
Submitted:
03 August 2023
Posted:
08 August 2023
You are already at the latest version
Abstract
The aim of the present study was to evaluate the effects on the presence and distribution of oxyntopeptic cells (OPs) and enteroendocrine cells (EECs) expressing GHR, NPY and SOM in the gastric mucosa of European seabass and seabream of dietary levels of bioactive peptides (BPs) obtained from salmon processing by-products (in silico approach). In this study, 27 seabass and 27 seabreams were divided into three experimental groups: a control group (CTR) fed a control diet and two groups fed different levels of BP to replace fishmeal: 5% BP (BP5%) and 10% BP (BP10%). The stomach of each fish was sampled and processed for immunohistochemistry. Some SOM, NPY and GHR-IR cells exhibited alternating “open type” and “closed type” EECs morphologies. In seabream gastric mucosa, the BP10% group (16.8 ± 7.5) showed a significant change in NPY-IR cell number compared to CTR (CTR 8.5 ± 4.8) and BP5% (BP10% vs. CTR P ≤ 0.01; BP10% vs. BP5% P ≤ 0.05). In seabream, SOM-IR cells in the BP 10% diet (16.8 ± 3.5) differed from those in CTR (12.5 ± 5) (CTR vs. BP 10% P ≤ 0.05) and BP 5% (12.9 ± 2.5) (BP 5% vs. BP 10% P ≤ 0.01). In seabass, EEC SOM-IR cells increased at 10 % BP (5.3 ± 0.7) compared to 5 % BP (4.4 ± 0.8) (5 % BP vs. 10 % BP P ≤ 0.05). The results obtained suggest that salmon BPs have a promising impact as circulating ingredients for seabass and seabream.
Keywords:
gilthead seabream and European seabass
; bioactive peptide
; oxyntopeptic cells
; neuropeptide Y
; somatostatin and ghrelin
1. Introduction
The fishing industry has grown steadily over the last decade. This growth has been accompanied by a high volume of protein-rich by-products. This waste includes whole or parts of fish such as fillets, skin and fins, bones, heads, guts and scales. In a context of circular economy, these by-products rich in proteins, which can be recovered / reused as functional ingredients in, feed industry. Recently, several studies have reported the utilisation of fish by-products by enzymatic hydrolysis for the recovery of various valuable components, and fish protein hydrolysates such as capelin, mackerel, hoki frame, jumbo squid, yellow stripe trevally and tuna liver have been shown to possess antioxidant activity with the ability to scavenge hydroxyl radical, superoxide anion radical, hydrogen peroxide and chelate metal ions [1,2,3,4,5,6]. Fishery proteins represent a potential source of biopeptides.
Previous studies have described the biological activity of marine protein hydrolysates produced from different species [7,8,9]. Due to their low molecular weight compounds and balanced amino acid profile, hydrolysates have been used as chemoattractants and fishmeal replacers in aquafeed [10,11,12,13,14].
State-of-the-art techniques in peptidomics and bioinformatics (often referred to as the in silico approach) have been used to identify peptides with predicted anti-inflammatory, immunostimulatory or antimicrobial properties in the different by-product fractions. In this context, interest in the in silico approach has increased because it is less costly and time consuming [15,16].
With its high nutritional value and potential pharmacological applications, farmed Atlantic salmon is a popular food around the world. The fileting operation generates large amounts of by-products consisting of head, backbone, skin and viscera. Mechanically separated salmon muscle is readily available as a low-cost by-product of the filleting process and can account for up to 60% (w/w) of filleting waste [17]. Several studies have evaluated the biochemical functional properties of salmon muscle protein hydrolysate (and their respective peptides) [18,19,20] and there is growing industrial interest in the utilization of bioactive peptides (BPs) within the fish feed and pet food industry. Recently, in European seabass the dietary inclusion level of bioactive peptide from farmed Atlantic salmon have shown the possibility of an almost total replacement of fish meal in a plant-based diet in terms of growth and feed efficiency [21].
The regulation of food intake relies heavily on the gut-brain axis. Several studies have highlighted the important role played by gut hormones in response to food digestion. [22,23]. These hormones are involved in appetite regulation as short-term peripheral satiety signals. They promote satiety, i.e., a decrease in appetite and a reduction in food intake, through endocrine and nervous pathways by activating various signalling pathways. [24,25,26,27]. In vertebrates, appetite and digestion are controlled by the enteroendocrine system [28,29]. “Gut-brain hormones such as ghrelin (GHR), neuropeptide Y (NPY), somatostatin (SOM), cholecystokinin, etc. are important factors in the control of feeding behaviour [28,29]. GHR, NPY and SOM have been found in a large number of fish species. Their tissue distribution supports the idea that GHR has an integrative role in the regulation of energy balance at both central nervous system and systemic levels [30]. NPY and GHR are reported to correlate positively with feed intake [28]. GHR, a small peptide hormone secreted by the stomach, is an appetite stimulator [31]. GHR levels increase before a meal and decrease postprandially [32] and are involved in the regulation of appetite, energy balance and body weight [33]. NPY is a potent, highly conserved, multi-functional peptide found in vertebrates, including fish. It plays an important role in the regulation of feeding behaviour, energy metabolism and digestive processes [34,35,36,37]. In fish, SOMs have many direct and indirect effects on intermediary metabolism and feeding behaviour. In general, SOM inhibit food intake and promote catabolic processes (e.g., mobilizing stored lipid and carbohydrate) [36,38].
Certain hormone-like peptides obtained by protein hydrolysis have the potential to affect gastrointestinal (GI) motility, endocrine metabolism, intake and animal performance [39]. Bioactive peptides (BPs) share some structural characteristics. These include a peptide residue length between 2-20 amino acids [40] and the presence of hydrophobic amino acids besides proline, lysine or arginine groups [41].
Regulatory peptides with hormone-like activity (hormone peptides) or the ability to modulate blood levels of certain hormones could also be obtained by enzymatic hydrolysis. The biological activity of hormone-like peptides is typically mediated by their interaction with G protein-coupled receptors (GPCRs) on the cell surface and further activation of the ligand-receptor signalling pathway to regulate various physiological functions of the body [42]. Fish proteins may also serve as a reservoir of hormone-like peptides. Interestingly, some authors have found neuropeptide immunorelated molecules and molecules capable of binding to specific hormone receptors on cell membranes in fish protein hydrolysates [43,44,45]. In this study, we investigated the effect of supplementing salmon by-products derived from enzymatic hydrolysis on the presence and distribution of oxytocic (OP) and enteroendocrine (EEC) cell subpopulations expressing GHR, NPY and SOM in the gastric mucosa of European seabass and seabream.
2. Materials and Methods
2.1. Protein hydrolysate
Fresh Atlantic salmon (Salmo salar) heads and backbones were purchased from Biomega (Øygarden, Norway). Grinded raw material was mixed with tap water (1:1), heated to 50 °C and added Chymotrypsin (0.1 % w/w; Enzyme Supplies, UK). Hydrolysis was carried out for 60 min, followed by inactivation of enzyme activity at T > 90 °C for 10 min. The water phase (hydrolysate) was separated from the lipid and bone phases and dried to a dry powder in a NIRO P-6.3 spray dryer (GEA, Denmark) with inlet and outlet temperatures of 200 and 92 °C, respectively.
2.2. Feed production
The diets were formulated with FM and with a mixture of vegetable ingredients that are currently used in aquafeed for European seabass and gilthead seabream [46,47]. The control diet (CTR) was formulated to resemble a commercial feed for European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata) (Table 1). Two experimental feed mixes were formulated with exchange of the fishmeal with the experimental fish protein hydrolysate at 5% (BP5) and 10% (BP10) level, respectively [21]. Composition of fish protein hydrolysate are reported in Table 2. The diets were conditioned in an atmospheric double differential preconditioner (Wenger Manufacturing Inc., Sabetha, KS) prior to extrusion on a TX-52 twin screw extruder (Wenger) and expanded through 2.5 mm dies to 3.2 mm pellets. The pellets were dried in a hot air double layer carousel dryer (Model 200.2, Paul Klockner GmbH, Nistertal, Germany) at constant air temperature and vacuum oil coated to achieve the final lipid content. Diets were formulated with FM and a mixture of vegetable ingredients currently used in aquafeed for European seabass and seabream [46,47].
2.3. Chemical analysis
The nitrogen content was analysed by the Kjeldahl method (ISO 5983-2, 2009) and the crude protein content was estimated on the basis of N × 6,25. Ash was determined by combustion of the raw material at 550 °C (ISO 5984-2, 2002). The dry matter content was determined by drying at 103 °C (ISO 6469-2, 1999). Fat content of the protein hydrolysate was analyzed by the EU method (Commission Directive 98/64/EC), while fat content of the raw material was analyzed based on ethyl acetate extraction (NS 9402). Peptide size distributions were measured by HPLC size exclusion chromatography (SEC) (1260 series HPLC Agilent Technologies) using a Superdex Peptide 10/300GL column (GE Healthcare, Uppsala, Sweden). The eluent was acetonitrile with TFA and UV detection at 190-600 nm [48].
2.4. Rearing condition and sampling
Juvenile European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata) were collected from an Italian commercial hatchery and reared in recirculating aquaculture systems (RAS) at the Aquaculture Laboratory of Cesenatico, Department of Veterinary Sciences, University of Bologna, Italy. At the beginning of the experimental procedures, sixty seabass and sixty seabreams were individually weighted and allocated, based on the species, in each of eighteen conical-bottomed tanks with a volume of 800 litters connected to a RAS supplied with natural sea water (overall water volume: 22 m3; oxygen level 8.0 ± 1.0 mg L-1; temperature 24 ± 0.5 °C, salinity 28-32 g L-1) according to Busti et al. [49]. Over a period of 58 days, each experimental diet was administered twice daily until full satiety using an overfeeding approach employed for both species as described in Parma et al. [46,47]. At the end of the trial, three fish per tank were taken at random for a total of 27 seabass (average weight 147.1 g) and 27 seabreams (average weight 168.76 g). Subsequently, the previously selected fish were sacrificed under anesthesia (excess of anesthetic MS222, 300 mg L-1). The GI tract from the esophagus to the posterior intestine was gently removed from each seabass and seabream and isolated from the coelomic cavity. Stomachs were isolated and fixed in formalin (pH 7.2) for 48 hours at room temperature. After fixation, the stomach was divided symmetrically by cutting along the long axis to obtain two equal halves. Stomach samples were dehydrated in a graded alcohol series, cleared in xylene and paraffin-embedded. Sections (6 µm thick) placed on polylysine slides were obtained from each block of paraffin.
The Ethical-Scientific Committee for Animal Experimentation of the University of Bologna, Italy (ID 113/2020-PR) evaluated and approved all experimental procedures.
2.5. Feed intake and growth calculation
The calculations used to determine the various performance parameters were as follows
Specific growth rate (SGR) (% day-1) = 100 * (ln FBW–ln IBW) / days (where FBW and IBW are the final and initial body weights, respectively);
Feed intake (FI) (g kg ABW-1 day-1) = ((100 ∗ total intake)/(ABW))/days) (where average body weight, ABW = (IBW + FBW)/2;
Feed conversion ratio (FCR) = feed intake/weight gain; protein efficiency.
2.6. Immunohistochemistry
Stomach sections were processed for double labelling immunofluorescence. Table 3 lists the primary and secondary antibodies used in this study. Sections were deparaffinised, rehydrated and incubated with appropriate normal serum (5% normal goat or donkey serum) and 1% BSA diluted in PBS (phosphate buffer saline 0.01M pH 7.4) for 1 hour at room temperature (RT) to reduce non-specific secondary antibody binding. Sections were then incubated for 48 h at 4°C in a humidity chamber with the following primary antibodies: rabbit anti Na+/K+-ATPase 1:600, rat anti SOM 1:500, mouse anti GHR 1:500 and goat anti NPY 1:1000. After washing, the sections were incubated for 1 h at RT with goat anti-rabbit AlexaFluor 594 at 1:1000, donkey anti-goat AlexaFluor 488 at 1:1200, donkey anti-mouse AlexaFluor 594 at 1:1200, or goat anti-rat at 1:1000. Finally, the sections were coverslipped with buffered glycerol, pH 8.6. After performing some preliminary tests, since we observed that some immunoreactive NPY cells coexpressed GHR, anti-NPY and anti-GHR antibodies were used simultaneously to evaluate the EECs that colocalized NPY / GHR, while the rat antiserum anti-SOM was used together with Na+K+-ATPase antibody, to characterize the distribution of OPs which express SOM.
2.7. Threshold binarization procedure
The following method was used to characterise the area occupied by the immunoreactive (-IR) of OPs in the gastric mucosa. Slides were scanned with a Nikon DS-Qi1Nc digital camera at 20× magnification using NIS Elements software BR 4.20.01 (Nikon Instruments Europe BV, Amsterdam, The Netherlands). Automated image binarisation was applied to a region of each selected stomach image using NIS Elements software BR 4.20.01. Image binarisation is a widely used method for distinguishing objects of interest from the background. In fact, it determines a grey threshold and assigns each pixel of a digital image to a class (image objects). If it is a grey value, it is greater than the threshold determined in relation to the other class (image background). Specifying the correct threshold limits is a crucial procedure in automated image analysis, used to determine which pixels are included in the binary layer and which are not. In our case, binarisation allowed us to separate the pixels representing the OPs-IR of the gastric surface (brighter pixels) from those representing the rest of the section. The measurement area can be restricted by a user-defined region of interest (ROI). Within the ROI, it is possible to binarise only the selected part and not include other parts of the section. This allowed quantification of the gastric surface covered by the OPs-IR area. Gastric morphometric assessments were performed in a blinded fashion by two investigators.
2.8. Antibody specificity
Specificity for Na+K+-ATPase, GHR, SOM antibodies was previously demonstrated by Mazzoni et al. [50]. The specificity of the NPY antibodies was demonstrated by the absence of immunostaining when the antibodies were preabsorbed with an excess of the homologous peptide. Omitting the primary antibody prevented the secondary antibody from binding.
2.9. Morphometric evaluations and statistical analysis
The specimens were examined using a Nikon Eclipse Ni microscope and images were taken using a Nikon DS-Qi1Nc digital camera and NIS Elements software BR 4.20.01 (Nikon Instruments Europe BV, Amsterdam, Netherlands). Minor adjustments to contrast and brightness were made using Corel Photo Paint, while figure panels were prepared using Corel Draw (Corel Photo Paint and Corel Draw, Ottawa, ON, Canada). The 20X objective was used for morphometric evaluation. In the gastric mucosa, the area occupied by OPs-IR in 4.1 mm2 (0.410 x 10 fields) was measured by binarisation (described above). In addition, the number of GHR, NPY and SOM IRs in 4.1 mm2 was counted in the gastric mucosa.
For each experimental group (CTR, BP 5% and BP 10%), the values obtained for the OPs-IR area and the number of EECs were grouped and the means were calculated. The results were expressed as mean ± standard deviation (SD). Data were analysed by one-way ANOVA (Graph Prism 4, GraphPad Software, Inc., La Jolla, CA, USA): Experimental group was considered as the main effect. In addition, the means were then separated using the Tukey-HSD test. A P ≤ 0.05 was considered statistically significant.
3. Results
3.1. Protein hydrolysate
The obtained spray dried protein hydrolysate contained 82.8% protein of which 82.5% was water soluble (Table 2). Fat level was comparable to a normal fishmeal; however, the ash level was lower due to the removal of bones by centrifugation after the hydrolysis process. No attempt was made to reduce the fat level further, however, an additional separation step has the potential to increase the protein level above 90%. Molecular weight distribution of the soluble protein fraction showed minor parts (5.2%) above 4 kDa and a high fraction (54%) below 1 kDa (Table 2).
3.2. Feed intake and growth
Table 4 presents data on feed intake and growth parameters. Feed intake and growth (final bodyweight, weight gain and SGR) were not significantly different between treatments or within species.
3.3. Seabream and seabass morphological features
The gastric mucosa of the seabass as well as that of the seabream is lined with prismatic epithelium composed of poorly stained cells with a nucleus in a basal position. These epithelial cells, probably responsible for the production of mucin, line the lumen of the stomach: these cells perform a protective function by interposing themselves between the gastric glands and the contents of the stomach. Below the epithelial layer, simple tubular gastric glands (in some cases bifid) have been observed over the entire surface of the stomach. Below the latter, a robust muscularis mucosae delimits the passage between the mucosa and submucosa.
Immunofluorescence with the Na+/K+-ATPase antibody confirmed the presence of OPs in all parts of the stomach: indeed, they showed intense immunoreactivity and appeared to be distributed along the simple tubular gastric glands (Figure 1B,C). However, the immunoreactivity of OPs is abruptly disrupted in the transition zone between the gastric and esophageal mucosa as well as at the gastrointestinal junction.
EECs-IR have been observed mainly above the glandular adenomers, while a smaller number of these cells are found mixed with OPs. Generally, EECs intermingled between the mucous cells tend to reach the endoluminal side (Figure 2 and Figure 3). These EECs located in correspondence of the endoluminal surface show an “open type” morphological aspect (“open type” EECs). These endocrine elements have an elongated or pyriform shape with two cytoplasmic extensions: one extension tends to reach the endoluminal side, insinuating itself between the epithelial cells, while the other, moving in the opposite direction, reaches the basement membrane. Other EECs exhibit a “closed type” morphological features (“closed type” EECs). These cells, unlike the previous ones, do not show any cytoplasmic extension and have a rounded shape. Generally, SOM- and NPY-IR cells show a morphological appearance of “open type” EECs, while most GHR-IR cells show, on the contrary, appearance of “closed type” EECs (Figure 2 and Figure 3). In addition, some NPY-IR cells were observed to co-express GHR and vice versa (Figure 3). Finally, these analyses surprisingly highlighted that the seabream gastric mucosa appears to have a much higher number of EECs
3.4. Seabream morphometric results
From the evaluation of OPs-IR, it emerged that the integration of 5% and 10% of BP did not change the area occupied by OPs in the gastric mucosa of seabream. In detail CTR 0.17 ± 0.02; BP 5% 0.14 ± 0.05; BP 10% 0.16 ± 004) (Figure 4 A).
Regarding the number of EECs, we observed that the 10% BP group showed a statistically higher average number of NPY-IR cells (16.8 ± 7.5) comparing with CTR group (CTR 8.5 ± 4.8) (CTR vs BP 10%; P ≤ 0.01), and, similarly with respect to the BP 5% group (10 ± 5.3) (BP 5% vs BP 10% P ≤ 0.05) (Figure 4 B). Likewise, compared to observed for the NPY-IR EECs, also the EECs expressing SOM were on average more numerous in the 10% BP diet (16.8 ± 3.5) and significantly different from the experimental CTR group (12.5 ± 5) (CTR vs BP 10% P ≤ 0.05) and to the experimental group 5% (12.9 ± 2.5) (BP 5% vs BP 10% P ≤ 0.01) (Figure 4 C). No significant differences were observed regarding the mean number of GHR-IR cells in the three experimental groups (CTR 10.3 ± 3; BP 5% 9.8 ± 2.9 and BP 10% 12.4 ± 3.4, respectively) (Figure 4 B). The percentage of colocalization, expressed as the number of NPY-IR cells that co-express GHR out of the total of GHR-IR cells, was 11.6% in the CTR group (84/723), 12% in the BP 5% group (84 / 702) while 13.6% in the BP 10% group (121/891).
3.5. Seabass morphometric results
As for the seabream, no statistical differences were observed in the seabass concerning the presence and distribution of OPs-IR within the three experimental groups. The following values were obtained: CTR 0.16 ± 0.02; BP 5% 0.17 ± 0.01; BP 10% 0.16 ± 0.01 (Figure 4 D).
Unlike what observed in seabream, no significant differences were observed in the seabass gastric mucosa regarding the EECs expressing NPY (CTR 1.6 ± 0.4; BP 5% 1.4 ± 0.3; BP 10% 1.6 ± 0.4) and GHR (CTR 4.1 ± 0.7; BP 5% 4 ± 0.6; BP 10% 4.2 ± 0.6); it should be noted that there was only a slight increase in GHR-IR cells in the 10% BP diet (Figure 4 E). We evaluated the percentage of cells co-expressing NPY / GHR on the total of EECs GHR-IR: (CTR 14.4%, 38/263; BP 5%, 28/289; BP 10%, 31/304).
EECs SOM-IR increased in the diet BP 10% (5.3 ± 0.7) compared to the BP 5% experimental group (4.4 ± 0.8) (BP 5% vs BP 10% P ≤ 0.05) (Figure 4 F).
4. Discussion
In aquaculture, fish protein hydrolysates have been evaluated as fishmeal replacers and attractants in several fish species [14,51,52,53,54,55]. Due to their good nutritional value and functional properties, protein hydrolysates derived from marine processing by-products have been considered as an excellent ingredient in aquafeed [7]. Previous studies have shown a significant improvement in the apparent digestibility of nutrients through the supplementation of dietary hydrolysates in the diets of red seabream [56].
Most of the EECs dispersed in the mucosa of the GI tract can directly sense nutrients at their apical pole, which faces the lumen of the gut, and can be induced to release hormones into the circulation from their basal pole. Receptors of the G-protein-coupled receptor (GPCR) class have been implicated in EEC protein/amino acid sensing [57,58]. After enzymatic digestion of dietary proteins in the gut lumen, these EECs may be an easy target for functional peptides. Some EEC subtypes (e.g., GHR, SOM and enterochromaffin cells) do not contact the gut lumen [57]. In this context, also free amino acids can exert similar effects on EECs [58,59,60,61]. Some in vitro studies have shown that intact proteins and partially hydrolysed proteins are more effective in inducing enteric hormone secretion than the products of their complete hydrolysis. Some selected synthetic peptides are more effective than the corresponding free amino acids [62,63,64].
The T1R1/T1R3 and T1R2/T1R3 heterodimers, calcium-sensing receptor (CaSR) and GPRC6A receptor, can sense amino acids in the extracellular milieu and show partial selectivity for ligands [58]. In addition, in EECs in the stomach and small intestine, the GPR39 taste receptor is highly expressed. It is suggested that the GPR39 taste receptor is activated more by peptides than by free amino acids [57,65].
Both long-term and short-term pathways have been identified to regulate energy intake: long-term pathways inform the brain about body mass fluctuations, whereas short-term pathways relate to energy available in the GI. Both pathways use hormones like insulin and leptin for long-term control and GHR, cholecystokinin and glucagon-like peptide-1 for short-term control [66].
It has been reported that the concentration of amino acids in the plasma is higher and rises more rapidly after the ingestion of peptide hydrolysates than after the ingestion of the corresponding whole proteins [67]. Moughan et al. [67] based their conclusions on the observation that solutions of peptide hydrolysates undergo a more rapid gastric emptying and intestinal absorption of their constituent amino acids than do the complete proteins from which they are derived.
One of the most highly conserved neuropeptides in vertebrates [69,70] is NPY, a peptide of 36 amino acid residues first isolated from pig brain [68]. In some Teleosts, two classes of NPY (NPYa and NPYb) have been identified. However, teleost fish such as goldfish (Carassius auratus) and zebrafish (Danio rerio) have only NPYa [71,72]. NPY is known to be mainly secreted by the neurosecretory cells of the hypothalamus and is secreted in response to hunger [73,74]. Its primary function as a signalling factor is the regulation of a variety of biological processes such as food intake, the circadian rhythm, neuroendocrine functions and glucose homeostasis [75]. The NPY gene has been shown to be expressed in many tissues of several Teleosts. It is expressed in the central nervous system, intestine, liver, spleen, skeletal muscle and adipose tissue of several fish species, including zebrafish, goldfish (Carassius auratus), Atlantic salmon (Salmo salar), catfish (Ictalurus punctatus) and tilapia (Oreochromis sp.) [76,77,78,79]. In addition, fish NPY receptors are located in the brain, but can also be found in peripheral tissues, including the eye and intestine [80,81,82,83]. The GI tract is the largest peripheral NPY-producing organ, with distribution in all layers. In mammals, the distribution is greater in the submucosal and myenteric plexus neurons [84,85], whereas in fish, the greater distribution is observed in the EECs of the first intestine segments, as reported in pejerrey, Odontesthes bonariensis [86], R. quelen [87], dorado, Salminus brasiliensis [88], milkfish, Chanos chanos [89] and Nile tilapia (O. niloticus) [90]. In a fasting-refeeding study carried out in the blunt-nosed seabream (M. amblycephala), no differences were found in the expression of intestinal NPY, whereas the expression levels in the brain increased after fasting and decreased after refeeding [91]. These authors suggested that central NPY would act as an orexigenic factor. However, intestinal NPY would not act as a brain-gut peptide to stimulate appetite [92]. A high number of NPY cells in the gut may indicate that this region would act as a primary source of peripheral signals to stimulate feeding in the absence of food in this location, according to Vigliano et al. [86]. This suggests that the regulation of fish feeding is the result of the integration of different endocrine responses. There may be fish and species-specific mechanisms of agonism and antagonism [93,94]. It is reasonable to assume that the salmon-derived BPs tested in this trial stimulated, in the gastric mucosa, EECs expressing NPY differently in seabream than in seabass.
In this study, no differences were observed regarding the number of EECs GHR-IR in the experimental diets compared to the CTR. In overweight/obese individuals, Jensen et al. [95] hypothesised that supplementation with cod protein hydrolysate for 8 weeks would have a suppressive effect on postprandial GHR levels. These potentially beneficial effects were hypothesised to occur due to the presence of small peptides, mainly di- and tripeptides, which are rapidly absorbed from the GI tract and may affect appetite regulation pathways. The authors showed that fasting and postprandial concentrations of acylated GHR were not affected by 4 g of cod protein hydrolysate for 8 weeks. Jensen et al. [95] reported the effect of low-dose cod protein hydrolysate supplementation on fasting and postprandial acylated GHR levels: they observed no effect on fasting or postprandial acylated GHR levels. On the other hand, a single dose of 20 mg/kg body weight of cod protein hydrolysate had no effect on postprandial acylated GHR levels or hunger-related sensations in comparison with the control group [33].
The increase in GHR observed in this study (although not statistically significant) is a positive aspect given the abundant evidence that GHR is an orexigenic peptide in several fish species, despite the data reported in the literature.
SOM has a primary role in the regulation of endocrine and exocrine secretion: it contributes to the reduction of gastric acid secretion and gastric motility and inhibits the secretion of several gastro-enteric and pancreatic hormones [96]. In the stomach, several regulatory peptides act in combination to regulate the secretion of acid and enzymes by the OPs [97,98] and the digestive processes in the small intestine, thereby optimising the digestive functions of the entire GI tract.
Luminal application of glutamate in the rat stomach has previously been reported to increase the activity of the afferent fibres of the gastric branch of the vagus nerve by Uneyama et al. [99]. Interestingly, although each of the amino acids responded to luminal application in the gut [100,101], other amino acids did not show such effects. Thus, of the 20 dietary amino acids that are part of the body’s proteins, only glutamate can transmit nutritional information from the stomach to the brain. However, the exact mechanism for the luminal sensing of glutamate is still unclear [102].
Nakamura et al. [102], observed, in D (SOM-producing) cells obtained by fractionation of rat gastric cells, highly expression of the CaSR, GPRC6A and Gi-coupling mGluRs receptors. Notably, CaSR expression was much higher on D cells than on parietal cells. The same authors also observed, in primary D cell cultures, that some specific CaSR amino acid (such as phenylalanine, tryptophan and histidine) significantly stimulated SOM release more than twofold, while other aminoacid receptor (e.g., GPRC6A) favourable for the lysine had no effect. They conclude that in the stimulation of SOM release in D cells, only the interaction between amino acids and specific receptors plays a functionally important role. Since SOM is an inhibitory regulator of gastric exocrine and endocrine secretion [103,104,105,106], it is reasonable to assume that inhibiting SOM release via amino acid coupled specific amino acid receptors (i.e., inhibiting the inhibitory effect) results in stimulating gastric secretion. This may partly explain the results we have obtained. In the gastric mucosa of seabream and seabass we did not observe any changes of OPs in fish fed with 10% BP (and BP 5%) despite the increase in the number of SOM-IR EECs.
5. Conclusions
For the first time, the number and distribution of NPY, SOM and GHR-IR cells in the gastric mucosa of seabass and seabream have been shown to be affected by the administration of a BP-supplemented diet. Based on the data obtained, it is plausible to hypothesize that BPs perform, directly or indirectly, an action that stimulates the production of NPY, GHR and SOM without modifying the expression of OPs.
To gain more knowledge and to better understand the mechanisms of action of different bioactive peptides on the morphology of the gastric mucosa of fish, further studies are certainly needed.
In summary, the data suggest that salmon-derived BPs have a promising future as dietary ingredients for European seabass and seabream. The study also provides new insights into the morpho-functional changes that occur in the digestive tract of these species in response to feeding stimuli.
Author Contributions
Conceptualization, M.M., G.L., A.B., L.P., S.B.; methodology, A.O., O.H.R., T.A., A.B., L.P., P.C..; software, M.M., G.L. A.B., L.P.; validation, M.M., A.B., L.P., formal analysis M.M., G.L., A.B., L.P., S.B.; investigation, M.M.., G.L., L.P., S.B.; resources, M.M., G.L., A.B., L.P., S.B. A.O., O.H.R., T.A.; Data curation, M.M., S.B., L.P., C.T., A.B; Writing—Original Draft, M.M.; writing—review & editing, M.M., L.P., P.C., P.G.; supervision, M.M., A.O., O.H.R., T.A., A.B., L.P., P.C.; project administration, A.B., A.O., O.H.R., T.A.; Funding acquisition, A.B., P.C., A.O., O.H.R., T.A. All authors have read and agreed to the published version of this manuscript.
Funding
This research was carried out as part of the Novofeed project, funded by Eranet–Marine Biotech under the European Commission’s 7th Framework Programme. The Norwegian Research Council is acknowledged for its support to the Aquafeed Technology Centre, ATC, with grants nos. 604814 and 245883/F50.
Institutional Review Board Statement
The Ethical and Scientific Committee for Animal Experimentation of the University of Bologna, Italy (ID 113/2020-PR) evaluated and approved all experimental procedures.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the datasets used and/or analyzed throughout the present study are available from the corresponding author on reasonable request.
Acknowledgments
We are grateful to Dr. Giacomo Vidotto, Department of Medical Veterinary Sciences, Alma Mater Studiorum–University of Bologna, Ozzano dell’Emilia, Italy, for his valuable technical assistance with immunohistochemistry and cell counting.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amarowicz, R.; Shahidi, F. Antioxidant activity of peptide fractions of capelin protein hydrolysates. Food Chem. 1997, 58, 355–359. [Google Scholar] [CrossRef]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar]
- Kim, S.Y; Je, J.Y; Kim, S.K. Purification and characterization of antioxidant peptide from Hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Byun, H.-G. ; Kim. S.-K. Investigation of Jumbo Squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef]
- Je, J.Y.; Lee, K.H.; Lee, M.H.; Ahn, C.B. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food Res. Int. 2009, 42, 1266–1272. [Google Scholar]
- Chalamaiah, M.; Dinesh Kumar, B.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: proximate composition, amino acid composition, antioxidant activities and applications: a review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-originating ACE inhibitors, including antihypertensive peptides, as preventive food components in blood pressure reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Aksnes, A.; Hope, B.; Høstmark, Ø.; Albrektsen, S. Inclusion of size fractionated fish hydrolysate in high plant protein diets for Atlantic cod, Gadus morhua. Aquaculture 261, 1102–1110. [CrossRef]
- Aksnes, A.; Hope, B.; Jönsson, E.; Björnsson, B.T.; Albrektsen, S. Size-fractionated fish hydrolysate as feed ingredient for rainbow trout (Oncorhynchus mykiss) fed high plant protein diets. I: Growth, growth regulation and feed utilization. Aquaculture, 2006; 261, 305–317. [Google Scholar]
- Cahu, C.; Infante, J.Z. Substitution of live food by formulated diets in marine fish larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Kolkovski, S.; Czesny, S.; Dabrowski, K. Use of krill hydrolysate as feed attractant for fish larvae and juveniles. J. World Aquacult. Soc. 2000, 31, 81–88. [Google Scholar] [CrossRef]
- Refstie, S.; Olli, J.J.; Standal, H. Feed intake, growth, and protein utilisation by post-smolt Atlantic salmon (Salmo salar) in response to graded levels of fish protein hydrolysate in the diet. Aquaculture 2004, 239, 331–349. [Google Scholar] [CrossRef]
- Panjaitan, F.C.A.; Gomez, H.L.R.; Chang, Y.W. In silico analysis of bioactive peptides released from giant grouper (Epinephelus lanceolatus) roe proteins identified by proteomics approach. Molecules 2018, 23, 2910. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Romero-Luna, H.E. Conventional and in silico approaches to select promising food-derived bioactive peptides: A review. Food Chem. 2022; X 13, 100183. [Google Scholar]
- Girgih, A.T.; Udenigwe, C.C.; Hasan, F.M.; Gill, T.A.; Aluko, R.E. Antioxidant properties of Salmon (Salmo salar) protein hydrolysate and peptide fractions isolated by reverse-phase HPLC. Food Res. Int. 2013, 52, 315–322. [Google Scholar]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [CrossRef]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Inhibition properties of dipeptides from salmon muscle hydrolysate on angiotensin I-converting enzyme. Int. J. Food Sci. Technol. 2006, 41, 383–386. [Google Scholar] [CrossRef]
- Gu, R.Z.; Li, C.Y.; Liu, W.Y.; Yi, W.X.; Cai, M.Y. Angiotensin I-converting enzyme inhibitory activity of low-molecular-weight peptides from Atlantic salmon (Salmo salar L.) skin. Food Res. Int. 2011, 44, 1536–1540. [Google Scholar]
- 21 Parma, L.; Busti, S.; Ciulli, S.; Volpe, E.; Errani, F.; Oterhals, Å.; Romarheim, O.H. , Aspevik, T. ; Dondi, F.; Gatta, P.P.; Bonaldo, A. Growth, plasma biochemistry and immune-related gene expression of European sea bass (Dicentrarchus labrax) fed bioactive peptides from farmed salmon by-products. Aquaculture 2023, 573, 738982. [Google Scholar]
- Cummings, D.E.; Overduin, J. Gastrointestinal regulation of food intake. J. Clin. Investig. 2007, 117, 13–23. [Google Scholar] [CrossRef]
- Sternini, C.; Anselmi, L.; Rozengurt, E. Enteroendocrine cells: a site of ‘taste’ in gastrointestinal chemosensing. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Moran, T.H.; Dailey, M.J. Gut peptides: targets for antiobesity drug development? Endocrinology 2009, 150, 2526–2530. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Lemmens, S.G.; Westerterp, K.R. Dietary protein–its role in satiety, energetics, weight loss and health. Br. J. Nutr. 2012, 108, S105–S112. [Google Scholar] [CrossRef] [PubMed]
- Bensaid, A.; Tomè, D.; Gietzen, D.; Even, P.; Morens, C.; Gaussercs, N.; Fromentin, G. Protein is more potent than carbohydrate for reducing appetite in rats. Physiol. Behav. 2002, 75, 577–582. [Google Scholar] [CrossRef]
- Caron, J.; Domenger, D.; Dhulster, P.; Ravallec, R.; Cudennec, B. Protein digestion-derived peptides and the peripheral regulation of food intake. Front. Endocrinol. 2017, 8, 85. [Google Scholar] [CrossRef] [PubMed]
- Chaudhri, O.; Small, C.; Bloom, S. Gastrointestinal hormones regulating appetite. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1187–1209. [Google Scholar] [CrossRef]
- Volkoff, H.; Xu, M.; MacDonald, E.; Hoskins, L. Aspects of the hormonal regulation of appetite in fish with emphasis on goldfish, Atlantic cod and winter flounder: notes on actions and responses to nutritional, environmental and reproductive changes. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2009, 153, 8–12. [Google Scholar] [CrossRef]
- Jönsson, E. The role of ghrelin in energy balance regulation in fish. Gen. Comp. Endocrinol. 2013, 187, 79–85. [Google Scholar] [CrossRef]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E. , Weigle D.S. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef]
- Cummings, D.E. Ghrelin and the short- and long-term regulation of appetite and body weight. Physiol. Behav. 2006, 89, 71–84. [Google Scholar] [CrossRef]
- MacDonald, E.; Volkoff, H. Cloning, distribution and effects of season and nutritional status on the expression of neuropeptide Y (NPY), cocaine and amphetamine regulated transcript (CART) and cholecystokinin (CCK) in winter flounder (Pseudopleuronectes americanus). Horm. Behav. 2009, 56, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. The role of neuropeptide Y, orexins, cocaine and amphetamine related transcript, cholecystokinin, amylin and leptin in the regulation of feeding in fish. Comp. Biochem. Physiol. 2006, 144, 325–331. [Google Scholar] [CrossRef]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerda-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, X.F.; Yuan, X.C.; Li, J.; He, Y.; Fang, L.; Guo, X.; Liu, L.; Li, B.; Shen, D. Neuropeptide Y stimulates food intake and regulates metabolism in grass carp, Ctenopharyngodon idellus. Aquaculture, 2013; 380–383, 52–61. [Google Scholar]
- Sheridan, M.A.; Kittilson, J.D. The role of somatostatins in the regulation of metabolism in fish. Comp. Biochem. Physio. l B Biochem. Mol. Biol. 2004, 138, 323–330. [Google Scholar] [CrossRef]
- Martínez-Alvarez, O. Hormone-like peptides obtained by marine-protein hydrolysis and their bioactivities. In: Marine proteins and peptides: biological activities and applications. Se-Kwon Kim, Eds.; John Wiley & Sons, Ltd., the Atrium, Southern Gate, Chichester, West Sussex, PO19 8SQ, UK, 2013; pp. 351–367. [CrossRef]
- 40 Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- Kitts, D.D.; Weiler, K. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Curr. Pharm. Des. 2003, 9, 1309–1323. [Google Scholar] [CrossRef] [PubMed]
- Teschemacher, H. Opioid receptor ligands derived from food proteins. Curr. Pharm. Des. 2003, 9, 1331–1344. [Google Scholar] [CrossRef]
- Fouchereau-Peron, M.; Duvail, L.; Michel, C.; Gildberg, A.; Batista, I.; Le Gal, Y. Isolation of an acid fraction from a fish protein hydrolysate with a calcitonin-gene-related-peptide-like biological activity. Biotechnol. Appl. Biochem. 1999, 29, 87–92. [Google Scholar]
- Martínez-Alvarez, O.; Guimas, L.; Delannoy, C.; Fouchereau-Peron, M. Occurrence of a CGRP like molecule in Siki (Centroscymnus coelolepsis) hydrolysate of industrial origin. J. Agric. Food Chem. 2007, 55, 5469–5475. [Google Scholar] [CrossRef]
- Šližytė, R.; Mozuraitytė, R.; Martínez-Alvarez, O.; Falch, E.; Fouchereau-Peron, M.; Rustad, T. Functional, bioactive and antioxidative properties of hydrolysates obtained from cod (Gadus morhua) backbones. Process Biochem. 2009, 44, 668–677. [Google Scholar] [CrossRef]
- 46: Parma, L.; Yúfera, M.; Navarro-Guillén, C.; Moyano, F.J.; Soverini, M.; D’Amico, F.; Candela, M.; Fontanillas, R.; Gatta, P.P.; Bonaldo, A. Effects of calcium carbonate inclusion in low fishmeal diets on growth, gastrointestinal pH, digestive enzyme activity and gut bacterial community of European sea bass (Dicentrarchus labrax L.) juveniles. Aquaculture 2019, 510, 283–292. [Google Scholar] [CrossRef]
- Parma, L.; Pelusio, N.F.; Gisbert, E.; Esteban, M.A.; D’Amico, F.; Soverini, M.; Candela, M.; Dondi, F.; Gatta, P.P.; Bonaldo, A. Effects of rearing density on growth, digestive conditions, welfare indicators and gut bacterial community of gilthead sea bream (Sparus aurata, L. 1758) fed different fishmeal and fish oil dietary levels. Aquaculture 2020, 518, 734854. [Google Scholar] [CrossRef]
- Oterhals, Å.; Samuelsen, T.A. Plasticization effect of solubles in fishmeal. Food Res. Int. 2015, 69, 313–321. [Google Scholar]
- Busti, S.; Bonaldo, A.; Dondi, F.; Cavallini, D.; Yúfera, M.; Gilannejad, N.; Moyano, F.J.; Gatta, P.P.; Parma, L. Effects of different feeding frequencies on growth, feed utilization, digestive enzyme activities and plasma biochemistry of gilthead sea bream (Sparus aurata) fed with different fishmeal and fish oil dietary levels. Aquaculture 2020, 529, 735616. [Google Scholar] [CrossRef]
- Mazzoni, M.; Lattanzio, G.; Bonaldo, A.; Tagliavia, C.; Parma, L.; Busti, S.; Gatta, P.P.; Bernardi, N.; Clavenzani, P. Effect of essential oils on the oxyntopeptic cells and somatostatin and ghrelin immunoreactive cells in the European Sea Bass (Dicentrarchus labrax) gastric mucosa. Animals 2021, 11, 3401. [Google Scholar] [CrossRef] [PubMed]
- Cahu, C.L.; Infante, J.Z.; Quazuguel, P.; Le Gall, M.M. Protein hydrolysate vs. fish meal in compound diets for 10-day old sea bass Dicentrarchus labrax larvae. Aquaculture 1999, 171, 109–119. [Google Scholar] [CrossRef]
- Hardy, R.W. Fish hydrolysates: production and use in aquaculture feeds. In: Proceeding of the Aquaculture Feed Processing and Nutrition Workshop; Akiyama, D.M., Tan, R.K.H., Eds.; American Soybean Association, Singapore, 1991; pp. 109–115.
- Bøgwald, J.; Dalmo, R.A.; Leifson, R.M.; Stenbern, E.; Gildberg, A. The stimulatory effect of a muscle protein hydrolysate from Atlantic cod, Gadus morhua L. on Atlantic salmon, Salmo salar L., head kidney leucocytes. Fish Shellfish Immunol. 1996, 6, 3–16. [Google Scholar] [CrossRef]
- Gildberg, A.; Bøgwald, J.; Johansen, A.; Stenverg, E. Isolation of acid peptide fractions from a fish protein hydrolysate with strong stimulatory effect on Atlantic salmon (Salmo salar) head kidney leucocytes. Comp. Biochem. Physiol. 1996; 114B, 97–101. [Google Scholar]
- Liang, M.; Wang, J.; Chang, Q.; Mai, K. Effects of different levels of fish protein hydrolysate in the diet on the nonspecific immunity of Japanese sea bass, Lateolabrax japonicus (Cuvier et Valenciennes, 1828). Aquac. Res. 2006, 37, 102–106. [Google Scholar] [CrossRef]
- Bui, H.T.D.; Khosravi, S.; Fournier, V.; Herault, M.; Lee, K.J. Growth performance, feed utilization, innate immunity, digestibility and disease resistance of juvenile red sea bream (Pagrus major) fed diets supplemented with protein hydrolysates. Aquaculture, 2014; 418–419, 11–16. [Google Scholar]
- Santos-Hernandez, M.S.; Miralles, B.; Amigo, L.; Recio, I. Intestinal signals of proteins and digestion-derived products relevant to satiety. J. Agric. Food Chem. 2018, 66, 10123–10131. [Google Scholar] [CrossRef]
- Wauson, E.M.; Lorente-Rodriguez, A.; Cobb, M.H. Minireview: nutrient sensing by G protein-coupled receptors. Mol. Endocrinol. 2013, 27, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Tulipano, G. Whey proteins and enteric hormones interplay between food components and the enteroendocrine system. Agro. Food Ind. Hi Tech. 2015, 26, 15–19. [Google Scholar]
- Liou, A.P.; Chavez, D.L.; Espero, E.; Hao, S.; Wank, S.A.; Raybould, H.E. Protein hydrolysate-induced cholecystokinin secretion from enteroendocrine cells is indirectly mediated by the intestinal oligopeptide transporter PepT1. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G895–G902. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Al-Rammahi, M.; Moran, A.; Marcello, M.; Ninomiya, Y.; Shirazi-Beechey, S.P. Sensing of amino acids by the gut expressed taste receptor T1R1-T1R3 stimulates CCK secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G271–G282. [Google Scholar] [CrossRef] [PubMed]
- Geraedts, M.C.P.; Troost, F.J.; Fisher, M.A.J.G.; Edens, L.; Saris, W.H.M. Direct induction of CCK and GLP-1 release from murine endocrine cells by intact dietary proteins. Mol. Nutr. Food Res. 2011, 55, 476–484. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Cacciamali, A.; Caroli, A.M. Food protein-derived peptide sensing by enteroendocrine cells compared to osteoblast-like cells: role of peptide length and peptide composition, focusing on products of beta-lactoglobulin hydrolysis. Int. Dairy J. 2017, 72, 55–62. [Google Scholar] [CrossRef]
- Tulipano, G. Role of bioactive peptide sequences in the potential impact of dairy protein intake on metabolic health. Int. J. Mol. Sci. 2020, 21, 8881. [Google Scholar] [CrossRef]
- Choi, S.; Lee, M.; Shiu, A.L.; Yo, S.J.; Hallden, G.; Aponte, G.W. GPR93 activation by protein hydrolysate induces CCK transcription and secretion in STC-1 cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1366–G1375. [Google Scholar] [CrossRef]
- Cudennec, B.; Ravallec-Plé, R.; Courois, E.; Fouchereau-Peron, M. Peptides from fish and crustacean by-products hydrolysates stimulate cholecystokinin release in STC-1 cells. Food Chem. 2008, 111, 970–975. [Google Scholar] [CrossRef]
- Moughan, P.J.; Cranwell, P.D.; Smith, W.C. An evaluation with piglets of bovine milk, hydrolysed bovine milk, and isolated soybean proteins included in infant milk formulas. II. Stomach emptying rate and the postprandial change in gastric pH and milk-clotting enzyme activity. J. Pediatr. Gastroenterol. Nutr. 1991, 12, 253–259. [Google Scholar] [CrossRef]
- Tatemoto, K.; Carlquist, M.; Mutt, V. Neuropeptide Y—A novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 1982, 296, 659–660. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, A.G.; Söderberg, C.; Lundell, I.; Milner, R.J.; Larhammar, D. Strong evolutionary conservation of neuropeptide Y: Sequences of chicken, goldfish, and Torpedo marmorata DNA clones. Proc. Natl. Acad. Sci. USA 1992, 89, 2350–2354. [Google Scholar] [CrossRef]
- Kurokawa, T.; Suzuki, T. Development of Neuropeptide Y-related peptides in the digestive organs during the larval stage of Japanese flounder, Paralichthys olivaceus. Gen. Comp. Endocrinol. 2002, 126, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Sundström, G.; Larsson, T.; Xu, B.; Heldin, J.; Larhammar, D. Interactions of zebrafish peptide YYb with the neuropeptide Y-family receptors Y4, Y7, Y8a, and Y8b. Front. Neurosci. 2013, 7, 29. [Google Scholar]
- Matsuda, K.; Azuma, M.; Kang, K.S. Orexin system in teleost fish. In: Vitamins and Hormones, 1st ed.; Harris, R.S., Lorraine, J.A., Munson, P.L., Glover, J., Aurbach, G.D., Thiman, T.V., Wool, I.G., Diczfalusy, E., Olsen, R., McCormick, D.B., Eds.; Elsevier, 32 Jamestown Road, London, UK, 2012; Volume 89, pp. 341–361. [CrossRef]
- Cerdá-Reverter, J.; Larhammar, D. Neuropeptide Y family of peptides: Structure, anatomical expression, function, and molecular evolution. Biochem. Cell Biol. 2000, 78, 371–392. [Google Scholar] [CrossRef]
- López-Patiño, M.A.; Guijarro, A.I.; Isorna, E.; Delgado, M.J.; Alonso-Bedate, M.; de Pedro, N. Neuropeptide Y has a stimulatory action on feeding behavior in Goldfish (Carassius Auratus). Eur. J. Pharmacol. 1999, 377, 147–153. [Google Scholar] [CrossRef]
- Tang, Z.; Sun, C.; Yan, A.; Wu, S.; Qin, C.; Zhang, Y.; Li, W. Genes involved in fatty acid metabolism: Molecular characterization and hypothalamic mRNA response to energy status and neuropeptide Y treatment in the orange-spotted grouper Epinephelus coioides. Mol. Cell. Endocrinol. 2013, 376, 114–124. [Google Scholar] [CrossRef]
- Murashita, K.; Kurokawa, T.; Nilsen, T.O.; Rønnestad, I. Ghrelin, cholecystokinin, and peptide YY in Atlantic salmon (Salmo salar): Molecular cloning and tissue expression. Gen. Comp. Endocrinol. 2009, 160, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Narnaware, Y.K.; Peyon, P.P.; Lin, X.; Peter, R.E. Regulation of food intake by neuropeptide Y in goldfish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, 1025–1034. [Google Scholar] [CrossRef]
- Liang, X.-F.; Li, G.-Z.; Yao, W.; Cheong, L.-W.; Liao, W.-Q. Molecular characterization of neuropeptide Y gene in Chinese perch, an acanthomorph fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 55–64. [Google Scholar] [CrossRef]
- Narnaware, Y.K.; Peter, R.E. Influence of diet composition on food intake and neuropeptide Y (NPY) gene expression in goldfish brain. Regul. Pept. 2022, 103, 75–83. [Google Scholar] [CrossRef]
- Fredriksson, R.; Larson, E.T.; Yan, Y.-L.; Postlethwait, J.H.; Larhammar, D. Novel Neuropeptide Y Y2-Like receptor subtype in zebrafish and frogs supports early vertebrate chromosome duplications. J. Mol. Evol. 2004, 58, 106–114. [Google Scholar] [CrossRef]
- Fredriksson, R.; Sjödin, P.; Larson, E.T.; Conlon, J.M.; Larhammar, D. Cloning and characterization of a zebrafish Y2 receptor. Regul. Pept. 2006, 133, 32–40. [Google Scholar] [CrossRef]
- Lundell, I.; Berglund, M.M.; Starback, P.; Salaneck, E.; Gehlert, D.R.; Larhammar, D. Cloning and characterization of a novel neuropeptide Y receptor subtype in the zebrafish. DNA Cell Biol. 1997, 16, 1357. [Google Scholar] [CrossRef] [PubMed]
- Assan, D.; Mustapha, U.F.; Chen, H.; Li, Z.; Peng, Y.; Li, G. The roles of neuropeptide Y (Npy) and peptide YY (Pyy) in Teleost food intake: A mini review. Life (Basel). 2021, 11, 547. [Google Scholar] [CrossRef]
- Bar, F.; Foh, B.; Pagel, R.; Schroder, T.; Schlichting, H.; Hirose, M.; Sina, C. Carboxypeptidase E modulates intestinal immune homeostasis and protects against experimental colitis in mice. PLoS One, 2014, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.A.; Englander, E.W.; Greeley, G.H. Postpyloric gastrointestinal peptides. In: Physiology of the Gastrointestinal Tract, Johnson L.R., Eds.; Elsevier Inc., New York, USA, 2012; Volume 1, pp. 155–198.
- Vigliano, F.A.; Munoz, L.; Hernandez, D.; Cerutti, P.; Bermudez, R.; Quiroga, M.I. An immunohistochemical study of the gut neuroendocrine system in juvenile pejerrey Odontesthes bonariensis (Valenciennes). J. Fish Biol. 2011, 78, 901–911. [Google Scholar] [CrossRef]
- Hernández, D.R.; Vigliano, F.A.; Sanchez, S.; Bermudez, R.; Domitrovic, H.A.; Quiroga, M.I. Neuroendocrine system of the digestive tract in Rhamdia quelen juvenile: An immunohistochemical study. Tissue Cell 2012, 44, 220–226. [Google Scholar] [CrossRef]
- Pereira, R.T.; Costa, L.S.; Oliveira, I.R.; Araujo, J.C.; Aerts, M.; Vigliano, F.A.; Rosa, P.V. Relative distribution of gastrin-, CCK-8-, NPY- and CGRP-immunoreactive cells in the digestive tract of dorado (Salminus brasiliensis). Tissue Cell 2015, 47, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, P.; Ou, Y.; Li, J.; Wen, J. An immunohistochemical study on endocrine cells in the neuroendocrine system of the digestive tract of milkfish Chanos chanos (Forsskal, 1775). Aquac. Res. 2017, 48, 1439–1449. [Google Scholar] [CrossRef]
- Pereira, R.T.; Rodriguez de Freitas, T.; Cardozo de Oliveira, I.R.; Costa, L.S.; Vigliano, F.A.; Rosa, P.V. Endocrine cells producing peptide hormones in the intestine of Nile tilapia: Distribution and effects of feeding and fasting on the cell density. Fish Physiol. Biochem. 2017, 43, 1399–1412. [Google Scholar] [CrossRef]
- Ji, W.; Ping, H.C.; Wei, K.J.; Zhang, G.R.; Shi, Z.C.; Yang, R.B.; Wang, W.M. Ghrelin, neuropeptide Y (NPY) and cholecystokinin (CCK) in blunt snout bream (Megalobrama amblycephala): cDNA cloning, tissue distribution and mRNA expression changes responding to fasting and refeeding. Gen. Comp. Endocrinol. 2015, 223, 108–119. [Google Scholar] [CrossRef]
- Hernández, D.R.; Barrios, C.E.; Santinón, J.J.; Sánchez, S.; Baldisserotto, B. Effect of fasting and feeding on growth, intestinal morphology and enteroendocrine cell density in Rhamdia quelen juveniles. Aquac. Res. 2018, 49, 1512–1520. [Google Scholar] [CrossRef]
- Gorissen, M.H.A.G.; Flik, G.; Huising, M. , Peptides and proteins regulating food intake: A comparative view. Anim. Biol. 2006, 56, 447–473. [Google Scholar] [CrossRef]
- Volkoff, H. The neuroendocrine regulation of food intake in fish: A review of current knowledge. Front. Neurosci. 2016, 10, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Dale, H.F.; Hausken, T.; Hatlebakk, J.G.; Brønstad, I.; Lied, G.A.; Hoff, D.A.L. The effect of supplementation with low doses of a cod protein hydrolysate on satiety hormones and inflammatory biomarkers in adults with metabolic syndrome: A randomized, double-blind study. Nutrients 2020, 12, 3421. [Google Scholar] [CrossRef]
- Günther, T.; Tulipano, G.; Dournaud, P.; Bousquet, C.; Csaba, Z.; Kreienkamp, H.J.; Lupp, A. , Korbonits, M.; Castaño, J.P.; Wester, H.J.; Culler, M.; Melmed, S.; Schulz S. International union of basic and clinical pharmacology. CV. Somatostatin receptors: structure, function, ligands, and new nomenclature. Pharmacol. Rev. 2018, 70, 763–835. [Google Scholar] [CrossRef]
- Holstein, B.; Cederberg, C. Effect of somatostatin on basal and stimulated gastric secretion in the cod, Gadus morhua. Am. J. Physiol. 1988, 254, G183–G188. [Google Scholar] [CrossRef]
- Holstein, B.; Humphrey, C.S. Stimulation of gastric acid secretion and suppression of VIP-like immunoreactivity by bombesin in the Atlantic codfish, Gadus morhua. Acta Physiol. Scand. 1980, 109, 217–223. [Google Scholar] [CrossRef]
- Uneyama, H.; Niijima; A. , San Gabriel, A.; Torii, K. Luminal amino acid sensing in the rat gastric mucosa. Am. J. Physiol. 2006, 291, G1163–G1170. [Google Scholar] [CrossRef]
- Niijima, A. Effects of oral and intestinal stimulation with umami substance on gastric vagus activity. Physiol. Behav. 1991, 49, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Niijima, A.; Torii, K.; Uneyama, H. Role played by vagal chemical sensors in the hepato-portal region and duodeno-intestinal canal: an electrophysiological study. Chem. Senses. 2005, 30, i178–i179. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, E.; Hasumura, M.; Uneyama, H.; Torii, K. Luminal amino acid-sensing cells in gastric mucosa. Digestion 2011, 83, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L. Gastric secretion. Curr. Opin. Gastroenterol. 2003, 19, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L. Gastric secretion. Curr. Opin. Gastroenterol. 2005, 21, 636–643. [Google Scholar] [CrossRef]
- Shamburek, R.D.; Schubert, M.L. Control of gastric acid secretion. Histamine H2-receptor antagonists and H+, K+-ATPase inhibitors. Gastroenterol. Clin. North. Am. 1992, 21, 527–550. [Google Scholar] [CrossRef] [PubMed]
- Van Op den Bosch, J.; Adriaensen, D.; Van Nassauw, L.; Timmermans, J.P. The role(s) of somatostatin, structurally related peptides and somatostatin receptors in the gastrointestinal tract: a review. Regul. Pept. 2009, 156, 1–8. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Seabream gastric mucosa. Image A shows the localization of immunoreactive (-IR) enteroendocrine cells (EECs) for somatostatin (SOM, arrows), while in B oxyntopeptic cells (asterisks) IR stained with Na+K+-ATPase antiserum. Occasionally, SOM-IR cells show a typical morphological appearance of ‘open type’ EECs (A and C, arrows), while other SOM-IR cells show a ‘closed type’ morphological appearance (A and C, arrowheads). Merge C shows the arrangement of SOM-IR cells in the gastric mucosa: most of the IR cells were positioned at the endoluminal side, while some SOM-IR cells are scattered along the adenomer of the gastric glands.
Figure 1.
Seabream gastric mucosa. Image A shows the localization of immunoreactive (-IR) enteroendocrine cells (EECs) for somatostatin (SOM, arrows), while in B oxyntopeptic cells (asterisks) IR stained with Na+K+-ATPase antiserum. Occasionally, SOM-IR cells show a typical morphological appearance of ‘open type’ EECs (A and C, arrows), while other SOM-IR cells show a ‘closed type’ morphological appearance (A and C, arrowheads). Merge C shows the arrangement of SOM-IR cells in the gastric mucosa: most of the IR cells were positioned at the endoluminal side, while some SOM-IR cells are scattered along the adenomer of the gastric glands.
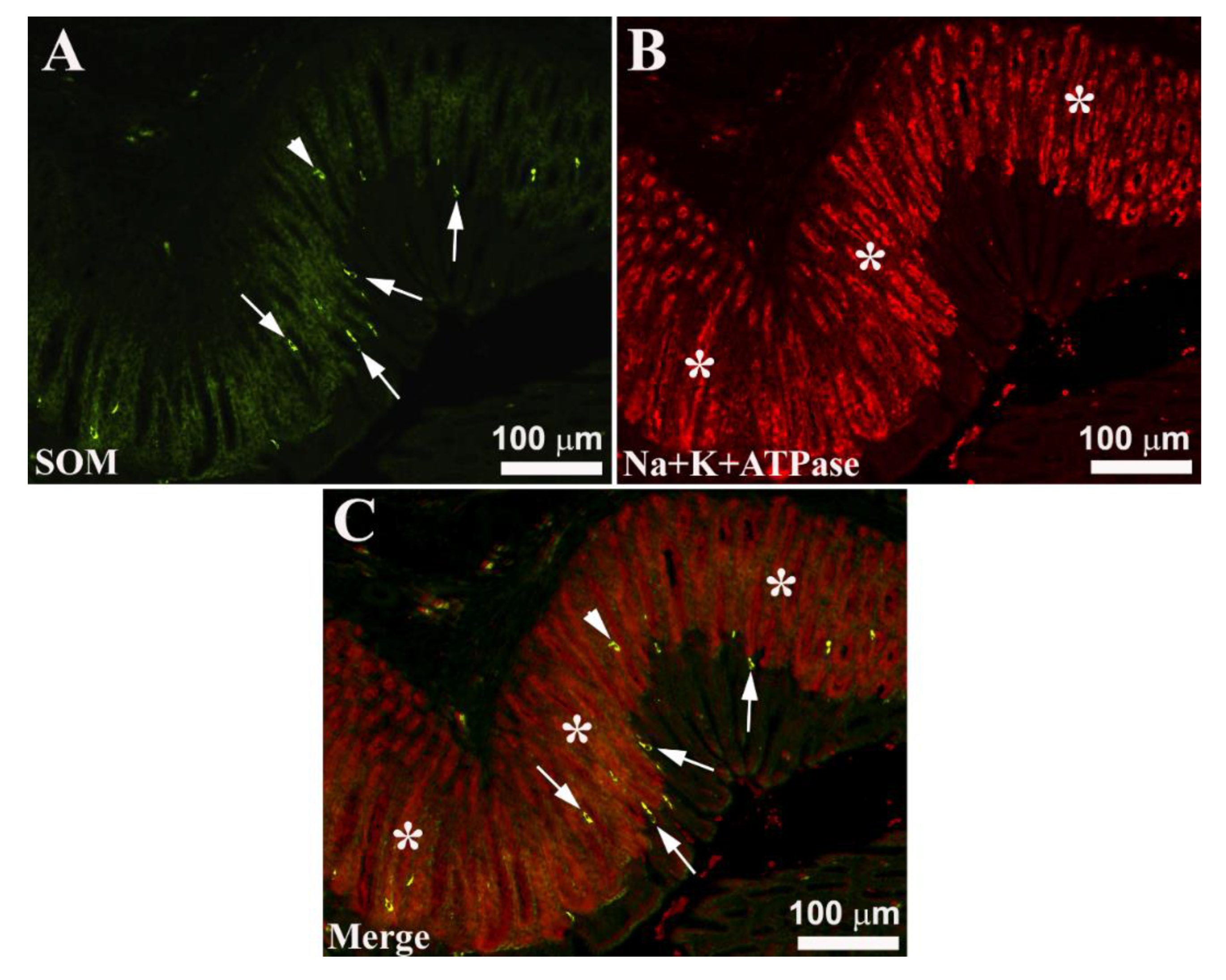
Figure 2.
Seabass gastric mucosa. In these images, obtained at higher magnification, both the different morphological aspect and the different position of the two subpopulations of immunoreactive (-IR) endocrine cells are represented. In the mucosa, neuropeptide Y (NPY, A) and ghrelin (GHR, B) -IR cells alternate which exhibit the two morphological types of “open type” (A and C, arrows) and “closed type” enteroendocrine cells (B and C, arrowheads). Merge C shows that not all enteroendocrine cells co-express NPY / GHR.
Figure 2.
Seabass gastric mucosa. In these images, obtained at higher magnification, both the different morphological aspect and the different position of the two subpopulations of immunoreactive (-IR) endocrine cells are represented. In the mucosa, neuropeptide Y (NPY, A) and ghrelin (GHR, B) -IR cells alternate which exhibit the two morphological types of “open type” (A and C, arrows) and “closed type” enteroendocrine cells (B and C, arrowheads). Merge C shows that not all enteroendocrine cells co-express NPY / GHR.
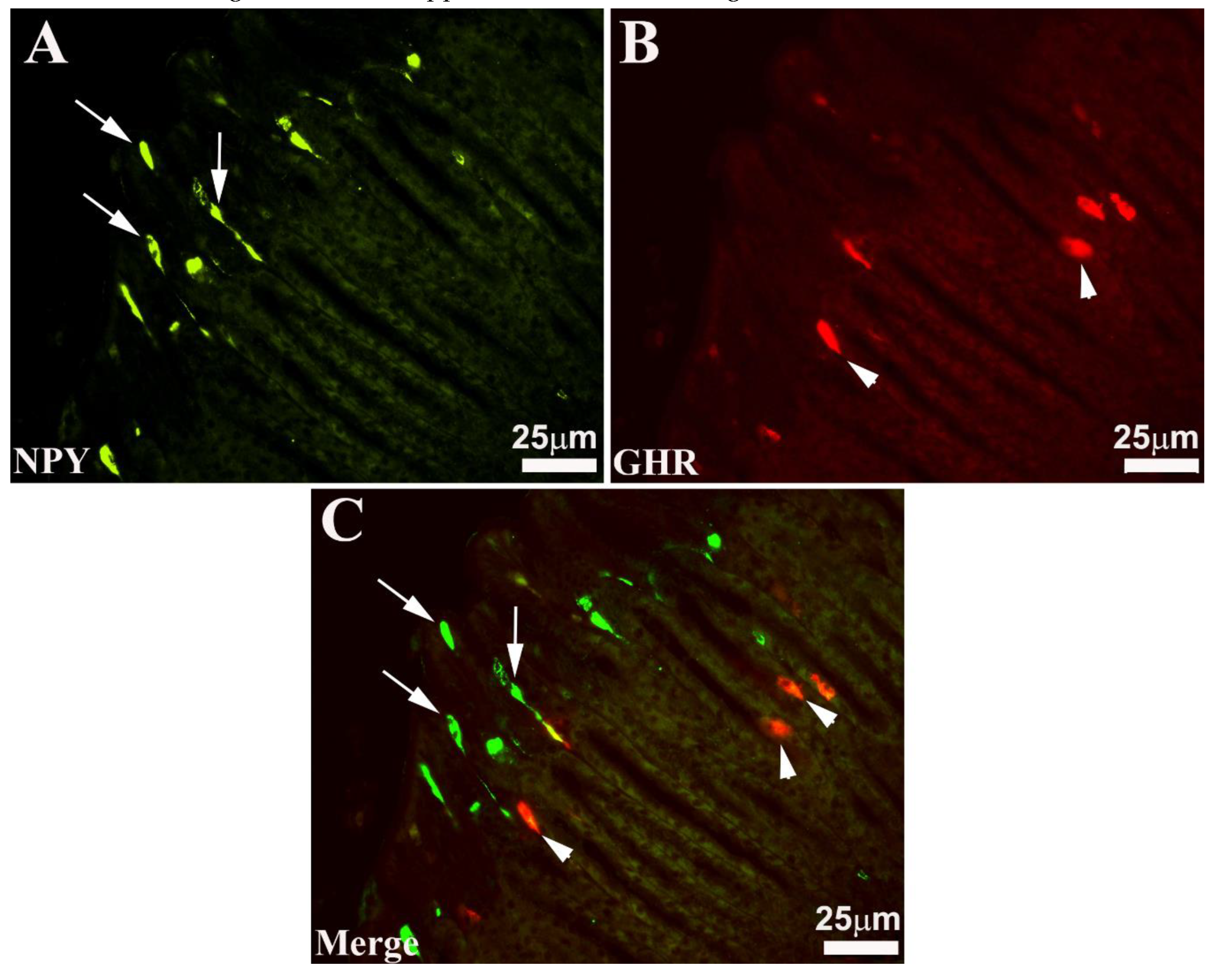
Figure 3.
Seabass gastric mucosa. Images A, B and C show the total overlapping exhibited by the enteroendocrine neuropeptide Y, (NPY) and ghrelin (GHR) immunoreactive (-IR) cells intermingled with oxyntopeptic cells. Note that some -IR enteroendocrine cells show morphological appearance of “open type” (A, B and C, arrows), while others of “closed type” (A, B and C, arrowheads). C merge.
Figure 3.
Seabass gastric mucosa. Images A, B and C show the total overlapping exhibited by the enteroendocrine neuropeptide Y, (NPY) and ghrelin (GHR) immunoreactive (-IR) cells intermingled with oxyntopeptic cells. Note that some -IR enteroendocrine cells show morphological appearance of “open type” (A, B and C, arrows), while others of “closed type” (A, B and C, arrowheads). C merge.
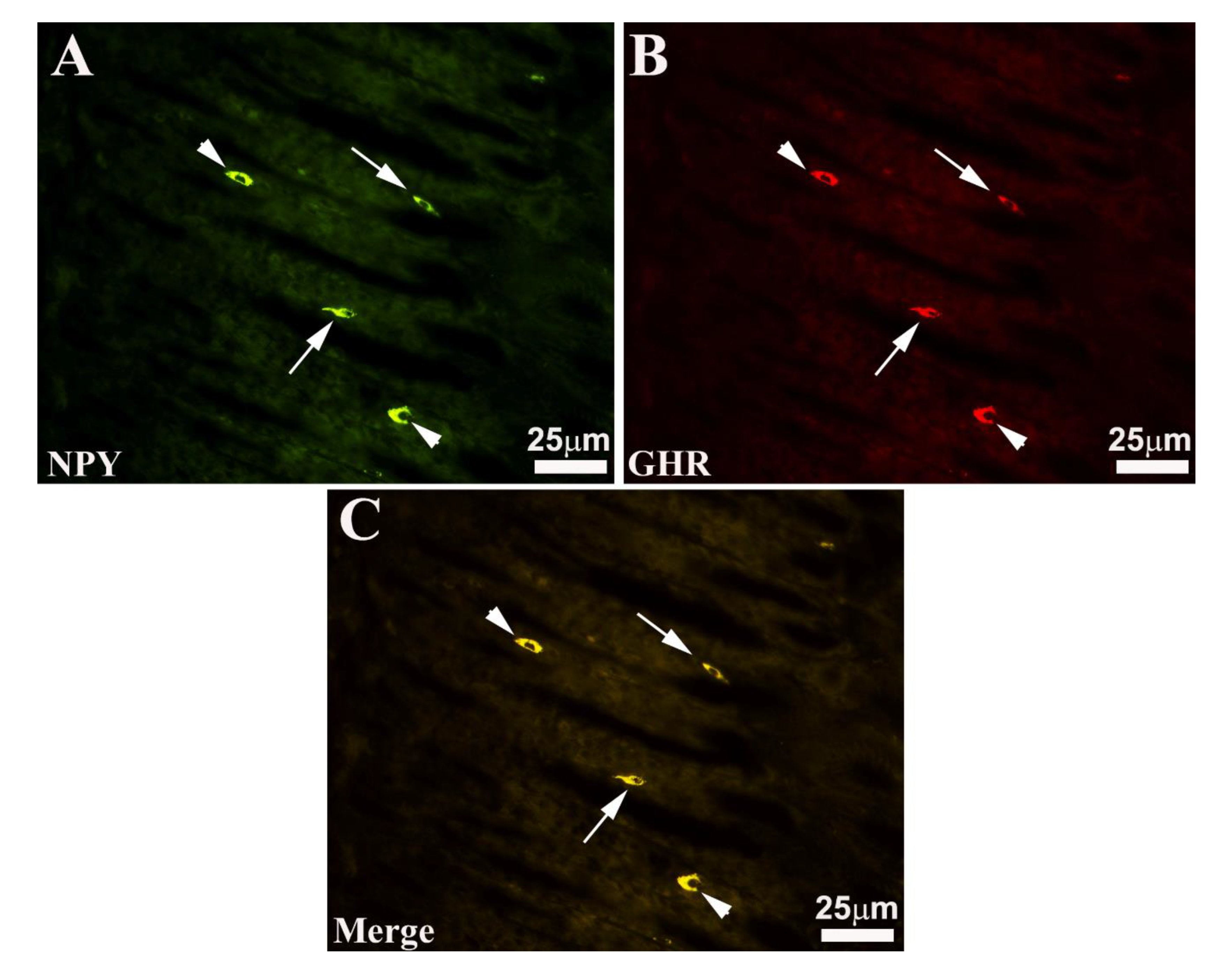
Figure 4.
The histograms show the area occupied by oxyntopeptic (OP) immunoreactive (-IR) cells evaluated in the gastric mucosa of seabream (A) and seabass (B). In the (C) (seabream) and (D) (seabass) were represented the mean number of neuropeptide Y (NPY) and ghrelin (GHR) -IR enteroendocrine cells (EECs), while in E and F the mean number of somatostatin-IR cells in the seabream (E) and seabass (F) gastric mucosa. Experimental groups: CTR (control group), BP 5% and BP 10%. * indicates P ≤ 0.05; ** indicates P ≤ 0.01. Values expressed as mean + SD.
Figure 4.
The histograms show the area occupied by oxyntopeptic (OP) immunoreactive (-IR) cells evaluated in the gastric mucosa of seabream (A) and seabass (B). In the (C) (seabream) and (D) (seabass) were represented the mean number of neuropeptide Y (NPY) and ghrelin (GHR) -IR enteroendocrine cells (EECs), while in E and F the mean number of somatostatin-IR cells in the seabream (E) and seabass (F) gastric mucosa. Experimental groups: CTR (control group), BP 5% and BP 10%. * indicates P ≤ 0.05; ** indicates P ≤ 0.01. Values expressed as mean + SD.
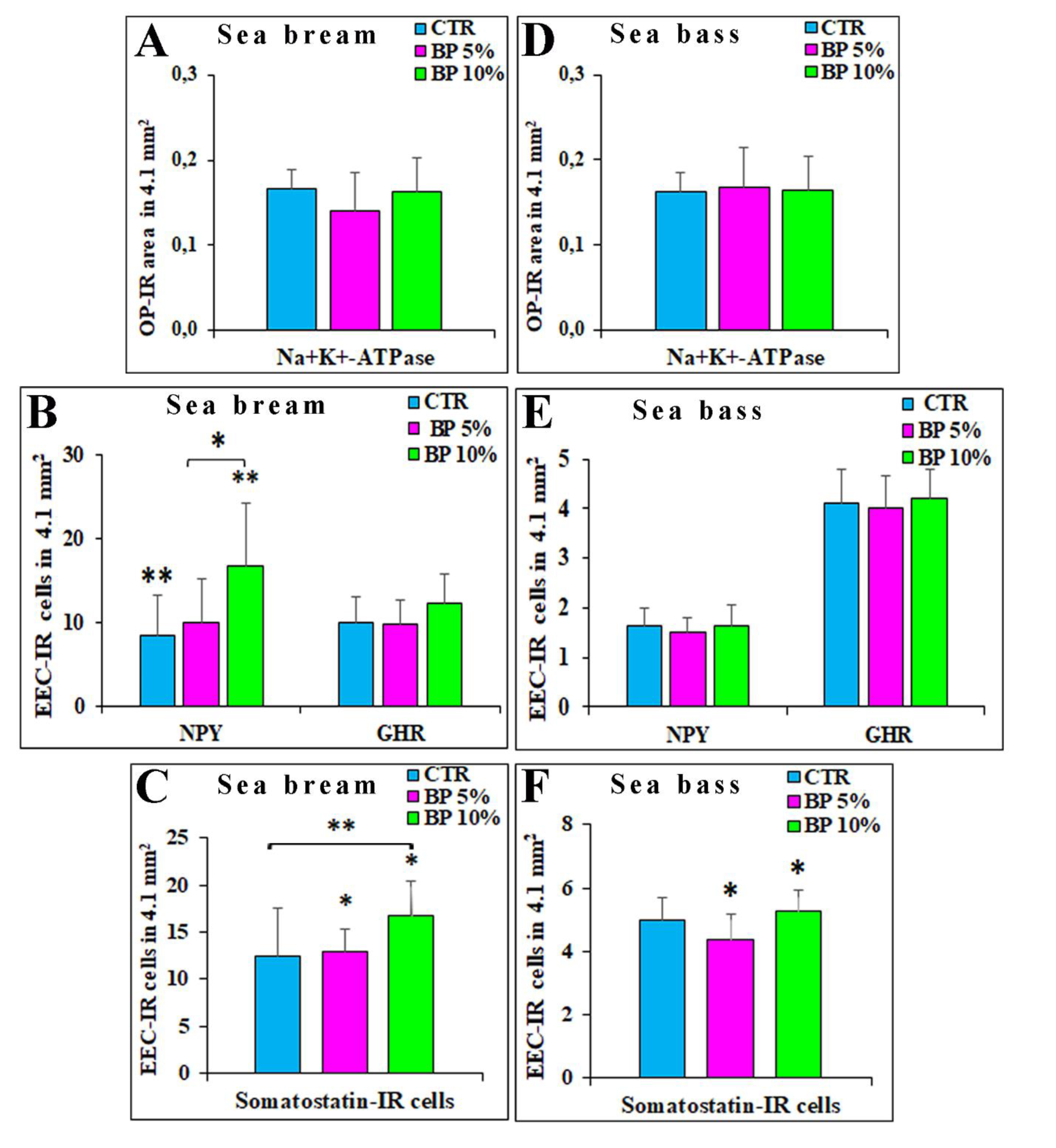

Table 1.
Components of the three experimental diets and their proximate composition.
| Experimental diets | |||
|---|---|---|---|
| CTR | BP5 | BP10 | |
| Ingredients, % | |||
| Salmon hydrolysate | - | 5.00 | 10.00 |
| Fish meal | 15.00 | 10.00 | 5.00 |
| Soybean meal | 15.00 | 15.00 | 15.00 |
| Wheat | 15.28 | 16.02 | 16.87 |
| Wheat gluten | 12.20 | 11.30 | 10.30 |
| Corn gluten | 8.00 | 8.00 | 8.00 |
| Soy protein concentrate | 5.40 | 5.40 | 5.40 |
| Fish oil | 10.00 | 9.90 | 9.80 |
| Rapeseed oil | 5.00 | 5.00 | 5.00 |
| Horse beans | 10.00 | 10.00 | 10.00 |
| Lecithin from rapeseed | 1.00 | 1.00 | 1.00 |
| *Vitamin premix | 0.50 | 0.50 | 0.50 |
| *Mineral premics | 0.50 | 0.50 | 0.50 |
| Monosodiumphosphate | 3.00 | 3.00 | 3.00 |
| L-Lysine | 0.40 | 0.40 | 0.40 |
| DL-Methionin | 0.05 | 0.05 | 0.05 |
| Proximate composition % | |||
| Moisture | 6.41 | 6.46 | 6.57 |
| Protein | 39.90 | 39.50 | 39.22 |
| Lipids | 19.20 | 18.75 | 18.04 |
| Ash | 6.79 | 6.29 | 5.73 |
* Vitamin and mineral premix; (fulfilling recommendations for marine fish species given by NRC, 2011.
Table 2.
Chemical composition (g/100 g) of the spray dried protein hydrolysate obtained from salmon raw material.
Table 2.
Chemical composition (g/100 g) of the spray dried protein hydrolysate obtained from salmon raw material.
| Protein hydrolysate | |
|---|---|
| Crude protein | 82.8 |
| Water soluble protein | 68.3 |
| Total dry matter | 98.3 |
| Ash | 6.3 |
| Fat | 11.2 |
| Molecular weight (kDa) % | |
| > 20 | 0.1 |
| 15-20 | < 0.1 |
| 10-15 | 0.1 |
| 8-10 | 0.2 |
| 6-8 | 0.9 |
| 4-6 | 3.9 |
| 2-4 | 16.5 |
| 1-2 | 24.3 |
| 0.5-1 | 19.4 |
| 0.2-0.5 | 13.9 |
| < 0.2 | 20.7 |
Table 3.
List and dilutions of primary and secondary antibodies.
| Primary antibodies | Code | Species | Dilution | Supplier |
| Ghrelin | AM26736PU-N | mouse | 1:500 | Acris |
| Somatostatin | ab16007 | rat | 1:500 | Enzo Life Sciences |
| Neuropeptide Y | NBP1-46535 | goat | 1:1000 | Novus Biological |
| Na+K+-ATPase | GLP-1(1-36) # 9153 | rabbit | 1:600 | Abcam |
| Secondary antibodies | Dilution | Supplier | ||
| goat anti-rat FITC | 1:1000 | Proteintech® | ||
| goat anti-rabbit Alexa Fluor® 594 | 1:1000 | Thermofisher / Invitrogen | ||
| donkey anti-mouse Alexa Fluor® 594 | 1.1000 | Thermofisher / Invitrogen | ||
| donkey anti-goat Alexa Fluor® 488 | 1:1200 | Thermofisher / Invitrogen | ||
Acris Antibodies GmbH OriGene Company, Schillerstraße 5, Herford, Germany. Enzo Life Sciences, New York, NY, USA. Novus Biological, Centennial, CO, USA. Abcam, Cambridge, UK. Proteintech® Group Inc, Rosemont, IL, USA. Thermo Fisher / Invitrogen Scientific, Waltham, MA, USA.
Table 4.
Growth performance for both species Dicentrarchus labrax and Sparus aurata fed the three experimental diets overall 58 days.
Table 4.
Growth performance for both species Dicentrarchus labrax and Sparus aurata fed the three experimental diets overall 58 days.
| Experimental diets | ||||||
|---|---|---|---|---|---|---|
| Dicentrarchus labrax | Sparus aurata | |||||
| CTR | BP5 | BP10 | CTR | BP5 | BP10 | |
| IBW(g) | 73.2 ± 0.6 | 73.3 ± 1.6 | 73.3 ± 0.3 | 72.7 ± 0.8 | 72.7 ± 0.8 | 72.7 ± 0.4 |
| FBW(g) | 145.1 ± 1.8 | 147.1 ± 3.8 | 149.2 ± 2.6 | 169.6 ± 2.9 | 166.7 ± 2.8 | 170.0 ± 2.2 |
| Weight gain(g) | 71.8 ± 2.3 | 73.8 ± 2.3 | 75.9 ± 2.3 | 96.9 ± 2.2 | 94.0 ± 2.1 | 97.3 ± 2.4 |
| SGR | 1.18 ± 0.3 | 1.20 ± 0.01 | 1.23 ± 0.02 | 1.46 ± 0.02 | 1.43 ± 0.01 | 1.46 ± 0.03 |
| FI | 1.47 ± 0.06 | 1.53 ± 0.01 | 1.53 ± 0.02 | 1.87 ± 0.02 | 1.84 ± 0.02 | 1.86 ± 0.03 |
Values are indicated as mean(n=3) ± SD. No significant differences among treatments (One-way Anova P > 0.05). IBW= Initial body weight. FBW= Final body weight. SGR (% day-1) (Specific growth rate) = 100 * (ln FBW- ln IBW) / days. FI (% ABW−1 day−1) (Feed intake) = ((100 ∗ total ingestion)/ (ABW))/days.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



