Submitted:
07 August 2023
Posted:
08 August 2023
You are already at the latest version
Abstract
Krill oil (KO) shows promise as a natural marine-derived ingredient for improving skin health. This study investigated its antioxidant, anti-inflammatory, anti-wrinkle, and moisturizing effects on skin cells and UVB-induced skin photoaging in hairless mice. In vitro assays on HDF, HaCaT, and B16/F10 cells, and in vivo experiments on 66 hairless mice, were conducted. Mice received oral KO administration (100, 200, or 400 mg/kg) once a day for 15 weeks and UVB radiation three times a week. In vitro, KO significantly countered UVB-induced oxidative stress, reduced wrinkles, and prevented skin water loss by enhancing collagen and hyaluronic synthesis. In vivo, all KO dosages showed dose-dependent inhibition of oxidative stress-induced inflammatory photoaging-related skin changes. Skin mRNA expressions for hyaluronan synthesis and collagen synthesis genes also increased dose-dependently after KO treatment. Histopathological analysis confirmed that Krill Oil (KO) ameliorated the damage caused by UVB-irradiated skin tissues. These findings suggest that KO may be a valuable intervention to mitigate UVB-induced skin photoaging and address various skin concerns.
Keywords:
Krill oil
; Ultraviolet
; Skin photoaging
; Marine-derived ingredients
1. Introduction
The human skin, consisting of three layers (epidermis, dermis, and subcutaneous) [1], serves as a vital barrier against environmental stressors like ultraviolet (UV) rays, physical trauma, and microorganisms. Direct exposure to UV radiation from sunlight can lead to acute effects such as DNA damage, suppression of DNA synthesis, cell death, and erythema. Additionally, it can also result in chronic effects like photoaging and epidermal cancer [2]. Photoaging, induced by repeated UV exposure, results in histological alterations, collagen fiber damage, and uneven pigmentation, leading to wrinkled and coarse skin [3,4,5,6]. Unlike the mechanism of UVA-induced pigmentation, Ultraviolet B (UVB), with a wavelength range of 280 to 315 nanometers, exhibits both beneficial and harmful effects on the skin. It can cause sunburn, premature aging, and an increased risk of skin cancer, among other characteristics. [7]. UVB penetrates the epidermis, leading to DNA damage and mutations over time, contributing to skin cancer development[8].
UV rays induce intracellular reactive oxygen species (ROS), causing extracellular matrix (ECM) components imbalance, inflammation, and immunosuppression, leading to skin photoaging [9]. Nutricosmetic products with vitamins, minerals, fatty acids, and other ingredients aim to inhibit oxidative stress and inflammation, improving photoaging and skin conditions [10]. Dietary supplementation with deficient vitamins, minerals, or essential fatty acids is believed to improve skin conditions, as skin function and attractiveness are nutrition-dependent [11]. Essential fatty acids, including alpha-linolenic acid (ALA) and gamma-linolenic acid, belong to omega-3 and omega-6 categories. Docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA), derived from ALA, play a crucial role in skin structure and function [10].
Krill oil (KO) is derived from krill (Euphausia superba), which is a small marine crustacean that feeds on marine algae. Unlike fish oil, which is primarily composed of triglycerides, KO consists of both phospholipids (30-65%) and triglycerides [12]. The main phospholipid found in KO is phosphatidylcholine (PC), and approximately 40% of the total fatty acids bound to PC are known to be EPA and DHA [13]. Therefore, it has been reported that EPA and DHA from KO exhibit higher bioavailability compared to other forms of n-3 PUFAs (such as ethyl-ester and re-esterified omega-3). Although KO has a smaller daily dose of n-3 PUFAs compared to fish oil, it demonstrates higher absorption rates. Hence, KO and fish oil can be considered comparable dietary sources of n-3 PUFAs, even though the EPA and DHA dose in krill oil is 62.8% of that in fish oil. [12]. Recently, KO has been studied not only for its role as a dietary supplement in improving human health but also for its potential in the prevention and/or treatment of skin-related conditions. It is suggested that KO may have beneficial effects on skin health and could be explored as a potential intervention for various skin concerns [14,15].
While KO has been studied for its antioxidant and anti-inflammatory properties, more research is needed to understand its specific effects on skin under environmental stress and its potential to mitigate their impact. This study aims to compare the positive effects of KO with vitamin C (L-ascorbic acid, L-AA) on skin health. L-AA enhances the skin barrier, reduces wrinkles, offers UV protection, inhibits melanogenesis, and provides moisturizing and antioxidant benefits [10]. This study supports KO's (SuperbaTM Boost) positive impact on skin health, including wrinkle improvement, skin moisturization, inflammation reduction, and antioxidant activities. Using a UVB-induced skin photoaging mouse model and in vitro experiments, the study investigates skin photoaging and evaluates KO's potential as a skin protection agent and nutricosmetic ingredient.
2. Results
2.1. In Vitro Evidence Anti-Aging
2.1.1. Cytotoxicity of KO in HDF, HaCaT, and B16/F10 Cells
After treating HDF, HaCaT, and B16/F10 cells with KO at concentrations of 0.25, 0.5, 1, 1.5, 2, 4, and 8 mg/ml for 48 hours, no cytotoxic effects of KO were observed compared to the control. The IC50 values for each cell type were observed to be greater than 8 mg/ml (Figure 1).
2.1.2. Free Radical Scavenging Activity of KO
The evaluation of DPPH radical scavenging activity for L-AA (1 mg/ml) and KO (0.25, 0.5, 1, 1.5, 2, 4, and 8 mg/ml) showed that L-AA exhibited a scavenging activity of 82.00±15.28%, and KO demonstrated significant increases in DPPH radical scavenging activity compared to the control: 27.17±11.14, 49.50±8.14, 59.67±9.20, 63.67±14.80, 75.33±12.23, 81.17±11.65, and 88.33±8.80% (p<0.01) (Figure 2).
2.1.3. Anti-Wrinkle Benefits of KO
PP (10 μM) showed a significant increase in elastase inhibitory activity by 38.17±8.68% compared to the control (p<0.01). Additionally, KO demonstrated a concentration-dependent increase in elastase inhibitory activity (31.50±9.73, 54.50±11.73, 64.50±13.03, 70.17±8.57, 77.50±9.65, and 85.67±7.45%) compared to the control (p<0.01) (Figure 3a). At TGF-β1 10 ng/ml, there was a significant increase of 165.33±31.33% in HDF cell procollagen synthesis compared to the control (p<0.01). Moreover, KO showed a concentration-dependent (KO 0.25 to 8 mg/ml) increase in HDF cell procollagen synthesis (38.17±18.40, 70.00±30.10, 109.33±26.52, 143.00±19.59, 159.33±24.74, 163.83±25.76, and 172.00±27.82%) compared to the control (p<0.01) (Figure 3b). Under UVB irradiation (50mJ/cm2), a significant increase in MMP-1 activity by 65.83±10.48% was observed compared to non-irradiated UVB control cells (p<0.01). However, when treated with RA at 1μM, a significant decrease in MMP-1 activity (-22.13±12.55%) was observed compared to UVB radiation (p<0.01). Similarly, treatment with KO from 0.5 mg/ml to the highest concentration of 8 mg/ml resulted in a significant decrease in MMP-1 activity compared to UVB radiation (-11.30±5.71, -17.92±10.39, -24.63±8.62, -27.67±11.60, -29.75±11.54, -33.12±7.51, and -33.85±10.42%) (p<0.01) (Figure 3c).
2.1.4. Moisturizing Benefits of KO
In HaCaT cells, the evaluation of KO's moisturizing activity through hyaluronan synthesis showed that RA at 1 μM resulted in a significant increase in hyaluronan synthesis (109.17±17.44%) compared to the control (p<0.01). Similarly, KO demonstrated a concentration-dependent (KO 0.25 to 8 mg/ml) increase in hyaluronan synthesis (13.50±7.34, 31.83±11.32, 53.17±13.36, 62.00±14.68, 66.67±12.09, 76.00±11.31, and 80.17±12.02%) compared to the control (p<0.01) (Figure 4).
2.2. In vivo Evidence Anti-Photoaging
2.2.1. Effects of KO on Body Weight Changes
The intact control group's average body weight was 24.93±1.23 g one day before oral administration, and it decreased to 21.77±1.21 g on the start day of administration (Day 0). During the treatment period of 105 days, the average body weight of the intact control group gradually increased to 32.82±2.21 g on the final day of administration (Day 104) (Figure 5). The increase in the average body weight of UVB-irradiated hairless mice group during the 105-day treatment period did not show a significant difference compared to the non-UVB-irradiated intact control group. Additionally, the administration of KO and L-AA groups did not result in significant changes in average body weight compared to the intact control or UVB group.
2.2.2. Effects of KO on UVB-Induced Skin Wrinkle Formation and Skin Moisturization
The efficacy of KO in examining wrinkle formation on UVB-irradiated skin was conducted, and the length and depth of skin wrinkles were evaluated using replicas of the dorsal back skin. Exposure to UVB radiation clearly induced skin wrinkles, but their severity was alleviated by oral administration of either KO or L-AA (Figure 6a). After exposure to UVB, the mean skin length (mm) and depth (µm) significantly increased (0.69±0.07mm, 99.16±17.37µm) compared to the intact control. However, both KO and L-AA significantly suppressed the formation of skin wrinkles in terms of length and depth (Figure 6b, c). Skin water content significantly decreased by 13.16±2.23% after UVB irradiation compared to the intact control group (37.56±5.47%) (p<0.01). However, both KO and L-AA treatments significantly increased skin water content (p<0.05) (Figure 6d). Key molecules for skin wrinkle formation and skin moisture maintenance, skin COL1 contents, and hyaluronic acid contents, were significantly increased by both L-AA and KO treatments compared to UVB irradiation alone (p<0.01) (Figure 6e,f). Moreover, the mRNA expression of hyaluronic acid and COL1 related genes COL1A1, COL1A2, Has1, Has2, and Has3 showed a significant increase compared to UVB irradiation alone (p<0.01) (Figure 6g,h). The mRNA expression of UVB-induced MMP1, MMP9, and MMP13 was significantly reduced by both L-AA and KO treatments (p<0.01) (Figure 6i). There was no significant difference in the treatment effects of UVB-induced skin wrinkle formation and skin moisturization between KO and L-AA treatments with an equal oral dosage.
2.2.3. Effects of KO on UVB-Induced Skin Inflammation
Continuous exposure to UVB irradiation can lead to skin inflammation, causing symptoms such as erythema, swelling, edema, and itching. The evaluation of KO's effect on skin edema is done by measuring the weight of 6mm diameter skin samples. The increased weight of skin samples due to UVB irradiation was significantly reduced by both L-AA and KO treatments (p<0.01) (Figure 7a). Furthermore, UVB irradiation can significantly increase MPO activity, but it was significantly decreased by both L-AA and KO treatments (p<0.01) (Figure 7b). IL-1β levels were significantly increased by UVB irradiation but were significantly suppressed by both L-AA and KO (p<0.01). On the other hand, IL-10 levels were significantly decreased by UVB irradiation but were significantly increased by L-AA and KO treatments (p<0.05) (Figure 7c). There was no significant difference in the treatment effects of UVB-induced skin inflammation between KO and L-AA treatments with an equal oral dosage.
2.2.4. Effects of KO on UVB-Induced Oxidative Stress
The evaluation of Krill Oil's antioxidant effects on skin tissue involved investigating alterations in levels of glutathione (GSH), malondialdehyde (MDA), and superoxide anions.UVB irradiation led to a significant decrease in GSH contents compared to the intact control, but administration of L-AA and KO significantly increased GSH levels (p<0.05) (Figure 8a). Additionally, the increased MDA and superoxide anion levels caused by UVB irradiation were significantly suppressed by the administration of L-AA and KO (p<0.01) (Figure 8b, c). GSH reductase mRNA expression, which was significantly reduced by UVB irradiation compared to the intact control, was significantly increased by the administration of L-AA and KO (p<0.01) (Figure 8d). On the other hand, UVB irradiation significantly increased the mRNA expression of NOX2, an enzyme related to NADPH oxidase-derived ROS generation, but it was significantly decreased by L-AA and KO (p<0.05) (Figure 8e). There was no significant difference in the treatment effects of UVB-induced skin oxidative stress between KO and L-AA treatments with an equal oral dosage.
2.2.5. Effects of KO on UVB-Induced Histopathological Changes in Skin Tissue
Histopathological analysis and Masson's trichrome staining revealed thickened epithelial tissue and abnormal collagen deposition in the dorsal back skin tissue due to UVB irradiation. However, the histopathological changes were significantly mitigated by L-AA and KO treatments (Figure 9a and Table 1). Additionally, KO treatment improved UVB-induced immunolabeled cells for oxidative stress markers (NT and 4-HNE), apoptosis markers (cleaved caspase 3 and cleaved PARP) in the epidermis, and immunoreactive cells for MMP9 in the dermis (Figure 9b and Table 2). However, there was no significant difference in the treatment effects of UVB-induced histopathological changes in skin tissue between KO and L-AA treatments with an equal oral dosage.
3. Discussion
Skin aging can be categorized into intrinsic aging, driven by hormonal changes and cellular aging, and extrinsic aging, caused by external factors like UV exposure, air pollution, and smoking [16]. As the demand for anti-skin aging solutions increases in the market, research and development of natural product-derived ingredients have advanced cutaneous science in skin beauty and health-related industries. Nutricosmetic products, including UV protectors, anti-wrinkle treatments, and moisturizers, are introduced to address these concerns [10]. However, the long-term consumption of cost-effective and functional products raises ongoing concerns about potential risks of adverse events, harmful chemicals, and toxins [17].
Krill oil (KO) is gaining attention for its high bioavailability of n-3 polyunsaturated fatty acids (PUFAs) like EPA and DHA in phospholipid form. Despite being more expensive than fish oil, its superior bioavailability of EPA/DHA has sparked interest. Some clinical studies [12,18,19] have reported minor adverse events, such as rash, headache, taste changes, diarrhea, and decreased appetite. However, KO is Generally Recognized as Safe (GRAS) by the American Food and Drug Administration and has received Novel Food status from the European Union, confirming its safety profile [13].
In our previous studies, KO has been recognized as a marine-derived natural substance with significant activation of nuclear factor E2-related factor 2 (Nrf2) transferase and potent antioxidant properties, making it a promising raw material for health functional foods with natural antioxidant benefits [20,21]. Although previous research on human immortalized keratinocyte lines and NC/Nga mice has suggested the potential benefits of KO in terms of antioxidants and anti-inflammatory effects[14,15], there is a lack of direct and detailed research on KO's role in improving skin wrinkle and moisturization. Hence, this study aims to scientifically evaluate the anti-skin aging effects of KO.
Skin aging is linked to the activation of matrix metalloproteinases (MMPs) triggered by inflammatory cytokines in skin tissue. Frequent exposure to UV radiation accelerates skin aging by causing DNA breakdown, ROS generation, and DNA damage [22]. Nutricosmetic products, known for their antioxidant and anti-inflammatory functions, are anticipated to prevent or improve skin aging. Enzymatic antioxidants have been shown to reduce UV-induced oxidative stress in skin tissue, suppressing inflammation and inhibiting apoptosis of skin cells [11,22]. KO may not be an enzymatic antioxidant, our data demonstrated its inhibition of apoptosis in UVB-exposed skin tissue of mice, suggesting KO's potential in suppressing skin inflammation from UV radiation. Moreover, KO's inhibitory effects may extend to the apoptosis pathway and cell cycle arrest caused by UV-induced DNA damage [23], specifically cyclobutane pyrimidine dimers (CPD) and pyrimidine 6-4 pyrimidine photoproducts (6-4PP). Further research on KO's effects on CPD and 6-4PP and its ability to modulate CPD-photolyases and 6-4PP-photolyases repairing mechanisms would provide a clearer understanding of KO's protection against UV-induced DNA damage and apoptosis. These investigations will enhance our understanding of KO's potential as a skin-protective agent.
Previous studies have reported KO's antioxidant and anti-inflammatory effects on human immortalized keratinocyte lines [14]. KO's potential to regulate ECM proteins and protect the skin through its antioxidant activity and anti-inflammatory effects was supported by its ability to suppress skin inflammation in NC/Nga mice using a phospholipid-enriched alkyl phospholipid from krill [15]. As a non-enzymatic antioxidant, KO builds an antioxidant defense network, protecting cells and tissues from ROS and benefiting skin health. In this study, we observed KO's concentration-dependent free radical scavenging activity. Furthermore, KO administration improved the UVB-induced decrease in GSH contents by upregulating GSH reductase mRNA expression. KO also inhibited UVB-induced lipid peroxidation and superoxide anion production through the transcriptional regulation of NOX2. The observed results, confirmed through immunohistochemical analysis using NT and 4-HNE staining, suggest that KO's antioxidant activity plays a significant role. Additionally, increased ROS due to UV exposure can activate the MAPK (mitogen-activated protein kinase) signaling pathway, leading to the activation of AP-1 (activated protein-1) and subsequently promoting the expression of MMPs (matrix metalloproteinases), which can strongly contribute to the breakdown of ECM proteins like collagen and elastin [24,25,26,27]. Indeed, the increased MMPs due to UV radiation can promote the degradation of ECM proteins and ultimately lead to the formation of skin wrinkles and photoaging [27]. While our study did not directly investigate the UV-induced MAPK pathway and AP-1 activation, we observed that KO's antioxidant activity effectively suppressed the mRNA expression of MMP-1, MMP-9, and MMP-13. This inhibition contributed to the regulation of ECM proteins and directly prevented the formation of skin wrinkles, as evidenced by mean length and depth of wrinkles.
Human skin contains 28 different types of MMPs, including collagenases (MMP-1 and MMP-13) and a gelatinase (MMP-9), which increase with UV exposure [28]. MMP-1 and MMP-13 not only promote ECM collagen degradation but also reduce collagen density in the dermal layer [26,29]. Our study demonstrated that KO reduced UVB-induced MMP-1 activity in HDF cells, indicating its potential to inhibit ECM collagen degradation. Moreover, KO suppressed the upregulation of MMP-1, MMP-9, and MMP-13 gene expression in skin tissue induced by UVB, leading to improved skin COL1 levels and COL1A1/2 expression. This suggests that KO may inhibit MMP expression, likely through its effect on local inflammatory and neutrophil responses to UVB [30]. Such MMP suppression could help maintain skin collagen levels and prevent excessive collagen degradation linked to photoaging. Additionally, our findings show that UV radiation induces an inflammatory response in the skin, with increased secretion of pro-inflammatory cytokine IL-1 and reduced expression of anti-inflammatory cytokine IL-10. KO administration appears to regulate this inflammatory state induced by UV, balancing pro- and anti-inflammatory cytokines, and contributing to its anti-inflammatory effects and potential skin health benefits.
In response to UVB-induced skin injury, polymorphneutrophils (PMNs) and neutrophils are recruited to the injured tissues through the action of oxygen metabolites [31]. MPO, released from PMNs, is a cytotoxic enzyme that activates inflammation [32]. The reduction of neutrophils infiltrating into the skin tissue can be confirmed by assessing MPO activity [33]. Our study revealed that KO alleviates the UVB-induced inflammatory state in the skin and directly inhibits MPO activity, leading to reduced neutrophil recruitment to inflammatory sites [34]. These findings suggest that KO administration modulates the inflammatory response induced by UVB exposure by decreasing MPO activity and subsequently limiting neutrophil infiltration into the skin.
Skin aging often leads to a reduction in hyaluronic acid, a vital component responsible for retaining water in the skin [35]. Fatty acids play a crucial role in maintaining skin hydration and barrier integrity [10], while PUFA deficiency can increase water loss through the skin barrier [36]. Our study found that UVB exposure and aging downregulated the genes responsible for hyaluronic acid synthesis (HAS1, HAS2, and HAS3) in the dermis [37]. However, oral administration of KO reversed this downregulation, resulting in increased hyaluronic acid content in the skin. These results suggest that KO enhances skin moisturization by promoting hyaluronic acid synthesis through regulation of HAS genes in response to UVB-induced water loss.
This study was conducted with the objective of investigating the protective effects of KO against UVB-induced skin photoaging. Existing literature has only offered limited insights into the potential skin health advantages of KO, with two or fewer studies referencing it. Our investigation demonstrated that oral administration of KO notably mitigated UVB-induced wrinkles, skin water loss, collagen degradation, and skin edema, comparable to L-AA (100 mg/kg) at the same dosage. These findings indicate the potential of KO as a functional product for preventing UVB-induced skin photoaging and enhancing skin moisturization. However, further clinical studies are necessary to comprehensively elucidate the diverse range of benefits provided by KO for skin health.
4. Materials and Methods
4.1. In Vitro
4.1.1. Preparation of KO
The commercial product of Antarctic KO (SuperbaTM Boost) was produced by Arker Biomarine (Houston, TX, USA) and was supplied by SC Science (Goyang, Korea) for the study. KO extracted Euphausia superba through steam heating followed by ethanol extraction. The solid particles were separated through filtration and then refined by adding ion exchange resin and NaOH. The refined KO was further processed at 60°C and 360 mmHg vapor pressure for 1 hour to remove ethanol. After centrifugation, a secondary evaporation and filtration were conducted to produce the final product, KO superbaTM Boost. The KO composition included 51.2% (wt/wt) phospholipids, comprising 44.9% phosphatidylcholine, 3.6% 1-palmitoyl-2-hydroxy-glycero-3-phosphocholine, 2.1% phosphatidylethanolamine, and 0.6% N-Acyl-phosphatidylethanolamine [21]. Additionally, EPA and DHA contents were analyzed in the KO. Methanol-treated KO was subjected to sodium hydroxide and boron trifluoride esterification, followed by dissolution in isooctane for analysis using Gas Chromatography (GC). A gas chromatograph (Agilent, USA) equipped with an SP®-2560 capillary GC column (100 m × 0.25 mm, 0.20 μm) and a Flame Ionization Detector (GC/FID) was used. Helium was employed as the carrier gas at a flow rate of 0.75 mL/min with a split ratio of 200:1. The injector and detector (FID) temperatures were set at 225°C and 285°C, respectively. The column temperature was maintained at 100°C for 4 minutes, followed by an increase to 240°C at a rate of 3°C/min. Quantification was achieved by calculating the peak areas of each fatty acid obtained from test and standard solutions, along with the peak area of internal standard substances. The EPA and DHA contents in KO were determined to be 296 mg/g. The KO was stored at -20°C and utilized consistently for in vitro and in vivo studies.
L-AA, RA, PP, and arbutin were purchased from Sigma-Aldrich (St. Louis, MO, USA), while TGF-β1 was obtained from R&D Systems (Minneapolis, MN, USA). The treatment concentrations for KO in this study were chosen as 0.25, 0.5, 1, 1.5, 2, 4, and 8 mg/ml, based on previously established conditions in our laboratory [38,39]. Similarly, the treatment concentrations for the standard references, L-AA (1 mg/ml), PP (10 μM), RA (1 μM), TGF-β1 (10 ng/ml), and arbutin (2 mM), were selected based on our previous reports[38,39], for use in the current experiment.
4.1.2. Cell Cultures
HaCaT (PCS-200-011) and B16/F10 melanoma (CRL-6475) cells were procured from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured following the manufacturer's protocols. The cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS; Lonza, Walkersville, MD, USA), 100 µg/mL streptomycin, and 100 U/ml penicillin (Sigma-Aldrich, St. Louis, MO, USA). For B16/F10 murine melanoma cells, 2 mM L-glutamine (Sigma-Aldrich, St. Louis, MO, USA) was additionally added to the culture medium. HDF cell lines (PCS-201-010) were also obtained from ATCC (Manassas, VA, USA) and cultured in fibroblast basal medium (FBM; PCS-201-030, ATCC, Manassas, VA, USA) supplemented with FBM low serum kit (PCS-201-041; ATCC, Manassas, VA, USA). For treatments, FBM serum-free kit (PCS-201-040; ATCC, Manassas, VA, USA) was used. All cells were cultured at 37°C in a fully humidified atmosphere with 5% CO2 using a commercial CO2 incubator (Model 311, Thermo Forma, Marietta, OH, USA) and were passaged approximately every other day.
4.1.3. Cell Viability Assay
HaCaT, HDF, and B16/F10 cells were seeded at 1 × 105 cells/well in 96-well plates and exposed to various concentrations of KO (0.25, 0.5, 1, 1.5, 2, 4, and 8 mg/ml) for 48 hours. Cell viability was assessed using MTT reagent (Sigma-Aldrich, St. Louis, MO, USA) with a concentration of 2 mg/ml. The plates were then incubated in a CO2 incubator at 37°C for 2 hours. To measure the cell viability, the absorbance (optical density, OD) of the wells at 450 nm was recorded using a microplate reader (Sunrise, TECAN, Männedorf, Switzerland). The relative cell viability (%) was calculated as [(ODs / ODc) × 100], where ODs represents the absorbance of the sample at 450 nm, and ODc is the absorbance of the vehicle control at 450 nm. The results were expressed in terms of IC50, which indicates the concentration at which cell viability reaches 50% of the control.
4.1.4. DPPH Radical Scavenging Activity Test
Samples were diluted with distilled water to achieve final KO concentrations of 0.25 to 8 mg/ml or a final L-AA concentration of 1 mg/ml. The DPPH radical scavenging activity was measured at 517 nm using a UV/Vis spectrophotometer after 10 minutes. The results were reported as IC50, representing the concentration required to reduce 50% of DPPH. L-AA at 1 mg/ml served as the positive control.
4.1.5. Elastase Inhibition Assay
The elastase inhibition assay measured the release of p-nitroaniline due to proteolysis of N-succinyl-(Ala)3-p-nitroanilide by human leucocyte elastase (Sigma-Aldrich, St. Louis, MO, USA) [40]. KO was tested at concentrations ranging from 0.25 to 8 mg/ml, while PP at 10 μM served as the standard. Elastase inhibitory activity was measured at 410 nm using a 96-well microplate reader. The results were reported as IC50, representing the concentration at which the percentage inhibition of elastase activity was 50%.
4.1.6. Procollagen Content Test.
HDF cells were cultured in 24-well plates and treated with 50 μg/ml medium containing various concentrations of KO (0.25 to 8 mg/ml) or TGF-β1 at 10 ng/ml. Procollagen contents in the culture supernatant were measured using a Procollagen type I-c-peptide (PIP) ELISA kit. The results were reported as EC50, representing the concentration at which the percentage increase in HDF cell procollagen synthesis was 2-fold higher.
4.1.7. MMP-1 Inhibition Test
The assay was conducted using a fluorescence microplate reader (RF-5301PC; Shimadzu Corp., Tokyo, Japan) following modified methods of Losso et al. [41]. HDF cells were treated with 50 µg/ml of KO (0.25 to 8 mg/ml) or 1 μM of RA in the absence or presence of UVB (5 mJ/cm2) for 2 minutes. Total MMP-1 protein levels in the cell culture supernatants were quantified using the Human Total MMP-1 ELISA kit (DY901; R&D Systems, Minneapolis, MN, USA). The results were reported as IC50, representing the concentration at which the percentage inhibition of UVB-induced HDF cell MMP-1 expressions was 50%.
4.1.8. Hyaluronan Production Assay
HaCaT cells (4 × 104 cells/well) were treated with KO (0.25-8 mg/ml) or 1 μM of RA for 24 hours. Hyaluronan synthesis was quantified using the Hyaluronan ELISA kit (DY3614, R&D Systems). Results were normalized based on total protein content. The relative hyaluronan synthesis (%) was calculated as [(Hyaluronan contents in experimental sample / Hyaluronan contents in unexposed control) × 100]. The results were reported as EC50, representing the concentration at which the percentage increase in hyaluronan synthesis was 2-fold higher.
4.2. In Vivo
A total of 66 SPF/VAF Outbred SKH1-hr hairless female mice (OrientBio, Seungnam, Korea) were obtained. After a 7-day acclimation period, mice with normal skin and stable body weight (average 24.91±1.14 g, range: 22.60 ~ 27.00 g) were divided into 6 groups, with 10 mice per group. The animal experiment was conducted with prior approval from the Institutional Animal Care and Use Committee of Daegu Haany University. [Approval No. DHU2021-070, August 18, 2021].
4.2.1. Preparation of Experimental Group
The mice were divided into a total of 6 groups, with 10 mice per group: 1) Normal control group (no UVB exposure; administered sterile distilled water orally); 2) UVB control group (UVB exposure control; administered sterile distilled water orally); 3) L-AA group (UVB exposure and L-AA 100 mg/kg oral administration); 4) KO100 group (UVB exposure and KO 100 mg/kg oral administration); 5) KO200 group (UVB exposure and KO 200 mg/kg oral administration); and 6) KO400 group (UVB exposure and KO 400 mg/kg oral administration).
4.2.2. Skin Photoaging
Following the methods used in previous studies [42,43], a UV Crosslinker system (CL-1000M, Analytik Jena, Upland, CA, USA) emitting wavelengths of 254 nm, 312 nm, and 365 nm (with 312 nm as the main wavelength) was used to induce skin photoaging in hairless mice. The mice were exposed to UVB at a dose of 0.18 J/cm2, three times per week, for 15 weeks. For the normal control group, the same environment stress was applied by leaving the UV Crosslinker system powered off under the same conditions for the same duration.
4.2.3. Experimental Substances and Administration
The experiment substances were prepared by dissolving KO at concentrations of 40, 20, and 10 mg/ml in sterile distilled water. They were administered orally using a metal gavage needle attached to a 1 ml syringe at doses of 10 ml/kg (100, 200, and 400 mg/kg) daily for 105 days, 1 hour after UVB exposure. L-AA was also dissolved in sterile distilled water at a concentration of 10 mg/ml and administered orally at a dose of 10 ml/kg (10 mg/kg) daily for 105 days, 1 hour after UVB exposure. In the normal control and UVB control groups, only sterile distilled water was administered orally at the same volume and duration as the experimental substances to apply the same handling stress. The experimental substances were prepared at least once a week and stored in a refrigerator at 4°C until use.
4.2.4. Antioxidant and Anti-Inflammatory
One week before UVB irradiation, the body weight of hairless mice was measured at one-week intervals. On the final sacrifice day, replicas of the skin near the dorsal back were collected to measure the average length and depth of formed wrinkles. The skin weight per unit area (6mm in diameter) was measured to assess skin edema. Changes in MPO activity, proinflammatory cytokines (IL-1β and IL-10) levels, along with histopathological changes, were measured to evaluate the anti-inflammatory effect. Modifications in the levels of glutathione (GSH), the extent of lipid peroxidation, the generation of superoxide anions, and the mRNA expression of Nox2 (gp91phox) and GSH reductase were monitored through real-time reverse transcription polymerase chain reaction (RT-PCR) analysis to assess alterations in the antioxidant defense systems. Moreover, RT-PCR was employed to detect variations in the mRNA expression of TGF-β1, p38 MAPK, AKT, MMP-1, MMP-9, and MMP-13. The primer sequences utilized for the RT-PCR are provided in Table S1. Immunoreactivity of MMP-9 in the dermis, and immunoreactivity of apoptosis markers - caspase-3 and PARP in epidermal keratinocytes were also observed.
4.2.5. Moisturizing
The moisture content, COL1, and hyaluronan levels near the dorsal back skin (6mm in diameter) of hairless mice were observed using RT-PCR to assess the moisturizing effect. The mRNA expression of hyaluronan synthesis-related genes Has1, Has2, and Has3, as well as collagen synthesis-related genes COL1A1 and COL1A2, were measured to evaluate the effects on hyaluronan and collagen synthesis.
4.2.6. Anti-Wrinkle
The number of microfold wrinkles formed on the skin tissue surface, average epidermal thickness, the number of infiltrating inflammatory cells in the dermis, and the percentage of collagen fibers in the dermis (%/mm2) were measured using histomorphometrical analysis with a tissue processing camera and an automated video analysis system. The number of immunoreactive cells (NT, 4-HNE, cleaved caspase-3, and PARP) per 100 epithelial cells, as well as the percentage of MMP-9 immunoreactive area in the dermis (%/mm2), were also quantified using the same method [42,43].
4.3. Statistical Analyses
All collected data were presented as mean ± SD. In vitro and in vivo data were subjected to multiple comparison tests, including the Levene test to assess variance homogeneity. One-way ANOVA followed by Tukey's HSD test was applied to data with no significant deviations from variance homogeneity, while Dunnett's T3 test was used for data with significant deviations. Nonparametric comparisons were performed using the Kruskal-Wallis H test and Mann-Whitney u test. Statistical significance was considered for p-values < 0.05. Statistical analysis was conducted using SPSS for Windows (Release 27.0).
5. Conclusions
In this investigation, we assessed KO's potential in mitigating UVB-induced skin photoaging. Our experimental outcomes demonstrated that oral administration of KO significantly attenuated UVB-induced wrinkle formation, skin water loss, and collagen degradation. These advantageous effects were attributed to the anti-inflammatory, anti-apoptotic, and antioxidant properties inherent in KO, which exhibited similarity to L-AA (100 mg/kg) at an equivalent oral dose level. Given these compelling results, KO emerges as a promising therapeutic candidate or natural resource for the development of functional products targeted at preventing skin photoaging.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: GC analysis of KO for EPA+DHA contents; Table S1. Primer sequences for real-time RT-PCR.
Author Contributions
Conceptualization, J.K. and S.-K.K; Methodology, S.-H.L. and S.-K.K; Software, J.K. and Y.-S.C; Validation, J.K. and N.L.; Formal Analysis, Y.-S.C. and J.K.; Investigation, S.-K.K.; Resources, J.K and S.-K.K; Data Curation, S.-K.K; Writing – Original Draft Preparation, J.K and N.L; Writing – Review & Editing, K.-S.K, J.K, and N.L.; Visualization, J.K., and N.L.; Supervision, S.-K.K.; Project Administration, J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (2018AR1A5A2025272) and by the KO functional food research and development support fund (KO2021A0907) from SC science.
Institutional Review Board Statement
The animal study protocol was conducted according to the national regulations for the usage and welfare of laboratory animals based on the NIH guideline and approved by the Institutional Animal Care and Use Committee of Daegu Haany University (Approval No. DHU2021-070, August 18, 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We extend our sincere appreciation for the administrative and technical support received from the College of Korean Medicine, Daegu Hanny University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kolarsick, P.A.; Kolarsick, M.A.; Goodwin, C. Anatomy and physiology of the skin. JDNA 2011, 3, 203–213. [Google Scholar] [CrossRef]
- McKnight, G.; Shah, J.; Hargest, R. Physiology of the skin. Surgery (Oxford) 2022, 40, 8–12. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The cutaneous serotoninergic/melatoninergic system: securing a place under the sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.; Morison, W.L.; Sauder, D.N. Photoaging: mechanisms and repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef]
- Kawada, S.; Ohtani, M.; Ishii, N. Increased oxygen tension attenuates acute ultraviolet-B-induced skin angiogenesis and wrinkle formation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R694–R701. [Google Scholar] [CrossRef]
- Juzeniene, A.; Moan, J. Beneficial effects of UV radiation other than via vitamin D production. Dermatoendocrinol. 2012, 4, 109–117. [Google Scholar] [CrossRef]
- Narayanan, D.L.; Saladi, R.N.; Fox, J.L. Ultraviolet radiation and skin cancer. Int. J. Dermatol. 2010, 49, 978–986. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Photoaging: UV radiation-induced inflammation and immunosuppression accelerate the aging process in the skin. Inflamm. Res. 2022, 71, 817–831. [Google Scholar] [CrossRef]
- Michalak, M.; Pierzak, M.; Kręcisz, B.; Suliga, E. Bioactive compounds for skin health: A review. Nutrients 2021, 13, 203. [Google Scholar] [CrossRef]
- Boelsma, E.; Hendriks, H.F.; Roza, L. Nutritional skin care: health effects of micronutrients and fatty acids. Am. J. Clin. Nutr. 2001, 73, 853–864. [Google Scholar] [CrossRef]
- Ulven, S.M.; Kirkhus, B.; Lamglait, A.; Basu, S.; Elind, E.; Haider, T.; Berge, K.; Vik, H.; Pedersen, J.I. Metabolic effects of krill oil are essentially similar to those of fish oil but at lower dose of EPA and DHA, in healthy volunteers. Lipids 2011, 46, 37–46. [Google Scholar] [CrossRef]
- Ramprasath, V.R.; Eyal, I.; Zchut, S.; Jones, P.J. Enhanced increase of omega-3 index in healthy individuals with response to 4-week n-3 fatty acid supplementation from krill oil versus fish oil. Lipids Health Dis. 2013, 12, 1–11. [Google Scholar] [CrossRef]
- Fasano, E.; Serini, S.; Mondella, N.; Trombino, S.; Celleno, L.; Lanza, P.; Cittadini, A.; Calviello, G. Antioxidant and anti-inflammatory effects of selected natural compounds contained in a dietary supplement on two human immortalized keratinocyte lines. Biomed Res. Int. 2014. [Google Scholar] [CrossRef]
- Watanabe, N.; Suzuki, T.; Yamazaki, Y.; Sugiyama, K.; Koike, S.; Nishimukai, M. Supplemental feeding of phospholipid-enriched alkyl phospholipid from krill relieves spontaneous atopic dermatitis and strengthens skin intercellular lipid barriers in NC/Nga mice. Biosc. Biotechnol. Biochem. 2019, 83, 717–727. [Google Scholar] [CrossRef]
- Farage, M.; Miller, K.; Elsner, P.; Maibach, H. Intrinsic and extrinsic factors in skin ageing: a review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Park, H.R.; Kim, J.K.; Lee, J.K.; Choi, B.R.; Ku, S.K.; Jegal, K.H. The Protective Effects of Unripe Apple (Malus pumila) Extract on Ultraviolet B-Induced Skin Photoaging Mouse Model. Appl. Sci. 2023, 13, 4788. [Google Scholar] [CrossRef]
- Berge, K.; Musa-Veloso, K.; Harwood, M.; Hoem, N.; Burri, L. Krill oil supplementation lowers serum triglycerides without increasing low-density lipoprotein cholesterol in adults with borderline high or high triglyceride levels. Nutr. Res. 2014, 34, 126–133. [Google Scholar] [CrossRef]
- Mödinger, Y.; Schön, C.; Wilhelm, M.; Hals, P.-A. Plasma kinetics of choline and choline metabolites after a single dose of SuperbaBoostTM krill oil or choline bitartrate in healthy volunteers. Nutrients 2019, 11, 2548. [Google Scholar] [CrossRef]
- Lee, H.J.; Seo, J.W.; Chun, Y.S.; Kim, J.; Lim, T.G.; Shim, S.M. Krill oil inhibited adipogenic differentiation by inducing the nuclear Nrf2 expression and the AMPK activity. Food Sci. Nutr. 2023. [Google Scholar] [CrossRef]
- Hwang, S.-M.; Kim, Y.U.; Kim, J.-K.; Chun, Y.-S.; Kwon, Y.-S.; Ku, S.-K.; Song, C.-H. Preventive and Therapeutic Effects of Krill Oil on Obesity and Obesity-Induced Metabolic Syndromes in High-Fat Diet-Fed Mice. Mar. Drugs 2022, 20, 483. [Google Scholar] [CrossRef] [PubMed]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory molecules associated with ultraviolet radiation-mediated skin aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, G.S.; Quayle, C.; Garcia, C.; Fotoran, W.L.; Dos Santos, J.F.; van der Horst, G.T.; Hoeijmakers, J.H.; Menck, C.F. Photorepair of either CPD or 6-4PP DNA lesions in basal keratinocytes attenuates ultraviolet-induced skin effects in nucleotide excision repair deficient mice. Front. Immunol. 2022, 13, 800606. [Google Scholar] [CrossRef]
- Karin, M. The regulation of AP-1 activity by mitogen-activated protein kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-activated protein kinases and reactive oxygen species: how can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Lin, T.-Y.; Wu, P.-Y.; Hou, C.-W.; Chien, T.-Y.; Chang, Q.-X.; Wen, K.-C.; Lin, C.-Y.; Chiang, H.-M. Protective effects of sesamin against UVB-induced skin inflammation and photodamage in vitro and in vivo. Biomolecules 2019, 9, 479. [Google Scholar] [CrossRef]
- Choi, H.-J.; Alam, M.B.; Baek, M.-E.; Kwon, Y.-G.; Lim, J.-Y.; Lee, S.-H. Protection against UVB-induced photoaging by Nypa fruticans via inhibition of MAPK/AP-1/MMP-1 signaling. Oxid. Med. Cell. Longev. 2020. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investiga. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation–a review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Sullivan, G.W.; Sarembock, I.J.; Linden, J. The role of inflammation in vascular diseases. J. Leukoc. Biol. 2000, 67, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Íşeri, S.O.; Sener, G.; Yuksel, M.; Contuk, G.; Cetinel, S.; Gedik, N.; Yeğen, B.C. Ghrelin against alendronate-induced gastric damage in rats. J. Endocrinol. 2005, 187, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.; Rao, V.; Carvalho, C.; Bezerra-Filho, J.; Fonseca, S.; Vale, M.; Montenegro, D.; Cunha, F.; Ribeiro, R.; Brito, G. Lippia sidoides and Myracrodruon urundeuva gel prevents alveolar bone resorption in experimental periodontitis in rats. J. Ethnopharmacol. 2007, 113, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Campanini, M.Z.; Pinho-Ribeiro, F.A.; Ivan, A.L.; Ferreira, V.S.; Vilela, F.M.; Vicentini, F.T.; Martinez, R.M.; Zarpelon, A.C.; Fonseca, M.J.; Faria, T.J. Efficacy of topical formulations containing Pimenta pseudocaryophyllus extract against UVB-induced oxidative stress and inflammation in hairless mice. J. Photochem. Photobiol. B. 2013, 127, 153–160. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Meguro, S.; Arai, Y.; Masukawa, Y.; Uie, K.; Tokimitsu, I. Relationship between covalently bound ceramides and transepidermal water loss (TEWL). Arch. Dermatol. Res. 2000, 292, 463–468. [Google Scholar] [CrossRef]
- Dai, G.; Freudenberger, T.; Zipper, P.; Melchior, A.; Grether-Beck, S.; Rabausch, B.; de Groot, J.; Twarock, S.; Hanenberg, H.; Homey, B. Chronic ultraviolet B irradiation causes loss of hyaluronic acid from mouse dermis because of down-regulation of hyaluronic acid synthases. Am. J. Pathol. 2007, 171, 1451–1461. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, S.J.; Lee, J.E.; Lee, Y.J.; Song, C.H.; Choi, S.H.; Ku, S.K.; Kang, S.J. Anti-skin-aging benefits of exopolymers from Aureobasidium pullulans SM2001. J. Cosmet. Sci. 2014, 65, 285–298. [Google Scholar]
- Kang, S.J.; Choi, B.R.; Kim, S.H.; Yi, H.Y.; Park, H.R.; Park, S.J.; Song, C.H.; Park, J.H.; Lee, Y.J.; Kwang, S. Inhibitory effects of pomegranate concentrated solution on the activities of hyaluronidase, tyrosinase, and metalloproteinase. J. Cosmet. Sci. 2015, 66, 145–159. [Google Scholar] [CrossRef]
- Jin-Chul, H.; Sang-Mi, A.; Chi-Young, Y.; Heung-Mook, S.; Taeg-Kyu, K. Anti-oxidant and anti-tumor activities of crude extracts by Gastrodia elata Blume. Korean J. Food Preserv. 2006, 13, 83–87. [Google Scholar]
- Losso, J.N.; Munene, C.N.; Bansode, R.R.; Bawadi, H.A. Inhibition of matrix metalloproteinase-1 activity by the soybean Bowman–Birk inhibitor. Biotechnol. lett. 2004, 26, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Kim, H.Y.; Oh, M.H.; Byun, S.; Lim, S.H.; Heo, Y.-S.; Kang, N.J.; Bode, A.M.; Dong, Z. Myricetin suppresses UVB-induced wrinkle formation and MMP-9 expression by inhibiting Raf. Biochem. Pharmacol. 2010, 79, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-I.; Oh, W.-S.; Song, P.H.; Yun, S.; Kwon, Y.-S.; Lee, Y.J.; Ku, S.-K.; Song, C.-H.; Oh, T.-H. Anti-photoaging effects of low molecular-weight fucoidan on ultraviolet B-irradiated mice. Mar. Drugs 2018, 16, 286. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cytotoxicity of KO on HDF (a), HaCaT cell (b), and B16F10 (c) B16/F10 Cells: Data are presented as the mean ± standard deviation (SD). KO, Krill oil (SuperbaTM Boost); HaCaT, Human keratinocytes; B16F10, Murine melanoma cells.
Figure 1.
Cytotoxicity of KO on HDF (a), HaCaT cell (b), and B16F10 (c) B16/F10 Cells: Data are presented as the mean ± standard deviation (SD). KO, Krill oil (SuperbaTM Boost); HaCaT, Human keratinocytes; B16F10, Murine melanoma cells.
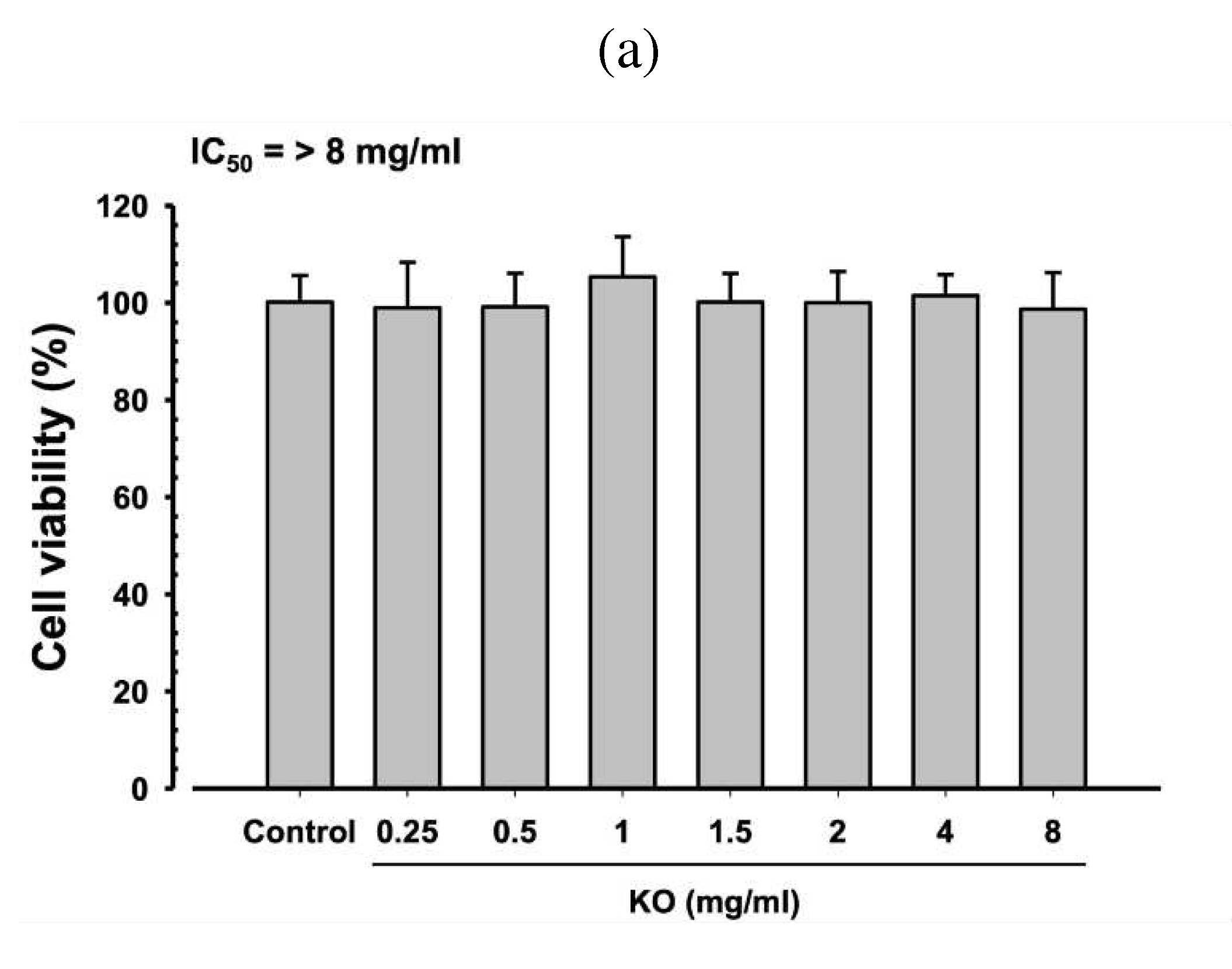
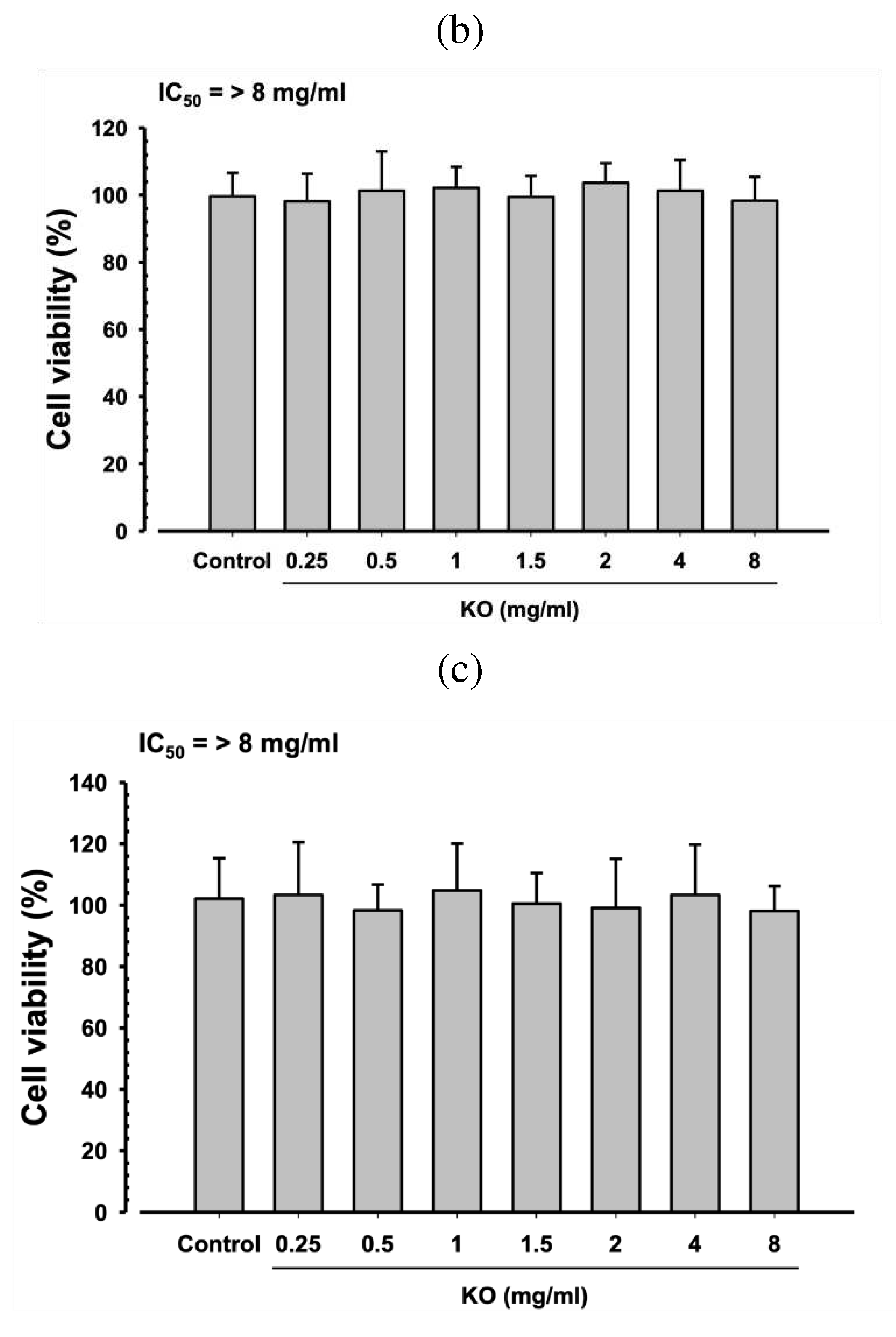
Figure 2.
The antioxidant characteristics of KO: Data are presented as the mean ± SD. KO, Krill oil (SuperbaTM Boost); L-AA, L-Ascorbic acid; DPPH, 1-Diphenyl-2-picryhydrazyl radical, 2,2-Diphenyl-1-(2,4,6-trinitrophemyl) hydrazyl. ** p<0.01 as compared with control cells.
Figure 2.
The antioxidant characteristics of KO: Data are presented as the mean ± SD. KO, Krill oil (SuperbaTM Boost); L-AA, L-Ascorbic acid; DPPH, 1-Diphenyl-2-picryhydrazyl radical, 2,2-Diphenyl-1-(2,4,6-trinitrophemyl) hydrazyl. ** p<0.01 as compared with control cells.
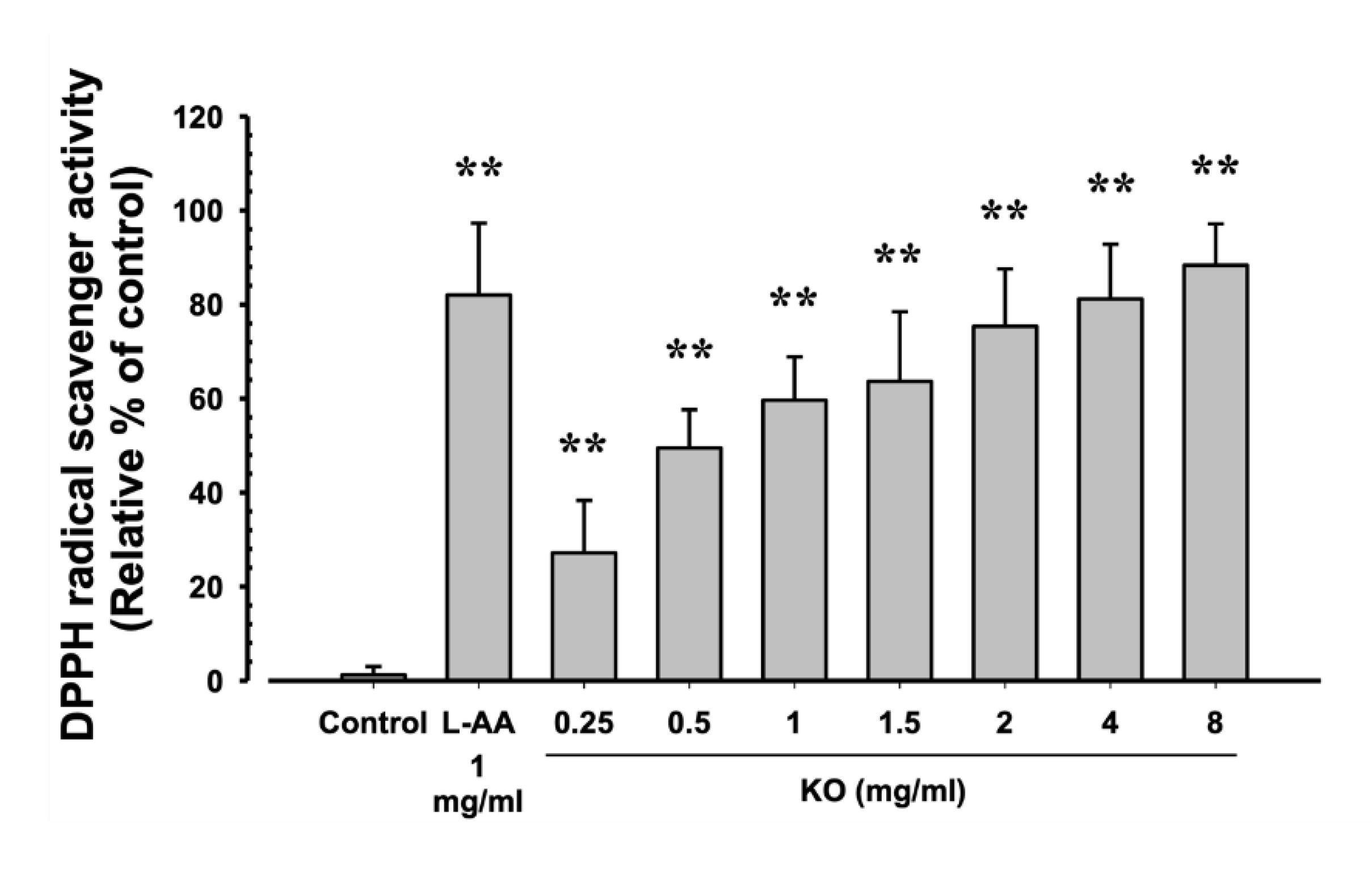
Figure 3.
Anti-winkle of KO: (a) elastase inhibitory activity; (b) collagen synthesis; (c) MMP-1 activity. Data are presented as the mean ± SD. KO, Krill oil (SuperbaTM Boost); PP, phosphoramidon disodium salt; TGF, transforming growth factor; MMP, matrix metalloproteinase; RA, retinoic acid; UVB, ultraviolet B; HDF, human dermal fibroblasts; * p<0.05 and ** p<0.01 as compared with control cells. ## p<0.01 as compared with UVB irradiated control cells.
Figure 3.
Anti-winkle of KO: (a) elastase inhibitory activity; (b) collagen synthesis; (c) MMP-1 activity. Data are presented as the mean ± SD. KO, Krill oil (SuperbaTM Boost); PP, phosphoramidon disodium salt; TGF, transforming growth factor; MMP, matrix metalloproteinase; RA, retinoic acid; UVB, ultraviolet B; HDF, human dermal fibroblasts; * p<0.05 and ** p<0.01 as compared with control cells. ## p<0.01 as compared with UVB irradiated control cells.
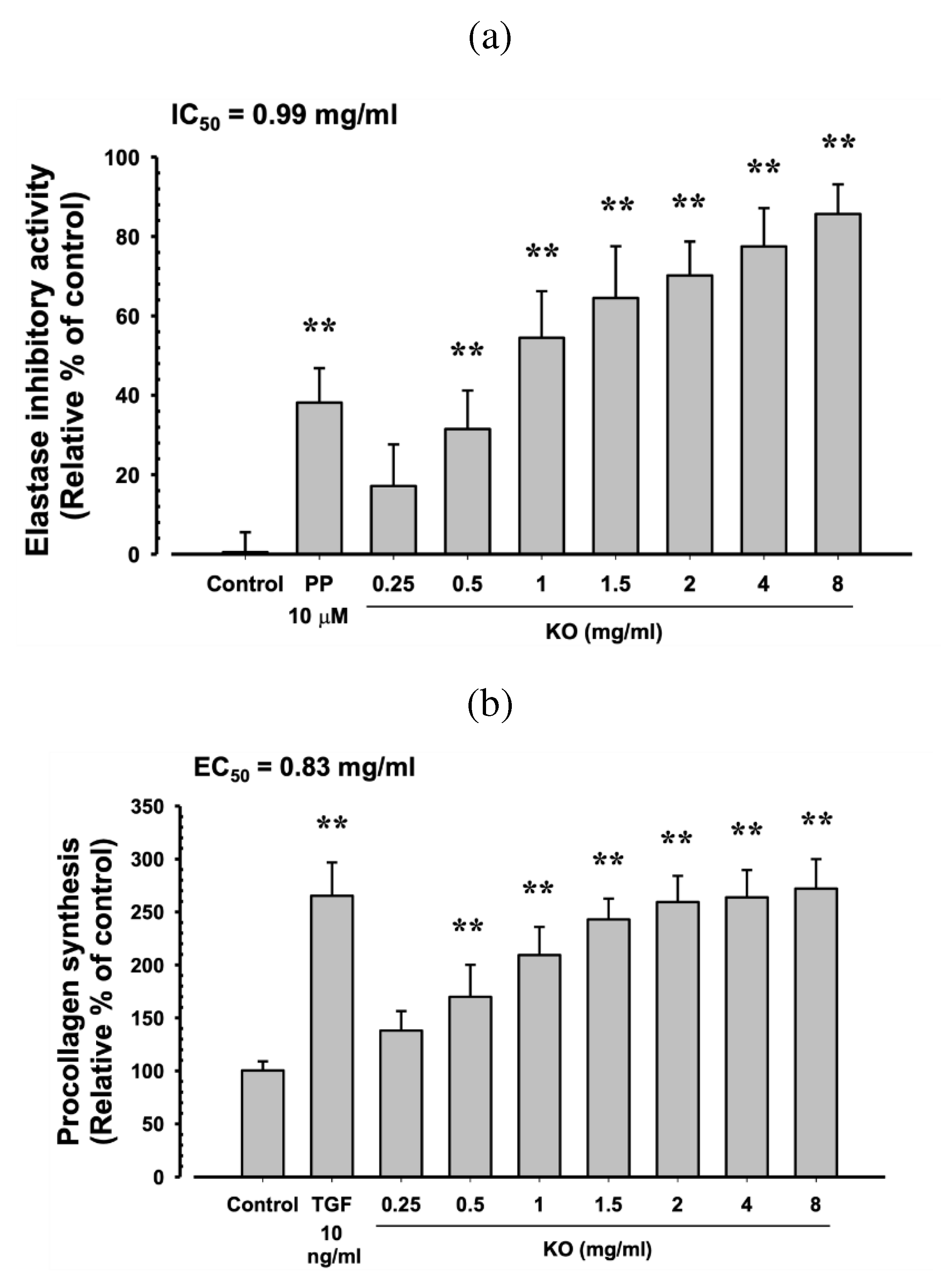
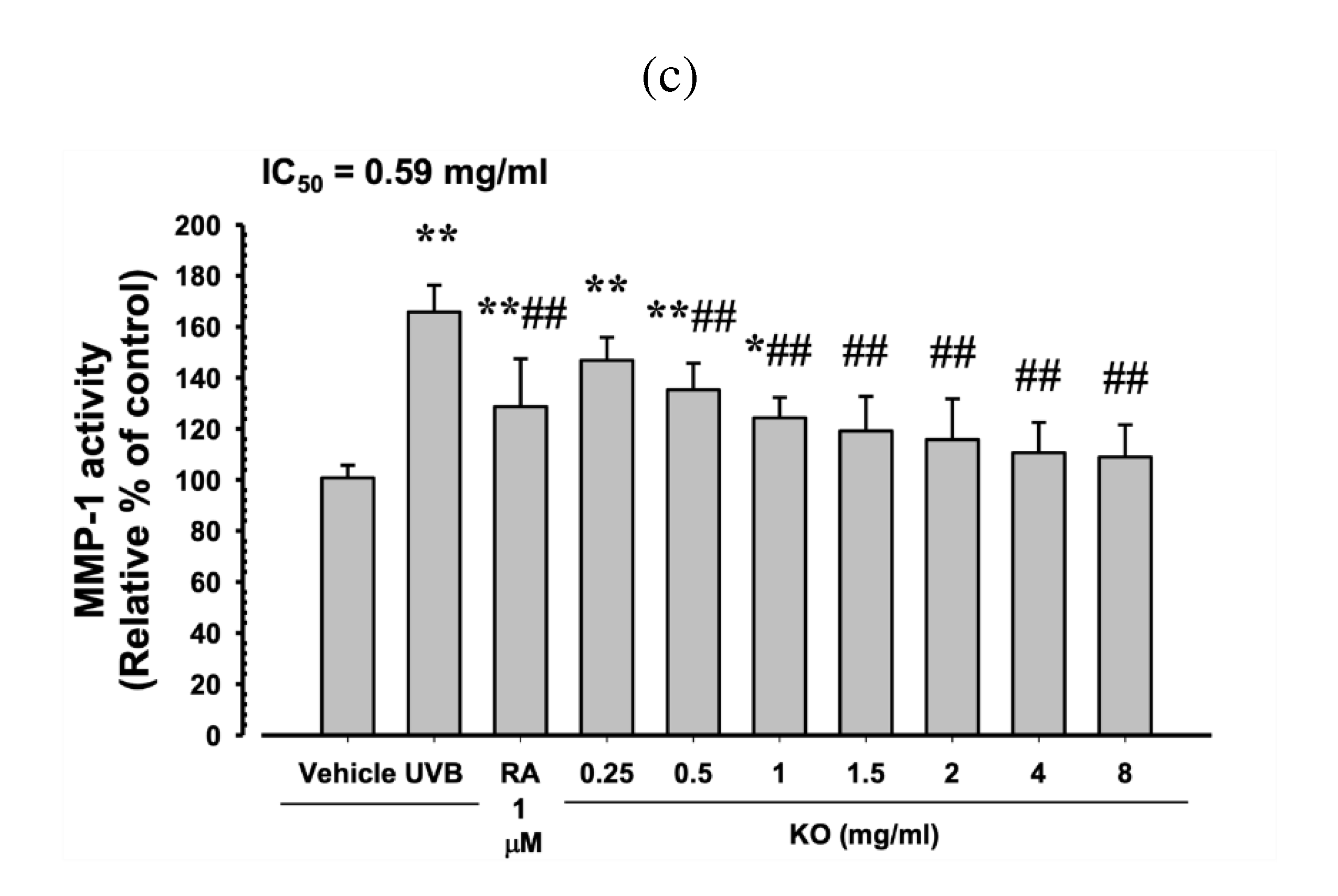
Figure 4.
The effects of KO on hyaluronan synthesis: Data are presented as the mean ± SD. KO, krill oil (SuperbaTM Boost); RA, retinoic acid; HaCaT, Human keratinocytes; ** p<0.01 as compared with control cells.
Figure 4.
The effects of KO on hyaluronan synthesis: Data are presented as the mean ± SD. KO, krill oil (SuperbaTM Boost); RA, retinoic acid; HaCaT, Human keratinocytes; ** p<0.01 as compared with control cells.
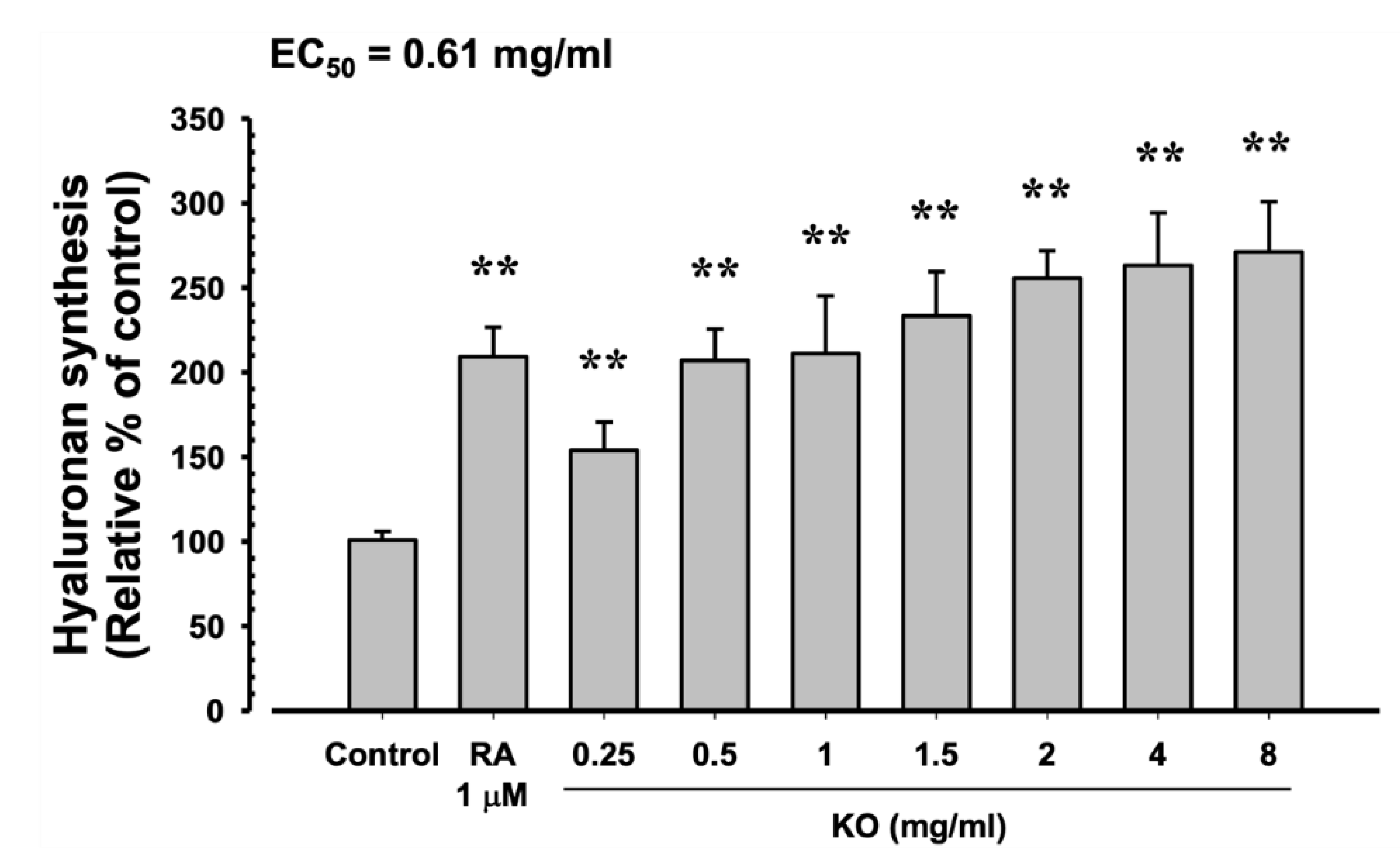
Figure 5.
Body weight changes on the days after UVB irradiation and oral administration: KO (100, 200, and 400 mg/kg) or L-AA (100 mg/kg) was orally administrated once a day for 105 days after 1h of UVB irradiation. The body weights were measured every week. Data are presented as the mean ± SD (n=10, significance compared with intact control mice).
Figure 5.
Body weight changes on the days after UVB irradiation and oral administration: KO (100, 200, and 400 mg/kg) or L-AA (100 mg/kg) was orally administrated once a day for 105 days after 1h of UVB irradiation. The body weights were measured every week. Data are presented as the mean ± SD (n=10, significance compared with intact control mice).
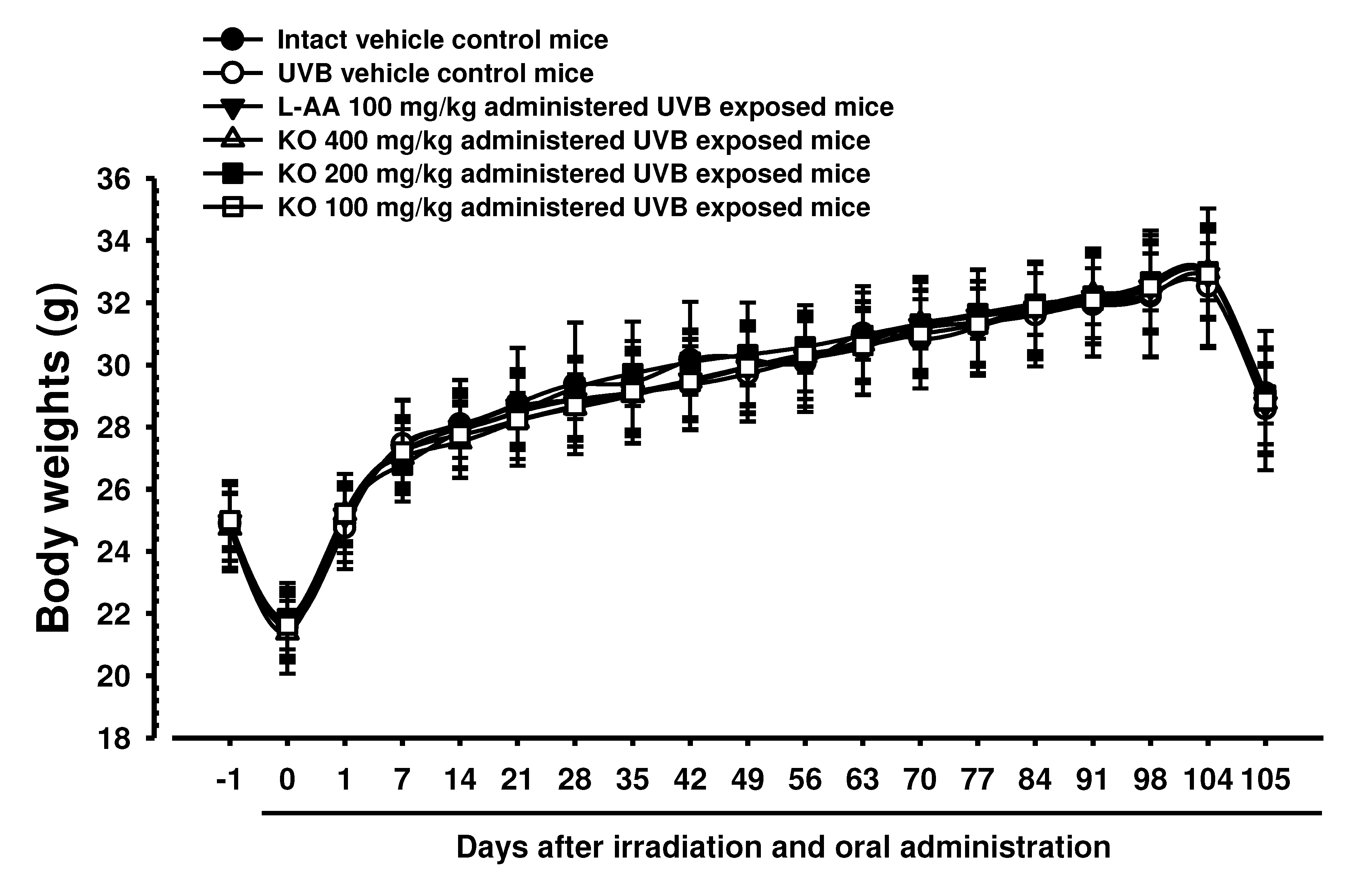
Figure 6.
Effects of KO on UVB-induced wrinkle formation in dorsal back skin: (a) Photograph of dorsal back skin (upper), monochrome image of skin replicas (lower). Scale bars indicate 10 mm. Wrinkle shadows were generated using an optic light source by a fixed intensity at a 40゜angle.; (b) and (c) Wrinkle length and depth; (d) Skin water contents (6mm-diameter skin); (e) skin COL1 contents (%, relative to intact); (f) skin hyaluronic acid contents; (g) COL1 synthetic (COL1A1 and COL1A2) in dorsal back skin tissue; (h) hyaluronic acid synthesis (HAS 1, HAS 2, and HAS 3) in dorsal back skin tissue;(i) MMP (MMP-1, MMP-9, and MMP13) gene expression in dorsal back skin tissue. Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
Figure 6.
Effects of KO on UVB-induced wrinkle formation in dorsal back skin: (a) Photograph of dorsal back skin (upper), monochrome image of skin replicas (lower). Scale bars indicate 10 mm. Wrinkle shadows were generated using an optic light source by a fixed intensity at a 40゜angle.; (b) and (c) Wrinkle length and depth; (d) Skin water contents (6mm-diameter skin); (e) skin COL1 contents (%, relative to intact); (f) skin hyaluronic acid contents; (g) COL1 synthetic (COL1A1 and COL1A2) in dorsal back skin tissue; (h) hyaluronic acid synthesis (HAS 1, HAS 2, and HAS 3) in dorsal back skin tissue;(i) MMP (MMP-1, MMP-9, and MMP13) gene expression in dorsal back skin tissue. Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
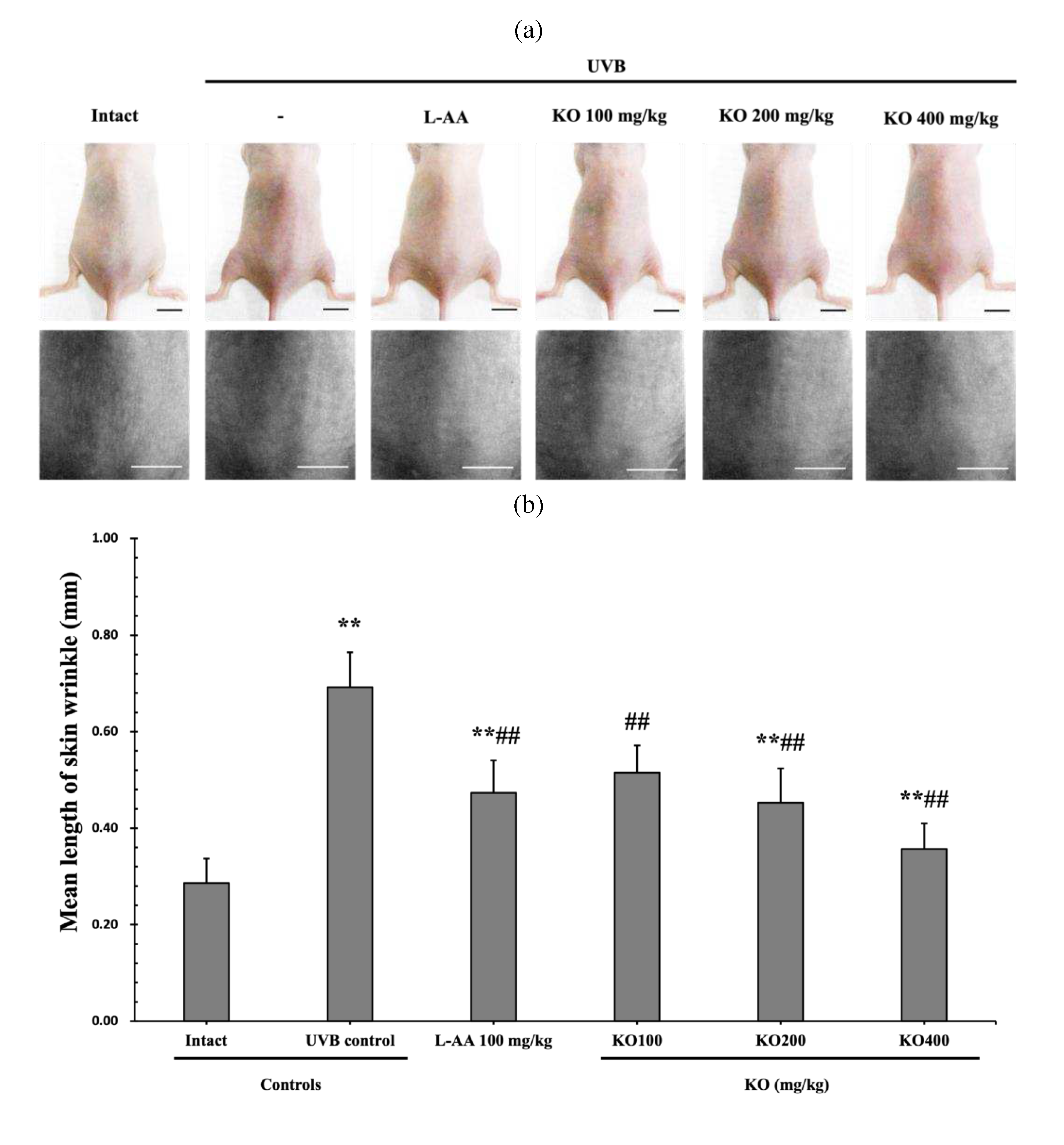
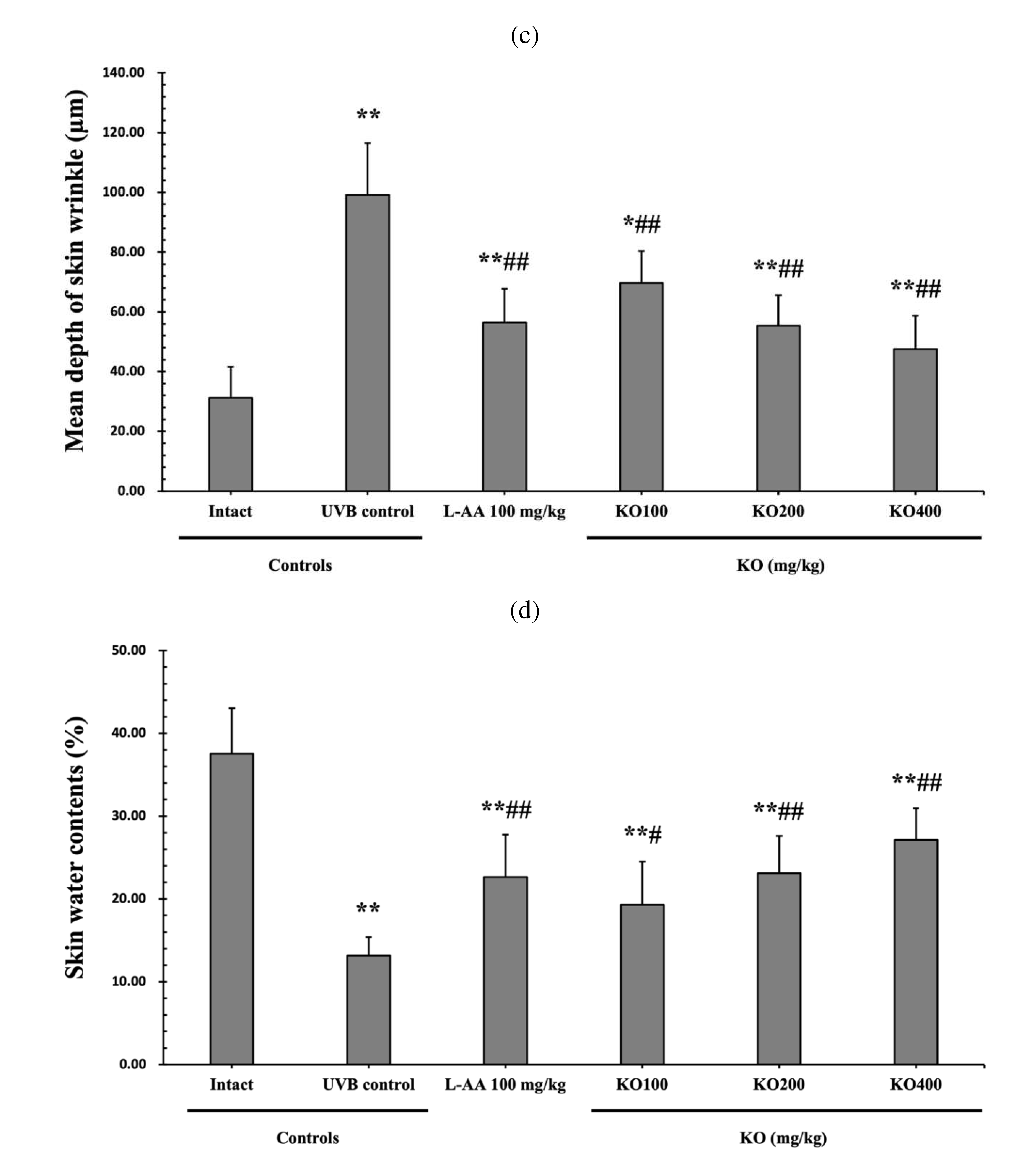
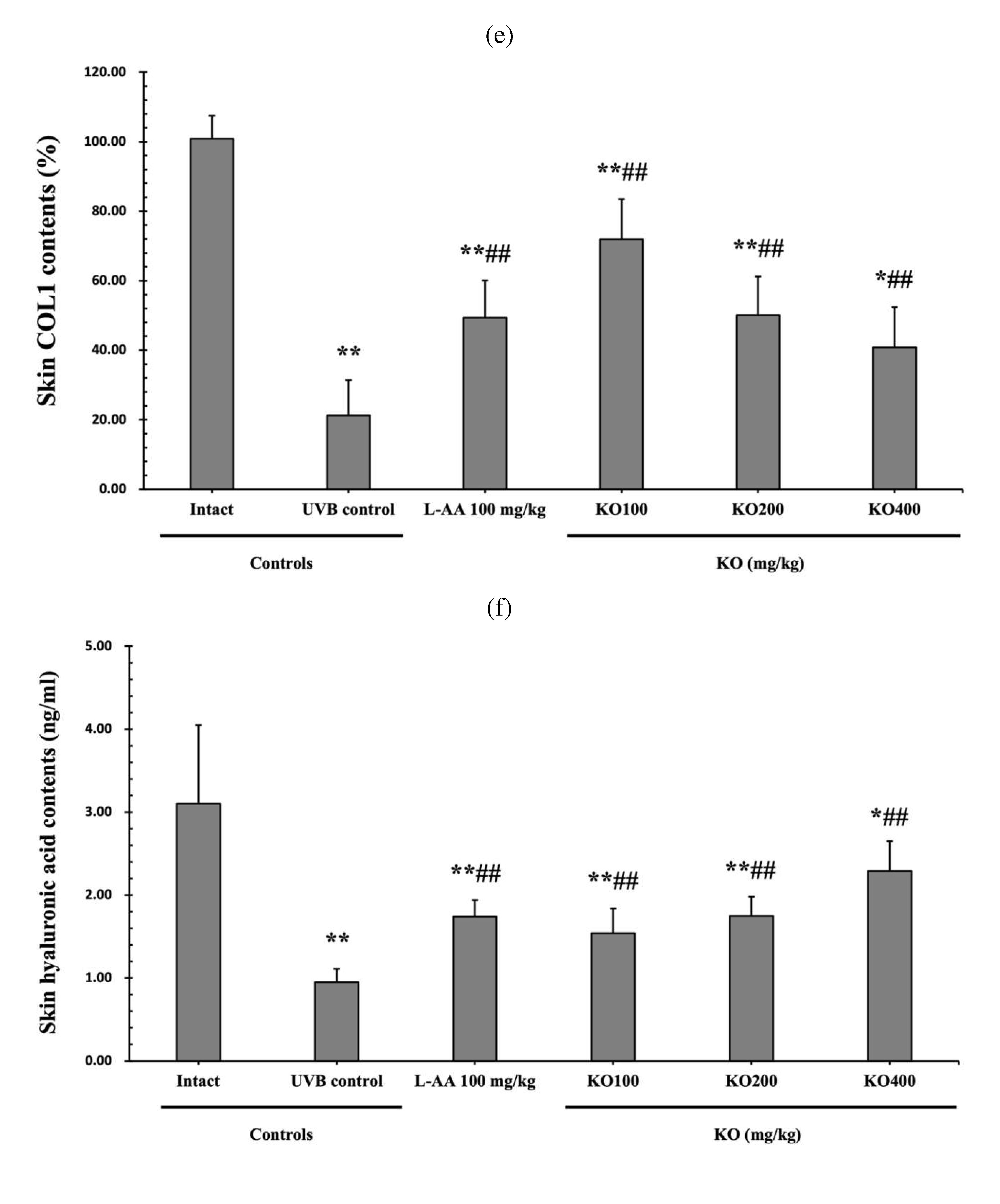
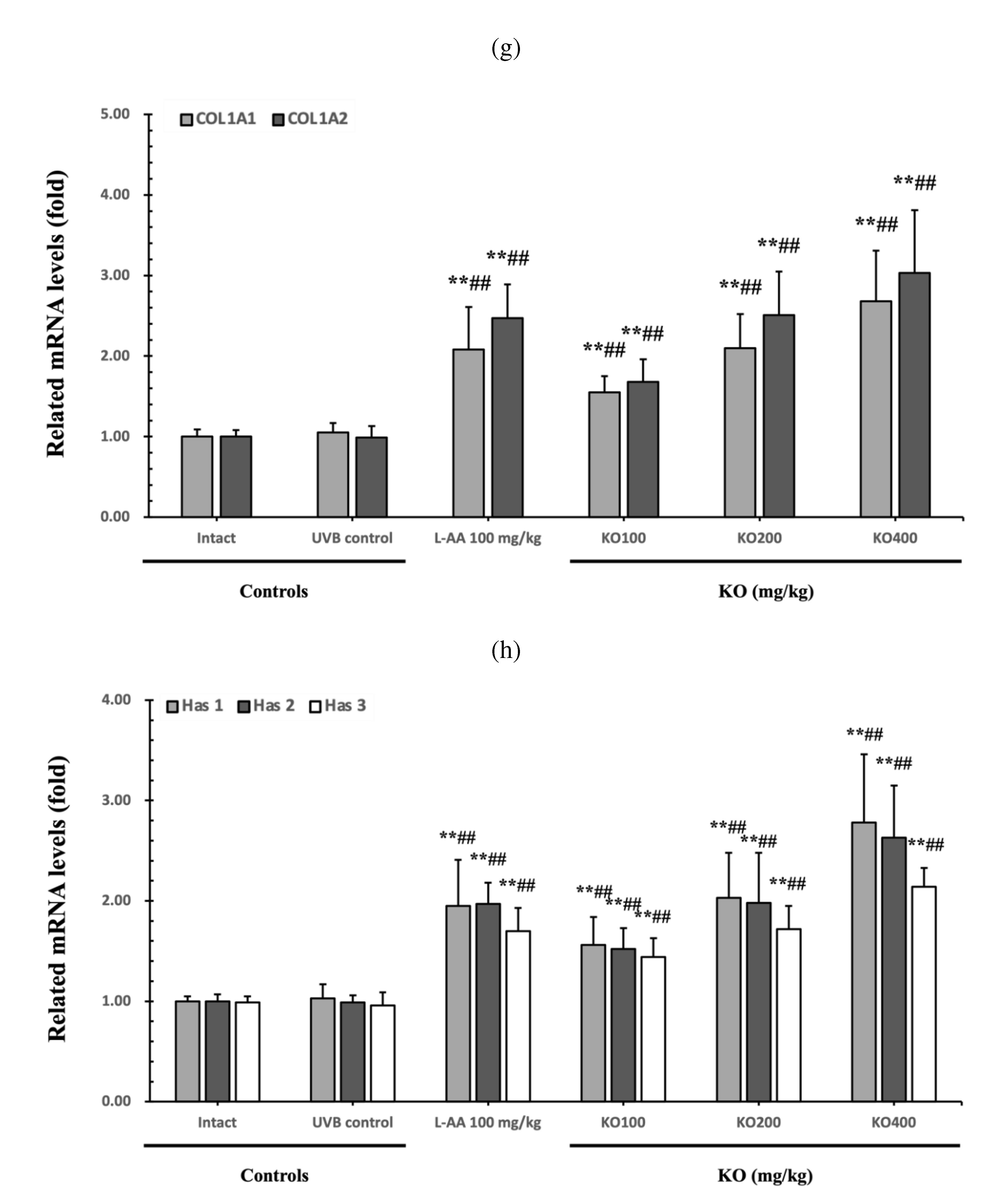
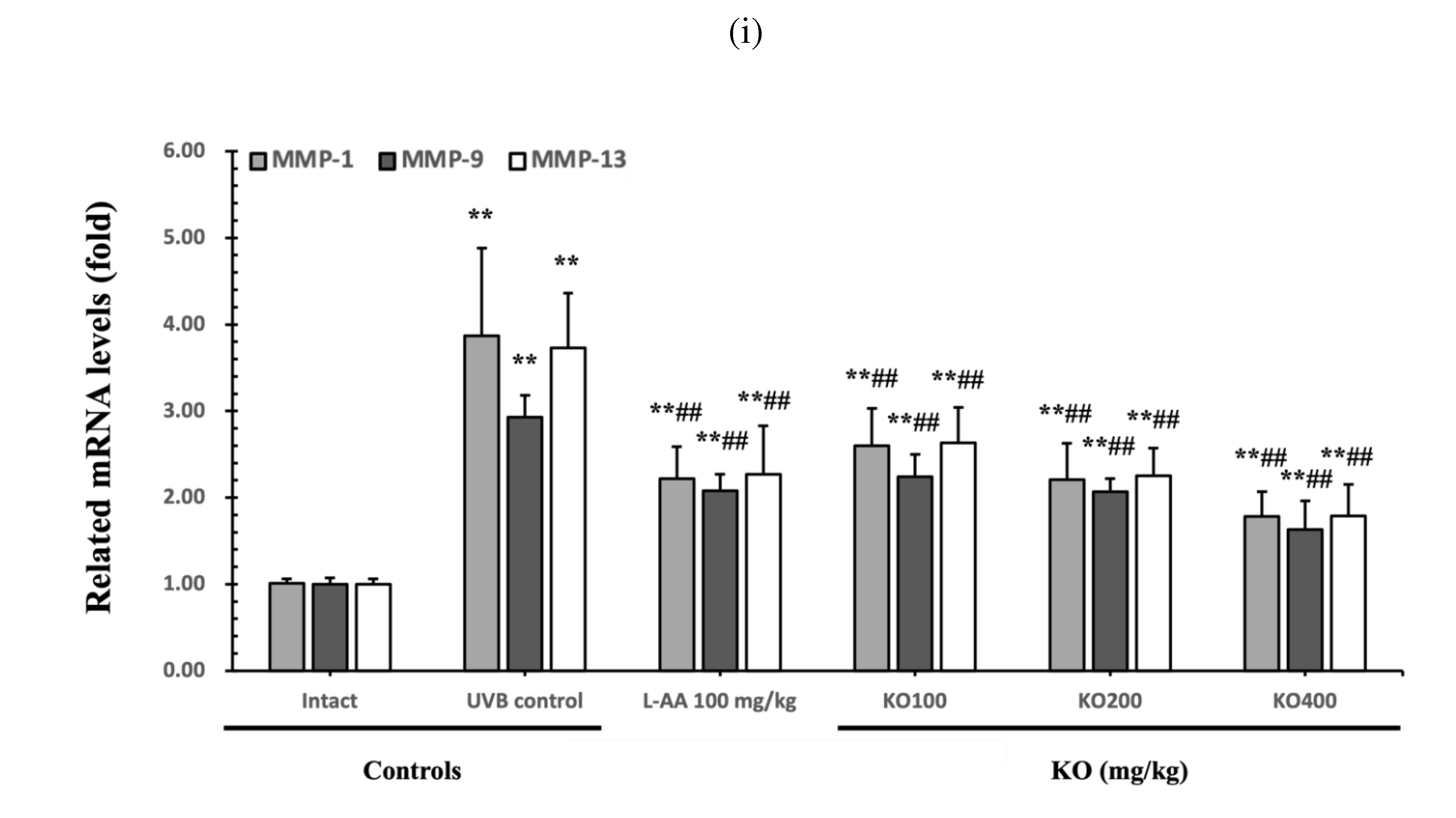
Figure 7.
Effects of KO on UVB-induced skin inflammation: (a) skin edema (weight of 6mm diameter skin sample); (b) Myeloperoxidase (MPO) activities for skin neutrophil content; (c) IL-1β and IL-10 levels in dorsal back skin tissue. Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
Figure 7.
Effects of KO on UVB-induced skin inflammation: (a) skin edema (weight of 6mm diameter skin sample); (b) Myeloperoxidase (MPO) activities for skin neutrophil content; (c) IL-1β and IL-10 levels in dorsal back skin tissue. Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
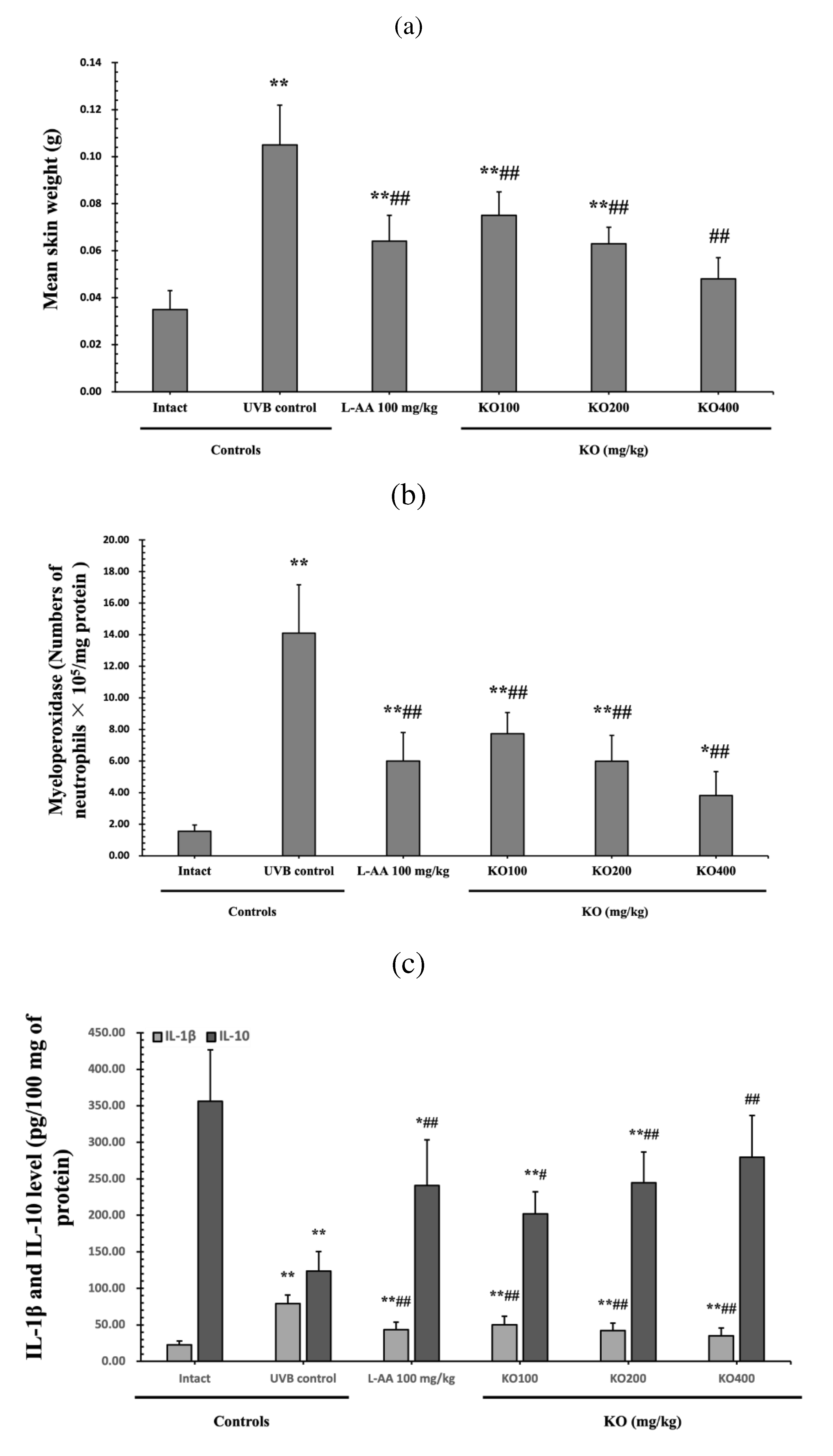
Figure 8.
Effects of KO on UVB-induced oxidative stress: (a) GSH contents in the skin tissue; (b) MDA level in the skin tissue; (c) superoxide anion production in the skin tissue; (d) GSH reductase mRNA expression level in the dorsal back skin tissue; (e) NOX2 mRNA expression level in the dorsal back skin tissue Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
Figure 8.
Effects of KO on UVB-induced oxidative stress: (a) GSH contents in the skin tissue; (b) MDA level in the skin tissue; (c) superoxide anion production in the skin tissue; (d) GSH reductase mRNA expression level in the dorsal back skin tissue; (e) NOX2 mRNA expression level in the dorsal back skin tissue Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
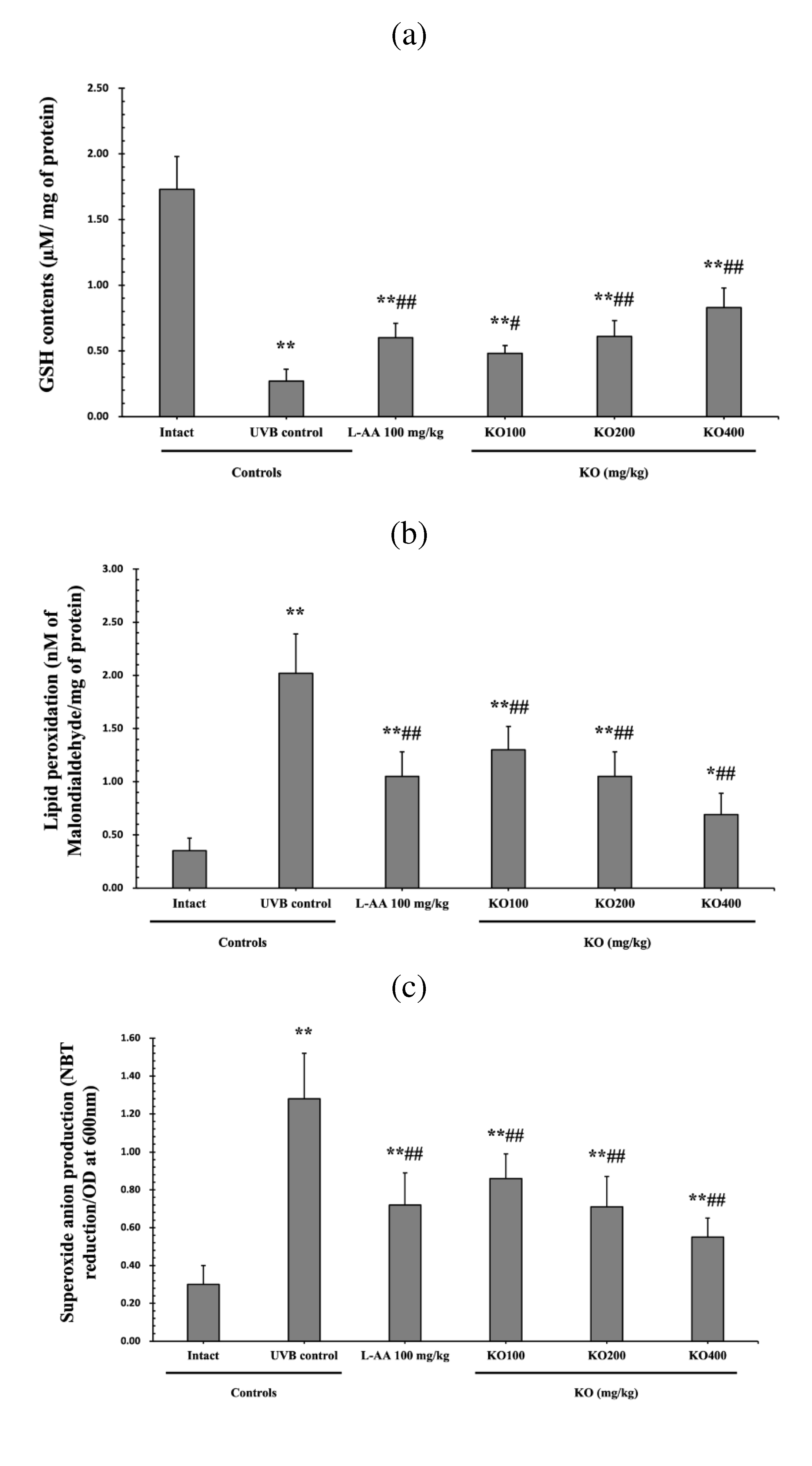
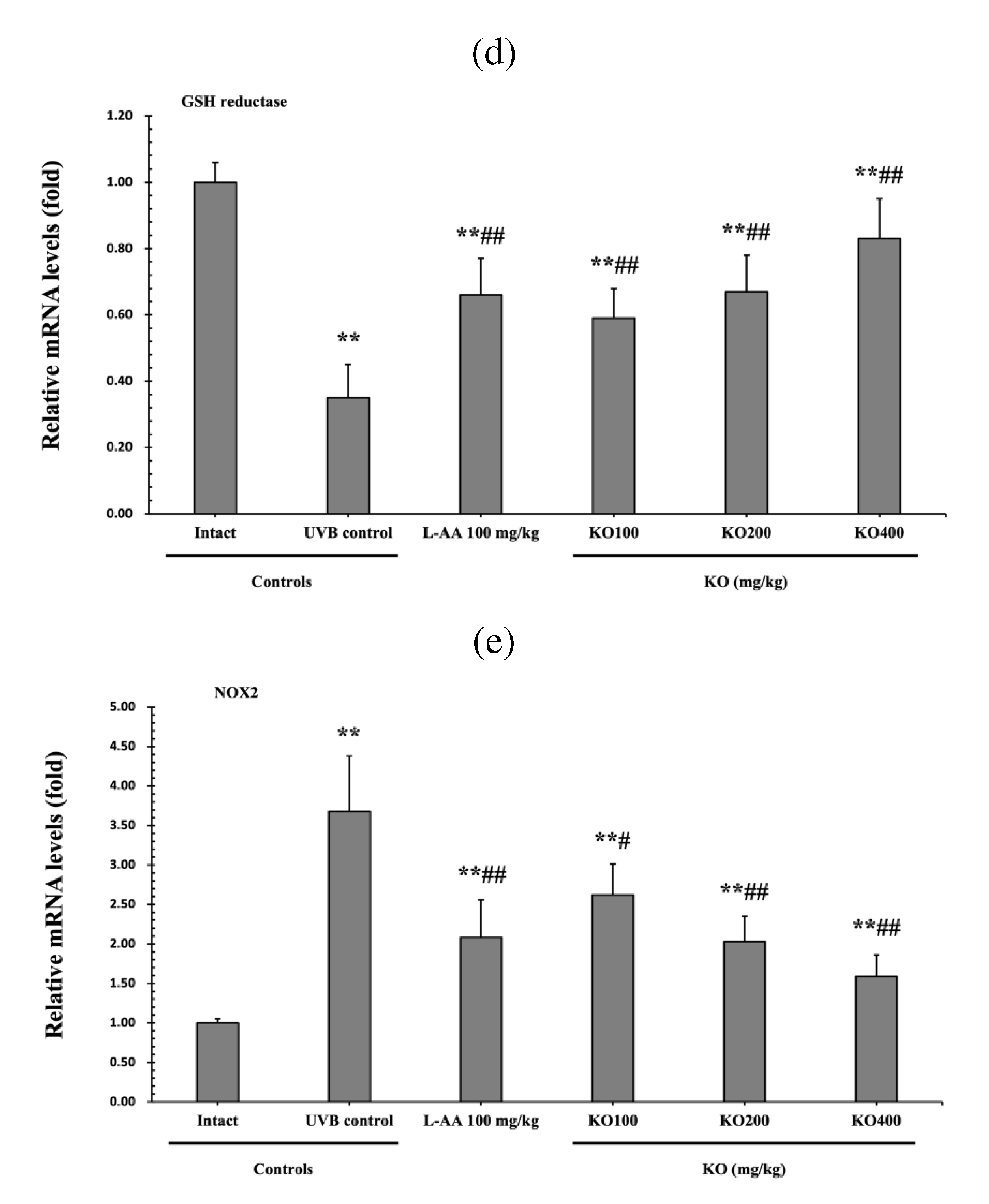
Figure 9.
(a) Representative images of stained skin with tissue with hematoxylin and eosin or Masson’s trichrome (MT). Arrows indicate microfolds in skin epithelial surface. Scale bars indicate 200 µm. (b) Immuno-stained skin tissue using nitrotyrosine (NT), 4-hydroxynonenal (4-HNE), cleaved caspase-3, cleaved PARP, and MMP9 antibodies. Scale bars indicate 100 µm. EP, epithelium; DE, dermis; CM, cutaneous muscle; SE, sebaceous gland; AC, adipocyte; Th, thickness.
Figure 9.
(a) Representative images of stained skin with tissue with hematoxylin and eosin or Masson’s trichrome (MT). Arrows indicate microfolds in skin epithelial surface. Scale bars indicate 200 µm. (b) Immuno-stained skin tissue using nitrotyrosine (NT), 4-hydroxynonenal (4-HNE), cleaved caspase-3, cleaved PARP, and MMP9 antibodies. Scale bars indicate 100 µm. EP, epithelium; DE, dermis; CM, cutaneous muscle; SE, sebaceous gland; AC, adipocyte; Th, thickness.
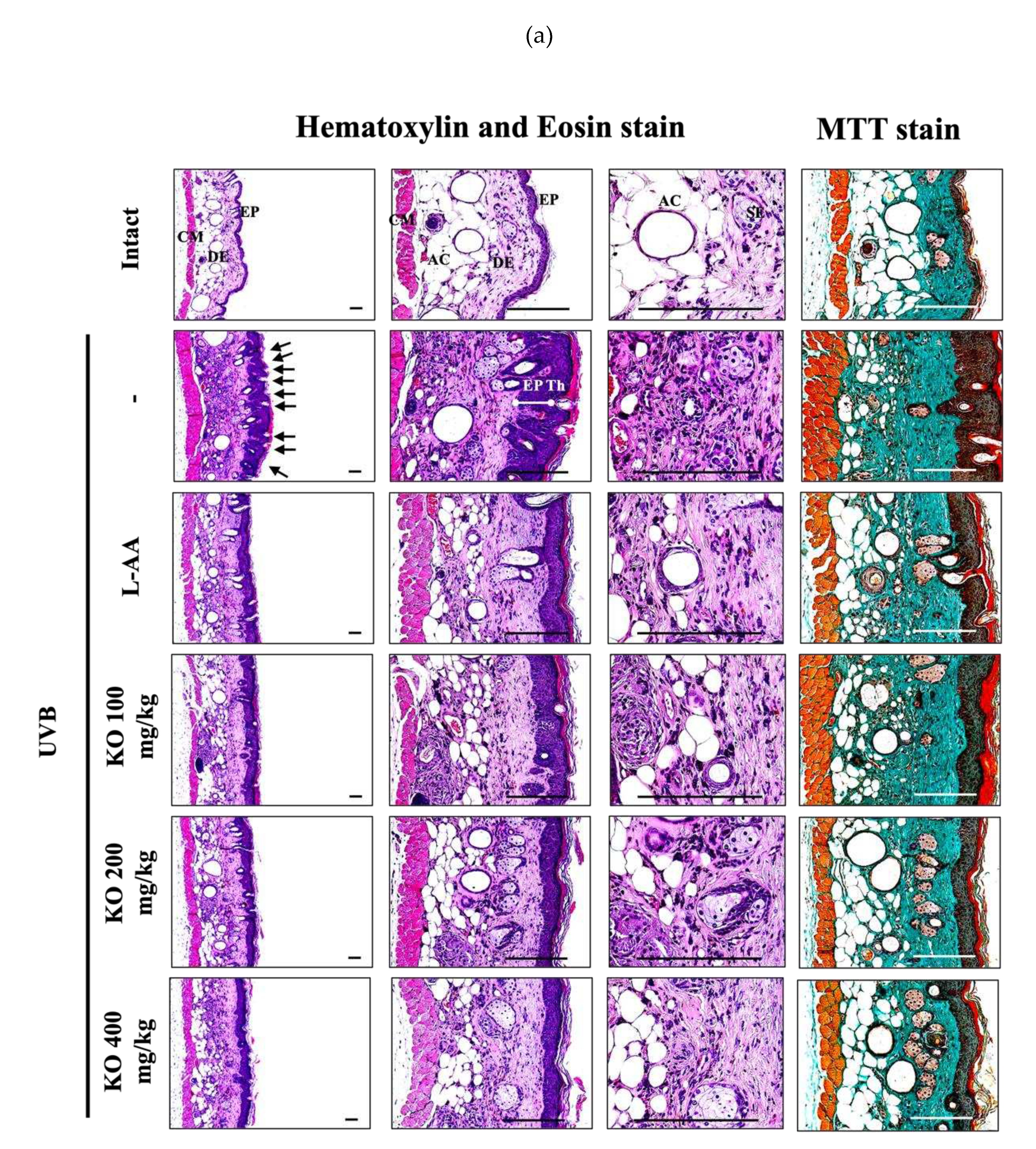
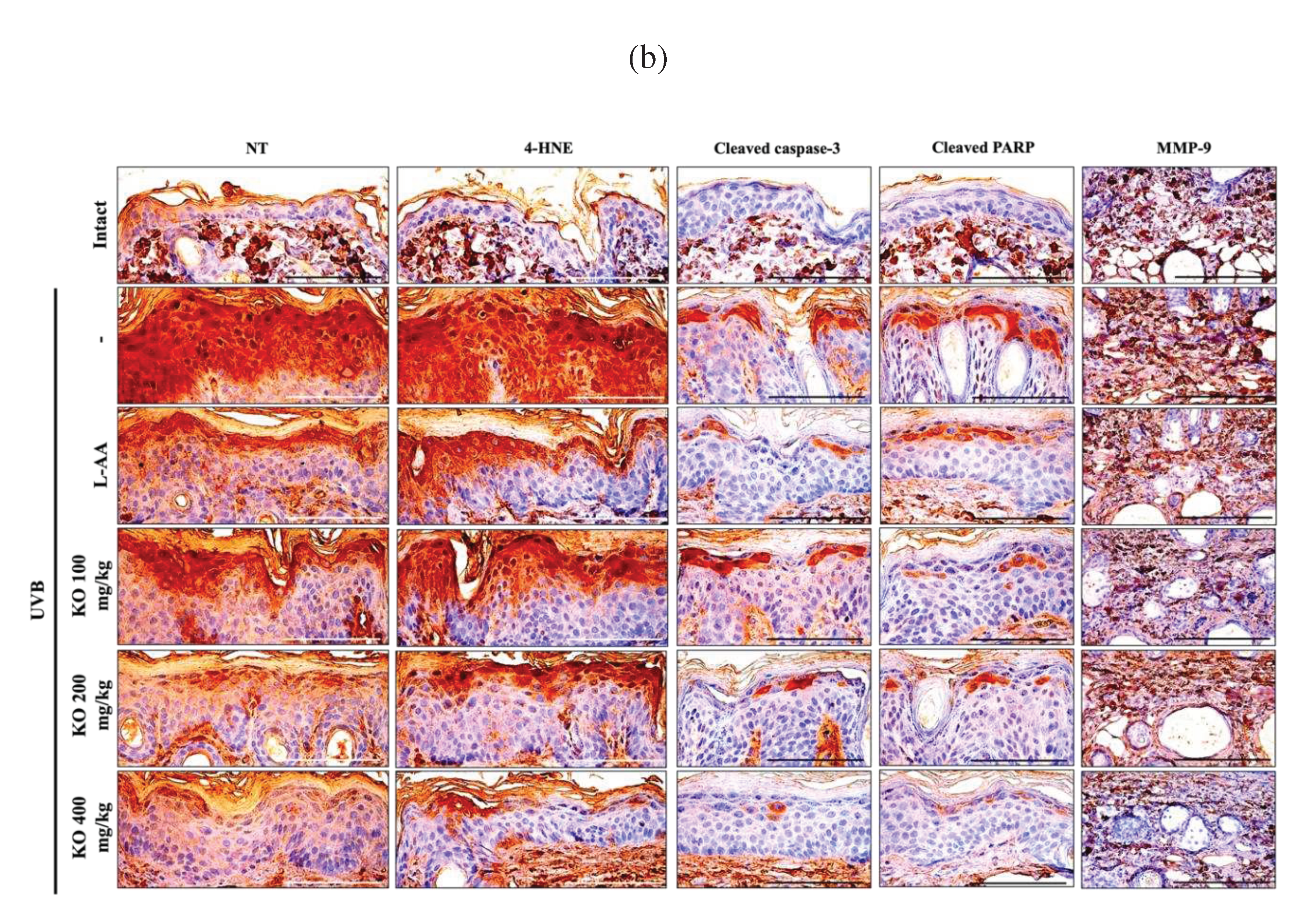

Table 1.
General histomorphometrical analysis of dorsal back skin, taken from unexposed intact or UVB-exposed hairless mice.
Table 1.
General histomorphometrical analysis of dorsal back skin, taken from unexposed intact or UVB-exposed hairless mice.
| Items (Unit) Groups |
Number of Microfolds (Folds/mm of Epidermis) |
Mean Epithelial Thickness (μm/Epidermis) | Mean Inflammatory Cells (Cells/mm2 of Dermis) |
Collagen Fiber Occupied Regions (%/mm2 of Dermis) |
|---|---|---|---|---|
| Controls | ||||
| Intact | 3.30±1.06 | 24.45±4.21 | 43.40±11.12 | 31.76±6.58 |
| UVB | 17.60±2.37** | 120.25±13.37** | 525.20±118.05** | 75.73±7.50** |
| Reference | ||||
| L-AA 100 mg/kg | 8.90±1.79**## | 65.94±10.15**## | 188.60±74.90**## | 51.47±8.96**## |
| Test materials | ||||
| KO 100 mg/kg | 12.10±1.60**## | 77.69±10.98**## | 242.80±67.05**## | 56.78±5.30**## |
| KO 200 mg/kg | 8.10±1.45**## | 65.03±11.28**## | 186.40±82.58**## | 51.43±7.23**## |
| KO 400 mg/kg | 6.10±1.29**## | 55.00±10.87**## | 134.20±38.05**## | 42.95±8.00*## |
significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01).
Table 2.
Immunohistomorphometrical analysis of dorsal back skin, taken from unexposed intact or UVB-exposed hairless mice.
Table 2.
Immunohistomorphometrical analysis of dorsal back skin, taken from unexposed intact or UVB-exposed hairless mice.
| Groups Items |
Controls | Reference | Test materials | |||
|---|---|---|---|---|---|---|
| Intact | UVB | L-AA 100 mg/kg | KO 100mg/kg | KO 200 mg/kg | KO 400 mg/kg | |
| Epidermis (cells/100 epithelial cells) |
||||||
| Nitrotyrosine | 18.20±4.47 | 80.20±7.80** | 41.40±10.98**## | 53.60±10.45**## | 40.80±11.52**## | 30.20±10.69## |
| 4-HNE | 14.00±2.49 | 86.20±4.94** | 52.00±6.86**## | 63.00±10.17*## | 51.20±10.92**## | 28.00±7.60**## |
| Cleaved caspase-3 | 4.60±1.90 | 36.60±4.22** | 18.40±5.40**## | 24.80±4.12**## | 18.00±5.25**## | 10.00±4.32## |
| Cleaved PARP | 4.60±1.90 | 41.00±5.68** | 22.00±3.77**## | 27.00±3.68**## | 21.00±3.92**## | 12.20±2.20**## |
| Dermis (%/mm2) | ||||||
| MMP-9 | 20.60±5.17 | 70.14±7.05** | 43.50±9.50**## | 53.95±8.33**## | 41.98±10.77**## | 32.75±9.27*## |
Data are presented as the mean ± SD (n=10, significance difference vs. intact control; *p<0.05, **p<0.01, vs. UVB-irradiated control mice; # p<0.05, ## p<0.01). 4-HNE, 4-Hydroxynonenal; PARP, Poly(ADP-ribose) polymerase; MMP, Matrix metalloprotease.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



