
Submitted:
09 August 2023
Posted:
10 August 2023
You are already at the latest version
Abstract
Beef meat production in Uruguay is based on pasture or concentrate feeding systems, leading to differences in meat quality. The objective of this study was to compare the oxidative and color stability during refrigerated retail display of beef meat from these two systems. For this, ten Aberdeen Angus steers were reared and fed on pasture and other ten were finished on a concentrate diet. After slaughter, the Semimembranosus muscle was cut into slices and put in a refrigerated showcase for 3, 6, and 9 days simulating commercial conditions. The β-carotene, α-tocopherol, and fatty acid content were determined before the display, and the lipid and protein oxidation, color parameters, and heme iron content were determined before and during the display. Meat from steers fed pasture had a lower content of intramuscular fat, monounsaturated fatty acids, n-6/n-3 ratio, lipid and protein oxidation, L* and a* values, and a higher content of linolenic acid, DHA, total n-3, β-carotene, and α-tocopherol. In conclusion, meat from pasture-fed steers was more stable during the retail display, which can probably be explained by a higher content of antioxidant compounds, such as β-carotene and α-tocopherol, and presented a healthier fatty acid profile.
Keywords:
lipid oxidation
; protein oxidation
; β-carotene
; α-tocopherol
; fatty acid
; heme iron
; beef meat
; pasture
; concentrate
1. Introduction
Oxidative processes are major non-microbiological factors involved in the quality deterioration of meat during refrigerated storage [1]. Indeed, oxidation induces modifications of muscle lipids and proteins and, therefore, affects the organoleptic and nutritional properties of meat and meat products. Oxidation of myoglobin and lipids reduces the color and flavor acceptability of fresh meat during refrigerated storage [2,3]. In fact, color is an important parameter for consumers, that has substantial influence on acceptability and purchasing decision at retail points [4], where a bright red color in beef is indicative of freshness [5]. When lipid peroxidation propagates in membranes, it promotes myoglobin oxidation resulting in color deterioration, formation of rancid odours and other off flavours in fresh meat [6]. This is why the beef industry concerns about product shelf life [7]. The stability of meat to oxidation is the result of the balance between prooxidants and antioxidants [1]. Transition metals, specifically iron, is considered a strong pro-oxidant specie [8]. Dietary antioxidants can be delivered to the muscle where, together with the endogenous defense systems, they counteract the action of prooxidants [9]. However, endogenous antioxidant concentrations depend on animal species, muscle type and diet [10,11,12]. As shown in other works [9,13,14], dietary treatments may directly affect the balance between antioxidant and prooxidant components in muscle.
Beef cattle production systems in Uruguay rely almost exclusively on grazed pastures. However, more recently intensive beef production systems have gained increased interest from some beef producers. These two systems produce meat with different antioxidant, prooxidant, and fatty acid content and composition [9,15,16]. Fresh herbage is rich in antioxidants such as tocopherols, carotenoids, ascorbic acid, and phenolic compounds [12], and grains contain polyphenols like proanthocyanidins and phytic acid [17]. Tocopherols and β-carotene constitute the main lipid-soluble free radical scavengers in meat. Tocopherols are a group of plant phenolic isomers deposited from dietary sources [18]. Several studies indicate that the antioxidant α-tocopherol delays myoglobin oxidation and extends color stability in retail beef [19,20,21]. β-carotene is another essential fat-soluble antioxidant that quenches sites localized within the hydrophobic region of biological membranes, contrasting with the scavenging activity of α-tocopherol close to the membrane surface [22].
Thus, the aim of this investigation was to study the oxidative and color stability of beef meat during refrigerated retail display simulating usual commercial conditions, and the α-tocopherol, β-carotene, and fatty acid content prior to the display, coming from animals produced in pastures and in concentrate finishing system.
2. Materials and methods
2.1. Animals and samples
Aberdeen Angus young steers (2–teeth) from 26–30 months of age, coming from two feeding systems prevalent in Uruguay were used for this study. All the steers came from commercial-run operations by the Breeders Society of Aberdeen Angus of Uruguay (SCAAU). Ten animals from one feeding system were randomly selected, with a final live weight average of 495.8 kg, which were reared under Uruguayan characteristic conditions based on the exploitation of natural resources with traditional extensive grazing. They grazed on natural pasture and were finished (100 days before slaughtering) on an improved pasture (40% natural and 60% cultivated grass) consisting in tall fescue (Festuca arundinacea), white clover (Trifolium repens) and birdsfoot trefoil (Lotus subbiflorus cv El Rincón). Another ten animals from the other feeding system were also randomly selected, with a final live weight average of 498.2 kg, which came from an intensive production farm. This farm exports HQB (high-quality beef) meat following the European Commission Regulation (Number 481/2012) with the last 100 days prior to slaughter finished with concentrate (1.4% body weight, 62% dry matter, 12.26 MJ/kg dry matter). The concentrate consisted of whole plant sorghum silage and wet grain sorghum and corn silage, sunflower pellets, mineral sources, urea, and ionophore. All animals were slaughtered the same day, in a commercial slaughterhouse named Frigorífico Breeders & Packers of Uruguay (BPU) located in the city of Durazno and classified following the rules for exportation. According to the official system of classification of INACUR-Uruguay, carcasses conformation and fatness were classified as A and 2, respectively, for both feeding systems [23]. Carcasses were kept refrigerated at 1-2 ºC for 36 hours postmortem and then the Semimembranosus muscle was sampled, vacuum packaged, and kept at 1-2 ºC. In the laboratory, muscle samples were cut into slices of 100 g and 1 cm thick and put in food-grade polyfoam trays and overwrapped with food-grade oxygen-permeable PVC (polyvinyl chloride) film (density 1.39 g/cm3; O2 permeability cm3 mm/m2 day atm). Trays with 3 slices of meat from each animal, were put in a commercial refrigerated showcase (CE, SS1500 model, 1.25 m height, 90 cm wide and 1.50 m long) at 2-8 ºC with artificial light (120 cm, 18 W, 2700 Kelvin and 1700 lm). After each display time was completed, 3, 6, and 9 days, a sample was tested for color and immediately vacuum packaged (105 µm, Lacor, LR69454, Spain) and frozen at -18 ºC until the other determinations were done (n=10 for each feeding system and display time). Before placing the meat samples (100 g and 1 cm thick, n=10) in the showcase (day 0), the color was tested and immediately frozen at -18 ºC until further determinations.
2.2. Determination of lipid oxidation
Lipid oxidation was determined in meat from day 0, and after 3, 6, and 9 days of display. Samples of 5 g frozen meat were homogenized in a Waring-Blender with 100 ml of extraction buffer (0.15 M KCl, 0.02 M EDTA, and 0.30 M BHT) at 12,000 rpm for 1 minute. Part of the homogenate was frozen for carbonyl and protein content assays, and part was used for the TBARS (thiobarbituric acid reactive species) test. The TBARS procedure for the determination of lipid oxidation was followed according to [24] and [25] as described in [15]. Results were expressed as mg malondialdehyde/kg of fresh meat.
2.3. Determination of protein oxidation
Protein oxidation was determined in meat from day 0, and after 3, 6, and 9 days of display, following the carbonyl protein assay according to [26] as described in [15]. Results were expressed as nmoles of dinitrophenylhydrazine/mg of protein. Protein content was determined at 280 nm in the extraction buffer using bovine serum albumin (BSA) from Sigma chemicals Co (St Louis, USA) as a protein standard, as described by [27].
2.4. Β-carotene and α-tocopherol content
Both compounds were extracted from meat from day 0 following [28], [18] for carotenoids, and [29] for tocopherol, with some modifications. Briefly, 0.5 g of frozen meat was homogenized with 11 ml of methanol: THF (1:1) in an Ultra-Turrax IKA T18 basic for 30 seconds at 12,000 rpm. Then, centrifugation was done at 12,000 rpm and 4 ºC for 1 minute. The supernatant was kept at -80 ºC and prior to measuring an aliquot of 1 ml was taken and evaporated using N2. The pellet was resuspended in 0.2 ml of the same extraction solution and 20 μl were injected in the HPLC (Thermo Separation Products, Spectral Series P100, Inc. USA) with detector UV2000. β-carotene content was measured at 450 nm and α-tocopherol at 290 nm, with a C30 thermostated column at 30 ºC. The mobile phase used was ethanol: methanol: THF (75:20:5) HPLC grade, flux 0.5 ml/min. Calibration curves were performed with β-carotene (Sigma Aldrich C9750) and DL-α-tocopherol (Sigma Aldrich T3251) standards. Results were expressed as μg/g fresh meat.
2.5. Determination of fat content and fatty acid composition
Total intramuscular fat was extracted from meat from day 0 following the chloroform: methanol (2:1; v:v) procedure of [30]. Extract containing approximately 40 mg of fat was methylated following the method of [31] and fatty acid analysis was performed by gas chromatography with a Clarus 500 (Perkin Elmer Instruments) split/splitless equipped with 100 meters CPSIL 88 capillary column. Hydrogen was the carrier gas (flow rate: 1 ml/min) and a FID detector was used. Fatty acids (FAMEs) were determined comparing the retention time to fatty acids standards. The atherogenic index was calculated as [C12:0+4*(C14:0) +C16:0]/[(ΣPUFA) + (ΣMUFA)] and the thrombogenic index as [C14:0+C16:0+C18:0]/[(0.5*ΣMUFA) + (0.5*n-6) + (3*n-3) + (n-3/n-6)].
2.6. Color measurement
Color parameters were measured in meat from day 0, and after 3, 6, and 9 days of display using a Minolta CR-10 colorimeter (Konica Minolta, Japan). The CIE L*a*b* system provides the values of three-color components: L* (0: black to 100: white component, lightness), a* (+ red to – green component), and b* (+ yellow to – blue component) [32]. All samples were allowed to bloom for 10 min before measuring.
2.7. Heme iron content
Total heme pigments in meat from day 0, and after 3, 6, and 9 days of display were determined as hemin after extraction with acidified acetone solution described by [33,34,35]. The heme iron content was calculated with the factor 0.0882 μg iron/μg hematin [33], and results were expressed as ppm. All samples were assayed in duplicate.
2.8. Statistical analysis
The data of TBARS, carbonyls, β-carotene, α-tocopherol, total fat content, fatty acid composition, color parameters, and heme iron content, were reported as mean ± standard error of the mean (SEM). To evaluate the feeding system and day of display effects, repeated measures ANOVA was followed. Samples from different feeding systems of the same day of display and samples from a different day of display in each feeding system were compared using a one-way ANOVA followed by the Tukey-Kramer multiple comparison test. The statistical analysis was done using the NCSS 2007 software (NCSS, 329 North 1000 East, Kaysville, UT 84037) and the level of significance was established at P < 0.05.
3. Results and discussion
3.1. Lipid and protein oxidation
Lipid and protein oxidation were lower (main effects, P < 0.0001 and P < 0.01, respectively) in meat from pasture-fed Aberdeen Angus steers, compared with meat from concentrate-fed Aberdeen Angus steers (Figure 1) in agreement with [4] in Psoas major steaks from crossbreed steers during 9 days in retail display. The same result was found for lipid oxidation by [36] in strip loins of Angus and Angus-Hereford cross steers between conventional grain finished beef and 25 months grass-fed beef, and by [37] in loin steaks from Tudanca bulls, both during 6 days in refrigerated retail display, and by [8] in Longissimus thoracis muscle from British and Zebu cross steers during 13 days of refrigerated storage. All these results observed are indicating that beef meat coming from pasture systems has a greater oxidative stability during refrigerated retail display conditions than beef meat coming from concentrate feeding systems.
The lower level of lipid and protein oxidation found in meat from pasture-fed steers could be due to a higher content of α-tocopherol and β-carotene (Table 1) in this meat before the refrigerated display. It is known that α-tocopherol and β-carotene act as antioxidant protection in meat, as observed in previous investigations [38,39]. Vitamin E (α-tocopherol) protects cells against the effects of free radicals and may also block the formation of nitrosamines [19]. Carotenes can quench singlet oxygen and scavenge toxic free radicals preventing or reducing damage to living cells. Carotenoids react chemically with the free radicals and the system of conjugated double bonds is directly destroyed. Due to their long-conjugated chains, carotenoids are highly reactive [40].
When the display day effect was considered, an increment in both lipid and protein oxidation (P < 0.0001) with the time of the display was found. This is in accordance with other research [4,41,42,43], and could be explained by the presence of oxygen in the air and the artificial light of the showcase, which favor oxidative processes [44]. However, despite this increment of lipid oxidation during the time of display, the maximum TBARS values reached in meat from both feeding systems, are lower than those proposed as a threshold of acceptability by consumers of 2 mg MDA/kg meat [45]. When the evolution of the lipid oxidation was analyzed day by day (Figure 1a), a significant increment in TBARS can be seen at the beginning of the display (day 0 vs day 3) in meat from concentrate-fed animals, and at day 6 and day 9 in meat from pasture-fed animals. In addition, the carbonyl content (Figure 1b) increased significantly from day 3 until day 6, and from day 6 until day 9 of the display in meat from concentrate-fed animals, and from day 6 until day 9 in meat from pasture-fed ones. These results show that oxidation processes started to increase earlier in meat from concentrate-fed Aberdeen Angus steers than in pasture-fed ones, during the refrigerated retail display conducted in the present investigation. A similar pattern was reported by [37] in loin steaks from Tudanca bulls during 6 days in refrigerated retail display and by [4] in Psoas major steaks from crossbreed steers during 9 days in refrigerated retail display. In the present investigation, these results could be due to a higher antioxidant protection from α-tocopherol and β-carotene in meat from pasture-fed animals, which can be delaying the oxidation processes in this meat since no significant differences in polyunsaturated fatty acids (PUFA) content were found between meat from both systems. It is relevant to mention that the lipids that contain PUFA are particularly susceptible to free radical attack.
3.2. β-carotene and α-tocopherol content
As observed in Table 1, β-carotene content measured before the display in meat from pasture-fed Aberdeen Angus steers was 6 times higher (1.73 ± 0.18 μg/g fresh meat) than in meat from concentrate-fed ones (0.27 ± 0.04 μg/g fresh meat) (P < 0.0001).
Other authors also found higher levels of β-carotene in animals fed pasture compared with those fed grains [4,46,47,48]. This occurs because β-carotene concentrations are higher in pastures, legumes, and other green plants, while seeds and whole plant silage have a low content of vitamins [49]. Regarding the α-tocopherol content, also measured before the display, it was almost 2-fold greater (P < 0.05) in meat from animals fed pasture (3.7 ± 0.5 μg/g fresh meat) than in meat from animals fed grains (1.9 ± 0.3 μg/g fresh meat) (Table 1). This result is supported by numerous publications [4,8,18,37,46,48,50,51] and can be explained by the greater concentration of α-tocopherol in green leaves tissues compared with grains [26]. As was mentioned by [52], desirable levels of α-tocopherol in muscle must be above 3.5 μg/g, in order to minimize lipid oxidation processes in meat. Therefore, this threshold level was exceded in meat from pasture-fed steers but was not reached in meat from concentrate-fed steers. This fact probably led to higher levels of lipid and protein oxidation in concentrate-fed beef.
3.3. Lipid content and fatty acid composition
Meat from Aberdeen Angus steers fed pasture presented a lower intramuscular fat content (Table 2) than meat from Aberdeen Angus steers fed concentrate (P < 0.0001), which is in accordance with several publications [15,16,37,46,48,53,54,55]. This result can be explained by a lower intramuscular fat deposition in animals in an extensive system based on a pasture diet and greater surface extensions where they can move freely and are not confined in a limited area like animals in feedlot system receiving a concentrate diet [19].
Regarding the fatty acid composition measured in meat before the display, no differences between feeding systems were found in the even chain saturated fatty acids (SFA) 12:0, 14:0, 16:0, and 18:0, in the MUFA 14:1, 16:1, 17:1, 18:1, and 20:1, and in the PUFA n-6 18:2 and 20:4, and n-3 EPA, and DPA content (Table 2). CLA content was also not significantly different between systems. A higher content of linolenic acid (C18:3), C20:3 and DHA, which are PUFA n-3, was observed in meat from pasture-fed animals compared with concentrate-fed ones (Table 2). Particularly, linolenic acid content was 3.4-fold and DHA 5-fold greater. This is a positive health aspect for meat consumers, as both fatty acids have anti thrombogenic and can prevent cardiovascular diseases [56]. Similar results regarding n-3 content in intramuscular fat of beef meat were found in another national [15,16,51] and international [57] research. Regarding total SFA, total PUFA, and total n-6 content, as well as PUFA/SFA ratio, no differences between feeding systems were observed (Table 2). Despite this, in general, higher PUFA concentrations were observed in pasture-fed animals compared to grain-finished ones [46,50,51,58,59]. Concerning PUFA/SFA ratio in meat from both feeding systems found in the present study (0.10 ± 0.02 in pasture and 0.09 ± 0.02 in concentrate), they were both under recommendations (0.45) from [60]. The PUFA/SFA ratio is nowadays recommended to be above 0.4-0.5 to prevent an excess of SFA, which have a negative effect on the level of plasmatic LDL cholesterol, and an excess of PUFA, which some of them are precursors of clotting agents and are involved in the etiology of some cancers [61]. Besides, total MUFA content was lower in meat from pasture system (P < 0.05) (Table 2), which is in accordance with other publications [7,46,51,58,62,63]. Furthermore, n-3 content was higher (P < 0.001) and the n-6/n-3 rate was lower (P < 0.05) in meat from pasture (Table 2), which is desirable because the balance between n-6 and n-3 PUFA is an important determinant in decreasing the risk for cardiovascular diseases and in the prevention of atherosclerosis [64]. This last result can be explained as corn grains of concentrate diet contain more n-6 than n-3 fatty acids, and in general, pasture-fed animals tend to accumulate more n-3 fatty acids [19,37] that are provided from pastures. These results were also observed in other research [46,59,65,66]. Indeed, the value observed for n-6/n-3 ratio in meat from steers fed concentrate (11.18 ± 3.96), was higher than recommendations (4/1 or 5/1) from [60] and exceeds the value of 10 which is not recommended for consumer´s health. Regarding the atherogenic and thrombogenic indexes, no significant differences were observed between feeding systems (Table 2). Concerning lipid content and fatty acid composition results, we can conclude that meat from Aberdeen Angus steers fed pasture is apparently more beneficial for consumer´s health than meat from animals in a concentrate system.
3.4. Color
When evaluating the main effects for the color parameters, a feeding system effect was found for L* (P < 0.01) and a* (P < 0.05), where meat from animals fed pasture showed lower values (Figure 2). This translates into a darker and less red meat compared with a concentrate-produced meat.
Similar results were found in other research [4,36,48,67,68,69,70], where beef meat from animals finished on pasture was darker (<L*) compared with those finished with grains. This darker color was explained by [68] by a higher myoglobin content in animals fed pasture. In our case, this cannot be the explanation because no differences between systems in heme iron content were observed (Table 3). Another hypothesis is that animals produced in an extensive system perform a higher physical activity compared with animals in a confined system [71]. Physical exercise elevates reactive oxygen species (ROS) concentrations in vivo, causing oxidative damage to tissues, including muscle. During the conversion of muscle into meat, the biochemical events favor oxidation, and intermediate free radicals that are generated, are similar to ROS and contribute to oxymyoglobin oxidation, causing loss of meat color [72]. Another hypothesis is that animals produced on pastures are older than those in the concentrate system, which is reflected in a darker meat, but in the present study, age difference between systems was of 30 days to reach a similar pre-slaughter weight. The greater redness observed in meat from animals concentrate-fed was opposite to what was found by [8] in Longissimus thoracis muscle from British and Zebu cross steers at day 10 and 13 of refrigerated storage, [4] in Psoas major steaks from crossbreed steers at 7 days of retail display, and [73] in Longissimus dorsi muscle of steers during the 10 days of the display. However, [74] and [51], did not found a feeding system effect (pasture vs grain) in a* parameter in the Longissimus dorsi muscle of steers. In the present study, no feeding system effect was found for the b* parameter (Figure 2(c)), which is in accordance with [51] in Longissimus dorsi muscle of Hereford steers, [75] in beef Longissimus dorsi muscle and [4] in Psoas major of steers. However, in the work of [8] steaks from steers fed pasture were significantly yellower when compared to grain steaks.
When the display day effect was evaluated, an effect was found for the three parameters (P < 0.0001, in all cases) (Figure 2). The L* value was higher on day 9 compared with the other days of the display, so meat becomes brighter with time (Figure 2a). In meat from steers fed pasture, L* values were relatively constant, only a significant increase was observed from day 3 to day 6. In meat from steers fed concentrate L* values decreased from day 0 until day 6 and then increased until day 9. Consequently, lightness is more stable in Semimembranosus muscle from Aberdeen Angus produced in pasture, during refrigerated display. Besides, a fall in a* value was observed (Figure 2b) in meat from both systems, so meat loses redness with the time of display which indicates myoglobin oxidation. This decline in redness was also observed in several publications [4,8,41,43,72,74,76] in beef meat in retail display conditions. The b* value was higher at the beginning and at the end of the display (Figure 2-c). In [74] b* values remain stable during the first 2 days and decreased after 6 days of storage, and in [4] this decrease was observed on day 6 of the display, but values stayed still until day 9, in animals from pasture and concentrate systems. In addition, a decrease in b value was also observed in meat with increasing days of storage by [8].
3.5. Heme iron content
The heme iron content results did not show any statistical difference between feeding systems (pasture vs concentrate) (Table 3).
Similar findings have been reported by [74] and [4] in Longissimus dorsi and Psoas major muscles in Charolais breed and crossbreed (British x Indicus), respectively. Even though a decrease in heme iron content in meat was expected during the 9 days of display in a refrigerated showcase, because meat loses fluids and consequently heme iron and myoglobin [76], heme iron content did not show differences between the days of the display (Table 3).
4. Conclusions
When meat was submitted to a process of refrigerated retail display simulating commercial conditions, the one that came from pasture-fed Aberdeen Angus steers was more stable, from the oxidative and color point of views, and the oxidation processes started afterwards than in meat from concentrate-fed steers, allowing it to achieve a longer shelf-life. This fact was true for lipid oxidation, even though meat from pasture-fed animals presented a higher content of two fatty acids very sensitive to oxidation, such as α-linolenic and DHA. This can probably be explained by the higher content of antioxidant compounds found in meat from pasture-fed steers, such as β-carotene and α-tocopherol, both determined in the present investigation. Besides, pasture feeding may improve the beef lipid profile and make it healthier for meat consumers, preventing the occurrence of cardiovascular diseases.
Author Contributions
Conceptualization, A.T., M.C.C., F.Z. and A.S.; methodology, A.T., M.C.C., F.Z. and A.S.; validation, A.T., M.C.C., F.Z. and A.S.; formal analysis, A.T., M.C.C., F.Z. and A.S.; investigation, A.T., M.C.C., F.Z. and A.S.; resources, A.T., M.C.C. and A.S.; writing—original draft preparation, A.T.; writing—review and editing, M.C.C. and A.S.; visualization, A.T.; supervision, M.C.C. and A.S.; project administration, M.C.C. and A.S.; funding acquisition, M.C.C. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We would like to thank the Breeders Society of Aberdeen Angus of Uruguay (SCAAU) and Frigorífico Breeders & Packers of Uruguay (BPU). Also, CSIC for the PhD scholarship given to Alejandra Terevinto.
Conflicts of Interest
the authors declare no conflict of interest.
References
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M.A. Comprehensive Review on Lipid Oxidation in Meat and Meat Products. Antiox. (Basel). 2019, 8(10), 429. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.I.; Tejeda, J.F.; Parra, V.; Andrés, A.I. Food Tech Evolution of the fatty acid composition and oxidative stability of Merino lamb meat stored under different modified atmospheres. Irish J. Agric. Food Res. 2013, 52, 81–92. [Google Scholar]
- Pereira, A.L.F.; Abreu, V.K.G. Lipid peroxidation in meat and meat products. In Lipid Peroxidation Research, IntechOpen Ed., 2018; pp. 1-15. https://www.intechopen.com/chapters/63958#:~:text=DOI%3A%2010.5772/intechopen. 8153. [Google Scholar]
- Insani, E.M.; Eyherabide, A.; Grigioni, G.; Sancho, A.M.; Pensel, N.A.; Descalzo, A.M. Oxidative stability and its relationship with natural antioxidants during refrigerated display of beef produced in Argentina. Meat Sci. 2008, 79, 444–452. [Google Scholar] [CrossRef]
- Ripoll, G.; Panea, B.; Albertí, P. Apreciación visual de la carne bovina y su relación con el espacio de color CIELab. Inf. Téc. Econ. Agr. 2012, 108, 222–232. [Google Scholar]
- Gorelik, S.; Kanner, J. Oxymyoglobin oxidation and membranal lipid peroxidation initiated by iron redox cycle. J. Agric. Food Chem. 2001, 49(12), 5939–5944. [Google Scholar] [CrossRef]
- Realini, C.E.; Duckett, S.K.; Windham, W.R. Effect of vitamin C addition to ground beef from grass-fed or grain-fed sources on color and lipid stability, and prediction of fatty acid composition by near-infrared reflectance analysis. Meat Sci. 2004, 68, 35–43. [Google Scholar] [CrossRef]
- Fruet, A.P.B.; De Mello, A.; Trombetta, F.; Stefanello, F.S.; Speroni, C.S.; De Vargas, D.P.; De Souza, A.N.M.; Rosado Júnior, A.G.; Tonetto, C.J.; Nörnberg, J.L. Oxidative stability of beef from steers finished exclusively with concentrate, supplemented, or on legume-grass pasture. Meat Sci. 2018, 145, 121–126. [Google Scholar] [CrossRef]
- Descalzo, A.M.; Sancho, A.M. A review of natural antioxidants and their effects on oxidative status, odor and quality of fresh beef produced in Argentina. Meat Sci. 2008, 79, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Livisay, S.A.; Zhou, S. Mechanisms of endogenous skeletal muscle antioxidants: chemical and physical aspects. In Antioxidants in Muscle Foods: Nutritional Strategies to Improve Quality; Decker, E.A., Faustman, C., Lopez-Bote, C.J., Eds.; John Wiley & Sons: New York, USA, 2000; pp. 25–60. [Google Scholar]
- Gregory, N.G. Animal Welfare and Meat Science; CABI Publishing: Oxon, UK, 1998. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M. Factors influencing fatty acids in meat and the role of antioxidants in improving meat quality. Br. J. Nutr. 1997, 78, S49–S60. [Google Scholar] [CrossRef] [PubMed]
- Luciano, G.; Moloney, A.P.; Priolo, A.; Rohrle, F.T.; Vasta, V.; Biondi, L.; López-Andrés, P.; Grasso, S.; Monahan, F. J. Vitamin E and polyunsaturated fatty acids in bovine muscle and the oxidative stability of beef from cattle receiving grass or concentrate-based rations. J. Anim Sci. 2011, 89(11), 3759–3768. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; McDonagh, M.B.; Jacobs, J.L.; Hopkins, D.L. Relationship between muscle antioxidant status, forms of iron, polyunsaturated fatty acids and functionality (retail colour) of meat in lambs. Meat Sci. 2012, 90, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Terevinto, A.; Cabrera, M.C.; Saadoun, A. Oxidative stability, fatty acid composition and health lipid indices of Longissimus dorsi muscle from Aberdeen Angus steers produced in different feeding systems. Ciênc. Rural. [CrossRef]
- Terevinto, A.; Saadoun, A.; Cabrera, M.C. From the fatty acid content perspective, is it healthier to eat a hindquarter or a forequarter cut? Angus steers in pasture or concentrate systems. CyTA – J. Food. [CrossRef]
- Jood, S.; Kapoor, A.C.; Singh, R. Polyphenol and phytic acid contents of cereal grains as affected by insect infestation. J. Agric. Food Chem. 1995, 43, 435–438. [Google Scholar] [CrossRef]
- Descalzo, A.M.; Rossetti, L.; Sancho, A.M.; García, P.T.; Biolatto, A.; Carduza, F.; Grigioni, G.M. Antioxidant consumption and development of oxidation during ageing of buffalo meat produced in Argentina. Meat Sci. 2008, 79, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Daley, C.A.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 10. [Google Scholar] [CrossRef]
- Faustman, C.; Chan, W.K.M.; Lynch, M.P.; Joo, S.T. Strategies for increasing oxidative stability of (fresh) meat color. In Proceedings of the 49th Annual Reciprocal Meat Conference, Kansas City, USA, 9-11 June 1996; pp. 73–78. [Google Scholar]
- Nassu, R.T.; Dugan, M.E.R.; Juárez, M.; Basarab, J.A.; Baron, V.S.; Aalhus, J.L. Effect of α-tocopherol tissue levels on beef quality. Animal. 2011, 5(12), 2010–2018. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, K. Dynamics of lipid peroxidation and antioxidion of α-tocopherol in membranes. J. Nutr. Sci. Vitaminol. 2008, 54(4), 273–285. [Google Scholar] [CrossRef]
- 23. INAC; Instituto Nacional de Carnes. Manual de cortes bovinos para abasto, /: 2008. pp. 1-107. Available at: https, 2008.
- Lynch, S.M.; Frei, B. Mechanisms of copper- and iron-dependent oxidative modification of human low-density lipoprotein. J. Lipid Res. 1993, 34, 1745–1751. [Google Scholar] [CrossRef] [PubMed]
- Gatellier, P.; Mercier, Y.; Renerre, M. Effect of diet finishing mode (pasture or mixed diet) on antioxidant status of Charolais bovine meat. Meat Sci. 2004, 67, 385–394. [Google Scholar] [CrossRef]
- Mercier, Y.; Gatellier, P.; Renerre, M. Lipid and protein oxidation in vitro, and antioxidant potential in meat from Charolais cows finished on pasture or mixed diet. Meat Sci. 2004, 66, 467–473. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of Protein. Meth. Enzymol. 1990, 182, 50–68. [Google Scholar] [CrossRef]
- Zaccari, F.; Cabrera, M.C.; Ramos, A.; Saadoun, A. In vitro bioaccessibility of β-carotene, Ca, Mg and Zn in landrace carrots (Daucus carota, L.). Food Chem. 2015, 166, 365–371. [Google Scholar] [CrossRef]
- Koprivnjak, J.F.; Lum, K.R.; Sisak, M.M.; Saborowski, R. Determination of α-, γ (+β)-, and δ-tocopherols in a variety of liver tissues by reverse-phase high pressure liquid chromatography. Comp. Biochem. Physiol. 1996, 113, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipid from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ichihara, K.; Shibahara, A.; Yamamoto, K.; Nakayama, T. An improved method for rapid analysis of the fatty acids of glycerolipids. Lipids. 1996, 31(5), 535–539. [Google Scholar] [CrossRef] [PubMed]
- 32. CIE. Commisssion Internationale de L`Eleclerige, 2: Colorimetry, 15, 2004.
- Hornsey, H.C. The colour of cooked cured pork. I.-Estimation of the nitric oxide-haem pigments. J. Sci. Food Agric. 1956, 7, 534–540. [Google Scholar] [CrossRef]
- Ramos, A.; Cabrera, M.C.; del Puerto, M.; Saadoun, A. Minerals, haem iron and non-haem iron contents of rhea meat. Meat Sci. 2009, 81, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.; Cabrera, M.C.; Saadoun, A. Bioaccessibility of Se, Cu, Zn, Mn and Fe, and haem iron content in unaged and aged meat of Hereford and Braford steers fed pasture. Meat Sci. 2012, 91, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Duarte, T.L.; Bolkenov, B.; Klopatek, S.C.; Oltjen, J.W.; King, D.A.; Shackelford, S.D.; Wheeler, T.L.; Yang, X. Evaluating the shelf life and sensory properties of beef steaks from cattle raised on different grass feeding systems in the Western United States. Foods. 2022, 11(14), 2141. [Google Scholar] [CrossRef]
- Humada, M.J.; Sañudo, C.; Serrano, E. Chemical composition, vitamin E content, lipid oxidation, color and cooking losses in meat from Tudanca bulls finished on semi-extensive or intensive systems and slaughtered at 12 or 14 months. Meat Sci. 2014, 96, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.N.; Arp, S.C.; Scheller, K.K.; Williams, S.N.; Schaefer, D.M. Tissue equilibration and subcellular distribution of vitamin E relative to myoglobin and lipid oxidation in displayed beef. J. Anim. Sci. 1993, 71, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Renerre, M. Oxidative processes and myoglobin. In Antioxidants in muscle foods; Decker, E., Faustman, C., Lopez-Bote, C., Eds.; Wiley & Sons: New York, USA, 2000; pp. 113–135. [Google Scholar]
- Srivastava, R. Physicochemical, antioxidant properties of carotenoids and its optoelectronic and interaction studies with chlorophyll pigments. Sci. Rep. 2021, 11(1), 1–14. [Google Scholar] [CrossRef]
- Arend, F.A.; Murdoch, G.K.; Doumit, M.E.; Chibisa, G.E. Inclusion of grape pomace in finishing cattle diets: carcass traits, meat quality and fatty acid composition. Animals. 2022, 12(19), 2597. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Kang, G.; Seong, P.; Park, B.; Kang, S.M. Effect of slaughter age on the antioxidant enzyme activity, color, and oxidative stability of Korean Hanwoo (Bos Taurus coreanae) cow beef. Meat Sci. 2015, 108, 44–49. [Google Scholar] [CrossRef]
- Mitsumoto, M.; Ozawa, S.; Mitsuhashi, T.; Koide, K. Effect of dietary vitamin E supplementation for one week before slaughter on drip, colour and lipid stability during display in Japanese black steer beef. Meat Sci. 1998, 49, 165–174. [Google Scholar] [CrossRef]
- Zainudin, M.A.M.; Poojary, M.M.; Jongberg, S.; Lund, M.N. Light exposure accelerates oxidative protein polymerization in beef stored in high oxygen atmosphere. Food Chem. 2019, 299, 125132. [Google Scholar] [CrossRef] [PubMed]
- Campo, M.; Nute, G.; Hughes, S.; Enser, M.; Wood, J.; Richardson, R. Flavour perception of oxidation in beef. Meat Sci. 2006, 72, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Descalzo, A.M.; Insani, E.M.; Biolatto, A.; Sancho, A.M.; García, P.T.; Pensel, N.A.; Josifovich, J.A. Influence of pasture or grain-based diets supplemented with vitamin E on antioxidant/oxidative balance of Argentine beef. Meat Sci. 2005, 70, 35–44. [Google Scholar] [CrossRef]
- Simmone, A.H.; Green, N.R.; Bransby, D.I. Consumer acceptability and β-carotene content of beef as related to cattle finishing diets. J. Food Sci. 1996, 61(6), 1254–1257. [Google Scholar] [CrossRef]
- Yang, A.; Brewster, M.J.; Lanari, M.C.; Tunne, R.K. Effect of vitamin E supplementation on α-tocopherol and β-carotene concentrations in tissues from pasture- and grain-fed cattle. Meat Sci. 2002, 60, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Putnam, M.E.; Comben, N. Vitamin E—review article. Vet. Rec. 1987, 121(23), 541–545. [Google Scholar] [PubMed]
- De la Fuente, J.; Díaz, M.T.; Álvarez, I.; Oliver, M.A.; Font i Furnols, M.; Sañudo, C.; Campo, M.M.; Montossi, F.; Nute, G.R.; Cañeque, V. Fatty acid and vitamin E composition of intramuscular fat in cattle reared in different production systems. Meat Sci. 2009, 82(3), 331–337. [Google Scholar] [CrossRef] [PubMed]
- Realini, C.E.; Duckett, S.K.; Brito, G.W.; Dalla Rizza, M.; De Mattos, D. Effect of pasture vs concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Sci. 2004, 66, 567–577. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Norng, S.; Burnett, V.F.; Dunshea, F.R.; Jacobs, J.L.; Hopkins, D.L. The synergism of biochemical components controlling lipid oxidation in lamb muscle. Lipids. 2014, 49, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Manner, W.N.; Maxwell, R.J.; Williams, J.E. Effects of dietary regimen and tissue site on bovine fatty acid profiles. J. Anim. Sci. 1984, 59(1), 109–121. [Google Scholar] [CrossRef]
- Terevinto, A.; Cabrera, M.C.; Saadoun, A. Catalase, SOD and GPx activities in Triceps brachii muscle from Aberdeen Angus steers finished on pasture, pasture and concentrate, or concentrate. Am. J. Food Nutr. [CrossRef]
- Terevinto, A.; Cabrera, M.C.; Saadoun, A. Influence of feeding system on lipids and proteins oxidation, and antioxidant enzymes activities of meat from Aberdeen Angus steers. J. Food Nutr. Res. /: 581 – 586. http.
- Deckelbaum, R.J. n-6 and n-3 fatty acids and atherosclerosis. Ratios or amounts? Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2325–2326. [Google Scholar] [CrossRef] [PubMed]
- Pordomingo, A.J.; García, T.P.; Volpi Lagreca, G. Effect of feeding treatment during the backgrounding phase of beef production from pasture on: II. Longissimus muscle proximate composition, cholesterol and fatty acids. Meat Sci. [CrossRef]
- García, P.T.; Pensel, N.A.; Sancho, A.M.; Latimori, N.J.; Kloster, A.M.; Amigone, M.A.; Casal, J.J. Beef lipids in relation to animal breed and nutrition in Argentina. Meat Sci. 2008, 79, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Venkata Reddy, B.; Sivakumar, A.D.; Jeong, D.W.; Woo, Y-B. ; Park, S-J.; Lee, S-Y.; Byun, J-Y.; Kim, C-H.; Cho, S-H.; Hwang, I. Beef quality traits of heifer in comparison with steer, bull and cow at various feeding environments. Anim. Sci. J. 2015, 86, 1–16. [Google Scholar] [CrossRef]
- FAO-WHO. Fats and fatty acids in human nutrition. Rome: FAO Food and nutrition paper # 91. Report of an expert consultation, Geneva, 2010, 10-. https://www.who. 14 November.
- Stajic, S.; Zivkovic, D.; Perunovic, M.; Sobajic, S.; Vranic, D. Cholesterol content and atherogenicity of fermented sausages made of pork meat from various breeds. In 11th International Congress on Engineering and Food. Procedia Food Sci. 2011, 1, 568–575. [Google Scholar] [CrossRef]
- Alfaia, C.P.M.; Alves, S.P.; Martins, S.I.V.; Costa, A.S.H.; Fontes, C.M.G.A.; Lemos, J.P.C.; Bessa, R.J.B.; Prates, J.A.M. Effect of feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 2009, 114, 939–946. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Mann, N.J.; Sinclair, A.J. Effect of feeding systems on omega-3 fatty acids, conjugated linoleic acid and trans fatty acids in Australian beef cuts, potential impact on human health. Asia Pac. J. Clin. Nutr. 2006, 15(1), 21–29. [Google Scholar] [PubMed]
- Song, Z.; Xia, H.; Yang, L.; Wang, S.; Sun, G. Lowering the n-6/n-3 PUFAs ratio inhibits the formation of THP-1 macrophage-derived foam cell. Lipids Health Dis. 2018, 17(125), 1–8. [Google Scholar] [CrossRef] [PubMed]
- García, P.T.; Pensel, N.A.; Margaria, C.A.; Olga Rossco, C.M. Intramuscular fat, cholesterol and 18:2 n-6/18:3 n-3 ratio in total lipids in two frame steers under different dietary regimen. In Proceedings of the 45th International Congress of Meat Science and Technology, Yokohama, Japan, 1-7 August 1999; pp. 76–77. [Google Scholar]
- Yang, A.; Lanari, M.C.; Brewster, M.; Tume, R.K. Lipid stability and meat colour of beef from pasture- and grain-fed cattle with or without vitamin E supplement. Meat Sci. 2002, 60, 41–50. [Google Scholar] [CrossRef]
- Bennet, L.L.; Hammon, A.C.; Williams, M.J.; Kunkle, W.E.; Johnson, D.D.; Preston, R.L. Performance, carcass yield, and carcass quality characteristics of steers finished on rhizoma peanut-tropical grass pasture or concentrate. J. Anim. Sci. 1995, 73, 1881–1887. [Google Scholar] [CrossRef] [PubMed]
- Bidner, T.D.; Schupp, A.R.; Mohamad, A.B.; Rumore, N.C.; Montgomery, R.E.; Bagley, C.P. Acceptability of beef from Angus-Hereford or Angus-Hereford-Brahman steers finished on all-forage or a high energy diet. J. Anim. Sci. 1986, 63, 381–387. [Google Scholar] [CrossRef]
- Schroeder, J.W.; Cramer, D.A.; Bowling, R.A.; Cook, C.W. Palatability, shelf-life and chemical differences between forage- and grain-finished beef. J. Anim. Sci. 1980, 50, 852–859. [Google Scholar] [CrossRef]
- Vestergaard, M.; Oksbjerg, N.; Henckel, P. Influence of feeding intensity, grazing and finishing feeding on muscle fibre characteristics and meat colour of Semitendinosus, Longissimus dorsi and Supraspinatus muscles of young Bulls. Meat Sci. 2000, 54(2), 177–185. [Google Scholar] [CrossRef] [PubMed]
- Dunne, P.G.; Monahan, F.J.; Moloney, A.P. Current perspectives on the darker beef often reported from extensively-managed cattle: Does physical activity play a significant role? Livest. Sci. 2011, 142, 1–22. [Google Scholar] [CrossRef]
- Dunne, P.G.; Monahan, F.J.; O`Mara, F.P.; Moloney, A.P. Color stability, under simulated retail display conditions, of M. longissimus dorsi and M. semimembranosus from steers given long-term daily exercise and supplemented with vitamin E. Meat Sci. [CrossRef]
- Sapp, P.H.; Williams, S.E.; McCann, M.A. Sensory attributes and retail display characteristics of pasture- and/or grain-fed beef aged 7, 14 or 21 days. J. Food Qual. 1998, 22, 257–274. [Google Scholar] [CrossRef]
- Gatellier, P.; Mercier, Y.; Juin, H.; Renerre, M. Effect of finishing mode (pasture- or mixed diet) on lipid composition, color stability and lipid oxidation in meat from Charolais cattle. Meat Sci. 2005, 69, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Razminowicz, R.H.; Kreuzer, M.; Scheeder, M.R.L. Quality of retail beef from two grass-based production systems in comparison with conventional beef. Meat Sci. 2006, 73, 351–361. [Google Scholar] [CrossRef]
- Purohit, A.; Singh, R.; Kerr, W.; Mohan, A. Effects of heme and nonheme iron on meat quality characteristics during retail display and storage. J. Food Meas. Charact. 2015, 9, 175–185. [Google Scholar] [CrossRef]
Figure 1.
Lipid oxidation (a) and protein oxidation (b) before and during retail display in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate. Values are means ± SEM (n=10). Different capital letters show significant differences between feeding systems in each day of the display (P<0.05). Different lowercase letters show significant differences between days of display in each feeding system (P<0.05).
Figure 1.
Lipid oxidation (a) and protein oxidation (b) before and during retail display in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate. Values are means ± SEM (n=10). Different capital letters show significant differences between feeding systems in each day of the display (P<0.05). Different lowercase letters show significant differences between days of display in each feeding system (P<0.05).
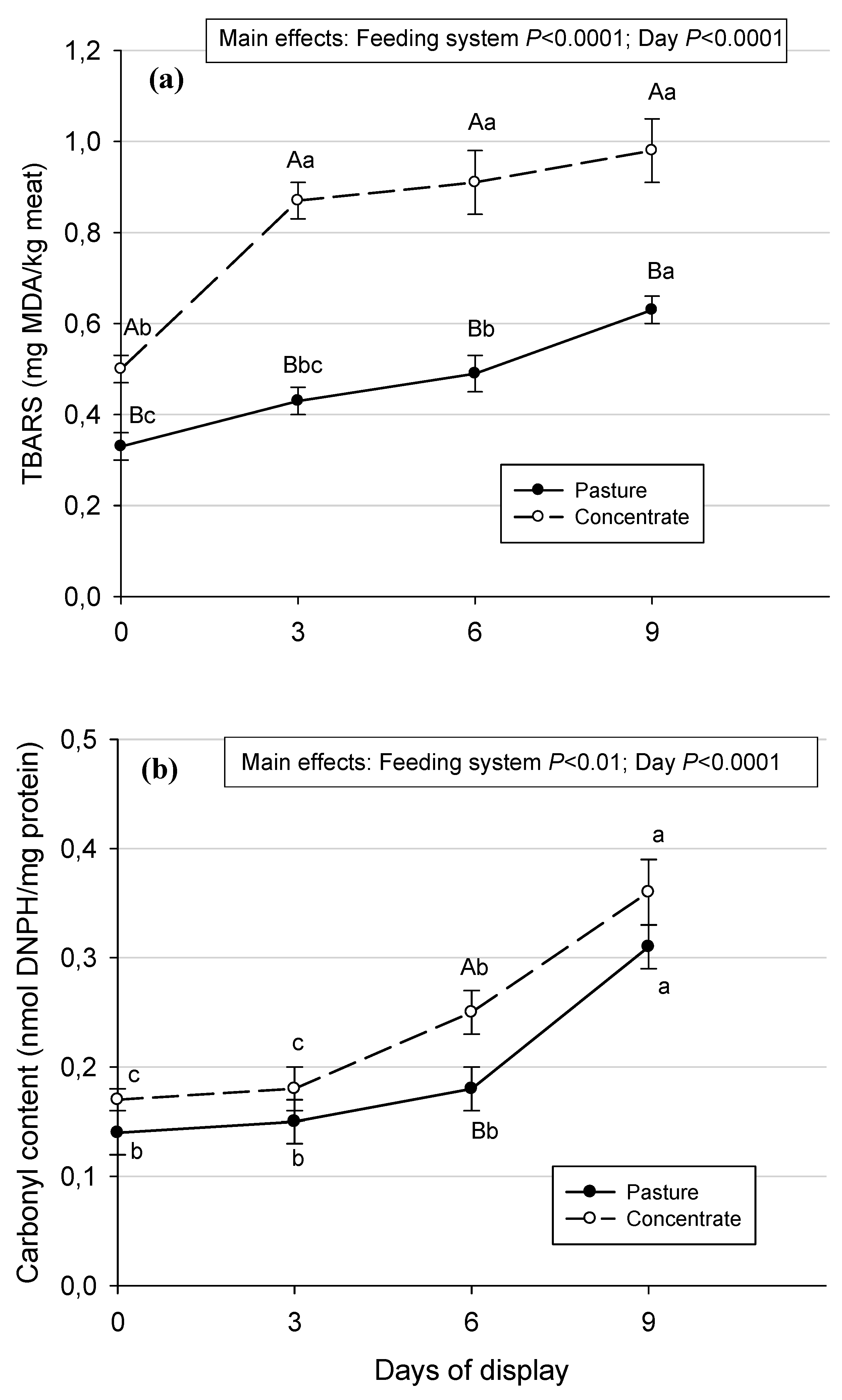
Figure 2.
Color parameters (L*, a*, b*) before and during retail display in Semimembranosus muscle of Aberdeen Angus steers fed pasture or concentrate. Values are means ± SEM (n=10). Different capital letters show significant differences between feeding systems in each day of the display (P<0.05). Different lowercase letters show significant differences between days of display in each feeding system (P<0.05). NS: not significant.
Figure 2.
Color parameters (L*, a*, b*) before and during retail display in Semimembranosus muscle of Aberdeen Angus steers fed pasture or concentrate. Values are means ± SEM (n=10). Different capital letters show significant differences between feeding systems in each day of the display (P<0.05). Different lowercase letters show significant differences between days of display in each feeding system (P<0.05). NS: not significant.
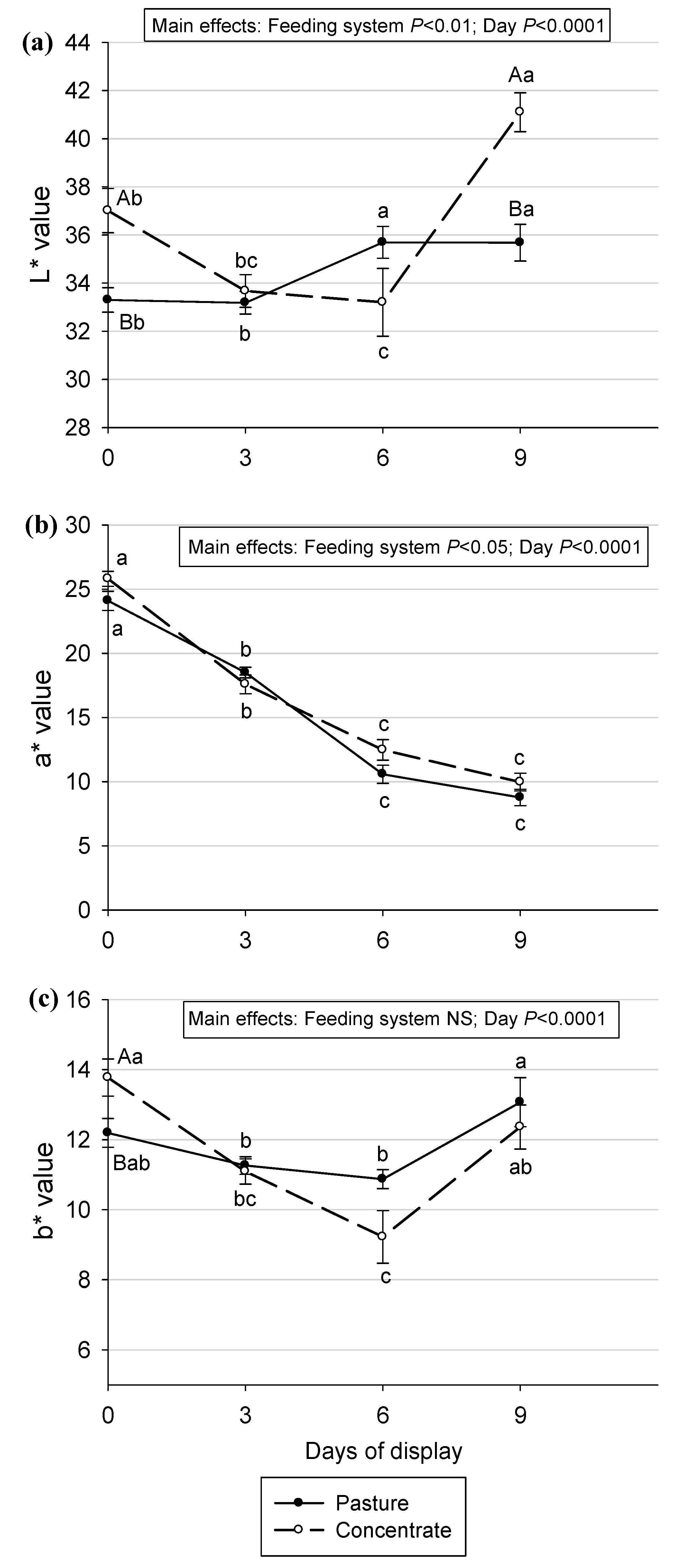

Table 1.
β-carotene and α-tocopherol content (μg/g fresh meat) in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate before the retail display.
Table 1.
β-carotene and α-tocopherol content (μg/g fresh meat) in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate before the retail display.
| Pasture | Concentrate CConcentratccConcentrat | p-value | |
|---|---|---|---|
| β-carotene | 1.73 ± 0.18 a | 0.27 ± 0.04 b | <0.0001 |
| α-tocopherol | 3.7 ± 0.5 a | 1.9 ± 0.3 b | <0.05 |
Values are means ± SEM (n=6). Different letters in the same row show significant differences between feeding systems (P < 0.05).
Table 2.
Total intramuscular fat content (%) and fatty acid composition (g/100 g fatty acids) in Semimembranosus muscle of Aberdeen Angus steers fed pasture or concentrate before the retail display.
Table 2.
Total intramuscular fat content (%) and fatty acid composition (g/100 g fatty acids) in Semimembranosus muscle of Aberdeen Angus steers fed pasture or concentrate before the retail display.
| Pasture | Concentrate | p-value | |
|---|---|---|---|
| % Fat | 1.78 ± 0.15b | 4.52 ± 0.46a | <0.0001 |
| C12:0 | 0.13 ± 0.04 | 0.08 ± 0.03 | NS |
| C14:0 | 3.46 ± 0.71 | 3.10 ± 0.85 | NS |
| C15:0i | 0.25 ± 0.04a | 0.12 ± 0.03b | 0.01 |
| C15:0ai | 0.26 ± 0.03 | 0.17 ± 0.04 | NS |
| C14:1 | 0.60 ± 0.12 | 0.59 ± 0.17 | NS |
| C15:0 | 0.72 ± 0.10 | 0.51 ± 0.11 | NS |
| C16:0i | 0.20 ± 0.03a | 0.10 ± 0.01b | <0.05 |
| C16:0 | 31.85 ± 2.71 | 28.75 ± 2.45 | NS |
| C16:1 | 4.16 ± 0.38 | 4.22 ± 0.35 | NS |
| C17:0 | 1.10 ± 0.04b | 1.27 ± 0.05a | 0.01 |
| C17:1 | 0.74 ± 0.24 | 0.94 ± 0.07 | NS |
| C18:0 | 12.12 ± 1.25 | 12.53 ± 1.08 | NS |
| C18:1 | 34.46 ± 2.91 | 41.53 ± 2.57 | NS |
| C18:2 n-6 | 2.84 ± 0.36 | 3.07 ± 0.22 | NS |
| C20:1 | 0.13 ± 0.04 | 0.21 ± 0.08 | NS |
| C18:3 n-3 | 0.61 ± 0.04a | 0.18 ± 0.03b | 0.001 |
| CLA | 0.31 ± 0.03 | 0.31 ± 0.09 | NS |
| C20:3 n-3 | 0.14 ± 0.04a | 0.03 ± 0.01b | 0.01 |
| C20:3 n-6 | 0.34 ± 0.10a | 0.11 ± 0.02b | 0.05 |
| C20:4 n-6 | 0.30 ± 0.12 | 0.21 ± 0.06 | NS |
| EPA n-3 | 0.04 ± 0.05 | 0.01 ± 0.01 | NS |
| DPA n-3 | 0.13 ± 0.08 | 0.04 ± 0.02 | NS |
| DHA n-3 | 0.41 ± 0.06a | 0.08 ± 0.07b | <0.01 |
| Others | 4.73 ± 1.37 | 1.92 ± 0.41 | …… |
| SFA | 50.09 ± 2.54 | 46.64 ± 2.39 | NS |
| MUFA | 40.08 ± 2.68b | 47.49 ± 2.19a | <0.05 |
| PUFA | 5.13 ± 0.62 | 4.02 ± 0.49 | NS |
| Σn-6 | 3.79 ± 0.41 | 3.69 ± 0.37 | NS |
| Σn-3 | 1.33 ± 0.24a | 0.33 ± 0.13b | <0.001 |
| n-6/n-3 | 2.85 ± 0.35b | 11.18 ± 3.96a | <0.05 |
| PUFA/SFA | 0.10 ± 0.02 | 0.09 ± 0.02 | NS |
| Atherogenic index | 1.02 ± 0.19 | 0.81 ± 0.15 | NS |
| Thrombogenic index | 1.82 ± 0.24 | 1.67 ± 0.19 | NS |
Values are means ± SEM (n=10). Different letters in the same row show significant differences between feeding systems (P<0.05). CLA: conjugated linoleic acid; EPA: eicosapentaenoic acid; DPA: docosapentaenoic acid; DHA: docosahexaenoic acid; SFA: saturated fatty acids; MUFA: monounsaturated fatty acids; PUFA: polyunsaturated fatty acids; NS: not significant. i:iso; ai:anteiso.
Table 3.
Heme iron content (ppm) in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate before and during the retail display.
Table 3.
Heme iron content (ppm) in Semimembranosus muscle from Aberdeen Angus steers fed pasture or concentrate before and during the retail display.
| Day 0 | Day 3 | Day 6 | Day 9 | p-value | |
|---|---|---|---|---|---|
| Pasture | 19.3 ± 1.0 | 20.5 ± 0.9 | 19.5 ± 0.7 | 19.2 ± 1.5 | NS |
| Concentrate | 20.2 ± 1.4 | 20.5 ± 2.5 | 21.0 ± 2.8 | 19.3 ± 2.8 | NS |
| p-value | NS | NS | NS | NS |
Values are means ± SEM (n=6).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



