
Submitted:
10 August 2023
Posted:
11 August 2023
You are already at the latest version
Abstract
Foliage-feeding fall armyworm (Spodoptera frugiperda) and root-feeding western corn rootworm (Diabrotica virgifera virgifera) are maize (Zea mays L.) pests that cause significant yield losses. Jasmonic acid (JA) play pivotal defense role against insects. 12-oxo-phytodienoic acid (12-OPDA) is converted into JA by peroxisome-localized OPDA reductases (OPR). However, little is known about the physiological functions of cytoplasmic OPRs. Here, we show that disruption of ZmOPR2 reduced wound-induced JA production and defense against FAW while accumulating more JA catabolites. Overexpression of ZmOPR2 in Arabidopsis enhanced JA production and defense against beet armyworm (Spodoptera exigua). In addition, lox10opr2 double mutants were more susceptible than either single mutant, suggesting that ZmOPR2 and ZmLOX10 uniquely and additively contributed to defense. In contrast to the defensive role of ZmOPR2 and ZmLOX10 in leaves, single mutants did not display any alteration in root herbivory defense against WCR. Feeding on lox10opr2 double mutants resulted in increased WCR mortality associated with greater herbivory-induced production of insecticidal death acids and ketols. Thus, ZmOPR2 and ZmLOX10 cooperatively inhibit synthesis of these metabolites during herbivory by WCR. We conclude that ZmOPR2 and ZmLOX10 regulate JA-mediated resistance in leaves against FAW while suppressing insecticidal oxylipin synthesis in roots during WCR infestation.
Keywords:
OPDA reductase
; lipoxygenase
; Spodoptera frugiperda
; Diabrotica virgifera virgifera
; ketols
; death acids
; jasmonic acid
; maize
1. Introduction
Maize (Zea mays L.) is a major cereal crop produced globally and a staple crop in many parts of the world. Maize is also a major feed source for livestock worldwide and is used to produce a variety of foods and industrial products. In 2021, U.S. maize production was worth close to $86 billion [1]. Maize production is constantly threatened by abiotic and biotic stressors, including attack of insect herbivores [2]. Fall armyworm (FAW, Spodoptera frugiperda) and western corn rootworm (WCR, Diabrotica virgifera virgifera) are among the world’s most important maize pests [3,4,5,6,7].
FAW causes devastating yield losses annually in maize [8]. Its larvae feed on foliage, stems, and reproductive tissues and maize is its preferred host. Aside from maize, it has been reported to attack >350 plant species including numerous crops, such as maize, sorghum, soybean, cotton, barley, and wheat [9,10]. FAW is native to the American tropics and subtropics and has recently invaded Africa, Asia, and Australia [8,10]. The rapid global spread of FAW has caused significant yield losses and threatened food security especially in African countries where maize is a staple and the most susceptible crop, and annual yield losses caused by FAW are estimated to exceed 9 billion dollars [11,12]. Resistance to FAW and other chewing insects is mediated mainly by jasmonic acid (JA)-mediated signaling [13,14,15].
WCR is a specialist herbivore of maize and the most important maize pest in the USA [16,17]. WCR larvae cause significant injury to maize roots and are difficult to control with insecticides because of their belowground feeding habit. WCR is proposed to have originated in Mexico or Guatemala and spread into North America with the spread of maize cultivation [18]. WCR was first identified as a pest in USA in the early 1900s and quickly expanded its range throughout North America, where it has caused significant economic losses for over a century, especially in the USA Corn Belt [16,18]. WCR was detected in Europe in 1992 and has now invaded 21 European countries [19]. WCR is known as the billion-dollar beetle, because annual costs associated with its control and lost yield exceed 1 billion dollars [20]. Maize resistance mechanisms against WCR are not well understood, and whether JA signaling plays any role in defense responses is unclear [21].
Lipoxygenases (LOXs) in plants incorporate molecular oxygen into linoleic acid (C18:2) and linolenic acid (C18:3) at either carbon position 9 or 13 of the 18-carbon chain and are functionally grouped into 9-LOXs and 13-LOXs [22,23]. Oxidized polyenoic fatty acids are collectively called oxylipins, and most of them are derivatives of seven downstream branches of lipoxygenase (LOX) pathways [22,23,24]. Jasmonates, including JA and its derivatives, are lipid-derived phytohormones that play pivotal roles in insect defense responses [25,26,27]. The initial step of JA biosynthesis occurs in chloroplasts where 13-lipoxygenases (13-LOX) oxidize α-linolenic acid (C18:3), and involves the ensuing enzymatic activities of allene oxide synthase (AOS) and allene oxide cyclase (AOC) to produce 12-oxo-phytodienoic acid (12-OPDA). 12-OPDA is then converted to JA by OPDA reductase (OPR) in the peroxisomes [22,24,28]. The formation of the biologically active jasmonoyl-isoleucine (JA-Ile) is catalyzed by JASMONATE RESISTANT 1 (JAR1) in the cytoplasm [29]. In addition to JA, AOS produces ketols, a group of C18-oxylipin compounds with hormone-like signaling activities that were recently shown to play essential roles in defense against fungal pathogens and chewing insects [30,31,32,33,34,35]. Some ketols, including 9-hydroxy-10-oxo-12(Z),15(Z)-octadecadienoic acid (9,10-KODA), 9-hydroxy-12-oxo-10(E),15(Z)-octadecadienoic acid (9-12-KODA), and 9-hydroxy-12-oxo-10(E)-octadecenoic acid (9-12-KOMA) have been shown to prime plants for defense against pathogens [34,35]. Moreover, ketol 9,10-KODA displayed hormone-like and insecticidal activities in defense against FAW and suppresses FAW larvae growth [31].
Plant OPRs are phylogenetically and functionally classified into OPRI and OPRII subfamilies depending on their substrate catalytic activity [36,37]. OPRII subfamily peroxisome-localized enzymes are JA-producing OPRs because they preferentially catalyze the conversion of the natural JA precursor, cis-(+)-OPDA. The members of the cytosolic OPRI subfamily were found to reduce cis-(-)-OPDA and were long-believed irrelevant in JA biosynthesis [38,39]. So far, several OPRII genes have been characterized. Arabidopsis intronic T-DNA insertional opr3 mutants are deficient in wound-induced JA production and more susceptible to cabbage looper caterpillars [40]. Disruption of both maize ZmOPR7 and ZmOPR8 results in JA deficiency and extreme susceptibility to root-rotting oomycete Pythium spp. and beet armyworm (BAW, Spodoptera exigua) [13]. Little is known about the biochemical and physiological functions of OPRI subfamily enzymes, and it is unknown whether they contribute to insect defense.
Previously, the OPRI subfamily member ZmOPR2 was shown to be localized to the cytoplasm [41] and functions in salicylic acid (SA)-mediated defense responses against maize biotrophic and hemibiotrophic pathogens via suppression of JA production in response to pathogen infection [42]. However, its role in wound-induced JA production and insect defense remains unexplored. This study aimed to partly fill this gap and explore the role, it any, of ZmOPR2 in insect defense in both above- and belowground tissues. We show here that ZmOPR2 contributes to aboveground defense against FAW by regulating wound-induced JA production and catabolism and interacting additively with ZmLOX10 in resistance to herbivory in leaves. Additionally, we show that while single opr2 or lox10 mutants did not affect maize defense against WCR, lox10opr2 double mutants markedly reduced the survivorship of WCR larvae. Metabolite profiling of the lox10opr2 double mutants under WCR herbivory revealed increased accumulations of multiple ketols and death acids known to have insecticidal activities when delivered via an artificial diet. Given these findings, we concluded that ZmOPR2 and ZmLOX10 together negatively regulate the production of the insecticidal death acids and ketols, which are likely a part of host defense strategy against herbivory by WCR. Also, our data suggested that JA may not have as significant function in herbivory defense in maize roots as it has in the leaves.
2. Materials and Methods
2.1. Plant materials
Mutator-insertional opr2-1 (PV 03 80 A-05), and opr2-3 (mu1079063::Mu; stock ID UFMu-08953) alleles were backcrossed to B73 inbred line and advanced to BC7 and BC1 stage, respectively. Mutants and their corresponding near-isogenic wild-type were identified from F2 segregating populations by PCR genotyping using Mu-terminal inverted repeat-specific and gene-specific primers as described previously [42]. opr2-1 and opr2-3 are knock-out alleles and were described in detail in Huang et al. [42]. Identification of the lox10-3 allele was previously described by Christensen et al. [14]. The double mutant lox10opr2 was generated by crossing single mutants lox10-3 and opr2-1 at the BC7 genetic stage [42]. Maize seeds were grown in conical pots (20.5 by 4 cm) filled with commercial potting mix (Jolly Gardener Pro Line C/20 potting mix) on light shelves at room temperature (22 to 24°C) with a 16-h light period. For generating Arabidopsis ZmOPR2-overexpression lines, Gateway vectors carrying the ZmOPR2 gene were transformed into rdr6-11 background as described in Tolley et al. [41] and grown under the same condition. Two T4 homozygous ZmOPR2-GFP overexpression lines, #32-9 and #39-1, and in this study were named OE1 and OE2 lines, respectively, and untransformed rdr6-11 plants was used as the WT controls.
2.2. Fall armyworm bioassays
The laboratory strain of FAW (Spodoptera frugiperda) was purchased from Benzon Research (Carlisle, PA, USA). The eggs laid on paper towels were hatched and reared on an artificial diet purchased from Southland Product Inc. (Arkansas, USA). To evaluate insect resistance and leaf damaged area, the fourth leaves of maize plants at the V4 stage were individually caged and infested with one 3rd-instar FAW larva per spot for approximately one hour and then moved toward the base afterward and so on. Leaves were scanned, and eaten areas were measured with ImageJ software (https://imagej.nih.gov/ij/). Larval weight gain experiments were performed by caging six FAW neonates with individual maize plants at the V3 developmental stage and allowed them to move and feed on the plant freely for 7 days. FAW larvae were removed from the plants, and total weight was determined 7 days post-infestation.
2.3. Beet armyworm bioassay
BAW (Spodoptera exigua) eggs were purchased from Benzon Research (Carlisle, PA, USA). After hatching, neonate larvae were reared on an artificial diet purchased from Southland Product Inc. (Arkansas, USA) for 5 days. Third-instar larvae were starved overnight and the initial weight was measured before transferring to a 0.8% agar plate. Mature rosette leaves were cut from 4-week-old Arabidopsis plants and placed into the plate with 4 BAW larvae per plate. Fresh leaves were provided two days after initial feeding and the experiment was terminated on day 4, and the larval weight gain was determined after subtracting the initial weight and normalized by the number of recovered larvae.
2.4. WCR bioassays
WCR (Diabrotica virgifera virgifera) eggs were provided by USDA-ARS North Central Agriculture Research Laboratory (Brookings, SD, USA) and stored at 4°C until use. The diapausing Tent strain was used for this study. Prior to use, eggs were washed and incubated on moist filter paper for 12 days at 27°C. Neonates were used within 24 hrs after the first emergence. For the WCR assay, maize seeds were grown in conical pots (20.5 by 4 cm) filled with commercial potting mix (Jolly Gardener Pro Line C/25 potting mix) in a climate-controlled, insect-free growth room under artificial full-spectrum growing lights (27°C, 50% Relative Humidity, 12:12 hour light:dark cycle, PPFD 450 µmol m-2 s-1). Experiments were performed in the same environmental conditions as the plants were grown. Plants were watered as needed and before application of WCR neonates. 2 to 3-week-old maize plants that had 3 fully expanded true leaves were infested with 10 WCR neonates. WCR larvae were removed from the soil and total weight gain was determined 10 days post-infestation. Recovery of WCR larvae is a proxy measure of survivorship. WCR head capsules were then imaged and measured using ImageJ to quantify larval instar stage (ImageJ, National Institute of Health, Bethesda, United States). The soil was then removed from root tissue and root fresh weight was measured. Root ratio was analyzed as larvae damaged over undamaged root mass to determine host plant tolerance as compared to WT.
2.5. Oxylipin profiling of wounded leaf tissue and WCR-infested root tissues
For wounding treatment in leaves, the third fully expanded leaves of seedlings at the V3 developmental stage were wounded seven times using a hemostat, with three wound sites on one side and four on the other side of the midvein and wound sites approximately 1 cm apart, in the middle portion of the leaf. For Arabidopsis, rosette leaves were wounded twice using forceps across the midvein. The wounded regions were then harvested in 2 mL screw-cap Fast-Prep tubes (Qbiogene, Carlsbad, CA, U. S. A.) in liquid nitrogen and stored in a -80 °C freezer. For WCR-infested root tissues, maize root tissues were washed at 0 (CTRL), 8, 24, and 48 hours after being exposed to WCR neonates and harvested in liquid nitrogen, and stored in a -80 °C freezer. Phytohormone and oxylipin extraction and profiling of wounded leaf tissue were performed by using a LC-MS/MS as described in Huang et al. [42].
2.6. Statistical Methods
Statistical analyses were performed using the software program R (R version 4.2.2, R Core Team, 2023), JMP Pro 17 (SAS Institute Inc.), and GraphPad Prism 10 (GraphPad Software, Boston, Massachusetts, USA). FAW leaf area consumed and larval weight gain were analyzed using Student’s t-test (Figure 1A and B). BAW larval weight gain was analyzed using ANOVA followed by Dunnett’s test (Figure 1C). Two-way ANOVAs, with genotype and time as the two factors, were used for leaf metabolite analysis of WT-mutant comparisons followed by Sidak’s multiple comparisons test for comparisons within time (Figure 2, 3, and 4). FAW larval weight gain and leaf area consumed (Figure 5) were compared using ANOVA followed by Tukey’s honestly significant differences (HSD) test. Root and shoot ratios, WCR larvae recovery, and WCR mass gain were analyzed using ANOVA followed by Tukey’s multiple comparison test (Figure 5A, B, and D). Larval instars were compared using ANOVA, followed by Dunnett’s multiple comparison test (Figure 5C). Root hormone analysis at 24 hpi were compared using Student’s t-test (Figure 7).
3. Results
3.1. ZmOPR2 promotes defense against FAW herbivory
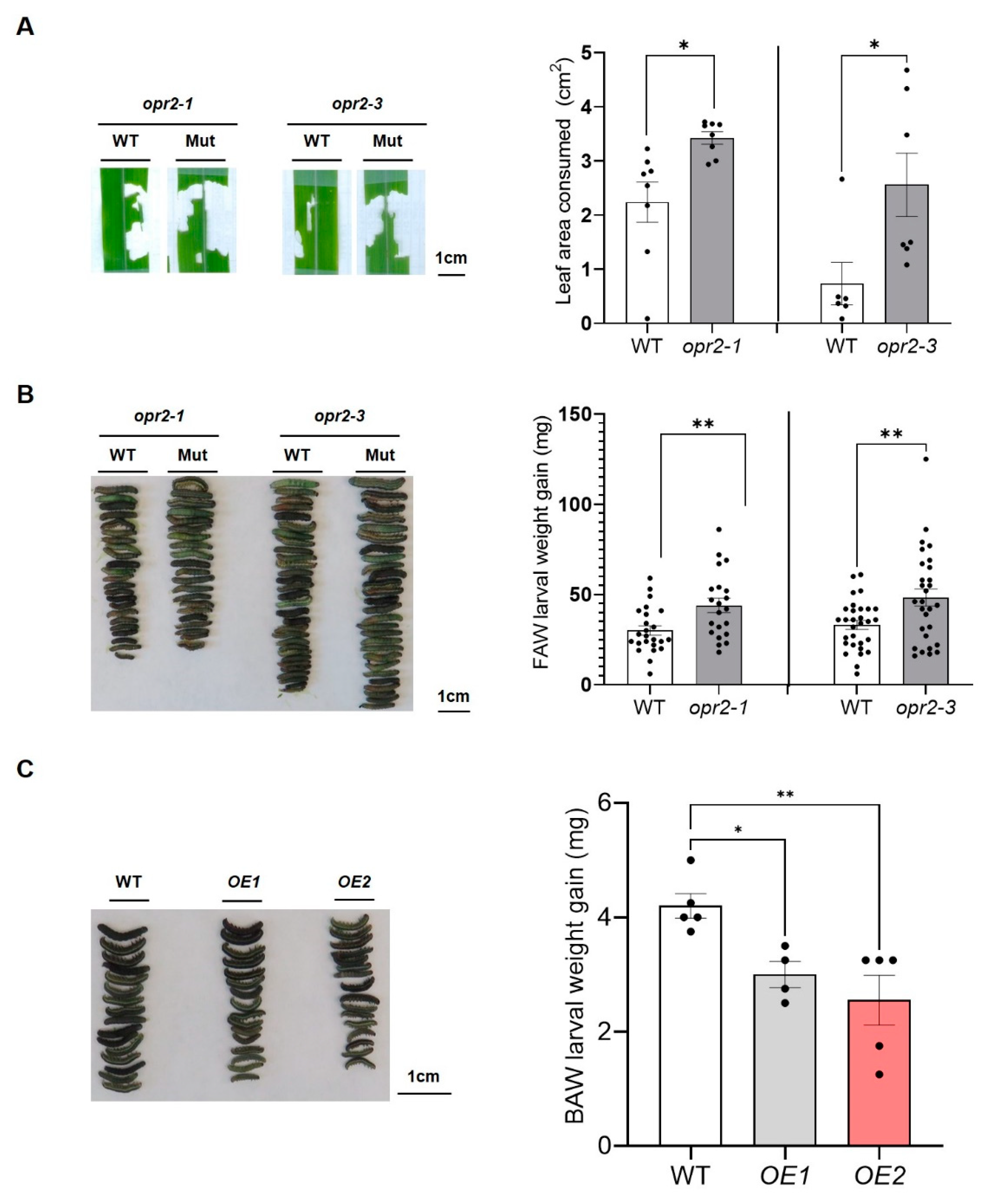
In a previous study, we reported that ZmOPR2 functions in SA-mediated defense responses against (hemi)biotrophic pathogens by suppressing the activity of JA-producing ZmLOX10 [42]. In this study, we tested whether ZmOPR2 is relevant to insect defense by caging single 3rd-instar FAW larvae on leaves of opr2-1 and opr2-3 mutants and their respective WTs at V4 developmental stage maize. After 6 hours, FAW larvae consumed significantly more leaf tissue in opr2-1 and opr2-3 mutant seedlings (~1.5- and 3.5-fold larger, respectively) compared to consumption in WT seedlings, suggesting that opr2 mutants are more susceptible to FAW (Figure 1A). Consistent with this result, FAW larvae weighed significantly more (>40%) after feeding on opr2-1 and opr2-3 mutants for 7 days than those fed on WT plants (Figure 1B). These results suggested that ZmOPR2 is involved in defense against aboveground herbivory by chewing insects. Transgenic Arabidopsis plants overexpressing ZmOPR2 were generated to corroborate whether ZmOPR2 enhances insect defense [41]. Third-instar BAW larvae were offered mature rosette leaves from ZmOPR2-OE Arabidopsis lines or control plants for 4 days. After 4 days, BAW larvae gained significantly less weight (29-39% reduction) after feeding on ZmOPR2-OE line leaves compared to larvae feeding on the WT leaves, suggesting that overexpression of ZmOPR2 in Arabidopsis enhanced defense against BAW (Figure 1C). Together, these results showed that ZmOPR2 promotes defense in above-ground tissues against chewing insects, such as FAW and BAW.
Figure 1.
ZmOPR2 promotes insect resistance. (A) Fourth leaves of opr2-1 and its WT at V4 stage were caged and infested with one 3rd-instar FAW larva per leaf. The leaves were scanned after experiment and eaten leaf areas were measured using ImageJ. (B) opr2-1 and opr2-3 mutants and their respective WT plants at V3 stage were caged with 6 FAW neonates per plant. FAW larvae were removed from the plants and larval weight was analyzed 7 days post infestation. Bars are means ± SE (n≥25 FAW larvae removed from 5 different maize plants). Asterisks represent statistically significant differences between WT and mutant (Student’s t-test, *P < 0.05, **P < 0.01). (C) BAW larval weight gain after feeding on ZmOPR2-OE lines and WT for 4 days. Bars are means ± SE (n≥4 replicates of BAW larvae feeding on leaves from different Arabidopsis plants). Asterisks represent statistically significant differences (Dunnett’s test, *P < 0.05, **P < 0.01).
Figure 1.
ZmOPR2 promotes insect resistance. (A) Fourth leaves of opr2-1 and its WT at V4 stage were caged and infested with one 3rd-instar FAW larva per leaf. The leaves were scanned after experiment and eaten leaf areas were measured using ImageJ. (B) opr2-1 and opr2-3 mutants and their respective WT plants at V3 stage were caged with 6 FAW neonates per plant. FAW larvae were removed from the plants and larval weight was analyzed 7 days post infestation. Bars are means ± SE (n≥25 FAW larvae removed from 5 different maize plants). Asterisks represent statistically significant differences between WT and mutant (Student’s t-test, *P < 0.05, **P < 0.01). (C) BAW larval weight gain after feeding on ZmOPR2-OE lines and WT for 4 days. Bars are means ± SE (n≥4 replicates of BAW larvae feeding on leaves from different Arabidopsis plants). Asterisks represent statistically significant differences (Dunnett’s test, *P < 0.05, **P < 0.01).
3.2. ZmOPR2 enhances wound-induced JA accumulation
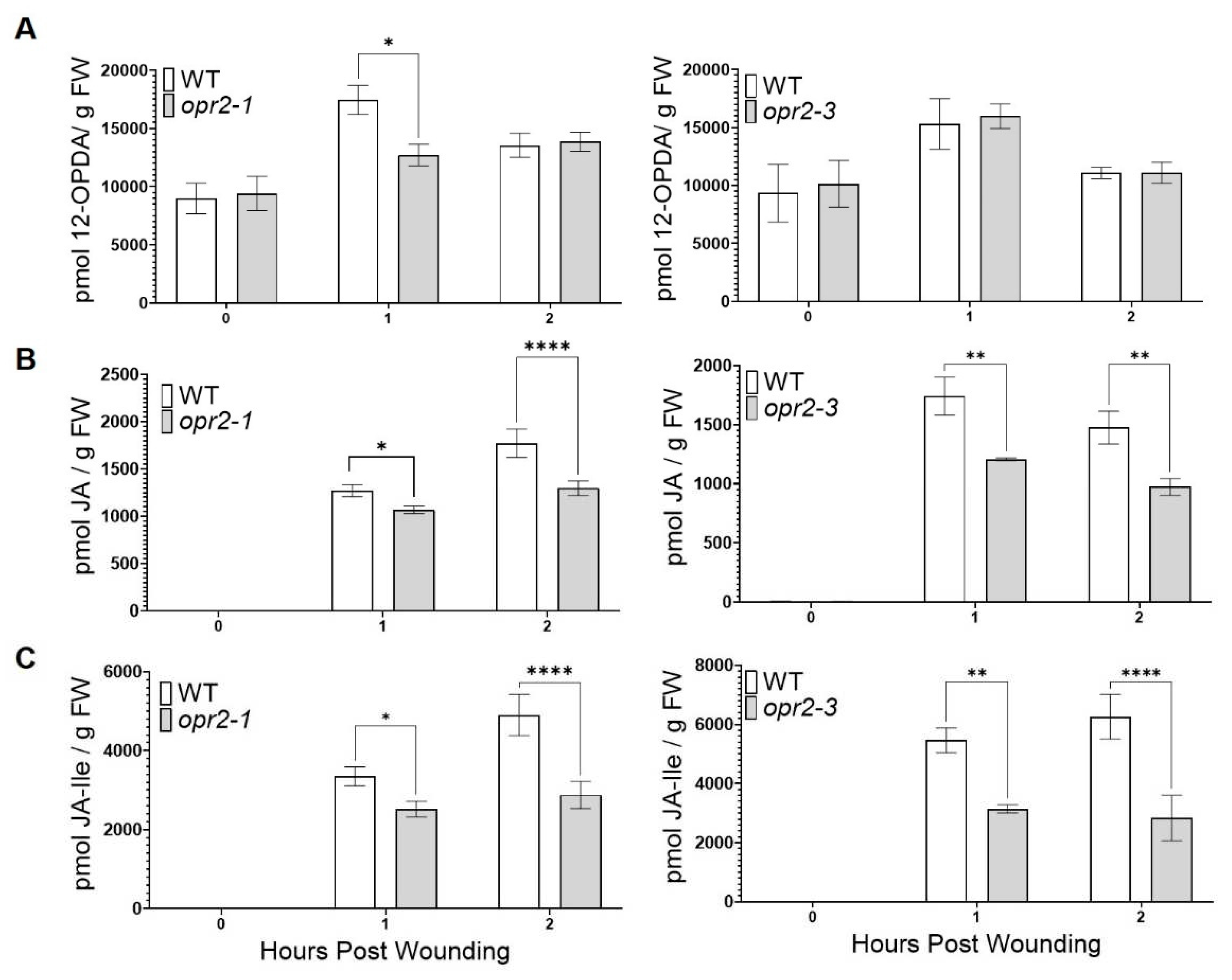
A previous study showed that disruption of ZmOPR2 in maize resulted in reduced resistance to hemibiotrophic Colletotrichum graminicola associated with greater infection-induced JA accumulation [42]. Surprisingly, in the present study, we found that opr2 mutants were more susceptible to FAW, which prompted us to test whether ZmOPR2 plays a role in wound-induced JA production. To this end, leaves of opr2-1 and opr2-3 mutants and WT were mechanically wounded and accumulation of jasmonates was measured at 0, 1, and 2 hours post wounding (hpw). We found that opr2-1 and opr2-3 mutants accumulated significantly lower levels of wound-induced JA at 1 and 2 hpw (16 and 27% reduction in opr2-1, 31 and 34% reduction in opr2-3, respectively) (Figure 2B) and JA-Ile (25 and 41% reduction in opr2-1, 42 and 55% reduction in opr2-3) (Figure 2C). Both alleles accumulated similar amounts of 12-ODPA compared to WT (Figure 2A).
Figure 2.
opr2 mutants accumulated lower levels of wound-induced JA and JA-Ile. Contents of (A) 12-OPDA, (B) JA and (C) JA-Ile were measured at 0, 1, and 2 hours post wounding (n≥ 4 maize plants per genotype). Asterisks represent statistically significant differences between WT and mutant (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.0001).
Figure 2.
opr2 mutants accumulated lower levels of wound-induced JA and JA-Ile. Contents of (A) 12-OPDA, (B) JA and (C) JA-Ile were measured at 0, 1, and 2 hours post wounding (n≥ 4 maize plants per genotype). Asterisks represent statistically significant differences between WT and mutant (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.0001).
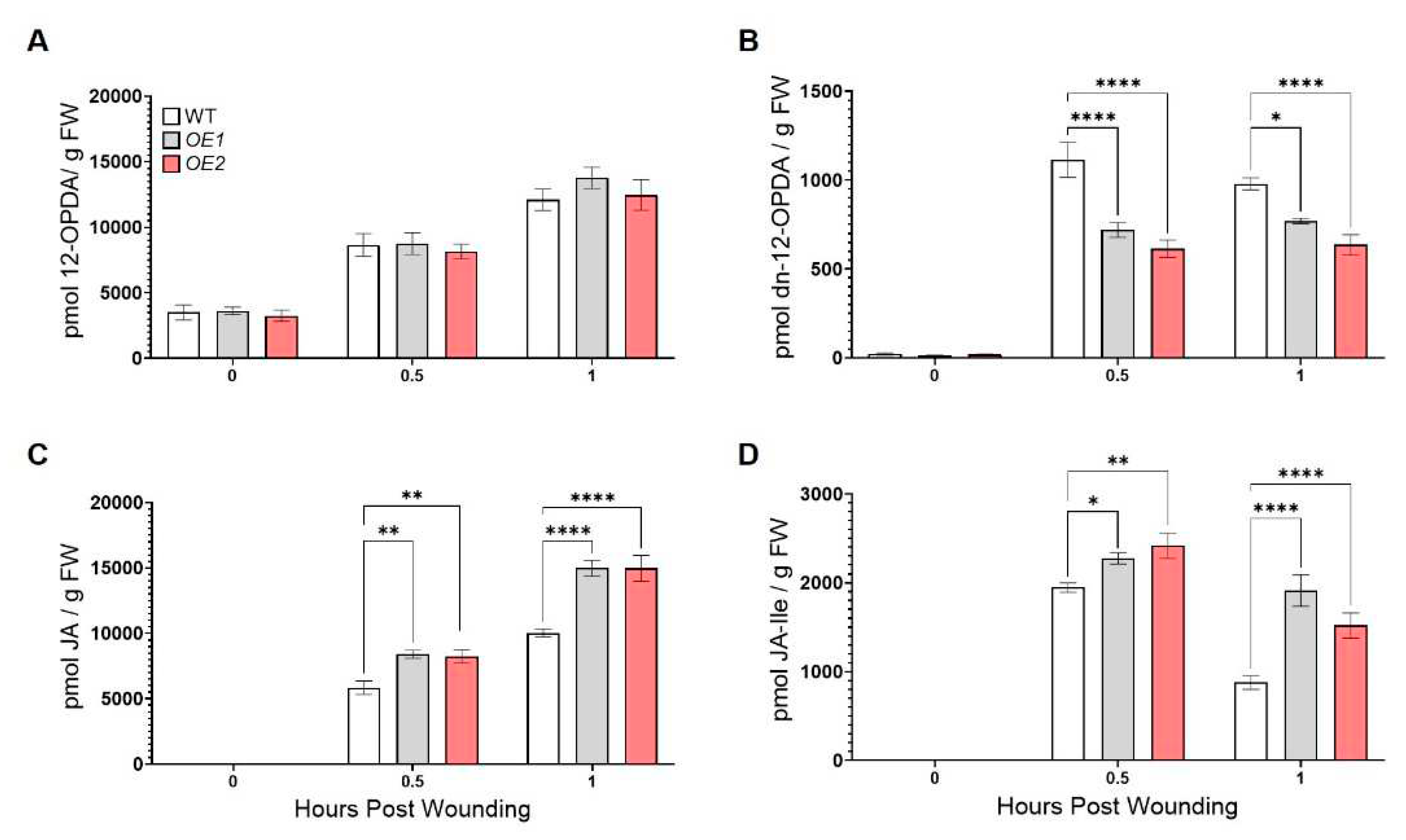
In support of the notion that ZmOPR2 promotes wound-induced JA synthesis, we found that Arabidopsis ZmOPR2-OE lines accumulated significantly higher levels of JA at 0.5 and 1 hpw (42% and 49% increase, respectively) (Figure 3C) and JA-Ile (~20% and >70% increase) (Figure 3D) while accumulating less dn-12-OPDA (~40% and >20% reduction) (Figure 3B) compared to WT. We found no significant difference in the levels of 12-OPDA (Figure 3A).
Figure 3.
ZmOPR2-OE lines accumulated higher levels of wound-induced JA and JA-Ile. Contents of (A) 12-OPDA, (B) dn-12-OPDA, (C) JA and (D) JA-Ile were measured at 0, 0.5, and 1 hours post wounding Bars are means ± SE (n≥ 5 plants per genotype). Asterisks represent statistically significant differences (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.01).
Figure 3.
ZmOPR2-OE lines accumulated higher levels of wound-induced JA and JA-Ile. Contents of (A) 12-OPDA, (B) dn-12-OPDA, (C) JA and (D) JA-Ile were measured at 0, 0.5, and 1 hours post wounding Bars are means ± SE (n≥ 5 plants per genotype). Asterisks represent statistically significant differences (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.01).
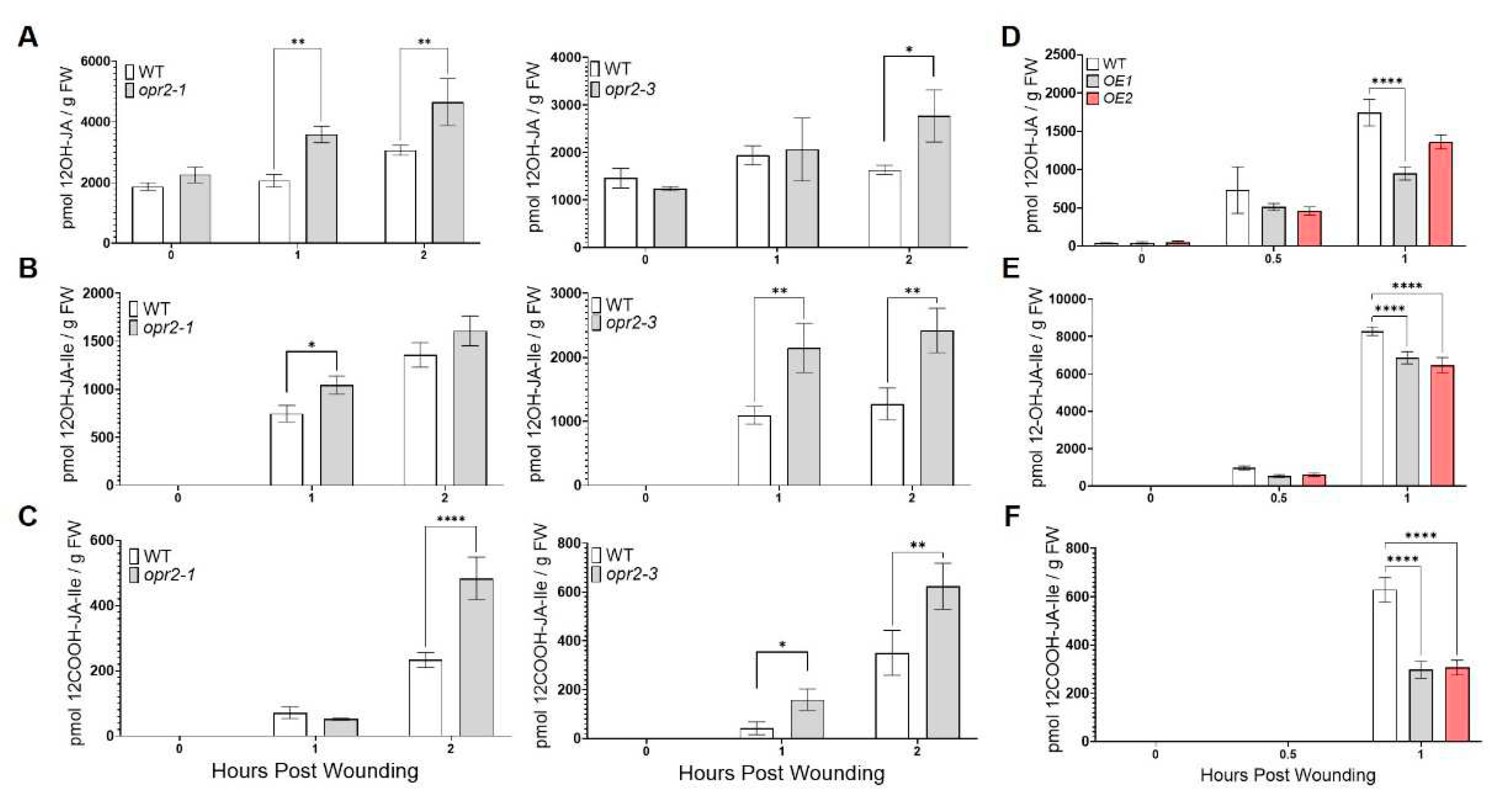
Accumulating evidence suggests that JA turnover and homeostasis is tightly regulated by JA catabolism under stresses-specific manner [43]. Therefore, we also measured the levels of JA catabolites, including 12-hydroxy-jasmonic acid (12OH-JA), 12-hydroxy-jasmonyl-L-isoleucine (12OH-JA-Ile), and 12-carboxy-jasmonoyl-L-isoleucine (12COOH-JA-Ile) in leaves upon wounding. Interestingly, we found that opr2-1 and opr2-3 mutants accumulated significantly higher levels of JA catabolites at 1 and 2 hpw, including 12-OH-JA (73 and 51% increase in opr2-1, 6 and 69% increase in opr2-3, respectively), 12-OH-JA-Ile (40 and 18% increase in opr2-1, 96 and 90% increase in opr2-3), and 12-COOH-JA-Ile (28% reduction and 107% increase in opr2-1, 280 and 78% increase in opr2-3) compared to WT (Figure 4A-C). These results suggest that ZmOPR2 not only regulates biosynthesis of biologically active JA but also JA catabolism after wounding. To corroborate these results, we also measured levels of JA catabolites in mechanically wounded Arabidopsis ZmOPR2-OE lines at 0.5 and 1 hpw. We found that at 0.5 and 1 hpw after wounding significantly lower levels of 12-OH-JA (30-37% and 22-46 % reduction, respectively), 12-OH-JA-Ile (38-45% and 17-22% reduction), and 12-COOH-JA-Ile (51-53 % reduction at 1 hpw) were accumulated in ZmOPR2-OE lines compared to WT (Figure 4D-F). Together, these results showed that ZmOPR2 enhances wound-induced JA and JA-Ile biosynthesis and regulates the catabolism of jasmonates, the first report of such a specific effect of any plant OPR on JA catabolism.
Figure 4.
ZmOPR2 regulates accumulation of JA catabolites in response to wounding. Contents of (A) 12OH-JA, (B) 12OH-JA-Ile, and (C) 12COOH-JA-Ile in opr2-1 and opr2-3 mutants and WT were measured at 0, 1, and 2 hours post wounding. Accumulation of (D) 12OH-JA, (E) 12OH-JA-Ile, and (F) 12COOH-JA-Ile were measured in ZmOPR2-OE lines and control plants at 0, 0.5, and 1 hours post wounding. Bars are means ± SE (n≥ 4 plants per genotype). Asterisks represent statistically significant differences (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.0001).
Figure 4.
ZmOPR2 regulates accumulation of JA catabolites in response to wounding. Contents of (A) 12OH-JA, (B) 12OH-JA-Ile, and (C) 12COOH-JA-Ile in opr2-1 and opr2-3 mutants and WT were measured at 0, 1, and 2 hours post wounding. Accumulation of (D) 12OH-JA, (E) 12OH-JA-Ile, and (F) 12COOH-JA-Ile were measured in ZmOPR2-OE lines and control plants at 0, 0.5, and 1 hours post wounding. Bars are means ± SE (n≥ 4 plants per genotype). Asterisks represent statistically significant differences (Sidak’s multiple comparisons test, *P < 0.05, **P < 0.01, ****P < 0.0001).
3.3. ZmOPR2 and ZmLOX10 have additive effect on defense against FAW
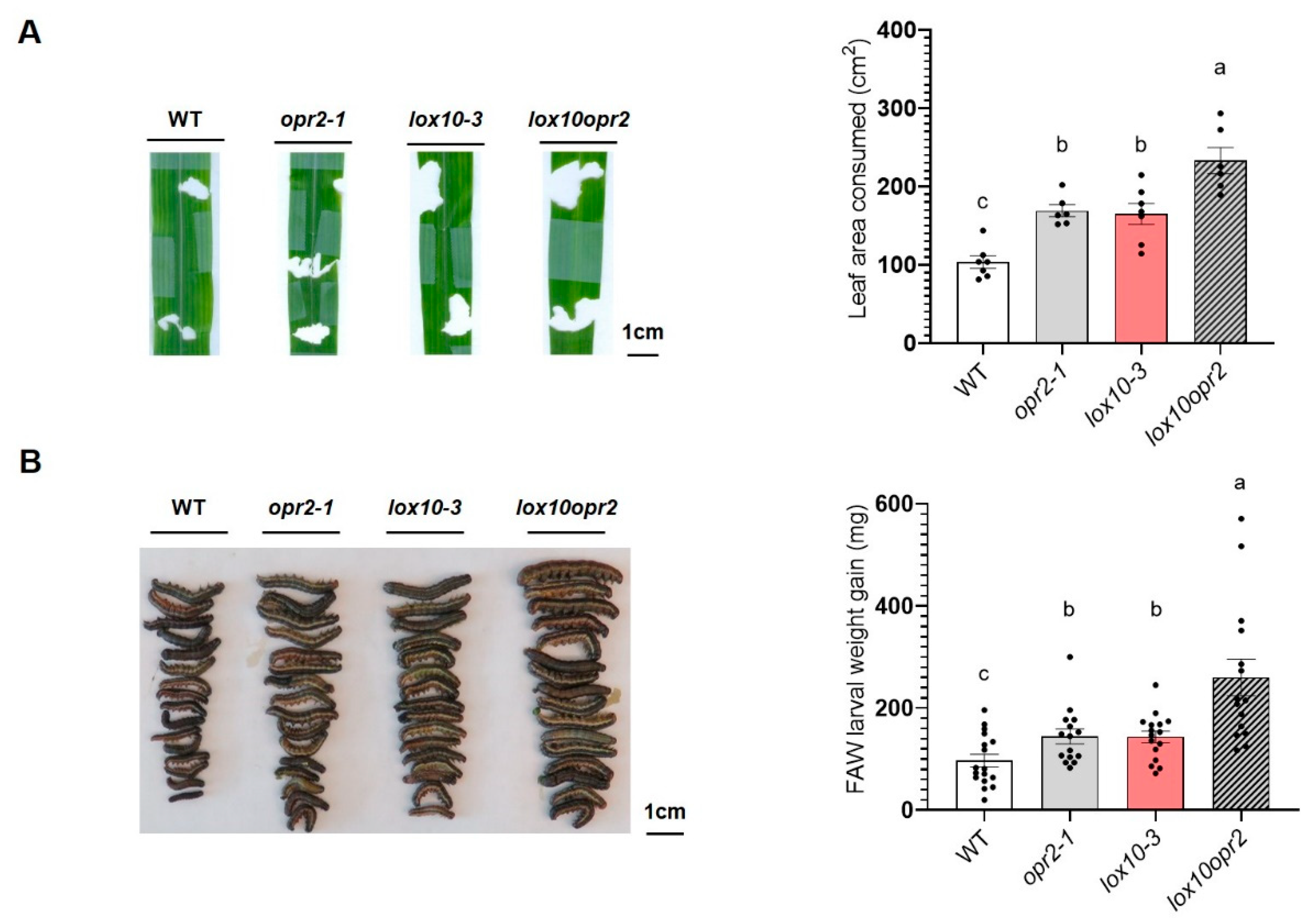
Previous reports showed that ZmLOX10 plays an important role in plant modulation of insect defense by taking part in the biosynthesis of green leaf volatiles (GLV) and JA in response to wounding and insect herbivory [14,44]. Although mutation of ZmOPR2 increased the production of ZmLOX10-mediated GLVs and accumulation of JA in response to pathogen infection [42], similar to lox10 mutants, opr2 mutants in the present study accumulated less wound-induced JA and JA-Ile and were more susceptible to a chewing insect, FAW. To explore the relative contributions of ZmOPR2 and ZmLOX10 to defense against herbivory, we performed FAW bioassays on single opr2-1 and lox10-3 mutants, lox10opr2 double mutants and corresponding WT controls. We found that while the 3rd-instar FAW larvae consumed 64% and 60% more leaf tissue in single opr2-1 and lox10-3 mutants, respectively, compared to WT, FAW larvae consumed twice as much (126%) lox10opr2 double mutant tissue (Figure 5A). Correspondingly, FAW larvae gained more weight after feeding on single opr2-1 and lox10-3 mutants (48% increase) compared to those fed on WT, and FAW larvae that fed on lox10opr2 double mutants showed the greatest increase in weight (167%) relative to WT (Figure 5B). Together, these results suggested that ZmOPR2 and ZmLOX10 acted additively in defense modulation against the chewing insect FAW, and that the contribution of ZmOPR2 to defense against chewing insects extends beyond its role in JA synthesis.
Figure 5.
ZmOPR2 and ZmLOX10 act synergistically in regulation of defense against FAW. (A) Average leaf consumed area of opr2-1, lox10-3 and lox10opr2 mutants and their WT after FAW infestation. Eaten leaf area is the sum of all the damaged areas on one leaf. Bars are means ± SE (n ≥ 6 maize plants). (B) Average FAW larval weight gain after feeding on opr2-1, lox10-3 and lox10opr2 mutants and WT for 7 days. Bars are means ± SE (n≥ 20 larvae from 5 different plants). Different letters indicate statistically significant differences on log-transformed data (Tukey’s HSD test, p < 0.05).
Figure 5.
ZmOPR2 and ZmLOX10 act synergistically in regulation of defense against FAW. (A) Average leaf consumed area of opr2-1, lox10-3 and lox10opr2 mutants and their WT after FAW infestation. Eaten leaf area is the sum of all the damaged areas on one leaf. Bars are means ± SE (n ≥ 6 maize plants). (B) Average FAW larval weight gain after feeding on opr2-1, lox10-3 and lox10opr2 mutants and WT for 7 days. Bars are means ± SE (n≥ 20 larvae from 5 different plants). Different letters indicate statistically significant differences on log-transformed data (Tukey’s HSD test, p < 0.05).
3.4. ZmOPR2 and ZmLOX10 negatively regulate production of insecticidal oxylipins during root herbivory by WCR
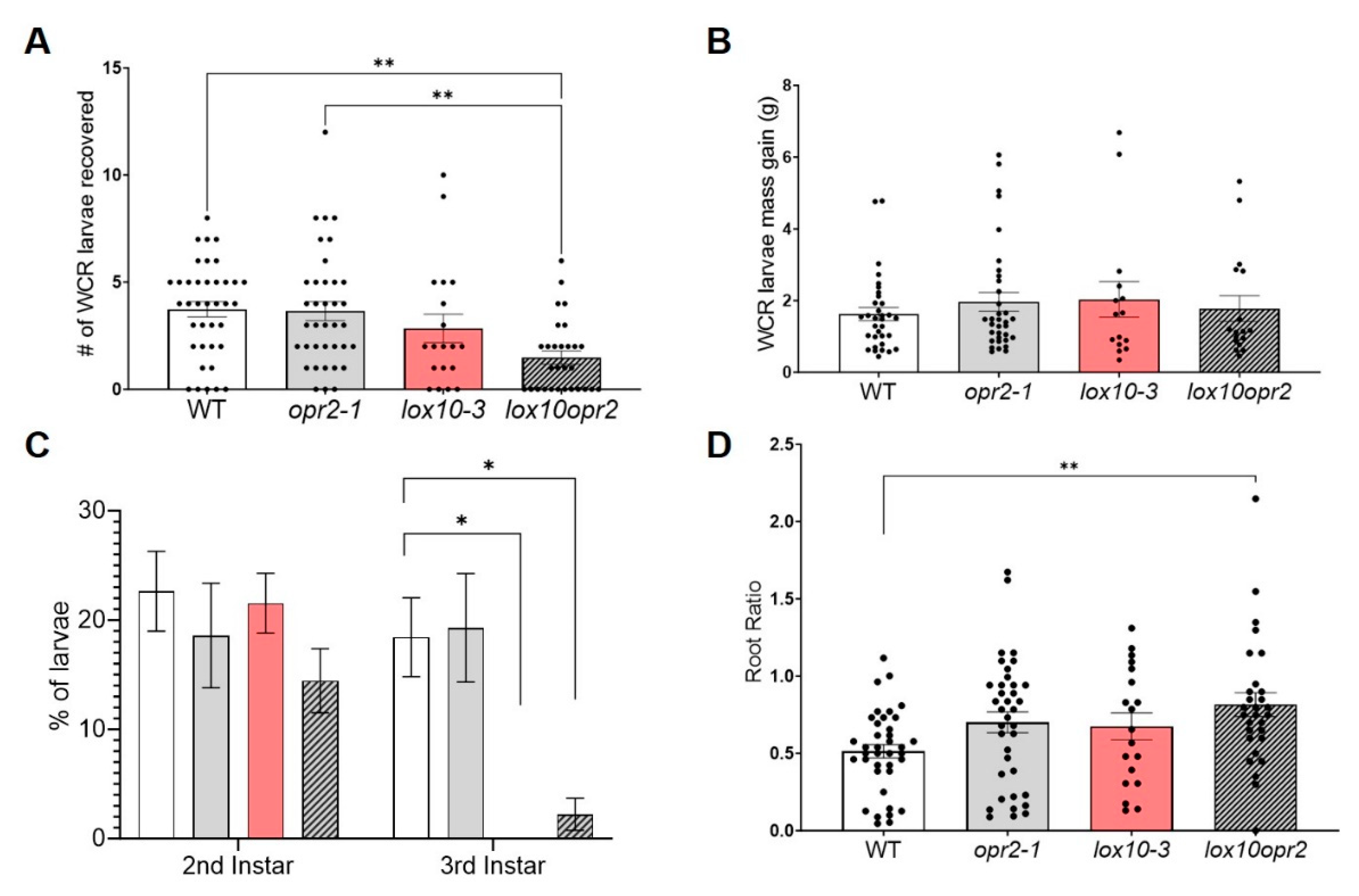
Multiple line of evidence, including the current study, confirmed that JA plays an essential role in regulating plant defense responses against chewing insects in aboveground tissues [13,45]. However, whether JA is required for WCR defense in roots remains largely unknown. Here, we tested whether ZmOPR2 and ZmLOX10 contribute to JA synthesis and defense against chewing insects in roots by assaying corresponding single and double mutants. We found that survivorship of WCR larvae feeding on roots of lox10opr2 mutants was lower in comparison to the survivorship of larvae feeding on roots of single opr2-1 or WT plants (ANOVA, F3,119= 6.11, P < 0.001) (Figure 6A) but showed no difference in comparison to single lox10-3 mutant. However, no significant difference in larval weights were found after larvae fed on opr2-1, lox10-3, or lox10opr2 mutants compared to larvae fed on WT (ANOVA, F3,95= 0.427, P = 0.733) (Figure 6B). Additionally, WCR larvae feeding on lox10-3 or lox10opr2 mutants developed slower compared to those feeding on opr2-1 mutant and WT plants (ANOVA, F3,45= 4.428, P = 0.008) (Figure 6C). Finally, lox10opr2 mutans showed greater tolerance (ability to recover tissues lost to herbivory) compared to WT, opr2-1, and lox10-3 mutants (ANOVA, F3,119= 3.907, P = 0.011), as indicated by their greater root mass ratio (ratio of root mass of plants exposed to WCR relative to root mass of plants not exposed to WCR) (Figure 6D). Together, these results showed that together ZmOPR2 and ZmLOX10 downregulated both direct defense and tolerance against the chewing insect WCR in maize roots.
Figure 6.
Mutation of both ZmOPR2 and ZmLOX10 suppressed WCR larvae survival and larvae development. (A) Larvae recovery and (B) Larvae mass were recorded 10 days post infestation and (C) the developmental stages of the larvae recovered were determined by measuring larvae head capsules using ImageJ (D) Root ratio was analyzed as larvae damaged over undamaged root mass to determine tolerance after compared to WT. Bars are means ± SE. Asterisks represent statistically significant differences (A,B,D - Tukey HSD and C – Dunnett’s Test , *P < 0.05, **P < 0.01).
Figure 6.
Mutation of both ZmOPR2 and ZmLOX10 suppressed WCR larvae survival and larvae development. (A) Larvae recovery and (B) Larvae mass were recorded 10 days post infestation and (C) the developmental stages of the larvae recovered were determined by measuring larvae head capsules using ImageJ (D) Root ratio was analyzed as larvae damaged over undamaged root mass to determine tolerance after compared to WT. Bars are means ± SE. Asterisks represent statistically significant differences (A,B,D - Tukey HSD and C – Dunnett’s Test , *P < 0.05, **P < 0.01).
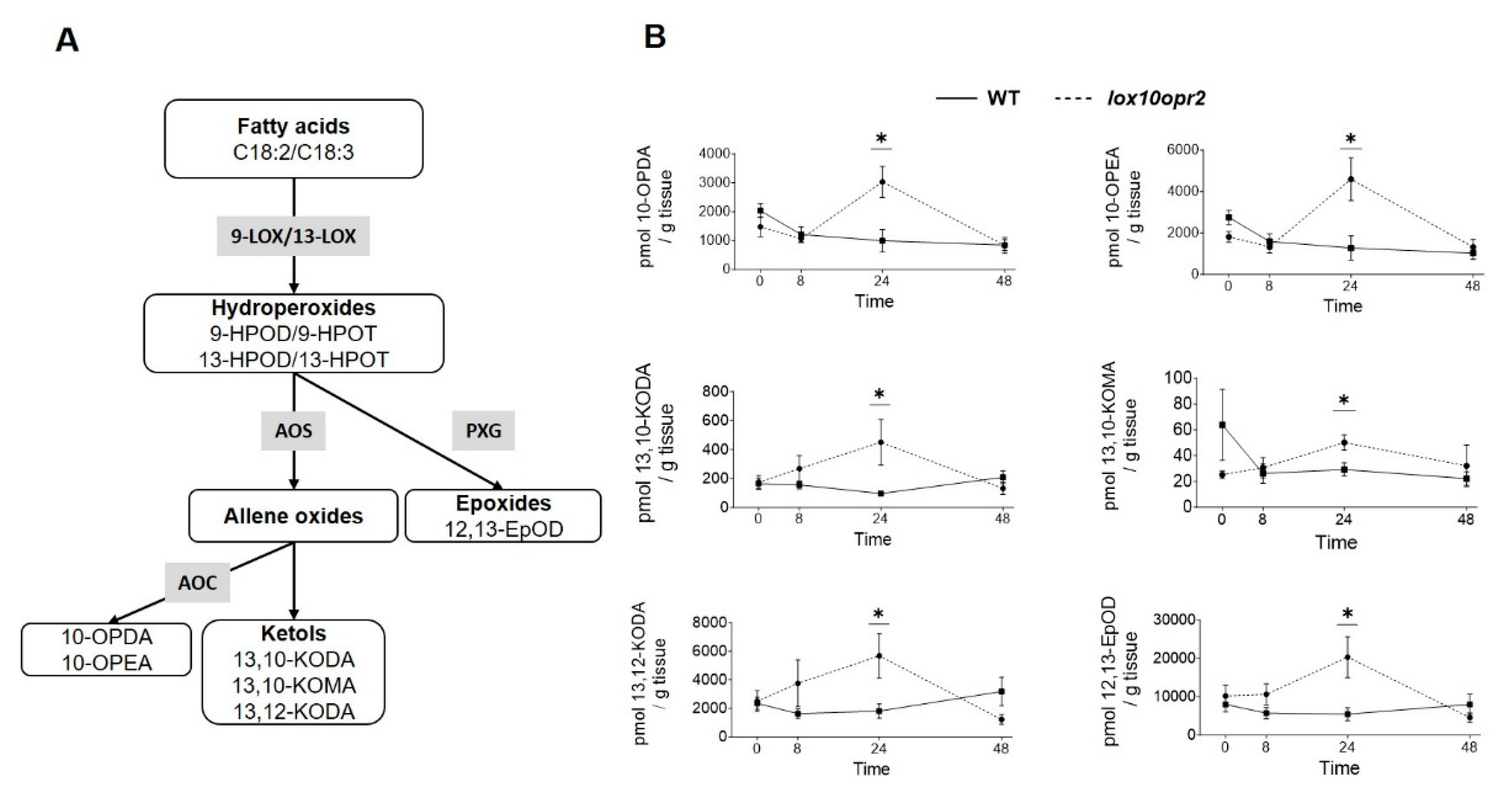
To investigate metabolites responsible for the enhanced defense and greater tolerance of lox10opr2, we quantified root metabolites of WT and lox10opr2 double mutant plants at 0-(no WCR herbivory), 8-, 24-, and 48 hours post-exposure to WCR larvae. The relative abundances of root metabolites in WT and lox10opr2 double mutant plants were visualized in two-way dendrogram and heatmap to reveal the temporal dynamics and variation of metabolite accumulation (Supplemental Figure S1). Examination of the heatmap suggested that, in aggregate, metabolite levels in WT decreased between 0 and 8 hours and remained at low levels through 48 hours after exposure to WCR larvae, while aggregate metabolite levels in lox10opr2 double mutants were low between 0 and 8 hours, increased at 24 hours, and decreased at 48 hours (Supplemental Figure S1). Interestingly, accumulation of JA and JA-Ile did not differ between WT and lox10opr2 double mutants at any of the time points, 0-48 hours, suggesting that, while phenotypic changes were evident in the double mutants, these changes were not due to changes in JA-related abundances (Supplemental Figures S1 and S2). After paired comparisons between WT-lox10opr2 mutants for all metabolites at 24 hours post exposure to WCR, we identified several compounds that accumulated at significantly higher levels in lox10opr2 mutant plants than WT at 24 hours post WCR infestation (Figure 7A). These included 4 oxylipins synthesized by the 9-LOX pathways: 10-OPDA (Student’s t-test, T= 2.583, P = 0.036), 10-OPEA (Student’s t-test, T= 2.899, P = 0.023), 13,10-KODA (Student’s t-test, T= 3.667, P = 0.014), and 13,10-KOMA (Student’s t-test, T= 2.793, P = 0.038) and 2 oxylipins synthesized in the 13-LOX pathways: 13,12-KODA (Student’s t-test, T= 2.546, P = 0.034) and 12,13-EpOD (Student’s t-test, T= 2.497, P = 0.037) (Figure 7B); and ABA (Student’s t-test, T= -3.0192, P = 0.016) (Supplemental Figure S2) relative to their WT controls before returning to WT levels at 48 hours post infestation. It is likely that these insecticidal and signaling oxylipins [31,46] underlie the increased mortality of larvae feeding on lox10opr2 roots and enhanced tolerance of lox10opr2 double mutants.
Figure 7.
Disruption of both ZmOPR2 and ZmLOX10 enhances ketol and insecticidal compounds production upon WCR infestation (A) Simplified lipoxygenase biosynthesis pathways showing the oxylipins with differential accumulation in the WT and lox10opr2 double mutants. Hydroperoxides derived from C18:2/C18:3 by 9- or 13-LOXs are used as substrates for the downstream branches. Enzyme symbols are represented by gray boxes while oxylipins are in white boxes and grouped by chemical class. Abbreviations are as follows: (Enzymes) AOC, allene oxide cyclase; AOS, allene oxide synthase; LOX, lipoxygenase; PXG, peroxygenase. (Oxylipins) C18:2, linoleic acid; C18:3 linolenic acid; 9-HPOD, (9S,10E,12Z)-9-hydroperoxy-10,12-octadecadienoic acid; 9-HPOT, (9S,10E,12Z,15Z)-9-hydroperoxy-10,12,15-octadecatrienoic acid; 10-OPDA, 10-oxo-11(Z),15(Z)-phytodienoic acid; 10-OPEA, 10-oxo-11(Z)-phytoenoic acid; 12,13-EpOD, cis-12,13-expoxy-9(Z),15(Z)-octadecenoic acid; 13,10-KODA, 13-hydroxy-10-oxo-11(E),15(Z)-octadecadienoic acid; 13,10-KOMA, 13-hydroxy-10-oxo-11(E)-octadecenoic acid; 13,12-KODA, 13-hydroxy-12-oxo-9(Z),15(Z)-octadecadienoic acid. (B) Accumulation of various oxylipins, including 10-OPDA, 10-OPEA, 13,10-KODA, 13,10-KOMA, 13,12-KODA, and 12,13-EpOD in WT and lox10opr2 mutans at 0, 8, 24, and 48 hours post WCR infestation. Bars are means ± SE. Asterisks represent statistically significant differences (Student’s t-test, *P < 0.05).
Figure 7.
Disruption of both ZmOPR2 and ZmLOX10 enhances ketol and insecticidal compounds production upon WCR infestation (A) Simplified lipoxygenase biosynthesis pathways showing the oxylipins with differential accumulation in the WT and lox10opr2 double mutants. Hydroperoxides derived from C18:2/C18:3 by 9- or 13-LOXs are used as substrates for the downstream branches. Enzyme symbols are represented by gray boxes while oxylipins are in white boxes and grouped by chemical class. Abbreviations are as follows: (Enzymes) AOC, allene oxide cyclase; AOS, allene oxide synthase; LOX, lipoxygenase; PXG, peroxygenase. (Oxylipins) C18:2, linoleic acid; C18:3 linolenic acid; 9-HPOD, (9S,10E,12Z)-9-hydroperoxy-10,12-octadecadienoic acid; 9-HPOT, (9S,10E,12Z,15Z)-9-hydroperoxy-10,12,15-octadecatrienoic acid; 10-OPDA, 10-oxo-11(Z),15(Z)-phytodienoic acid; 10-OPEA, 10-oxo-11(Z)-phytoenoic acid; 12,13-EpOD, cis-12,13-expoxy-9(Z),15(Z)-octadecenoic acid; 13,10-KODA, 13-hydroxy-10-oxo-11(E),15(Z)-octadecadienoic acid; 13,10-KOMA, 13-hydroxy-10-oxo-11(E)-octadecenoic acid; 13,12-KODA, 13-hydroxy-12-oxo-9(Z),15(Z)-octadecadienoic acid. (B) Accumulation of various oxylipins, including 10-OPDA, 10-OPEA, 13,10-KODA, 13,10-KOMA, 13,12-KODA, and 12,13-EpOD in WT and lox10opr2 mutans at 0, 8, 24, and 48 hours post WCR infestation. Bars are means ± SE. Asterisks represent statistically significant differences (Student’s t-test, *P < 0.05).
4. Discussion
JA has been extensively studied in aboveground tissue for its role in herbivory defense against chewing insects in numerous plant species, including in maize [13,14,47,48]. For instance, suppression of JA-producing OPRII enzymes resulted in reduced resistance to insect herbivory in aboveground Arabidopsis and maize tissues [13,40]. In contrast, there are no studies reporting the roles of any OPRI subfamily member in insect defense. The maize genome contains eight OPR genes, six of which belong to the OPRI subfamily [37]. Green leaf volatiles, mechanical wounding, and insect elicitor treatment have been shown to induce expression of ZmOPR1/2 [37,49], and feeding by either BAW or FAW induced ZmOPR2 expression in maize [15,50], suggesting that ZmOPR2 may be involved in insect defense. Elevated pathogen-induced JA contents in opr2 mutants [42] led us to test whether opr2 mutants also produce higher wound-induced jasmonates and are more resistant to FAW. However, the results showed that opr2 mutants unexpectedly accumulated lower levels of wound-induced JA and JA-Ile (Figure 2B and C), which was associated with increased susceptibility to FAW as manifested by greater consumption of leaf tissue by FAW and greater weight gain in FAW larvae after feeding on opr2 mutants compared to those feeding on WT. Correspondingly, overexpression of ZmOPR2 in Arabidopsis enhanced the production of wound-induced JA and JA-Ile (Figure 3C and D) and resistance to BAW. ZmOPR2 belongs to the OPRI subfamily that preferentially catalyzes the reduction of cis-(-) OPDA over the JA precursor cis-(+) OPDA [36], and localizes in the cytoplasm [41]. Therefore, ZmOPR2 was believed to not be directly involved in providing substrate for JA biosynthesis. In maize, only ZmOPR7 and ZmOPR8 belong to the OPRII subfamily, which preferentially catalyzes cis-(+) OPDA over cis-(-) OPDA to form the JA precursor OPC 8:0, and disruption of these two genes results in JA-deficiency and increased susceptibility to chewing insects and necrotrophic pathogens [13,51]. However, there are trace amounts of JA accumulation in young leaves of mature opr7opr8 plants [13], suggesting the occurrence of at least another OPR enzyme capable of producing JA under specific conditions. In Arabidopsis, the OPRII enzyme AtOPR3 was considered the only JA-producing OPR enzyme [52,53]. AtOPR2, an OPRI subfamily member, was ruled out for playing a direct role in JA biosynthesis because it is not localized to peroxisome and its low efficiency in catalyzing reduction on cis-(+) OPDA. Recently, a novel AtOPR3-independent pathway for JA biosynthesis was discovered that involves cytosolic AtOPR2 and uses 4,5-didehydrojasmonate as a JA substrate derived from dn-12-OPDA rather than cis-(+) OPDA [54]. In addition, increased dosage or transgenic overexpression of OPRI subfamily genes reduced seminal root growth in wheat and was associated with higher accumulation of JA [55]. In agreement with these findings, we observed that ZmOPR2-OE lines accumulated reduced levels of wound-induced dn-12-OPDA presumably due to its utilization as the primary substrate for JA synthesis (Figure 3B). Thus, it is likely that ZmOPR2 is also involved in wound-induced JA production in the pathway identified for Arabidopsis AtOPR2. In maize, the existence of a similarly alternative JA synthesis pathway is supported by our previous finding that normally JA-deficient opr7opr8 mutants accumulated significant levels of JA in response to C. graminicola at 7 days post-infection [56].
Other than directly producing JA, decreased wound-induced JA found in opr2 mutants may be due to increased catabolism of JA based on our finding that opr2 mutants accumulated significantly higher levels of JA catabolites, such as 12-OH-JA, 12-OH-JA-Ile, and 12-COOH-JA-Ile (Figure 4A-C). In contrast, after C. graminicola infection, susceptible opr2 mutants showed greater levels of JA and JA-Ile [42] accompanied by significantly lower pathogen-induced accumulation of 12-OH-JA (Supplemental Figure S3), while 12-OH-JA-Ile and 12-COOH-JA-Ile were undetectable. Together, these data suggest that ZmOPR2 may not only contribute to JA biosynthesis directly but also regulates JA catabolism under various stress conditions by a yet unknown mechanism.
Our results provided evidence that, in addition to the important role in JA synthesis and catabolism, ZmOPR2 appears to contribute to defense by an additional yet not understood mechanism. This conclusion is based on our finding that lox10opr2 double mutants are more susceptible than single lox10 or opr2 mutants (Figure 5), This result suggests that ZmOPR2 and ZmLOX10 interact positively to regulate defense against chewing insects, and though the increased susceptibility of the double mutant cannot be explained by lower JA or GLV production alone, as reported for lox10-3 single mutants [14]. Based on the well-reported biochemical activities of the OPRI subfamily, ZmOPR2 may also contribute to defense by detoxifying multiple α, β-unsaturated carbonyls [57].
Plants likely orchestrate divergent defense responses to aboveground and belowground insect herbivory. While JA and related metabolites play essential roles in mediation of defense responses against herbivory in aboveground tissues, several studies reported a much lower degree of the herbivory- or wounding-induced accumulation of JA and JA related metabolites and lower expression of JA-dependent genes in infested roots [58,59,60]. ZmOPR1 and ZmOPR2 are both significantly induced after WCR infestation, while expression of JA-producing ZmOPR7 and ZmOPR8 is not affected or minimally so per RNAseq transcriptome analyses of maize-WCR interactions [58,60]. Additionally, in the present study, we showed that JA accumulation upon WCR infestation was not significantly changed through 48 hours post-infestation, suggesting that ZmOPR1 and ZmOPR2 may contribute to resistance to WCR via a JA-independent pathway, and that JA may have little relevance to WCR defense. Given the clear evidence presented here that both ZmOPR2 and ZmLOX10 are required for defense against leaf herbivory by FAW, it was surprising to find that neither lox10-3 nor opr2-1 single mutants was affected in the resistance to WCR but lox10opr2 double mutants were more tolerant to herbivory as evidenced by less tissue consumed by WCR larvae. Notably, feeding on lox10opr2 double mutant roots resulted in greater mortality of WCR larvae. In the metabolite analysis of roots in response to WCR infestation we found that the more tolerant lox10opr2 mutants accumulated higher levels of several oxylipins at 24 hours post WCR infestation, including death acids, 10-OPDA, 10-OPEA, and several ketols, 13,10-KODA, 13,10-KOMA, and 13,12-KODA (Figure 7). Greater accumulation of these metabolites may underlie the greater mortality of WCR larvae that we found. 10-OPDA and 10-OPEA possess insecticidal activity and strongly suppress larval growth [46]. For instance, fatty acid-derived ketols were shown to contain hormone-like signaling activities to induce systemic resistance against pathogens and insects [30,31,34,35]. Moreover, Yuan et al. [31] showed that in addition to a strong signaling activity, the ketol 9,10-KODA displayed insecticidal activity against FAW. Whether the ketols 13,10-KODA, 13, 10-KOMA, and 13,12-KODA that accumulated to greater levels in lox10opr2 mutants also possess insecticidal activities or suppress larvae growth requires further study. Nevertheless, these data suggest that ZmOPR2 and ZmLOX10 negatively regulate WCR resistance by suppressing the biosynthesis of lipid-derived insecticidal death acids and ketols.
5. Conclusions
In this study, we showed that disruption of ZmOPR2 resulted in reduced accumulation of wound-induced JA, elevated levels of JA derivatives and increased susceptibility to leaf herbivory by FAW. Overexpression of ZmOPR2 in Arabidopsis enhanced wound-induced JA production, suppressed JA catabolism, and increased resistance to BAW. Together, these data demonstrated that ZmOPR2, a member of OPRI subfamily enzymes, modulates defense against leaf herbivores by regulating JA homeostasis. Moreover, we found that ZmOPR2 and ZmLOX10 synergistically contribute to insect defense in the aboveground tissue. In contrast, ZmOPR2 and ZmLOX10 cooperatively suppress resistance to WCR by inhibiting synthesis of death acids and ketols in roots upon WCR infestation. The results suggest that these metabolites may play a major role in defense against root herbivory.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
P.-C.H., J.M.G., J.S.B. and M.V.K. designed the research and analyzed data. P.-C.H. performed the FAW and BAW bioassays and drafted the article. J.M.G. conducted the WCR bioassays and helped draft the manuscript. J.P.T. and H.K. generated the Arabidopsis overexpression lines. P.-C.H., J.M.G. and M.V.K. wrote the article with input from and revisions by J.P.T., J.S.B. and H.K.
Funding
This study was supported by USDA-NIFA Grant No. 2015-67013-22816 awarded to M.V.K. and H.K., and USDA-NIFA 2017-67013-26524 awarded to M.V.K. and 2021-67013-33568 grants awarded to M.V.K. and J.S.B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Andie Miller, Department of Entomology, Texas A&M University for kindly shares FAW eggs/neonates and Katherine Berg-Falloure for help with phytohormone and oxylipin profiling via LC-MS/MS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- USDA, Crop production 2021 Summary. 2022.
- De La Fuente, G. N.; Barrero, I.; Murray, S. C.; Isakeit, T.; Kolomiets, M. V., Improving Maize Production under Drought Stress Traits, Screening Methods, and Environments. In Handbook of Plant and Crop Physiology, CRC Press: 2014.
- de Lange, E. S.; Balmer, D.; Mauch-Mani, B.; Turlings, T. C. J. , Insect and pathogen attack and resistance in maize and its wild ancestors, the teosintes. New Phytologist 2014, 204, 329–341. [Google Scholar] [CrossRef]
- Fontes-Puebla, A. A.; Borrego, E. J.; Kolomiets, M. V.; Bernal, J. S. , Maize biochemistry in response to root herbivory was mediated by domestication, spread, and breeding. Planta 2021, 254, 70. [Google Scholar] [CrossRef] [PubMed]
- Lima, A. F.; Bernal, J.; Venâncio, M. G. S.; de Souza, B. H. S.; Carvalho, G. A. , Comparative Tolerance Levels of Maize Landraces and a Hybrid to Natural Infestation of Fall Armyworm. Insects 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J. S.; Helms, A. M.; Fontes-Puebla, A. A.; DeWitt, T. J.; Kolomiets, M. V.; Grunseich, J. M. , Root volatile profiles and herbivore preference are mediated by maize domestication, geographic spread, and modern breeding. Planta 2022, 257, 24. [Google Scholar] [CrossRef] [PubMed]
- Warburton, M. L.; Woolfolk, S. W.; Smith, J. S.; Hawkins, L. K.; Castano-Duque, L.; Lebar, M. D.; Williams, W. P. , Genes and genetic mechanisms contributing to fall armyworm resistance in maize. The plant genome 2023, 16, e20311. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M. B., Giovanni; Biondi, Antonio; Calatayud, Paul-André; Day, Roger; Desneux, Nicolas; Harrison, Rhett D; Kriticos, Darren; Rwomushana, Ivan; van den Berg, Johnnie; Verheggen, François; Zhang, Yong-Jun; Agboyi, Lakpo Koku; Ahissou, Régis Besmer; Ba, Malick N; Wu, Kongming, Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomologia Generalis 2023, 43, 55.
- Montezano, D. G.; Specht, A.; Sosa-Gómez, D. R.; Roque-Specht, V. F.; Sousa-Silva, J. C.; Paula-Moraes, S. V.; Peterson, J. A.; Hunt, T. E. Host Plants of <i>Spodoptera frugiperda</i> (Lepidoptera: Noctuidae) in the Americas. African Entomology 2018, 26, 286–300. [Google Scholar]
- Overton, K.; Maino, J. L.; Day, R.; Umina, P. A.; Bett, B.; Carnovale, D.; Ekesi, S.; Meagher, R.; Reynolds, O. L. , Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): A review. Crop Protection 2021, 145, 105641. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. , Fall armyworm: impacts and implications for Africa. Outlooks on Pest Management 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Eschen, R.; Beale, T.; Bonnin, J. M.; Constantine, K. L.; Duah, S.; Finch, E. A.; Makale, F.; Nunda, W.; Ogunmodede, A.; Pratt, C. F. , Towards estimating the economic cost of invasive alien species to African crop and livestock production. CABI Agriculture and Bioscience 2021, 2, 1–18. [Google Scholar]
- Yan, Y.; Christensen, S.; Isakeit, T.; Engelberth, J.; Meeley, R.; Hayward, A.; Emery, R. J.; Kolomiets, M. V. , Disruption of OPR7 and OPR8 reveals the versatile functions of jasmonic acid in maize development and defense. Plant Cell 2012, 24, 1420–36. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S. A.; Nemchenko, A.; Borrego, E.; Murray, I.; Sobhy, I. S.; Bosak, L.; DeBlasio, S.; Erb, M.; Robert, C. A.; Vaughn, K. A.; Herrfurth, C.; Tumlinson, J.; Feussner, I.; Jackson, D.; Turlings, T. C.; Engelberth, J.; Nansen, C.; Meeley, R.; Kolomiets, M. V. , The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. The Plant journal: for cell and molecular biology 2013, 74, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luthe, D. , Key Genes in the JAZ Signaling Pathway Are Up-Regulated Faster and More Abundantly in Caterpillar-Resistant Maize. Journal of chemical ecology 2022, 48, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Gray, M. E.; Sappington, T. W.; Miller, N. J.; Moeser, J.; Bohn, M. O. , Adaptation and invasiveness of western corn rootworm: intensifying research on a worsening pest. Annual review of entomology 2009, 54, 303–321. [Google Scholar] [CrossRef]
- Meinke, L. J.; Sappington, T. W.; Onstad, D. W.; Guillemaud, T.; Miller, N. J.; Komáromi, J.; Levay, N.; Furlan, L.; Kiss, J.; Toth, F. , Western corn rootworm (Diabrotica virgifera virgifera LeConte) population dynamics. Agricultural and Forest Entomology 2009, 11, 29–46. [Google Scholar] [CrossRef]
- Lombaert, E.; Ciosi, M.; Miller, N. J.; Sappington, T. W.; Blin, A.; Guillemaud, T. , Colonization history of the western corn rootworm (Diabrotica virgifera virgifera) in North America: insights from random forest ABC using microsatellite data. Biological Invasions 2018, 20, 665–677. [Google Scholar] [CrossRef]
- Bažok, R.; Lemić, D.; Chiarini, F.; Furlan, L. , Western Corn Rootworm (Diabrotica virgifera virgifera LeConte) in Europe: Current Status and Sustainable Pest Management. Insects 2021, 12, 195. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, S.; Smith, D. , Has resistance taken root in US corn fields? Demand for insect control. American Journal of Agricultural Economics 2018, 100, 1136–1150. [Google Scholar] [CrossRef]
- Castano-Duque, L.; Loades, K. W.; Tooker, J. F.; Brown, K. M.; Paul Williams, W.; Luthe, D. S. , A Maize Inbred Exhibits Resistance Against Western Corn Rootwoorm, Diabrotica virgifera virgifera. Journal of chemical ecology 2017, 43, (11–12). [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C. , The lipoxygenase pathway. Annual review of plant biology 2002, 53, 275–97. [Google Scholar] [CrossRef]
- Wasternack, C.; Feussner, I. , The Oxylipin Pathways: Biochemistry and Function. Annual review of plant biology 2018, 69, 363–386. [Google Scholar] [CrossRef] [PubMed]
- Borrego, E. J.; Kolomiets, M. V. , Synthesis and Functions of Jasmonates in Maize. Plants (Basel, Switzerland) 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Howe, G. A.; Jander, G. , Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Meldau, S.; Howe, G. A. , Role of phytohormones in insect-specific plant reactions. Trends in plant science 2012, 17, 250–259. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, F.; Melotto, M.; Yao, J.; He, S. Y. , Jasmonate signaling and manipulation by pathogens and insects. J Exp Bot 2017, 68, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Strnad, M. , Jasmonates: News on Occurrence, Biosynthesis, Metabolism and Action of an Ancient Group of Signaling Compounds. International journal of molecular sciences 2018, 19, 2539. [Google Scholar] [CrossRef]
- Suza, W. P.; Staswick, P. E. , The role of JAR1 in Jasmonoyl-L: -isoleucine production during Arabidopsis wound response. Planta 2008, 227, 1221–32. [Google Scholar] [CrossRef]
- Berg-Falloure, K. M.; Kolomiets, M. V. , Ketols Emerge as Potent Oxylipin Signals Regulating Diverse Physiological Processes in Plants. Plants (Basel, Switzerland) 2023, 12. [Google Scholar] [CrossRef]
- Yuan, P.; Borrego, E.; Park, Y. S.; Gorman, Z.; Huang, P. C.; Tolley, J.; Christensen, S. A.; Blanford, J.; Kilaru, A.; Meeley, R.; Koiwa, H.; Vidal, S.; Huffaker, A.; Schmelz, E.; Kolomiets, M. V. , 9,10-KODA, an α-ketol produced by the tonoplast-localized 9-lipoxygenase ZmLOX5, plays a signaling role in maize defense against insect herbivory. Molecular plant 2023. [Google Scholar] [CrossRef]
- Haque, E.; Osmani, A. A.; Ahmadi, S. H.; Ogawa, S.; Takagi, K.; Yokoyama, M.; Ban, T. , KODA, an α-ketol derivative of linolenic acid provides wide recovery ability of wheat against various abiotic stresses. Biocatalysis and Agricultural Biotechnology 2016, 7, 67–75. [Google Scholar] [CrossRef]
- Wang, S.; Saito, T.; Ohkawa, K.; Ohara, H.; Shishido, M.; Ikeura, H.; Takagi, K.; Ogawa, S.; Yokoyama, M.; Kondo, S. , α-Ketol linolenic acid (KODA) application affects endogenous abscisic acid, jasmonic acid and aromatic volatiles in grapes infected by a pathogen (Glomerella cingulata). Journal of plant physiology 2016, 192, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, K. D.; Borrego, E. J.; Kenerley, C. M.; Kolomiets, M. V. , Oxylipins Other Than Jasmonic Acid Are Xylem-Resident Signals Regulating Systemic Resistance Induced by Trichoderma virens in Maize. Plant Cell 2020, 32, 166–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, K. D.; Gorman, Z.; Huang, P. C.; Kenerley, C. M.; Kolomiets, M. V. , Trichoderma virens colonization of maize roots triggers rapid accumulation of 12-oxophytodienoate and two ᵧ-ketols in leaves as priming agents of induced systemic resistance. Plant signaling & behavior 2020, 15, 1792187. [Google Scholar]
- Schaller, F.; Hennig, P.; Weiler, E. W. , 12-Oxophytodienoate-10,11-reductase: occurrence of two isoenzymes of different specificity against stereoisomers of 12-oxophytodienoic acid. Plant physiology 1998, 118, 1345–51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Simmons, C.; Yalpani, N.; Crane, V.; Wilkinson, H.; Kolomiets, M. , Genomic analysis of the 12-oxo-phytodienoic acid reductase gene family of Zea mays. Plant Mol Biol 2005, 59, 323–43. [Google Scholar] [CrossRef] [PubMed]
- Schaller, F.; Biesgen, C.; Müssig, C.; Altmann, T.; Weiler, E. W. , 12-Oxophytodienoate reductase 3 (OPR3) is the isoenzyme involved in jasmonate biosynthesis. Planta 2000, 210, 979–84. [Google Scholar] [CrossRef]
- Strassner, J.; Fürholz, A.; Macheroux, P.; Amrhein, N.; Schaller, A. , A homolog of old yellow enzyme in tomato. Spectral properties and substrate specificity of the recombinant protein. The Journal of biological chemistry 1999, 274, 35067–73. [Google Scholar] [CrossRef]
- Chehab, E. W.; Kim, S.; Savchenko, T.; Kliebenstein, D.; Dehesh, K.; Braam, J. , Intronic T-DNA Insertion Renders Arabidopsis opr3 a Conditional Jasmonic Acid-Producing Mutant. Plant physiology 2011, 156, 770–778. [Google Scholar] [CrossRef]
- Tolley, J. P.; Nagashima, Y.; Gorman, Z.; Kolomiets, M. V.; Koiwa, H. , Isoform-specific subcellular localization of Zea mays lipoxygenases and oxo-phytodienoate reductase 2. Plant Gene 2018, 13, 36–41. [Google Scholar] [CrossRef]
- Huang, P. C.; Tate, M.; Berg-Falloure, K. M.; Christensen, S. A.; Zhang, J.; Schirawski, J.; Meeley, R.; Kolomiets, M. V. , A non-JA producing oxophytodienoate reductase functions in salicylic acid-mediated antagonism with jasmonic acid during pathogen attack. Molecular plant pathology 2023, 24, 725–741. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. , Jasmonates: News on Occurrence, Biosynthesis, Metabolism and Action of an Ancient Group of Signaling Compounds. International journal of molecular sciences 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Borrego, E. J.; Gorman, Z.; Huang, P. C.; Kolomiets, M. V. , Relative contribution of LOX10, green leaf volatiles and JA to wound-induced local and systemic oxylipin and hormone signature in Zea mays (maize). Phytochemistry 2020, 174, 112334. [Google Scholar] [CrossRef] [PubMed]
- McConn, M.; Creelman, R. A.; Bell, E.; Mullet, J. E.; Browse, J. , Jasmonate is essential for insect defense in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 1997, 94, 5473–7. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S. A.; Huffaker, A.; Kaplan, F.; Sims, J.; Ziemann, S.; Doehlemann, G.; Ji, L.; Schmitz, R. J.; Kolomiets, M. V.; Alborn, H. T.; Mori, N.; Jander, G.; Ni, X.; Sartor, R. C.; Byers, S.; Abdo, Z.; Schmelz, E. A. , Maize death acids, 9-lipoxygenase–derived cyclopente(a)nones, display activity as cytotoxic phytoalexins and transcriptional mediators. Proceedings of the National Academy of Sciences 2015, 112, 11407–11412. [Google Scholar] [CrossRef] [PubMed]
- Verhage, A.; Vlaardingerbroek, I.; Raaijmakers, C.; Van Dam, N.; Dicke, M.; Van Wees, S.; Pieterse, C. , Rewiring of the Jasmonate Signaling Pathway in Arabidopsis during Insect Herbivory. Frontiers in plant science 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Campos, M. L.; Kang, J. H.; Howe, G. A. , Jasmonate-triggered plant immunity. Journal of chemical ecology 2014, 40, 657–75. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, J.; Seidl-Adams, I.; Schultz, J. C.; Tumlinson, J. H. , Insect elicitors and exposure to green leafy volatiles differentially upregulate major octadecanoids and transcripts of 12-oxo phytodienoic acid reductases in Zea mays. Molecular plant-microbe interactions: MPMI 2007, 20, 707–16. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Hojo, Y.; Strickler, S. R.; Bartsch, L. J.; Archer, C. M.; Ahern, K. R.; Zhou, S.; Christensen, S. A.; Galis, I.; Mueller, L. A.; Jander, G. , Rapid defense responses in maize leaves induced by Spodoptera exigua caterpillar feeding. Journal of Experimental Botany 2017, 68, 4709–4723. [Google Scholar] [CrossRef]
- Christensen, S. A.; Nemchenko, A.; Park, Y. S.; Borrego, E.; Huang, P. C.; Schmelz, E. A.; Kunze, S.; Feussner, I.; Yalpani, N.; Meeley, R.; Kolomiets, M. V. , The novel monocot-specific 9-lipoxygenase ZmLOX12 is required to mount an effective jasmonate-mediated defense against Fusarium verticillioides in maize. Molecular plant-microbe interactions: MPMI 2014, 27, 1263–76. [Google Scholar] [CrossRef]
- Stintzi, A.; Browse, J. , The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis. Proceedings of the National Academy of Sciences of the United States of America 2000, 97, 10625–30. [Google Scholar] [CrossRef]
- Sanders, P. M.; Lee, P. Y.; Biesgen, C.; Boone, J. D.; Beals, T. P.; Weiler, E. W.; Goldberg, R. B. , The arabidopsis DELAYED DEHISCENCE1 gene encodes an enzyme in the jasmonic acid synthesis pathway. Plant Cell 2000, 12, 1041–61. [Google Scholar] [CrossRef] [PubMed]
- Chini, A.; Monte, I.; Zamarreno, A. M.; Hamberg, M.; Lassueur, S.; Reymond, P.; Weiss, S.; Stintzi, A.; Schaller, A.; Porzel, A.; Garcia-Mina, J. M.; Solano, R. , An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis. Nat Chem Biol 2018, 14, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Gabay, G.; Wang, H.; Zhang, J.; Moriconi, J. I.; Burguener, G. F.; Gualano, L. D.; Howell, T.; Lukaszewski, A.; Staskawicz, B.; Cho, M.-J.; Tanaka, J.; Fahima, T.; Ke, H.; Dehesh, K.; Zhang, G.-L.; Gou, J.-Y.; Hamberg, M.; Santa-María, G. E.; Dubcovsky, J. , Dosage differences in 12-OXOPHYTODIENOATE REDUCTASE genes modulate wheat root growth. Nature Communications 2023, 14, 539. [Google Scholar] [CrossRef] [PubMed]
- Gorman, Z.; Christensen, S. A.; Yan, Y.; He, Y.; Borrego, E.; Kolomiets, M. V. , Green leaf volatiles and jasmonic acid enhance susceptibility to anthracnose diseases caused by Colletotrichum graminicola in maize. Molecular plant pathology 2020, 21, 702–715. [Google Scholar] [CrossRef] [PubMed]
- Kohli, R. M.; Massey, V. , The oxidative half-reaction of Old Yellow Enzyme. The role of tyrosine 196. The Journal of biological chemistry 1998, 273, 32763–70. [Google Scholar] [CrossRef]
- Ye, W.; Bustos-Segura, C.; Degen, T.; Erb, M.; Turlings, T. C. J. , Belowground and aboveground herbivory differentially affect the transcriptome in roots and shoots of maize. Plant Direct 2022, 6, e426. [Google Scholar] [CrossRef]
- Erb, M.; Flors, V.; Karlen, D.; De Lange, E.; Planchamp, C.; D’Alessandro, M.; Turlings, T. C. J.; Ton, J. , Signal signature of aboveground-induced resistance upon belowground herbivory in maize. The Plant Journal 2009, 59, 292–302. [Google Scholar] [CrossRef]
- Pingault, L.; Basu, S.; Vellichirammal, N. N.; Williams, W. P.; Sarath, G.; Louis, J. , Co-Transcriptomic Analysis of the Maize-Western Corn Rootworm Interaction. Plants 2022, 11, 2335. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



