
Submitted:
01 August 2023
Posted:
03 August 2023
You are already at the latest version
Abstract
There is a growing body of evidence supporting the significant role of bacterial biofilms in the pathogenesis of various human diseases including cancer. Biofilms are polymicrobial communities enclosed within an extracellular matrix composed of polysaccharides, proteins, extracellular DNA, and lipids. This complex matrix provides protection against antibiotics and host immune responses, enabling the microorganisms to establish persistent infections. Moreover, biofilms induce anti-inflammatory responses and metabolic changes in the host, further facilitating their survival. Many of these changes are comparable to those observed in cancer cells. This review will cover recent research on the role of bacterial biofilms in carcinogenesis, especially in colorectal (CRC) and gastric cancers, emphasizing the shared physical and chemical characteristics of biofilms and cancer. This review will also discuss the interactions between bacteria and the tumor microenvironment, which can facilitate oncogene expression and cancer progression. This information will provide insight into developing new therapies to identify and treat biofilm-associated cancers, such as utilizing bacteria as delivery vectors, using bacteria to upregulate immune function, or more selectively targeting biofilms and cancer for their shared traits.
Keywords:
extracellular polymeric substance (EPS)
; tumor microenvironment (TME)
; tumor microbiome (TM)
; tumor-associated macrophages (TAM)
; colorectal cancer (CRC)
; Fusobacterium nucleatum (Fn)
; Helicobacter py
1. Introduction
Biofilms are estimated to be involved in ~80% of all human infections as well as various types of human pathologies, including cancer [1]. Biofilms are communities, often multispecies communities, of microorganisms enclosed within a self-constructed matrix of extracellular polymeric substances such as polysaccharides, proteins, extracellular DNA, and lipids [2]. This matrix can either adhere to a surface, such as skin, wound tissue, or indwelling medical devices, or become suspended within a secondary matrix of host-derived pus or mucus [3]. Over time, biofilms mature into a mushroom shaped structure, and at the end of their development can release microorganisms to colonize new surfaces and serve as reservoirs of infection.
Biofilm contains various microorganisms, even multicellular organisms. The physiological states of these organisms are heavily contingent upon their location both within the body and within the biofilm. The limited diffusion of chemicals within a biofilm creates gradients for O2, pH and nutrients, with the center zones often being hypoxic and nutrient poor. Microorganisms within the biofilm may, therefore, be actively dividing, non-dividing, metabolically active, or inactive depending on their conditions within the biofilm microenvironment [4]. Organisms in a biofilm can also respond to environmental stimuli via quorum sensing, using small signaling molecules that can up- or down-regulate stress tolerance, metabolic, or virulence genes in the colony [5].
Emerging research suggests a link between biofilm and cancer, even beyond the conventionally accepted links to colorectal cancer (CRC) and gastric cancer. Malignant cells in vivo are supported by a tumor microenvironment (TME) that consists of various immune cells, fibroblasts, vascular endothelial cells, and adipocytes that communicate via chemokines, cytokines, growth factors, exosomes, proteoglycans, and glycoproteins [6]. The TME thus contributes to the cellular and genetic heterogeneity encapsulating tumors, immune cells, and bacteria, creating an abnormal environment that remodels the extracellular matrix (ECM) [7]. The altered ECM triggers changes in gene expression, leading to epithelial-to-mesenchymal transition (EMT), a hallmark of tumor cell migration and invasion. Moreover, the TME modulates immune cell functions, shifting them from pro-inflammatory to anti-inflammatory states. Regulatory T cells suppress immune responses, hindering the effector T cells’ ability to attack tumors. Myeloid-derived suppressor cells and tumor-associated macrophages also inhibit T cells and natural killer cells, further suppressing immune responses. In addition, tumor cells and other cells in the TME release immunosuppressive cytokines.
As a multicellular aggregation, cancer cells must be able to efficiently distribute nutrients, requiring metabolic changes collectively known as the Warburg effect. This metabolic transition in cancer cells allows sufficient ATP generation under hypoxic conditions and provides elevated level of glutamine, a viral substrate for cell growth. These changes can be accomplished by increased expression of transporters for the substrate and enzymes involved in its metabolism [8]. Similarly, biofilms are biological structures formed through cooperative interactions among microorganisms to ensure their survival. Within biofilms, microorganisms have developed specialized adaptive mechanisms to thrive in extreme environments, akin to the Warburg effect observed in cancer cells. These microorganisms efficiently metabolize glucose to derive energy and maintain the stability of the biofilm, even in oxygen-limited conditions [9]. The commonality between biofilms and cancer cells lies in their unique metabolic adaptations for nutrient distribution, supporting their survival and growth. Cancer cells sustain rapid proliferation by employing the Warburg effect to acquire high levels of substrates like glutamine [10]. Understanding the mechanisms behind these metabolic adaptations is a crucial challenge in cancer therapy and biofilm control.
Biofilm can promote caner through various mechanisms: 1) Biofilms can trigger inflammation that often fails to clear biofilm-associated pathogens. Persistent inflammation can cause DNA damage and promote the growth of cancer cells [1]. 2) Biofilms can modulate and limit the efficacy of the host immune response [11], creating an environment that supports cancer development [12]. 3) Some bacteria in biofilms can produce toxins functioning as carcinogens which increase cancer risk [13]. 4) New evidence increasingly suggests that bacteria within the TME, referred to as the tumor microbiome (TM), are involved in cancer progression [14]. Biofilms can alter the microenvironment surrounding cancer cells. The altered ECM protect cancer cells and facilitate their invasion and metastasis. Given the prevalence of biofilm formation in the human body and its detrimental impact on infections and cancer progression, there is an increasing emphasis on having a deeper understanding of bacteria behavior in cancer. Elucidating the mechanisms of biofilm contribution to cancer will allow us to improve diagnosis and treatment of biofilm-associated cancers [15], and will also open up new and innovative treatment options.
2. Biofilm and Colorectal Cancer (CRC)
Colorectal cancer (CRC) is one of the leading causes of cancer-related death worldwide [16]. Among various contributing factors, dysbiosis, an imbalance in the microbiota, appears to play a key role in its pathogenesis [17]. Research has found that patients with CRC have distinct microbiota compositions compared to healthy controls. CRC tissue contains elevated levels of an invasive anaerobe, Fusobacterium nucleatum (Fn), compared to healthy tissue, which is linked to lymph node metastasis [18,19]. In addition to dysbiosis, CRC is also associated with a compromised mucosal barrier [20]. The colon is lined with a dense layer of mucus that separates the microbiota from the human colonic epithelial cells. This mucus layer prevents direct contact between the microorganisms and the cells, but these mucosal barriers can be compromised when bacteria form biofilms, leading to chronic mucosal inflammation that eventually damages colonic epithelial cells (Figure 1A,B). Disruption of the colonic epithelial cells can result in the loss of E-cadherin, a cell-cell junction responsible for connecting epithelial cells, from the apical zonula adherens [21]. This condition is also accompanied by elevated levels of pro-inflammatory cytokines such as IL-6, increasing the likelihood of CRC development, particularly in individuals with obesity, diabetes mellitus, and smoking habits [22].
Damage to the mucus barrier allows microorganisms directly contact with colonic epithelial cells and can alter the mechanisms by which microorganisms metabolize, dramatically affecting nearby host cells. Heightened levels of acetylated polyamine metabolites such as N 1-acetylspermidine and N 1-acetylspermine have been found in biofilms-concentrated human colon cancer, driven by altered levels of bacterial acetyltransferases [23]. Polyamines are naturally occurring polycations in eukaryotes that can stabilize or distort the conformation of negatively charged macromolecules in cells, such as DNA, RNA, proteins, and acidic lipids containing phosphate groups. They play a critical role in cell growth and differentiation [24]. The polyamine pathway is a downstream target for many oncogenes, such as MYC transcription factor, p53, RAS, MEK, AKT, and mTOR [24]. Dysregulation of a polyamine metabolism is frequently observed in other cancer cells such as lung, breast, prostate, and gastric cancers [24]. This indicates that bacteria in biofilm manipulate the host polyamine metabolism, ultimately accelerating cell growth and tumor formation.
Deoxycholic acid (DCA) is another important metabolic modulator found in human colon biopsies and mouse models mediating CRC progression [25]. DCA promotes DNA damage and oxidative stress, enhancing proliferation of tumor cells, and suppressing apoptosis, functioning as a naturally produced carcinogen [26]. It is one of the secondary bile acids produced by the host but modified by intestinal bacteria. Biofilms can provide an optimal microenvironment where bile acids are changed into DCA through deconjugation, dehydroxylation and dehydrogenation [27] Colonic epithelial cells covered with biofilm, therefore, are exposed to significantly higher levels of DCA, initiating or aggravating CRC. Further use of the metabolomics analysis is an exciting area of study for investigating direct or indirect metabolic interactions of bacteria and hosts. Taken together, the available evidence strongly suggests that bacterial biofilms are a critical contributor to CRC. Aggregating and invasive pathogens like Fn in biofilm can initiate and progress CRC by eliciting prolonged inflammation and manipulating the host metabolism, which could serve as a biomarker, allowing for earlier drug interventions.
One of the objectives of the human microbiome project has been to identify bacteria and toxins related to CRC. For instance, Bacteroides fragilis and Enterobacteriaceae bacteria have been found in the mucosal biofilms of patients with inflammatory bowel disease (IBD), increasing host susceptibility to CRC [28]. Fn is a prevalent commensal but opportunistic pathogen in human oral cavities and can co-aggregate with almost all types of oral bacteria species associated with oral plaque-forming biofilm [29]. Along with other anaerobic bacteria like Campylobacter and Leptotrichia, Fn is a key pathogen associated with CRC [30,31]. Fn produces several key virulence factors. One such factor is outer membrane vesicles (OMVs) that contain proteins, lipids, and adhesion molecules. OMVs of Fn alone were shown to co-aggregate bacteria proved to be a powerful biofilm former [32]. OMVs also play a role in bacterial communication and interaction with environments including host cells and immune cells. Fn also attaches to and transports non-invasive bacteria into human epithelial cells, functioning as a shuttle vector for other pathogens [33]. Fn produces a toxin called Fusobacterium adhesin A (FadA) that is correlated with CRC tumorigenesis [13], lower survival rate [34], metastasis [35], and recurrence [36]. The toxin is responsible for bacterial adhesion, activation of signaling pathways, inflammation, and CRC cell proliferation [37]. Other toxins, such as genotoxins called polyketide synthase of Escherichia coli and enterotoxigenic Bacteroides fragilis toxin have been linked to colon tumors in mouse models [38], but what they cause has yet to be established.
3. Biofilm-associated Helicobacter pylori and gastric cancer
Helicobacter pylori (Hp), classified by the WHO as a class I carcinogen, plays a critical role in the development of gastric cancer, the fifth most prevalent cancer worldwide [39]. Gastric cancer was responsible for ~800,000 cancer-related deaths in 2020 [40]. Chronic Hp infection significantly increases the risk of developing gastric cancer and mucosa-associated lymphoid tissue (MALT) lymphoma [41,42]. Hp can survive in the low pH of the stomach through its use of urease and pH-sensing Bab A [43]. Chronic Hp infections create persistent inflammation, provoking the development of gastric cancer. Hp also expresses a toxin called cytotoxin-associated antigen A (CagA), which is internalized into host cells via the bacterial type IV secretion system and phosphorylated by kinases like Fyn and Lyn. It targets and interacts with several factors involved in inflammation and cytoskeleton rearrangement. CagA also activates NF-kB through interaction with a protein called YWHAE (14-3-3ε), influencing cell cycle regulation, signal transduction, and malignant transformation [44,45]. Additionally, CagA has been shown to promote EMT, which contributes to the generation of cancer stem cells [46,47].
The virulent mechanism of Hp in the harsh gastric niche appears to be closely linked to its adaptive ability to form biofilms. Various biomedical in vivo systems have demonstrated the existence of Hp biofilms in gastric biopsies. Recent transcriptomic and proteomic analyses have provided valuable insights into the transition from planktonic Hp to the biofilm phase, including higher gene expression of adhesins, flagella, toxins, efflux pumps, lipopolysaccharides (LPS), the Type 4 secretion system, urease, and hydrogenase for obtaining alternative energy sources. Conversely, factors involved in quorum sensing, metabolism and, translation were reduced in biofilm [48]. Hp also exhibits adaptive responses during biofilm formation, such as the secretion of OMVs, morphological changes from spiral to dormant but more resistant coccoid form, matrix production, and increased efflux pump activity [49]. The altered gene expression of Hp is also modulated depending on the stages of biofilm formation, with genes in the initial attachment stage showing different expression patterns compared to the maturation stage of biofilm. Besides the factors mentioned above, there are many other regulators that affect the physiology of Hp and promote the transition of planktonic Hp to biofilm form.
Antibiotic treatment of Hp is able to reduce risk of the cancer in asymptomatic patients [24], and in individuals with a family history of gastric cancer [50]. It can also reduce tumor size in gastric MALT lymphoma patients [51]. Biofilm-associated Hp, however, is resistant to clarithromycin, a commonly used antibiotic for Hp infections. Hp biofilms treated with clarithromycin in vitro displayed increased minimum bactericidal concentration and minimum inhibitory concentration, as well as a higher mutation rate, which contributes to their already significant genotypic variation [52,53]. There is also emerging resistance of Hp to other antibiotics such as levofloxacin and metronidazole [54]. In addition to becoming multi-drug resistant, Hp biofilm successfully defend against host immune response. For example, LPS of Hp are recognized via the Toll-like receptor (TLR) 2 system instead of a common TLR4 used by most of bacteria. The TLR2 pathway mediates a weaker host innate and acquired T-cell responses in the gastric mucosa allowing Hp to escape effective immune detection [55].
4. Biofilm in tumor microenvironment
Contrary to a once-believed medical dogma that cancer tissue is sterile, a growing body of evidence has demonstrated the presence of bacteria in various cancerous tumors, including breast, lung, and CRC [19,56,57]. Recent compelling evidence suggests that bacterial communities, known as tumor microbiome (TM), play a crucial role in the TME of many different types of cancer. Intriguingly, bacteria have been identified both in the extracellular compartment and inside cancer cells and immune cells [58]. Immunohistochemistry experiments have demonstrated the presence of Gram-negative bacteria LPS in both the nucleus and cytoplasm of tumor cells, while fluorescence in situ hybridization has detected bacterial 16S rRNA primarily in the cytoplasm. Correlative light and electron microscopy studies have provided visual evidence of bacteria near the nuclear membrane in human breast cancer samples [58]. Moreover, several studies have reported the presence of cell-wall deficient (L-form) bacteria within breast cancer cells, supporting the notion of their intracellular existence [58,59]. These intracellular bacteria within breast cancer cells can activate Rho-ROCK signaling, reorganize the cytoskeleton, and potentially facilitate metastasis and dissemination to distant parts of the body [59]. Similarly, studies have demonstrated the invasion of Fn in esophageal squamous cell carcinoma and its activation of the NFκB signaling pathway, thereby promoting tumor progression [60]. A toxin like FadA can also activates beta-catenin signaling inside the cancer cells, resulting in increased expression of transcription factors, oncogenes, and inflammatory genes, thereby promoting CRC cell proliferation [61].
A recent study utilized a cutting-edge technique called spatial transcriptomics to detect differential gene expression in slices of tumor tissue found the oral bacterium Fn in both CRC and oral squamous tumor cells. This suggests a potential link between oral health and cancers in different parts of the body (Figure 2), [62]. The areas colonized by bacteria exhibited a reduced number of cancer-killing T cells and showed suppressed immune responses. Furthermore, the cancer cells within these immunosuppressive microniches displayed significantly lower levels of p53, a tumor suppressor protein, indicating that the bacteria resided within highly transformed cancer cells in the TME. The oral epithelial cancer cells harboring bacteria showed significant upregulation of genes involved in signaling pathways such as EMT and interferon-mediated response. The study also revealed that the bacteria-containing epithelial cells were detached from the spheroids and had invaded the surrounding tissue as single cells, thereby promoting cancer progression. In addition, intratumoral microbiota compromised the effect of a common chemotherapy drug like 5-fluorouracil (5-FU), which also possesses antimicrobial activity [63]. 5-FU was expected to negatively affect Fusobacterium in the TME and enhance the effectiveness of anti-cancer therapy. Instead, E. coli, a member of intratumoral microbiota, played a protective role in CRC by metabolizing the drug. These findings suggest that intratumoral microbes actively contribute to cancer development, progression, and response to treatment. However, it remains unclear if bacteria simply exploit the pre-established immunosuppressive regions of the TME or if they actively contribute to the immunosuppression within cancer-promoting niches to further thrive. It is also unclear whether a specific intratumoral bacteria or a heterogeneous microbial community like biofilm is more important in promoting cancer, or if this relationship works differently in different types of cancer. Other reports have demonstrated the presence of fungal species and viruses in the TME as well, suggesting that this effect may not be limited to bacteria [64,65].
As research on biofilms in the tumor microenvironment is still in its early stages, many questions remain unanswered. The composition, dynamics, and functional significance of biofilms can vary significantly between different cancer types, stages, and individual patients. Therefore, a comprehensive understanding of biofilm-mediated effects in cancer will require further investigation through advanced techniques such as spatial transcriptomics, single-cell sequencing, and high-resolution imaging. Identifying monoclonal bacterial strains functioning as key cancer promoters would be beneficial to the anti-tumor therapeutics. Ultimately, elucidating the intricate interactions between biofilms, the tumor microenvironment, and cancer cells holds great potential for the development of innovative therapeutic strategies that can target these microbial communities and enhance the efficacy of existing cancer treatments. Such knowledge may lead to the discovery of novel biomarkers, the development of personalized treatment regimens, and the advancement of precision medicine in the fight against cancer.
5. Therapeutics
Biofilm-associated cancer therapeutics face a variety of challenges, but there are a variety of emerging therapies that seek to target their joint weakness. One such strategy would be to target an integral bacterium that drive cancer proliferation, which is complicated by the antibiotic resistance offered by the biofilm itself. One promising therapeutic strategy has been to use mucolytic agents such as N-acetyl cysteine (NAC). NAC has been shown to disperse H. pylori biofilms in in vivo infections [66], and when used in conjunction with a dual therapy of clarithromycin and a proton pump inhibitor, it has resulted in a significantly greater reduction in H. pylori load than the dual therapy alone [67]. These results suggest that NAC may be an effective way to clear carcinogenic infections, but further studies are needed to determine its potential application in antimicrobial therapies.
Other treatment options would attempt to target biofilms and cancer simultaneously. N-Acyl-homoserine lactones (AHL) are quorum sensing molecules important for biofilm formation for many bacteria and have been implicated in carcinogenesis for oral squamous cell carcinomas OSCC) by repressing NFκB signaling [2,68]. Certain bacteria such as Pseudomonas aeruginosa use AHL signaling to inhibit bacterial competitors and can even modulate eukaryotic immune cell function [69]. Some synthetic analogs of the AHLs produced by P. aeruginosa have demonstrated antimicrobial properties and antiproliferative, or even apoptotic effects on OSCC cells [69,70]. Biofilms especially seem to provide a protective coating for tumors. Extratumoral E. coli biofilms can reduce the efficacy of antibiotics and anti-cancer drugs, create closer cell-cell contacts, improve cancer cell viability, and increase metastatic potential [71]. Treatment with antibiofilm agents, in association with antibiotic and anti-cancer drugs, improved cancer eradication outcomes, demonstrating that the biofilm offered important protection for both the bacteria and the cancer cells [71].
Bacteria are also known to modulate the efficacy and toxicity of existing cancer treatments. For instance, the efficacy of monoclonal antibodies that blockade CTLA-4, such as ipilimumab, on preventing tumor growth are contingent upon T cell interactions with specific Bacteroides species [72]. Tumors in germ free mice models were unresponsive to treatment, but when B. fragilis and/or B. thetaiotaomicron were reintroduced via oral cultures or fecal material transfers, treatment efficacy was restored [72]. Similar effects are observed with cyclophosphamide, oxaliplatin and cisplatin, wherein microbiota depletion was associated with lower anti-tumor efficacy [73,74]. Bacteria can also minimize the collateral damage induced by conventional cancer treatments. For instance, treatment with Lactobacillus probiotics after radiotherapy reduces the severity of associated diarrhea [75,76,77].
While bacteria and the biofilms they inhabit can pose significant problems for cancer patients, there is increasing research into ways bacteria can be utilized to treat cancer. One potential benefit of this is that bacteria can be used to selectively target cancer cells, minimizing the general toxicity of conventional treatments. Research in this area is exploring the possibility of using those bacteria as a delivery system for cancer-killing compounds. For instance, Salmonella typhimurium are chemoattracted to the microenvironment within the quiescent and necrotic areas of solid tumor models, where they preferentially grow [78]. Attenuated S. typhimurium are able to create large necrotic zones in the center of the tumors and stunt melanoma growth [79]. Other research has used S. typhimurium as a vector to deliver prodrug enzymes, immunomodulatory molecules, or toxins that further reduced tumor growth or even induced tumor regression [80]. For example, a thymidine kinase expressed by S. typhimurium in conjunction with additional prodrugs allows for targeted treatment of breast and colon carcinomas, resulting in greater reductions in tumor growth than bacteria treatment alone [81]. Modified S. typhimurium that expressed interleukin-2 were able to induce tumor regression or slow tumor growth in melanomas and osteosarcomas, and prevent the formation of metastases [82,83,84].
By harnessing the immunomodulatory properties of certain bacterial components, researchers aim to enhance the body’s natural defenses against cancer. Bacterial peptidoglycan, LPS, flagella, nucleic acids are recognized by pattern recognition receptors on macrophages, dendritic cells and neutrophils, triggering immune responses [85]. The activated immune response includes activation of NF-κB and STAT-3 that enhance anti-tumor activity by stimulating cytokine production from macrophages, dendritic cells, natural killer cells and B cells, and induces proliferation of immune cells [86,87]. This bacteria-based immunotherapy can be an effective treatment that modulate host immune system to recognize and target cancer cells with minimal side effects. Bacteria can also be engineered to express specific immune-stimulating molecules to further boost their immunotherapeutic potential. Immune checkpoint inhibitors have shown promising results in enhancing efficacy of cancer immunotherapies [88]. Immune checkpoints proteins are present on cells including tumor cells and are recognized and bound to proteins on immune cells like T cells. This interaction prevents strong immune response from destroying cancer cells as well as healthy cells. Immune checkpoint inhibitors are designed to block checkpoint proteins from binding with their partner proteins, allowing T cells to target cancer cells. Current immune checkpoint inhibitors are against checkpoint proteins, CTLA-4 and programmed death-ligand (PD-1) or its partner protein PD-L1. However, this area of research is still in its early stages, and more investigations are required to optimize the approach and ensure safety in clinical settings
Nanoparticles are tiny particles with sizes typically in the range of 1 to 100 nanometers. Due to their small size and unique properties, nanoparticles can be engineered to target specific tissues, including tumor tissues and biofilms, offering a more effective and localized delivery of therapeutic agents. The nanoparticles can be designed to penetrate the biofilm matrix to deliver drugs directly to biofilm-infected tumor sites, overcoming one of the major challenges of biofilm treatment – limited drug penetration. Moreover, nanoparticles can carry multiple agents simultaneously, allowing for combination therapies that target both the biofilm and cancer cells. This strategy has the potential to enhance treatment efficacy and reduce the risk of drug resistance. As with other emerging therapeutic approaches, nanoparticle-based therapies are still in preclinical stages of research and development.
It is important to note that research in this field is still in its early stages, and many of these approaches require further study and validation. The complexity of biofilm interactions and the heterogeneity of cancer types present significant challenges. Nevertheless, biofilm-associated cancer therapeutics offer a promising avenue for developing more effective and targeted treatments to improve cancer patient outcomes
6. Conclusion
Until recently, only dozens of declared bacteria like Fn and H. pylori have been considered to cause carcinogenesis by inducing inflammation and altering signal transduction that influence mucosal cells. Biotechnology utilizing single cell genomics and multi-omics have provided a new concept that TM is present within many tumor tissues and interact with host cells. TM is found in tumor regions where T cell recruitment and function are suppressed promoting tumor progression. Given the fact that the immune-privileged and hypoxic TME can provide an ideal environment where obligate and facultative pathogenic anaerobes bacteria can proliferate and colonize, it is presumed to contain biofilms.
Biofilm is one of the successful forms of life for microorganisms. It provides protection for microorganisms from immune detection and a favorable environment to adjust to environment. Now it is well recognized that biofilm plays a key role in the cancer progression as well as persistence of many bacterial infections. Although it is now widely accepted that the TME contains many types of microorganisms inside and outside of tumor cells and immune cells, it is not yet determined the lifestyle of the microorganisms. The unique complexity and adaptation of biofilm may bring changes in host’s peripheral environment, metabolism, and immune responses, facilitating host cells’ transformation, tumor progression. We suggest future cancer therapeutics consider to combine cancer chemotherapy and bacteriotherapy along with biofilm-disrupting agents. The innovative therapeutic agents can help defeat tumor progression, metastasis, and drug resistance more effectively.
Author Contributions
Conceptualization, E.C. and S.C.; writing—original draft preparation, E.C. and B. M.; figure and proofreading, L.H.; writing—review and editing, E.C. and S.C.; supervision, S.C.; funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2016R1D1A1B01009752 S.C.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Laura Hailey for figure production.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vestby, L.K.; Gronseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Ghosh, A.; Jayaraman, N.; Chatterji, D. Small-Molecule Inhibition of Bacterial Biofilm. ACS Omega 2020, 5, 3108–3115. [Google Scholar] [CrossRef] [PubMed]
- Sonderholm, M.; Kragh, K.N.; Koren, K.; Jakobsen, T.H.; Darch, S.E.; Alhede, M.; Jensen, P.O.; Whiteley, M.; Kuhl, M.; Bjarnsholt, T. Pseudomonas aeruginosa Aggregate Formation in an Alginate Bead Model System Exhibits In Vivo-Like Characteristics. Appl Environ Microbiol 2017, 83. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.D.; Olson, M.E. Current concepts in biofilm formation of Staphylococcus epidermidis. Future Microbiol 2010, 5, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [PubMed]
- Elgundi, Z.; Papanicolaou, M.; Major, G.; Cox, T.R.; Melrose, J.; Whitelock, J.M.; Farrugia, B.L. Cancer Metastasis: The Role of the Extracellular Matrix and the Heparan Sulfate Proteoglycan Perlecan. Front Oncol 2019, 9, 1482. [Google Scholar] [CrossRef] [PubMed]
- Cluntun, A.A.; Lukey, M.J.; Cerione, R.A.; Locasale, J.W. Glutamine Metabolism in Cancer: Understanding the Heterogeneity. Trends Cancer 2017, 3, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Wan, N.; Wang, H.; Ng, C.K.; Mukherjee, M.; Ren, D.; Cao, B.; Tang, Y.J. Bacterial Metabolism During Biofilm Growth Investigated by (13)C Tracing. Front Microbiol 2018, 9, 2657. [Google Scholar] [CrossRef]
- Gong, T.; Zheng, C.; Ou, X.; Zheng, J.; Yu, J.; Chen, S.; Duan, Y.; Liu, W. Glutamine metabolism in cancers: Targeting the oxidative homeostasis. Front Oncol 2022, 12, 994672. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis--the ’accidental’ pathogen. Nat Rev Microbiol 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Ellermann, M.; Sartor, R.B. Intestinal bacterial biofilms modulate mucosal immune responses. J Immunol Sci 2018, 2, 13–18. [Google Scholar] [PubMed]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Ciernikova, S.; Sevcikova, A.; Stevurkova, V.; Mego, M. Tumor microbiome - an integral part of the tumor microenvironment. Front Oncol 2022, 12, 1063100. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M. Molecular Mechanism of Helicobacter pylori-Induced Gastric Cancer. J Gastrointest Cancer 2021, 52, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Kang, M.; Martin, A. Microbiome and colorectal cancer: Unraveling host-microbiota interactions in colitis-associated colorectal cancer development. Semin Immunol 2017, 32, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Burns, M.B.; Lynch, J.; Starr, T.K.; Knights, D.; Blekhman, R. Virulence genes are a signature of the microbiome in the colorectal tumor microenvironment. Genome Med 2015, 7, 55. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Genua, F.; Raghunathan, V.; Jenab, M.; Gallagher, W.M.; Hughes, D.J. The Role of Gut Barrier Dysfunction and Microbiome Dysbiosis in Colorectal Cancer Development. Front Oncol 2021, 11, 626349. [Google Scholar] [CrossRef]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Mark Welch, J.L.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc Natl Acad Sci U S A 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Xi, Y.; Huang, Z.; Xu, P. Linking Obesity with Colorectal Cancer: Epidemiology and Mechanistic Insights. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Dejea, C.M.; Edler, D.; Hoang, L.T.; Santidrian, A.F.; Felding, B.H.; Ivanisevic, J.; Cho, K.; Wick, E.C.; Hechenbleikner, E.M.; et al. Metabolism links bacterial biofilms and colon carcinogenesis. Cell Metab 2015, 21, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Meng, Y.; Wu, X.; Sun, Y. Polyamines and related signaling pathways in cancer. Cancer Cell Int 2020, 20, 539. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, C.; Holubec, H.; Bhattacharyya, A.K.; Nguyen, H.; Payne, C.M.; Zaitlin, B.; Bernstein, H. Carcinogenicity of deoxycholate, a secondary bile acid. Arch Toxicol 2011, 85, 863–871. [Google Scholar] [CrossRef]
- Cao, H.; Luo, S.; Xu, M.; Zhang, Y.; Song, S.; Wang, S.; Kong, X.; He, N.; Cao, X.; Yan, F.; et al. The secondary bile acid, deoxycholate accelerates intestinal adenoma-adenocarcinoma sequence in Apc (min/+) mice through enhancing Wnt signaling. Fam Cancer 2014, 13, 563–571. [Google Scholar] [CrossRef]
- Guzior, D.V.; Quinn, R.A. Review: microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- Tomkovich, S.; Dejea, C.M.; Winglee, K.; Drewes, J.L.; Chung, L.; Housseau, F.; Pope, J.L.; Gauthier, J.; Sun, X.; Muhlbauer, M.; et al. Human colon mucosal biofilms from healthy or colon cancer hosts are carcinogenic. J Clin Invest 2019, 129, 1699–1712. [Google Scholar] [CrossRef]
- Allen-Vercoe, E.; Strauss, J.; Chadee, K. Fusobacterium nucleatum: an emerging gut pathogen? Gut Microbes 2011, 2, 294–298. [Google Scholar] [CrossRef]
- Warren, R.L.; Freeman, D.J.; Pleasance, S.; Watson, P.; Moore, R.A.; Cochrane, K.; Allen-Vercoe, E.; Holt, R.A. Co-occurrence of anaerobic bacteria in colorectal carcinomas. Microbiome 2013, 1, 16. [Google Scholar] [CrossRef]
- Wirbel, J.; Pyl, P.T.; Kartal, E.; Zych, K.; Kashani, A.; Milanese, A.; Fleck, J.S.; Voigt, A.Y.; Palleja, A.; Ponnudurai, R.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat Med 2019, 25, 679–689. [Google Scholar] [CrossRef]
- Kinder, S.A.; Holt, S.C. Localization of the Fusobacterium nucleatum T18 adhesin activity mediating coaggregation with Porphyromonas gingivalis T22. J Bacteriol 1993, 175, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.M.; Grossman, T.J.; Rudney, J.D. Fusobacterium nucleatum transports noninvasive Streptococcus cristatus into human epithelial cells. Infect Immun 2006, 74, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Kunzmann, A.T.; Proenca, M.A.; Jordao, H.W.; Jiraskova, K.; Schneiderova, M.; Levy, M.; Liska, V.; Buchler, T.; Vodickova, L.; Vymetalkova, V.; et al. Fusobacterium nucleatum tumor DNA levels are associated with survival in colorectal cancer patients. Eur J Clin Microbiol Infect Dis 2019, 38, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/beta-catenin modulator Annexin A1. EMBO Rep 2019, 20. [Google Scholar] [CrossRef]
- Dejea, C.M.; Sears, C.L. Do biofilms confer a pro-carcinogenic state? Gut Microbes 2016, 7, 54–57. [Google Scholar] [CrossRef]
- World Cancer Research Fund International. Stomach cancer statistics. Available online: https://www.wcrf.org/cancer-trends/stomach-cancer-statistics (accessed on 02 February 2023). (0228),.
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Han, Z.; Liu, J.; Zhang, W.; Kong, Q.; Wan, M.; Lin, M.; Lin, B.; Ding, Y.; Duan, M.; Li, Y.; et al. Cardia and non-cardia gastric cancer risk associated with Helicobacter pylori in East Asia and the West: A systematic review, meta-analysis, and estimation of population attributable fraction. Helicobacter 2023, 28, e12950. [Google Scholar] [CrossRef]
- Parsonnet, J.; Hansen, S.; Rodriguez, L.; Gelb, A.B.; Warnke, R.A.; Jellum, E.; Orentreich, N.; Vogelman, J.H.; Friedman, G.D. Helicobacter pylori infection and gastric lymphoma. N Engl J Med 1994, 330, 1267–1271. [Google Scholar] [CrossRef]
- Bugaytsova, J.A.; Bjornham, O.; Chernov, Y.A.; Gideonsson, P.; Henriksson, S.; Mendez, M.; Sjostrom, R.; Mahdavi, J.; Shevtsova, A.; Ilver, D.; et al. Helicobacter pylori Adapts to Chronic Infection and Gastric Disease via pH-Responsive BabA-Mediated Adherence. Cell Host Microbe 2017, 21, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Aitken, A. 14-3-3 proteins: a historic overview. Semin Cancer Biol 2006, 16, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Jan, Y.J.; Ko, B.S.; Liang, S.M.; Liou, J.Y. Involvement of 14-3-3 Proteins in Regulating Tumor Progression of Hepatocellular Carcinoma. Cancers (Basel) 2015, 7, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Sougleri, I.S.; Papadakos, K.S.; Zadik, M.P.; Mavri-Vavagianni, M.; Mentis, A.F.; Sgouras, D.N. Helicobacter pylori CagA protein induces factors involved in the epithelial to mesenchymal transition (EMT) in infected gastric epithelial cells in an EPIYA- phosphorylation-dependent manner. FEBS J 2016, 283, 206–220. [Google Scholar] [CrossRef]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat Rev Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Hathroubi S., Z. J., Ottemann K.M. Helicobacter pylori Biofilm Cells are Metabolically Distinct, Express Flagella, and Antibiotic Tolerant. bioRxiv 2019. [Google Scholar] [CrossRef]
- Krzyzek, P.; Grande, R.; Migdal, P.; Paluch, E.; Gosciniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020, 9, 50. [Google Scholar] [CrossRef]
- Choi, I.J.; Kim, C.G.; Lee, J.Y.; Kim, Y.I.; Kook, M.C.; Park, B.; Joo, J. Family History of Gastric Cancer and Helicobacter pylori Treatment. N Engl J Med 2020, 382, 427–436. [Google Scholar] [CrossRef]
- Stathis, A.; Chini, C.; Bertoni, F.; Proserpio, I.; Capella, C.; Mazzucchelli, L.; Pedrinis, E.; Cavalli, F.; Pinotti, G.; Zucca, E. Long-term outcome following Helicobacter pylori eradication in a retrospective study of 105 patients with localized gastric marginal zone B-cell lymphoma of MALT type. Ann Oncol 2009, 20, 1086–1093. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Hanawa, T.; Kurata, S.; Ochiai, K.; Kamiya, S. Impact of Helicobacter pylori biofilm formation on clarithromycin susceptibility and generation of resistance mutations. PLoS One 2013, 8, e73301. [Google Scholar] [CrossRef]
- Waskito, L.A.; Salama, N.R.; Yamaoka, Y. Pathogenesis of Helicobacter pylori infection. Helicobacter 2018, 23 Suppl 1, e12516. [Google Scholar] [CrossRef]
- Dascalu, R.I.; Bolocan, A.; Paduaru, D.N.; Constantinescu, A.; Mitache, M.M.; Stoica, A.D.; Andronic, O. Multidrug resistance in Helicobacter pylori infection. Front Microbiol 2023, 14, 1128497. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, M.; El-Zataari, M.; Owyang, S.Y.; Eaton, K.A.; Liu, M.; Chang, Y.M.; Zou, W.; Kao, J.Y. TLR2 mediates Helicobacter pylori-induced tolerogenic immune response in mice. PLoS One 2013, 8, e74595. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.; Shamonki, J.M.; Chung, A.; Dinome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS One 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the microbiome and TP53 in human lung cancer. Genome Biol 2018, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef]
- Fu, A.; Yao, B.; Dong, T.; Chen, Y.; Yao, J.; Liu, Y.; Li, H.; Bai, H.; Liu, X.; Zhang, Y.; et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022, 185, 1356–1372 e26. [Google Scholar] [CrossRef]
- Yang, Y.; Weng, W.; Peng, J.; Hong, L.; Yang, L.; Toiyama, Y.; Gao, R.; Liu, M.; Yin, M.; Pan, C.; et al. Fusobacterium nucleatum Increases Proliferation of Colorectal Cancer Cells and Tumor Development in Mice by Activating Toll-Like Receptor 4 Signaling to Nuclear Factor-kappaB, and Up-regulating Expression of MicroRNA-21. Gastroenterology 2017, 152, 851–866. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/beta-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Galeano Nino, J.L.; Wu, H.; LaCourse, K.D.; Kempchinsky, A.G.; Baryiames, A.; Barber, B.; Futran, N.; Houlton, J.; Sather, C.; Sicinska, E.; et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature 2022, 611, 810–817. [Google Scholar] [CrossRef] [PubMed]
- LaCourse, K.D.; Zepeda-Rivera, M.; Kempchinsky, A.G.; Baryiames, A.; Minot, S.S.; Johnston, C.D.; Bullman, S. The cancer chemotherapeutic 5-fluorouracil is a potent Fusobacterium nucleatum inhibitor and its activity is modified by intratumoral microbiota. Cell Rep 2022, 41, 111625. [Google Scholar] [CrossRef] [PubMed]
- Inamura, K. Beyond Bacteria: Fungi in the Tumor Microbiome. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Broecker, F.; Moelling, K. The Roles of the Virome in Cancer. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Cammarota, G.; Branca, G.; Ardito, F.; Sanguinetti, M.; Ianiro, G.; Cianci, R.; Torelli, R.; Masala, G.; Gasbarrini, A.; Fadda, G.; et al. Biofilm demolition and antibiotic treatment to eradicate resistant Helicobacter pylori: a clinical trial. Clin Gastroenterol Hepatol 2010, 8, 817–820 e3. [Google Scholar] [CrossRef]
- Gurbuz, A.K.; Ozel, A.M.; Ozturk, R.; Yildirim, S.; Yazgan, Y.; Demirturk, L. Effect of N-acetyl cysteine on Helicobacter pylori. South Med J 2005, 98, 1095–1097. [Google Scholar] [CrossRef]
- Sathish Sankar a 1, M.Y.b. , Pitchaipillai Sankar Ganesh. Acyl homoserine lactone inhibitors for oral squamous cell carcinoma – Novel insights and therapeutic perspective. Medical Hypotheses 2023, 170, 69. [Google Scholar] [CrossRef]
- Chai, H.; Hazawa, M.; Shirai, N.; Igarashi, J.; Takahashi, K.; Hosokawa, Y.; Suga, H.; Kashiwakura, I. Functional properties of synthetic N-acyl-L-homoserine lactone analogs of quorum-sensing gram-negative bacteria on the growth of human oral squamous carcinoma cells. Invest New Drugs 2012, 30, 157–163. [Google Scholar] [CrossRef]
- Chai, H.; Hazawa, M.; Hosokawa, Y.; Igarashi, J.; Suga, H.; Kashiwakura, I. Novel acridine-based N-acyl-homoserine lactone analogs induce endoreduplication in the human oral squamous carcinoma cell line SAS. Biol Pharm Bull 2012, 35, 1257–1263. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, S.Y.; Chua, S.L.; Khoo, B.L. The effects of biofilms on tumor progression in a 3D cancer-biofilm microfluidic model. Biosens Bioelectron 2021, 180, 113113. [Google Scholar] [CrossRef]
- Vetizou, M.; Pitt, J.M.; Daillere, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillere, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Salminen, E.; Elomaa, I.; Minkkinen, J.; Vapaatalo, H.; Salminen, S. Preservation of intestinal integrity during radiotherapy using live Lactobacillus acidophilus cultures. Clin Radiol 1988, 39, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Delia, P.; Sansotta, G.; Donato, V.; Messina, G.; Frosina, P.; Pergolizzi, S.; De Renzis, C.; Famularo, G. Prevention of radiation-induced diarrhea with the use of VSL#3, a new high-potency probiotic preparation. Am J Gastroenterol 2002, 97, 2150–2152. [Google Scholar] [CrossRef]
- Urbancsek, H.; Kazar, T.; Mezes, I.; Neumann, K. Results of a double-blind, randomized study to evaluate the efficacy and safety of Antibiophilus in patients with radiation-induced diarrhoea. Eur J Gastroenterol Hepatol 2001, 13, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Kasinskas, R.W.; Forbes, N.S. Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol Bioeng 2006, 94, 710–721. [Google Scholar] [CrossRef]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-targeted Salmonella as a novel anticancer vector. Cancer Res 1997, 57, 4537–4544. [Google Scholar]
- Leschner, S.; Weiss, S. Salmonella-allies in the fight against cancer. J Mol Med (Berl) 2010, 88, 763–773. [Google Scholar] [CrossRef]
- Friedlos, F.; Lehouritis, P.; Ogilvie, L.; Hedley, D.; Davies, L.; Bermudes, D.; King, I.; Martin, J.; Marais, R.; Springer, C.J. Attenuated Salmonella targets prodrug activating enzyme carboxypeptidase G2 to mouse melanoma and human breast and colon carcinomas for effective suicide gene therapy. Clin Cancer Res 2008, 14, 4259–4266. [Google Scholar] [CrossRef]
- Saltzman, D.A.; Heise, C.P.; Hasz, D.E.; Zebede, M.; Kelly, S.M.; Curtiss, R., 3rd; Leonard, A.S.; Anderson, P.M. Attenuated Salmonella typhimurium containing interleukin-2 decreases MC-38 hepatic metastases: a novel anti-tumor agent. Cancer Biother Radiopharm 1996, 11, 145–153. [Google Scholar] [CrossRef]
- Sorenson, B.S.; Banton, K.L.; Frykman, N.L.; Leonard, A.S.; Saltzman, D.A. Attenuated Salmonella typhimurium with IL-2 gene reduces pulmonary metastases in murine osteosarcoma. Clin Orthop Relat Res 2008, 466, 1285–1291. [Google Scholar] [CrossRef]
- al-Ramadi, B.K.; Fernandez-Cabezudo, M.J.; El-Hasasna, H.; Al-Salam, S.; Bashir, G.; Chouaib, S. Potent anti-tumor activity of systemically-administered IL2-expressing Salmonella correlates with decreased angiogenesis and enhanced tumor apoptosis. Clin Immunol 2009, 130, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gorjifard, S.; Goldszmid, R.S. Microbiota-myeloid cell crosstalk beyond the gut. J Leukoc Biol 2016, 100, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-kappaB, STAT3 or AP-1 activation. Mol Med Rep 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Kathera, C.; Janardhan, A.; Praveen Kumar, A.; Viswanath, B. Surfacing role of probiotics in cancer prophylaxis and therapy: A systematic review. Clin Nutr 2017, 36, 1465–1472. [Google Scholar] [CrossRef]
- Xu, J.; Saklatvala, R.; Mittal, S.; Deshmukh, S.; Procopio, A. Recent Progress of Potentiating Immune Checkpoint Blockade with External Stimuli-an Industry Perspective. Adv Sci (Weinh) 2020, 7, 1903394. [Google Scholar] [CrossRef]
Figure 1.
Biofilm-mediated CRC carcinogenesis. (A, upper) In healthy individuals, the colon is lined with a dense layer of mucus that separates the microbiota from the human colonic epithelial cells (B, bottom) Biofilm causes mucous depletion and closely contact colonic epithelial cells. This leads to chronic mucosal inflammation, increased level of IL-6, resulting in host cell’s transformation and tumor progression.
Figure 1.
Biofilm-mediated CRC carcinogenesis. (A, upper) In healthy individuals, the colon is lined with a dense layer of mucus that separates the microbiota from the human colonic epithelial cells (B, bottom) Biofilm causes mucous depletion and closely contact colonic epithelial cells. This leads to chronic mucosal inflammation, increased level of IL-6, resulting in host cell’s transformation and tumor progression.

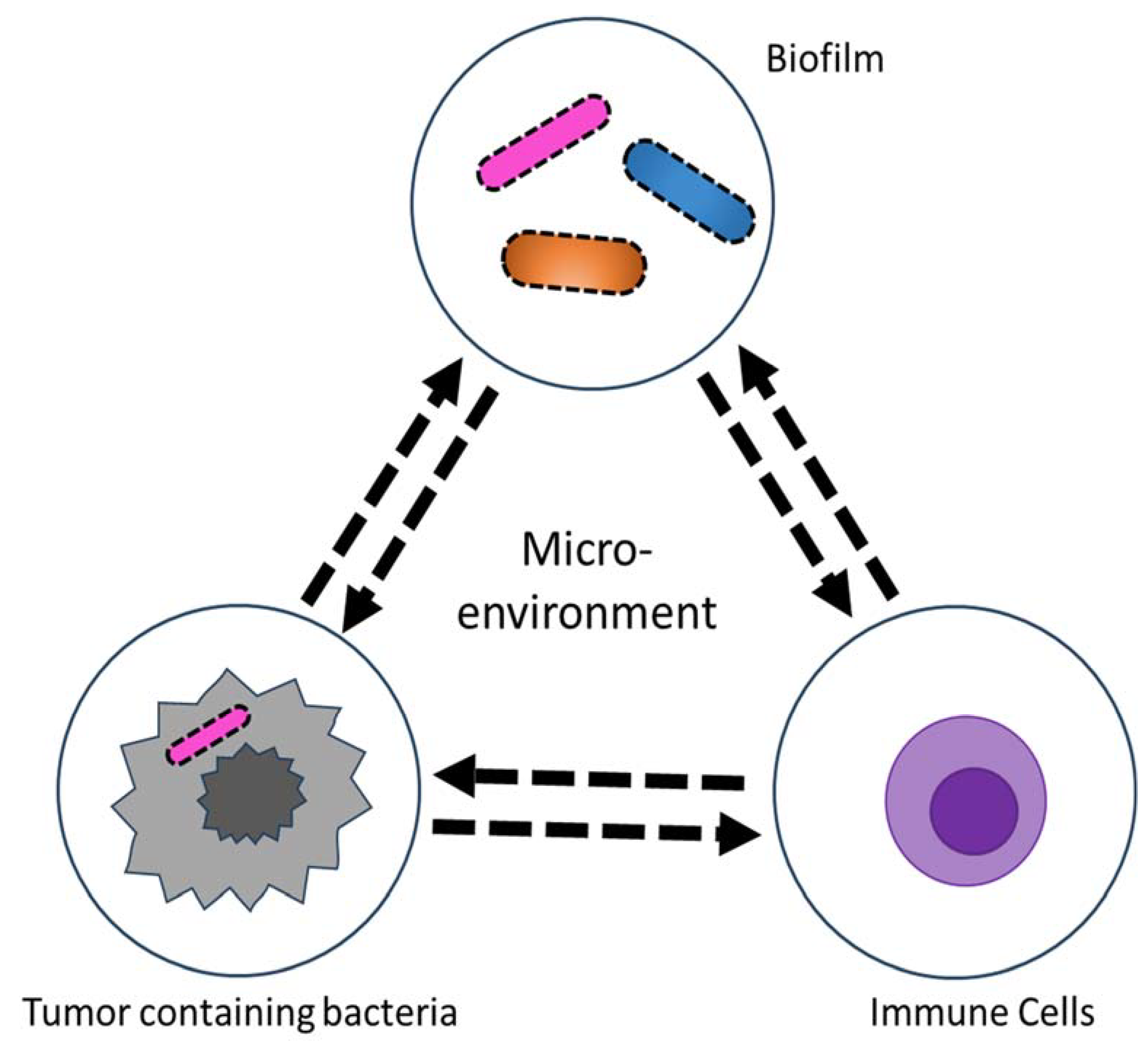
Figure 2.
Schematic of human tumor microenvironment that contains tumor microbiome. The tumor microbiome provides a unique environment where intratumoral or extratumoral microorganisms cross-talk with tumor cells and immune cells.
Figure 2.
Schematic of human tumor microenvironment that contains tumor microbiome. The tumor microbiome provides a unique environment where intratumoral or extratumoral microorganisms cross-talk with tumor cells and immune cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



