
Submitted:
10 August 2023
Posted:
11 August 2023
You are already at the latest version
Abstract
The pathogen Cryptococcus neoformans is a major cause of fungal meningitis in humans. As with other melanotic microorganisms associated with human diseases, the cell wall-associated melanin of C. neoformans is a major virulence factor that contributes to its ability to evade host immune responses. The levels of melanin substrate and the regulation of melanin formation could be influenced by the microbiota-gut-brain axis. Moreover, recent studies show that C. neoformans infection causes dysbiosis in the human gut microbiome. In this review, we discuss the potential association between cryptococcal meningitis and the gut microbiome. Additionally, we emphasize the promising prospects of targeting the gut microbiome as a means of diagnosing and treating this debilitating disease.
Keywords:
cryptococcal meningitis
; melanin
; catecholamines
; nutritional signals
; microbiota-gut-brain axis
1. Introduction
Cryptococcus neoformans is a human pathogenic fungus, classified within the phylum Basidiomycota [1,2]. Although it possesses a clearly defined bipolar mating system, consisting of two distinct mating types, MATa and MATα [3,4], it is also capable of undergoing self-fertilization within cells of the same mating type [5,6]. The fungi is broadly distributed in the environment, particularly in bird guano, soil and trees [7]. C. neoformans is especially dangerous for immunocompromised hosts including the HIV/AIDS population and organ transplant recipients due to its propensity to cause meningoencephalitis [8,9,10,11,12,13]. For example, C. neoformans is responsible for around 223,100 new cases and 181,000 deaths annually among individuals with HIV/AIDS [14,15]. Importantly, the fungus is classified as one of the four species in the highest priority group of fungal pathogens affecting humans, as identified by the World Health Organization (WHO). C. neoformans infections commonly occur through the inhalation of fungal spores or desiccated yeast cells in the environment [16,17], as well as through colonization of skin wounds [18]. The pathogen is able to overcome host defenses and enter the bloodstream, where it can disseminate to various organs including the central nervous system (CNS), causing meningitis [16,19]. The mortality rate of cryptococcal meningitis is high, particularly in developing countries where access to effective treatments is limited. Despite all the efforts being made to create an effective vaccine, there is currently no reliable method for preventing infection. Even the most advanced treatment methods, which use a combination of antifungal medications, can only bring down the 10-week mortality rate to 24% [20]. Treatment of C. neoformans infections typically involves a combination of antifungal drugs, such as amphotericin B and flucytosine [8,20,21,22]. Successful phase III trial results on the combination therapy of amphotericin B and flucytosine have led to the WHO updating its treatment guidelines for cryptococcal disease in HIV-positive patients [22,23]. However, the emergence of drug-resistant strains of C. neoformans is a growing concern, highlighting the need for continued research into new therapies and preventative measures [20,24,25,26,27,28].
The diagnosis of Cryptococcal meningitis typically involves a comprehensive evaluation of clinical symptoms and signs, as well as the use of various diagnostic methods [21,29,30,31]. These methods involve evaluating the patient’s medical history and conducting a physical examination to identify symptoms associated with Cryptococcal meningitis. Additionally, a lumbar puncture is commonly performed to collect and analyze cerebrospinal fluid (CSF) for the presence of Cryptococcus. Laboratory tests, including India ink staining of CSF, culture of C. neoformans, PCR, or antigen testing, are also employed to confirm the presence of Cryptococcus infection. Various neuroimaging techniques, such as computed tomography (CT) imaging and magnetic resonance imaging (MRI), can be used to examine the brain for signs of meningitis or other abnormalities. The integration of these methods is essential for an accurate diagnosis and to rule out other potential causes. Research suggests that the composition of the gut microbiome may have a potential role in the diagnosis of cryptococcal meningitis. Studies have shown that alterations in the gut microbiome may be associated with fungal infections, including Cryptococcus [32]. By analyzing the microbial communities in the gut, it may be possible to identify specific changes or markers that can aid in the diagnosis of cryptococcal meningitis. However, further research is needed to validate and establish the use of gut microbiome testing as a diagnostic tool for cryptococcal meningitis.
The C. neoformans melanin in the wall, the major virulence factor, has various functions such as protecting against oxidative stress, reducing the efficiency of antifungal drugs, and affecting interactions with phagocytic cells [16,33,34]. Researchers believe that melanin could be a potential target for the development of drugs aimed at combating C. neoformans [20,35]. One distinguishing characteristic of the C. neoformans laccase enzyme is its inability to generate melanin pigments from endogenously synthesized compounds, such as tyrosine. This sets it apart from other melanotic organisms that commonly utilize tyrosine as a precursor for melanin production [36]. In the brain tissue, C. neoformans produces melanin pigments by oxidizing exogenous catecholamines (ie, dopamine, norepinephrine, and epinephrine) through the laccase enzymes (Lac1 and Lac2) [37,38,39]. The neurotropism of C. neoformans is notably linked to the presence of catecholamines in the brain, along with low concentrations of glucose in the CNS. This is because under these conditions, laccase production is overregulated [38,40,41]. Recent research suggests that microbiota-gut-brain axis (MGBA) can regulate the levels of C. neoformans melanin substrates in the brain [42]. MGBA can also potentially influence the host’s nutritional signals, such as glucose [43], that regulate melanin formation [41]. Understanding the complex interplay between C. neoformans melanization and the MGBA may be crucial for the development of effective treatments for this fungal meningitis. In this review, we provide an overview of the existing knowledge regarding the possible links between cryptococcal meningitis and the gut microbiome. By gaining a deeper understanding of the connection, we may be able to develop more effective strategies for tests or treatments for cryptococcal meningitis.
2. C. neoformans infection causes gut microbiome disruption
Recent advancements in our understanding and analysis of the gut microbiota have unveiled the significant impact of alterations on human health. The gut microbiota, an intricate and diverse microbial ecosystem, plays a vital role in various aspects of the host’s physiology. It actively participates in the host’s immune response, influences metabolism, facilitates biosynthesis, and defends against the pathogenic yeast infections [44,45,46,47]. In instances, the composition of the gut bacterial microbiota has been found to influence the generation of the pulmonary IL-17 response during the opportunistic human fungal pathogen Aspergillus fumigatus infection in mice. Mice with a specific composition of gut bacteria exhibit a more robust IL-17 response, which is crucial for combating fungal infections [48]. Furthermore, systemic infection with the invasive fungal pathogen Candida albicans has been shown to negatively impact the composition and diversity of the gut microbiota. This disturbance in microbial diversity can disrupt the delicate equilibrium within the gut ecosystem, potentially affecting overall gut health and immune function [49]. Similarly, studies have demonstrated the critical role of the microbiota in the host’s defense against Cryptoccocus gattii infection. Germ-free mice, lacking a microbiota, exhibit heightened susceptibility to C. gattii infection, resulting in lower survival rates and increased fungal burden in the brain and lungs. Furthermore, they display reduced levels of key immune factors such as IL-17, interleukin (IL)-1β, and interferon-γ, as well as decreased phosphorylation of the nuclear factor κB p65, compared to their wild-type counterparts [50]. In summary, the commensal microbiota plays a significant role in modulating immune responses during invasive fungal infections. Manipulating its composition holds promise as a potential therapeutic approach.
Recently, Li and colleagues reported that Cryptoccocus infection induces alterations in the gut microbiota of humans [32]. The authors performed alpha and beta diversity analyses to compare the gut microbiota of patients with cryptococcal meningitis to healthy controls. The results showed that patients with cryptococcal meningitis had significantly lower alpha diversity compared to healthy controls, indicating gut dysbiosis. Totally, they identified 72 differentially abundant bacterial and 8 differentially abundant fungal species between these two groups. For example, the patients with cryptococcal meningitis had a higher abundance of bacteria Enterococcus avium, Microbacterium foliorum and Bacteroides spp and of fungi Pyricularia spp, Cytospora leucostoma and Wallemia ichthyophaga, and a lower abundance of bacteria Prevotella spp, Coprococcus spp and Arthrobacter woluwensis and of fungi Jimgerdemannia flammicorona, Metschnikowia aff. pulcherrima and Pyricularia pennisetigena as compared to the healthy controls. Interestingly, anti-fungal treatment had only minor effects on the gut microbiota composition, suggesting that C. neoformans infection causes long-lasting gut microbiota dysbiosis in the cryptococcal meningitis patients. To further explore the potential correlations between bacteria, fungi and clinical indicators of cryptococcal meningitis, the authors performed correlation analyses. Several bacterial and fungal taxa were found to be positively or negatively correlated with the disease-related symptoms, such as visual disorders and auditory symptoms. Overall, Li et al’s study provides valuable insights into the significant impact of C. neoformans infection on the gut microbiota and the potential associations between cryptococcal meningitis and gut microbiome disruption.
3. C. neoformans melanization in human brain tissue
For C. neoformans, the cell surface features that contribute to pathogenesis include the deposition of melanin in the cell wall [51,52]. Melanin production offers several advantages to the pathogen [53]. Firstly, melanin acts as a protective barrier against host immune responses, including phagocytosis by immune cells. Melanin has been shown to inhibit the production of reactive oxygen species and diminish the activity of antifungal agents, thus increasing the resistance of C. neoformans to host defenses. Moreover, melanin has been implicated in the dissemination of C. neoformans within the host. Melanized fungal cells have been detected in various organs, including the brain, and lungs, indicating that melanization plays a role in the invasion and establishment of infections in different tissues [54]. The primary indication of C. neoformans melanization during infection came from the identification of acid-resistant melanin ghost particles. These particles were isolated from infected animal and human tissue, as well as from cells cultivated on agar plates with tissue homogenate [55,56]. The nervous system of mammalian serves as an abundant source of precursors in the form of catecholamines, which are nitrogen-containing diphenolic compounds. These catecholamines include neurotransmitters like dopamine, epinephrine, and norepinephrine [57,58]. C. neoformans produces melanin by catalyzing the oxidation of exogenous catecholamine substrates using laccase enzymes [59,60,61]. Melanized C. neoformans cells were detected in brain tissue samples from patients with cryptococcal meningitis [55]. The melanin synthesized by C. neoformans in brain tissue may vary in different anatomical regions due to its ability to incorporate multiple catecholamines simultaneously (Figure 1). This is because the relative proportions of these neurotransmitters can differ significantly from one area of the brain to another [57,58]. The chemical structure of the substrate added to the media dictates the variability of the synthesized pigment type. Importantly, Baker et al., reported that C. neoformans could exploit mixture of human brain catecholamine brain mixture (0.6 mM dopamine, 0.33 mM norepinephrine, and 0.07 mM epinephrine) to produce polytypic melanin [37]. Interestingly, C. neoformans infections show a notable concentration in the basal ganglia region of the brain, where dopamine levels are highest [62]. This finding raises intriguing possibilities, such as the potential resistance of melanized cells to immune clearance mechanisms. Additionally, this preference for the basal ganglia region aligns with the hypothesis that the melanin produced by C. neoformans in the brain is primarily derived from dopamine. Taken together, the detection of melanized fungal cells in human brain tissue samples highlights the ability of C. neoformans to undergo melanization within the brain. The presence of multiple catecholamines within the human brain provides substrates for polytypic melanin synthesis (Figure 1), thereby augmenting the virulence and survivability of the fungus. The extent of melanization varied among different brain regions, with higher levels observed in the basal ganglia and thalamus. This suggests that melanization may play a role in the localization and dissemination of C. neoformans within the brain.
4. The possible impact of gut microbiome on C. neoformans melanization in brain
4.1. Gut microbiome influences the levels of melanin substrates
The gut-brain axis is a bidirectional communication system connecting the gut microbiome and the central nervous system [63,64]. It is widely recognized that the gut microbiome has the ability to impact various aspects of brain function, such as mood, behavior, and cognition [65,66]. This is believed to happen through different pathways, such as the production of neurotransmitters and other signaling molecules that can influence the functioning of the central nervous system [65,67]. While there is no direct evidence linking the gut-brain axis to the synthesis of C. neoformans melanin, evidence suggests that the gut microbiome has the potential to influence levels of catecholamines (Figure 2), such as dopamine and norepinephrine, which are the melanin substrate for C. neoformans in brain tissue. For instance, recent studies have indicated that the level of striatal dopamine is regulated by the colonization of intestinal microbes. Metabolites produced by gut bacteria are absorbed into the bloodstream and can cross the blood-brain barrier, where they have an impact on the dopamine system in the brain. Germ-free mice, which were colonized with a modified strain of Escherichia coli containing genes responsible for fatty acid amides (FAAs) biosynthesis, exhibited improved running performance on both exercise wheels and treadmills compared to the control group. Furthermore, the activation of the endocannabinoid receptor CB1, which is expressed in neurons that also express vanilloid receptor TRPV1, by FAAs led to an increase in sensory neuron activity, the release of dopamine in the striatum, and an overall enhancement in exercise performance [42]. Additionally, germ-free mice have higher turnover rates of catecholamines dopamine and norepinephrine in the brain [66]. The levels of norepinephrine in the hippocampus were significantly lower in mice with antibiotics-induced depression compared to normal mice [68]. In conclusion, the gut microbiome has the potential to influence C. neoformans melanin substrate levels.
4.2. Gut microbiome impact on nutritional signals that regulate melanization
The ability to quickly adapt to fluctuating external conditions is vital for the survival and propagation of microorganisms. This is especially significant for pathogenic microbes as they need to navigate the shift from the environment to the host milieu and initiate an appropriate response to establish an infection. Hosts pose challenging conditions, such as varying nutrient availability, oxygen levels, pH levels, and temperature, along with the potential threats posed by the host immune response [69,70]. It is interesting to note that the mechanisms of adapting to nutrient availability not only facilitate the proliferation of microorganisms but also play a role in regulating their virulence [71]. Glucose levels, for example, have been found to be a key factor in promoting melanin synthesis in C. neoformans [41]. When the fungus is starved of glucose, it responds by increasing its melanin production. Signal transduction pathways are crucial in facilitating microbial adaptation. The cAMP-PKA nutrient sensing pathway has also been discovered to play a crucial role in regulating melanin synthesis in C. neoformans. A glucose and methionine mixture has been shown to increase cAMP accumulation and PKA activation, leading to increased melanin synthesis [72,73].
The gut microbiome can regulate the availability of nutrients such as glucose and amino acids for C. neoformans [43]. Imbalances or alterations in the gut microbiome composition may therefore affect the levels of these nutritional signals, potentially influencing melanization processes in the body (Figure 2). The gut microbiome, consisting of trillions of microorganisms including bacteria, viruses, and fungi, is crucial in maintaining a healthy nutritional status [74]. It interacts with the host’s diet in complex ways, including breaking down complex carbohydrates and fibers that are not easily digestible by the host. Fermentation of these fibers results in the production of short-chain fatty acids, which play a crucial role in regulating various metabolic pathways in the gut and throughout the body, including the liver, adipose tissue, muscles, and brain [75,76,77,78,79]. These microbial metabolites are now recognized for their significant contributions to a wide range of physiological effects. These effects include the modulation of glucose metabolism, energy homeostasis, inflammation, as well as impacts on the immune system and cancer [78]. Additionally, the gut microbiome produces vitamins and amino acids that are essential for various metabolic and physiological processes. It can also enhance the bioavailability of minerals by producing chelators that bind to them. However, dysbiosis, alterations in the gut microbiome, can lead to malnutrition and deficiencies due to impaired digestion and absorption of nutrients, inflammation, and damage to the gut lining. The use of interventions like probiotics and prebiotics may be beneficial in promoting better nutrient absorption and overall health. Therefore, a healthy gut microbiome is essential for maintaining a healthy nutritional status. Research has shown that the gut microbiome can also influence glucose metabolism through the production of hormones like glucagon-like peptide-1 (GLP-1) and peptide YY (PYY), which regulate glucose levels and appetite [80,81]. These hormones are produced by specialized cells in the intestines, called enteroendocrine cells, which are in direct contact with the gut microbiome. The gut microbiome can also modulate the release of insulin by affecting the function of pancreatic beta cells, which are responsible for producing and releasing insulin in response to glucose levels in the blood. Furthermore, recent studies have suggested that dysbiosis in the gut microbiome may be linked to the development of insulin resistance and type 2 diabetes, as well as other metabolic disorders. For example, changes in the gut microbiome composition, such as an increase in harmful bacteria or a decrease in beneficial bacteria, have been associated with the development of insulin resistance. This highlights the importance of maintaining a healthy gut microbiome for the regulation of glucose metabolism and the prevention of metabolic disorders. In summary, the gut microbiome plays a crucial role in maintaining a healthy nutritional status by breaking down complex carbohydrates and fibers, producing essential nutrients, and enhancing mineral bioavailability. It also influences glucose metabolism through the production of enzymes, hormones, and the modulation of insulin release [43]. Therefore, maintaining a healthy gut microbiome is essential for overall health and preventing the development of metabolic disorders, including type 2 diabetes and cardiovascular disease. It is well-established that diet plays a crucial role in shaping the gut microbiome composition. Certain dietary components, such as prebiotics and probiotics, have been shown to modulate the gut microbiome and improve brain function. Therefore, exploring the effects of dietary interventions on melanin substrate levels and C. neoformans melanin synthesis could provide insights into potential therapeutic interventions for C. neoformans infection.
5. Conclusions
The current understanding of the relationship between cryptococcal meningitis and the gut microbiome is limited, but recent studies have shed light on potential connections. While no direct link has been established, evidence suggests that the presence of C. neoformans infection can have a significant impact on the composition and function of the gut microbiota. Studies have shown that the gut microbiome may play a role in influencing C. neoformans melanin formation. Melanin is a key component of the cryptococcal cell wall and is known to contribute to the pathogenicity of the fungus. It has been proposed that certain gut microbial species may modulate the availability of melanin precursors or provide metabolic support for melanin synthesis in C. neoformans. This hypothesis opens up new avenues for research into the specific microbial species and their potential involvement in melanin production. While the correlations between cryptococcal meningitis and the gut microbiome are still being actively investigated, it is clear that further studies are needed to fully understand the underlying mechanisms of this interaction. Future research should focus on elucidating the specific microbial species that are associated with C. neoformans infection and determining their functional roles in modulating disease progression. The potential implications of this research are significant. By gaining a better understanding of the relationship between cryptococcal meningitis and the gut microbiome, novel strategies for the prevention and treatment of C. neoformans infections can be developed. Targeting the gut microbiome through probiotics, prebiotics, or other interventions may offer a new approach to enhance the host’s immune response against the fungus or interfere with its pathogenic mechanisms. Furthermore, studying the gut microbiome in the context of cryptococcal meningitis may also provide insights into the broader field of infectious diseases. The gut microbiota has been implicated in the regulation of immune responses and the maintenance of overall health. Therefore, understanding how alterations in the gut microbiome influence the development and progression of cryptococcal meningitis could have implications beyond this specific infection. While the exact connection between cryptococcal meningitis and the gut microbiome remains unclear, recent studies have highlighted the impact of C. neoformans infection on the gut microbiota. Further research is needed to explore the role of specific gut microbial species in modulating melanin formation and to unravel the underlying mechanisms of this connection. This area of research holds promise for the development of innovative approaches to prevent and treat C. neoformans infections by targeting the gut microbiome.
Author Contributions
Y.-Y.M., X.-Y.Z. and P.X. were the major contributors in writing the manuscript. Y.-Y.M., L.Y., M.-N.J., X.-Y.Z. and P.X. read, corrected and approved the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 82173554), Natural Science Foundation of Jiangsu Province (BK20201444), Qing Lan Project for Excellent Young Key Teachers of Colleges and Universities of Jiangsu Province (2020) and Nantong University student innovation training program project (202310304049Z).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.; Janbon, G.; Idnurm, A.; Bahn, Y.S. , Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harbor perspectives in medicine 2014, 4, a019760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lin, X. , Cryptococcus neoformans: Sex, morphogenesis, and virulence. Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases 2021, 89, 104731. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J. , A new species of Filobasidiella, the sexual state of Cryptococcus neoformans B and C serotypes. Mycologia 1976, 68, 943–6. [Google Scholar] [PubMed]
- Kwon-Chung, K.J. , A new genus, filobasidiella, the perfect state of Cryptococcus neoformans. Mycologia 1975, 67, 1197–200. [Google Scholar] [CrossRef]
- Sun, S.; Heitman, J. From Two to One: Unipolar Sexual Reproduction. Fungal Biol Rev 2015, 29, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lin, X. , Mechanisms of unisexual mating in Cryptococcus neoformans. Fungal genetics and biology : FG & B 2011, 48, 651–60. [Google Scholar]
- Lin, X.; Heitman, J. , The biology of the Cryptococcus neoformans species complex. Annu Rev Microbiol 2006, 60, 69–105. [Google Scholar] [CrossRef]
- May, R.C.; Stone, N.R.; Wiesner, D.L.; Bicanic, T.; Nielsen, K. , Cryptococcus: from environmental saprophyte to global pathogen. Nature Reviews Microbiology 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Okurut, S.; Boulware, D.R.; Olobo, J.; Meya, D.B. , Landmark clinical observations and immunopathogenesis pathways linked to HIV and Cryptococcus fatal central nervous system co-infection. Mycoses 2020, 63, 840–853. [Google Scholar] [CrossRef]
- Shroufi, A.; Chiller, T.; Jordan, A.; Denning, D.W.; Harrison, T.S.; Govender, N.P.; Loyse, A.; Baptiste, S.; Rajasingham, R.; Boulware, D.R. , Ending deaths from HIV-related cryptococcal meningitis by 2030. The Lancet Infectious Diseases 2021, 21, 16–18. [Google Scholar] [CrossRef]
- Murphy, L.S.; Lacy, A.J.; Smith, A.T.; Shah, K.S. , Cryptococcal meningitis in an immunocompetent patient. The American journal of emergency medicine 2020, 38, 2492.e1–2492.e3. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.M.; Montrief, T.; Ramzy, M. , Cryptococcal meningitis: a review for emergency clinicians. 2021, 16, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ye, L.; Zhao, F.; Zhang, L.; Lu, Z.; Chu, T.; Wang, S.; Liu, Z.; Sun, Y.; Chen, M.; Liao, G.; Ding, C.; Xu, Y.; Liao, W.; Wang, L. Cryptococcus neoformans, a global threat to human health. 2023, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. , Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. The Lancet infectious diseases 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Mourad, A.; Perfect, J.R. , The war on cryptococcosis: A Review of the antifungal arsenal. Memorias do Instituto Oswaldo Cruz 2018, 113, e170391. [Google Scholar] [CrossRef] [PubMed]
- Kronstad, J.W.; Attarian, R.; Cadieux, B.; Choi, J.; D’souza, C.A.; Griffiths, E.J.; Geddes, J.M.; Hu, G.; Jung, W.H.; Kretschmer, M. , Expanding fungal pathogenesis: Cryptococcus breaks out of the opportunistic box. Nature reviews Microbiology 2011, 9, 193–203. [Google Scholar] [CrossRef]
- Botts, M.R.; Hull, C.M. , Dueling in the lung: how Cryptococcus spores race the host for survival. Curr Opin Microbiol 2010, 13, 437–42. [Google Scholar] [CrossRef]
- Lenz, D.; Held, J.; Goerke, S.; Wagner, D.; Tintelnot, K.; Henneke, P.; Hufnagel, M. , Primary cutaneous cryptococcosis in an eight-year-old immunocompetent child: how to treat? Klinische Padiatrie 2015, 227, 41–4. [Google Scholar] [CrossRef]
- Dang, E.V.; Lei, S.; Radkov, A.; Volk, R.F. , Secreted fungal virulence effector triggers allergic inflammation via TLR4. 2022, 608, 161–167. [Google Scholar] [CrossRef]
- Iyer, K.R.; Revie, N.M.; Fu, C.; Robbins, N.; Cowen, L.E. , Treatment strategies for cryptococcal infection: challenges, advances and future outlook. Nature Reviews Microbiology 2021, 19, 454–466. [Google Scholar] [CrossRef]
- Ngan, N.T.T.; Flower, B.; Day, J.N. Treatment of Cryptococcal Meningitis: How Have We Got Here and Where are We Going? 2022, 82, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Molloy, S.F.; Kanyama, C.; Heyderman, R.S.; Loyse, A.; Kouanfack, C.; Chanda, D.; Mfinanga, S.; Temfack, E.; Lakhi, S.; Lesikari, S.; Chan, A.K.; Stone, N.; Kalata, N.; Karunaharan, N.; Gaskell, K.; Peirse, M.; Ellis, J.; Chawinga, C.; Lontsi, S.; Ndong, J.G.; Bright, P.; Lupiya, D.; Chen, T.; Bradley, J.; Adams, J.; van der Horst, C.; van Oosterhout, J.J.; Sini, V.; Mapoure, Y.N.; Mwaba, P.; Bicanic, T.; Lalloo, D.G.; Wang, D.; Hosseinipour, M.C.; Lortholary, O.; Jaffar, S.; Harrison, T.S. , Antifungal Combinations for Treatment of Cryptococcal Meningitis in Africa. The New England journal of medicine 2018, 378, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for the diagnosis, prevention and management of cryptococcal disease in HIV-infected adults, adolescents and children (WHO, 2018).
- Arastehfar, A.; Gabaldón, T. , Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. 2020, 9. [Google Scholar] [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A. , Tackling the emerging threat of antifungal resistance to human health. 2022, 20, 557–571. [Google Scholar] [PubMed]
- Kano, R.; Sugita, T.; Kamata, H. , Antifungal Susceptibility of Clinical Isolates and Artificially Produced Multi-azole-resistant Strains of Cryptococcus neoformans (formerly: Cryptococcus grubii) to Ravuconazole. Medical mycology journal 2020, 61, 11–13. [Google Scholar] [CrossRef]
- Bermas, A.; Geddes-McAlister, J. , Combatting the evolution of antifungal resistance in Cryptococcus neoformans. Molecular microbiology 2020, 114, 721–734. [Google Scholar] [CrossRef]
- Zafar, H.; Altamirano, S.; Ballou, E.R.; Nielsen, K. , A titanic drug resistance threat in Cryptococcus neoformans. Curr Opin Microbiol 2019, 52, 158–164. [Google Scholar] [CrossRef]
- Poplin, V.; Boulware, D.R.; Bahr, N.C. , Methods for rapid diagnosis of meningitis etiology in adults. 2020, 14, 459–479. [Google Scholar]
- Rajasingham, R.; Wake, R.M.; Beyene, T.; Katende, A.; Letang, E.; Boulware, D.R. , Cryptococcal Meningitis Diagnostics and Screening in the Era of Point-of-Care Laboratory Testing. J Clin Microbiol 2019, 57. [Google Scholar] [CrossRef]
- Fang, W.; Wu, J.; Cheng, M.; Zhu, X.; Du, M.; Chen, C.; Liao, W.; Zhi, K.; Pan, W. , Diagnosis of invasive fungal infections: challenges and recent developments. Journal of biomedical science 2023, 30, 42. [Google Scholar] [CrossRef]
- Li, H.; Zhang, L.; Zhang, K.; Huang, Y.; Liu, Y.; Lu, X.; Liao, W.; Liu, X.; Zhang, Q.; Pan, W. , Gut microbiota associated with cryptococcal meningitis and dysbiosis caused by anti-fungal treatment. Front Microbiol 2023, 13, 1086239. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.F.; Casadevall, A. , The role of melanin in fungal pathogenesis for animal hosts. Fungal Physiology and Immunopathogenesis 2019, 1–30. [Google Scholar]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. , The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Frontiers in microbiology 2020, 10, 2993. [Google Scholar] [CrossRef]
- Wang, Y.; Casadevall, A. , Susceptibility of melanized and nonmelanized Cryptococcus neoformans to the melanin-binding compounds trifluoperazine and chloroquine. Antimicrobial agents and chemotherapy 1996, 40, 541–5. [Google Scholar] [CrossRef] [PubMed]
- Williamson, P.R. , Biochemical and molecular characterization of the diphenol oxidase of Cryptococcus neoformans: identification as a laccase. J Bacteriol 1994, 176, 656–64. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.P.; Chrissian, C.; Stark, R.E.; Casadevall, A. , Cryptococcus neoformans melanization incorporates multiple catecholamines to produce polytypic melanin. Journal of Biological Chemistry 2022, 298. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Jang, E.-H.; Lee, M.; Kim, S.-W.; Lee, Y.; Lee, K.-T.; Bahn, Y.-S. , Unraveling melanin biosynthesis and signaling networks in Cryptococcus neoformans. MBio 2019, 10, e02267–e19. [Google Scholar] [CrossRef]
- Williamson, P.R.; Wakamatsu, K.; Ito, S. , Melanin biosynthesis in Cryptococcus neoformans. J Bacteriol 1998, 180, 1570–2. [Google Scholar] [CrossRef]
- Li, Z.; Bi, J.; Yang, J.; Pan, J.; Sun, Z.; Zhu, X. , Requirement of a Tsp2-type tetraspanin for laccase repression and stress resistance in the basidiomycete Cryptococcus neoformans. Applied and environmental microbiology 2012, 78, 21–7. [Google Scholar] [CrossRef]
- Nurudeen, T.A.; Ahearn, D.G. , Regulation of melanin production by Cryptococcus neoformans. J Clin Microbiol 1979, 10, 724–9. [Google Scholar] [CrossRef]
- Dohnalová, L.; Lundgren, P.; Carty, J.R.; Goldstein, N.; Wenski, S.L.; Nanudorn, P.; Thiengmag, S.; Huang, K.-P.; Litichevskiy, L.; Descamps, H.C. , A microbiome-dependent gut–brain pathway regulates motivation for exercise. Nature 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Grasset, E.; Burcelin, R. , The gut microbiota to the brain axis in the metabolic control. Reviews in endocrine & metabolic disorders 2019, 20, 427–438. [Google Scholar]
- Erny, D.; Dokalis, N.; Mezö, C.; Castoldi, A.; Mossad, O.; Staszewski, O.; Frosch, M.; Villa, M.; Fuchs, V.; Mayer, A.; Neuber, J.; Sosat, J.; Tholen, S.; Schilling, O.; Vlachos, A.; Blank, T.; Gomez de Agüero, M.; Macpherson, A.J.; Pearce, E.J.; Prinz, M. , Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab 2021, 33, 2260–2276. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile acids and the gut microbiota: metabolic interactions and impacts on disease. 2023, 21, 236–247. [Google Scholar]
- Mizgier, M.; Jarzabek-Bielecka, G. , The role of diet and probiotics in prevention and treatment of bacterial vaginosis and vulvovaginal candidiasis in adolescent girls and non-pregnant women. 2020, 91, 412–416. [Google Scholar] [CrossRef] [PubMed]
- d’Enfert, C.; Kaune, A.K.; Alaban, L.R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Fróis-Martins, R.; Morelli, M.; Rosati, D. , The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: current knowledge and new perspectives. 2021, 45. [Google Scholar]
- McAleer, J.P.; Nguyen, N.L.; Chen, K.; Kumar, P. , Pulmonary Th17 Antifungal Immunity Is Regulated by the Gut Microbiome. 2016, 197, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xu, D.; Zhou, Z.; Zhu, J.; Wang, D.; Tang, J. , Alterations in the gut microbiota and metabolic profiles coincide with intestinal damage in mice with a bloodborne Candida albicans infection. Microbial pathogenesis 2021, 154, 104826. [Google Scholar] [CrossRef]
- Costa, M.C.; Santos, J.R.; Ribeiro, M.J.; Freitas, G.J.; Bastos, R.W.; Ferreira, G.F.; Miranda, A.S.; Arifa, R.D.; Santos, P.C.; Martins Fdos, S.; Paixão, T.A.; Teixeira, A.L.; Souza, D.G.; Santos, D.A. , The absence of microbiota delays the inflammatory response to Cryptococcus gattii. International journal of medical microbiology : IJMM 2016, 306, 187–95. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R. , The structural unit of melanin in the cell wall of the fungal pathogen Cryptococcus neoformans. 2019, 294, 10471–10489. [Google Scholar] [CrossRef]
- Xue, P.; Hu, G.; Jung, W.H.; Kronstad, J.W. , Metals and the cell surface of Cryptococcus neoformans. Curr Opin Microbiol 2023, 74, 102331. [Google Scholar] [CrossRef]
- Casadevall, A.; Rosas, A.L.; Nosanchuk, J.D. , Melanin and virulence in Cryptococcus neoformans. Curr Opin Microbiol 2000, 3, 354–8. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.P.; Casadevall, A. , Reciprocal modulation of ammonia and melanin production has implications for cryptococcal virulence. 2023, 14, 849. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Rosas, A.L.; Lee, S.C.; Casadevall, A. , Melanisation of Cryptococcus neoformans in human brain tissue. Lancet (London, England) 2000, 355, 2049–50. [Google Scholar] [CrossRef]
- Rosas, A.L.; Nosanchuk, J.D.; Feldmesser, M.; Cox, G.M.; McDade, H.C.; Casadevall, A. , Synthesis of polymerized melanin by Cryptococcus neoformans in infected rodents. Infect Immun 2000, 68, 2845–53. [Google Scholar] [CrossRef]
- Herregodts, P.; Michotte, Y.; Ebinger, G. , Regional differences in the distribution of norepinephrine and epinephrine in human cerebral cortex: a neurochemical study using HPLC and electrochemical detection. Neuroscience letters 1989, 98, 321–6. [Google Scholar] [CrossRef]
- Herregodts, P.; Ebinger, G.; Michotte, Y. , Distribution of monoamines in human brain: evidence for neurochemical heterogeneity in subcortical as well as in cortical areas. Brain research 1991, 542, 300–6. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.; Tom, W.; Costa, J. , Utilization of indole compounds by Cryptococcus neoformans to produce a melanin-like pigment. Journal of clinical microbiology 1983, 18, 1419–1421. [Google Scholar] [CrossRef]
- Panepinto, J.C.; Williamson, P.R. , Intersection of fungal fitness and virulence in Cryptococcus neoformans. FEMS yeast research 2006, 6, 489–498. [Google Scholar] [CrossRef]
- Polacheck, I.; Hearing, V.J.; Kwon-Chung, K.J. , Biochemical studies of phenoloxidase and utilization of catecholamines in Cryptococcus neoformans. J Bacteriol 1982, 150, 1212–20. [Google Scholar] [CrossRef]
- Lee, S.C.; Casadevall, A.; Dickson, D.W. , Immunohistochemical localization of capsular polysaccharide antigen in the central nervous system cells in cryptococcal meningoencephalitis. The American journal of pathology 1996, 148, 1267–74. [Google Scholar] [PubMed]
- Agirman, G.; Yu, K.B. Signaling inflammation across the gut-brain axis. 2021, 374, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. , The Gut-Brain Axis. Annual review of medicine 2022, 73, 439–453. [Google Scholar] [CrossRef]
- Strandwitz, P. , Neurotransmitter modulation by the gut microbiota. Brain research 2018, (Pt B), 128–133. [Google Scholar] [CrossRef]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. , Normal gut microbiota modulates brain development and behavior. Proceedings of the National Academy of Sciences of the United States of America 2011, 108, 3047–52. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; Guzzetta, K.E.; Jaggar, M.; Long-Smith, C.M.; Lyte, J.M.; Martin, J.A.; Molinero-Perez, A.; Moloney, G.; Morelli, E.; Morillas, E.; O’Connor, R.; Cruz-Pereira, J.S.; Peterson, V.L.; Rea, K.; Ritz, N.L.; Sherwin, E.; Spichak, S.; Teichman, E.M.; van de Wouw, M.; Ventura-Silva, A.P.; Wallace-Fitzsimons, S.E.; Hyland, N.; Clarke, G.; Dinan, T.G. , The Microbiota-Gut-Brain Axis. Physiol Rev 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Fan, X.; Deng, H.; Qiu, J.; Ji, H.; Shen, X. , Antibiotics-induced depression in mice via the microbiota-gut-brain axis. Journal of affective disorders 2022, 318, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C.; Frawley, E.R.; Tapscott, T.; Vázquez-Torres, A. , Bacterial Stress Responses during Host Infection. Cell Host Microbe 2016, 20, 133–43. [Google Scholar] [CrossRef]
- Brown, A.J.; Haynes, K.; Quinn, J. , Nitrosative and oxidative stress responses in fungal pathogenicity. Curr Opin Microbiol 2009, 12, 384–91. [Google Scholar] [CrossRef]
- Rutherford, J.C.; Bahn, Y.S.; van den Berg, B.; Heitman, J.; Xue, C. , Nutrient and Stress Sensing in Pathogenic Yeasts. Front Microbiol 2019, 10, 442. [Google Scholar] [CrossRef]
- Choi, J.; Jung, W.H.; Kronstad, J.W. , The cAMP/protein kinase A signaling pathway in pathogenic basidiomycete fungi: Connections with iron homeostasis. Journal of Microbiology 2015, 53, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Caza, M.; Kronstad, J.W. , The cAMP/Protein Kinase a Pathway Regulates Virulence and Adaptation to Host Conditions in Cryptococcus neoformans. Front Cell Infect Microbiol 2019, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- de Vos, W.M.; Tilg, H. Gut microbiome and health: mechanistic insights. 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; Carling, D.; Swann, J.R.; Gibson, G.; Viardot, A.; Morrison, D.; Louise Thomas, E.; Bell, J.D. , The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun 2014, 5, 3611. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. , Short-chain fatty acids in control of body weight and insulin sensitivity. Nature reviews. Endocrinology 2015, 11, 577–91. [Google Scholar] [CrossRef]
- Rastelli, M.; Cani, P.D.; Knauf, C. , The Gut Microbiome Influences Host Endocrine Functions. Endocrine reviews 2019, 40, 1271–1284. [Google Scholar] [CrossRef]
- Cani, P.D.; Jordan, B.F. , Gut microbiota-mediated inflammation in obesity: a link with gastrointestinal cancer. Nature reviews. Gastroenterology & hepatology 2018, 15, 671–682. [Google Scholar]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. , Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Current nutrition reports 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Cani, P.D.; Knauf, C.; Iglesias, M.A.; Drucker, D.J.; Delzenne, N.M.; Burcelin, R. , Improvement of glucose tolerance and hepatic insulin sensitivity by oligofructose requires a functional glucagon-like peptide 1 receptor. Diabetes 2006, 55, 1484–90. [Google Scholar] [CrossRef]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; de Vos, W.M.; Delzenne, N.M.; Schrenzel, J.; Cani, P.D. , Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–86. [Google Scholar] [CrossRef]
Figure 1.
Cryptococcus neoformans melanization in the different brain tissue. The composition of melanin generated during infection exhibits variations contingent upon the catecholamine composition of the tissue. It is highly probable that the in vivo synthesis of melanin pigment arises from the polymerization of a diverse assortment of precursor compounds [37]. Catecholamine distribution in the brain: red domain represents dopamine rich, green domain represents norepinephrine rich, yellow domain represents overlapping between dopamine and norepinephrine.
Figure 1.
Cryptococcus neoformans melanization in the different brain tissue. The composition of melanin generated during infection exhibits variations contingent upon the catecholamine composition of the tissue. It is highly probable that the in vivo synthesis of melanin pigment arises from the polymerization of a diverse assortment of precursor compounds [37]. Catecholamine distribution in the brain: red domain represents dopamine rich, green domain represents norepinephrine rich, yellow domain represents overlapping between dopamine and norepinephrine.
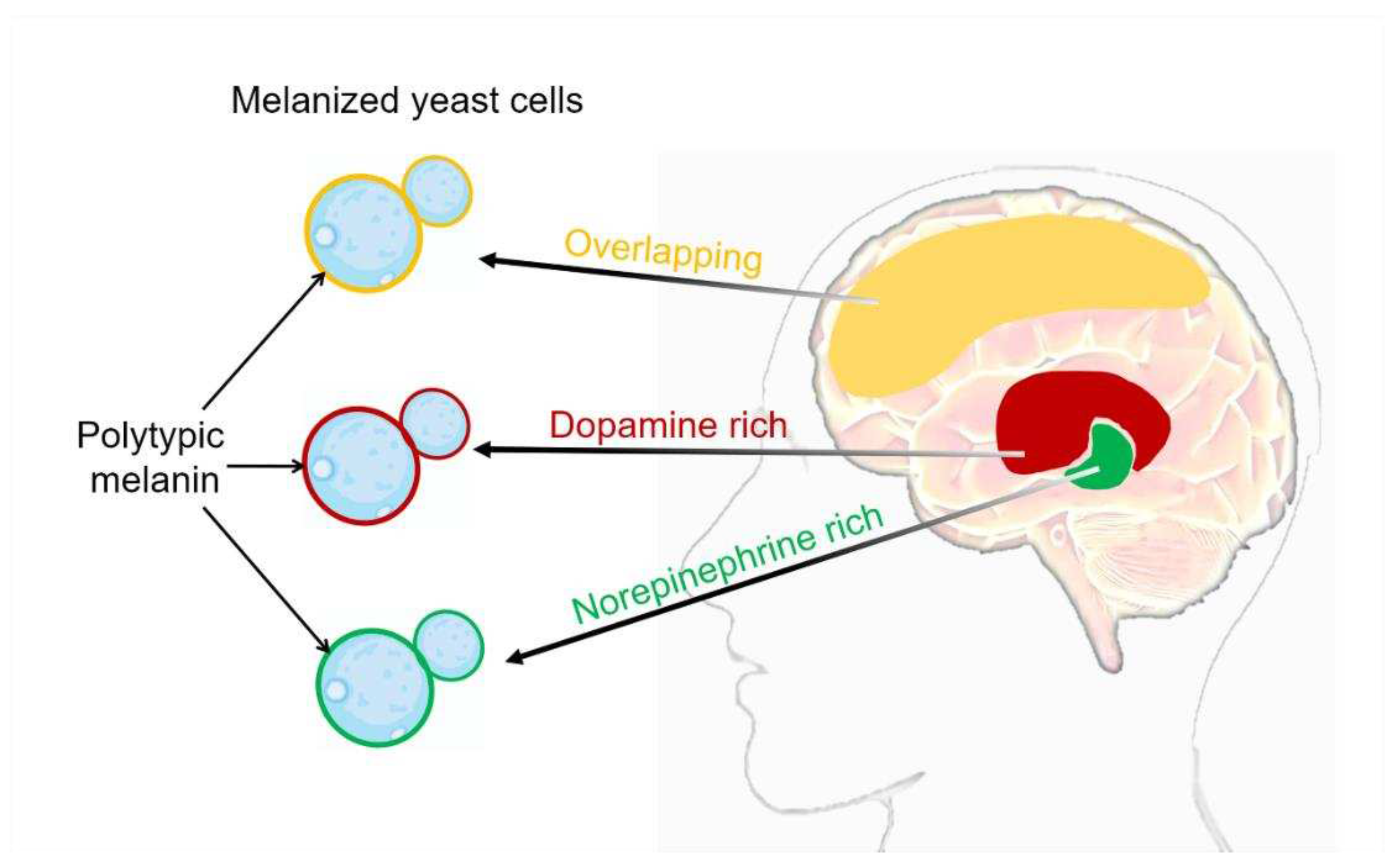
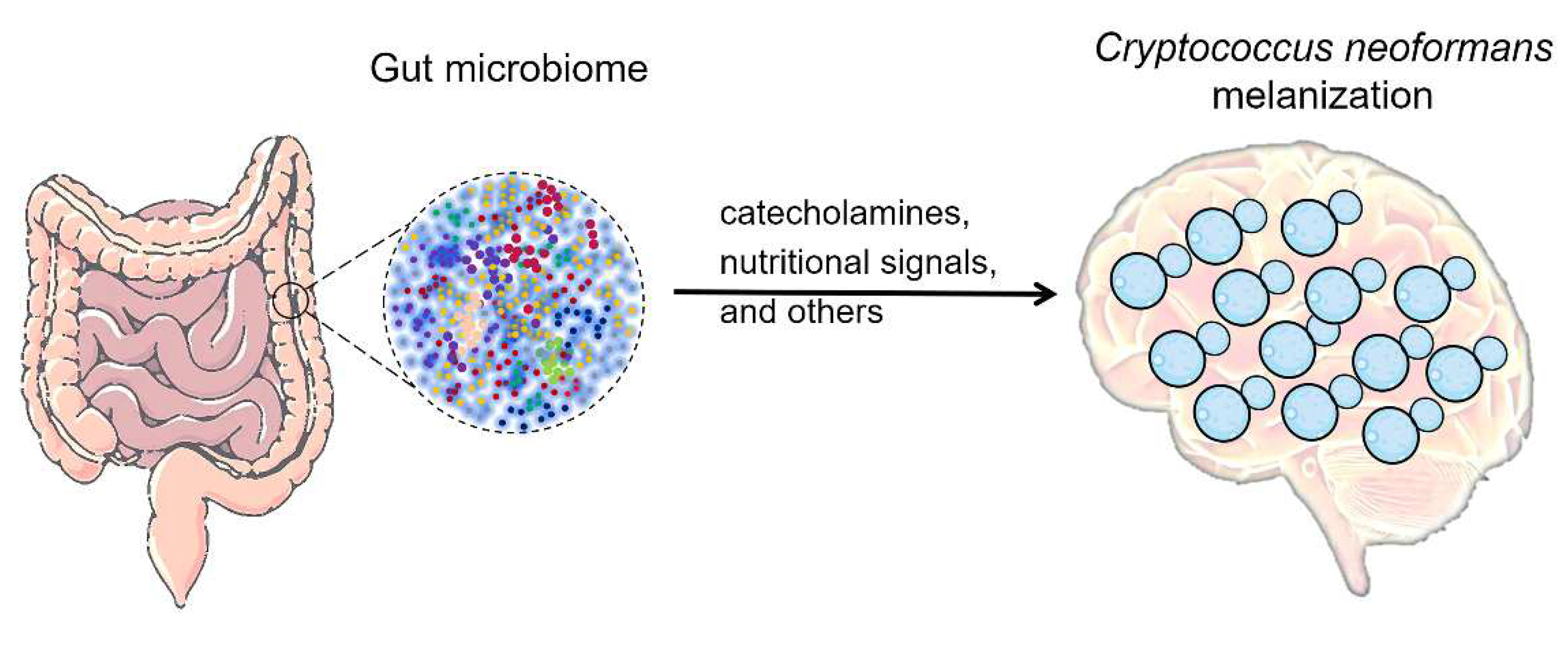
Figure 2.
The potential influence of gut microbiome on the melanization of C. neoformans in the brain. The microbiota-gut-brain axis may regulate the levels of melanin substrates catecholamines (e.g., dopamine or norepinephrine), as well as the nutritional signals (e.g., glucose or amino acids) that modulate melanin formation.
Figure 2.
The potential influence of gut microbiome on the melanization of C. neoformans in the brain. The microbiota-gut-brain axis may regulate the levels of melanin substrates catecholamines (e.g., dopamine or norepinephrine), as well as the nutritional signals (e.g., glucose or amino acids) that modulate melanin formation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



