
Submitted:
03 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
Notch signaling is a highly conserved pathway which occurs between adjacent cells to distinguish them. The Notch signaling cascade operates in a simple mechanism, yet Notch plays remarkably diverse roles in development to regulate cell fate determination, organ growth and tissue patterning. Originally isolated and characterized in the model insect Drosophila melanogaster, recent studies in various insect species have revealed that Notch signaling is broadly involved in the construction of insect tissues. This review focuses on providing a molecular mechanistic and developmental picture regarding the roles of Notch pathway in insects. The functions of Notch in the formation and patterning of insect embryo, wing, leg, ovary and several specific structures as well as in stress responses are summarized. These results are discussed in developmental context with an eye toward understanding the functionality of Notch signaling pathway in different insect species.
Keywords:
Notch
; insect
; development
; Drosophila
1. Introduction of the Notch signaling pathway
A small number of signaling pathways are iteratively used to regulate cell fate determination, organ growth and tissue patterning during insect development. One of the most important pathways is mediated by Notch, which functions to distinguish adjacent cells in numerous developmental processes [1]. The Notch signaling cascade operates in a remarkably linear fashion without signal amplification steps which are normally found in other pathways [2]. The core components of Notch signaling pathway include the ligands of Delta and Serrate (known as Jagged in vertebrates), the Notch receptor and the transcription factor of the CBF1/Su(H)/LAG1 (CSL) family [3]. Both Notch and Delta/Serrate (Dl/Ser) proteins contain large extracellular domain composed of epidermal growth factor-like repeats which function to mediate their direct contact. Notch recognizes Dl/Ser presented at the surface of neighboring cells, and binding with the ligands triggers proteolytic cleavage of the Notch protein [4]. As a result, the intracellular domain of Notch (NICD) is released and translocated into the nucleus, whereas NICD forms a transcription activation complex with CSL to drive the transcription of downstream target genes [5]. In the absence of signal input, NICD could not be produced and CSL recruits a variety of co-repressor factors to suppress the expression of Notch targets. Thus, Notch senses the information sent by neighboring cells, transduces the signal and adjusts the cell state accordingly (Figure 1).
Notch signaling is highly conserved across the animal kingdom, and a functional Notch pathway with true homologs of receptors and ligands could be identified in most metazoan species with available genomic information [6,7,8]. Notch signaling plays crucial roles in various developmental events such as cell fate determination, cell cycle progression, cell death and stem cell homeostasis [1,2,3,4,5]. Therefore, it is not surprising that mutations of genes encoding Notch signaling components are involved in various human diseases, including birth defects and malignant tumors [9,10,11,12]. Many aspects of the Notch signaling pathway, such as the signal transduction machinery, the regulatory network, as well as the involvement in human diseases have been elaborately reviewed in previous articles [13,14,15,16,17,18,19,20]. This review will focus on the roles of Notch signaling in different stages and tissues during insect development, coving recent findings in various insect species.
2. A brief history of Notch signaling studies in Drosophila
Over 100 years ago, Thomas H. Morgan chose Drosophila melanogaster, the fruit fly, as a model animal to study the fundamental law of heredity. The famous white eyed mutant fly led to a series of significant discoveries, and ultimately the establishment of the modern theory of the gene [21,22]. Along with the eye color mutants, various mutants affecting body color and wing morphology were isolated and characterized by Morgan and his students [23]. In 1911, Morgan reported the generation of “beaded wings” mutants by radium ray irradiation, and recorded that the marginal vein and wing blade in these mutants were eliminated to various degrees [24]. In 1914, John S. Dexter isolated one mutant strain from the beaded stocks which he named as “Perfect Notched”, and demonstrated that the wing notches in the tips is caused by a dominant sex-linked factor [25]. However, the “Perfect Notched” stock was lost and Dexter was not able to further examine which factor causes the notched wing phenotype. Later on, Morgan and his students identified additional Notch mutants and Calvin B. Bridges mapped the approximate position of Notch in the X chromosome [23]. In 1919, Otto L. Mohr recovered a novel Notch allele, and showed for the first time that deficiency of a small region of the X chromosome likely caused the Notch loss-of-function mutation [26]. In the next seven decades, the number of Notch alleles and related phenotypes continued to grow, while the biochemical nature of Notch protein remained a mystery [27]. Molecular cloning of the Notch locus marked a new stage of Notch signaling research, and the finding that Notch gene encoded a transmembrane receptor inspired the subsequent studies about cell interaction and signal transduction [28,29,30,31]. Thereafter, detailed molecular genetic analyses have defined the core components and the canonical signal transduction cascade of the Notch signaling pathway.
Mutations of Notch pathway components are found to affect a broad spectrum of developmental events such as cell differentiation, tissue pattern formation and stem cell self-renew [32,33,34]. Now there is a common understanding that Notch signaling is crucial for the development of most, if not all, organs in the fly [35,36,37,38]. Three major modes of Notch actions have been summarized, including lateral inhibition, boundary induction and lineage decision [32]. A historical description of the “lateral inhibition” concept appears in study of cuticle patterning in the bloodsucking bug Rhodnius prolixus [39]. In the dorsal abdomen of R. prolixus, bristles arise from little mounds of smooth cuticle termed plaques, and each plaque appears to exert an inhibitory influence to prevent the development of new plaques within a certain radius [39]. Lateral inhibition takes place within a cluster of cells with initially similar fate and potential, whereas Notch signaling amplifies small differences within them. As a result, one cell with low/no Notch activity acquires a specific fate and prevents the surrounding cells from adopting the same developmental route [32]. When occurring between two populations of cells, Notch signaling establishes a boundary to segregate the two cell groups, which is generally utilized to subdivide developmental fields during tissue formation. Through asymmetrical distribution of signaling regulators which leads to differential signaling activity, Notch controls binary cell fate specification between daughter cells in various cell lineages [32]. It has become evident that Notch functions at different stages and in many tissues during fly development, sometimes with distinct modes of action even within one tissue. Benefited from the abundant genetic resources and exquisite genetic tools, systematic in vivo screens have identified large sets of genes that are involved in Notch signaling in different developmental contexts [40,41,42,43,44,45]. Investigation of the molecular mechanisms between Notch and these genes will help to elucidate how Notch signaling fulfills such sophisticated roles.
The diversified developmental functions of Notch signal have been explored in depth in the model insect D. melanogaster. Genes encoding the receptor and ligands of Notch pathway are found in major groups of insects (Figure 2). Recent advances in genomic resources and methodologies have allowed investigations of Notch function among a broad range of insect species, which will be discussed in the following chapters.
3. Notch signaling in insect embryo development
Donald F. Poulson is generally regarded as the pioneer who established the link between Notch and insect embryo development. In the 1930s, Poulson described a unique “neurogenic” phenotype in Notch mutant D. melanogaster embryos, which failed to develop mesodermal and endodermal tissue while produced hypertrophic nervous system [46,47]. Genetic screens looking for similar neurogenic phenotypes led to the identification of Dl, mastermind (mam), and Enhancer of Split (E(spl)), which were later proven to encode core Notch signaling components [48,49]. Further studies revealed that Notch signaling acts during multiple stages to control embryonic nervous system development, including selection of neural progenitor cells (neuroblasts; NBs), control of NBs daughter cells proliferation, neuronal cell fate specification, glia development and axon pathfinding [50]. NBs are selected from a uniform layer of cells called neuroectoderm by the lateral inhibition process [51]. Expression of proneural genes defines stereotypically spaced “proneural clusters” in the neuroectodermal cell sheet, each proneural cluster consists of 6-8 cells with similar potential to develop into NBs [52]. Inside the proneural clusters, the cellular interactions mediated by Notch signal results in the selection of one cell with the lowest Notch activity to become NB. Activation of Notch signaling in cells surrounding NB results in the expression of the E(spl) gene complex encoded transcription factors which directly repress the proneural genes and further prevent these cells from becoming NBs [52]. Thus, in mutants of Notch and many other Notch pathway genes, excessive NBs are formed due to the lack of lateral inhibition inside the proneural cluster [53].
NBs were recognized as distinct cell population with specific characteristics of cell size, cell shape, and nuclear position by histological methods in the embryos of several insect species more than 130 years ago [54]. Similar number and arrangement of NBs were found in the embryos of insects with different developmental modes and life histories, including D. melanogaster, the bloodsucking bug R. prolixus, cockroaches (Blatta germanica and Periplaneta americana), locusts (Locusta migratoria, Melanoplus femurrubrum, Schistocerca americana and Schistocerca gregaria), potato beetle (Doryphora decemlineata), red flour beetle (Tribolium castaneum), stick insect (Carausius morosus), silverfish (Ctenolepisma longicaudata), tobacco hornworm (Manduca sexta) and yellow mealworm (Tenebrio molitor) [54,55,56,57,58,59,60,61,62,63,64,65,66]. These observations indicate that selection of NBs during insect embryonic development might be mediated by conserved mechanisms [67,68,69]. The landmark laser ablation experiments performed in locust (S. americana) embryo demonstrated that lateral inhibition by the enlarging NB ensures development of only one NB in each proneural cluster [59,66]. In cricket (Gryllus bimaculatus) and cockroach (P. americana) embryos, RNAi knock-down of Notch and/or Dl caused classic neurogenic phenotype in early stages and cell apoptosis in later stages [70,71,72]. Knock-down of Notch and E(spl) by RNAi in red flour beetle embryo led to similar neurogenic phenotype with increased number of NBs [73]. Computational analyses suggest that the E(spl) gene family is quite conserved among insects [74,75,76,77,78,79,80]. It is possible that Notch signaling and the E(spl) gene family are generally utilized to determine NB fate in insect embryos [81].
Other than the common role in neurogenesis, Notch signaling also regulates different developmental events in different insects. It appears that Notch signaling is critical for embryo segmentation in several insect species, but is dispensable for this process in other insects [82]. Segmentation is a fundamental process that divides the developing body into separate units which can undergo independent developmental programs [83]. Two distinct modes of embryo segmentation have been reported in insects [84]. In long germ insects such as D. melanogaster, all segments are specified almost simultaneously within the blastoderm before gastrulation takes place. On the other hand, only segments of the head region are specified in the blastoderm in short and intermediate germ insects, whereas the remaining segments arise sequentially from a posterior segment addition zone using a clock and wave front mechanism similar as that in vertebrates [84]. Originally identified from a genetic screen in D. melanogaster, many segmentation factors are found to function in a conserved fashion among insects with different germ band types [82,83,84,85]. In vertebrates and several sequential segmenting arthropods such as brine shrimp, water flea, centipede and spider, Notch signaling plays critical roles in segmentation [82]. It is proposed that Notch signaling represents an ancestral mechanisms controlling segmentation in arthropods and vertebrates [86]. However, whether and how Notch signaling is involved in embryo segmentation in insects is still under debating.
It is well accepted that Notch signal is not required for embryo segmentation in Drosophila. The segment morphology and expression pattern of segmentation factors were not affected in Notch mutant fly embryos, despite that remarkable neurogenesis defects would emerge in later stage [87,88]. In another long germ insect, the honeybee (Apis mellifera), Notch signaling was not involved in segmentation as well [89]. In the short germ hemipteran milkweed bug Oncopeltus fasciatus, Dl was not expressed in the right time and right space to regulate embryo segmentation [90,91]. Importantly, RNAi knock-down of Dl in O. fasciatus embryo did not affect the expression pattern of other segmentation factors [92]. In the short germ red flour beetle, no evidence could support a role of Notch signaling in segmentation [93]. The fringe (fng) gene encodes a conserved modifier of the Notch receptor [94,95], and is required for segmentation in mice and chicken [96,97]. Expression of fng is not detected until after segment boundaries are established in the embryo of the short germ locust S. gregaria, ruling out a role in embryo segmentation [98]. The germ band morphology is highly dynamic in the silkworm Bombyx mori, but the available molecular data suggest that the majority of segments are not patterned prior to the onset of gastrulation, which fits with the short germ type [84]. In B. mori embryos, Notch RNAi caused patterning defects without affecting the formation of segments [99], but Dl RNAi led to loss of posterior segments and disruption of segment boundaries [100]. The cockroach P. americana belongs to short germ type, and Notch RNAi resulted in segment morphology defects along with changes of segmentation factor expression pattern [72,86]. In the intermediate germ cricket G. bimaculatus, Notch signaling was maternally required for morphogenesis of segments and formation of posterior segments in the embryo [70]. However, a subsequent study argued that zygotic Notch signaling was not required for early segment patterning or morphological formation of segment boundaries, and the authors pointed out that apoptosis and neurogenesis defects in early stages might cause secondary defects in segment morphologies [71]. In conclusion, remarkable evolutionary flexibility exists among the insects in terms of dividing segments in the embryo, and there is no corresponding relationship between the germ band type and the involvement of Notch signaling in embryo segmentation.
4. Notch signaling in insect wing development and patterning
Insects are the only group of invertebrates that have wings, which are considered as a key evolutionary innovation that allowed insects to become the most diverse and abundant animal group [101]. The evolutionary origin of insect wings is an important yet highly debated mystery, but the key steps and signaling pathways underlying wing development are quite conserved among winged insects [101,102,103]. Our understanding of how Notch signal regulates insect wing development largely come from studies in D. melanogaster. In the wings of fruit fly, the developmental events regulated by Notch signal include, but are not limited to wing margin formation, wing growth, vein patterning and sensory organ specification [104,105].
The Notch gene is named after the phenotype of “one or more incisions at the end of wings”, and nicked wing margin is arguably the most common and conspicuous defect observed in survivable Notch mutant flies [22]. But it took more than a century to figure out how Notch signal regulates various aspects of fly wing development at the cellular and molecular level [23,104]. As a typical holometabolous insect, the fruit fly undergoes complete metamorphosis which means that the larvae have no resemblance to the adult and the transformation to adult occurs during the pupal stage. The precursors of adult wing are maintained during larval stages as distinct clusters of undifferentiated cells called wing imaginal disc (also known as wing disc) [105]. Despite the significant difference in cell number, cell size, cell identity and tissue morphology between wing disc and adult wing, most of the patterning events take place in the wing discs during the larval stage [104]. In the developing wing disc, cells utilize Notch signaling to establish the boundary between the dorsal and ventral (D/V) compartment. Cells in the two compartments develop into two apposed epithelial sheets in the adult wing blade, while the wing margin is formed by the D/V boundary cells [105]. Notch is activated along both sides of the D/V boundary by two different ligands, Dl activates Notch in dorsal boundary cells and Ser activates Notch in ventral boundary cells [106,107,108]. Glycosylation in the extracellular domain by fng sensitizes Notch towards binding with Dl while inhibits binding with Ser, thus differentiating its affinity and responses from the two ligands [109,110,111,112,113,114,115,116]. The expression of both Ser and fng is under the control of a dorsal-specific transcription factor apterous (ap), and feedback loops among these genes further strengthen the D/V boundary [110,117,118,119,120]. In the D/V boundary cells, Notch signal promotes their proliferation and survival through activating the expression of target genes such as vestigial (vg), wingless (wg) and cut [106,107,121,122,123,124,125,126,127,128,129,130,131]. In addition to the cell autonomous functions, vg and wg also regulate the growth of cells away from the D/V boundary [106,121,128,129]. Mutations impairing Notch signal activity disrupt the segregation of D/V compartment as well as the overall growth in the wing [105].
The fly wing blade consists two main cell types—vein and intervein, and the veins serve as structural supports for the wing blade and as vessels for trachea, nerves and hemolymph [104]. Notch signaling promotes intervein fate and inhibits vein fate so that the boundary between the two types of cells could be established [132,133,134,135]. Mutations dampening Notch signal activity during the larval and pupal stages result in uniformly thickened veins with deltas at the tips, while abnormal activation of Notch signaling can remove adult veins [118,132,133,134,136]. The wing disc contains small number of sensory organ precursor (SOP) cells which will form sensory bristles in the notum and along the anterior edge of the wing margin in adult flies [105]. SOPs are selected through a lateral inhibition process mediated by Notch signaling, after which they undergo stereotyped asymmetric divisions to form the mechanosensory organ and the fates of daughter cells are also regulated by Notch signaling [37,122,137,138,139]. Disruption of Notch activity at different developmental stages could disrupt bristle pattern, bristle number as well as cell fate specification [1,139]. The phenotypes in wing margin, veins and sensory bristles have been used as easy-to-spot and reliable readout for genetic screens that aim to identify Notch signaling modulators [140,141,142].
Notch signaling is required for wing development in several Dipteran insects. In two fly species closely related with D. melanogaster, Drosophila hydei and Drosophila virilism, mutant alleles of Notch and other genes in the pathway led to similar wing margin, vein and bristle defects [143,144,145,146,147,148,149,150]. Mutation stocks displaying nicked wing margins have been isolated in the housefly Musca domestica, which were later mapped as Notch and cut mutants [151,152,153]. With the completion of M. domestica genome sequencing and success of Cas9-mediated genome editing, further molecular genetics analysis will provide insights about the roles of Notch signaling in house fly development [154,155]. Many mutations affecting wing development have been isolated and characterized in the Australian sheep blowfly, Lucilia cuprina [156,157,158]. The Scalloped wings (Scl) loss-of-function mutants displayed wing notching, vein thickening and bristle abnormalities, and the Scl gene has been molecularly identified as the homolog of Notch [159,160].
Lepidopteran insects such as butterflies and moths normally have two pair of wings (forewings and hindwings) covered by microscopic dust-like scales. Although the wing structure and morphology are very different from the flies, a series of works have demonstrated the importance of Notch signaling during B. mori wing development. The flügellos (fl) mutant of the silkworm produce wingless pupae and moths because wing discs cannot respond to ecdysone during metamorphosis. The fl mutant wing discs develop normally until the fourth larval instar and developmental defects emerge in the fifth larval instar and pupae [161,162,163,164]. Molecular mapping and cloning have shown that the gene at the fl locus is actually the homolog of fng [165]. Whole-mount in situ hybridization confirmed that fng is expressed in the dorsal layer, while BmWnt1, the homolog of wg, is expressed along the D/V boundary in wing discs [165,166]. In fl/fng mutant wing discs, the expression of BmWnt1 is diminished [165]. The microRNA mir-2 was found to target fng, and over-expression of mir-2 and somatic mutagenesis of fng using the CRISPR/Cas9 system resulted in similar defects of wing morphology and inhibition of BmWnt1 expression [167]. The selector gene ap was found to be expressed in the dorsal layer of wing disc and vg mutants exhibit small wing phenotype similar as seen in the fly [168]. These findings indicate that B. mori likely employs the same regulatory cascade which includes the compartment selector (ap), Notch signaling and downstream targets (vg and wg) to control D/V boundary formation and wing growth.
Butterflies are well known for their colorful wings with pattern elements of many types and colors. In the best studied model species Precis coenia, the buckeye butterfly, the expression of ap was restricted in dorsal cells while wg was expressed in cells along the future wing margin in fifth instar wing discs [169]. Such expression pattern is comparable to that of fruit fly and silkworm [165,169]. Recent studies have shown that the up-regulation of Notch likely represents an early developmental event in the formation of eyespots and other wing color patterns in multiple butterfly species [170,171,172,173,174,175,176,177]. The expression pattern of Notch protein in the pupal wing of Heliconius erato indicates that Notch mediated lateral inhibition might play a role in organizing butterfly wing scales [178]. However, functional studies are required to demonstrate the exact roles of Notch signaling during butterfly wing development.
The highly specialized forewings called elytra is considered as an important trait driving the successful radiation of Coleoptera (beetles), one of the most successful animal groups on the planet [179]. Compared with the hindwings, elytra in beetles consist thick, hardened, and pigmented cuticles and many morphologically distinct features. Nevertheless, the expression pattern of many key regulatory genes in the elytra and hindwings of T. castaneum was surprisingly similar [180]. The expression of Notch target genes wg and cut were found in D/V boundary cells in both elytra and hindwing discs [180]. Comparative developmental analysis found that the abrupt gene encodes a novel regulator of Notch signaling and plays an essential role for proper wing vein patterning in both the red flour beetle and fruit fly [181]. The beetle E(spl) proteins suppressed the formation of bristles and veins when ectopically expressed in fly wing disc [73]. The red flour beetle has emerged as an important model insect with elaborate genetic toolkit and imaging techniques [182,183]. In the foreseeable future, studies in T. castaneum will bring up new insights about the roles of Notch signaling in different types of insect wings.
A recent study revealed that in the brown planthopper, Nilaparvata lugens, the Notch locus contains a complex alternatively spliced gene encoding a series of protein isoforms with different expression patterns [184]. When dsRNAs targeting different transcripts were injected into the planthopper nymphs, a certain degree of functional differences among Notch variants was observed. Several planthopper Notch variants were involved in bristle, vein and wing blade development [184]. Expression of Notch pathway genes was detected in the wing discs of various insect species, but their functions in wing development need further exploration [185,186].
5. Notch signaling in insect leg development
Notch pathway plays diverse and fundamental roles in D. melanogaster leg patterning and growth [187]. The fruit fly legs are composed of ten segments, and each segment is separated by a flexible joint. Fusion of leg segments and reduction of leg growth have been noticed as common defects in Notch, Dl and Ser mutants [188,189,190]. On the other hand, ectopic activation of Notch within the leg were sufficient to induce formation of extra segment borders (joints) and local cell growth [191,192,193]. The separation between leg segments is determined in the larval stage, at which time Notch, Ser, Dl, and fringe are expressed in a segmentally repeated pattern in the imaginal leg disc [191,192,193,194]. Interestingly, Notch is asymmetrically activated at the distal side of the Ser and Dl-expressing domain, forming nine rings along the Proximal-Distal (P-D) axis in the leg disc [195]. Such polarized activation of Notch is critical for proper leg development, continuously activation of Notch across segments led to shortened legs with segmentation defects [191,192,193]. Numerous factors and pathways were found to interact with Notch signaling to establish the boundaries between leg segments, but the underlying mechanisms are still not fully elucidated [196,197,198,199,200,201,202,203,204,205,206,207,208,209]. Proper activation of Notch pathway is also instructive for the formation of the joints and for the correct growth of the leg [210]. The fly leg joints can be subdivided into two classes, the proximal “true joints” and the distal “tarsal joints”. The morphology of true joints varies from one to another, while the tarsal joints are identical and are composed of a “socket” in the proximal part and an interlocking “ball” in the distal part [195]. Both types of joints are shaped by Notch signaling, while distinct target genes are separately activated in true joints and tarsal joints [187,195,211]. During tarsal joints morphogenesis, Notch signaling is required for the fate specification of socket cells and ball cells, as well as for the extensive cell shape changes and cell movements which are essential for forming the joint structures [212,213]. How Notch controls leg growth are mostly unknown, while some reports indicate that the interaction with Hippo pathway is involved [192,197].
The roles of Notch in leg segmentation, joint formation and leg growth are quite conserved among different insect species. Fusion of the tarsal elements in the leg was found in D. hydei Notch mutants [146]. In the cricket G. bimaculatus, RNAi knock-down of Notch led to reduction of leg length and loss of joints [70]. In the red flour beetle, Notch RNAi eliminated most joints from the leg but the leg length was not affected, while Ser RNAi eliminated nearly all the joints and reduced the overall length of the leg [214,215]. The nubbin (nub) gene was expressed in a series of concentric rings in fly leg disc, and mutation of nub resulted in shortened legs [216,217]. Notch signaling directly regulates the expression of nub in fly leg disc [192,203]. Expression of nub homologs were found in the developing legs in several insect species, including the cockroach P. americana, the milkweed bug O. fasciatus and the primitively wingless firebrat Thermobia domestica [218]. RNAi knock-down of nub in O. fasciatus embryos led to shortened thoracic legs and growth of ectopic appendages on abdominal segments [219]. In the house cricket Acheta domesticus and the cockroach P. americana, nub was required for the growth of specific leg segments and joint formation [220]. The finding that the levels of nub expression were reduced after Notch RNAi indicates that Notch signaling might also regulate nub expression in P. americana [220]. Several signaling pathways, including Notch were upregulated during regenerative patterning and growth in ladybird beetle (Harmonia axyridis) legs [221]. Notch mediated segmentation of the appendages has been proposed as a defining character and molecular phylotypic trait of arthropods, which could be further tested in other insect species [222].
6. Notch signaling in insect reproduction
Notch signaling is essential for ovary development in D. melanogaster, particularly during egg chamber formation and the assembly and maintenance of the ovarian germline stem cell (GSC) niche [33,223,224].
Oogenesis in fruit fly begins in the germarium located at the anterior tip of the ovariole, where GSCs divide asymmetrically to give rise to cystoblasts. Each cystoblast undergoes four rounds of incomplete cell division to produce a germline cyst containing 16 interconnected cells. The germline cyst is then encapsulated by somatically derived follicle cells, and the collection of germline and follicle cells at this point is known as an “egg chamber.” In the egg chamber, only one cell within a germline cyst will develop into the oocyte, while the remaining 15 cells become nurse cells to provide RNAs and proteins for the oocyte. An egg chamber progresses through numerous developmental stages while moving toward the posterior of the ovary before finally becoming a mature egg and entering the oviduct [224]. Using temperature sensitive mutant alleles, mutation of Notch and Dl were found to significantly inhibit the number of eggs laid by female flies, and defects in both follicle cell development and the establishment of anterior-posterior polarity in the oocyte were observed [225]. Subsequent studies demonstrated that Notch activity is required in the follicle cells for their transition from mitosis to endocycling, which is regulated by Dl expressed in the germline cells [226,227,228,229,230,231,232,233,234,235,236,237,238]. The egg chamber possesses intrinsic anterior-posterior (AP) polarity, with the nurse cells being anterior and the oocyte posterior. This stereotypical position plays a key role in generating AP polarity in both the oocyte and the surrounding follicle cells [239]. Interestingly, such AP polarity is established through a relay mechanism which propagates asymmetry from older to younger cysts. During early stage of oogenesis, a germline cyst signals through the Dl-Notch pathway to induce the formation of the anterior polar cells when it buds from the germarium. The anterior polar cells in turn express the JAK/STAT signaling ligand Unpaired and signal toward the adjacent anterior stalk/polar cell precursors to induce the stalk cell fate. The stalk induces the positioning of the oocyte at the posterior pole in the adjacent younger cyst by adhesive interactions. As the oocyte is being positioned, a new round of Dl-Notch signaling takes place in that cyst to induce the anterior polar cells [240,241]. Thus, each cyst polarizes the next cyst through a series of posterior to anterior induction events [242,243].
The concept of stem cell niche was proposed around half a century ago to describe the tissue microenvironment capable of supporting the self-renew and maturation of haemopoietic stem cells, which was later extended to other tissues that contain stem cells. However, the ability of the niche to support stemness was firstly demonstrated by studies of fly ovarian GSC niche [244]. At the anterior tip of the germarium, the terminal filament cells and the cap cells form the niche which supports two to three ovarian GSCs. The formation of both terminal filament cells and cap cells rely on proper Notch signaling [244,245,246,247,248,249,250,251,252,253]. Importantly, the strength of Notch signaling controls the size of the ovarian GSC niche. Hyperactivation of Notch signaling leads to the formation of more cap cells and bigger niches, which support more GSCs. Whereas dampened Notch signaling results in decreased cap cell number and niche size, and consequently less GSCs [246]. In mature adult flies, Notch signaling also transmits the effects of diet and age on GSC niche activity and GSC maintenance [254,255,256,257,258].
Two major types of ovarioles, panoistic ovarioles and meroistic ovarioles, are found in insects. In panoistic ovarioles, all progenies of germline stem cell become functional oocytes and no nurse cells are formed. Meroistic ovarioles contain nurse cells and can be further classified into polytrophic ovarioles and telotrophic ovarioles by morphological characters. In polytrophic ovarioles, nurse cells are found in the egg chamber and transport mRNA and proteins to oocyte through ring canals. While in teleotrophic ovarioles, nurse cells are located in the germarium and are connected to oocytes in early stages by nutritive cords in a distance [259]. Although it is generally believed that the panoistic ovaries represent the ancestral type from which the meroistic types had derived, there is no exact correspondence between ovariole type and phylogenetic position [260]. In the panoistic ovariole of the cockroach Blattella germanica, inhibition of Notch signaling activity caused defects in stalk formation, follicle cell proliferation and follicle cell differentiation [261,262,263,264]. In the telotrophic ovariole of T. castaneum, Notch signaling is required for stalk formation, follicle cell proliferation and AP axis establishment [265,266]. Interestingly, the role of Notch signaling in follicle cells differ significantly in distinct insect species. In D. melanogaster, Notch promotes the switch from mitosis to the endocycle in follicle cells, whereas in B. germanica and T. castaneum Notch signaling is required for maintaining the mitotic cycle of follicle cells. The relationship between the role of Notch in follicle cells and ovariole structure remains an open question. Notch was found to regulate vitellogenesis in the fat body of L. migratoria, which is crucial for oocyte maturation and ovarian growth [267]. Vitellogenesis is generally required for insect oogenesis and egg production, whether Notch signaling is involved in this basic physiological event in other insects is worth for further examinations [268]. In the honeybee A. mellifera, oogenesis in the germarium of worker bees were repressed by Notch signaling [269,270]. In the honeybee queens, Notch pathway genes were dynamically expressed in the ovariole at different oviposition phases, but their functions were not examined [271,272]. Highly abundant Notch-like proteins were detected during the initial stages of ovary development in Bactrocera dorsalis, indicating a potential role in oriental fruit fly oogenesis [273]. Taken together, Notch signaling is likely generally required for oogenesis in insect species, but we have just scratched the surfaces on how Notch executes specific roles in distinct cell types, developmental stages and insect species during ovary development and reproduction.
7. Notch signaling in insect stress responses
Insects encounter numerous stresses in the real world, including but not limited to temperature fluctuation, nutrition deficiency, pathogen infection and toxin exposure. Recent studies have discovered that Notch signaling is involved in stress responses under several conditions.
Nutrition is one of the most important environmental variables impacting insect life cycles, and insects adjust their behavior, physiological activities and metabolic programs in respond to changes of food quality and quantity [274]. The insulin pathway functions as nutritional sensor to coordinate the metabolic requirement and other biological events [268]. In female insects, reproduction requires a massive input of nutrition resources to produce eggs with a large amount of nutrient reserves [275]. Insulin signal pathway relays the effect of diet on reproduction via regulation of Notch signaling in D. melanogaster ovary [254,255,256,258,276]. In somatic tissues such as gut, muscle and neuronal system, insulin signaling affects Notch activity by various means [277,278,279]. A recent study found that dietary cholesterol affects the level and duration of Notch signaling through modulating Dl and Notch stability and trafficking, which further influences cell differentiation in fly adult midgut and alters the metabolic program [280]. The expression of Notch pathway genes was upregulated when high quality diet was provided for honeybee A. mellifera, but the underlying mechanisms and the functional importance is still unknown [281]. In the larval guts of Asian honey bee, Apis cerana, the Notch pathway appeared to be targeted by miRNAs and piRNAs at different developmental stages [282,283]. The lifespan of fruit fly adults was extended by supplementation of pterostilbene, fucoxanthin and a traditional Chinse herb Cistanche tubulosa, and the expression of Notch pathway genes was increased upon ingestion of these substances [284,285,286]. These studies highlight the role of Notch signaling during response to nutrition status in insects.
Many viruses are dangerous pathogens for human beings and could be transmitted by vector insects such as mosquitoes [287]. After infection by the alphavirus Sindbis, expression of Notch pathway genes was up-regulated in S2 cells, leading to the presumption that Notch signaling may be involved in establishment of virus persistence in insect cells [288]. Induction of Notch pathway genes was also observed upon Dengue virus infection in Aedes albopictus cells [289]. Infection of Dengue virus and Chikungunya virus resulted in up-regulation of Notch pathway genes and midgut cell division in the vector mosquito Aedes aegypti [290,291,292]. Knocking-down the expression of Dl by RNAi inhibited midgut cell division induced by infection, but significantly increased the susceptibility of refractory Aedes aegypti strain to Dengue virus [293]. The human malaria parasite Plasmodium berghei also induced midgut cell division and Notch pathway activation in the vector mosquito Anopheles albimanus [294,295]. In the midguts of Anopheles gambiae, Plasmodium falciparum infection induced chromatin changes in regulatory elements of Notch pathway genes, but the significance of these observations needs to be further explored [296]. When wild silk moth Antheraea yamamai was infected by the pathogenic nucleopolyhedrovirus, expression of several Notch pathway genes was up-regulated in the larval midgut [297]. Inhibition of Notch signaling was associated with midgut development defects in locust L. migratoria and the yellow fever mosquito Aedes aegypti [298,299]. In fruit fly midgut, Notch signaling promotes asymmetric division in the intestine stem cells to control tissue homeostasis and responses to various stimulations [300,301,302,303,304]. Whether virus and pathogen infection would trigger Notch related responses in intestine stem cells could be further tested in other insects.
Notch plays a central role in hematopoietic cell fate determination and maintenance in larvae lymph gland, a vital organ of the immune defense system in D. melanogaster [35,305,306,307,308]. Fungal infection and wasp parasitization led to downregulation of Notch signaling and triggered specific immune responses in fruit fly [308,309]. Stimulation by Gram-negative bacteria led to up-regulation of Notch pathway genes in honey bee workers [310]. Several lncRNAs were found to control immune priming in T. castaneum likely by regulating expression of Notch pathway genes [311]. These studies indicate that Notch signaling might be involved in certain immune responses upon pathogen infection in insects.
Using the developing wing as a model system, the anthrax toxins and cholera toxins were found to inhibit endocytic trafficking of Notch signaling components and impair Notch activity in D. melanogaster [312,313]. In a Zika virus infection model, the virus non-structural protein NS4A was shown to restrict fly eye growth through regulation of JAK/STAT signaling and inhibit wing growth via affecting Notch activity [314]. The heavy metal mercury led to neurogenesis defects in fly embryo and marginal nicks in the wing strongly through inhibiting Notch cleavage and NICD production [315]. Treatment with methylmercury, an organic form of mercury easily absorbed by the intestinal trac and an environmental pollutant, led to increased levels of Notch signaling in fly cells and embryos [316,317,318,319]. The response of Notch target genes to mercury showed variations in different cell types, upon treatment of organic or inorganic forms, and was sometimes independent of the Notch receptor [316,317,318,319]. Pesticide exposure also affects Notch signaling in several insects. Feeding fly larvae with sublethal level of chlorfenapyr resulted in wing and leg developmental defects and disruption of Notch signal activity [320]. Exposure to four different adverse environmental factors, including low dose of gamma-irradiation, formaldehyde, toluene and dioxin, commonly affected Notch signaling in adult flies [321]. Harmine, a natural β-carboline, inhibits Drosophila development by negatively affecting the developmental signaling network including Notch signal pathway [322]. Fipronil exposure decreases the expression and activity of Notch in the brain and Malpighian tubules of the stingless bee Partamona helleri [323,324]. Sublethal level of fluralaner impaired larval development of the common cutworm Spodoptera litura and led to wing notches in the surviving adults, but whether Notch signaling was affected is not examined [325]. The Notch pathway was responsive to ultraviolet irradiation and metamorphosis oxidative stress in B. mori [326]. Together, these results suggest that Notch signaling may be involved in response to hazardous factors in insects.
Insects would go through functional hypoxia when oxygen supply is insufficient for metabolism demand, and respond to hypoxia via multiple strategies [327]. Hypoxia tolerance is regulated by Notch signaling in D. melanogaster, flies with impaired Notch activity had a significant reduction in hypoxia tolerance, but those with hyperactivated Notch signaling had a dramatic opposite effect [328,329,330,331,332]. It would be exciting to examine whether such strong effects of Notch signaling in hypoxia tolerance is also present in other insects, especially those experience environmental hypoxia at certain stages in their life history.
8. Notch signaling in less studied but more interesting tissues
Insects have evolved to acquire many fascinating novel structures while keeping a steady basic body plan, and the mechanisms underlying the evolution of morphological novelties remain enigmatic [333]. From a developmental biology point of view, as one of the fundamental regulator units of tissue development and growth, the involvement of Notch signaling in construction of such morphological novelties is actually not surprising at all.
The bull-headed dung beetle Onthophagus taurus and many other scarab beetle species possess rigid projections of the exoskeleton referred to as horns, which is a highly diversified and evolutionarily novel morphological structure evolved from wing serial homologs [334]. Notch pathway genes were expressed in the developing dung beetle horn [335,336]. Importantly, Notch signaling was identified as a regulator of the dramatic diversity of horn sizes and shapes across dung beetle males [337]. In the Asian rhinoceros beetle Trypoxylus dichotomus, Notch RNAi disturbed horn primordial furrow depth which led to defects in the horn shape in the pupae [338].
The dorsomedial and the abdominal support structurea are two types of body wall projections commonly observed in the pupae of scarab beetles. These two structures were identified as wing serial homologues in Onthophagus taurus, and Ser RNAi disrupted the formation of both structures [339]. Gin-trap is as structure found exclusively on pupae of the closely related beetle families Tenebrionidae and Colydiidae, which is believed to be evolved after the radiation of holometabolous insects. Gin-traps function as defensive organs to grasp the appendages of predators in the pupae. In the beetle T. castaneum, RNAi knock-down of Notch pathway components disrupted the formation of gin-traps [340].
In dung beetles, the fore tibia has become a specialized digging tool which may have allowed them to access the compacted soil as habitat. The dung beetle fore tibia is a flattened and enlarged segment characterized by four to five prominent tibial teeth which could enhance the digging performance. RNAi of Ser and downstream genes of Notch pathway resulted in fusion of all leg segments accompanied by a reduction of tibial teeth in Onthophagus taurus [341]. These findings indicate that Notch signaling is repeatedly recruited to regulate the development of evolutionarily novel morphological structures in beetles.
As the major olfactory sensory organ, antennae are crucial for insects to locate food resources, mating partners and oviposition sites, as well as to evade predators and toxic substances. The insect antennae are highly diversified with astonishing variations in shapes, structures and sizes [342]. In D. melanogaster, the fate of antenna cells is determined by several selector genes, while Notch signaling regulates cell proliferation, tissue growth and formation of boundary between antenna segments [192,203,343,344,345]. In the beetle T. castaneum, Ser RNAi resulted in strong reduction in antenna length and complete absence of joints, while Notch RNAi led to the absence of most antennal joints without significant reduction of antenna growth [346]. However, Notch RNAi, but not Ser RNAi decreased the density of sensory bristles on the antenna [346]. The expression pattern of Notch and Dl in the developing antenna in the cricket G. bimaculatus was suggestive for a role in segmentation [70].The nub gene is a down-stream target of Notch signaling during leg development in D. melanogaster and P. americana [220]. In the milkweed bug O. fasciatus, nub RNAi resulted in deficient sensory bristles formation without significant affecting segmentation and growth [219]. Depletion of nub by RNAi in Acheta domesticus and P. americana led to fusion of segments in antenna [220]. The Expression of Notch target gene E(spl)mβ was detected in specific segments in B. mori larval antennal primordium, which develops as the feather like antenna in adults [347]. Upon Notch RNAi, the B. mori antenna segments were drastically fused and the overall length along the PD axis was extensively reduced, and milder defects such as fusion of neighboring lateral branches were also observed [348]. These findings support a general requirement of Notch signaling in insects antenna growth and segmentation.
Insects show a remarkable diversity of mouthpart morphology, and the genetic regulatory network for mouthpart development is not completely understood. RNAi knock-down of Dl in the silkworm and honeybee caused mild alterations in embryonic labrum shape [89,100]. Knockdown of two components of the Notch pathway, Ser and mib1, led to loss of the labrum in T. castaneum larvae likely due to defects in cell proliferation [215]. RNAi of Notch and Dl also disrupted sensory organ development in the T. castaneum labrum [349]. A role of Notch signaling during mouthpart construction in the fruit fly and other insects is expected but has not been reported yet.
The diverse color patterns of insects often function as camouflage to protect them from predators [350]. In the Asian swallowtail butterfly, Papilio xuthus, young larvae from the first to the fourth instar show black and brown patterns to mimic bird droppings but change their color pattern to mimic host plants in the final instar. Such color pattern switch in Papilio xuthus caterpillar was initiated at the early fourth instar by the juvenile hormone, and both Dl and E(spl)mβ were found to be specifically expressed in the epidermis of certain color marking region during the pattern switch period [351,352]. Functional analysis demonstrated that Notch signaling defines the edge and pigmentation area in the final color patterns [353]. RNAi knock-down of Notch and Dl led to expansion of pigmentation area and disruption of border lines in the fifth instar larvae. RNAi knock-down of Dl in Papilio machaon, a species closely related to Papilio xuthus, led to similar, albeit more subtle phenotypic changes of larvae color pattern. Interestingly, in the silkworm L mutant, which shows pairs of black brown twin spots on each larva body segment, knockdown of Notch but not Dl, Ser or fringe caused pigmentation loss in the twin spots [353]. These findings demonstrated the importance of Notch signaling in the adaptive evolution of camouflage formation in caterpillars, which encourages further examination of the roles of Notch signaling during color pattern formation in other insect species.
9. Conclusions
It has been more than one century since the first Notch mutant was isolated in the fruit fly, which has been used as a leading model system to investigate the developmental functions of Notch signaling. Comprehensive studies in this little inconspicuous insect has led to impressive progresses in Notch signaling mechanisms. The crucial components, signal transduction cascade and the major modes of action of Notch pathway are likely highly conserved among insect species, and the involvement of Notch signaling during the development of various insect tissues have been validated.
Nevertheless, in distinct insect species there are too many fascinating developmental events which are absent in the fruit fly [58,84,342,350,354,355]. Recent advances in genome editing and other functional genetics technologies have made it possible to identify novel genetic factors and evaluate the contribution of Notch signaling pathway underlying diverse developmental phenomena in ‘non-model’ insect species (Figure 3). Future studies will undoubtedly help us to better understand the extensive roles of Notch in the formation and evolution of insect tissues, and may also reveal novel regulators, functions and signaling mechanisms. On the other hand, these knowledges could be readily converted to design genetic control methods such as RNA pesticide [356], genetic sexing [357] and gene drive system [358] to protect crops and humans from insect pests.
Author Contributions
Conceptualization, T-C.Y., J.S. and J.Z.; formal analysis, Y.C.; resources, Y.C.; data curation, H.L.; writing—original draft preparation, Y.C. and H.L.; writing—review and editing, T-C.Y., J.S. and J.Z.; supervision, T-C.Y. and J.S.; project administration, J.Z.; funding acquisition, J.S. and J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31970478 to J.Z; 32030012 to J.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Dr. Yonggang Hu for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bray, S.J. Notch signalling in context. Nat Rev Mol Cell Biol 2016, 17, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C. Notch signaling: control of cell communication and cell fate. Development 2004, 131, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Kovall, R.A.; Gebelein, B.; Sprinzak, D.; Kopan, R. The Canonical Notch Signaling Pathway: Structural and Biochemical Insights into Shape, Sugar, and Force. Dev Cell 2017, 41, 228–241. [Google Scholar] [CrossRef]
- Sachan, N.; Sharma, V.; Mutsuddi, M.; Mukherjee, A. Notch signaling: Multifaceted role in development and disease. FEBS J 2023. [Google Scholar] [CrossRef]
- Henrique, D.; Schweisguth, F. Mechanisms of Notch signaling: a simple logic deployed in time and space. Development 2019, 146. [Google Scholar] [CrossRef]
- Gazave, E.; Lapebie, P.; Richards, G.S.; Brunet, F.; Ereskovsky, A.V.; Degnan, B.M.; Borchiellini, C.; Vervoort, M.; Renard, E. Origin and evolution of the Notch signalling pathway: an overview from eukaryotic genomes. BMC Evol Biol 2009, 9, 249. [Google Scholar] [CrossRef]
- Babonis, L.S.; Martindale, M.Q. Phylogenetic evidence for the modular evolution of metazoan signalling pathways. Philos Trans R Soc Lond B Biol Sci 2017, 372. [Google Scholar] [CrossRef]
- Vlachakis, D.; Papageorgiou, L.; Papadaki, A.; Georga, M.; Kossida, S.; Eliopoulos, E. An updated evolutionary study of the Notch family reveals a new ancient origin and novel invariable motifs as potential pharmacological targets. PeerJ 2020, 8, e10334. [Google Scholar] [CrossRef]
- Chimento, A.; D'Amico, M.; Pezzi, V.; De Amicis, F. Notch Signaling in Breast Tumor Microenvironment as Mediator of Drug Resistance. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- D'Assoro, A.B.; Leon-Ferre, R.; Braune, E.B.; Lendahl, U. Roles of Notch Signaling in the Tumor Microenvironment. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Ntziachristos, P.; Lim, J.S.; Sage, J.; Aifantis, I. From fly wings to targeted cancer therapies: a centennial for notch signaling. Cancer Cell 2014, 25, 318–334. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: architecture, disease, and therapeutics. Signal Transduct Target Ther 2022, 7, 95. [Google Scholar] [CrossRef]
- Gao, J.; Fan, L.; Zhao, L.; Su, Y. The interaction of Notch and Wnt signaling pathways in vertebrate regeneration. Cell Regen 2021, 10, 11. [Google Scholar] [CrossRef]
- Papagiannouli, F. Endocytosis at the Crossroad of Polarity and Signaling Regulation: Learning from Drosophila melanogaster and Beyond. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Kiesel, V.A.; Stan, S.D. Modulation of Notch Signaling Pathway by Bioactive Dietary Agents. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, S.; Muskavitch, M.A. Notch: the past, the present, and the future. Curr Top Dev Biol 2010, 92, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Shim, Y.S.; Lee, H.S.; Hwang, J.S. Aberrant Notch Signaling Pathway as a Potential Mechanism of Central Precocious Puberty. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Shen, W.; Huang, J.; Wang, Y. Biological Significance of NOTCH Signaling Strength. Front Cell Dev Biol 2021, 9, 652273. [Google Scholar] [CrossRef]
- Sprinzak, D.; Blacklow, S.C. Biophysics of Notch Signaling. Annu Rev Biophys 2021, 50, 157–189. [Google Scholar] [CrossRef] [PubMed]
- Ballhause, T.M.; Jiang, S.; Baranowsky, A.; Brandt, S.; Mertens, P.R.; Frosch, K.H.; Yorgan, T.; Keller, J. Relevance of Notch Signaling for Bone Metabolism and Regeneration. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. Sex limited inheritance in Drosophila. Science 1910, 32, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. The theory of the gene. Am Nat 1917, 51, 513–544. [Google Scholar] [CrossRef]
- Morgan, T.H.; Bridges, C.B. Sex-linked inheritance in Drosophila; Carnegie Institution of Washington: Washington, 1916; pp. 87, 81 p. incl. illus., tables. [Google Scholar]
- Morgan, T.H. The Origin of Nine Wing Mutations in Drosophila. Science 1911, 33, 496–499. [Google Scholar] [CrossRef]
- Dexter, J.S. The analysis of a case of continuous variation in Drosophila by a study of its linkage relations. Am Nat 1914, 48, 712–758. [Google Scholar] [CrossRef]
- Mohr, O.L. Character Changes Caused by Mutation of an Entire Region of a Chromosome in Drosophila. Genetics 1919, 4, 275–282. [Google Scholar] [CrossRef]
- Guruharsha, K.G.; Kankel, M.W.; Artavanis-Tsakonas, S. The Notch signalling system: recent insights into the complexity of a conserved pathway. Nat Rev Genet 2012, 13, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, S.; Muskavitch, M.A.; Yedvobnick, B. Molecular cloning of Notch, a locus affecting neurogenesis in Drosophila melanogaster. Proc Natl Acad Sci U S A 1983, 80, 1977–1981. [Google Scholar] [CrossRef]
- Kidd, S.; Lockett, T.J.; Young, M.W. The Notch locus of Drosophila melanogaster. Cell 1983, 34, 421–433. [Google Scholar] [CrossRef]
- Wharton, K.A.; Johansen, K.M.; Xu, T.; Artavanistsakonas, S. Nucleotide-Sequence from the Neurogenic Locus Notch Implies a Gene-Product That Shares Homology with Proteins Containing Egf-Like Repeats. Cell 1985, 43, 567–581. [Google Scholar] [CrossRef]
- Kidd, S.; Kelley, M.R.; Young, M.W. Sequence of the Notch Locus of Drosophila-Melanogaster - Relationship of the Encoded Protein to Mammalian Clotting and Growth-Factors. Mol Cell Biol 1986, 6, 3094–3108. [Google Scholar] [CrossRef]
- Bray, S.J. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol 2006, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Zamfirescu, A.M.; Yatsenko, A.S.; Shcherbata, H.R. Notch signaling sculpts the stem cell niche. Front Cell Dev Biol 2022, 10, 1027222. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.L.; Heckscher, E.S. Development of motor circuits: From neuronal stem cells and neuronal diversity to motor circuit assembly. Curr Top Dev Biol 2021, 142, 409–442. [Google Scholar] [CrossRef]
- Lan, W.; Liu, S.; Zhao, L.; Su, Y. Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Jiang, H.; Edgar, B.A. Intestinal stem cells in the adult Drosophila midgut. Exp Cell Res 2011, 317, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Barad, O.; Hornstein, E.; Barkai, N. Robust selection of sensory organ precursors by the Notch-Delta pathway. Curr Opin Cell Biol 2011, 23, 663–667. [Google Scholar] [CrossRef]
- Udolph, G. Notch signaling and the generation of cell diversity in Drosophila neuroblast lineages. Adv Exp Med Biol 2012, 727, 47–60. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. Local and general factors in the development of "pattern" in Rhodnius prolixus (hemiptera). J Exp Biol 1940, 17, 180–U189. [Google Scholar] [CrossRef]
- Mummery-Widmer, J.L.; Yamazaki, M.; Stoeger, T.; Novatchkova, M.; Bhalerao, S.; Chen, D.; Dietzl, G.; Dickson, B.J.; Knoblich, J.A. Genome-wide analysis of Notch signalling in Drosophila by transgenic RNAi. Nature 2009, 458, 987–992. [Google Scholar] [CrossRef]
- Saj, A.; Arziman, Z.; Stempfle, D.; van Belle, W.; Sauder, U.; Horn, T.; Durrenberger, M.; Paro, R.; Boutros, M.; Merdes, G. A combined ex vivo and in vivo RNAi screen for notch regulators in Drosophila reveals an extensive notch interaction network. Dev Cell 2010, 18, 862–876. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, M.; Su, Y.; Du, J.; Zhu, A.J. A targeted in vivo RNAi screen reveals deubiquitinases as new regulators of Notch signaling. G3 (Bethesda) 2012, 2, 1563–1575. [Google Scholar] [CrossRef] [PubMed]
- Mo, D.; Shen, J.; Zhang, J. Use of FLP/FRT System to Screen for Notch Signaling Regulators in the Drosophila Wing. Methods Mol Biol 2022, 2472, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Mo, D.; Li, Y.; Liu, T.; Yin, H.; Jiang, N.; Zhang, J. A genetic mosaic screen identifies genes modulating Notch signaling in Drosophila. PLoS One 2018, 13, e0203781. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Sachan, N.; Mutsuddi, M.; Mukherjee, A. Somatic Clonal Analyses Using FLP/FRT and MARCM System to Understand Notch Signaling Mechanism and Its Regulation. Methods Mol Biol 2022, 2472, 83–94. [Google Scholar] [CrossRef]
- Poulson, D.F. Chromosomal Deficiencies and the Embryonic Development of Drosophila Melanogaster. Proc Natl Acad Sci U S A 1937, 23, 133–137. [Google Scholar] [CrossRef]
- Poulson, D.F. The effects of certain X-chromosome deficiencies on the embryonic development of drosophila melanogaster. J. Exp. Zool. 1940, 83, 271–325. [Google Scholar] [CrossRef]
- Lehmann, R.; Dietrich, U.; Jimenez, F.; Campos-Ortega, J.A. Mutations of early neurogenesis in Drosophila. Wilehm Roux Arch Dev Biol 1981, 190, 226–229. [Google Scholar] [CrossRef]
- Lehmann, R.; Jimenez, F.; Dietrich, U.; Campos-Ortega, J.A. On the phenotype and development of mutants of early neurogenesis in Drosophila melanogaster. Wilehm Roux Arch Dev Biol 1983, 192, 62–74. [Google Scholar] [CrossRef]
- Bahrampour, S.; Thor, S. The Five Faces of Notch Signalling During Drosophila melanogaster Embryonic CNS Development. Adv Exp Med Biol 2020, 1218, 39–58. [Google Scholar] [CrossRef]
- Skeath, J.B.; Thor, S. Genetic control of Drosophila nerve cord development. Curr Opin Neurobiol 2003, 13, 8–15. [Google Scholar] [CrossRef]
- Crews, S.T. Drosophila Embryonic CNS Development: Neurogenesis, Gliogenesis, Cell Fate, and Differentiation. Genetics 2019, 213, 1111–1144. [Google Scholar] [CrossRef] [PubMed]
- Hartenstein, V.; Wodarz, A. Initial neurogenesis in Drosophila. Wiley Interdiscip Rev Dev Biol 2013, 2, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, W.M. Neuroblasts in the Arthropod embryo. J. Morphol. 1891, 4, 337–343. [Google Scholar] [CrossRef]
- Truman, J.W.; Ball, E.E. Patterns of embryonic neurogenesis in a primitive wingless insect, the silverfish, Ctenolepisma longicaudata: comparison with those seen in flying insects. Dev Genes Evol 1998, 208, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.B.; Bastiani, M.J.; Bate, M.; Goodman, C.S. From grasshopper to Drosophila: a common plan for neuronal development. Nature 1984, 310, 203–207. [Google Scholar] [CrossRef]
- Bate, C.M. Embryogenesis of an insect nervous system. I. A map of the thoracic and abdominal neuroblasts in Locusta migratoria. J Embryol Exp Morphol 1976, 35, 107–123. [Google Scholar]
- Biffar, L.; Stollewerk, A. Conservation and evolutionary modifications of neuroblast expression patterns in insects. Dev Biol 2014, 388, 103–116. [Google Scholar] [CrossRef]
- Doe, C.Q.; Goodman, C.S. Early events in insect neurogenesis. II. The role of cell interactions and cell lineage in the determination of neuronal precursor cells. Dev Biol 1985, 111, 206–219. [Google Scholar] [CrossRef]
- Doe, C.Q.; Goodman, C.S. Early events in insect neurogenesis. I. Development and segmental differences in the pattern of neuronal precursor cells. Dev Biol 1985, 111, 193–205. [Google Scholar] [CrossRef]
- Hartenstein, V.; Campos-Ortega, J.A. Early neurogenesis in wild-type Drosophila melanogaster. Wilehm Roux Arch Dev Biol 1984, 193, 308–325. [Google Scholar] [CrossRef]
- Kuwada, J.Y.; Goodman, C.S. Neuronal determination during embryonic development of the grasshopper nervous system. Dev Biol 1985, 110, 114–126. [Google Scholar] [CrossRef]
- Urbach, R.; Technau, G.M. Early steps in building the insect brain: neuroblast formation and segmental patterning in the developing brain of different insect species. Arthropod Struct Dev 2003, 32, 103–123. [Google Scholar] [CrossRef]
- Urbach, R.; Technau, G.M.; Breidbach, O. Spatial and temporal pattern of neuroblasts, proliferation, and Engrailed expression during early brain development in Tenebrio molitor L. (Coleoptera). Arthropod Struct Dev 2003, 32, 125–140. [Google Scholar] [CrossRef]
- Nazar, A.P.; Delgado, M.J.; Lavore, A. Empty-spiracles is maternally expressed and essential for neurodevelopment and early embryo determination in Rhodnius prolixus. Dev Biol 2022, 490, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Taghert, P.H.; Doe, C.Q.; Goodman, C.S. Cell determination and regulation during development of neuroblasts and neurones in grasshopper embryo. Nature 1984, 307, 163–165. [Google Scholar] [CrossRef]
- Pop, S.; Chen, C.L.; Sproston, C.J.; Kondo, S.; Ramdya, P.; Williams, D.W. Extensive and diverse patterns of cell death sculpt neural networks in insects. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Boyan, G.; Williams, L. Embryonic development of the insect central complex: insights from lineages in the grasshopper and Drosophila. Arthropod Struct Dev 2011, 40, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Stollewerk, A. A flexible genetic toolkit for arthropod neurogenesis. Philos Trans R Soc Lond B Biol Sci 2016, 371, 20150044. [Google Scholar] [CrossRef]
- Mito, T.; Shinmyo, Y.; Kurita, K.; Nakamura, T.; Ohuchi, H.; Noji, S. Ancestral functions of Delta/Notch signaling in the formation of body and leg segments in the cricket Gryllus bimaculatus. Development 2011, 138, 3823–3833. [Google Scholar] [CrossRef]
- Kainz, F.; Ewen-Campen, B.; Akam, M.; Extavour, C.G. Notch/Delta signalling is not required for segment generation in the basally branching insect Gryllus bimaculatus. Development 2011, 138, 5015–5026. [Google Scholar] [CrossRef]
- Pueyo, J.I.; Lanfear, R.; Couso, J.P. Ancestral Notch-mediated segmentation revealed in the cockroach Periplaneta americana. Proc Natl Acad Sci U S A 2008, 105, 16614–16619. [Google Scholar] [CrossRef] [PubMed]
- Kux, K.; Kiparaki, M.; Delidakis, C. The two Tribolium E(spl) genes show evolutionarily conserved expression and function during embryonic neurogenesis. Mech Dev 2013, 130, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, R.; Maier, D. The Enhancer of split and Achaete-Scute complexes of Drosophilids derived from simple ur-complexes preserved in mosquito and honeybee. BMC Evol Biol 2005, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Maeder, M.L.; Polansky, B.J.; Robson, B.E.; Eastman, D.A. Phylogenetic footprinting analysis in the upstream regulatory regions of the Drosophila enhancer of split genes. Genetics 2007, 177, 1377–1394. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.J.; Dearden, P.K. Evolution of a genomic regulatory domain: the role of gene co-option and gene duplication in the Enhancer of split complex. Genome Res 2010, 20, 917–928. [Google Scholar] [CrossRef]
- Baker, R.H.; Kuehl, J.V.; Wilkinson, G.S. The Enhancer of split complex arose prior to the diversification of schizophoran flies and is strongly conserved between Drosophila and stalk-eyed flies (Diopsidae). BMC Evol Biol 2011, 11, 354. [Google Scholar] [CrossRef]
- Delidakis, C.; Monastirioti, M.; Magadi, S.S. E(spl): genetic, developmental, and evolutionary aspects of a group of invertebrate Hes proteins with close ties to Notch signaling. Curr Top Dev Biol 2014, 110, 217–262. [Google Scholar] [CrossRef]
- Liu, M.; Wang, C.; Li, D.; Liu, Y.; Sheng, Q.; Lv, Z.; Yu, W.; Wang, D.; Zhang, Y.; Nie, Z. Cloning and expression characteristics of the notch-associated gene BmE(spl)mgamma from silkworm, Bombyx mori. Appl Biochem Biotechnol 2014, 173, 2065–2075. [Google Scholar] [CrossRef]
- Dearden, P.K. Origin and evolution of the enhancer of split complex. BMC Genomics 2015, 16, 712. [Google Scholar] [CrossRef]
- Honda, H.; Tanemura, M.; Yoshida, A. Estimation of neuroblast numbers in insect neurogenesis using the lateral inhibition hypothesis of cell differentiation. Development 1990, 110, 1349–1352. [Google Scholar] [CrossRef]
- Clark, E.; Peel, A.D.; Akam, M. Arthropod segmentation. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.K.; Oates, A.C. Delta-Notch signalling in segmentation. Arthropod Struct Dev 2017, 46, 429–447. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.K.; Patel, N.H. Short, long, and beyond: molecular and embryological approaches to insect segmentation. Annu Rev Entomol 2002, 47, 669–699. [Google Scholar] [CrossRef]
- Nusslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Chesebro, J.E.; Pueyo, J.I.; Couso, J.P. Interplay between a Wnt-dependent organiser and the Notch segmentation clock regulates posterior development in Periplaneta americana. Biol Open 2013, 2, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Menne, T.V.; Klambt, C. The formation of commissures in the Drosophila CNS depends on the midline cells and on the Notch gene. Development 1994, 120, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Nusse, R. wingless signaling in the Drosophila eye and embryonic epidermis. Development 1996, 122, 2801–2812. [Google Scholar] [CrossRef]
- Wilson, M.J.; McKelvey, B.H.; van der Heide, S.; Dearden, P.K. Notch signaling does not regulate segmentation in the honeybee, Apis mellifera. Dev Genes Evol 2010, 220, 179–190. [Google Scholar] [CrossRef]
- Auman, T.; Vreede, B.M.I.; Weiss, A.; Hester, S.D.; Williams, T.A.; Nagy, L.M.; Chipman, A.D. Dynamics of growth zone patterning in the milkweed bug Oncopeltus fasciatus. Development 2017, 144, 1896–1905. [Google Scholar] [CrossRef]
- Auman, T.; Chipman, A.D. Growth zone segmentation in the milkweed bug Oncopeltus fasciatus sheds light on the evolution of insect segmentation. BMC Evol Biol 2018, 18, 178. [Google Scholar] [CrossRef]
- Stahi, R.; Chipman, A.D. Blastoderm segmentation in Oncopeltus fasciatus and the evolution of insect segmentation mechanisms. Proc Biol Sci 2016, 283. [Google Scholar] [CrossRef]
- Aranda, M.; Marques-Souza, H.; Bayer, T.; Tautz, D. The role of the segmentation gene hairy in Tribolium. Dev Genes Evol 2008, 218, 465–477. [Google Scholar] [CrossRef]
- Panin, V.M.; Papayannopoulos, V.; Wilson, R.; Irvine, K.D. Fringe modulates Notch-ligand interactions. Nature 1997, 387, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Moloney, D.J.; Panin, V.M.; Johnston, S.H.; Chen, J.; Shao, L.; Wilson, R.; Wang, Y.; Stanley, P.; Irvine, K.D.; Haltiwanger, R.S.; et al. Fringe is a glycosyltransferase that modifies Notch. Nature 2000, 406, 369–375. [Google Scholar] [CrossRef]
- Evrard, Y.A.; Lun, Y.; Aulehla, A.; Gan, L.; Johnson, R.L. lunatic fringe is an essential mediator of somite segmentation and patterning. Nature 1998, 394, 377–381. [Google Scholar] [CrossRef]
- Zhang, N.; Gridley, T. Defects in somite formation in lunatic fringe-deficient mice. Nature 1998, 394, 374–377. [Google Scholar] [CrossRef]
- Dearden, P.; Akam, M. A role for Fringe in segment morphogenesis but not segment formation in the grasshopper, Schistocerca gregaria. Dev Genes Evol 2000, 210, 329–336. [Google Scholar] [CrossRef]
- Liu, W. Functional analyses in the silkworm, Bombyx mori, support a role for Notch signaling in appendage development but not the groucho-dependent pair-rule process. J Exp Zool B Mol Dev Evol 2012, 318, 651–662. [Google Scholar] [CrossRef]
- Liu, W. Bmdelta phenotype implies involvement of Notch signaling in body segmentation and appendage development of silkworm, Bombyx mori. Arthropod Struct Dev 2013, 42, 143–151. [Google Scholar] [CrossRef]
- Tomoyasu, Y. What crustaceans can tell us about the evolution of insect wings and other morphologically novel structures. Curr Opin Genet Dev 2021, 69, 48–55. [Google Scholar] [CrossRef]
- Prokop, J.; Rosova, K.; Krzeminska, E.; Krzeminski, W.; Nel, A.; Engel, M.S. Abdominal serial homologues of wings in Paleozoic insects. Curr Biol 2022, 32, 3414–3422. [Google Scholar] [CrossRef] [PubMed]
- Ohde, T.; Mito, T.; Niimi, T. A hemimetabolous wing development suggests the wing origin from lateral tergum of a wingless ancestor. Nat Commun 2022, 13, 979. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.S. Wing vein patterning in Drosophila and the analysis of intercellular signaling. Annu Rev Cell Dev Biol 2007, 23, 293–319. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.K.; Irvine, K.D. The wing imaginal disc. Genetics 2022, 220. [Google Scholar] [CrossRef]
- Diaz-Benjumea, F.J.; Cohen, S.M. Serrate signals through Notch to establish a Wingless-dependent organizer at the dorsal/ventral compartment boundary of the Drosophila wing. Development 1995, 121, 4215–4225. [Google Scholar] [CrossRef]
- de Celis, J.F.; Garcia-Bellido, A.; Bray, S.J. Activation and function of Notch at the dorsal-ventral boundary of the wing imaginal disc. Development 1996, 122, 359–369. [Google Scholar] [CrossRef]
- Doherty, D.; Feger, G.; Younger-Shepherd, S.; Jan, L.Y.; Jan, Y.N. Delta is a ventral to dorsal signal complementary to Serrate, another Notch ligand, in Drosophila wing formation. Genes Dev 1996, 10, 421–434. [Google Scholar] [CrossRef]
- Irvine, K.D.; Wieschaus, E. fringe, a Boundary-specific signaling molecule, mediates interactions between dorsal and ventral cells during Drosophila wing development. Cell 1994, 79, 595–606. [Google Scholar] [CrossRef]
- Kim, J.; Irvine, K.D.; Carroll, S.B. Cell recognition, signal induction, and symmetrical gene activation at the dorsal-ventral boundary of the developing Drosophila wing. Cell 1995, 82, 795–802. [Google Scholar] [CrossRef]
- Fleming, R.J.; Gu, Y.; Hukriede, N.A. Serrate-mediated activation of Notch is specifically blocked by the product of the gene fringe in the dorsal compartment of the Drosophila wing imaginal disc. Development 1997, 124, 2973–2981. [Google Scholar] [CrossRef]
- Klein, T.; Arias, A.M. Interactions among Delta, Serrate and Fringe modulate Notch activity during Drosophila wing development. Development 1998, 125, 2951–2962. [Google Scholar] [CrossRef]
- Bruckner, K.; Perez, L.; Clausen, H.; Cohen, S. Glycosyltransferase activity of Fringe modulates Notch-Delta interactions. Nature 2000, 406, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Freeman, M. The notch signalling regulator fringe acts in the Golgi apparatus and requires the glycosyltransferase signature motif DXD. Curr Biol 2000, 10, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Charng, W.L.; Rana, N.A.; Kakuda, S.; Jaiswal, M.; Bayat, V.; Xiong, B.; Zhang, K.; Sandoval, H.; David, G.; et al. A mutation in EGF repeat-8 of Notch discriminates between Serrate/Jagged and Delta family ligands. Science 2012, 338, 1229–1232. [Google Scholar] [CrossRef]
- Pandey, A.; Harvey, B.M.; Lopez, M.F.; Ito, A.; Haltiwanger, R.S.; Jafar-Nejad, H. Glycosylation of Specific Notch EGF Repeats by O-Fut1 and Fringe Regulates Notch Signaling in Drosophila. Cell Rep 2019, 29, 2054–2066 e2056. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Benjumea, F.J.; Cohen, S.M. Interaction between dorsal and ventral cells in the imaginal disc directs wing development in Drosophila. Cell 1993, 75, 741–752. [Google Scholar] [CrossRef] [PubMed]
- de Celis, J.F.; Bray, S. Feed-back mechanisms affecting Notch activation at the dorsoventral boundary in the Drosophila wing. Development 1997, 124, 3241–3251. [Google Scholar] [CrossRef]
- Yan, S.J.; Gu, Y.; Li, W.X.; Fleming, R.J. Multiple signaling pathways and a selector protein sequentially regulate Drosophila wing development. Development 2004, 131, 285–298. [Google Scholar] [CrossRef]
- LeBon, L.; Lee, T.V.; Sprinzak, D.; Jafar-Nejad, H.; Elowitz, M.B. Fringe proteins modulate Notch-ligand cis and trans interactions to specify signaling states. Elife 2014, 3, e02950. [Google Scholar] [CrossRef]
- Couso, J.P.; Knust, E.; Martinez Arias, A. Serrate and wingless cooperate to induce vestigial gene expression and wing formation in Drosophila. Curr Biol 1995, 5, 1437–1448. [Google Scholar] [CrossRef]
- Rulifson, E.J.; Blair, S.S. Notch regulates wingless expression and is not required for reception of the paracrine wingless signal during wing margin neurogenesis in Drosophila. Development 1995, 121, 2813–2824. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sebring, A.; Esch, J.J.; Kraus, M.E.; Vorwerk, K.; Magee, J.; Carroll, S.B. Integration of positional signals and regulation of wing formation and identity by Drosophila vestigial gene. Nature 1996, 382, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Neumann, C.J.; Cohen, S.M. A hierarchy of cross-regulation involving Notch, wingless, vestigial and cut organizes the dorsal/ventral axis of the Drosophila wing. Development 1996, 122, 3477–3485. [Google Scholar] [CrossRef] [PubMed]
- Rulifson, E.J.; Micchelli, C.A.; Axelrod, J.D.; Perrimon, N.; Blair, S.S. wingless refines its own expression domain on the Drosophila wing margin. Nature 1996, 384, 72–74. [Google Scholar] [CrossRef]
- Micchelli, C.A.; Rulifson, E.J.; Blair, S.S. The function and regulation of cut expression on the wing margin of Drosophila: Notch, Wingless and a dominant negative role for Delta and Serrate. Development 1997, 124, 1485–1495. [Google Scholar] [CrossRef]
- Go, M.J.; Eastman, D.S.; Artavanis-Tsakonas, S. Cell proliferation control by Notch signaling in Drosophila development. Development 1998, 125, 2031–2040. [Google Scholar] [CrossRef]
- Klein, T.; Arias, A.M. The vestigial gene product provides a molecular context for the interpretation of signals during the development of the wing in Drosophila. Development 1999, 126, 913–925. [Google Scholar] [CrossRef]
- Giraldez, A.J.; Cohen, S.M. Wingless and Notch signaling provide cell survival cues and control cell proliferation during wing development. Development 2003, 130, 6533–6543. [Google Scholar] [CrossRef]
- Rafel, N.; Milan, M. Notch signalling coordinates tissue growth and wing fate specification in Drosophila. Development 2008, 135, 3995–4001. [Google Scholar] [CrossRef]
- Jia, D.; Bryant, J.; Jevitt, A.; Calvin, G.; Deng, W.M. The Ecdysone and Notch Pathways Synergistically Regulate Cut at the Dorsal-Ventral Boundary in Drosophila Wing Discs. J Genet Genomics 2016, 43, 179–186. [Google Scholar] [CrossRef]
- Sturtevant, M.A.; Bier, E. Analysis of the genetic hierarchy guiding wing vein development in Drosophila. Development 1995, 121, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Huppert, S.S.; Jacobsen, T.L.; Muskavitch, M.A. Feedback regulation is central to Delta-Notch signalling required for Drosophila wing vein morphogenesis. Development 1997, 124, 3283–3291. [Google Scholar] [CrossRef] [PubMed]
- de Celis, J.F.; Bray, S.; Garcia-Bellido, A. Notch signalling regulates veinlet expression and establishes boundaries between veins and interveins in the Drosophila wing. Development 1997, 124, 1919–1928. [Google Scholar] [CrossRef]
- De Celis, J.F. Pattern formation in the Drosophila wing: The development of the veins. Bioessays 2003, 25, 443–451. [Google Scholar] [CrossRef]
- Crozatier, M.; Glise, B.; Khemici, V.; Vincent, A. Vein-positioning in the Drosophila wing in response to Hh; new roles of Notch signaling. Mech Dev 2003, 120, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Dambly-Chaudiere, C.; Ghysen, A. The emergence of sense organs in the wing disc of Drosophila. Development 1991, 111, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Troost, T.; Binshtok, U.; Sprinzak, D.; Klein, T. Cis-inhibition suppresses basal Notch signaling during sensory organ precursor selection. Proc Natl Acad Sci U S A 2023, 120, e2214535120. [Google Scholar] [CrossRef] [PubMed]
- Schweisguth, F. Asymmetric cell division in the Drosophila bristle lineage: from the polarization of sensory organ precursor cells to Notch-mediated binary fate decision. Wiley Interdiscip Rev Dev Biol 2015, 4, 299–309. [Google Scholar] [CrossRef]
- Li, Y.; Liu, T.; Zhang, J. The ATPase TER94 regulates Notch signaling during Drosophila wing development. Biol Open 2019, 8. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Y.; Shen, J.; Zhang, J. The Ubiquitin Conjugating Enzyme UbcD1 is Required for Notch Signaling Activation During Drosophila Wing Development. Front Genet 2021, 12, 770853. [Google Scholar] [CrossRef]
- Mo, D.; Liu, C.; Chen, Y.; Cheng, X.; Shen, J.; Zhao, L.; Zhang, J. The mitochondrial ribosomal protein mRpL4 regulates Notch signaling. EMBO Rep 2023, 24, e55764. [Google Scholar] [CrossRef] [PubMed]
- van Breugel, F.M. The action of the Notch locus in Drosophila hydei. Genetica 1971, 42, 25–41. [Google Scholar] [CrossRef] [PubMed]
- van Breugel, F.M.; Vermet-Rozeboom, E.; Gloor, H. Phenocopies in Drosophila hydei induced by actinomycin D and fluorouracil with special reference to Notch mutants. Wilehm Roux Arch Dev Biol 1975, 178, 309–320. [Google Scholar] [CrossRef]
- van Breugel, F.M.; van den Broek, H.C.; Grond, C.; den Hertog, F. Bristle patterns, clones and cell competition along the anterior margin of Notch wings of Drosophila hydei. Wilehm Roux Arch Dev Biol 1981, 190, 40–48. [Google Scholar] [CrossRef]
- Van Breugel, F.M.; Langhout, B.V. The NOTCH Locus of DROSOPHILA HYDEI: Alleles, Phenotypes and Functional Organization. Genetics 1983, 103, 197–217. [Google Scholar] [CrossRef]
- Gubenko, I.S.; Subbota, R.P.; Kozereretskaia, I.A.; Vagin Iu, V. [The Delta locus in Drosophila virilis: alleles, wing phenotypes and genetic interactions]. Tsitol Genet 1998, 32, 22–34. [Google Scholar]
- Gubenko, I.S.; Zelentsova, E.S.; Kozeretskaia, I.A.; Bagin Iu, V. [Genetic characteristics of the wing mutation Odd(22) and its interaction with alleles of the Delta locus in Drosophila virilis]. Genetika 2000, 36, 1049–1054. [Google Scholar]
- Rybtsova, N.N.; Zelentsova, E.S.; Lezin, G.T.; Evgen’ev, M.B.; Korochkin, L.I. Comparison of the structure and embryonic expression of Delta in Drosophila virilis and D. melanogaster. Molecular Biology 2000, 34, 673–679. [Google Scholar] [CrossRef]
- Gubenko, I.S.; Subbota, R.P.; Zelentsova, E.S. Notch122, mutation of Abruptex-type Notch locus in Drosophila virilis: Peculiarities of genetic interactions. Cytol Genet 2010, 44, 170–173. [Google Scholar] [CrossRef]
- Hiroyoshi, T. Some new mutants and linkage groups of the house fly. J Econ Entomol 1960, 53, 985–990. [Google Scholar] [CrossRef]
- Nickel, C.A. , Wagoner, D. E. Some New Mutants of House Flies and Their Linkage Groups and Map positions. J Econ Entomol 1970, 63, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Hiroyoshi, T. Some new mutants and revised linkage maps of the housefly, Musca domestica L. Japan J Genetics 1977, 52, 275–288. [Google Scholar] [CrossRef]
- Scott, J.G.; Warren, W.C.; Beukeboom, L.W.; Bopp, D.; Clark, A.G.; Giers, S.D.; Hediger, M.; Jones, A.K.; Kasai, S.; Leichter, C.A.; et al. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol 2014, 15, 466. [Google Scholar] [CrossRef]
- Heinze, S.D.; Kohlbrenner, T.; Ippolito, D.; Meccariello, A.; Burger, A.; Mosimann, C.; Saccone, G.; Bopp, D. CRISPR-Cas9 targeted disruption of the yellow ortholog in the housefly identifies the brown body locus. Sci Rep 2017, 7, 4582. [Google Scholar] [CrossRef] [PubMed]
- Foster, G. , Whitten, M., Konovalov, C., Arnold, J., Maffi, G.. Autosomal genetic maps of the Australian Sheep Blowfly, Lucilia cuprina dorsalis R.-D. (Diptera: Calliphoridae), and possible correlations with the linkage maps of Musca domestica L. and Drosophila melanogaster (Mg.). Genet Res 1981, 37, 55–69. [Google Scholar] [CrossRef]
- Foster, G.G.; Weller, G.L.; Bedo, D.G. Homozygous-viable pericentric inversions for genetic control of Lucilia Cuprina. Theor Appl Genet 1991, 82, 681–689. [Google Scholar] [CrossRef]
- Weller, G.L.; Foster, G.G. Genetic maps of the sheep blowfly Lucilia cuprina: linkage-group correlations with other dipteran genera. Genome 1993, 36, 495–506. [Google Scholar] [CrossRef]
- Davies, A.G.; Game, A.Y.; Chen, Z.; Williams, T.J.; Goodall, S.; Yen, J.L.; McKenzie, J.A.; Batterham, P. Scalloped wings is the Lucilia cuprina Notch homologue and a candidate for the modifier of fitness and asymmetry of diazinon resistance. Genetics 1996, 143, 1321–1337. [Google Scholar] [CrossRef]
- Chen, Z.; Newsome, T.; McKenzie, J.A.; Batterham, P. Molecular characterization of the Notch homologue from the Australian sheep blowfly, Lucilia cuprina. Insect Biochem Mol Biol 1998, 28, 601–612. [Google Scholar] [CrossRef]
- Fujiwara, H.; Hojyo, T. Developmental profiles of wing imaginal discs of flugellos(fl), a wingless mutant of the silkworm, Bombyx mori. Dev Genes Evol 1997, 207, 12–18. [Google Scholar] [CrossRef]
- Hojyo, T.; Fujiwara, H. Reciprocal transplantation of wing discs between a wing deficient mutant (fl) and wild type of the silkworm, Bombyx mori. Dev Growth Differ 1997, 39, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Fujiwara, H. Expression of ecdysteroid-regulated genes is reduced specifically in the wing discs of the wing-deficient mutant (fl) of Bombyx mori. Dev Genes Evol 2000, 210, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, T.M.; Fujiwara, H. Identification and characterization of genes abnormally expressed in wing-deficient mutant (flugellos) of the silkworm, Bombyx mori. Insect Biochem Mol Biol 2002, 32, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Matsunaga, T.M.; Futahashi, R.; Kojima, T.; Mita, K.; Banno, Y.; Fujiwara, H. Positional cloning of a Bombyx wingless locus flugellos (fl) reveals a crucial role for fringe that is specific for wing morphogenesis. Genetics 2008, 179, 875–885. [Google Scholar] [CrossRef]
- Kango-Singh, M.; Singh, A.; Gopinathan, K.P. The wings of Bombyx mori develop from larval discs exhibiting an early differentiated state: a preliminary report. J Biosci 2001, 26, 167–177. [Google Scholar] [CrossRef]
- Ling, L.; Ge, X.; Li, Z.; Zeng, B.; Xu, J.; Chen, X.; Shang, P.; James, A.A.; Huang, Y.; Tan, A. MiR-2 family targets awd and fng to regulate wing morphogenesis in Bombyx mori. RNA Biol 2015, 12, 742–748. [Google Scholar] [CrossRef]
- Fujii, T.; Abe, H.; Katsuma, S.; Shimada, T. Identification and characterization of the fusion transcript, composed of the apterous homolog and a putative protein phosphatase gene, generated by 1.5-Mb interstitial deletion in the vestigial (Vg) mutant of Bombyx mori. Insect Biochem Mol Biol 2011, 41, 306–312. [Google Scholar] [CrossRef]
- Carroll, S.B.; Gates, J.; Keys, D.N.; Paddock, S.W.; Panganiban, G.E.; Selegue, J.E.; Williams, J.A. Pattern formation and eyespot determination in butterfly wings. Science 1994, 265, 109–114. [Google Scholar] [CrossRef]
- Reed, R.D.; Serfas, M.S. Butterfly wing pattern evolution is associated with changes in a Notch/Distal-less temporal pattern formation process. Curr Biol 2004, 14, 1159–1166. [Google Scholar] [CrossRef]
- Reed, R.D.; Chen, P.H.; Frederik Nijhout, H. Cryptic variation in butterfly eyespot development: the importance of sample size in gene expression studies. Evol Dev 2007, 9, 2–9. [Google Scholar] [CrossRef]
- Saenko, S.V.; Marialva, M.S.; Beldade, P. Involvement of the conserved Hox gene Antennapedia in the development and evolution of a novel trait. Evodevo 2011, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Shirai, L.T.; Saenko, S.V.; Keller, R.A.; Jeronimo, M.A.; Brakefield, P.M.; Descimon, H.; Wahlberg, N.; Beldade, P. Evolutionary history of the recruitment of conserved developmental genes in association to the formation and diversification of a novel trait. BMC Evol Biol 2012, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.C.; Tong, X.L.; Gall, L.F.; Piel, W.H.; Monteiro, A. A single origin for nymphalid butterfly eyespots followed by widespread loss of associated gene expression. PLoS Genet 2012, 8, e1002893. [Google Scholar] [CrossRef]
- Oliver, J.C.; Ramos, D.; Prudic, K.L.; Monteiro, A. Temporal gene expression variation associated with eyespot size plasticity in Bicyclus anynana. PLoS One 2013, 8, e65830. [Google Scholar] [CrossRef] [PubMed]
- Connahs, H.; Rhen, T.; Simmons, R.B. Transcriptome analysis of the painted lady butterfly, Vanessa cardui during wing color pattern development. BMC Genomics 2016, 17, 270. [Google Scholar] [CrossRef] [PubMed]
- Wee, J.L.Q.; Murugesan, S.N.; Wheat, C.W.; Monteiro, A. The genetic basis of wing spots in Pieris canidia butterflies. BMC Genomics 2023, 24, 169. [Google Scholar] [CrossRef]
- Reed, R.D. Evidence for Notch-mediated lateral inhibition in organizing butterfly wing scales. Dev Genes Evol 2004, 214, 43–46. [Google Scholar] [CrossRef]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; John, O.S.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S.; et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef]
- Tomoyasu, Y.; Arakane, Y.; Kramer, K.J.; Denell, R.E. Repeated co-options of exoskeleton formation during wing-to-elytron evolution in beetles. Curr Biol 2009, 19, 2057–2065. [Google Scholar] [CrossRef]
- Ravisankar, P.; Lai, Y.T.; Sambrani, N.; Tomoyasu, Y. Comparative developmental analysis of Drosophila and Tribolium reveals conserved and diverged roles of abrupt in insect wing evolution. Dev Biol 2016, 409, 518–529. [Google Scholar] [CrossRef]
- Schmidt-Ott, U.; Lynch, J.A. Emerging developmental genetic model systems in holometabolous insects. Curr Opin Genet Dev 2016, 39, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Klingler, M.; Bucher, G. The red flour beetle T. castaneum: elaborate genetic toolkit and unbiased large scale RNAi screening to study insect biology and evolution. Evodevo 2022, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.B.; Xiao, S.; Wang, Y.; Hua, H.X. Notch is an alternative splicing gene in brown planthopper, Nilaparvata lugens. Arch Insect Biochem Physiol 2022, 110, e21894. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wei, W.; Chu, Y.; Zhang, L.; Shen, J.; An, C. De novo transcriptome analysis of wing development-related signaling pathways in Locusta migratoria manilensis and Ostrinia furnacalis (Guenee). PLoS One 2014, 9, e106770. [Google Scholar] [CrossRef]
- Feindt, W.; Oppenheim, S.J.; DeSalle, R.; Goldstein, P.Z.; Hadrys, H. Transcriptome profiling with focus on potential key genes for wing development and evolution in Megaloprepus caerulatus, the damselfly species with the world's largest wings. PLoS One 2018, 13, e0189898. [Google Scholar] [CrossRef]
- Cordoba, S.; Estella, C. Role of Notch Signaling in Leg Development in Drosophila melanogaster. Adv Exp Med Biol 2020, 1218, 103–127. [Google Scholar] [CrossRef]
- Shellenbarger, D.L.; Mohler, J.D. Temperature-sensitive periods and autonomy of pleiotropic effects of l(1)Nts1, a conditional notch lethal in Drosophila. Dev Biol 1978, 62, 432–446. [Google Scholar] [CrossRef]
- Parody, T.R.; Muskavitch, M.A. The pleiotropic function of Delta during postembryonic development of Drosophila melanogaster. Genetics 1993, 135, 527–539. [Google Scholar] [CrossRef]
- Speicher, S.A.; Thomas, U.; Hinz, U.; Knust, E. The Serrate locus of Drosophila and its role in morphogenesis of the wing imaginal discs: control of cell proliferation. Development 1994, 120, 535–544. [Google Scholar] [CrossRef]
- de Celis, J.F.; Tyler, D.M.; de Celis, J.; Bray, S.J. Notch signalling mediates segmentation of the Drosophila leg. Development 1998, 125, 4617–4626. [Google Scholar] [CrossRef]
- Rauskolb, C.; Irvine, K.D. Notch-mediated segmentation and growth control of the Drosophila leg. Dev Biol 1999, 210, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.A.; Klein, T.; Arias, A.M.; Couso, J.P. Composite signalling from Serrate and Delta establishes leg segments in Drosophila through Notch. Development 1999, 126, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Agrawal, N.; Banerjee, S.; Sardesai, D.; Dalal, J.S.; Bhojwani, J.; Sinha, P. Spatial regulation of DELTA expression mediates NOTCH signalling for segmentation of Drosophila legs. Mech Dev 2001, 105, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Suzanne, M. Molecular and cellular mechanisms involved in leg joint morphogenesis. Semin Cell Dev Biol 2016, 55, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Kerber, B.; Monge, I.; Mueller, M.; Mitchell, P.J.; Cohen, S.M. The AP-2 transcription factor is required for joint formation and cell survival in Drosophila leg development. Development 2001, 128, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Buckles, G.R.; Rauskolb, C.; Villano, J.L.; Katz, F.N. Four-jointed interacts with dachs, abelson and enabled and feeds back onto the Notch pathway to affect growth and segmentation in the Drosophila leg. Development 2001, 128, 3533–3542. [Google Scholar] [CrossRef]
- de Celis Ibeas, J.M.; Bray, S.J. Bowl is required downstream of Notch for elaboration of distal limb patterning. Development 2003, 130, 5943–5952. [Google Scholar] [CrossRef]
- Hao, I.; Green, R.B.; Dunaevsky, O.; Lengyel, J.A.; Rauskolb, C. The odd-skipped family of zinc finger genes promotes Drosophila leg segmentation. Dev Biol 2003, 263, 282–295. [Google Scholar] [CrossRef]
- Campbell, G. Regulation of gene expression in the distal region of the Drosophila leg by the Hox11 homolog, C15. Dev Biol 2005, 278, 607–618. [Google Scholar] [CrossRef]
- Ciechanska, E.; Dansereau, D.A.; Svendsen, P.C.; Heslip, T.R.; Brook, W.J. dAP-2 and defective proventriculus regulate Serrate and Delta expression in the tarsus of Drosophila melanogaster. Genome 2007, 50, 693–705. [Google Scholar] [CrossRef]
- Shirai, T.; Yorimitsu, T.; Kiritooshi, N.; Matsuzaki, F.; Nakagoshi, H. Notch signaling relieves the joint-suppressive activity of Defective proventriculus in the Drosophila leg. Dev Biol 2007, 312, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, L.; Hatini, V. Essential roles for lines in mediating leg and antennal proximodistal patterning and generating a stable Notch signaling interface at segment borders. Dev Biol 2009, 330, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Pueyo, J.I.; Couso, J.P. Tarsal-less peptides control Notch signalling through the Shavenbaby transcription factor. Dev Biol 2011, 355, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Capilla, A.; Johnson, R.; Daniels, M.; Benavente, M.; Bray, S.J.; Galindo, M.I. Planar cell polarity controls directional Notch signaling in the Drosophila leg. Development 2012, 139, 2584–2593. [Google Scholar] [CrossRef] [PubMed]
- Guarner, A.; Manjon, C.; Edwards, K.; Steller, H.; Suzanne, M.; Sanchez-Herrero, E. The zinc finger homeodomain-2 gene of Drosophila controls Notch targets and regulates apoptosis in the tarsal segments. Dev Biol 2014, 385, 350–365. [Google Scholar] [CrossRef]
- Cordoba, S.; Requena, D.; Jory, A.; Saiz, A.; Estella, C. The evolutionarily conserved transcription factor Sp1 controls appendage growth through Notch signaling. Development 2016, 143, 3623–3631. [Google Scholar] [CrossRef]
- Fan, Z.; Zhang, J.; Wang, D.; Shen, J. T-box transcription factors Dorsocross and optomotor-blind control Drosophila leg patterning in a functionally redundant manner. Insect Biochem Mol Biol 2021, 129, 103516. [Google Scholar] [CrossRef]
- Guntur, A.R.; Venkatanarayan, A.; Gangula, S.; Lundell, M.J. Zfh-2 facilitates Notch-induced apoptosis in the CNS and appendages of Drosophila melanogaster. Dev Biol 2021, 475, 65–79. [Google Scholar] [CrossRef]
- Kojima, T. Developmental mechanism of the tarsus in insect legs. Curr Opin Insect Sci 2017, 19, 36–42. [Google Scholar] [CrossRef]
- Cordoba, S.; Estella, C. The bHLH-PAS transcription factor dysfusion regulates tarsal joint formation in response to Notch activity during drosophila leg development. PLoS Genet 2014, 10, e1004621. [Google Scholar] [CrossRef]
- Tajiri, R.; Misaki, K.; Yonemura, S.; Hayashi, S. Dynamic shape changes of ECM-producing cells drive morphogenesis of ball-and-socket joints in the fly leg. Development 2010, 137, 2055–2063. [Google Scholar] [CrossRef]
- Tajiri, R.; Misaki, K.; Yonemura, S.; Hayashi, S. Joint morphology in the insect leg: evolutionary history inferred from Notch loss-of-function phenotypes in Drosophila. Development 2011, 138, 4621–4626. [Google Scholar] [CrossRef] [PubMed]
- Angelini, D.R.; Smith, F.W.; Jockusch, E.L. Extent With Modification: Leg Patterning in the Beetle Tribolium castaneum and the Evolution of Serial Homologs. G3 (Bethesda) 2012, 2, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Siemanowski, J.; Richter, T.; Dao, V.A.; Bucher, G. Notch signaling induces cell proliferation in the labrum in a regulatory network different from the thoracic legs. Dev Biol 2015, 408, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Diaz-Benjumea, F.J.; Cohen, S.M. Nubbin encodes a POU-domain protein required for proximal-distal patterning in the Drosophila wing. Development 1995, 121, 589–599. [Google Scholar] [CrossRef]
- Cifuentes, F.J.; Garcia-Bellido, A. Proximo-distal specification in the wing disc of Drosophila by the nubbin gene. Proc Natl Acad Sci U S A 1997, 94, 11405–11410. [Google Scholar] [CrossRef]
- Li, H.; Popadic, A. Analysis of nubbin expression patterns in insects. Evol Dev 2004, 6, 310–324. [Google Scholar] [CrossRef]
- Hrycaj, S.; Mihajlovic, M.; Mahfooz, N.; Couso, J.P.; Popadic, A. RNAi analysis of nubbin embryonic functions in a hemimetabolous insect, Oncopeltus fasciatus. Evol Dev 2008, 10, 705–716. [Google Scholar] [CrossRef]
- Turchyn, N.; Chesebro, J.; Hrycaj, S.; Couso, J.P.; Popadic, A. Evolution of nubbin function in hemimetabolous and holometabolous insect appendages. Dev Biol 2011, 357, 83–95. [Google Scholar] [CrossRef]
- Zhou, H.; Ma, Z.; Wang, Z.; Yan, S.; Wang, D.; Shen, J. Hedgehog signaling regulates regenerative patterning and growth in Harmonia axyridis leg. Cell Mol Life Sci 2021, 78, 2185–2197. [Google Scholar] [CrossRef]
- Prpic, N.M.; Damen, W.G. Notch-mediated segmentation of the appendages is a molecular phylotypic trait of the arthropods. Dev Biol 2009, 326, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Klusza, S.; Deng, W.M. At the crossroads of differentiation and proliferation: precise control of cell-cycle changes by multiple signaling pathways in Drosophila follicle cells. Bioessays 2011, 33, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Roth, S. Drosophila oogenesis: coordinating germ line and soma. Curr Biol 2001, 11, R779–R781. [Google Scholar] [CrossRef] [PubMed]
- Ruohola, H.; Bremer, K.A.; Baker, D.; Swedlow, J.R.; Jan, L.Y.; Jan, Y.N. Role of neurogenic genes in establishment of follicle cell fate and oocyte polarity during oogenesis in Drosophila. Cell 1991, 66, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Bender, L.B.; Kooh, P.J.; Muskavitch, M.A. Complex function and expression of Delta during Drosophila oogenesis. Genetics 1993, 133, 967–978. [Google Scholar] [CrossRef]
- Larkin, M.K.; Holder, K.; Yost, C.; Giniger, E.; Ruohola-Baker, H. Expression of constitutively active Notch arrests follicle cells at a precursor stage during Drosophila oogenesis and disrupts the anterior-posterior axis of the oocyte. Development 1996, 122, 3639–3650. [Google Scholar] [CrossRef]
- Jackson, S.M.; Blochlinger, K. cut interacts with Notch and protein kinase A to regulate egg chamber formation and to maintain germline cyst integrity during Drosophila oogenesis. Development 1997, 124, 3663–3672. [Google Scholar] [CrossRef]
- Zhao, D.; Clyde, D.; Bownes, M. Expression of fringe is down regulated by Gurken/Epidermal Growth Factor Receptor signalling and is required for the morphogenesis of ovarian follicle cells. J Cell Sci 2000, 113 Pt 21, 3781–3794. [Google Scholar] [CrossRef]
- Lopez-Schier, H.; St Johnston, D. Delta signaling from the germ line controls the proliferation and differentiation of the somatic follicle cells during Drosophila oogenesis. Genes Dev 2001, 15, 1393–1405. [Google Scholar] [CrossRef]
- Deng, W.M.; Althauser, C.; Ruohola-Baker, H. Notch-Delta signaling induces a transition from mitotic cell cycle to endocycle in Drosophila follicle cells. Development 2001, 128, 4737–4746. [Google Scholar] [CrossRef]
- Grammont, M.; Irvine, K.D. fringe and Notch specify polar cell fate during Drosophila oogenesis. Development 2001, 128, 2243–2253. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, V.; Althauser, C.; Shcherbata, H.R.; Deng, W.M.; Ruohola-Baker, H. Notch-dependent Fizzy-related/Hec1/Cdh1 expression is required for the mitotic-to-endocycle transition in Drosophila follicle cells. Curr Biol 2004, 14, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Shcherbata, H.R.; Althauser, C.; Findley, S.D.; Ruohola-Baker, H. The mitotic-to-endocycle switch in Drosophila follicle cells is executed by Notch-dependent regulation of G1/S, G2/M and M/G1 cell-cycle transitions. Development 2004, 131, 3169–3181. [Google Scholar] [CrossRef] [PubMed]
- Adam, J.C.; Montell, D.J. A role for extra macrochaetae downstream of Notch in follicle cell differentiation. Development 2004, 131, 5971–5980. [Google Scholar] [CrossRef]
- Sun, J.; Deng, W.M. Notch-dependent downregulation of the homeodomain gene cut is required for the mitotic cycle/endocycle switch and cell differentiation in Drosophila follicle cells. Development 2005, 132, 4299–4308. [Google Scholar] [CrossRef]
- Sun, J.; Deng, W.M. Hindsight mediates the role of notch in suppressing hedgehog signaling and cell proliferation. Dev Cell 2007, 12, 431–442. [Google Scholar] [CrossRef]
- Sun, J.; Smith, L.; Armento, A.; Deng, W.M. Regulation of the endocycle/gene amplification switch by Notch and ecdysone signaling. J Cell Biol 2008, 182, 885–896. [Google Scholar] [CrossRef]
- Gonzalez-Reyes, A.; St Johnston, D. Role of oocyte position in establishment of anterior-posterior polarity in Drosophila. Science 1994, 266, 639–642. [Google Scholar] [CrossRef]
- Torres, I.L.; Lopez-Schier, H.; St Johnston, D. A Notch/Delta-dependent relay mechanism establishes anterior-posterior polarity in Drosophila. Dev Cell 2003, 5, 547–558. [Google Scholar] [CrossRef]
- Assa-Kunik, E.; Torres, I.L.; Schejter, E.D.; Johnston, D.S.; Shilo, B.Z. Drosophila follicle cells are patterned by multiple levels of Notch signaling and antagonism between the Notch and JAK/STAT pathways. Development 2007, 134, 1161–1169. [Google Scholar] [CrossRef]
- Roth, S.; Lynch, J.A. Symmetry breaking during Drosophila oogenesis. Cold Spring Harb Perspect Biol 2009, 1, a001891. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gridley, T. Notch Signaling during Oogenesis in Drosophila melanogaster. Genet Res Int 2012, 2012, 648207. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Spradling, A.C. A niche maintaining germ line stem cells in the Drosophila ovary. Science 2000, 290, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.J.; Shcherbata, H.R.; Reynolds, S.H.; Fischer, K.A.; Hatfield, S.D.; Ruohola-Baker, H. Stem cells signal to the niche through the Notch pathway in the Drosophila ovary. Curr Biol 2006, 16, 2352–2358. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Call, G.B.; Kirilly, D.; Xie, T. Notch signaling controls germline stem cell niche formation in the Drosophila ovary. Development 2007, 134, 1071–1080. [Google Scholar] [CrossRef]
- Dai, W.; Peterson, A.; Kenney, T.; Burrous, H.; Montell, D.J. Quantitative microscopy of the Drosophila ovary shows multiple niche signals specify progenitor cell fate. Nat Commun 2017, 8, 1244. [Google Scholar] [CrossRef]
- Panchal, T.; Chen, X.; Alchits, E.; Oh, Y.; Poon, J.; Kouptsova, J.; Laski, F.A.; Godt, D. Specification and spatial arrangement of cells in the germline stem cell niche of the Drosophila ovary depend on the Maf transcription factor Traffic jam. PLoS Genet 2017, 13, e1006790. [Google Scholar] [CrossRef]
- Shimizu, H.; Wilkin, M.B.; Woodcock, S.A.; Bonfini, A.; Hung, Y.; Mazaleyrat, S.; Baron, M. The Drosophila ZO-1 protein Polychaetoid suppresses Deltex-regulated Notch activity to modulate germline stem cell niche formation. Open Biol 2017, 7. [Google Scholar] [CrossRef]
- Yatsenko, A.S.; Shcherbata, H.R. Stereotypical architecture of the stem cell niche is spatiotemporally established by miR-125-dependent coordination of Notch and steroid signaling. Development 2018, 145. [Google Scholar] [CrossRef]
- Zhao, S.; Wu, C.; Gao, Z.; Li, X.; Guo, Z.; Wang, Z. Notch signaling governs the expression of glypican Dally to define the stem cell niche. Biol Open 2020, 9. [Google Scholar] [CrossRef]
- Yatsenko, A.S.; Shcherbata, H.R. Distant activation of Notch signaling induces stem cell niche assembly. PLoS Genet 2021, 17, e1009489. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Gao, Y.; Xiao, G. The expression of Catsup in escort cells affects Drosophila ovarian stem cell niche establishment and germline stem cells self-renewal via Notch signaling. Biochem Biophys Res Commun 2023, 641, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.J.; Drummond-Barbosa, D. Insulin levels control female germline stem cell maintenance via the niche in Drosophila. Proc Natl Acad Sci U S A 2009, 106, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.J.; Drummond-Barbosa, D. Insulin signals control the competence of the Drosophila female germline stem cell niche to respond to Notch ligands. Dev Biol 2011, 350, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.A.; Wang, W.D.; Chen, C.T.; Tseng, C.Y.; Chen, Y.N.; Hsu, H.J. FOXO/Fringe is necessary for maintenance of the germline stem cell niche in response to insulin insufficiency. Dev Biol 2013, 382, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.Y.; Kao, S.H.; Wan, C.L.; Cho, Y.; Tung, S.Y.; Hsu, H.J. Notch signaling mediates the age-associated decrease in adhesion of germline stem cells to the niche. PLoS Genet 2014, 10, e1004888. [Google Scholar] [CrossRef]
- Bonfini, A.; Wilkin, M.B.; Baron, M. Reversible regulation of stem cell niche size associated with dietary control of Notch signalling. BMC Dev Biol 2015, 15, 8. [Google Scholar] [CrossRef]
- Song, J.; Zhou, S. Post-transcriptional regulation of insect metamorphosis and oogenesis. Cell Mol Life Sci 2020, 77, 1893–1909. [Google Scholar] [CrossRef]
- Belles, X.; Piulachs, M.D. Ecdysone signalling and ovarian development in insects: from stem cells to ovarian follicle formation. Biochim Biophys Acta 2015, 1849, 181–186. [Google Scholar] [CrossRef]
- Irles, P.; Piulachs, M.D. Unlike in Drosophila Meroistic Ovaries, hippo represses notch in Blattella germanica Panoistic ovaries, triggering the mitosis-endocycle switch in the follicular cells. PLoS One 2014, 9, e113850. [Google Scholar] [CrossRef]
- Elshaer, N.; Piulachs, M.D. Crosstalk of EGFR signalling with Notch and Hippo pathways to regulate cell specification, migration and proliferation in cockroach panoistic ovaries. Biol Cell 2015, 107, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Irles, P.; Elshaer, N.; Piulachs, M.D. The Notch pathway regulates both the proliferation and differentiation of follicular cells in the panoistic ovary of Blattella germanica. Open Biol 2016, 6, 150197. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Chelemen, F.; Pagone, V.; Elshaer, N.; Irles, P.; Piulachs, M.D. Eyes absent in the cockroach panoistic ovaries regulates proliferation and differentiation through ecdysone signalling. Insect Biochem Mol Biol 2020, 123, 103407. [Google Scholar] [CrossRef] [PubMed]
- Baumer, D.; Strohlein, N.M.; Schoppmeier, M. Opposing effects of Notch-signaling in maintaining the proliferative state of follicle cells in the telotrophic ovary of the beetle Tribolium. Front Zool 2012, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Teuscher, M.; Strohlein, N.; Birkenbach, M.; Schultheis, D.; Schoppmeier, M. TC003132 is essential for the follicle stem cell lineage in telotrophic Tribolium oogenesis. Front Zool 2017, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Li, W.; Zhao, H.; Zhou, S. Clustered miR-2, miR-13a, miR-13b and miR-71 coordinately target Notch gene to regulate oogenesis of the migratory locust Locusta migratoria. Insect Biochem Mol Biol 2019, 106, 39–46. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory Mechanisms of Vitellogenesis in Insects. Front Cell Dev Biol 2020, 8, 593613. [Google Scholar] [CrossRef]
- Duncan, E.J.; Hyink, O.; Dearden, P.K. Notch signalling mediates reproductive constraint in the adult worker honeybee. Nat Commun 2016, 7, 12427. [Google Scholar] [CrossRef]
- Duncan, E.J.; Leask, M.P.; Dearden, P.K. Genome Architecture Facilitates Phenotypic Plasticity in the Honeybee (Apis mellifera). Mol Biol Evol 2020, 37, 1964–1978. [Google Scholar] [CrossRef]
- Wilson, M.J.; Abbott, H.; Dearden, P.K. The evolution of oocyte patterning in insects: multiple cell-signaling pathways are active during honeybee oogenesis and are likely to play a role in axis patterning. Evol Dev 2011, 13, 127–137. [Google Scholar] [CrossRef]
- Chen, X.; Ma, C.; Chen, C.; Lu, Q.; Shi, W.; Liu, Z.; Wang, H.; Guo, H. Integration of lncRNA-miRNA-mRNA reveals novel insights into oviposition regulation in honey bees. PeerJ 2017, 5, e3881. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Li, R.; Zhang, M.Y.; Liu, Y.W.; Zhang, Z.; Smagghe, G.; Wang, J.J. Comparative Proteomic Profiling Reveals Molecular Characteristics Associated with Oogenesis and Oocyte Maturation during Ovarian Development of Bactrocera dorsalis (Hendel). Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Chowanski, S.; Walkowiak-Nowicka, K.; Winkiel, M.; Marciniak, P.; Urbanski, A.; Pacholska-Bogalska, J. Insulin-Like Peptides and Cross-Talk With Other Factors in the Regulation of Insect Metabolism. Front Physiol 2021, 12, 701203. [Google Scholar] [CrossRef] [PubMed]
- Smykal, V.; Raikhel, A.S. Nutritional Control of Insect Reproduction. Curr Opin Insect Sci 2015, 11, 31–38. [Google Scholar] [CrossRef]
- Jouandin, P.; Ghiglione, C.; Noselli, S. Starvation induces FoxO-dependent mitotic-to-endocycle switch pausing during Drosophila oogenesis. Development 2014, 141, 3013–3021. [Google Scholar] [CrossRef]
- Foronda, D.; Weng, R.; Verma, P.; Chen, Y.W.; Cohen, S.M. Coordination of insulin and Notch pathway activities by microRNA miR-305 mediates adaptive homeostasis in the intestinal stem cells of the Drosophila gut. Genes Dev 2014, 28, 2421–2431. [Google Scholar] [CrossRef]
- Aradhya, R.; Zmojdzian, M.; Da Ponte, J.P.; Jagla, K. Muscle niche-driven Insulin-Notch-Myc cascade reactivates dormant Adult Muscle Precursors in Drosophila. Elife 2015, 4. [Google Scholar] [CrossRef]
- Dutriaux, A.; Godart, A.; Brachet, A.; Silber, J. The insulin receptor is required for the development of the Drosophila peripheral nervous system. PLoS One 2013, 8, e71857. [Google Scholar] [CrossRef]
- Obniski, R.; Sieber, M.; Spradling, A.C. Dietary Lipids Modulate Notch Signaling and Influence Adult Intestinal Development and Metabolism in Drosophila. Dev Cell 2018, 47, 98–111. [Google Scholar] [CrossRef]
- Rutter, L.; Carrillo-Tripp, J.; Bonning, B.C.; Cook, D.; Toth, A.L.; Dolezal, A.G. Transcriptomic responses to diet quality and viral infection in Apis mellifera. BMC Genomics 2019, 20, 412. [Google Scholar] [CrossRef]
- Xu, Y.J.; Long, Q.; Fan, X.X.; Ye, Y.P.; Zhang, K.Y.; Zhang, J.X.; Zhao, H.D.; Yao, Y.T.; Fu, Z.M.; Chen, D.F.; et al. Transcriptome-Wide Characterization of piRNAs during the Developmental Process of European Honey-Bee Larval Guts. Genes (Basel) 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhang, W.; Guo, S.; Zhu, L.; Zhang, Y.; Zhao, H.; Gao, X.; Jiang, H.; Zhang, T.; Chen, D.; et al. Expression Profile, Regulatory Network, and Putative Role of microRNAs in the Developmental Process of Asian Honey Bee Larval Guts. Insects 2023, 14. [Google Scholar] [CrossRef]
- Lin, W.Y.; Yao, C.; Cheng, J.; Kao, S.T.; Tsai, F.J.; Liu, H.P. Molecular pathways related to the longevity promotion and cognitive improvement of Cistanche tubulosa in Drosophila. Phytomedicine 2017, 26, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, A.; Shaposhnikov, M.; Zemskaya, N.; Belyi, A.; Dobrovolskaya, E.; Patova, A.; Guvatova, Z.; Lukyanova, E.; Snezhkina, A.; Kudryavtseva, A. Transcriptome analysis reveals mechanisms of geroprotective effects of fucoxanthin in Drosophila. BMC Genomics 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Beghelli, D.; Zallocco, L.; Barbalace, M.C.; Paglia, S.; Strocchi, S.; Cirilli, I.; Marzano, V.; Putignani, L.; Lupidi, G.; Hrelia, S.; et al. Pterostilbene Promotes Mean Lifespan in Both Male and Female Drosophila Melanogaster Modulating Different Proteins in the Two Sexes. Oxid Med Cell Longev 2022, 2022, 1744408. [Google Scholar] [CrossRef]
- Zimmerman, O.; Holmes, A.C.; Kafai, N.M.; Adams, L.J.; Diamond, M.S. Entry receptors - the gateway to alphavirus infection. J Clin Invest 2023, 133. [Google Scholar] [CrossRef]
- Mudiganti, U.; Hernandez, R.; Brown, D.T. Insect response to alphavirus infection--establishment of alphavirus persistence in insect cells involves inhibition of viral polyprotein cleavage. Virus Res 2010, 150, 73–84. [Google Scholar] [CrossRef]
- Li, M.; Xing, D.; Su, D.; Wang, D.; Gao, H.; Lan, C.; Gu, Z.; Zhao, T.; Li, C. Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Serrato-Salas, J.; Izquierdo-Sanchez, J.; Arguello, M.; Conde, R.; Alvarado-Delgado, A.; Lanz-Mendoza, H. Aedes aegypti antiviral adaptive response against DENV-2. Dev Comp Immunol 2018, 84, 28–36. [Google Scholar] [CrossRef]
- Serrato-Salas, J.; Hernandez-Martinez, S.; Martinez-Barnetche, J.; Conde, R.; Alvarado-Delgado, A.; Zumaya-Estrada, F.; Lanz-Mendoza, H. De Novo DNA Synthesis in Aedes aegypti Midgut Cells as a Complementary Strategy to Limit Dengue Viral Replication. Front Microbiol 2018, 9, 801. [Google Scholar] [CrossRef]
- Shin, D.; Kang, S.; Smartt, C.T. Profiling Transcripts of Vector Competence between Two Different Aedes aegypti Populations in Florida. Viruses 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Taracena, M.L.; Bottino-Rojas, V.; Talyuli, O.A.C.; Walter-Nuno, A.B.; Oliveira, J.H.M.; Anglero-Rodriguez, Y.I.; Wells, M.B.; Dimopoulos, G.; Oliveira, P.L.; Paiva-Silva, G.O. Regulation of midgut cell proliferation impacts Aedes aegypti susceptibility to dengue virus. PLoS Negl Trop Dis 2018, 12, e0006498. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Garduno, J.; Rodriguez, M.C.; Hernandez-Martinez, S.; Martinez-Barnetche, J.; Alvarado-Delgado, A.; Izquierdo, J.; Herrera-Ortiz, A.; Moreno-Garcia, M.; Velazquez-Meza, M.E.; Valverde, V.; et al. Plasmodium berghei induced priming in Anopheles albimanus independently of bacterial co-infection. Dev Comp Immunol 2015, 52, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Maya-Maldonado, K.; Cardoso-Jaime, V.; Hernandez-Martinez, S.; Recio-Totoro, B.; Bello-Garcia, D.; Hernandez-Hernandez, F.C.; Lanz-Mendoza, H. Plasmodium exposure alters midgut epithelial cell dynamics during the immune memory in Anopheles albimanus. Dev Comp Immunol 2022, 133, 104424. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.L.; Yerbanga, R.S.; Lefevre, T.; Ouedraogo, J.B.; Corces, V.G.; Gomez-Diaz, E. Chromatin changes in Anopheles gambiae induced by Plasmodium falciparum infection. Epigenetics Chromatin 2019, 12, 5. [Google Scholar] [CrossRef]
- Feng, M.; Ren, F.; Zhou, Y.; Zhang, N.; Lu, Q.; Swevers, L.; Sun, J. Correlation in Expression between LTR Retrotransposons and Potential Host Cis-Targets during Infection of Antherea pernyi with ApNPV Baculovirus. Viruses 2019, 11. [Google Scholar] [CrossRef]
- Liu, X.J.; Liang, X.Y.; Guo, J.; Shi, X.K.; Merzendorfer, H.; Zhu, K.Y.; Zhang, J.Z. V-ATPase subunit a is required for survival and midgut development of Locusta migratoria. Insect Mol Biol 2022, 31, 60–72. [Google Scholar] [CrossRef]
- Zhao, B.; Lucas, K.J.; Saha, T.T.; Ha, J.; Ling, L.; Kokoza, V.A.; Roy, S.; Raikhel, A.S. MicroRNA-275 targets sarco/endoplasmic reticulum Ca2+ adenosine triphosphatase (SERCA) to control key functions in the mosquito gut. PLoS Genet 2017, 13, e1006943. [Google Scholar] [CrossRef]
- Ohlstein, B.; Spradling, A. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature 2006, 439, 470–474. [Google Scholar] [CrossRef]
- Micchelli, C.A.; Perrimon, N. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature 2006, 439, 475–479. [Google Scholar] [CrossRef]
- Ohlstein, B.; Spradling, A. Multipotent Drosophila intestinal stem cells specify daughter cell fates by differential notch signaling. Science 2007, 315, 988–992. [Google Scholar] [CrossRef]
- Jiang, H.; Patel, P.H.; Kohlmaier, A.; Grenley, M.O.; McEwen, D.G.; Edgar, B.A. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 2009, 137, 1343–1355. [Google Scholar] [CrossRef]
- Mathur, D.; Bost, A.; Driver, I.; Ohlstein, B. A transient niche regulates the specification of Drosophila intestinal stem cells. Science 2010, 327, 210–213. [Google Scholar] [CrossRef]
- Duvic, B.; Hoffmann, J.A.; Meister, M.; Royet, J. Notch signaling controls lineage specification during Drosophila larval hematopoiesis. Curr Biol 2002, 12, 1923–1927. [Google Scholar] [CrossRef]
- Lebestky, T.; Jung, S.H.; Banerjee, U. A Serrate-expressing signaling center controls Drosophila hematopoiesis. Genes Dev 2003, 17, 348–353. [Google Scholar] [CrossRef]
- Krzemien, J.; Dubois, L.; Makki, R.; Meister, M.; Vincent, A.; Crozatier, M. Control of blood cell homeostasis in Drosophila larvae by the posterior signalling centre. Nature 2007, 446, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Small, C.; Ramroop, J.; Otazo, M.; Huang, L.H.; Saleque, S.; Govind, S. An unexpected link between notch signaling and ROS in restricting the differentiation of hematopoietic progenitors in Drosophila. Genetics 2014, 197, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.H.; Choi, J.K.; Kim, B.; Cho, H.S.; Kim, J.; Kim-Ha, J.; Kim, Y.J. Requirement of Split ends for epigenetic regulation of Notch signal-dependent genes during infection-induced hemocyte differentiation. Mol Cell Biol 2009, 29, 1515–1525. [Google Scholar] [CrossRef]
- Richard, F.J.; Holt, H.L.; Grozinger, C.M. Effects of immunostimulation on social behavior, chemical communication and genome-wide gene expression in honey bee workers (Apis mellifera). BMC Genomics 2012, 13, 558. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Abd El Halim, H.M. Re-thinking adaptive immunity in the beetles: Evolutionary and functional trajectories of lncRNAs. Genomics 2020, 112, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Guichard, A.; McGillivray, S.M.; Cruz-Moreno, B.; van Sorge, N.M.; Nizet, V.; Bier, E. Anthrax toxins cooperatively inhibit endocytic recycling by the Rab11/Sec15 exocyst. Nature 2010, 467, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Guichard, A.; Cruz-Moreno, B.; Aguilar, B.; van Sorge, N.M.; Kuang, J.; Kurkciyan, A.A.; Wang, Z.; Hang, S.; Pineton de Chambrun, G.P.; McCole, D.F.; et al. Cholera toxin disrupts barrier function by inhibiting exocyst-mediated trafficking of host proteins to intestinal cell junctions. Cell Host Microbe 2013, 14, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Harsh, S.; Fu, Y.; Kenney, E.; Han, Z.; Eleftherianos, I. Zika virus non-structural protein NS4A restricts eye growth in Drosophila through regulation of JAK/STAT signaling. Dis Model Mech 2020, 13. [Google Scholar] [CrossRef]
- Alattia, J.R.; Kuraishi, T.; Dimitrov, M.; Chang, I.; Lemaitre, B.; Fraering, P.C. Mercury is a direct and potent gamma-secretase inhibitor affecting Notch processing and development in Drosophila. FASEB J 2011, 25, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Bland, C.; Rand, M.D. Methylmercury induces activation of Notch signaling. Neurotoxicology 2006, 27, 982–991. [Google Scholar] [CrossRef]
- Rand, M.D.; Bland, C.E.; Bond, J. Methylmercury activates enhancer-of-split and bearded complex genes independent of the notch receptor. Toxicol Sci 2008, 104, 163–176. [Google Scholar] [CrossRef]
- Engel, G.L.; Delwig, A.; Rand, M.D. The effects of methylmercury on Notch signaling during embryonic neural development in Drosophila melanogaster. Toxicol In Vitro 2012, 26, 485–492. [Google Scholar] [CrossRef]
- Engel, G.L.; Rand, M.D. The Notch target E(spl)mdelta is a muscle-specific gene involved in methylmercury toxicity in motor neuron development. Neurotoxicol Teratol 2014, 43, 11–18. [Google Scholar] [CrossRef]
- Yan, S.; Yin, H.; Li, N.; Chen, Y.; Ji, C.D.; Jiang, Q.H.; Du, J.; Yin, M.Z.; Shen, J.; Zhang, J.Z. Combination of a nanocarrier delivery system with genetic manipulation further improves pesticide efficiency: a case study with chlorfenapyr. Environ Sci-Nano 2022, 9, 2020–2031. [Google Scholar] [CrossRef]
- Moskalev, A.; Shaposhnikov, M.; Snezhkina, A.; Kogan, V.; Plyusnina, E.; Peregudova, D.; Melnikova, N.; Uroshlev, L.; Mylnikov, S.; Dmitriev, A.; et al. Mining gene expression data for pollutants (dioxin, toluene, formaldehyde) and low dose of gamma-irradiation. PLoS One 2014, 9, e86051. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, H.; Jiang, Z.; Zhang, J.; Sun, Z.; Zhong, G. Natural harmine negatively regulates the developmental signaling network of Drosophila melanogaster (Drosophilidae: Diptera) in vivo. Ecotoxicol Environ Saf 2020, 190, 110134. [Google Scholar] [CrossRef] [PubMed]
- Farder-Gomes, C.F.; Fernandes, K.M.; Bernardes, R.C.; Bastos, D.S.S.; Oliveira, L.L.; Martins, G.F.; Serrao, J.E. Harmful effects of fipronil exposure on the behavior and brain of the stingless bee Partamona helleri Friese (Hymenoptera: Meliponini). Sci Total Environ 2021, 794, 148678. [Google Scholar] [CrossRef] [PubMed]
- Farder-Gomes, C.F.; Santos, A.A.; Fernandes, K.M.; Bernardes, R.C.; Martins, G.F.; Serrao, J.E. Fipronil exposure compromises respiration and damages the Malpighian tubules of the stingless bee Partamona helleri Friese (Hymenoptera: Apidae). Environ Sci Pollut Res Int 2022, 29, 88101–88108. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jia, Z.Q.; Peng, Y.C.; Sheng, C.W.; Tang, T.; Xu, L.; Han, Z.J.; Zhao, C.Q. Toxicity and sublethal effects of fluralaner on Spodoptera litura Fabricius (Lepidoptera: Noctuidae). Pestic Biochem Physiol 2018, 152, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Nojima, Y.; Bono, H.; Yokoyama, T.; Iwabuchi, K.; Sato, R.; Arai, K.; Tabunoki, H. Superoxide dismutase down-regulation and the oxidative stress is required to initiate pupation in Bombyx mori. Sci Rep 2019, 9, 14693. [Google Scholar] [CrossRef]
- Harrison, J.F.; Greenlee, K.J.; Verberk, W. Functional Hypoxia in Insects: Definition, Assessment, and Consequences for Physiology, Ecology, and Evolution. Annu Rev Entomol 2018, 63, 303–325. [Google Scholar] [CrossRef]
- Zhou, D.; Xue, J.; Lai, J.C.; Schork, N.J.; White, K.P.; Haddad, G.G. Mechanisms underlying hypoxia tolerance in Drosophila melanogaster: hairy as a metabolic switch. PLoS Genet 2008, 4, e1000221. [Google Scholar] [CrossRef]
- Zhou, D.; Udpa, N.; Gersten, M.; Visk, D.W.; Bashir, A.; Xue, J.; Frazer, K.A.; Posakony, J.W.; Subramaniam, S.; Bafna, V.; et al. Experimental selection of hypoxia-tolerant Drosophila melanogaster. Proc Natl Acad Sci U S A 2011, 108, 2349–2354. [Google Scholar] [CrossRef]
- Azad, P.; Zhou, D.; Zarndt, R.; Haddad, G.G. Identification of genes underlying hypoxia tolerance in Drosophila by a P-element screen. G3 (Bethesda) 2012, 2, 1169–1178. [Google Scholar] [CrossRef]
- Ronen, R.; Udpa, N.; Halperin, E.; Bafna, V. Learning natural selection from the site frequency spectrum. Genetics 2013, 195, 181–193. [Google Scholar] [CrossRef]
- Zhou, D.; Stobdan, T.; Visk, D.; Xue, J.; Haddad, G.G. Genetic interactions regulate hypoxia tolerance conferred by activating Notch in excitatory amino acid transporter 1-positive glial cells in Drosophila melanogaster. G3 (Bethesda) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Linz, D.M.; Parker, E.S.; Schwab, D.B.; Casasa, S.; Macagno, A.L.M.; Moczek, A.P. Developmental bias in horned dung beetles and its contributions to innovation, adaptation, and resilience. Evol Dev 2020, 22, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Linz, D.M.; Moczek, A.P. Beetle horns evolved from wing serial homologs. Science 2019, 366, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Kijimoto, T.; Costello, J.; Tang, Z.; Moczek, A.P.; Andrews, J. EST and microarray analysis of horn development in Onthophagus beetles. BMC Genomics 2009, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kijimoto, T.; Snell-Rood, E.; Tae, H.; Yang, Y.; Moczek, A.P.; Andrews, J. Gene discovery in the horned beetle Onthophagus taurus. BMC Genomics 2010, 11, 703. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.R.; Macagno, A.L.M.; Moczek, A.P.; Rohner, P.T.; Hu, Y. Notch signaling patterns head horn shape in the bull-headed dung beetle Onthophagus taurus. Dev Genes Evol 2020, 230, 213–225. [Google Scholar] [CrossRef]
- Adachi, H.; Matsuda, K.; Niimi, T.; Kondo, S.; Gotoh, H. Genetical control of 2D pattern and depth of the primordial furrow that prefigures 3D shape of the rhinoceros beetle horn. Sci Rep 2020, 10, 18687. [Google Scholar] [CrossRef]
- Hu, Y.; Moczek, A.P. Wing serial homologues and the diversification of insect outgrowths: insights from the pupae of scarab beetles. Proc Biol Sci 2021, 288, 20202828. [Google Scholar] [CrossRef]
- Hu, Y.; Schmitt-Engel, C.; Schwirz, J.; Stroehlein, N.; Richter, T.; Majumdar, U.; Bucher, G. A morphological novelty evolved by co-option of a reduced gene regulatory network and gene recruitment in a beetle. Proc Biol Sci 2018, 285. [Google Scholar] [CrossRef]
- Linz, D.M.; Hu, Y.; Moczek, A.P. The origins of novelty from within the confines of homology: the developmental evolution of the digging tibia of dung beetles. Proc Biol Sci 2019, 286, 20182427. [Google Scholar] [CrossRef]
- Elgar, M.A.; Zhang, D.; Wang, Q.; Wittwer, B.; Thi Pham, H.; Johnson, T.L.; Freelance, C.B.; Coquilleau, M. Insect Antennal Morphology: The Evolution of Diverse Solutions to Odorant Perception. Yale J Biol Med 2018, 91, 457–469. [Google Scholar] [PubMed]
- Kenyon, K.L.; Ranade, S.S.; Curtiss, J.; Mlodzik, M.; Pignoni, F. Coordinating proliferation and tissue specification to promote regional identity in the Drosophila head. Dev Cell 2003, 5, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Ku, H.Y.; Sun, Y.H. Notch-dependent epithelial fold determines boundary formation between developmental fields in the Drosophila antenna. PLoS Genet 2017, 13, e1006898. [Google Scholar] [CrossRef]
- Zhu, J.; Palliyil, S.; Ran, C.; Kumar, J.P. Drosophila Pax6 promotes development of the entire eye-antennal disc, thereby ensuring proper adult head formation. Proc Natl Acad Sci U S A 2017, 114, 5846–5853. [Google Scholar] [CrossRef] [PubMed]
- Angelini, D.R.; Kikuchi, M.; Jockusch, E.L. Genetic patterning in the adult capitate antenna of the beetle Tribolium castaneum. Dev Biol 2009, 327, 240–251. [Google Scholar] [CrossRef]
- Ando, T.; Kojima, T.; Fujiwara, H. Dramatic changes in patterning gene expression during metamorphosis are associated with the formation of a feather-like antenna by the silk moth, Bombyx mori. Dev Biol 2011, 357, 53–63. [Google Scholar] [CrossRef]
- Ando, T.; Fujiwara, H.; Kojima, T. The pivotal role of aristaless in development and evolution of diverse antennal morphologies in moths and butterflies. BMC Evol Biol 2018, 18, 8. [Google Scholar] [CrossRef]
- Smith, F.W.; Angelini, D.R.; Gaudio, M.S.; Jockusch, E.L. Metamorphic labral axis patterning in the beetle Tribolium castaneum requires multiple upstream, but few downstream, genes in the appendage patterning network. Evol Dev 2014, 16, 78–91. [Google Scholar] [CrossRef]
- Tong, X.; Qiao, L.; Luo, J.; Ding, X.; Wu, S. The evolution and genetics of lepidopteran egg and caterpillar coloration. Curr Opin Genet Dev 2021, 69, 140–146. [Google Scholar] [CrossRef]
- Futahashi, R.; Fujiwara, H. Juvenile hormone regulates butterfly larval pattern switches. Science 2008, 319, 1061. [Google Scholar] [CrossRef]
- Jin, H.; Seki, T.; Yamaguchi, J.; Fujiwara, H. Prepatterning of Papilio xuthus caterpillar camouflage is controlled by three homeobox genes: clawless, abdominal-A, and Abdominal-B. Sci Adv 2019, 5, eaav7569. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Yoda, S.; Liu, L.; Kojima, T.; Fujiwara, H. Notch and Delta Control the Switch and Formation of Camouflage Patterns in Caterpillars. iScience 2020, 23, 101315. [Google Scholar] [CrossRef]
- Patel, N.H. The ancestry of segmentation. Dev Cell 2003, 5, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Truman, J.W. The Evolution of Insect Metamorphosis. Curr Biol 2019, 29, R1252–R1268. [Google Scholar] [CrossRef]
- Mysore, K.; Sun, L.; Hapairai, L.K.; Wang, C.W.; Roethele, J.B.; Igiede, J.; Scheel, M.P.; Scheel, N.D.; Li, P.; Wei, N.; et al. A Broad-Based Mosquito Yeast Interfering RNA Pesticide Targeting Rbfox1 Represses Notch Signaling and Kills Both Larvae and Adult Mosquitoes. Pathogens 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.M.; Choo, A.; Baxter, S.W. Lessons from Drosophila: Engineering Genetic Sexing Strains with Temperature-Sensitive Lethality for Sterile Insect Technique Applications. Insects 2021, 12. [Google Scholar] [CrossRef]
- Akbari, O.S.; Chen, C.H.; Marshall, J.M.; Huang, H.; Antoshechkin, I.; Hay, B.A. Novel synthetic Medea selfish genetic elements drive population replacement in Drosophila; a theoretical exploration of Medea-dependent population suppression. ACS Synth Biol 2014, 3, 915–928. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A simplified model of the Notch signaling transduction cascade in the fruit fly. The signal sending cell is in blue and the signal receiving cell is in earthy yellow. Upon binding with the Delta/Serrate ligands, the Notch receptor sequentially processed by metalloproteases of the ADAM/TACE family and the γ-secretase, resulting the release of NICD from the membrane. In the nucleus, NICD assembles with the CSL family protein Su(H) and the co-activator Mam to form a complex that regulates target gene expression.
Figure 1.
A simplified model of the Notch signaling transduction cascade in the fruit fly. The signal sending cell is in blue and the signal receiving cell is in earthy yellow. Upon binding with the Delta/Serrate ligands, the Notch receptor sequentially processed by metalloproteases of the ADAM/TACE family and the γ-secretase, resulting the release of NICD from the membrane. In the nucleus, NICD assembles with the CSL family protein Su(H) and the co-activator Mam to form a complex that regulates target gene expression.
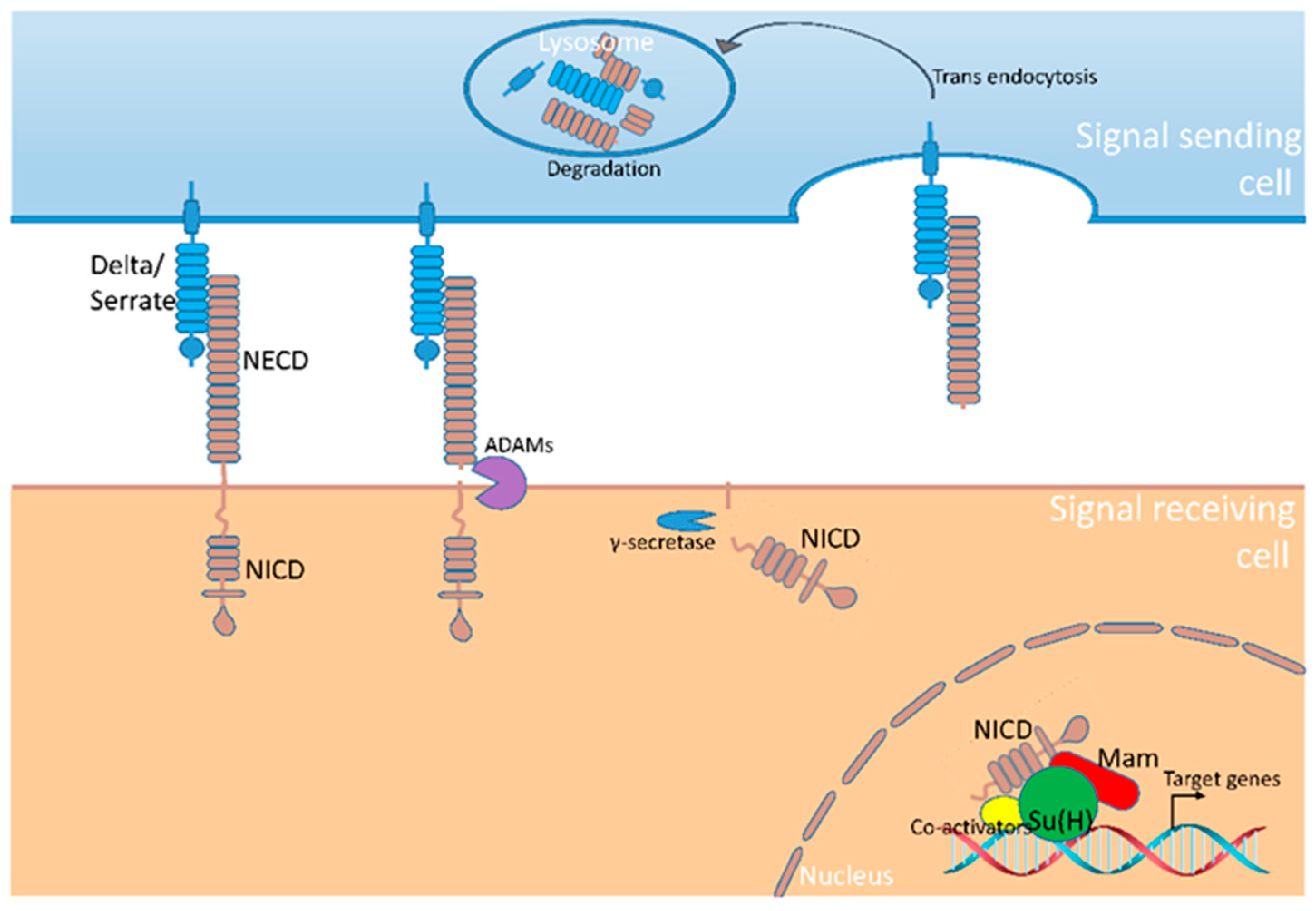
Figure 2.
Notch signaling is highly conserved among insects. The cladogram shows that homologs of Notch, Delta and Serrate are found in major groups of insects (red dots).
Figure 2.
Notch signaling is highly conserved among insects. The cladogram shows that homologs of Notch, Delta and Serrate are found in major groups of insects (red dots).
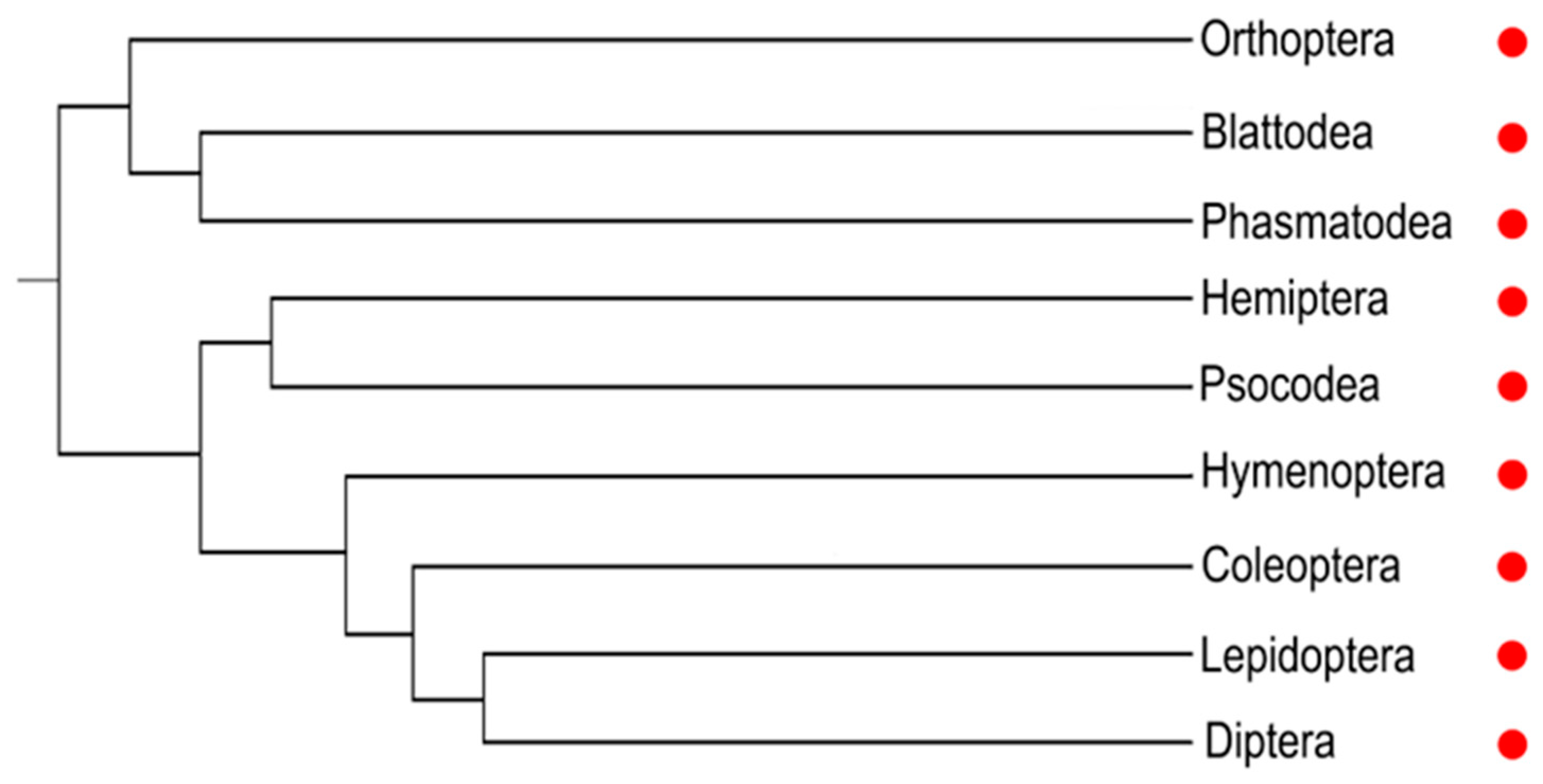
Figure 3.
Roles of Notch signaling in insect development. The Notch signaling pathway primarily functions through three major modes, including lateral inhibition, boundary formation, and lineage determination. Widely present in major insect taxa, Notch signaling plays a crucial role in the development of embryos, wings, legs, ovaries and antennae, as well as in the insect's response to stress.
Figure 3.
Roles of Notch signaling in insect development. The Notch signaling pathway primarily functions through three major modes, including lateral inhibition, boundary formation, and lineage determination. Widely present in major insect taxa, Notch signaling plays a crucial role in the development of embryos, wings, legs, ovaries and antennae, as well as in the insect's response to stress.
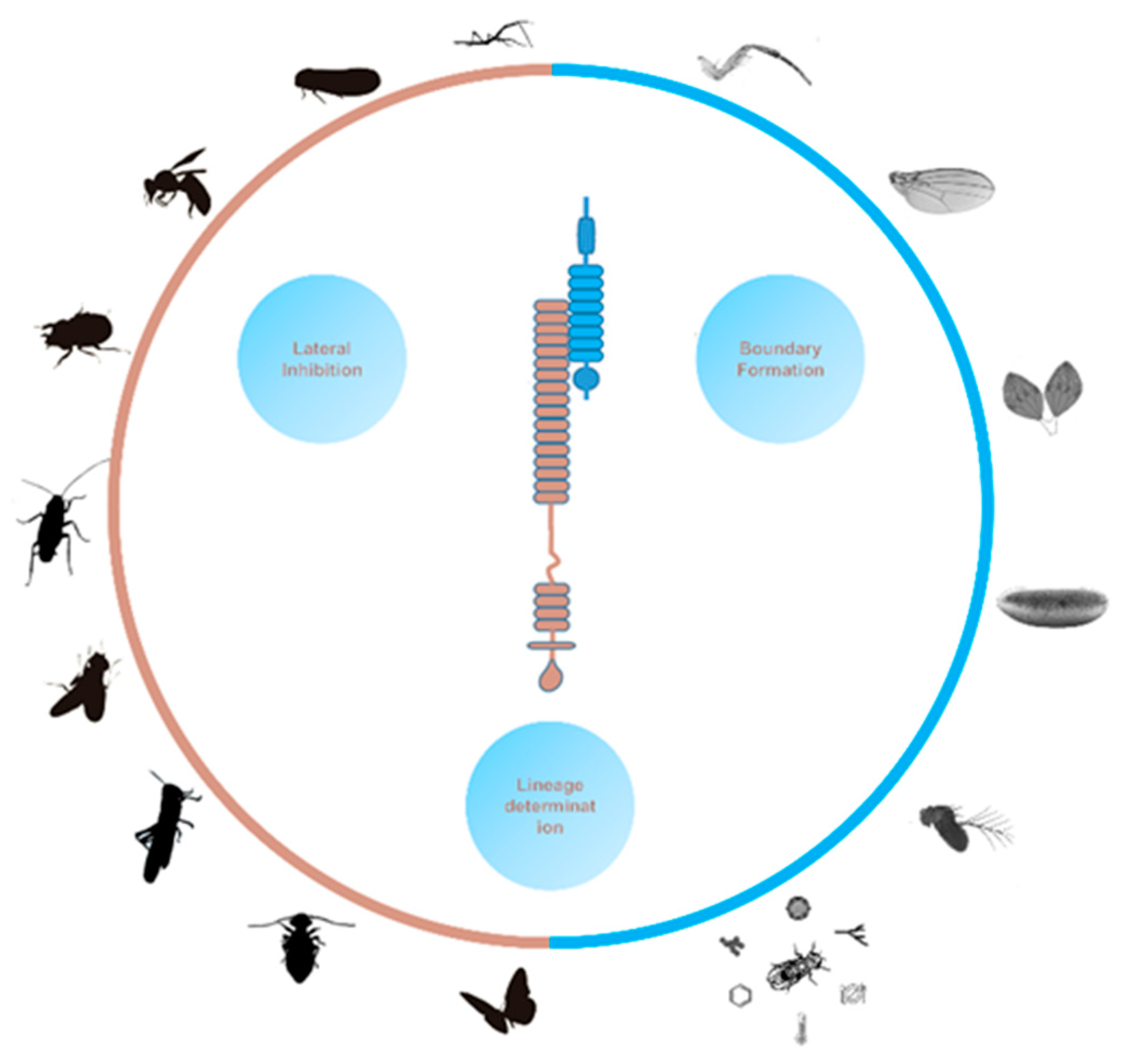
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



