
Submitted:
03 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
Unlike trees, shrubs (i.e., multiple-stemmed woody plants) do not need evenly spaced large diameter structural roots and thus can spread further per unit belowground biomass. We therefore hypothesized that compared to trees, shrubs respond more to asymmetric distributions of nutrients and reach nutrient-rich patches of soil faster and with less below-ground biomass. To test these hypotheses we planted individual seedlings of shrubs (Cornus racemosa, Rhus glabra, and Viburnum dentatum) and trees (Acer rubrum, Betula populifera, and Fraxinus americana) in the centers of sand-filled rectangular boxes. In one direction we created stepwise gradients of increasing soil nutrients with slow-release fertilizer; in the other direction no fertilizer was added. Seedlings were harvested when their first root reached the plexiglass-covered fertilized end of their box; time taken as well as above- and below-ground biomass by nutrient segment were determined. There were no consistent differences between the shrubs and trees in belowground architectural plasticity (= ratio of biomass in fertilized and unfertilized volumes of soil) or in the rate and efficiency of lateral growth. Interspecific variation appeared more related to edaphic characteristics of the native habitats than to growth form. The fastest and most efficient roots were produced by the shrub (R. glabra) and the tree (B. populifolia) that are characteristic of poor soils. Root foraging by R. glabra was also facilitated by rapid rhizomatous expansion. The tested shrubs and trees did not consistently differ in nutrient foraging efficiencies or rates but further study is warranted with larger plants and more species.
Keywords:
environmental heterogeneity
; functional traits
; root architecture
; root foraging
; vegetative expansion
1. Introduction
Where soils are temporarily heterogeneous in nutrient availability, competitive benefits are likely to accrue to plants that locate fertile patches rapidly. In an experiment similar to the one described herein, liana roots were shown to consistently reach nutrient baits more quickly than tree roots [1]. The explanation offered for this result is that because lianas do not need to mechanically support their stems, they do not invest in large diameter roots and hence can efficiently and rapidly explore soil. This finding may help explain why per unit biomass, lianas often outcompete trees for below-ground resources [2]. Here we test whether shrub roots are similarly efficient and rapid in their soil exploration. Like lianas, shrubs are often strong competitors with trees especially on nutrient-poor and drought-prone sites [3]. Also like lianas, most shrub stems are slender [4] and do not need large diameter structural roots [5] nor do they do they need to evenly distribute roots around their stems for biomechanical purposes; these characteristics should increase the soil exploration capacities of shrubs per unit biomass invested below-ground. Finally, vegetative production of new stems by shrubs may depend on prior root or rhizome expansion, thus providing a further benefit for rapid and efficient lateral growth below-ground.
2. Materials and Methods
This experiment was conducted with one year-old nursery-grown seedlings in a greenhouse in Millbrook, New York (41°50′N, 73°45′W). Randomly selected bare root seedlings of trees (Acer rubrum, Betula populifera, and Fraxinus americana) and shrubs (Cornus racemosa, Rhus glabra, and Viburnum dentatum) acquired at a local nursery (N = 8 per species) were planted in early June in the centers of 60 x 15 x 40 cm (length x breadth x depth) plywood boxes filled with washed sand over a 2 cm thick layer of coarse quartz gravel (Figure 1). One end of each box was made from plexiglass that was kept dark other than when checking for the presence of roots. Between each seedling and the root observation window we created a four-step gradient of increasing nutrient availability by adding increasing amounts of a complete nutrient slow-release fertilizer (Sierra Chemical Company, California) to the sand when the boxes were being filled. Starting 6 cm from the plant the 6 cm wide (3600 cm3) bands of sand contained 1, 2, 4, and 8 g of fertilizer which corresponds with 7.5, 15, 30, and 60% of the recommended quantity for sensitive nursery stock. In the opposite direction we added no fertilizer. The plants were irrigated twice weekly or as needed.
Plants were grown until the first root reached the observation window (range = 49-182 d) at which time the entire plant was harvested. Final sample sizes ranged from 4-8 plants per species due to mortality. Roots were harvested by section from the soil fertility gradient by cutting down through the soil with a sharpened blade. Roots were removed from the soil by hand followed by washing in a Gillison Hydropneumatic Elutriation System. Roots and shoots were then dried to constant weight at 80 °C and weighed. Initial plant biomass and root:shoot biomass ratios were estimated using eight randomly selected individuals of each species from the original planting stock.
Differences between trees and shrubs were examined using analysis of variance (PROC GLM in SAS) [6], with species nested within growth form. Because species are the true level of replication for tests of the hypothesis that 2 growth forms differ in root growth and architecture, F statistics for effects of growth form were computed using the growth form * species interaction term as the error mean square. Since the effects of growth form were not significant for any of the dependent variables, further analyses were conducted to test for differences among the 6 species combined (simple 1-way analysis of variance, with species as a random effect, using PROC GLM in SAS). Pairwise differences in species means were tested using Tukey’s Studentized Range Test at P < 0.05 to control experiment-wise error [6].
The species used in this experiment overlap considerably in ecological ranges and are frequently sympatric in southern New England. Cornus racemosa Lam. is a clonal shrub that grows to be 2-3 m tall and forms dense, laterally spreading clumps. Viburnum dentatum L. is similar in growth form to C. racemosa but generally does not grow to be as large or dense; both species are characteristic of mesic, nutrient-rich, alluvial soils, but C. racemosa also occurs on poorer sites. Rhus glabra L., the third shrub species in the experiment, forms larger and more diffuse clumps in drier and more nutrient-poor sites than the other two species. All three tree species used (Acer rubrum L., Fraxinus americana L., and Betula populifolia Marsh) are fairly light demanding and geographically widespread, but B. populifolia is particularly characteristic of rocky areas with nutrient-poor and severely drained soils.
3. Results and Discussion
Contrary to our predictions there were no consistent differences between the shrubs and trees in root foraging efficiencies, root growth rates, or root:shoot ratios (Table 1, Figure 2). All six species generally produced more roots in the fertilized than in the unfertilized portions of the planting frames (Figure 2a), but only R. glabra showed this tendency to a significant degree. Lateral growth efficiency (maximum root growth distance/root biomass), another expression of root foraging efficiency, was quite variable among species (Figure 2b) but R. glabra was again the most efficient; the other two shrub species were markedly less efficient than any of the tree species tested. Among the trees, Acer rubrum roots reached the nutrient-rich end of the planting frames with smaller investments in root biomass than observed in the other two species (Figure 2b). R. glabra and B. populifolia grew most rapidly up the nutrient gradient (Figure 2c) and grew roots the fastest (Figure 2d); roots of four R. glabra and two B. populifolia seedlings reached the observation window within about three weeks after transplanting. Shoot growth rates by R. glabra were markedly higher than in any other tree or shrub species (Figure 2e). Among the three shrubs tested, the species with the highest root foraging efficiency and the most rapid lateral root growth rate (R. glabra) also had the lowest root/shoot ratio (Figure 2f). In contrast, a tree with rapid root growth toward the nutrient-rich bait (B. populifolia) had a high mean root/shoot ratio compared with the other tree species tested. This suggests that root-shoot ratio is not necessarily a good predictor of root foraging efficiency.
Root systems of species with high rates of growth along the soil fertility gradient (e.g., R. glabra and B. populifolia) tended not to proliferate in low fertility zones of soil (Figure 2a). This explanation for differences in root growth efficiency is not sufficient, however, to explain all the species differences observed. A. rubrum, for example, showed high in lateral root growth efficiency (Figure 2b) but fairly low growth rates up the nutrient gradient (Figure 2c) at least partially due to low root growth rates (Figure 2d).
Although our study was not designed to test hypotheses relating rooting habits to characteristics of the species’ natural habitats, the results do not support the idea that the root systems of plants from nutrient-rich sites are architecturally more plastic and more actively forage for nutrients than plants from infertile sites [7]. For example, R. glabra, the species that expanded the fastest below-ground with the most architectural plasticity and highest efficiency is characteristically found on infertile, rocky, and otherwise poor soils. Similarly, B. populifolia, the tree species with the most rapid root growth rates up the nutrient gradient (Figure 2c), is common on degraded abandoned farmland and on rocky outcrops where it roots in small pockets of soil that collect in spatially isolated cracks and crevices. In a related study of soil resource heterogeneity in the study region, Kelly and Canham [8] found the highest spatial variation in sites with the lowest average resource availability. This suggests that root foraging efficiency will be most beneficial for species that typically occur on poor and spatially heterogeneous soils. Thus, the apparent inefficiencies in root foraging of the shrubs V. dentatum and C. racemose and the trees F. americana and A. rubrum may be related to their occurrence in more fertile and relatively homogeneous alluvial soils.
Complicating interpretation of the main findings of this study that shrubs and trees do not differ in nutrient foraging speeds or efficiencies are other, often size-dependent contributions to of below-ground structures to plant survival and growth. For example, in addition to discovery of nutrient patches, rhizomes facilitate extensive clonal expansion by R. glabra, one of the traits that renders a similar species of Rhus (R. typhina) invasive where introduced in China [9]. Also, because we worked only with seedlings we missed any shrub-tree differences that may become apparent when the latter grow large stems and the former stay small.
Author Contributions
E.P. and C.D.C. conceptualized the study, S.V.O. assisted with data collection, C.D.C. analyzed the data, F.E.P. wrote the first draft that was revised by C.D.C. and S.V.O.
Funding
This work was supported by the Central Hudson Gas and Electric Corporation, the Empire State Electric Energy Research Corporation, NSF Grant BSR-902003, NSF Grant DEB-9220620, and the Mary Flagler Cary Charitable Trust.
Data Availability Statement
Upon acceptance for publication, the data will be published by the Environmental Data Initiative (EDI).
Acknowledgments
We thank R. Ostertag and P.J. Burton for comments on an earlier version of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Putz, F.E. Climbing plants beat trees to soil nutrient patches. Current Biology 2023, 33, R659–R676. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Aceves, T. Above- and belowground competition between lianas and trees. In Ecology of Lianas; Schnitzer, S.A., Bongers, F., Burnham, R.J., Putz, F.E., Eds.; John Wiley and Sons: Hoboken, New Jersey, USA, 2015. [Google Scholar]
- Putz, F.E.; Canham, C.D. Mechanisms of arrested succession in shrublands: root and shoot competition between shrubs and tree seedlings. Forest Ecology and Management 1992, 49, 267–275. [Google Scholar] [CrossRef]
- Larjavaara, M. Trees and shrubs differ biomechanically. Trends in Ecology and Evolution 2015, 30, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Götmark, F.; Götmark, E.; Jensen, A.M. Why be a shrub? A basic model and hypotheses for the adaptive values of a common growth form. Frontiers in Plant Science 2016, 1095. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute, Inc. SAS/STAT Guide for Personal Computers; Version 6 Edition; SAS Institute, Inc.: Cary, North Carolina, USA, 1987. [Google Scholar]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley: Chichester, United Kingdom, 1979. [Google Scholar]
- Kelly, V.R.; Canham, C.D. Resource heterogeneity in oldfields. Journal of Vegetation Science 1992, 3, 545–552. [Google Scholar] [CrossRef]
- Wang, G.; Jiang, G.; Yu, S.; Li, Y.; Liu, H. Invasion possibility and potential effects of Rhus typhina on Beijing municipality. Journal of Integrative Plant Biology 2008, 50, 522–530. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The experimental planting boxes with nutrient gradients after several plants reached the plexiglass-covered end wall and were harvested.
Figure 1.
The experimental planting boxes with nutrient gradients after several plants reached the plexiglass-covered end wall and were harvested.

Figure 2.
Growth characteristics of seedlings planted in the middles of sand-filled boxes with no fertilizer added to one side and a gradient of increasing fertilizer added to the other (N = 4-8 per species). Error bars are ± 1 S. E. of the mean. Species with different letters above the bars are significantly different at p < 0.05 using Tukey’s Studentized Range Test. (A). Ratio of root plus rhizome biomass in the fertilized and unfertilized sides of the planting boxes. (B). Maximum lateral extension of the root system (cm away from the above-ground stem) per gram of root tissue in the fertilized side of the boxes. (C). Daily below-ground growth rates (cm/day) up the fertility gradient. (D). Total below-ground growth rate (g/day). (E). Daily above-ground growth rate (g/day). (F). The ratio of below-ground to above-ground biomass (including initial biomass). The three species in the left half of each box are trees (FRAM = Fraxinus americana, ACRU = Acer rubrum, BEPO = Betula populifolia), while the 3 species in the right half of each figure are shrubs (VIDE = Viburnum dentatum, CORA = Cornus racemosa, and RHGL = Rhus glabra).
Figure 2.
Growth characteristics of seedlings planted in the middles of sand-filled boxes with no fertilizer added to one side and a gradient of increasing fertilizer added to the other (N = 4-8 per species). Error bars are ± 1 S. E. of the mean. Species with different letters above the bars are significantly different at p < 0.05 using Tukey’s Studentized Range Test. (A). Ratio of root plus rhizome biomass in the fertilized and unfertilized sides of the planting boxes. (B). Maximum lateral extension of the root system (cm away from the above-ground stem) per gram of root tissue in the fertilized side of the boxes. (C). Daily below-ground growth rates (cm/day) up the fertility gradient. (D). Total below-ground growth rate (g/day). (E). Daily above-ground growth rate (g/day). (F). The ratio of below-ground to above-ground biomass (including initial biomass). The three species in the left half of each box are trees (FRAM = Fraxinus americana, ACRU = Acer rubrum, BEPO = Betula populifolia), while the 3 species in the right half of each figure are shrubs (VIDE = Viburnum dentatum, CORA = Cornus racemosa, and RHGL = Rhus glabra).
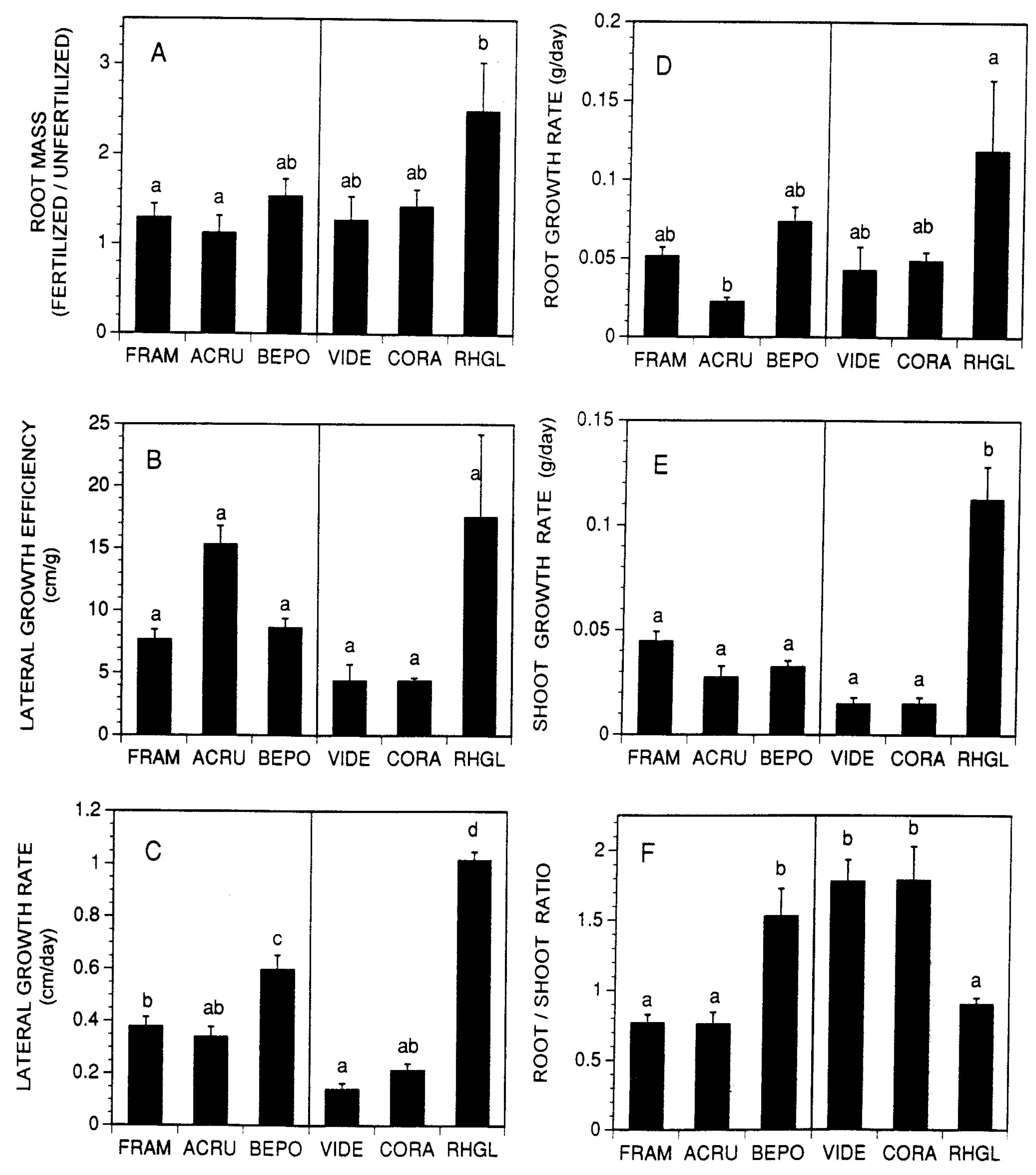

Table 1.
Results of 1-way ANOVAs on root and shoot growth characteristics of species of trees and shrubs (without regard to growth form).
Table 1.
Results of 1-way ANOVAs on root and shoot growth characteristics of species of trees and shrubs (without regard to growth form).
| Dependent Variable | df | F | p |
|---|---|---|---|
| Root Mass: Fertilized/Unfertilized (g/g) | 5.31 | 3.02 | 0.025 |
| Lateral Root Growth Efficiency (cm/g root) | 5.31 | 3.46 | 0.013 |
| Lateral Root Growth Rate (cm/day) | 5.32 | 53.00 | <0.001 |
| Root Growth Rate (g/day) | 5.31 | 2.98 | 0.026 |
| Shoot Growth Rate (g/day) | 5.32 | 20.46 | <0.001 |
| Root/Shoot Ratio (g/g) | 5.31 | 11.70 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



