
Submitted:
03 August 2023
Posted:
07 August 2023
You are already at the latest version
Abstract
Inflammatory activation within the brain was linked to a decrease in cognitive abilities, however the molecular mechanisms implicated in the development of inflammatory-related cognitive dysfunction and its prevention are poorly understood. This study compared responses of hippocampal transcriptomes 3 months after the striatal infusion of lipopolysaccharide, alone (LPS; 30 µg) resulting in memory loss, or with dexamethasone (DEX; 5 mg/kg intraperitoneal) pretreatment, which abolished the long-term LPS-induced memory impairment. After LPS alone, a significant elevation in the expression of immunity/inflammatory-linked genes, including chemokine (Cxcl13), cytokines (Il1b, Tnfsf13b), and major histocompatibility complex (MHC) class II members (Cd74, RT1-Ba, RT1-Bb, RT1-Da, RT1-Db1) was revealed. DEX pretreatment did not change expression of these genes, but significantly affected expression of genes encoding ion channels, primarily calcium and potassium channels, regulators of glutamate (Slc1a2, Grm5, Grin2a) and GABA (Gabrr2, Gabrb2) neurotransmission which enriched in such GO biological processes as “Regulation of transmembrane transport”, “Cognition”, “Learning”, “Neurogenesis”, and “Nervous system development”. Taken together, the data suggest that: (1) pretreatment with DEX did not markedly affect LPS-induced prolonged inflammatory response; (2) DEX pretreatment can affect processes associated with glutamatergic signaling and nervous system development, possibly involved by that in the recovery of memory impairment induced by LPS
Keywords:
lipopolysaccharide
; dexamethasone
; hippocampus
; RNA- sequencing
; memory
; inflammation
; glutamate
1. Introduction
Numerous reports indicate the involvement of inflammatory responses within the brain in the pathogenesis of cognitive impairment during neurodegenerative pathologies, including Alzheimer’s and Parkinson’s diseases [1,2,3,4]. Neuroinflammation was also linked to a decrease in cognitive abilities after traumatic brain injuries [5] and ischemia [6,7,8]. However, the molecular mechanisms underlying the cognitive impairment induced by neuroinflammatory activation, as well as possible ways to prevent or reduce development of this disfunction are poorly understood.
Animal studies showed that inflammatory response induced by central administration of lipopolysaccharide (LPS), a cell wall component of Gram-negative bacteria, was accompanied with a cognitive damage evidenced by an inability to increase the recognition index during the object recognition test in both short (48 h) [9] and delayed (3 months) [10] periods after LPS. The glucocorticoid dexamethasone (DEX) can suppress the immune system and reduce inflammation [11], and therefore, the action of this steroid should reduce neuroinflammation-induced cognitive impairment. Indeed, DEX intraperitoneally administered at 30 min before ischemia has been demonstrated to attenuate the acute impairment in cognitive ability of mice and reduce expression of TLR4, NF-kB, and CD68 in their hippocampus [12].
Identification of changes in brain gene expression after LPS is essential for understanding molecular pathways involved in the development of inflammation-induced cognitive and affective disturbances [13]. As was shown in this and our previous study [14], central injections of LPS resulted in hippocampal gene expression changes at 24 h after endotoxin administration. Considering the important role of hippocampus in recognition memory, in the present study, we tested the hypothesis that long-term cognitive impairment occurred along with LPS-induced changes in gene expression in the hippocampus can be prevented by DEX. Transcriptomic changes in the hippocampus associated with long-term behavioral effects of DEX given before LPS have not yet been reported.
2. Materials and Methods
2.1. Animals
Male Wistar rats of 7 weeks (weight 150-180 g) at the beginning of the experiments were used in our study. The rats were housed under 14-h light/10-h dark cycles with free access to food and water. All animal used procedures were approved by the ethics committee of the Institute of Cytology and Genetics in accordance with the guidelines of the Russian Ministry of Health regulations on Good Laboratory Practice (supplement to order No. 199n of 1 April 2016) and the European Council Directive (86/609/EEC).
2.2. Experimental Protocols
In order to activate neuroinflammation, lipopolysaccharide (LPS) from Escherichia coli serotype 055:B5 (Sigma-Aldrich Corp., St Louis, MO, USA) at 30 µg in 4 µL of sterile saline was infused stereotactically into the right striatum under isoflurane anesthesia (4% isoflurane for induction, 2.5% for maintenance in O2 at a flow rate of 1 L/min) using the coordinates: AP = + 0.5 mm, ML = + 3 mm, DV = −5.5/4.5 mm [14,15]. Control animals received an equivalent volume of saline (SAL). Intra-brain LPS exposure can superimpose on the damaging effect of the central injection per se [16]. In our work, the region of the apoptosis activation evaluated by tissue volume containing active caspase-3-positive cells determined by serial sections of the right striatum was approximately 0.5 mm3 after SAL and 7.3 mm3 after LPS showing a very weak impact of the central injection per se.
In the first set of investigations, twenty-four hours after the central administration of LPS (n=6-8) or SAL (n=6-8), rats were quickly decapitated and brain tissue samples (left and right striatum and hippocampus) were collected for mRNAs and proteins of pro-inflammatory and pro-apoptotic markers determination. The part of rats (n=4 per group) were used with the same purposes for immunohistochemical analysis.
In the second set of investigations, the rats were also randomly divided into two groups (n = 20 per group) according to intra-striatal treatment with LPS or SAL. Thirty minutes before the central administration of the drugs, half of the animals from each group received intraperitoneal injections of dexamethasone (DEX; KRKA Slovenia; 5 mg/kg) or vehicle (dimethyl sulfoxide; DMSO; Sigma Aldrich USA). Four groups of animals were created: LPS, SAL, DEX+LPS, and DEX+SAL. Twenty-four hours after the drug administration, rats were tested in the neurological test of Garcia [17]. Three months later, the same rats were tested in the Novel Object Recognition (NOR) test. On the next day after the NOR test, the rats were sacrificed by rapid decapitation. Brains were quickly extracted and ipsilateral hippocampi (n = 4 for each group) were rapidly isolated and each was placed in an Eppendorf tube with 1 mL of buffer containing an RNase inhibitor (RNAlater) at room temperature. After that, the tube was transferred to ice and after the end of hippocampal collection, stored overnight at +4 ◦C and then at −80 ◦C until the analysis of gene expression patterns.
2.3. Neurological Test
Neurologic testing was performed using the Garcia Neurologic Test [17]. This test is based on the 18-point behavioral scale and includes the evaluation of the spontaneous activity, symmetry in the movement of four Limbs, forepaw outstretching, climbing, body proprioception and response to vibrissae touch (each scored between 0 and 3, the maximum is 18).
2.4. Novel Object Recognition (NOR) Test
The NOR test was performed 3 months after drug administration to determine if LPS impacts recognition memory, and dexamethasone can alter LPS-induced behavioral effects. The test consisted of 3 sessions. During the first session, habituation, the rats were allowed to explore the open field arena (100 cm × 100 cm) for 10 min. In the next session 24 h later, two identical objects were placed on the OF (60 cm apart from each other), and the rats were allowed to explore them for 5 min. After that, rats were kept in the home cage for 1 h and then, discrimination session was performed, during which rats were allowed to explore the old and a novel object for 5 min. The OF arena was carefully cleaned with ethanol 10% between all testing periods. The time spent by the rats exploring each object and the number of contacts them were measured, and in order to analyze the cognitive performance, a recognition index was calculated in each session: the number of contacts with novel object / the number of contacts with both objects.
2.5. RNA-Sequencing and Data Analysis
RNA-seq was performed in JSC Genoanalytica (Moscow, Russia; http://genoanalytica.ru). For this, 3 months after drug administration, total RNA was extracted from ipsilateral rat hippocampus with Trisol reagent according to manufacture instruction (n = 4 in each group). Quality was checked with BioAnalyser and RNA 6000 Nano Kit (Agilent, Santa Clara, CA, USA). PolyA RNA was purified with Dynabeads® mRNA Purification Kit (Ambion, Thermo Fisher Scientific, Waltham, MA, USA). Illumina library was made from polyA NEBNext® Ultra™II RNA Library Prep (NEB, Ipswich, MA, USA), according to manual. Sequencing was performed on HiSeq1500 with 50 bp read length. At least 20 million reads were generated for each sample.
The raw reads from RNA-seq experiments were trimmed for quality (phred ≥ 20) and length (bp ≥ 32) using Trimmomatic v3.2.2 [18]. Reads were mapped to the Rnor_6.0 genome with STAR aligner [19] and differentially expressed transcripts were inferred by DESeq2.0 [20]. Genes with │log2 FC (fold change)│≥ 1 and an adjusted p-value (padj) less than 0.05 were classified as significantly differentially expressed genes (DEGs).
The Database for Annotation, Visualization and Integrated Discovery (DAVID; version 6.8; https://david.ncifcrf.gov/ [21] was used to perform functional annotation.
2.6. Real-Time PCR (RT-PCR)
Total RNA was isolated from the brain tissue samples by a one-step guanidine isothiocyanate method. cDNA were synthesized from 3 µg total RNA by M-MuLV reverse transcriptase (SybEnzyme, Russia) and oligo(dT)15 for 1 hours at 42◦C. The IL-1β mRNA was quantified relative the actb mRNA using TaqMan® Gene Expression Assay primers/probes (Rn00580432_m1 for IL-1β, and Rn00667869_m1 for actb; Thermo Fisher Scientific, USA) and VIIA™ 7 Real-Time PCR System (Thermo Fisher Scientific, USA). The mRNA levels were calculated using the ΔΔCt method in triplicate for each sample.
2.7. Immunofluorescence Assay
One day after LPS or SAL administration into striatum, four male rats from each group were selected for immunohistochemistry assay. Animals were deeply anesthetized by avertin, and their brains were removed after transcardial perfusion with ice-cold 0.02 M PBS followed by 4% paraformaldehyde (PFA) in 0.02 M phosphate-buffer saline (PBS). The perfused brains were post-fixed in the same fixative at 4°C for 4 hours and then cryoprotected by complete saturation in a 25% sucrose solution in 0.1 M phosphate buffer (PB; pH 7.4) at 4°C. After that, the brains were quickly frozen using powdered dry ice, cut into 20-μm-thick coronal sections on a cryostat and stored at −60°C before immunohistochemistry. The sections were rinsed twice in 0.02 M PBS with 0.1% Triton X-100 (PBST), and nonspecific binding sites were blocked with a 2.5% BSA (bovine serum albumin) in 0.02 M PBST for 1 h at room temperature. Subsequently, sections were incubated with primary antibodies against cleaved Caspase-3 (AB 9664, Cell Signalling, rabbit; dilution 1:200) and Iba-1 (ab 5076, Abcam, goat; dilution 1:200) for 24 h at 4°C. After that, the sections were washed with 0.02M PBS and incubated with secondary antibodies, F(ab')₂ donkey anti-rabbit IgG conjugated with Cy3 (711-166-152, Jackson ImmunoResearch; dilution 1:300) and F(ab')₂ donkey anti-goat IgG conjugated with Alexa Fluor 488 (705-546-147, Jackson ImmunoResearch; dilution 1:300), for 2 h at room temperature. For the negative controls, sections were incubated without primary antibodies. All sections were then washed with 0.02 M PBS and mounted using MOWIOL mounting medium with DAPI. Images were acquired on a confocal microscope (LSM 780 NLO) equipped with 405 nm, 488 nm and 561 nm lasers, using a Plan-Apochromat 20 objective (0.8 numerical aperture). Caspase-3 and Iba-1 immunoreactivities were quantified by counting amount of their immunopositive cells on sections. At least seven sections were analyzed for each rat with the open sourced program QuPath-0.2.3. The striatum regions for the protein expression analyses were determined using the atlas of Paxinos and Watson (from Bregma -0.8 to +1.6 mm).
2.8. Western Blot
Western blot was performed as described previously [22]. The brain samples were homogenized in lysis buffer containing 150 mM NaCl, 50 mM Tris, 1% Triton X-100 and protease inhibitors: 2 mM PMSF and 2 μg/ml leupeptin, pepstatin and aprotinin. Protein samples (50 μg) were separated by SDS electrophoresis in Mini-Protean 3 Dodeca Cell (Bio-Rad Laboratories, USA) in 12% polyacrylamide gel. The resolved proteins were transferred to 0.45-µм nitrocellulose membrane by Trans-Blot system (Bio-Rad Laboratories, USA). The proteins were stained with primary antibodies: rabbit monoclonal antibodies Iba-1(1:500, EPR16589, ab178847, Abcam, Cambridge, MA), cleaved caspase-3 (1:500, #9664, Cell Signaling, USA) and rabbit polyclonal antibodies β-actin (1:20000, I-19, sc-1616, Santa Cruz Biotechnology, USA). Secondary anti-rabbit IgG (Bio-Rad Laboratories, USA) were used at 1:1000 dilutions for Iba-1, cleaved caspase-3 staining and 1:10000 dilution for β-actin staining. The blots were developed with a SuperSignalTM West Femto Maximum Sensitivity Substrate chemiluminescence kit (Thermo Fisher Scientific, USA) and quantified after scanning with a ChemidocTM Touch Imaging System (Bio-Rad Laboratories, USA) using the Scion Image 4.0.3.2 program (Scion Corporation, USA). The levels of Iba-1 and cleaved caspase-3 proteins were expressed in relative units to β-actin in the same sample.
2.9. Statistical Analysis
For Western blot, PCR, immunohistochemical and behavioral data, two-way analysis of variance (ANOVA) and Tukey’s post hoc test were used to analyze between-group differences. Significant differences were set at p < 0.05.
3. Results
3.1. Ecute Effects of LPS on Striatum and Hippocampus
To verify the acute activation of neuroinflammation and neurodegeneration, at 24 h after LPS administration into the right striatum, expression of Iba1, interleukin-1beta and cleaved caspase-3 was investigated in the right and left striatum, the ipsilateral and contralateral hippocampus by real-time PCR, immunoblotting and immunohistochemically.
3.1.1. Striatum
As shown in Figure 1, LPS caused a rapid pro-inflammatory (a) and pro-apoptotic (b) responses in the right striatum that was a site of endotoxin administration. On the next day after LPS, in the right striatum, PCR and immunoblot analyses showed a significant increase in gene expression of the main pro-inflammatory cytokine interleukin-1beta (effect of LPS: F(1,22) = 13.897, p = 0.0011) (a) and protein of cleaved caspase-3 (effect of LPS: F(1,28) =30.702, p = 0.000006) (b), when compared with appropriate SAL groups. At this time point, effects were not observed in the left striatum (interaction LPS x striatum side: F(1,22) = 12.379, p = 0.0019 for interleukin-1beta; and F(1,28) = 7.865, p = 0.0091 for caspase-3).
It is well-known that LPS activates microglial cells. To evaluate the activation of microglia in the right striatum and caspase-3 in these cells after LPS, antibodies to Iba1 (macrophagic-specific calcium-binding protein) and caspase-3 were employed for immunohistochemical staining. Contrary to our expectation, no significant differences were found between LPS and SAL groups in the number of cells that were immuno-positive only for Iba1. At the same time, the number of cells showed immunoreactivities for both Iba-1 and caspase-3 was significantly increased after LPS (Figure 1c,d).
3.1.2. Hippocampus
In contrast to striatum, in which changes in parameters investigated were observed mainly on the side of LPS administration, short-term pro-inflammatory responses to endotoxin were similar in both the right and the left hippocampus. As compared to appropriate controls, PCR and immunoblot analyses showed increased levels of the pro-inflammatory cytokine interleukin-1beta mRNA one day after LPS injection: effect of LPS: F(1, 32) = 21.385, p = 0.00006; interaction LPS x hippocampal side: (F(1, 32) = 0.234, p = 0.6321) (Figure 2a). However at this time-point, an increase in expression of Iba-1 protein (effect of LPS: F(1, 28) = 8.584, p = 0.0067; interaction LPS x hippocampal side: (F(1, 28) = 1.354, p = 0.2544), reached statistical significance only in contralateral hippocampus (Figure 2b).
3.2. Behavioral Effects of Intra-Striatal LPS and Pretreatment with DEX at 24 h and 3 Months
3.2.1. A Single Pretreatment with DEX Attenuated the Acute LPS-Induced Neurological Deficit
The Garcia score in animals of the LPS group was significantly lower than in SAL group, while pretreatment of LPS animals with DEX, a classic anti-inflammatory drug, attenuated this decrease. As a result, the neurological score in the DEX+LPS group was higher than in the LPS group (Figure 3a).
3.2.2. A Single Pretreatment with DEX Prevented LPS-Induced Long-Lasting Memory Impairment
In the discrimination session of the NOR test that were performed 3 months after drug administration on the same animals used for neurological deficit evaluation described above, the recognition index was significantly decreased in LPS group, whereas this effect was absent, when DEX was injected 30 min before LPS (interaction LPS x DEX: F(1, 36) = 6.7903, p = 0,0132) (Figure 3b).
3.3. Transcriptomic Analyses
In an attempt to identify the molecular mechanisms that underlie the delayed behavioral effects of LPS and their modification by DEX pretreatment, transcriptomic analyses of hippocampus were performed 3 months after the drugs administration. To examine the effect of LPS, LPS-administered rats were compared with SAL group. Effect of DEX pretreatment was examined by comparing DEX+LPS and DEX+ SAL as well as DEX+LPS and LPS groups.
According to the criteria for screening of differentially expressed genes (DEGs): |log2 FC (fold change)| ≥ 1 and an adjusted P value (padj) < 0.05, 33 DEGs, 29 up-regulated and 4 down-regulated (Alox5, LOC500035, Tbx19, H1f4), were revealed in LPS vs. SAL (Supplementary Materials, Table S1). Pretreatment of LPS rats with DEX increased the total number of DEGs that were 161 (128 up-regulated and 33 down-regulated) in DEX+LPS vs. DEX+SAL (Supplementary Materials, Table S2), whereas no DEGs were detected in DEX+ SAL vs. SAL. Comparison of DEX+LPS and LPS groups revealed 112 DEGs (110 up-regulated and 2 down-regulated). All DEGs of three compared pairs of groups, LPS and SAL, DEX+LPS and DEX+SAL, DEX+LPS and LPS, at the significant level padj < 0.05, but │log2 FC (fold change)│ ≥ 0, presented in volcano plots (Figure 4). According to these criteria, only 3 DEGs, Ccdc88a, Ptprb and RT1-N2, were found in DEX+ SAL vs. SAL (volcano plot not shown).
Using the Venn diagrams (http://bioin forma tics.psb.ugent.be/webtools/Venn/), 22 common genes were revealed between DEGs among 33 DEGs in LPS vs. SAL and 161 DEGs in DEX+LPS vs. DEX+SAL, as well as 84 common genes among 161 DEGs in DEX+LPS vs. DEX+SAL and 112 DEGs in DEX+LPS vs. LPS (Figure 5a; Supplementary Materials, Table S3). Figure 5b shows expression profiles of 33 DEGs for LPS vs. SAL. Data were expressed as log2 transformation of fold changes. Asterisks indicate 22 DEGs shared between LPS vs. SAL and DEX+LPS vs. DEX+SAL.
Gene ontology (GO) analysis showed that DEGs after LPS administration (LPS vs. SAL) were predominantly associated with immune and inflammatory responses (Supplementary Materials, Table S4; p-value < 0.05). The 13 most enriched GO terms for biological processes (FDR (false discovery rate) < 0.05) were “antigen processing and presentation of exogenous peptide antigen via MHC class II” (GO:0019886), “antigen processing and presentation of peptide or polysaccharide antigen via MHC class II” (GO:0002504), “response to interferon-gamma“ (GO:0034341), “peptide antigen assembly with MHC class II protein complex” (GO:0002503), “positive regulation of immune response” (GO:0050778), “positive regulation of T cell activation” (GO:0050870), “adaptive immune response” (GO:0002250), “inflammatory response” (GO:0006954), “antigen processing and presentation” (GO:0019882), “immune response” (GO:0006955), “positive regulation of neutrophil chemotaxis” (GO:0090023), “immunoglobulin mediated immune response” (GO:0016064), “humoral immune response” (GO:0006959). Genes that were linked to these biological process terms included the major histocompatibility complex (MHC) class II members (Cd74, RT1-Ba, RT1-Bb, RT1-Da, RT1-Db1, RT1-Db2) and their expression transactivator (Ciita), adhesion receptor (Adgre1), chemoattractants for B lymphocyte (Cxcl13), complement C3 (C3), neutrophil cytosolic factor 1 (Ncf1), proinflammatory cytokines (Il1b, Tnfsf13b), regulators of cellular responses to cytokines (Csf2rb) and inflammatory response to IFNG/IFN-gamma (Mefv), catalyzer of the first step in leukotriene biosynthesis (Alox5), Ras-related C3 botulinum toxin substrate 2 (Rac2), and ubiquitin-like protein modifier (Ubd).
Contrary to our expectation, pretreatment with DEX did not significantly affect the responses of most of these immune/inflammatory-related genes to LPS. Figure 6 shows similar changes in expression of several representative genes such as MHCII genes and Ciita (a), Il1b and Tnfsf13b (b) after LPS alone and with DEX pretreatment. The RNA-sec results were confirmed by quantitative PCR for the IL1b chosen for verification (c). Significant correlation (r = +0.93) was found between sec and PCR IL1b expression values.
Due to this, GO analysis (DAVID) of DEGs that were revealed between DEX+LPS and DEX+SAL also showed that the top 8 biological processes (FDR < 0.05) mainly related to immune response (Supplementary Materials, Table S5): “antigen processing and presentation of exogenous peptide antigen via MHC class II” (GO:0019886), “immunoglobulin production involved in immunoglobulin mediated immune response” (GO:0002381), “peptide antigen assembly with MHC class II protein complex” (GO:0002503), “antigen processing and presentation of peptide or polysaccharide antigen via MHC class II” (GO:0002504), “positive regulation of T cell activation” (GO:0050870), “antigen processing and presentation” (GO:0019882), “response to interferon-gamma”(GO:0034341), “adaptive immune response” (GO:0002250).
To explore the candidate genes and pathways that might play important roles in preventing LPS-induced memory impairment by DEX, STRING (the Search Tool for the Retrieval of Interacting Genes/Proteins, https://string-db.org/) analyses was performed for common and unique DEGs between LPS vs. SAL and DEX+LPS vs. DEX+SAL.
Among the 33 DEGs from LPS vs. SAL, 3 genes (Cd4, Cd74, Il1b) ranked with the interaction degree ≥ 10 connections/interactions, were served as the top hub genes (Table). Among DEGs from comparison of DEX+LPS and DEX+SAL, the number of top hub genes was 6, the list of which in addition to similar with LPS vs. SAL-related DEGs (Cd4, Cd74, Il1b), included Ciita, Creb1 and RT1-Bb (Table). The high level of connections (n = 9) was also observed for Grin2a gene. Of these hub genes, Cd4 exhibited the highest degree.
Of the 33 DEGs revealed between LPS and SAL, 9 did not fall into the Protein-protein interaction (PPI) network (the number of connections/interactions = 0), which, when made from only other DEGs, was constructed of 24 nodes and 62 edges. K-means clustering of these DEGs outlined two clusters depicted in Figure 7a, the biological functions of each of that were characterized by the Gene Ontology (GO). The cluster 1 (green) included 17 DEGs together with Cd4 and Cd74 hub genes (Cd22, Cd4, Cd74, Ciita, Csf2rb, Cxcl13, Emr1, Itgal, Mx1, Plac8, RT1-Ba, RT1-Bb, RT1-Da, RT1-Db1, RT1-Db2, Rac2, Tnfsf13b). These DEGs were mainly enriched in immune response and immune system processes. The second cluster consisted of 7 DEGs together with Il1b hub gene (Alox5, C3, F10, Ifnlr1, Il1b, Mefv, Ncf1), which were involved in inflammatory response and response to stress.
From 22 DEGs shared between LPS vs. SAL and DEX+LPS vs. DEX+SAL, 16 DEGs together with Cd4, Cd74 and Il1b hub genes (Alox5, C3, Cd22, Cd4, Cd74, Ciita, Cxcl13, Emr1, Il1b, Itgal, Ncf1, RT1-Ba, RT1-Bb, RT1-Da, RT1-Db1, Tnfsf13b) that felled into the PPI network, were also predominantly related to immune and inflammatory responses (Figure 7b).
In addition to similar with LPS vs. SAL GO biological processes associated with immune-inflammatory activation that were revealed by DAVID analyses, DEGs from DEX+LPS vs. DEX+SAL also enriched (p-value < 0.05) in “regulation of ion transmembrane transport” (GO:0034765), “cellular response to growth factor stimulus”(GO:0071363), “regulation of membrane potential” (GO:0042391), “regulation of long-term neuronal synaptic plasticity” (GO:0048169), “potassium ion transmembrane transport” (GO:0071805), “intracellular signal transduction” (GO:0035556), “calcium ion transport” (GO:0006816) (Supplementary Materials, Table S5). Genes associating with these terms changed their expression specifically in LPS-administered rats pretreated with DEX, but not in rats received LPS or DEX alone. The list of these genes included calcium channels (Cacna1e, Cacna2d1), Cyclic AMP-responsive element-binding protein 1 (Creb1), Gamma-aminobutyric acid receptor subunits (Gabrb2, Gabrr2), Glutamate ionotropic receptor nmda type subunit 2a (Grin2a) and Metabotropic glutamate receptor 5 (Grm5), Potassium voltage-gated channels (Kcnh5, Kcnh7, Kcnq3), Mitogen activated protein kinase kinase kinase 2 (Map3k2). Figure 8 illustrates the specific increasing effects of DEX on representative genes in LPS-treated rats.
Of 84 specific for DEX+LPS vs. DEX+SAL DEGs, 39 DEGs were felled into the string PPI network. Using k-means clustering, 21 DEGs, including Grin2a hub gene, were grouped (Figure 9). These genes were associated with such GO biological processes, as “Regulation of transmembrane transport” (GO:0034762), “Cognition” ( GO:0050890), “Learning” (GO:0007612), “Neurogenesis” (GO:0022008), “Nervous system development” (GO:0007399).
4. Discussion
The main finding of the present study is that pretreatment with DEX can affect processes associated with glutamatergic signaling and nervous system development, possibly involved by that in the recovery of memory impairment induced by LPS.
4.1. Modeling of Inflammation
LPS, a bacterial mimetic, is widely used as an effective approach to induce neuroinflammation [23] and sickness behavior in rodents [24,25]. Neuroinflammation resulted from systemic or intra-brain LPS administration can induce and aggravate central degeneration [16,26]. We infused LPS into striatum that is among the most damaged areas of the brain after stroke as found in rat models of clinical relevance [27]. Striatum damage is often accompanied by secondary injury of the hippocampus [28]. Twenty four hours after LPS administration, we found the pro-inflammatory and pro-apoptotic activation in the part of striatum that was the site of LPS injection, but not in the other part, supporting the previous results [15], which authors even suggested to use the contralateral region as a control. However, in contrast to the striatum, our data demonstrated that short-term pro-inflammatory responses to intra-striatal LPS were similar in both the ipsilateral and contralateral hippocampi.
The activation of brain glial cells is considered as a critical event for mediating the neuroinflammatory and neurodegenerative responses to LPS. In the injected striatal side and both hippocampi, we found a significant elevation in gene expression of the main pro-inflammatory cytokine Il1b at 24 h after endotoxin administration. In the hippocampus, this effect was associated with the increase in expression of Iba-1 protein, a well-known marker for the activated microglia [29]. In the right striatum at 24 h after LPS, a significant increase in the number of microglial cells immunopositive for both Iba-1 and active caspase-3 was also found. An increase in expression of caspase-3 may reflect its involvement in regulating microglia activation by endotoxin as, for example, was suggested from observations at 24 h after injection of LPS into the rat substantia nigra [30].
4.2. Acute and Delayed Behavioral Effects of LPS: Influence of DEX Pretreatment
At 24 h, the Garcia score (evaluating neurological deficit) of LPS-treated rats was significantly lower than in animals injected with SAL. It should be noted that the Garcia test was designed for assessing function in rodents after ischemic brain injury [17], however this test is also used in other animal models, in the first place, in traumatic brain injury model [31,32]. A decrease in neurologic scores after brain injury can depend on LPS as they were markedly worse by the endotoxin [32]. In our study, pretreatment with such anti-inflammatory synthetic glucocorticoid as DEX attenuated LPS-induced decrease in neurological scores.
Brain damage resulted from injury or inflammatory disease can lead to the development of dementia, the most common symptom of which is memory loss [33]. However, in a short time after a single LPS systemic or intracerebroventricular injection, the data of acute behavioral effects are mixed indicating a decrease [9,34] or no changes [35] in the NOR test. One reason for these inconsistencies might be that specific neural processes underlying impairment of this form of memory was not yet developed. In agreement with such possibility, rats spent significantly less time exploring the novel object than control animals at day 70, but not at day 35 after administration of LPS on postnatal day 7 and 9 [36]. The rats that were given intraperitoneal injections of LPS once a week for 7 weeks exhibited a delay-dependent impairment in spatial memory [37]. Moreover, even in the absence of ongoing neuroimmune activation, memory deficits including object recognition impairment can persist for at least eight weeks after the LPS intraperitoneal injection [38]. Similar to our preliminary study [10], in the present work, the intra-striatal LPS administration also resulted in object recognition task impairment at 3 months.
Given the accepted participation of inflammatory response in the neurocognitive disturbances and anti-inflammatory action of DEX, we thought that DEX could reduce delayed memory impairment after LPS. The results of the present study support this idea. In 3-month, the recognition index significantly decreased in LPS group, and this effect was significantly attenuated by DEX pretreatment. This outcome is in agreement with data from an acute research. The impairment in cognitive ability of mice in the Morris water maze test and development of neuroinflammation 24 h after ischemia-reperfusion were prevented by DEX when intraperitoneally administered at 30 min before the ischemia [12].
4.3. Influence of DEX Pretreatment on Transcriptomic LPS Effects
For the first time, it was shown that a single intra-brain LPS injection up-regulated and down-regulated hippocampal genes expression 3 month later. DEGs after LPS administration were predominantly associated with immune and inflammatory responses. Some long-term gene and protein expression changes associated with peripheral LPS administration have been reported previously. Continued immune/inflammatory activation after LPS was in agreement with those from previous studies, in which even a single peripheral LPS injection can impact the central nervous system for the long time. For example, TNFα protein remained elevated in rodent brain [39], including hippocampus [40], 10 months after LPS intraperitoneal administration. In other study, three months after the end of intraperitoneal LPS injection, several neuroimmune-related genes including Cd74, also showed persistent upregulation in the hippocampus of male rats [41]. Although how LPS mediates the memory impairment is still poorly understood, the changes in expression of some inflammatory-related genes, especially Il1b, but not obviously in expression of MHC II-related genes [42], theoretically may be linked with memory impairment. The novel object recognition performance in the NOR test was inversely correlated with the levels of IL1b in the hippocampus of rats one week after intraperitoneal injection of LPS [43]. On the other hand, IL1b knock-down in the hippocampus significantly attenuated the memory deficits induced in mice by LPS injection [44]. However, in our study, pretreatment of LPS-exposed rats with DEX, which led to increase in recognition index, did not affect expression of IL1b gene suggesting that DEX involved some other mechanisms in the recovery of memory under neuroinflammatory conditions.
GO biological process enrichment analysis showed that Cacna1e, Creb1, Grin2a and Grm5 were significantly enriched in cognition and learning in our study. Changes in expression of modulators of ion channels, in particular, of Ca2+ ion signaling could be espessially important. Administration of felodipine, the L-type Ca2+ channel blocker, to C57BL/6 mice for 9 days significantly reduced LPS-induced spatial memory impairment in the Y-maze test by modulating the formation of hippocampal dendritic spine, but, nevertheless, did not significantly change the novel object preference in the NOR test on days 8-9 of the treatment regimen [45].
The activating effects of glutamate, the main excitatory neurotransmitter in the mammalian brain, on post-synaptic neurons occur through ionotropic and metabotropic receptors and often include Ca2+ influx [46]. Functional properties of the Ionotropic receptors are determined by the composition of the subunits, each of that is encoded by a separate gene [47,48]. GRIN2A (Glutamate Ionotropic Receptor NMDA Type Subunit 2A)-containing hippocampal NMDARs (N-methyl-d-aspartate receptors) were shown to play an important role in synaptic plasticity, learning and memory [49]. These receptors are involved in object recognition memory reconsolidation [50], and GRIN2A subunit knockout mice exhibited discrimination learning impairments [51]. In light of these data, our findings of increased Grin2a expression along with improved memory in DEX-pretreated animals suggest involvement of this receptor subunit in the prevention of endotoxin-induced long-term memory loss by glucocorticoids. Possible involvement of increase in expression of Grm5 gene, coding metabotropic glutamate receptor 5, in the mechanisms of prevention of LPS-induced long-term memory decline by DEX is supported by numerous data suggesting the use of this receptor as a therapeutic target for improving memory deficits [52]. For example, aged rats exposed to one month of environmental enrichment demonstrated an increase in learning and memory in the Morris water maze and novel object recognition behavioral tasks that was accompanied by enhanced hippocampal function including an increase in the level of mGluR5 [53]. In agreement with these data and reductions in hippocampal mGluR5 binding in early Alzheimer's disease that were revealed with help of positron emission tomography [54], our results may expand the understanding of Grm5 role in memory disturbances and correction.
In addition to receptors, in the paper of Brothers with co-authors [55], it was demonstrated the capacity of the brain to compensate for the presence of chronic neuroinflammation through enhanced clearance of extracellular glutamate. Excitotoxicity associated with glutamate accumulation in the extracellular space is regarded among the reasons of memory impairment. Increased expression of Slc1a2 gene encoding transporter for glutamate reuptake from the synaptic cleft was previously observed in the hippocampus one day after the single central administration of LPS [48], but not at 3 months in the present study. At the same time, in LPS-exposed rats pretreated with DEX, expression in Slc1a2 was significantly increased 3 months after the drug administration, indicating enhanced glutamate clearance in DEX-pretreated animals.
The natural aging process is associated with an increased inflammatory response of the brain, so it is not surprising that this model is often used to analyze the mechanisms of memory impairment associated with inflammatory activation. Inflammation and cognitive decline were investigated in old rats and young animals repetedly injected (once a week for 7 weeks) with LPS. Animals from both groups demonstrated similar increase in expression of genes associated with neuroinflammation, as well as impaired spatial memory [37]. In this experiments, however, in contrast to the age-related decrease in transcription of synaptic genes, after LPS, animals exhibited increased expression of genes that support the growth and maintenance of synapses, suggesting ivolvement of some synaptic processis in response to inflammation. In our study, DEX pretreatment affected expression of genes that were associated with such biological processes as Neurogenesis (9 DEGs: Cdkl5, Chl1, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Unc5d) and Nervous system development (10 DEGs: Chl1, Cdkl5, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Slc1a2, Unc5d). Among the proteins coded by listed genes, Cyclin-dependent kinase-like 5 (CDKL5) was suggested as an important regulator of synaptic function in glutamatergic neurons, and ablation of its expression impaired the hippocampal-dependent memory in mice [56]. Dysfunction of NRG1/ErbB4 signaling in the hippocampus was shown to mediate a long-term memory decline in a mouse model of systemic inflammation induced by repeated LPS injections [57].
In contrast to DEX transcriptomic effects in LPS-treated group, no differences in gene expression were observed between DEX+SAL and SAL groups. Poor penetration of dexamethasone into the brain [58] may be among the possible reasons of the lack of transcriptome response to glucocorticoid in the SAL group, while the well-known LPS-induced increase in the blood-brain barrier permeability appears to enhance brain susceptibility to DEX.
It should be also noted that strong criteria for screening of DEGs used in our work could lead to the loss of some functionally important genes, including, for example, gene coding the receptor for glucocorticoids (GR). Comparison of the RNA-sec data for DEX+LPS and DEX+SAL showed a moderate increase in expression of Nr3c1 (log2(FC) = 0.24; p-value = 2.29E-04; padj = 5.03E-03), in DEX+LPS. Changes in expression of GR that is involved in regulation of transcription of numerous genes may also play a role in induction of DEX pretreatment effects, including memory recovery. For example, the protein encoded by Ncf1 (Neutrophil Cytosolic Factor 1) is a cytosolic subunit of NADPH oxidase, which through reactive oxygen species produced by hippocampal microglia influences learning and memory [59]. NADPH oxidase activities, in its turn, can be affected by GR ligands [60].
In overall, the results of the present study showed that pretreatment with DEX did not markedly affect LPS-induced prolonged inflammatory response, and attenuated effect of glucocorticoid on the cognitive impairment may occur partly through changes in glutamatergic system.
5. Conclusion
Taken together, the data suggest that: (1) pretreatment with DEX did not markedly affect LPS-induced prolonged inflammatory response; (2) DEX pretreatment can activate processes associated with glutamatergic signaling and nervous system development, possibly involved by that in recovery of the memory impairment induced by LPS.
Supplementary Materials
Tables S1–S5: Table S1: List of differentially expressed genes with padj <0.05 and |log2FC| ≥ 1 in the hippocampus of LPS vs. SAL; Table S2: List of differentially expressed genes with padj <0.05 and |log2FC| ≥ 1 in the hippocampus of DEX+LPS vs. DEX+SAL; Table S3: venn_result27926; Table S4: Enriched GO biological process terms for LPS vs. SAL DEGs; Table S5: Enriched GO biological process terms for DEX+LPS vs. DEX+SAL DEGs.
Author Contributions
Conceptualization: Conceptualization: G.T.S.; Methodology: G.T.S., T.S.K., D.A.L. and N.N.D.; Investigation: T.S.K., D.A.L., V.V.B., N.P.K., A.V.B., U.S.D.; Writing – original draft preparation: G.T.S. and N.N.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by grant from the Russian Science Foundation Foundation (Grant number 20-64-47013 awarded to G.T.S.) and the state budget project (№ FWNR-2022-0023: care of animals).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the ethic committee of the Institute of Cytology and Genetics in accordance with the guidelines of the Ministry of Public Health of Russia (supplement to order N 267 of June 19, 2003).
Acknowledgments
Confocal microscopy data were acquired at the Advanced Neural Imaging Center at the Institute of Cytology and Genetics. We thank JSC Genoanalytica (Moscow, Russia) for providing the primary bioinformatic analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Öberg, M.; Fabrik, I.; Fabrikova, D.; Zehetner, N.; Härtlova, A. The role of innate immunity and inflammation in Parkinson´s disease. Scand. J. Immunol. 2021, 93, e13022. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Deng, M.; Hu, G.; Li, N.; Yuan, H.; Zhou, Y. New Insights into Microglial Mechanisms of Memory Impairment in Alzheimer’s Disease. Biomolecules 2022, 12, 1722. [Google Scholar] [CrossRef]
- McNicholas, K.; François, M.; Liu, J.-W.; Doecke, J.D.; Hecker, J.; Faunt, J.; Maddison, J.; Johns, S.; Pukala, T.L.; Rush, R.A.; et al. Salivary inflammatory biomarkers are predictive of mild cognitive impairment and Alzheimer’s disease in a feasibility study. Front. Aging Neurosci. 2022, 14, 1019296. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Si, Z.-Z.; Zou, C.-J.; Mei, X.; Li, X.-F.; Luo, H.; Shen, Y.; Hu, J.; Li, X.-X.; Wu, L. Targeting neuroinflammation in Alzheimer’s disease: from mechanisms to clinical applications. Neural Regen. Res. 2022, 18, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Lozano, D.; Schimmel, S.J.; Acosta, S. Neuroinflammation in traumatic brain injury: A chronic response to an acute injury. Brain Circ. 2017, 3, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Thiel, A.; Cechetto, D.F.; Heiss, W.-D.; Hachinski, V.; Whitehead, S.N.; S, E.; J, E.; S, S.; D, W.; J, B.; et al. Amyloid Burden, Neuroinflammation, and Links to Cognitive Decline After Ischemic Stroke. Stroke 2014, 45, 2825–2829. [Google Scholar] [CrossRef]
- Rosenberg, G.A. Extracellular matrix inflammation in vascular cognitive impairment and dementia. Clin. Sci. 2017, 131, 425–437. [Google Scholar] [CrossRef]
- Shishkina, G.T.; Kalinina, T.S.; Gulyaeva, N.V.; Lanshakov, D.A.; Dygalo, N.N. Changes in Gene Expression and Neuroinflammation in the Hippocampus after Focal Brain Ischemia: Involvement in the Long-Term Cognitive and Mental Disorders. Biochem. (Moscow) 2021, 86, 657–666. [Google Scholar] [CrossRef]
- Da Ré, C.; Souza, J.M.; Fróes, F.; Taday, J.; dos Santos, J.P.; Rodrigues, L.; Sesterheim, P.; Gonçalves, C.A.; Leite, M.C. Neuroinflammation induced by lipopolysaccharide leads to memory impairment and alterations in hippocampal leptin signaling. Behav. Brain Res. 2019, 379, 112360. [Google Scholar] [CrossRef]
- Shishkina, G.; Kalinina, T.; Lanshakov, D.; Komysheva, N.; Sukhareva, E.; Dygalo, N. P. 0127 Acute and delayed behavioral effects of lipopolysaccharide with a focus on hippocampal interleukin-1β. Eur. Neuropsychopharmacol. 2021, 53, S93. [Google Scholar] [CrossRef]
- Brown, E.S. Effects of Glucocorticoids on Mood, Memory, and the Hippocampus. Ann. New York Acad. Sci. 2009, 1179, 41–55. [Google Scholar] [CrossRef]
- Cheng, W.; Song, Y.; Liu, Y.; Sun, X.; Ren, W. Impact of Dexamethasone Preconditioning on Prevention of Development of Cognitive Impairment following Acute Inflammation. Contrast Media Mol. Imaging 2022, 2022, 1–8. [Google Scholar] [CrossRef]
- Bonow, R.H.; Aïd, S.; Zhang, Y.; Becker, K.G.; Bosetti, F. The brain expression of genes involved in inflammatory response, the ribosome, and learning and memory is altered by centrally injected lipopolysaccharide in mice. Pharmacogenomics J. 2008, 9, 116–126. [Google Scholar] [CrossRef]
- Shishkina, G.T.; Gulyaeva, N.V.; Lanshakov, D.A.; Kalinina, T.S.; Onufriev, M.V.; Moiseeva, Y.V.; Sukhareva, E.V.; Babenko, V.N.; Dygalo, N.N. Identifying the Involvement of Pro-Inflammatory Signal in Hippocampal Gene Expression Changes after Experimental Ischemia: Transcriptome-Wide Analysis. Biomedicines 2021, 9, 1840. [Google Scholar] [CrossRef]
- Ory, D.; Planas, A.; Dresselaers, T.; Gsell, W.; Postnov, A.; Celen, S.; Casteels, C.; Himmelreich, U.; Debyser, Z.; Van Laere, K.; et al. PET imaging of TSPO in a rat model of local neuroinflammation induced by intracerebral injection of lipopolysaccharide. Nucl. Med. Biol. 2015, 42, 753–761. [Google Scholar] [CrossRef]
- Sauvage, M.C.-D.; Maatouk, L.; Arnoux, I.; Pasco, M.; Diez, A.S.; Delahaye, M.; Herrero, M.T.; A Newman, T.; Calvo, C.F.; Audinat, E.; et al. Potent and multiple regulatory actions of microglial glucocorticoid receptors during CNS inflammation. Cell Death Differ. 2013, 20, 1546–1557. [Google Scholar] [CrossRef]
- Garcia, J.H.; Wagner, S.; Liu, K.-F.; Hu, X.-J.; Ho, K.-L.; Ellsworth, J.L.; Garcia, R.; Yu, J.; Kindy, M.S.; Gerriets, T.; et al. Neurological Deficit and Extent of Neuronal Necrosis Attributable to Middle Cerebral Artery Occlusion in Rats. Stroke 1995, 26, 627–635. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinformatics 2015, 51, 11.14.1–11.14.19. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; A Lempicki, R. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2008, 4, 44–57. [Google Scholar] [CrossRef]
- Shishkina, G.T.; Bannova, A.V.; Komysheva, N.P.; Dygalo, N.N. Anxiogenic-like effect of chronic lipopolysaccharide is associated with increased expression of matrix metalloproteinase 9 in the rat amygdala. Stress 2020, 23, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; De Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef]
- Dantzer, R.; O'Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.R.; Donck, L.V.; et al. Systemic Immune Activation Leads to Neuroinflammation and Sickness Behavior in Mice. Mediat. Inflamm. 2013, 2013, 1–14. [Google Scholar] [CrossRef]
- Choi, S.; Langenbach, R.; Bosetti, F. Genetic deletion or pharmacological inhibition of cyclooxygenase-1 attenuate lipopolysaccharide- induced inflammatory response and brain injury. FASEB J. 2007, 22, 1491–1501. [Google Scholar] [CrossRef]
- Shah, F.A.; Li, T.; Al Kury, L.T.; Zeb, A.; Khatoon, S.; Liu, G.; Yang, X.; Liu, F.; Yao, H.; Khan, A.-U.; et al. Pathological Comparisons of the Hippocampal Changes in the Transient and Permanent Middle Cerebral Artery Occlusion Rat Models. Front. Neurol. 2019, 10. [Google Scholar] [CrossRef]
- Uchida, H.; Fujita, Y.; Matsueda, M.; Umeda, M.; Matsuda, S.; Kato, H.; Kasahara, J.; Araki, T. Damage to Neurons and Oligodendrocytes in the Hippocampal CA1 Sector after Transient Focal Ischemia in Rats. Cell. Mol. Neurobiol. 2010, 30, 1125–1134. [Google Scholar] [CrossRef]
- Hoogland, I.C.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic inflammation and microglial activation: systematic review of animal experiments. J. Neuroinflammation 2015, 12, 114. [Google Scholar] [CrossRef]
- Burguillos, M.A.; Deierborg, T.; Kavanagh, E.; Persson, A.; Hajji, N.; Garcia-Quintanilla, A.; Cano, J.; Brundin, P.; Englund, E.; Venero, J.L.; et al. Caspase signalling controls microglia activation and neurotoxicity. Nature 2011, 472, 319–324. [Google Scholar] [CrossRef]
- McBride, D.W.; Wang, Y.; Adam, L.; Oudin, G.; Louis, J.-S.; Tang, J.; Zhang, J.H. Correlation Between Subacute Sensorimotor Deficits and Brain Edema in Rats after Surgical Brain Injury. 2016, 121, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Jacovides, C.L.; Ahmed, S.; Suto, Y.; Paris, A.J.; Leone, R.; McCarry, J.; Christofidou-Solomidou, M.; Kaplan, L.J.; Smith, D.H.; Holena, D.N.; et al. An inflammatory pulmonary insult post-traumatic brain injury worsens subsequent spatial learning and neurological outcomes. J. Trauma: Inj. Infect. Crit. Care 2019, 87, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Bishnoi, I.R.; Cloutier, C.J.; Tyson, C.-D.; Matic, V.M.; Kavaliers, M.; Ossenkopp, K.-P. Infection, learning, and memory: Focus on immune activation and aversive conditioning. Neurosci. Biobehav. Rev. 2022, 142, 104898. [Google Scholar] [CrossRef] [PubMed]
- Hennigan, A.; Trotter, C.; Kelly, M. Lipopolysaccharide impairs long-term potentiation and recognition memory and increases p75NTR expression in the rat dentate gyrus. Brain Res. 2007, 1130, 158–166. [Google Scholar] [CrossRef]
- Czerniawski, J.; Miyashita, T.; Lewandowski, G.; Guzowski, J.F. Systemic lipopolysaccharide administration impairs retrieval of context–object discrimination, but not spatial, memory: Evidence for selective disruption of specific hippocampus-dependent memory functions during acute neuroinflammation. Brain, Behav. Immun. 2015, 44, 159–166. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Harte, M.K.; Stenson, G.; Reynolds, G.P. Neonatal lipopolysaccharide induces pathological changes in parvalbumin immunoreactivity in the hippocampus of the rat. Behav. Brain Res. 2009, 205, 355–359. [Google Scholar] [CrossRef]
- Barter, J.; Kumar, A.; Rani, A.; Colon-Perez, L.M.; Febo, M.; Foster, T.C. Differential Effect of Repeated Lipopolysaccharide Treatment and Aging on Hippocampal Function and Biomarkers of Hippocampal Senescence. Mol. Neurobiol. 2020, 57, 4045–4059. [Google Scholar] [CrossRef]
- Tchessalova, D.; Tronson, N.C. Memory deficits in males and females long after subchronic immune challenge. Neurobiol. Learn. Mem. 2019, 158, 60–72. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.-S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef]
- Bossù, P.; Cutuli, D.; Palladino, I.; Caporali, P.; Angelucci, F.; Laricchiuta, D.; Gelfo, F.; De Bartolo, P.; Caltagirone, C.; Petrosini, L. A single intraperitoneal injection of endotoxin in rats induces long-lasting modifications in behavior and brain protein levels of TNF-α and IL-18. J. Neuroinflammation 2012, 9, 101–101. [Google Scholar] [CrossRef]
- Tchessalova, D.; Tronson, N.C. Enduring and Sex-specific Changes in Hippocampal Gene Expression after a Subchronic Immune Challenge. Neuroscience 2020, 428, 76–89. [Google Scholar] [CrossRef]
- VanGuilder, H.D.; Bixler, G.V.; Brucklacher, R.M.; A Farley, J.; Yan, H.; Warrington, J.P.; E Sonntag, W.; Freeman, W.M. Concurrent hippocampal induction of MHC II pathway components and glial activation with advanced aging is not correlated with cognitive impairment. J. Neuroinflammation 2011, 8, 138–138. [Google Scholar] [CrossRef]
- Kawano, T.; Morikawa, A.; Imori, S.; Waki, S.; Tamura, T.; Yamanaka, D.; Yamazaki, F.; Yokoyama, M. Preventive effects of multisensory rehabilitation on development of cognitive dysfunction following systemic inflammation in aged rats. J. Anesthesia 2014, 28, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, C.; Yu, H.; Cai, X.; Shen, X.; Sun, X.; Wang, J.; Zhang, Y.; Wang, C. Lentivirus-mediated interleukin-1β (IL-1β) knock-down in the hippocampus alleviates lipopolysaccharide (LPS)-induced memory deficits and anxiety- and depression-like behaviors in mice. J. Neuroinflammation 2017, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jeon, S.G.; Jeong, H.-R.; Park, H.; Kim, J.-I.; Hoe, H.-S. L-Type Ca2+ Channel Inhibition Rescues the LPS-Induced Neuroinflammatory Response and Impairments in Spatial Memory and Dendritic Spine Formation. Int. J. Mol. Sci. 2022, 23, 13606. [Google Scholar] [CrossRef] [PubMed]
- Pregnolato, S.; Chakkarapani, E.; Isles, A.R.; Luyt, K. Glutamate Transport and Preterm Brain Injury. Front. Physiol. 2019, 10, 417. [Google Scholar] [CrossRef]
- Dygalo, N.N.; Kalinina, T.S.; Shishkina, G.T. Stress-induced expression pattern of glutamate signaling genes associated with anhedonia. Stress 2020, 23, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, T.S.; Shishkina, G.T.; Lanshakov, D.A.; Sukhareva, E.V.; Onufriev, M.V.; Moiseeva, Y.V.; Gulyaeva, N.V.; Dygalo, N.N. Comparative Investigation of Expression of Glutamatergic and GABAergic Genes in the Rat Hippocampus after Focal Brain Ischemia and Central LPS Administration. Biochem. (Moscow) 2023, 88, 539–550. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, X.; Zhang, L.; Hu, J.; Chen, Y.; Zhan, L.; Gao, Z. The Functional and Molecular Properties, Physiological Functions, and Pathophysiological Roles of GluN2A in the Central Nervous System. Mol. Neurobiol. 2016, 54, 1008–1021. [Google Scholar] [CrossRef]
- Rossato, J.I.; Radiske, A.; Gonzalez, M.C.; Apolinário, G.; de Araújo, R.L.; Bevilaqua, L.R.; Cammarota, M. NMDARs control object recognition memory destabilization and reconsolidation. Brain Res. Bull. 2023, 197, 42–48. [Google Scholar] [CrossRef]
- Brigman, J.L.; Feyder, M.; Saksida, L.M.; Bussey, T.J.; Mishina, M.; Holmes, A. Impaired discrimination learning in mice lacking the NMDA receptor NR2A subunit. Learn. Mem. 2008, 15, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elrahman, K.S.; Ferguson, S.S.G. Noncanonical Metabotropic Glutamate Receptor 5 Signaling in Alzheimer's Disease. Annu Rev Pharmacol Toxicol. 2022, 62, 235–254. [Google Scholar] [CrossRef]
- Cortese, G.P.; Olin, A.; O'Riordan, K.; Hullinger, R.; Burger, C. Environmental enrichment improves hippocampal function in aged rats by enhancing learning and memory, LTP, and mGluR5-Homer1c activity. Neurobiol. Aging 2018, 63, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mecca, A.P.; McDonald, J.W.; Michalak, H.R.; Godek, T.A.; Harris, J.E.; Pugh, E.A.; Kemp, E.C.; Chen, M.-K.; Salardini, A.; Nabulsi, N.B.; et al. PET imaging of mGluR5 in Alzheimer’s disease. Alzheimer's Res. Ther. 2020, 12, 15–10. [Google Scholar] [CrossRef] [PubMed]
- Brothers, H.M.; Wenk, G.L.; Bardou, I.; Hopp, S.C.; Marchalant, Y.; Turner, S.M.; Mitchem, M.R.; Kigerl, K.; Kaercher, R.M. Time-Dependent Compensatory Responses to Chronic Neuroinflammation in Hippocampus and Brainstem: The Potential Role of Glutamate Neurotransmission. J. Alzheimer's Dis. Park. 2013, 3, 1–9. [Google Scholar] [CrossRef]
- Tang, S.; Wang, I.-T.J.; Yue, C.; Takano, H.; Terzic, B.; Pance, K.; Lee, J.Y.; Cui, Y.; Coulter, D.A.; Zhou, Z. Loss of CDKL5 in Glutamatergic Neurons Disrupts Hippocampal Microcircuitry and Leads to Memory Impairment in Mice. J. Neurosci. 2017, 37, 7420–7437. [Google Scholar] [CrossRef]
- Gao, Y.Z.; Wu, X.M.; Zhou, Z.Q.; Liu, P.M.; Yang, J.J.; Ji, M.H. Dysfunction of NRG1/ErbB4 Signaling in the Hippocampus Might Mediate Long-term Memory Decline After Systemic Inflammation. Mol. Neurobiol. 2023, 60, 3210–3226. [Google Scholar] [CrossRef]
- De Kloet, E.R. Why Dexamethasone Poorly Penetrates in Brain. Stress 1997, 2, 13–20. [Google Scholar] [CrossRef]
- Wang, K.; Shi, Y.; Liu, W.; Liu, S.; Sun, M.-Z. Taurine improves neuron injuries and cognitive impairment in a mouse Parkinson’s disease model through inhibition of microglial activation. NeuroToxicology 2021, 83, 129–136. [Google Scholar] [CrossRef]
- Cuong, T.T.; Yang, C.-S.; Yuk, J.-M.; Lee, H.-M.; Ko, S.-R.; Cho, B.-G.; Jo, E.-K. Glucocorticoid receptor agonist compound K regulates dectin-1-dependent inflammatory signaling through inhibition of reactive oxygen species. Life Sci. 2009, 85, 625–633. [Google Scholar] [CrossRef]
Figure 1.
Pro-inflammatory and pro-apoptotic effects of LPS in the striatum. In the right striatum that was the site of endotoxin infusion, LPS significantly (*p < 0.05) increased gene expression of pro-inflammatory cytokine interleukin-1beta (a) and protein of active caspase-3 (b) compared with SAL group at 24 h. In the right striatum, immunohistochemical analysis, using Iba-1 as a specific marker for microglia, did not reveal any differences between LPS and SAL group in the number of cells that were only Iba1 immuno-positive, whereas the number of cells immunoreactivities for both Iba-1 and caspase-3 was significantly increased (*p < 0.05) (c). (d) Iba-1 (green) and caspase-3 (red) immunofluorescence of representative images from the right striatum after SAL (up) and LPS (down).
Figure 1.
Pro-inflammatory and pro-apoptotic effects of LPS in the striatum. In the right striatum that was the site of endotoxin infusion, LPS significantly (*p < 0.05) increased gene expression of pro-inflammatory cytokine interleukin-1beta (a) and protein of active caspase-3 (b) compared with SAL group at 24 h. In the right striatum, immunohistochemical analysis, using Iba-1 as a specific marker for microglia, did not reveal any differences between LPS and SAL group in the number of cells that were only Iba1 immuno-positive, whereas the number of cells immunoreactivities for both Iba-1 and caspase-3 was significantly increased (*p < 0.05) (c). (d) Iba-1 (green) and caspase-3 (red) immunofluorescence of representative images from the right striatum after SAL (up) and LPS (down).
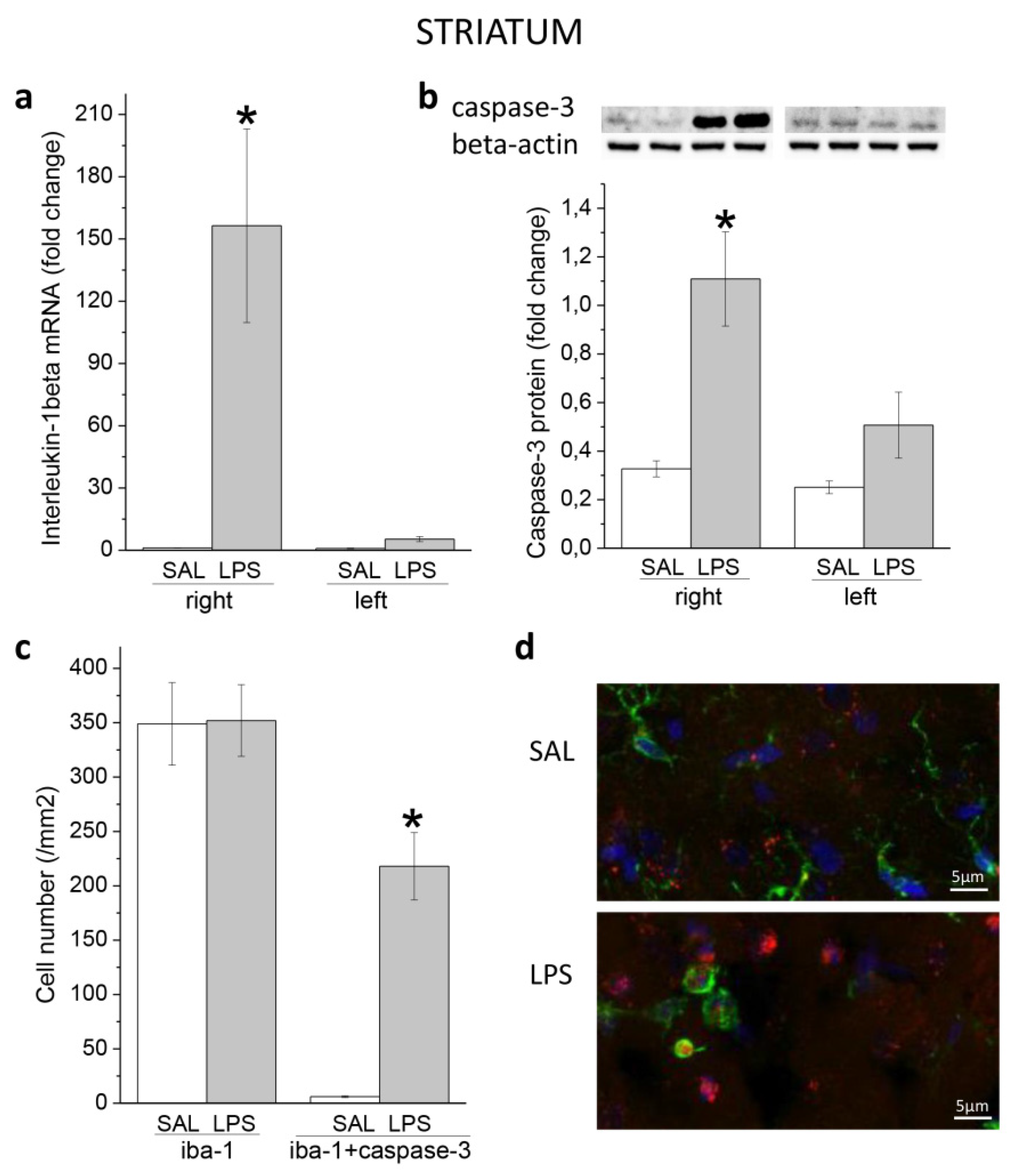
Figure 2.
Expression levels of interleukin-1beta mRNA (a) and protein of Iba-1 (b) in ipsilateral and contralateral hippocampi 24 h following administration of LPS into right striatum. * p < 0.05 compared with an appropriate SAL group.
Figure 2.
Expression levels of interleukin-1beta mRNA (a) and protein of Iba-1 (b) in ipsilateral and contralateral hippocampi 24 h following administration of LPS into right striatum. * p < 0.05 compared with an appropriate SAL group.
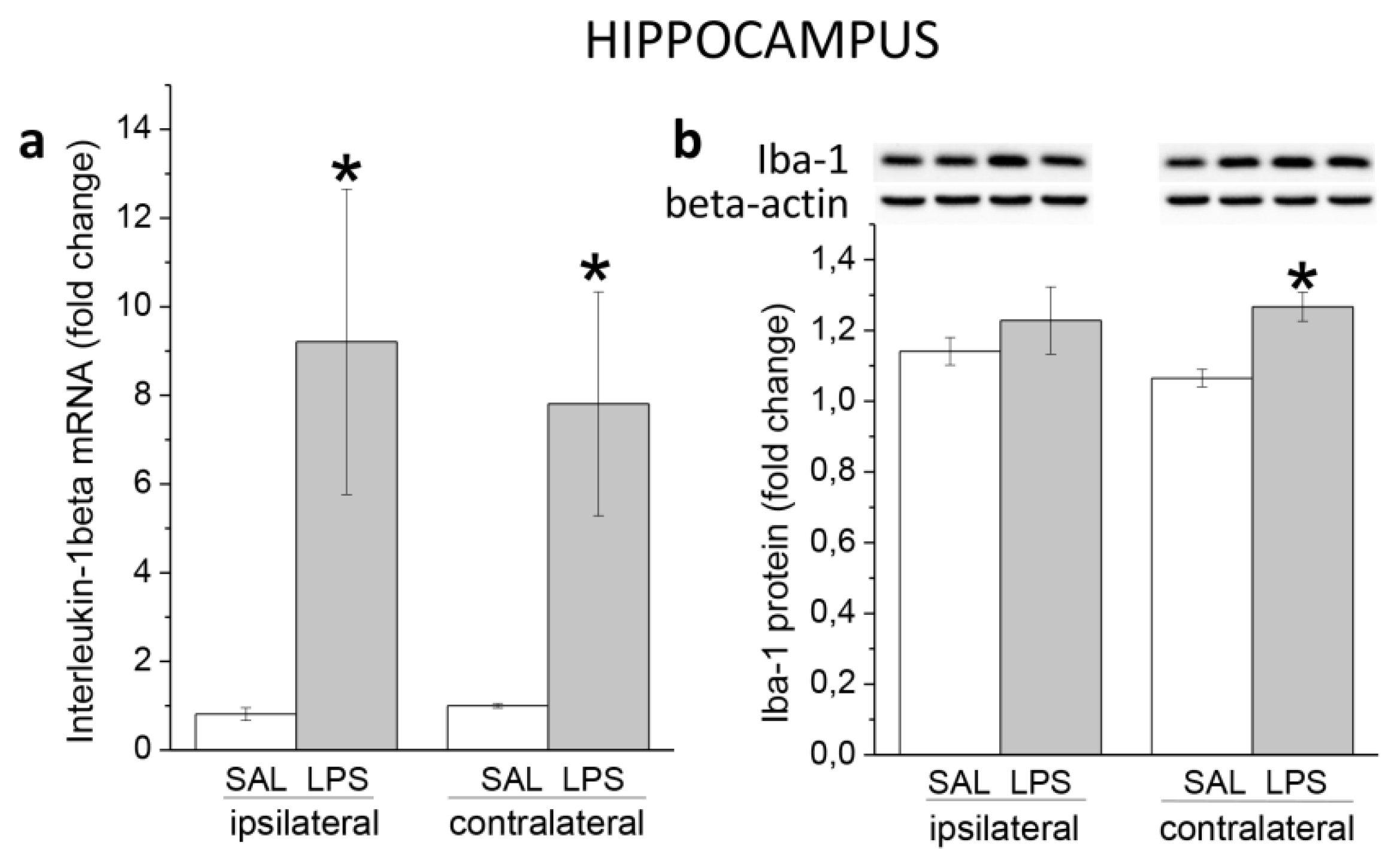
Figure 3.
Short-term and delayed behavioral effects of LPS administered alone or with DEX. (a) The Garcia score at 24 h. Neurological scale of the LPS-treated rats significantly decreased compared to that of the SAL-treated rats. When animals were pretreated with DEX, the neurological scale of the DEX+LPS group was higher than after LPS alone. (b) The recognition index assessed 3 months after the drugs administration. LPS caused memory impairment, and this effect was prevented by DEX pretreatment. Each group of animals that were the same on (a) and (b) consisted of 10 rats. * p < 0.05 compared with an appropriate control group (SAL or DEX+SAL); # p < 0.05 compared with LPS group.
Figure 3.
Short-term and delayed behavioral effects of LPS administered alone or with DEX. (a) The Garcia score at 24 h. Neurological scale of the LPS-treated rats significantly decreased compared to that of the SAL-treated rats. When animals were pretreated with DEX, the neurological scale of the DEX+LPS group was higher than after LPS alone. (b) The recognition index assessed 3 months after the drugs administration. LPS caused memory impairment, and this effect was prevented by DEX pretreatment. Each group of animals that were the same on (a) and (b) consisted of 10 rats. * p < 0.05 compared with an appropriate control group (SAL or DEX+SAL); # p < 0.05 compared with LPS group.
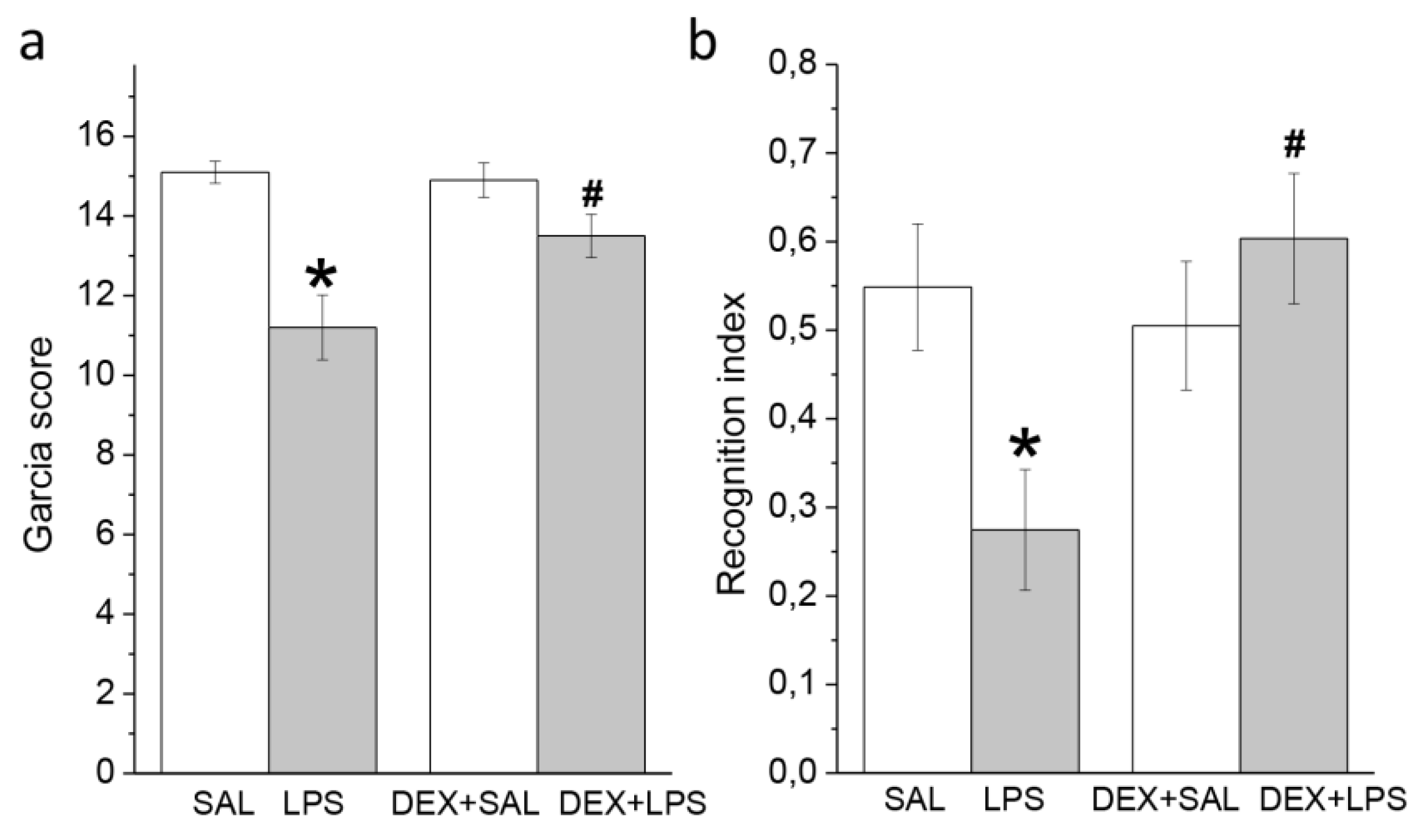
Figure 4.
Volcano plots show the differentially expressed genes (padj < 0.05). Red represents up-regulated genes, and blue represents down-regulated genes with│log2 (fold change)│ ≥ 1.
Figure 4.
Volcano plots show the differentially expressed genes (padj < 0.05). Red represents up-regulated genes, and blue represents down-regulated genes with│log2 (fold change)│ ≥ 1.
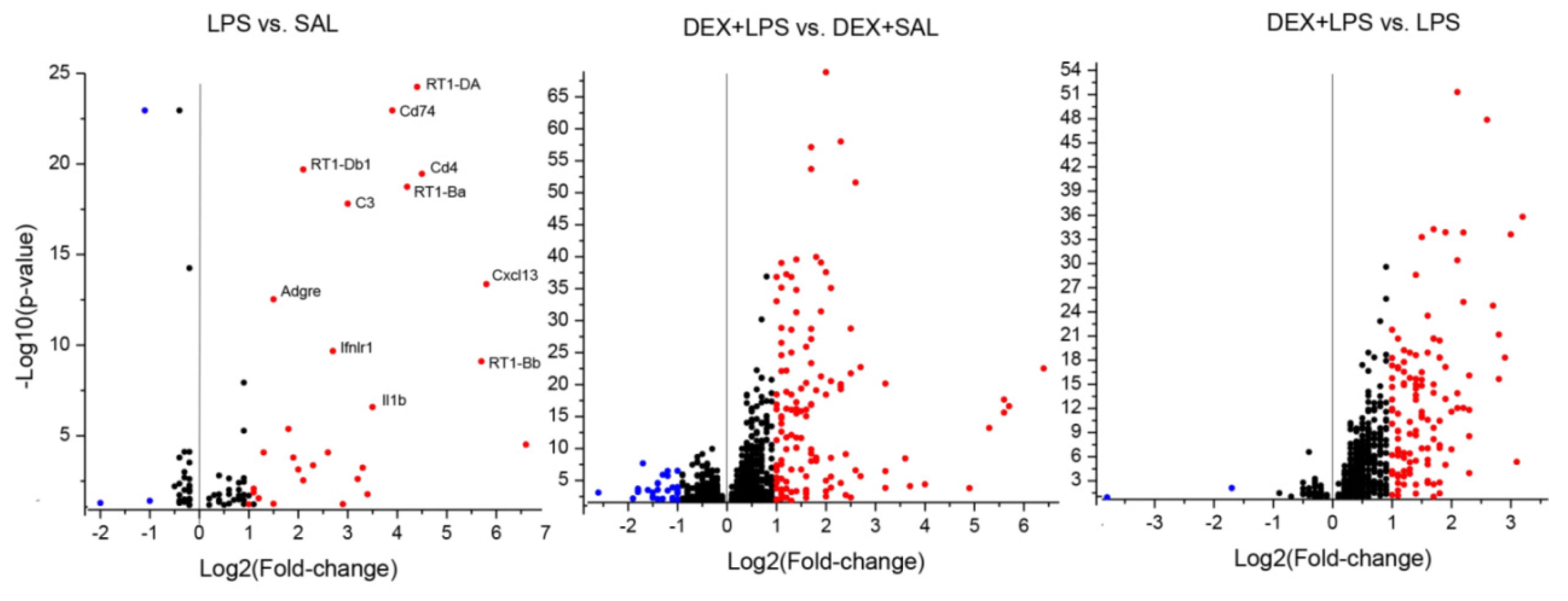
Figure 5.
(a) Common genes among DEGs in LPS vs. SAL and DEX+LPS vs. DEX+SAL (22 genes), as well as DEX+LPS vs. DEX+SAL and DEX+LPS vs. LPS (84 genes). (b) Expression profiles of DEGs for LPS vs. SAL. Data were expressed as log2 transformation of fold changes. Asterisks indicate 22 DEGs common for LPS vs. SAL and DEX+LPS vs. DEX+SAL.
Figure 5.
(a) Common genes among DEGs in LPS vs. SAL and DEX+LPS vs. DEX+SAL (22 genes), as well as DEX+LPS vs. DEX+SAL and DEX+LPS vs. LPS (84 genes). (b) Expression profiles of DEGs for LPS vs. SAL. Data were expressed as log2 transformation of fold changes. Asterisks indicate 22 DEGs common for LPS vs. SAL and DEX+LPS vs. DEX+SAL.
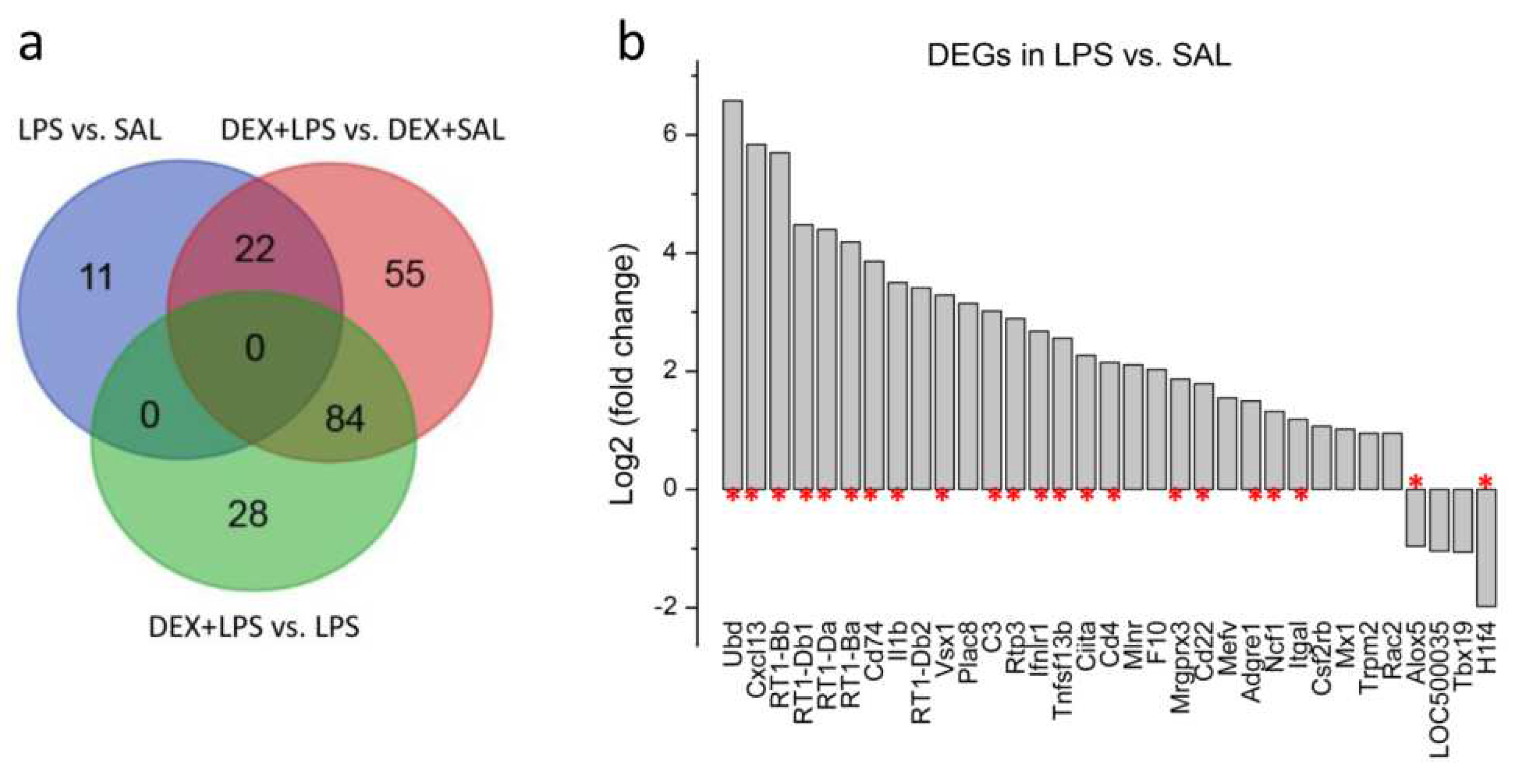
Figure 6.
Responses of representative genes to LPS alone and with DEX pretreatment (relative to SAL group): (a) MHCII-related genes (RT1-Db1, RT1-Da, RT1-Ba, Cd74), Ciita; (b) Il1b and Tnfsf13b; (c) verification of the RNA-sec results for IL1b by quantitative PCR.
Figure 6.
Responses of representative genes to LPS alone and with DEX pretreatment (relative to SAL group): (a) MHCII-related genes (RT1-Db1, RT1-Da, RT1-Ba, Cd74), Ciita; (b) Il1b and Tnfsf13b; (c) verification of the RNA-sec results for IL1b by quantitative PCR.
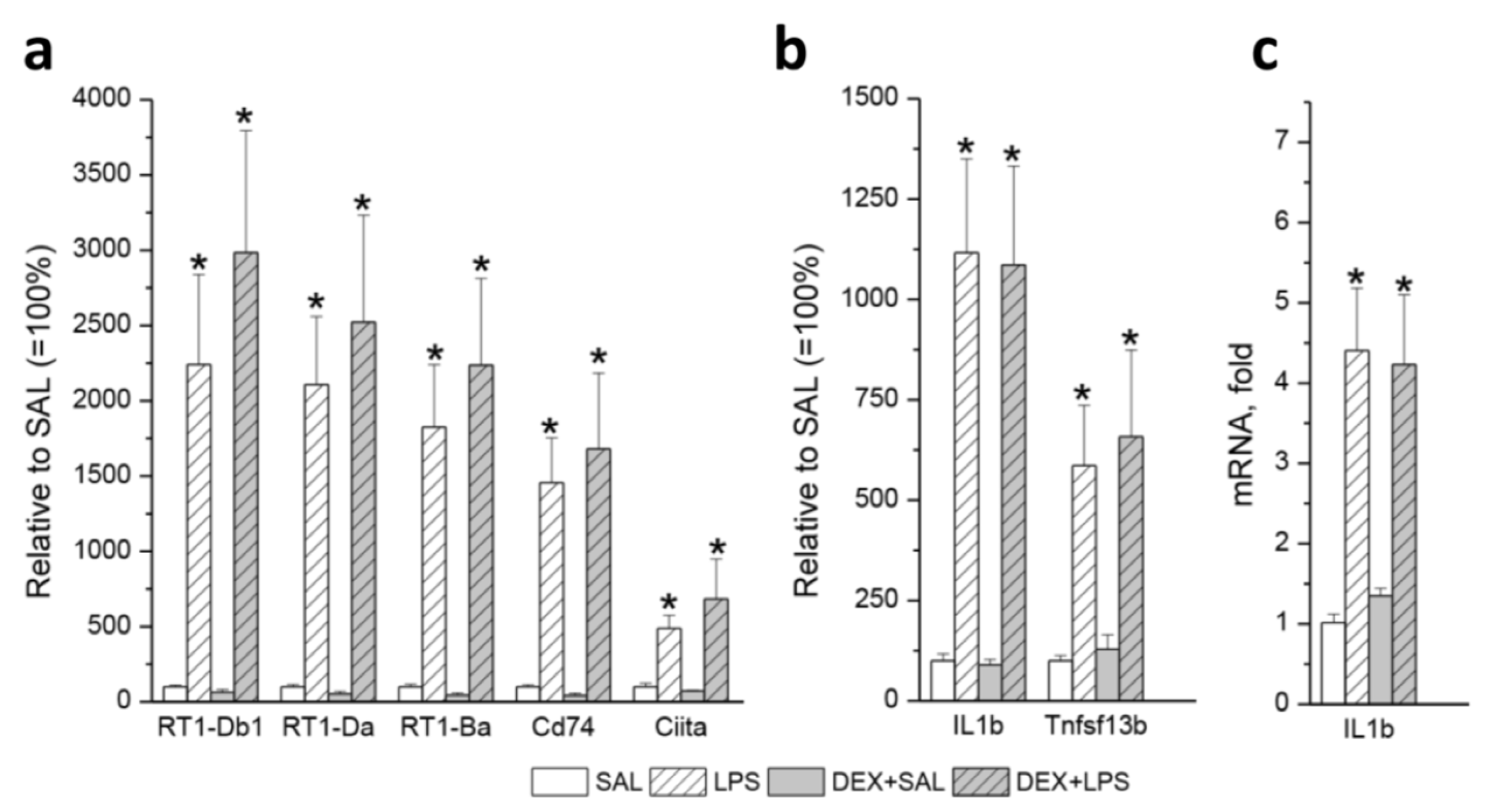
Figure 7.
Protein-protein interaction (PPI) networks: (a) Two k-means clusters of 24 DEGs between LPS and SAL. (b) Network of the 16 DEGs shared between LPS vs. SAL and DEX+LPS vs. DEX+SAL. Colors indicate association of DEGs with several GO biological processes: red - GO:0006955 Immune response, 14 DEGs; FDR = 1.74e-11; blue - GO:0006952 Defense response, 11 DEGs, FDR = 1.63e-07; green - GO:0006954 Inflammatory response, 7 DEGs, FDR = 2.53e-05; yellow - GO:0051716 Cellular response to stimulus, 14 DEGs, FDR = 0.0024; violet - GO:0051384 Response to glucocorticoid, 4 DEGs, FDR = 0.0048. FDR: false discovery rate. .
Figure 7.
Protein-protein interaction (PPI) networks: (a) Two k-means clusters of 24 DEGs between LPS and SAL. (b) Network of the 16 DEGs shared between LPS vs. SAL and DEX+LPS vs. DEX+SAL. Colors indicate association of DEGs with several GO biological processes: red - GO:0006955 Immune response, 14 DEGs; FDR = 1.74e-11; blue - GO:0006952 Defense response, 11 DEGs, FDR = 1.63e-07; green - GO:0006954 Inflammatory response, 7 DEGs, FDR = 2.53e-05; yellow - GO:0051716 Cellular response to stimulus, 14 DEGs, FDR = 0.0024; violet - GO:0051384 Response to glucocorticoid, 4 DEGs, FDR = 0.0048. FDR: false discovery rate. .
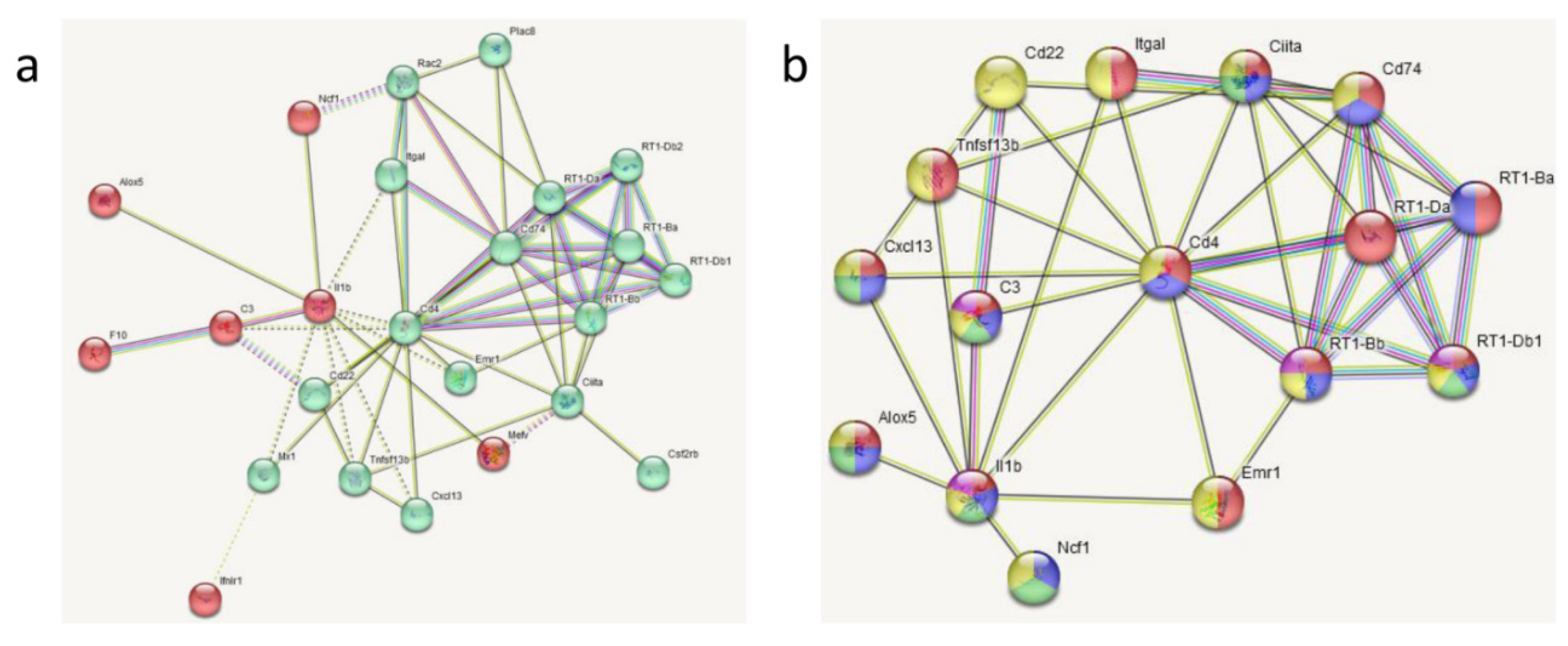
Figure 8.
Responses of representative genes to LPS or DEX alone, as well as to LPS with DEX pretreatment (relative to SAL group). (a) Data from RNA-sec; (b) verification of the RNA-sec results for Slc1a2 by quantitative PCR (correlation between sec and PCR expression values was 0.63, p < 0.05).
Figure 8.
Responses of representative genes to LPS or DEX alone, as well as to LPS with DEX pretreatment (relative to SAL group). (a) Data from RNA-sec; (b) verification of the RNA-sec results for Slc1a2 by quantitative PCR (correlation between sec and PCR expression values was 0.63, p < 0.05).
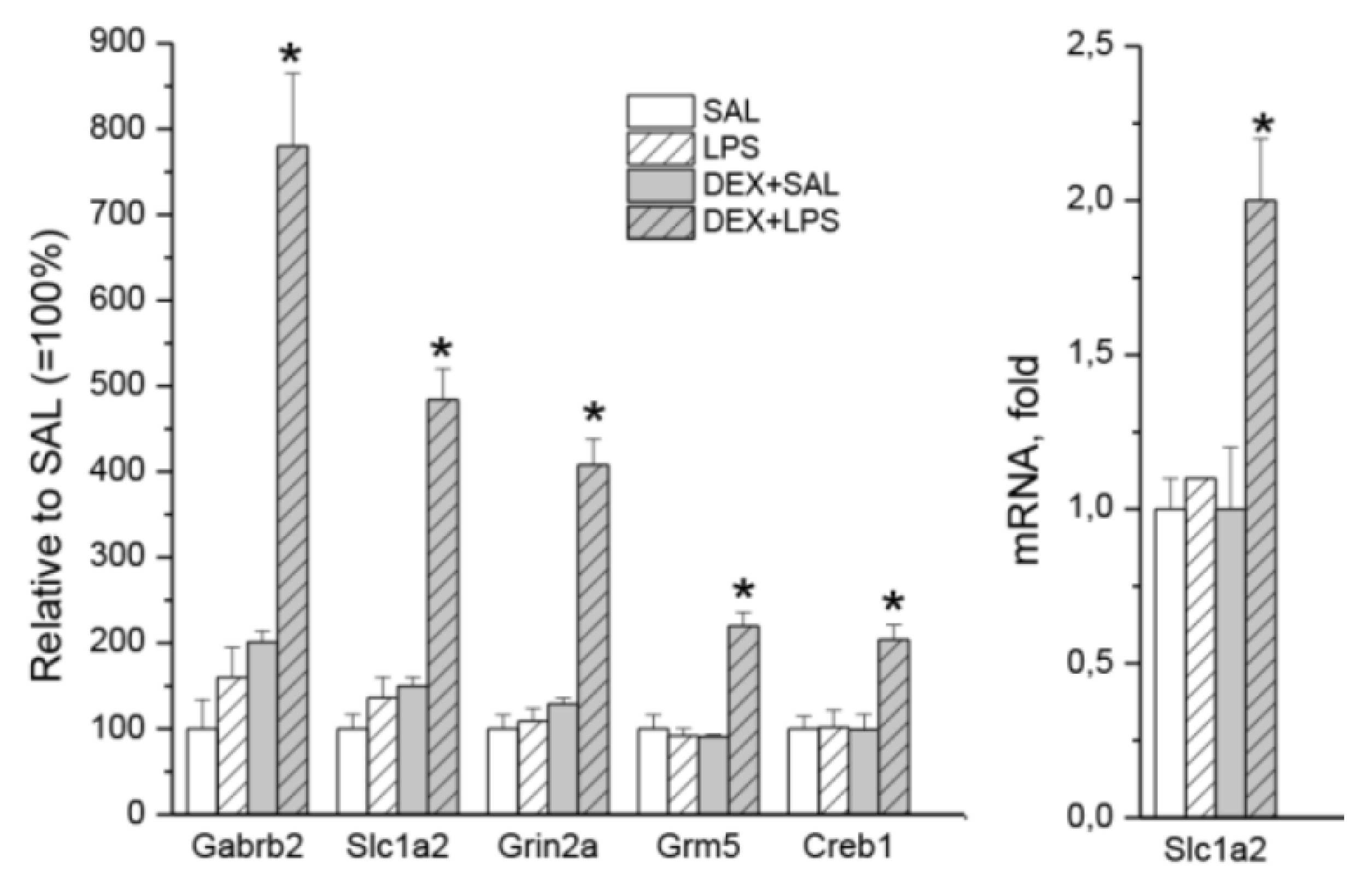
Figure 9.
Protein-protein interaction (PPI) network of the DEGs specific for DEX+LPS vs. DEX+SAL (k-means cluster 1, 21 DEGs). Colors indicate association of DEGs with several GO biological processes: red – GO:0034762 Regulation of transmembrane transport, 10 DEGs (Cacna1e, Cacna2d1, Erbb4, Grin2a, Grm5, Kcnh5, Kcnh7, Kcnq3, Slc1a2, Wnk3), FDR = 1.11e-06; blue – GO:0050890 Cognition, 5 DEGs (Cacna1e, Chl1, Creb1, Grin2a, Grm5) , FDR = 0.0165; violet - GO:0007612 Learning, 4 DEGs (Cacna1e, Creb1, Grin2a, Grm5) , FDR = 0.0198; green - GO:0022008 Neurogenesis, 9 DEGs (Cdkl5, Chl1, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Unc5d), FDR = 0.0223; yellow - GO:0007399 Nervous system development, 10 DEGs (Chl1, Cdkl5, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Slc1a2, Unc5d), FDR = 0.0340. FDR: false discovery rate.
Figure 9.
Protein-protein interaction (PPI) network of the DEGs specific for DEX+LPS vs. DEX+SAL (k-means cluster 1, 21 DEGs). Colors indicate association of DEGs with several GO biological processes: red – GO:0034762 Regulation of transmembrane transport, 10 DEGs (Cacna1e, Cacna2d1, Erbb4, Grin2a, Grm5, Kcnh5, Kcnh7, Kcnq3, Slc1a2, Wnk3), FDR = 1.11e-06; blue – GO:0050890 Cognition, 5 DEGs (Cacna1e, Chl1, Creb1, Grin2a, Grm5) , FDR = 0.0165; violet - GO:0007612 Learning, 4 DEGs (Cacna1e, Creb1, Grin2a, Grm5) , FDR = 0.0198; green - GO:0022008 Neurogenesis, 9 DEGs (Cdkl5, Chl1, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Unc5d), FDR = 0.0223; yellow - GO:0007399 Nervous system development, 10 DEGs (Chl1, Cdkl5, Creb1, Erbb4, Gabrb2, Grin2a, Grm5, Kcnq3, Slc1a2, Unc5d), FDR = 0.0340. FDR: false discovery rate.
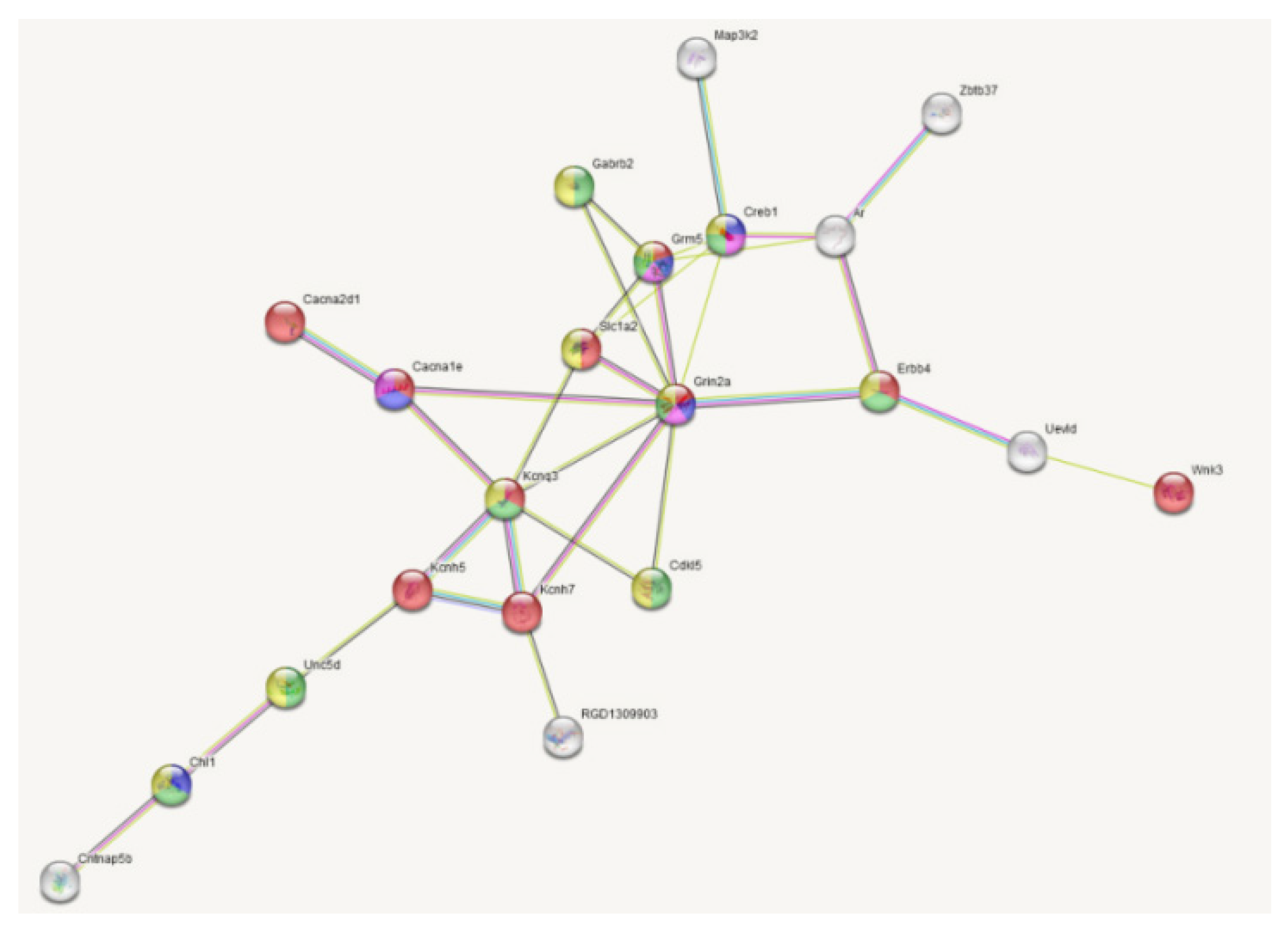

Table 1.
The top genes ranked with the interaction degree method.
| Gene | LPS vs. SAL (33) | DEX+LPS vs. DEX+SAL (161) | Common DEGs (22) | Specific DEGs (84) |
|---|---|---|---|---|
| Cd4 | 16 | 21 | 13 | - |
| Cd74 | 11 | 14 | 8 | - |
| Ciita | 8 | 11 | 6 | - |
| Creb1 | - | 11 | - | 7 |
| Il1b | 10 | 15 | 8 | - |
| RT1-Bb | 8 | 11 | 7 | - |
| RT1-Da | 9 | 9 | 6 | - |
| Grin2a | - | 9 | - | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



