, R.; Valero, L.; Oleaga, A.; Encinas-Grandes, A.; Pérez-Sánchez, R. An insight into the proteome of the saliva of the argasid tick Ornithodoros moubata reveals important differences in saliva protein composition between the sexes. J Proteomics 2013, 80, 216–235. [Google Scholar] [CrossRef]
- Manzano-Román, R.; Díaz-Martín, V.; Oleaga, V.; Obolo-Mvoulouga, P.; Pérez-Sánchez, R. TSGP4 from Ornithodoros moubata: molecular cloning, phylogenetic analysis and vaccine efficacy of a new member of the lipocalin clade of cysteinyl leukotriene scavengers. Vet. Parasitol. 2016, 227, 130–137. [Google Scholar] [CrossRef]
- Kim, T.; Radulovic, Z.; Mulenga, A. Target validation of highly conserved Amblyomma americanum tick saliva serine protease inhibitor 19. Ticks Tick Borne Dis. 2016, 7, 405–414. [Google Scholar] [CrossRef]
- Rangel, A.; Pereira, F.; Casquero, R.; Valério, M.; Andreotti, R. Design of the ATAQ peptide and its evaluation as an immunogen to develop a Rhipicephalus vaccine. Vet. Parasitol. 2016, 221, 30–38. [Google Scholar] [CrossRef]
- Alzugaray, M.; Parizi, L.; Seixas, A.; Benavides, U.; da Silva Vaz, I. Molecular and functional characterization of Bm05br antigen from Rhipicephalus microplus. Ticks Tick Borne Dis. 2017, 8, 320–329. [Google Scholar] [CrossRef]
- Sabadin, G.; Parizi, F.; Kiio, I.; Amaral, M.; da Silva, R.; Camargo-Mathias, M.; Wangombe, N.; Nene, V.; da Silva Vaz, I. Effect of recombinant glutathione S-transferase as vaccine antigen against Rhipicephalus appendiculatus and Rhipicephalus sanguineus infestation. Vaccine 2017, 35, 6649–6656. [Google Scholar] [CrossRef]
- Obolo-Mvoulouga, P.; Oleaga, A.; Manzano-Román, R.; Pérez-Sánchez, R. Evaluation of the protective efficacy of Ornithodoros moubata midgut membrane antigens selected using omics and in silico prediction algorithms. Ticks Tick Borne Dis 2018, 9, 1158–1172. [Google Scholar] [CrossRef]
- Asadollahi, Z.; Nabian, S.; Taheri, M.; Ebrahimzadeh, E. Introducing a new anti-Rhipicephalus (Boophilus) microplus tick recombinant vaccine candidate using cathepsin and tropomyosin multi-epitope gene. Vet Res Forum 2021, 12, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Ge, T.; Hu, E.; Fan, X.; Zhang, Y.; Zhai, X.; Li, M.; Zhang, W.; Wu, L.; Ka, A.; Cheung, L.; Chahan, B. Recombinant cysteine proteinase as anti-tick targeting Hyalomma asiaticum infestation. Exp. Parasitol. 2022, 235, 108234. [Google Scholar] [CrossRef]
- Adjou, P.; Naomasa, S.; Tuvshintulga, B.; Sato, N.; Okado, K.; Zheng, W.; Lee, S.; Mosqueda, J.; Suzuki, H.; Xuan, X.; Umemiya-Shirafuji, R. Identification and characterization of Rhipicephalus microplus ATAQ homolog from Haemaphysalis longicornis ticks and its immunogenic potential as an anti-tick vaccine candidate molecule. Microorganisms 2023, 11, 822. [Google Scholar] [CrossRef]
- Duo, D.; Ru, L.; Ya-Xue, W.; Xiang-Yuan, F.; Xiao-Ya, L.; Feng, Y.; Tian-Tian, Z.; Jing-Yi, M. Molecular characterization of hexokinase (HK) in Haemaphysalis longicornis and evaluation of HK protein- and DNA-based vaccines against adult ticks. Pest Manag. Sci. 2023, 79, 1721–1730. [Google Scholar] [CrossRef]
- Carnero-Morán, A.; Oleaga, A.; Cano-Argüelles, A.; Pérez-Sánchez, R. Function-guided selection of salivary antigens from Ornithodoros erraticus argasid ticks and assessment of their protective efficacy in rabbits. Ticks Tick Borne Dis. 2023, 14, 102218. [Google Scholar] [CrossRef]
- Kemp, D.; Pearson, R.; Gough, J.; Willadsen, P. Vaccination against Boophilus microplus: localization of antigens on the tick gut cells and their interaction with the host immune system. Exp. Appl. Acarol. 1989, 7, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Imamura, S.; Namangala, B.; Tajima, T.; Tembo, M.; Yasuda, J.; Ohashi, K.; Onuma, M. Two serine protease inhibitors (serpins) that induce a bovine protective immune response against Rhipicephalus appendiculatus ticks. Vaccine 2006, 24, 2230–2237. [Google Scholar] [CrossRef]
- Vargas-Hernandez, V.; Montero-Espinosa, C.; Sánchez-Villaurrutia, D.; Duarte, C.; Henrique, G.; Fuentes-Castillo, A.; Ancisar, J.; Suárez-Alba, J.; Mosqueda-Lobaina, O.; Suárez-Pedroso, M. Infestation of rabbits with just-molted adults of the cattle tick Rhipicephalus microplus: biological parameters and efficiency. Rev. Bras. DE Parasitol. Vet. 2023, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Daemon, E.; Prata, M.; Faccini, J. Goats as alternative hosts of Boophilus microplus (Acari: ixodidae). Rev. Bras. DE Parasitol. Vet. 1998, 7, 123–128. [Google Scholar]
- Franque, M.; Santos, H.; Silva, G.; Tajiri, J.; Massard, C. Biological characteristics of Boophilus microplus (Acari: Ixodidae) on dog under experimental infestation. Rev. Bras. DE Parasitol. Vet. 2007, 16, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Franque, M.; Santos, H.; Linarez, F.; Massard, C. Experimental infestation of horses by Rhipicephalus (Boophilus) microplus. Cienc Rural 2009, 39, 2117–2122. [Google Scholar] [CrossRef]
- Guimarães, C.; Wilwerth, D.; Daemon, E.; Faccini, J. Effect of the number of Boophilus microplus (Can., 1887) (Acari: lxodidae) larvae on the recovery of engorged females in rabbits. Rev. Bras. DE Parasitol. Vet. 1999, 3, 59–61. [Google Scholar] [CrossRef]
- Hitcheock, L. Studies on the parasitic stages of the cattle tick Boophilus microplus (Canestrini) (Acarina: Ixodidae). Aust. J. Zool. 1955a, 3, 145–155. [Google Scholar] [CrossRef]
- Nuttall, P.; Labuda, M. Tick-host interactions: saliva-activated transmission. Parasitology 2004, 129, S177–S189. [Google Scholar] [CrossRef]
- Silva, C.; Cunha, D.; Daemon, E.; Faccini, J. Effect of the number of Boophilus microplus (Can., 1887) larvae (Acari: lxodidae) on the recovery of engorged females in rabbits. Rev. Bras. DE Parasitol. Vet. 1996, 3, 59–61. [Google Scholar] [CrossRef]
- Zacarias, M.; Azevedo, M.; Daemon, E.; Furlong, J. Biological parameters of cattle ticks fed on rabbits. Rev. Bras. DE Parasitol. Vet. 2012, 21, 22–27. [Google Scholar]
- Francischetti, I. The role of saliva in tick feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef]
- Mudenda, L.; Pierlé, S.; Turse, J.; Scoles, G.; Purvine, S.; Nicora, C.; Clauss, T.; Ueti, M.; Brown, W.; Brayton, K. Proteomics informed by transcriptomics identifies novel secreted proteins in Dermacentor andersoni saliva. Int. J. Parasitol. 2014, 44, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Tirloni, L.; Reck, J.; Terra, R.; Martins, J.; Mulenga, A.; Sherman, N.; Fox, J.; Yates, J.; Termignoni, C.; Pinto, A.; da Silva Vaz, I. Proteomic analysis of cattle tick Rhipicephalus (Boophilus) microplus saliva: A comparison between partially and fully engorged females. PLoS ONE 2014, 9, e94831. [Google Scholar] [CrossRef]
- Tirloni, L.; Islam, M.; Kim, T.; Diedrich, J.; Yates, J.; Pinto, A.; Mulenga, A.; You, M.; da Silva, I. Saliva from nymph and adult females of Haemaphysalis longicornis: a proteomic study. Parasit. Vectors 2015, 8, 338. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Tirloni, L.; Pinto, A.; Moresco, J.; Yates, J.; da Silva Vaz, I.; Mulenga, A. Ixodes scapularis tick saliva proteins sequentially secreted every 24 h during blood feeding. PLoS. Negl. Trop. Dis. 2016b, 10, e0004323. [Google Scholar] [CrossRef]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S. The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef] [PubMed]
- Leboulle, G.; Rochez, C.; Louahed, J.; Ruti, B.; Brossard, M. Isolation of Ixodes ricinus salivary gland mRNA encoding factors induced during blood feeding. Am. J. Trop. Med. Hyg. 2022, 66, 225–233. [Google Scholar] [CrossRef]
- Francischetti, I.; Sa-Nunes, A.; Mans, B.; Santos, I.; Ribeiro, J. The role of saliva in tick feeding. Front. Biosci. 2009, 14, 2051–2088. [Google Scholar] [CrossRef]
- Mans, B. Evolution of vertebrate hemostatic and inflammatory control mechanisms in blood-feeding arthropods. J Innate Immun. 2011, 3, 41–51. [Google Scholar] [CrossRef]
- Ribeiro, J.; Francischetti, I. Role of arthropod saliva in blood feeding: sialome and post-sialome perspectives. Annu. Rev. Entomol. 2003, 48, 73–88. [Google Scholar] [CrossRef]
- Wikel, S. Host immunity to ticks. Annu. Rev. Entomol. 1996, 41, 1–22. [Google Scholar] [CrossRef]
- Tirloni, L.; Kim, T.; Pinto, A.; Yates, J.; da Silva Vaz, I.; Mulenga, A. Tick-Host range adaptation: Changes in protein profiles in unfed adult Ixodes scapularis and Amblyomma americanum saliva stimulated to feed on different hosts. Front. Cell. Infect. Microbiol. 2017, 7, 517. [Google Scholar] [CrossRef] [PubMed]
- Mulenga, A.; Blandon, M.; Khumthong, R. The molecular basis of the Amblyomma americanum tick attachment phase. Exp. Appl. Acarol. 2007, 41, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Lew-Tabor, A.; Moolhuijzen, P.; Vance, M.; Kurscheid, S.; Valle, M.; Jarrett, S.; Minchin, C.; Jackson, L.; Jonsson, N.; Bellgard, M.; Guerrero, F. Suppressive subtractive hybridization analysis of Rhipicephalus (Boophilus) microplus larval and adult transcript expression during attachment and feeding. Vet. Parasitol. 2010, 167, 304–320. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Tirloni, L.; Pinto, A.; Diedrich, J.; Moresco, J.; Yates, J.; da Silva Vaz, I.; Mulenga, A. Time resolved proteomic profile of Amblyomma americanum tick saliva during feeding. PLoS. Negl. Trop. Dis. 2020, 14, e0007758. [Google Scholar] [CrossRef]
- Schwarz, A.; von Reumont, B.; Erhart, J.; Chagas, A.; Ribeiro, J.; Kotsyfakis, M. De novo Ixodes ricinus salivary gland transcriptome analysis using two nextgeneration sequencing methodologies. FASEB J. 2013, 27, 4745–4756. [Google Scholar] [CrossRef]
- Imamura, S.; da Silva Vaz, I.; Konnai, S.; Yamada, Y.; Nakajima, C.; Onuma, M.; Ohashi, K. Effect of vaccination with a recombinant metalloprotease from Haemaphysalis longicornis. Exp. Appl. Acarol. 2009, 48, 345–358. [Google Scholar] [CrossRef]
Figure 1.
Tick-derived proteins evaluated in tick vaccination trials using rabbits as an animal model.
Figure 1.
Tick-derived proteins evaluated in tick vaccination trials using rabbits as an animal model.
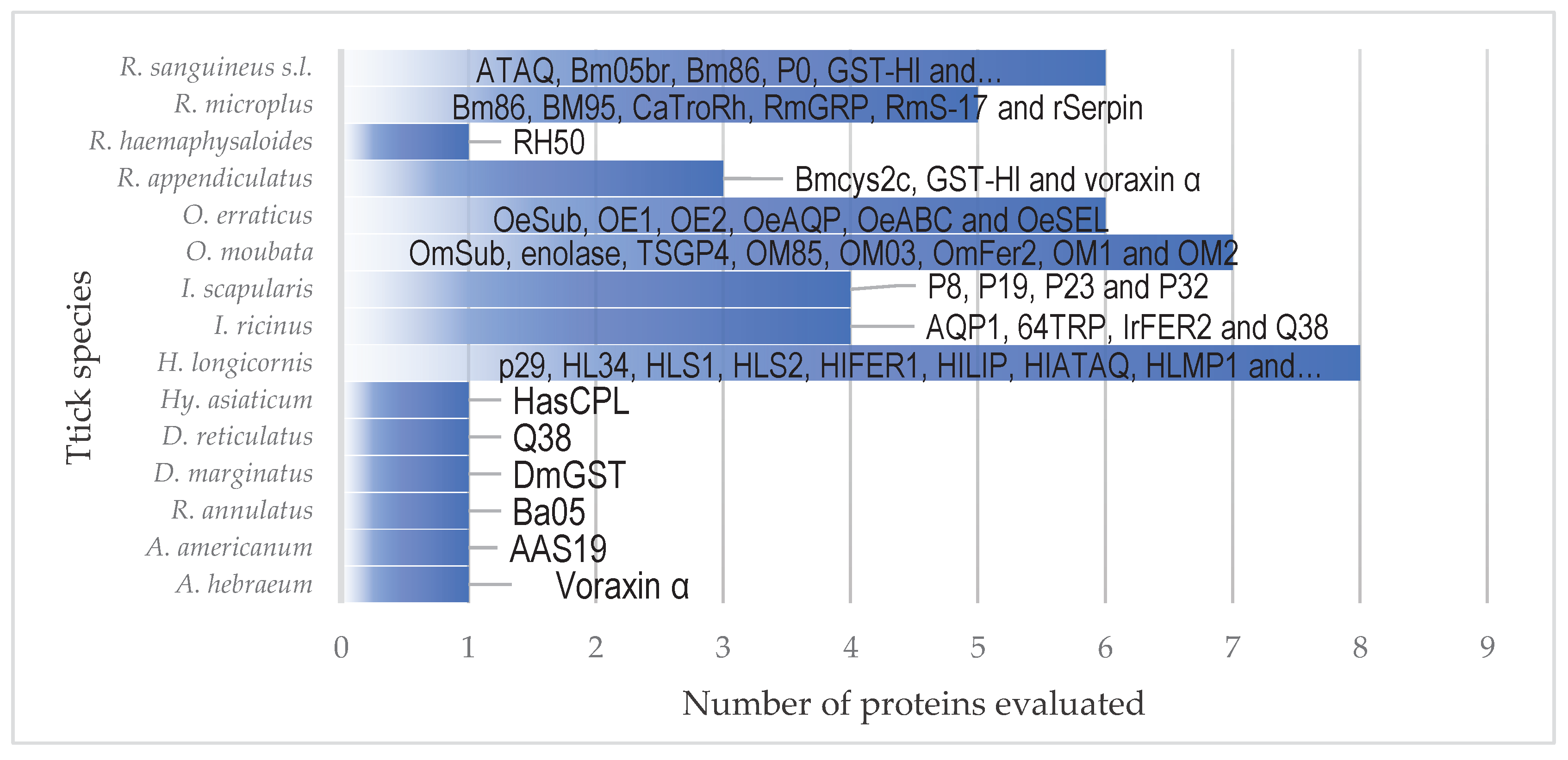
Figure 2.
Different tick species and how often they are evaluated in studies using rabbits as models for vaccination experiments.
Figure 2.
Different tick species and how often they are evaluated in studies using rabbits as models for vaccination experiments.
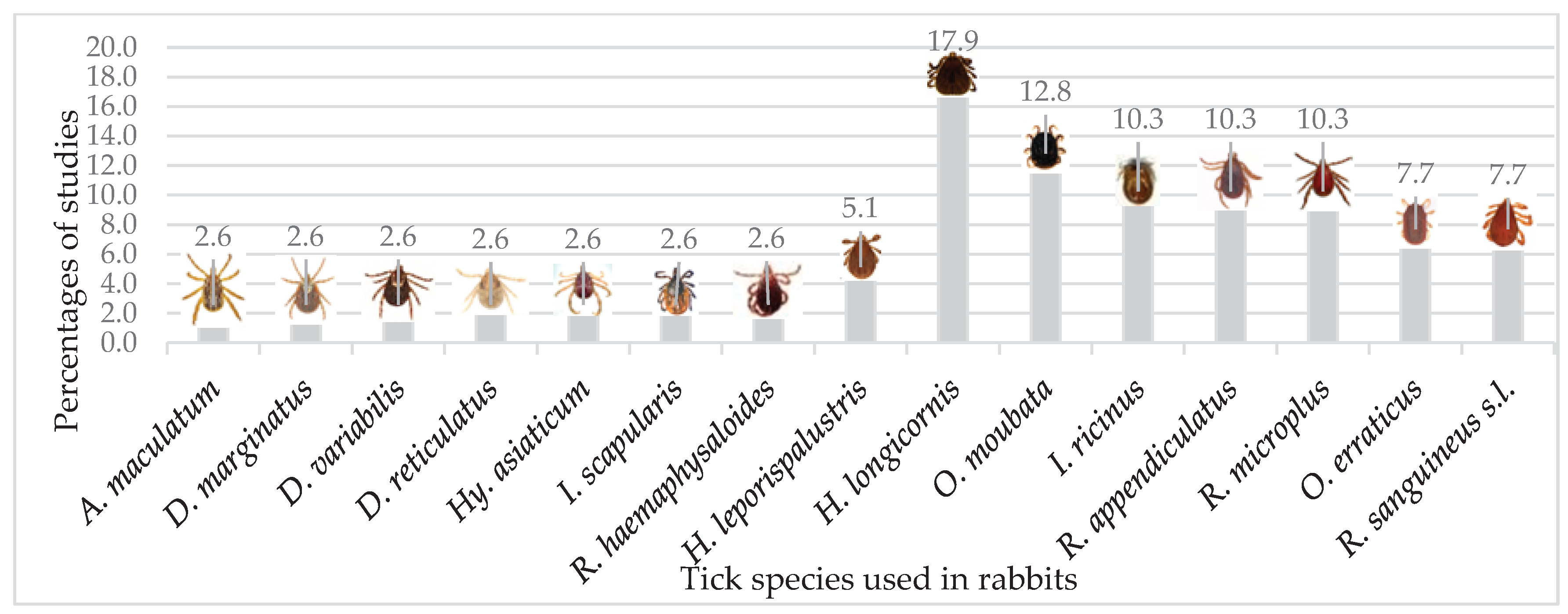
Figure 3.
Vaccine efficiency (percent reduction of tick infestation) against different ticks, according to the breed of rabbits used for immunization. NZ: New Zealand. JW: Japanese white. C: Californians. FL: Flemish lop-eared rabbit.
Figure 3.
Vaccine efficiency (percent reduction of tick infestation) against different ticks, according to the breed of rabbits used for immunization. NZ: New Zealand. JW: Japanese white. C: Californians. FL: Flemish lop-eared rabbit.
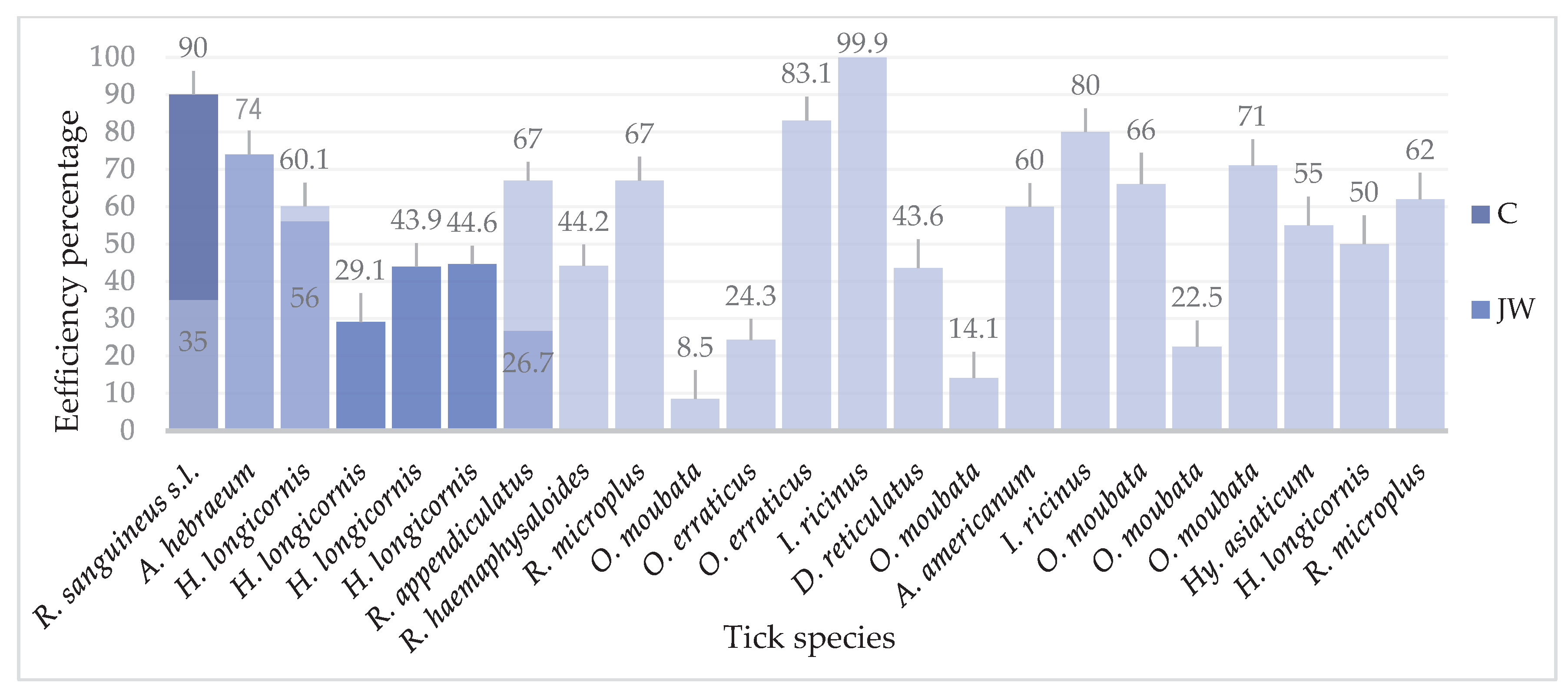
Figure 4.
Geographical distribution of studies using rabbits as animal models to test anti-tick vaccines. (Parts of the figures were drawn by using pictures from Servier Medical Art: http://smart.servier.com/.
Figure 4.
Geographical distribution of studies using rabbits as animal models to test anti-tick vaccines. (Parts of the figures were drawn by using pictures from Servier Medical Art: http://smart.servier.com/.
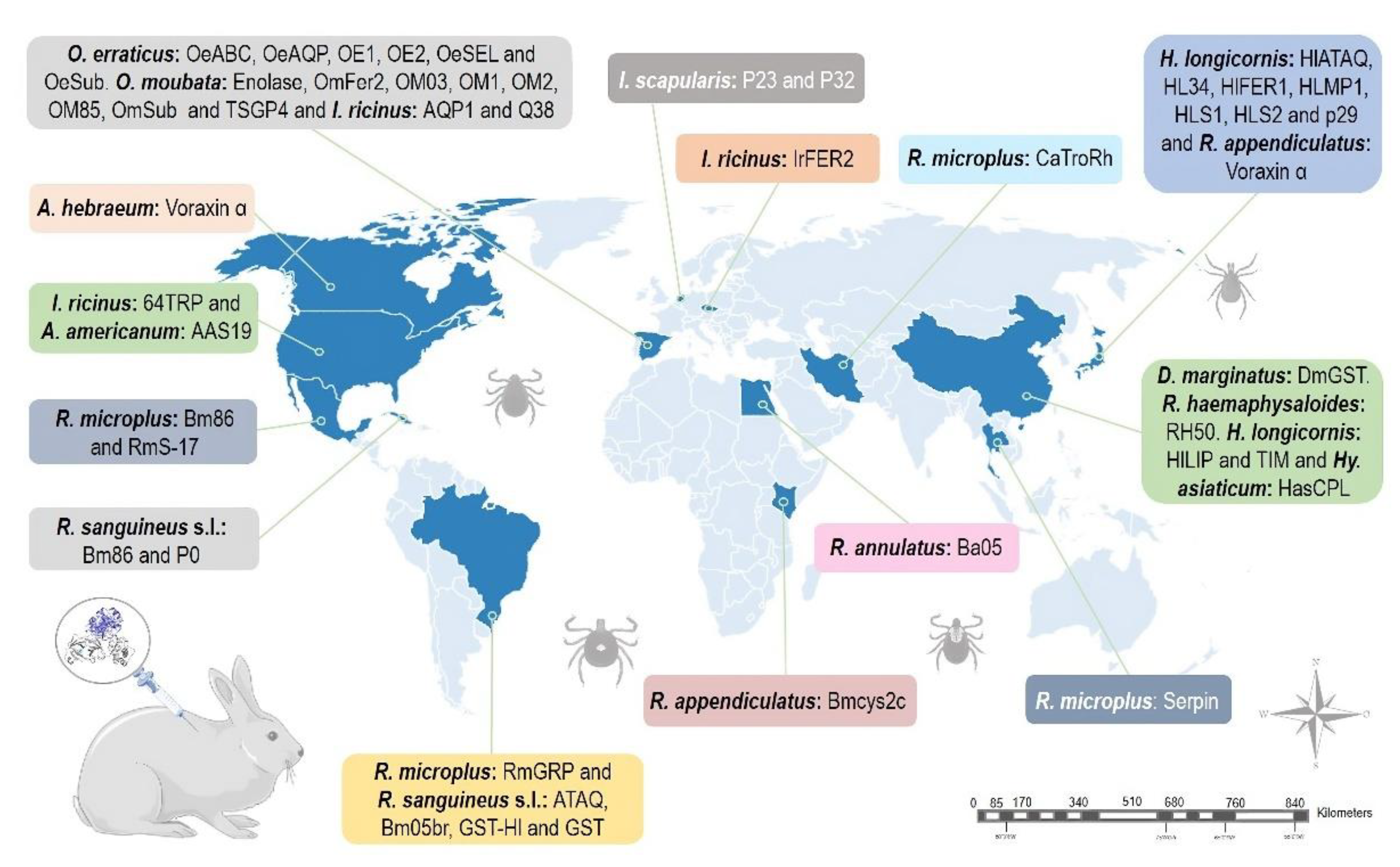
Figure 5.
Comparison of different models of tick infestation in rabbits. 1. Larval stage tick infestation, 2. Nymphal stage tick infestation, 3. Adult stage tick infestation, and 4. Nymphal and adult stage tick infestation (Parts of the figures were drawn by using pictures from Servier Medical Art: http://smart.servier.com/.
Figure 5.
Comparison of different models of tick infestation in rabbits. 1. Larval stage tick infestation, 2. Nymphal stage tick infestation, 3. Adult stage tick infestation, and 4. Nymphal and adult stage tick infestation (Parts of the figures were drawn by using pictures from Servier Medical Art: http://smart.servier.com/.
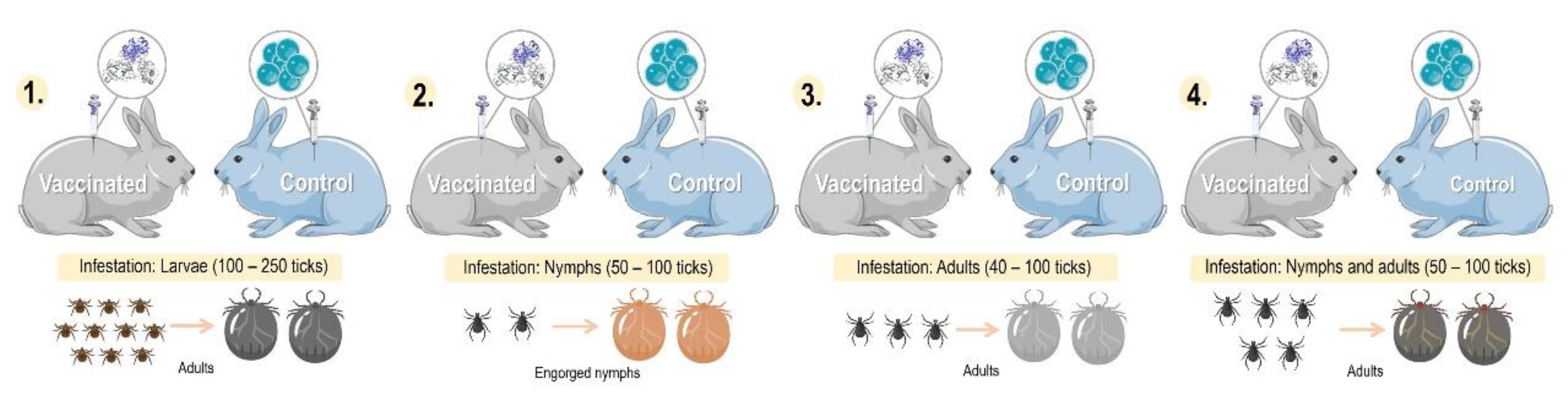

Table 1.
Use of rabbits as animal models for anti-tick vaccine experiment.
| No | Experiment/Molecule Name | # of Rabbits | Rabbitsbreed | Tick Species | Tick Stages | Immunization | Tick per Rabbit | % Reduction | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Evaluation of the immune response | 6 | NR | D. variabilis and H. leporispalustris | Larvae | 1 | 159 | NA | [61] |
| 2 | Evaluation of the immune response | NR | NZ | H. leporispalustris | Nymphs | 1 | NR | NA | [52] |
| 3 | Whole tick tissues collected from Amblyomma maculatum | 8 | NZ | A. maculatum | Nymphs and adults | 2 | 75 | NA | [53] |
| 4 | Attachment sites of Rhipicephalus appendiculatus | 5 | NZ | R. appendiculatus | Adults | 3 | 80 | NA | [54] |
| 5 | Tick egg yolk protein (vitellin) | 4 | JW | O. moubata | Nymphs and adults | 4 | 112 | NA | [104] |
| 6 | Native protein (p29) | 10 | JW | H. longicornis | Nymphs and adults | 3 | 2110 | 56 | [63] |
| 7 | Haemaphysalis longicornis protein 34 (HL34) | 4 | JW | H. longicornis | Nymphs and adults | 2 | 115 | 29,1 | [71] |
| 8 | Haemaphysalis longicornis serpin 1 (HLS1) | 4 | JW | H. longicornis | Nymphs and adults | 2 | 120 | 43,9 | [64] |
| 9 | Voraxin of Amblyomma hebraeum | 2 | FL | A. hebraeum | Adults | 3 | 62 | 74 | [105] |
| 10 | 64TRP | 14 | NR | I. ricinus | Adults | 3 | 30 | NA | [48] |
| 11 | Haemaphysalis longicornis serpin-2 (HLS2) | 4 | JW | H. longicornis | Nymphs and adults | 2 | 160 | 44,6 | [103] |
| 12 | RH50 | 6 | NZ | R. haemaphysaloides | Nymphs and adults | 3 | 120 | 74,7 | [89] |
| 13 | Protein 05 from Boophilus annulatus (Ba05) | 1 | NR | B. annulatus | Larvae | NA | NA | NA | [106] |
| 14 | Recombinant BM95-MSP1a fusion protein and Bm86 | 16 | NZ | R. microplus | Adults | 1 | 50 | 65.5 and 55.9 | [107] |
| 15 | rVoraxin from Rhipicephalus appendiculatus | 3 | JW | R. appendiculatus | Adults | 3 | 60 | 26,7 | [86] |
| 16 | IrFER2 | 4 | NR | I. ricinus | Nymphs | 3 | 50 | 98 | [97] |
| 17 | Haemaphysalis longicornis metalloprotease (HLMP1) | 3 | NR | H. longicornis | Nymphs and adults | 3 | 120 | 15,6 and 14,6 | [147] |
| 18 | Serpin | 6 | NZ | R. microplus | Larvae | 3 | 500 | 67 | [88] |
| 19 | Salivary antigens P8, P19, P23 and P32 | 3 | NZ | I. scapularis | Nymphs | 3 | 50 | NA | [95] |
| 20 | P0 protein and Bm86 | 10 | C | R. sanguineus s.l. | Nymphs and adults | 4 | 400 | 90 | [57] |
| 21 | Subolesin Ornithodoros erraticus and Ornithodoros moubata (rOeSub and rOmSub) | 9 | NZ | O. erraticus and O. moubata | Nymphs and adults | 3 | 90 | 8,5 and 24,3 | [79] |
| 22 | REnolase | 3 | NZ | O. moubata | Adults | 3 | 90 | NA | [108] |
| 23 | Haemaphysalis longicornis ferretin 1 (HlFER1) | 3 | JW | H. longicornis | Adults | 1 | 50 | NA | [65] |
| 24 | Subolesin/akirin orthologues of Ornithodoros erraticus (OE1, OE2 and OM1) | 3 | NZ | O. erraticus | Adults and nymphs | 3 | 200 | 48,6, 83,1 and 50,3 | [80] |
| 25 | Q38 | 3 | NZ | I. ricinus and D. reticulatus | Larvae | 2 | 200 | 99,9 and 43,6 | [39] |
| 26 | Ornithodoros moubata salivary lipocalin (TSGP4) | 6 | NZ | O. moubata | Adults and nymphs | 3 | 100 | 14,1 | [109] |
| 27 | Amblyomma americanum serine protease inhibitor 19 (AAS19) | 2 | NZ | Amblyomma americanum | Adults | 2 | 40 | 60 | [110] |
| 28 | ATAQ protein from Rhipicephalus microplus | 9 | NZ | R. sanguineus s.l. | Adults | 3 | NR | 47 | [111] |
| 29 | Rhipicephalus microplus ticks from Brazil (Bm05br) | 1 | NZ | R. sanguineus s.l. | Adults | 3 | NR | NA | [112] |
| 30 | CoAQP | 6 | NZ | I. ricinus | Larvae | 2 | 200 | 32 and 80 | [96] |
| 31 | Glutathione S-transferase from Haemaphysalis longicornis (GST-Hl) | 14 | NZ | R. sanguineus s.l. and R. appendiculatus | Nymphs and adults | 3 | 190 | 67 | [113] |
| 32 | RmGRP | N/A | NZ | R. microplus | N/A | 9 | N/A | NA | [87] |
| 33 | OM85 and OM03 | 6 | NZ | O. moubata | Nymphs and adults | 3 | 40 | 20,7 and 66,1 | [114] |
| 34 | Aquaporin of Ornithodoros erraticus (OeAQP) and selenoprotein T of Ornithodoros moubata (OeSEL) | 9 | NZ | O. erraticus and O. moubata | Nymphs and adults | 3 | 180 | 47,5 and 22,5 | [35] |
| 35 | Glutathione S-transferase GST-cocktail | 6 | NZ | R. sanguineus s.l. | Adults | 3 | 60 | 35 | [28] |
| 36 | Bmcys2c | 6 | NZ | R. appendiculatus | Nymphs and adults | 3 | 250 | 11,5 | [23] |
| 37 | Dermacentor marginatus S-transferase (DmGST) | 6 | NZ | D. marginatus | Nymphs and adults | 3 | 110 | 43,6 | [30] |
| 38 | Haemaphysalis longicornis lipocalin (HlLIP) | 6 | NZ | H. longicornis | Adults | 3 | 46 | 60,1 | [24] |
| 39 | Cathepsin L and tropomyosin proteins derived from Rhipicephalus microplus (CaTroRh) | 6 | NZ | R. microplus | NA | 3 | NA | NA | [115] |
| 40 | Ferritin 2 in Ornithodoros moubata (OmFer2) | 6 | NZ | O. moubata | Nymphs and adults | 3 | 95 | 71 | [78] |
| 41 | athepsin L rom Hyalomma asiaticum (HasCPL) | 6 | NZ | Hy. asiaticum | Larvae | 3 | 250 | 55 | [116] |
| 42 | Triosephosphate isomerase homologue from Haemaphysalis longicornis (HlTIM) | 27 | NZ | H. longicornis | Adults | 1 | 92 | 50 | [66] |
| 43 | RmS-17, and Bm86 | 6 | NZ | R. microplus | Adults | 3 | 120 | 79 and 62 | [47] |
| 44 | ATAQ in Haemaphysalis longicornis (HlATAQ) | 2 | JW | H. longicornis | Adults | 2 | 30 | NA | [117] |
| 45 | Hexokinase of Haemaphysalis longicornis (HlHK) | 12 | NZ | H. longicornis | Adults | 3 | 46 | 65.6 | [118] |
| 46 | Acid tail salivary protein (OeATSP), multiple coagulation factor deficiency protein 2 homolog (OeMCFD2), Cu/Zn-superoxide dismutase (OeSOD) and sulfotransferase (OeSULT) of Ornithodoros erraticus | 6 | NZ | O. erraticus | Nymphs and adults | 3 | 95 | 58.3 | [119] |
NR: Not reported. NA: Not applicable. NZ: New Zealand. JW: Japanese white. C: Californians. FL: Flemish lop-eared rabbit, and No: Number of ticks per rabbit.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



