
Submitted:
02 August 2023
Posted:
04 August 2023
You are already at the latest version
Abstract
Detecting the trends of species and populations is fundamental to identify taxa with high conservation priority. Unfortunately, long-term monitoring programs are challenging and often lacking. The Italian agile frog Rana latastei is endemic to Northern Italy and adjacent countries, is considered vulnerable by the IUCN, and is protected at the European level. However, quantitative estimates of its decline are extremely scarce. In this study, we document the trends in abundance and distribution of Rana latastei within the Monza Park, which currently represents the area closer to the type locality of the species. Wetlands within the park were monitored from 2000 to 2023; counts of egg clutches were taken as a measure of reproductive output and of the abundance of breeding females. In 2000, the species occurred over a significant proportion of the park. Total abundance showed strong yearly variation but remained rather constant from 2000 to 2019. However, Rana latastei disappeared from the park around 2021 and was never detected in 2022-23. The decline is probably related to the joint effect of multiple factors, including the conversion of breeding sites for farming, inappropriate water management, invasive alien species, and severe drought. The local extinction of Rana latastei occurred despite legal protection, highlighting the need for more effective and stringent tools for the conservation of European biodiversity.
Keywords:
Global amphibian decline
; climate change
; drought
; invasive alien species
; Procambarus clarkii
; EU Habitat Directive
; amphibian conservation
1. Introduction
The dramatic global decline of amphibians has been detected more than 30 years ago now [1,2,3], with amphibians consistently indicated as the animals suffering the strongest declines and with the highest proportion of threatened species [4]. Despite multiple calls and a growing number of ongoing conservation actions, we have been so far unable to halt the global amphibian crisis.
Documenting amphibian declines and extinction is pivotal to identify drivers and conservation priorities. However, ascertaining declines and extinctions is difficult, as they can be confused with natural oscillations of populations [5,6]. For instance, the Costa Rica golden toad, Incilius periglenes, was last observed in 1989. Nevertheless, uncertainty persisted regarding the rate and timing of the observed decline, and several years were required to understand that its extinction was most likely determined by complex interactions between climate change and other stressors [7,8,9,10]. In 1996, eight years after the last sighting and despite huge monitoring efforts, the golden toad was still considered "critically endangered" by the IUCN. Only in 2004 the golden toad was officially listed as "extinct" [11], after 15 years of regular quantitative monitoring without any detection [10]. Regular, long-term monitoring programs are thus necessary to document demographic trends and propose management actions. Unfortunately, implementing and maintaining extensive and long-term monitoring is often challenging, making them jeopardized and rarely covering significant portion of the species range.
The Italian agile frog Rana latastei is endemic to Northern Italy and adjacent countries, and is classified as "vulnerable" by the IUCN because it is threatened by the joint effect of habitat loss and fragmentation, pollution, and interactions with invasive predators [12,13]. The type locality of the Italian agile frog is “around Milan”, in Northern Italy [14], but this area currently is one of the most heavily human-dominated regions of Europe, with very few remaining natural habitats and wetlands potentially suitable for amphibians. Amphibians inhabiting this area are surveyed since the 1990’s [15,16,17,18], but it is only since 2009 that broad-scale, regular monitoring is ongoing [19,20,21]. Information from the whole amphibian community suggest an increase in environmental pressure that can negatively affect fitness [19,20,21,22,23]. Nevertheless, so far, no quantitative estimates of long-term population trends are available for the natural areas closest to Milan city [but see 23].
The aim of this study was to assess the long-term trend of the Italian agile frog in a protected area of Northern Italy: the Monza Park (southern portion of the Lambro Valley Regional Park) (Figure 1). This is the area hosting the R. latastei population that is closest to the type locality of the species (i.e., Milan). By combining data from more than 20 years of surveys, we quantify the dramatic decline and the possible local extinction of the species despite its legal protection [24].
2. Methods
Study area and study species
The Monza Park is located in Northern Italy, in the Lombardy region (45.35N, 9.16E; Figure 1) and is a 750-ha urban park within the Lambro Valley Regional Park. The area (altitude: 162–200 m a.s.l.) is covered by fragmented forests and meadows. This park is known to host significant populations of the Italian agile frog, Rana latastei. As its boundary is approx. 10-km N of the municipality of Milan, it should represent the area closest to the type locality of the species [14,25].
The Italian agile frog is a small frog species that exhibits explosive breeding behavior. Mating season generally lasts a few days, typically spanning from mid-February to mid-April, depending on rainfall patterns and temperature regime [25]. Each female produces a single egg clutch per year (approx. 1200 eggs per clutch) [25,26,27]. Clutches are globular and are deposited onto submerged woods and tree roots within 1 m of the water surface [25]. Therefore, clutches can be easily spotted and recognized and their number corresponds to the number of breeding females in a population. Detection probability analyses showed that the detection probability of clutches is >95%; therefore, clutch counts enable reliable estimates of population trends [28]. Preferred sites for egg-laying are perennial and slow-flowing waterbodies, where hatching typically occurs within 2 or 3 weeks, and is followed by a relatively long tadpole stage which, depending on water conditions, lasts for at least three months [25]. Despite the small range, several populations have developed local adaptations to local environmental conditions (e.g. climate, presence of predators), thus some isolated populations have been proposed as evolutionary significant units, deserving special conservation attention [17,26,29].
Field monitoring and data analyses
Starting in 2000, we monitored seven freshwater sites (ponds and ditches with slow streams) in the Monza Park (Figure 1) to assess the abundance of breeding R. latastei females by counting the number of egg clutches laid during the reproductive season. Field surveys were conducted in ten different years, from 2000 to 2023: 2000, 2001, 2004, 2005, 2006, 2011, 2017, 2019, 2022, and 2023. Surveys were conducted during daytime during the peak of the breeding season of the Italian agile frog (February-April), in order to perform visual counts of the egg clutches present at each site, following Dalpasso, Ficetola, Giachello, Lo Parrino, Manenti, Muraro and Falaschi [23]. For each year, each site was surveyed multiple times (range: 3–7) between February and April. During surveys, we carefully inspected the whole surface of sites and counted the number of egg clutches seen. As we performed multiple clutch counts in the same year, we reported the highest count for each breeding site, which represents the minimum number of breeding females in each wetland [28,30]. The minimum yearly abundance was then calculated as the sum of the highest count across all sites within a given year.
Estimating decline in species abundance
Several approaches can be used to assess species decline. For instance, the IUCN allows to assess declines on the basis of the decline in the number of individuals, or in the area of occupancy and extent of occurrence of the species [31].
First, we used the number of clutches as a measure of the abundance of breeding females [28,30,32]. Amphibian populations can undergo large fluctuations in abundance over time, due to both deterministic and stochastic reasons [6,33]. Hence, population abundance in a single year may bear little information about long-term trends. Therefore, to obtain a more general perspective on the abundance of Italian agile frogs over time, we aggregated the counts on a base of 4-year periods. The average longevity of females for this species is about 3 years [34], thus using 4-year periods allows to limit potential fluctuations deriving from some females skipping reproduction in certain years (e.g. particularly dry periods). Samples were aggregated as follows: 2000/2003, 2004/2007, 2016/2019, 2020/2023; surveys were also performed during 2011, but during this year we did not follow the standard protocol for counting clutches. Therefore, for 2011 we only report the occurrence of clutches, without providing estimates of the overall abundance of the species. During 2020-21 performing complete surveys was not possible due to travel restrictions related to COVID-19. We calculated the maximum clutch count per year and then the minimum and maximum abundance for each 4-year period, that is the highest and lowest yearly counts respectively. Given that the species was never found in the last period (see results), we ran a generalised linear mixed model (GLMM) to assess whether the population was showing any significant decline in abundance before its putative extinction in 2022. GLMM was run following Barker, et al. [35]. Simulations and real-world analyses showed that GLMMs using observed abundances can provide good estimates of population trends even for species with imperfect detection [35]. In the GLMM, we used the maximum yearly clutch count in each site as dependent variable, year as independent variable, and site identity as random effect. Before running the model, the year variable was scaled to mean = 0 and SD = 1 to improve model convergence. The number of clutches per site showed strong overdispersion, thus GLMM were run in a Bayesian framework using the brms R package with a negative binomial family [36]. The model was run with three chains for 4,000 iterations, with 1,000 iterations discarded as a burn-in and a thinning interval of 3, in order to sample 1,000 posteriors for each chain. Convergence was ensured by Rhat values being <1.001.
Second, we estimated the range of the Italian agile frog within the park as the minimum convex polygon comprising all the breeding sites used by Italian agile frogs within a given 4-year period (extent of occurrence) [31,37]. Again, we considered a wetland occupied if breeding occurred during at least one year within each period. We then estimated declines as the proportional decline in the extent of occurrence.
3. Results
At the beginning of our study, Italian agile frogs laid clutches in up to seven wetlands spread across the whole park (Figure 2).
Number of breeding females
The number of clutches per site showed high variability among sites and years, ranging from 0 to 207 (Figure 2). The abundance of the frog was highly skewed across sites, with just one site (site D in Figure 1) accounting for >50% of clutches in all years except 2004. During some years, >95% of breeding females laid clutches in that specific site (Figure 2).
Besides the last two years, the total number of clutches across all sites ranged from 45 to 224 (Figure 2). Within each 4-year period, the number of breeding females varied between a minimum of 45-72 to a maximum of 192-224 (Figure 3). Despite variation between adjacent years, the overall population size appeared to be rather stable until 2016-2019. The generalized linear model used to assess the possible decline of the species before its putative extinction from the study area showed an average regression coefficient between year and abundance of -0.04 (95 % credible interval: -0.16 – 0.08), suggesting that no evidence of population decline was detectable prior to the disappearance of the species from the study area.
Despite repeated surveys, no frogs or frog clutches were detected in 2022 and 2023, suggesting that the species went extinct in the park after 2019.
Extent of occurrence within the park
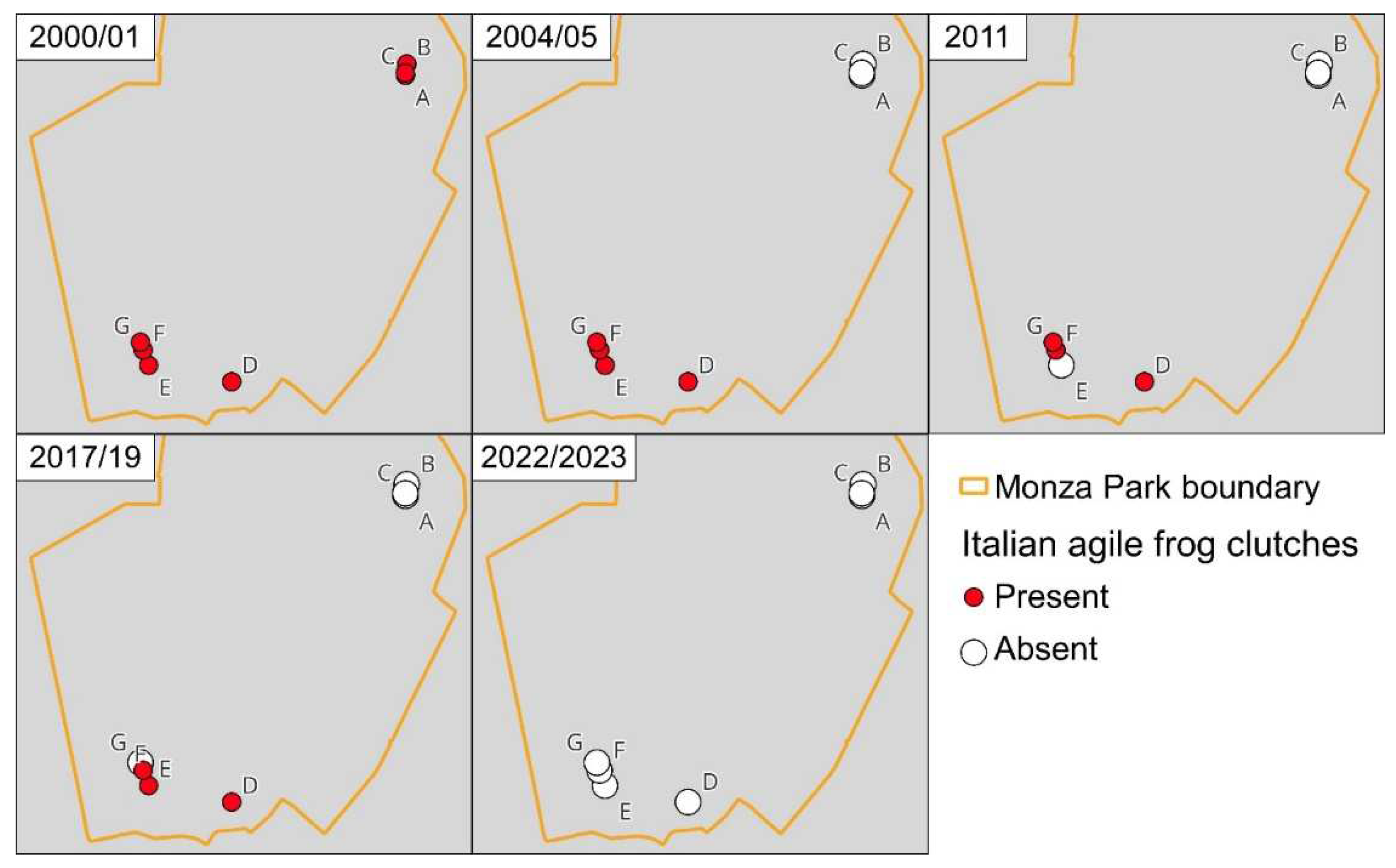
At the beginning of the study, frogs occurred both in Northern and in Southern sites (Figure 1, Figure 4), with a total extent of occurrence of 0.73 km2. However, between 2000 and 2001, the three northernmost sites (A, B, C in Figure 1) were destroyed with the conversion of the small waterbodies into dunghills, leading to the complete disappearance of frogs in the area. The destruction of these sites determined a 88% decline in the extent of occurrence. The extent of occurrence remained stable from 2004 to 2019, but was then followed by the extinction of all the populations in the southern portion of the Park (Figure 4).
4. Discussion
Our long-term monitoring study assessed the temporal changes in the abundance of the Italian agile frog, detecting its possible local extinction from an area that was probably the closest known occurrence to the type locality.
When we used the number of clutches as an estimate of abundance, the population of the Monza Park showed a stable trend from the beginning of our long-term sampling until 2019 (Figure 2, Figure 3), with the number of clutches consistently fluctuating between 40 and 200 (Figure 3). Then, in 2022 the species completely disappeared from all the study sites, and also in 2023 neither clutches nor adults were found during field surveys (Figure 3). The number of clutches was clearly not stable between 2000 and 2019, as it showed striking oscillation between consecutive years. Nevertheless, years with few active females (and thus, presumably, poor breeding success) were generally followed by years with a much larger number of breeding females. For instance, in 2004 just 49 clutches were detected, but this value showed a four-fold increase in 2005. Strong variations in the number of breeding amphibians are well known to occur and are generally related to the interplay between habitat variability, weather conditions, and stochastic demographic factors [6,38,39,40]. Weather conditions and water availability are key determinants of variation in the number of breeding individuals [40], as during dry years lower water availability in wetlands limits the number of breeding adults, but also affect larval stage mortality by increasing desiccation risk [23,41]. The effects of these oscillations can be exacerbated in small and isolated populations. Unfortunately, the Monza Park is severely isolated from other sites hosting Italian agile frogs. The nearest known populations occur ~15 km from the park. Given that the known dispersal distance of these frogs is not expected to exceed 1-km [42], dispersal events between this park and nearby populations are highly unlikely, as confirmed by population genetic analyses [43].
Nevertheless, the number of active individuals is just one possible parameter providing estimates of population size. Measuring population reduction on the basis of the extent of occurrence provided a much earlier evidence of decline (Figure 4). This occurred because habitat loss due to the expansion of farming activities determined the loss of three breeding sites, i.e. nearly 50% of all the breeding sites. Even if these sites were small and did not account for a large number of clutches (Figure 2), their unsuitability strongly reduced the range of the species within the park. Given the limited dispersal ability of amphibians, this can have long-term effects with crucial impacts on population dynamics. For instance, small isolated populations can rescue spatially-structured populations if the largest source populations become temporarily unsuitable (see below for a specific discussion).
The causes of the possible local extinction of the Italian agile frog from the Monza Park are complex, and this extinction was somewhat unexpected given the stable trend of the number of breeding females between 2000 and 2019. The local extinction of this species has also been reported in other core areas of its range. For instance, [23] documented the disappearance of this species from the Groane Regional Park, another protected area located near the type locality of the Italian agile frog. While models of population dynamics were not able to clearly identify the causes of this decline, the analyses suggest that the species nearly disappeared from the Groane Park probably due to high sensitivity to drought, short lifespan, and demographic stochasticity. The lack of explicit environmental data and incomplete sampling hampered testing the role of these processes. Nevertheless, we argue that the possible extinction of the Italian agile frog from the Monza Park may also be related to the joint effect of inappropriate management, drought, and invasive alien species. Many amphibian populations, including the Italian agile frog, survive within meta-population networks [16,22,39]. Our data do not have the temporal completeness needed to develop explicit models of spatially structured populations [22]. The concentration of most breeding individuals in a single pond (Figure 2) is in agreement with the idea that this site generally acts as a source within a meta-population [44]. Unfortunately, unsuitability of source sites for a few years can have dramatic impacts on the dynamics of the overall metapopulations, especially in short life-span species, such as R. latastei. Additionally, the invasive red swamp crayfish (Procambarus clarkii) was introduced in the area around 2005, possibly causing further impacts on this frog, as this crayfish exerts strong predation pressure on amphibian tadpoles [19,21,45].
The Monza Park is actively managed and, during spring 2021, the main breeding sites were without water because of maintenance work. Subsequently, the whole study area was subject to a severe drought between winter 2022 and early spring 2023 [46]. The appropriate management of waterbodies is a crucial task to ensure the persistence of amphibian populations in human-dominated landscapes. The persistence of water in breeding sites during spring is essential to ensure the complete development of amphibian larvae and several methods are available to provision water, among which the most effective are the redirection of water through flow-regulating structures and water pumping [47,48]. However, when a certain region is subject to drought, water use for the conservation of biodiversity and for human use (mainly agriculture) may collide [49]. Therefore, in areas where drought events are expected to be more frequent in the future, the optimization of water use for agriculture and other purposes should also consider the importance of maintaining water resources to ensure the correct development of semi-aquatic organisms, should we want to avoid losses of irreplaceable pieces of biodiversity [50].
Our analyses showed that even in the absence of a decline, populations that were self-sustaining over a long period can rapidly decrease and even disappear (Figure 3). The local extinction of a population can be particularly problematic in suburban areas, where the natural environment is more fragmented, and barriers created by urban and agricultural lands can prevent the recolonization of unoccupied sites [51]. This means that, after dealing with the possible factors that caused the local extinction of a population, human-mediated translocations would be required to re-establish a self-sustaining population in an isolated area such as the Monza Park. However, previous analyses have shown that the long-term isolation of the Monza Park population determined the development of striking local adaptations that allow them to withstand the local environmental challenges. For instance, these populations evolved a fast development rate in response to cold microclimate and the occurrence of alien predators [17,26,29]. As the persistence of the Italian agile frog is strongly related to these adaptations, the re-introductions of individuals belonging to the same species, but lacking specific adaptations, can be challenging. We, therefore, stress that biodiversity-friendly practices (e.g., appropriate water management and preservation of ecologically-relevant sites) should be paired with regular monitoring of biodiversity, in order to promptly identify threats and implement appropriate and timely conservation interventions, avoiding local extinctions.
In principle, the results of our sampling do not necessarily imply a complete extinction of the Italian agile frog from the Monza Park, since some adult individuals might be still present in the available terrestrial habitats, or we might have missed a very small fraction of clutches. Nevertheless, this is rather unlikely, given the high monitoring efforts to which the park is subjected, and considering the high detectability and distinctiveness of R. latastei. This putative extinction is probably related to the join teffect of inappropriate management and severe droughts. While future surveys are still needed to verify the actual local extinction of the Italian agile frog, we point out that eventual reintroduction efforts should be closely followed by appropriate management of the territory by local authorities, to avoid repeating the observed drastic decline that would erase any conservation action.
The EU Habitats Directive (Directive 92/43/EEC) is one of the major tools for the conservation of biodiversity in Europe, as it lists more than 1200 strictly protected species for which also habitats and breeding sites need to be protected. The Habitats Directive aims at “ensure the long term survival of Europe's most valuable and threatened species and habitats” [52], and has developed several tools to halt the decline of listed species. Despite 30 years of efforts, a large proportion of listed species are experiencing a worsening of their status [24]. We join recent calls for the development of more effective and stringent tools for the conservation of European biodiversity.
Author Contributions
Conceptualization, G.F.F., M.F.; R.M.; methodology and data collection, all the authors; writing—original draft preparation, G.F.F., M.F., C.G.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
GFF, MF, MM, RM, ELP, BB, VM and AM were funded by scholarship of the University of Milano. We thank the Monza Park Administration for logistic support during surveys. Part of this research was funded by the European Union – NextGenerationEU, Italian National Recovery and Resilience Plan (Mission 4, Component 2, Investment 1.5 ‘Innovation Ecosystems'), project MUSA.This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Upon acceptance, all data will be uploaded on Figshare (https://figshare.com/)
Acknowledgements
This paper is dedicated to the memory of our friend and colleague M. Barattieri. We thank S. Scali and F. De Bernardi for help during early stages of this work, and M. Maino for providing important unpublished information.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barinaga, M. Where have all the froggies gone? Science 1990, 247, 1033-1034.
- Vitt, L.J.; Caldwell, J.P.; Wilbur, H.M.; Smith, D.C. Amphibians as harbingers of decay. Bioscience 1990, 40, 418. [CrossRef]
- Wake, D. Declining amphibian populations. Science 1991, 253, 860. [CrossRef]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Bohm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The impact of conservation on the status of the world's vertebrates. Science 2010, 330, 1503-1509. [CrossRef]
- Salvidio, S. Detecting amphibian population cycles: The importance of appropriate statistical analyses. Biol. Conserv. 2009, 142, 455–461. [CrossRef]
- Pechmann, J.H.K.; Scott, D.E.; Semlitsch, R.D.; Caldwell, J.P.; Vitt, L.J.; Gibbons, J.W. Declining amphibian populations: the problem of separating human impacts from natural fluctuations. Science 1991, 253, 892-895. [CrossRef]
- Pounds, J.A.; Crump, M.I. Amphibian declines and climate disturbance: the case of the golden toad and the harlequin frog. Conserv. Biol. 1994, 8, 72-85. [CrossRef]
- Pechmann, J.H.K.; Wilbur, H.M. Putting declining amphibian populations in perspective - Natural fluctuations and human impact. Herpetologica 1994, 50, 65-84.
- Pounds, J.A.; Fogden, M.P.L.; Campbell, J.A. Biological response to climate change on a tropical mountain. Nature 1999, 398, 611-615. [CrossRef]
- Ochoa-Ochoa, L.M.; Whittaker, R.J.; Ladle, R.J. The Demise of the Golden Toad and the Creation of a Climate Change Icon Species. Conserv. Soc. 2013, 11, 291-319. [CrossRef]
- IUCN SSC Amphibian Specialist Group. Incilius periglenes. In The IUCN red list of threatened species 2020; 2020; p. e.T3172A54357699.
- Andreone, F.; Garner, T.W.J.; Schmidt, B.R.; Corti, C.; Vogrin, M. Rana latastei. In Threatened amphibians of the world, Stuart, S.N., Hoffman, M., Chanson, J.S., Cox, N.A., Berridge, R.J., Ramani, P., Young, B.E., Eds.; Lynx Editions, IUCN and Conservation International: Barcelona, 2008; p. 506.
- IUCN SSC Amphibian Specialist Group. Rana latastei. In The IUCN red list of threatened species 2022; 2022; p. e.T19156A215527729.
- Boulenger, G.A. Etude sur les grenouilles rousses Ranae temporariae et description d'espèces nouvelles ou méconues. Bulletin de la Société Zoologique de France Paris 1879, 4, 158-193.
- Scali, S. Amphibian and reptiles of Groane Regional Park (Lombardy, NW Italy). First census and ecological notes. Scientia herpetologica 1995, 307-311.
- Ficetola, G.F.; De Bernardi, F. Amphibians in an human-dominated landscape: the community structure is related to habitat features and isolation. Biol. Conserv. 2004, 119, 219-230. [CrossRef]
- Ficetola, G.F.; De Bernardi, F. Supplementation or in situ conservation? Evidence of local adaptation in the Italian agile frog Rana latastei and consequences for the management of populations. Anim. Conserv. 2005, 8, 33-40.
- Bernini, F.; Bonini, L.; Ferri, V.; Gentilli, A.; Razzetti, E.; Scali, S., (Eds.) Atlante degli Anfibi e dei Rettili della Lombardia. Provincia di Cremona: Cremona, 2004.
- Falaschi, M.; Giachello, S.; Lo Parrino, E.; Muraro, M.; Manenti, R.; Ficetola, G.F. Long-term drivers of survival and colonization dynamics in spatially-structured amphibian populations. Conserv. Biol. 2021, 35, 1530–1539. [CrossRef]
- Falaschi, M.; Muraro, M.; Gibertini, C.; Delle Monache, D.; Lo Parrino, E.; Faraci, F.; Belluardo, F.; Di Nicola, M.R.; Manenti, R.; Ficetola, G.F. Explaining declines of newt abundance in northern Italy. Freshw. Biol. 2022. [CrossRef]
- Ficetola, G.F.; Siesa, M.E.; Manenti, R.; Bottoni, L.; De Bernardi, F.; Padoa-Schioppa, E. Early assessment of the impact of alien species: differential consequences of an invasive crayfish on adult and larval amphibians. Divers. Distrib. 2011, 17, 1141-1151. [CrossRef]
- Manenti, R.; Falaschi, M.; Delle Monache, D.; Marta, S.; Ficetola, G.F. Network-scale effects of invasive species on spatially-structured amphibian populations. Ecography 2020, 43, 119-127. [CrossRef]
- Dalpasso, A.; Ficetola, G.F.; Giachello, S.; Lo Parrino, E.; Manenti, R.; Muraro, M.; Falaschi, M. Similar species, different fates: abundance dynamics in spatially structured populations of common and threatened frogs. Divers. Distrib. 2022, 28, 770-781.
- Engelhardt, E.K.; Bowler, D.E.; Hof, C. European Habitats Directive has fostered monitoring but not prevented species declines. Conservation Letters 2023, 16, e12948.
- Barbieri, F.; Mazzotti, S. Rana latastei Boulenger, 1879. In Fauna d'Italia, Vol. XLII: Amphibia, Lanza, B., Andreone, F., Bologna, M.A., Corti, C., Razzetti, E., Eds.; Calderini: Bologna, 2007; pp. 412-416.
- Muraro, M.; Romagnoli, S.; Barzaghi, B.; Falaschi, M.; Manenti, R.; Ficetola, G.F. Invasive predators induce plastic and adaptive responses during embryo development in a threatened frog. NeoBiota 2021, 70, 69-86. [CrossRef]
- Seglie, D.; Marzona, E.; Ficetola, G.F.; Giacoma, C. Comportamento riproduttivo e vocale della rana di Lataste, Rana latastei (Amphibia: Anura). In Herpetologia Sardiniae, Corti, C., Ed.; Belvedere: Latina, 2008; pp. 439-443.
- Falaschi, M.; Gibertini, C.; Lo Parrino, E.; Muraro, M.; Barzaghi, B.; Manenti, R.; Ficetola, G.F. Assessing population trends of species with imperfect detection: Double count analyses and simulations confirm reliable estimates in brown frogs. Animals 2022, 12, 2085. [CrossRef]
- Melotto, A.; Manenti, R.; Ficetola, G.F. Rapid adaptation to invasive predators overwhelms natural gradients of intraspecific variation. Nat. Commun. 2020, 11, 3608.
- Ficetola, G.F.; Padoa-Schioppa, E.; Wang, J.; Garner, T.W.J. Polygyny, census and effective population size in the threatened frog, Rana latastei. Anim. Conserv. 2010, 13 (suppl. 1), 82-89. [CrossRef]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1.; IUCN: Gland, Switzerland and Cambridge, UK, 2001; p. ii+30.
- Lodé, T.; Holveck, M.-J.; Lesbarrères, D. Asynchronous arrival pattern, operational sex ratio and occurrence of multiple paternities in a territorial breeding anuran, Rana dalmatina Biol. J. Linn. Soc. 2005, 86, 191–200.
- Cayuela, H.; Griffiths, R.A.; Zakaria, N.; Arntzen, J.W.; Priol, P.; Léna, J.-P.; Besnard, A.; Joly, P. Drivers of amphibian population dynamics and asynchrony at local and regional scales. J. Anim. Ecol. 2020, 89, 1350-1364. [CrossRef]
- Guarino, F.M.; Lunardi, S.; Carlomagno, M.; Mazzotti, S. A skeletochronological study of growth, longevity, and age at sexual maturity in a population of Rana latastei (Amphibia, Anura). J. Biosciences 2003, 28, 101-108.
- Barker, R.J.; Schofield, M.R.; Link, W.A.; Sauer, J.R. On the reliability of N-mixture models for count data. Biometrics 2018, 74, 369–377. [CrossRef]
- Bürkner, P.-C. Advanced Bayesian Multilevel Modeling with the R Package brms. The R Journal 2018, 10, 395-411. [CrossRef]
- IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 15.1.; http://www.iucnredlist.org/documents/RedListGuidelines.pdf: IUCN, 2022.
- Richter, S.C.; Young, J.E.; Johnson, G.N.; Seigel, R.A. Stochastic variation in reproductive success of a rare frog, Rana sevosa: implications for conservation and for monitoring amphibian populations. Biol. Conserv. 2003, 111, 171-177.
- Marsh, D.M.; Trenham, P.C. Metapopulations dynamics and Amphibian conservation. Conserv. Biol. 2001, 15, 40-49.
- Ficetola, G.F.; Maiorano, L. Contrasting effects of temperature and precipitation change on amphibian phenology, abundance and performance. Oecologia 2016, 181, 683–693. [CrossRef]
- Curtis, R.A.; Sterling, W.B.; Ashley, D.; Andrew, B.; David, R.C. Early Spring and Early Vanishing Wetlands as Harbingers of the Future? The Climate Change Trap for Ephemeral Pond-Breeding Frogs. Northwest Science 2019, 93, 52-65. [CrossRef]
- Smith, M.A.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: are all amphibian populations metapopulations? Ecography 2005, 28, 110-128.
- Ficetola, G.F.; Garner, T.W.J.; De Bernardi, F. Genetic diversity, but not hatching success, is jointly affected by post glacial colonization and isolation in the threatened frog, Rana latastei. Mol. Ecol. 2007, 16, 1787-1797.
- Hanski, I.; Gaggiotti, O.E., (Eds.) Ecology, genetics and evolution of metapopulations. Elsevier Academic Press: San Diego, USA, 2004; p. 696. [CrossRef]
- Lo Parrino, E.; Ficetola, G.F.; Manenti, R.; Falaschi, M. Thirty years of invasion: the distribution of the invasive crayfish Procambarus clarkii in Italy. Biogeographia 2020, 35, 43-50.
- Bonaldo, D.; Bellafiore, D.; Ferrarin, C.; Ferretti, R.; Ricchi, A.; Sangelantoni, L.; Vitelletti, M.L. The summer 2022 drought: a taste of future climate for the Po valley (Italy)? Reg. Envir. Chang. 2022, 23, 1. [CrossRef]
- Mathwin, R.; Wassens, S.; Young, J.; Ye, Q.; Bradshaw, C.J.A. Manipulating water for amphibian conservation. Conserv. Biol. 2021, 35, 24-34.
- Mathwin, R.; Wassens, S.; Gibbs, M.S.; Young, J.; Ye, Q.; Saltré, F.; Bradshaw, C.J.A. Modeling the effects of water regulation on the population viability of a threatened amphibian. Ecosphere 2023, 14, e4379.
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; McCracken, D.; Moritz, R.F.A.; Niemelä, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe–A review. Agriculture, Ecosystems & Environment 2008, 124, 60-71.
- Donati, G.F.A.; Bolliger, J.; Psomas, A.; Maurer, M.; Bach, P.M. Reconciling cities with nature: Identifying local Blue-Green Infrastructure interventions for regional biodiversity enhancement. Journal of Environmental Management 2022, 316, 115254.
- Hamer, A.J.; McDonnell, M.J. Amphibian ecology and conservation in the urbanising world: A review. Biol. Conserv. 2008, 141, 2432-2449. [CrossRef]
- European Commission. EU's biodiversity strategy for 2030; https://ec.europa.eu/environment/strategy/biodiversity-strategy-2030_en: 2021.
Figure 1.
Location of the study sites in the Monza Park (Northern Italy). a) Location of the Monza Park in the Lombardy region (grey) within Northern Italy. b) The two main areas (red squares) hosting breeding sites for Rana latastei in the Monza Park. c) Northeastern breeding sites, destroyed due to the conversion into dunghills between 2000 and 2001. d) Southern breeding sites, with site D which was the one consistently hosting the largest number of clutches until 2019 (see Figure 2).
Figure 1.
Location of the study sites in the Monza Park (Northern Italy). a) Location of the Monza Park in the Lombardy region (grey) within Northern Italy. b) The two main areas (red squares) hosting breeding sites for Rana latastei in the Monza Park. c) Northeastern breeding sites, destroyed due to the conversion into dunghills between 2000 and 2001. d) Southern breeding sites, with site D which was the one consistently hosting the largest number of clutches until 2019 (see Figure 2).
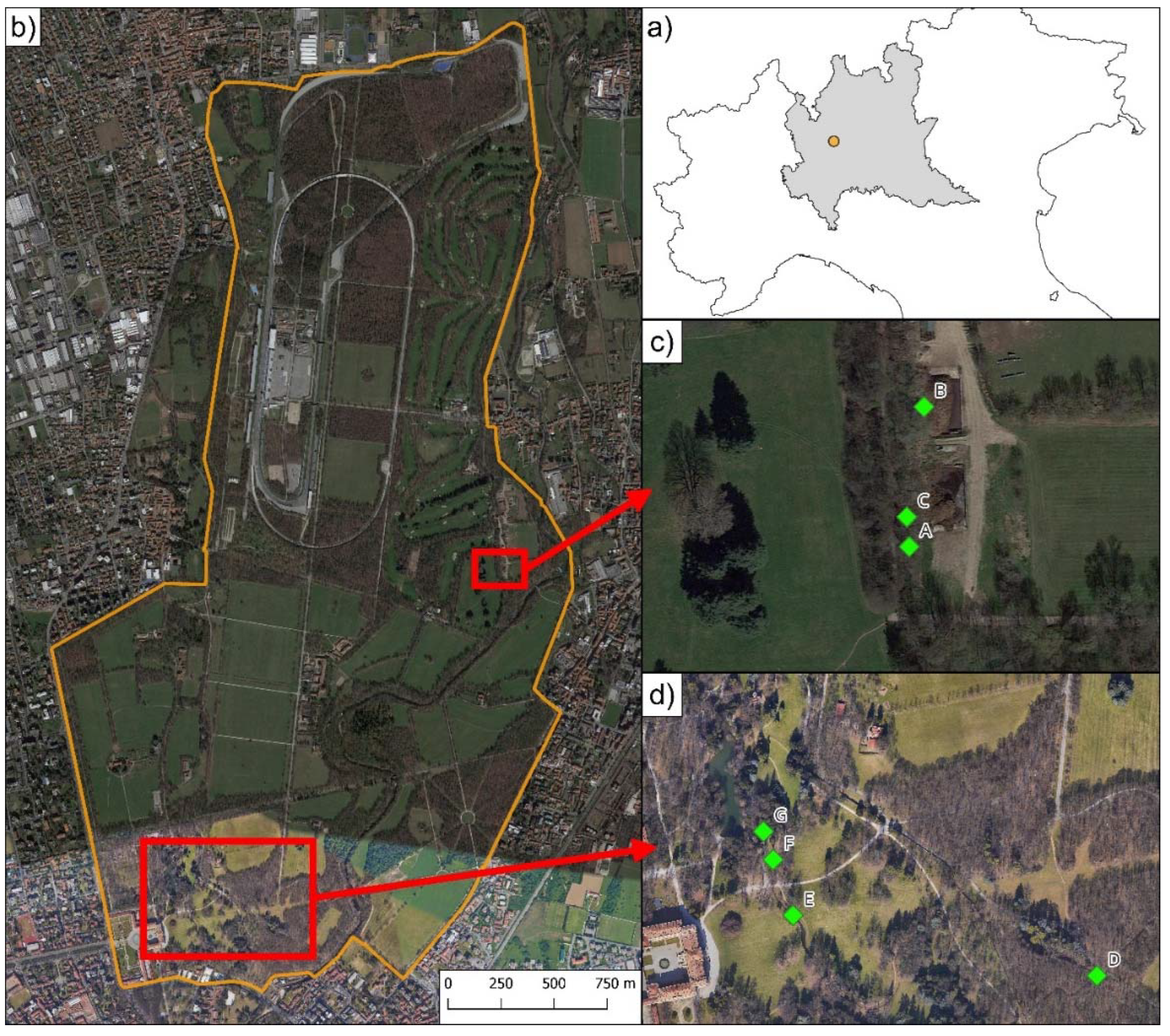
Figure 2.
Counts of Italian agile frog egg clutches present at breeding sites in the Monza Park between 2000 and 2023. For each site, the maximum number of clutches registered for each year is reported. Site codes are provided in Figure 1.
Figure 2.
Counts of Italian agile frog egg clutches present at breeding sites in the Monza Park between 2000 and 2023. For each site, the maximum number of clutches registered for each year is reported. Site codes are provided in Figure 1.
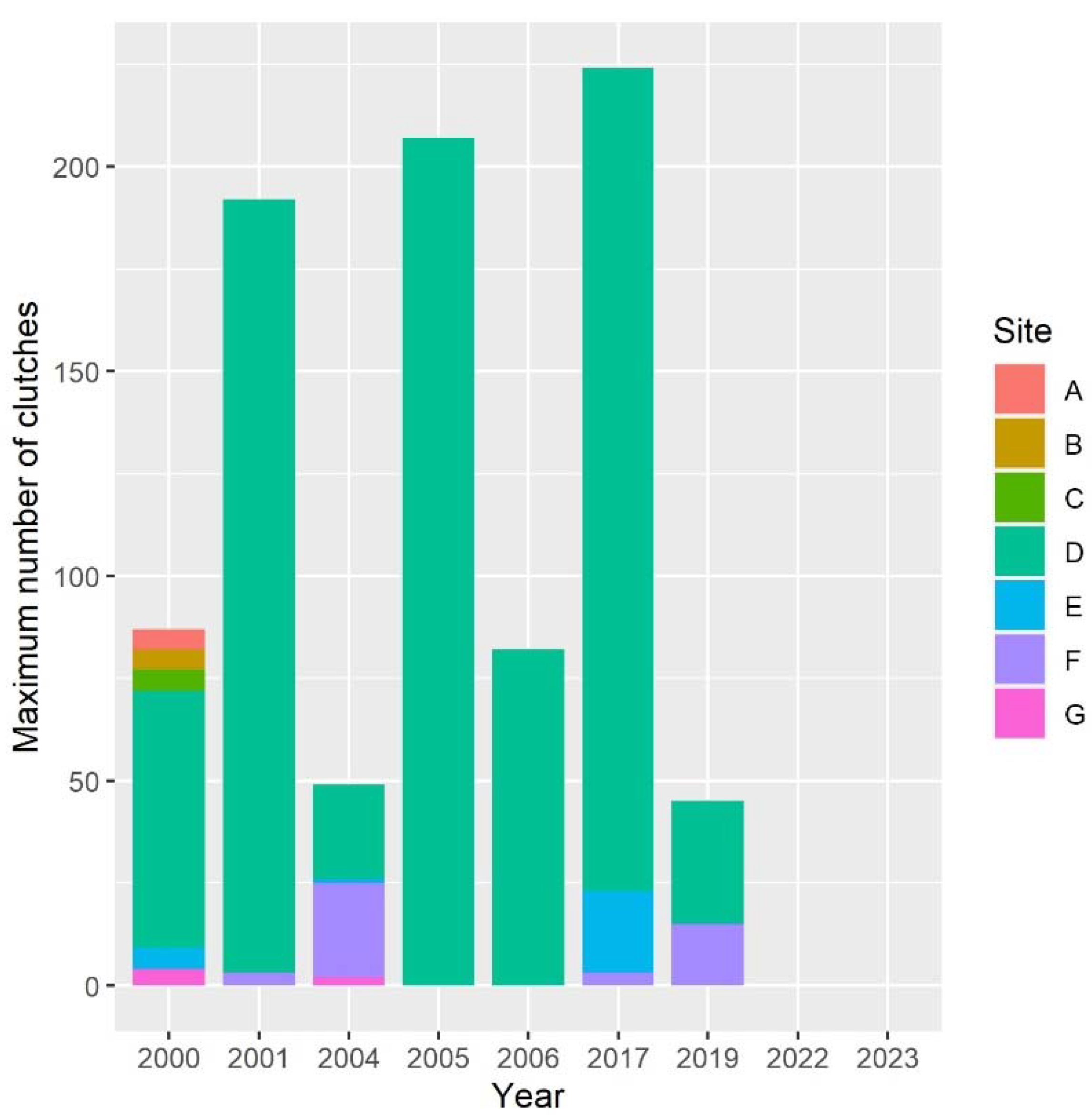
Figure 3.
Maximum (black dots) and minimum (empty dots) number of Italian agile frog clutches in four 4-year periods in the Monza Park. Within each period, the minimum and maximum values were calculated as the highest and lowest yearly abundances, therefore they are an indication of the range of abundance within each 4-year period.
Figure 3.
Maximum (black dots) and minimum (empty dots) number of Italian agile frog clutches in four 4-year periods in the Monza Park. Within each period, the minimum and maximum values were calculated as the highest and lowest yearly abundances, therefore they are an indication of the range of abundance within each 4-year period.
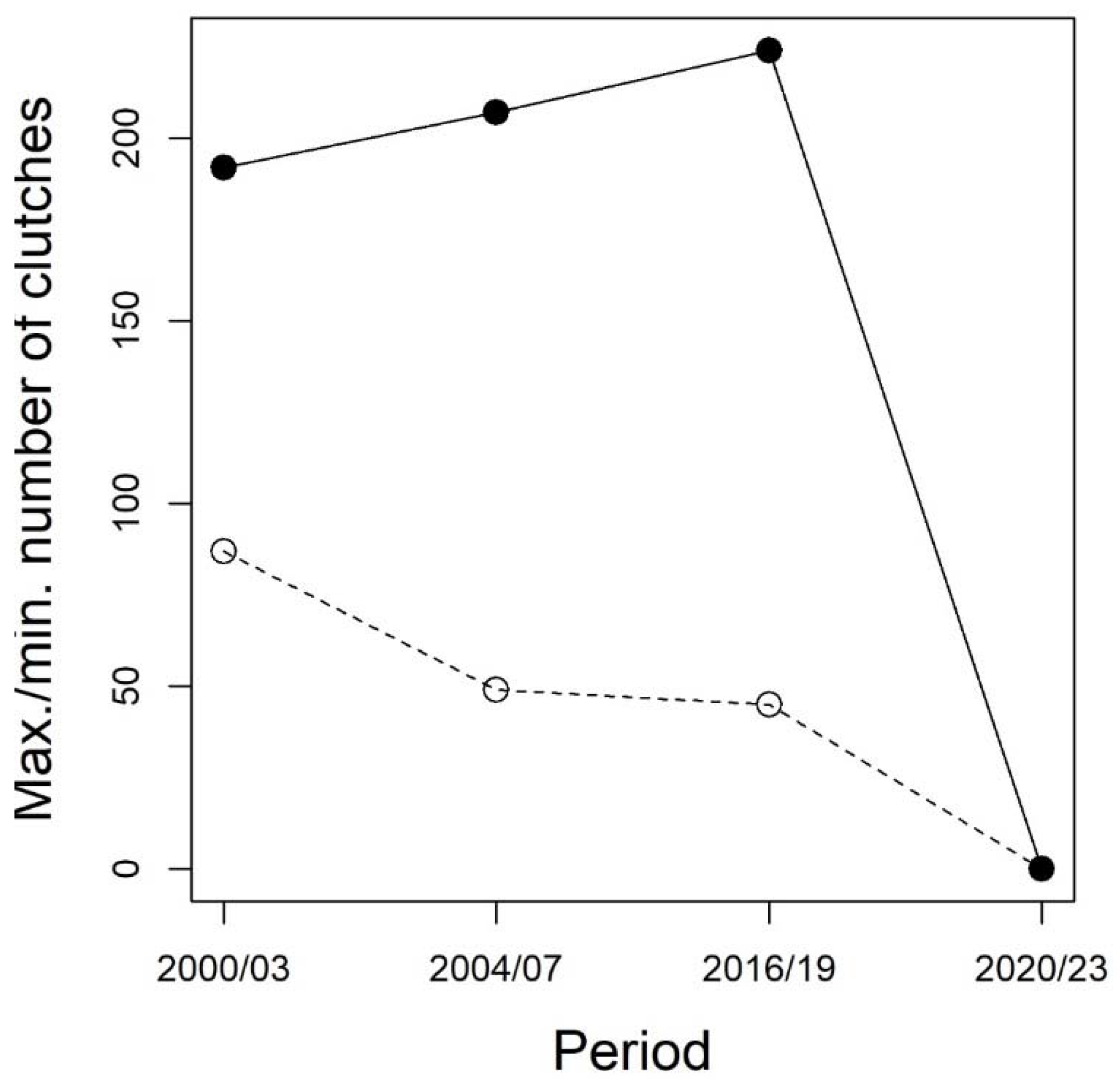
Figure 4.
Changes in distribution of the Italian Agile Frog within the Monza Park from 2000 to 2022. Red dots indicate sites where frog clutches were observed in each period, white dots indicate populations where clutches were not observed.
Figure 4.
Changes in distribution of the Italian Agile Frog within the Monza Park from 2000 to 2022. Red dots indicate sites where frog clutches were observed in each period, white dots indicate populations where clutches were not observed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



