
Submitted:
13 August 2023
Posted:
14 August 2023
You are already at the latest version
Abstract
Scientific evidence increasingly supports the strong link between diet and health, acknowledging that a well-balanced diet plays a crucial role in preventing chronic diseases such as obesity, diabetes, cardiovascular issues, and certain types of cancer. This perspective opens the door to developing precision diets, particularly tailored for individuals at risk of developing cancer. It encompasses a vast research area and involves the study of an expanding array of compounds with multi-level "omics" compositions, including genomics, transcriptomics, proteomics, epigenomics, miRNomics, and metabolomics. We review here the components of the Southern European Atlantic Diet (SEAD) from both a chemical and pharmacological standpoint. The information sources consulted, complemented by crystallographic data from the Protein Data Bank, establish a direct link between SEAD and its anticancer properties. The data collected strongly suggest that SEAD offers an exceptionally healthy profile, particularly due to the presence of beneficial biomolecules in its foods. The inclusion of olive oil and paprika in this diet provides numerous health benefits, and science supports the anti-cancer properties of dietary supplements with biomolecules sourced from vegetables of the brassica genus. Nonetheless, further research is warranted in this field to gain deeper insights into the potential benefits of SEAD's bioactive compounds against cancer.
Keywords:
Southern Atlantic Diet
; Anticancer
; glucosinolates
; sulforaphane
; phenolic compounds
; flavonoids
; antioxidant
; chemoprevention
; nutragenomic
; nutraceutical
1. Introduction
According to the World Health Organization [1], cancer is the second leading cause of death worldwide, and between 30% and 50% of cancer deaths could be prevented by modifying or avoiding key risk factors, primarily through proper nutrition, maintaining a healthy weight, and adopting certain healthy habits. In addition, the International Agency for Research on Cancer (IARC) [2] estimates that around one-third of cancer cases are related to dietary and nutritional factors. These studies have found associations between a high intake of ultra-processed foods, such as those commonly found in fast food, and an increased risk of developing certain types of cancer, such as colon, breast, and prostate cancer [3]. It is evident that adopting healthy eating patterns based on a balanced diet rich in fresh foods, such as fruits, vegetables, whole grains, and lean proteins, is crucial to reducing the incidence of cancer worldwide. Epigenetics is the study of changes in gene expression (phenotype) that do not involve modifications to the DNA sequence (genotype). It investigates how our genes can be activated or deactivated by various external factors such as the environment, lifestyle, physical activity, exposure to toxins, or diet [4].
Currently, the growing scientific evidence shows that nutritional and natural agents, known as nutraceuticals, could modulate the expression of miRNAs and, as a result, influence cellular responses in both health and disease conditions, including cancer. Among these mechanisms, it has been demonstrated that diet can affect DNA methylation, which is a chemical modification that can regulate gene activity. Therefore, the considerable significance of dietary biomolecules becomes evident, given their participation as functional bioactive compounds in crucial chemical processes, particularly, as antioxidants in cancer prevention by protecting against DNA damage and regulating DNA methylation [5,6].
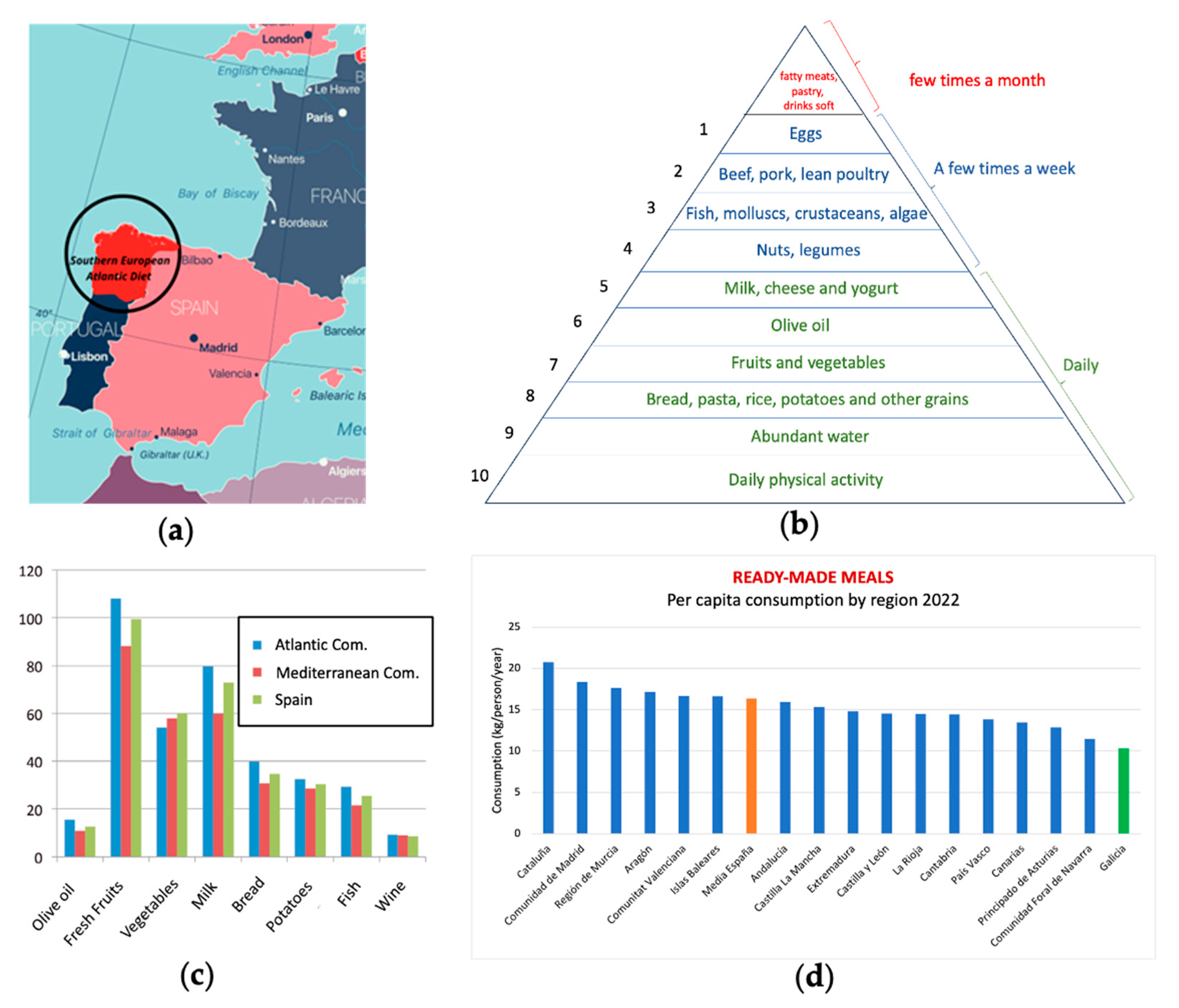
The Southern European Atlantic Diet (SEAD) [7,8,9,10] is a dietary pattern traditionally followed by populations inhabiting coastal regions of the South Atlantic, such as Galicia and Asturias in northwest Spain, including the province of León, areas of Cantabria, and northern Portugal (Figure 1a). The increasing interest in the SEAD dietary pattern is an essential component in understanding the significant flow of tourists and pilgrims who arrive every year to undertake the well-known “Camino de Santiago.” In fact, a recent study by Carral and coworkers analyzes the relationship between Gastronomy and Tourism: Socioeconomic and Territorial Implications in Santiago de Compostela-Galicia (NW Spain) [11].
SEAD is characterized by its focus on fresh and local foods, especially those from the Atlantic Ocean and its surrounding regions. It is based on three fundamental pillars. Firstly, a food chain rich in phytoplankton originating a wide variety of quality marine products, including fish, seafood, and seaweed [12]. Secondly, distinctive inland products, represented by vegetables of the brassica genus, and a high consumption of olive oil. Lastly, culinary techniques that respect the nutritional value of raw materials, with the most characteristic being stewing and roasting.
Compared to the Mediterranean diet, the South European Atlantic Diet (SEAD) presents some differences. While the Mediterranean diet is characterized by regular consumption of olive oil, nuts, legumes, and whole grains, the SEAD places greater emphasis on marine products, local vegetables, bread, dairy products, and a higher intake of olive oil, as shown in the nutritional pyramid and the key food consumption graph (Figure 1b, c) [13]. Additionally, the influence of the Atlantic climate in this region determines culinary preferences and the ingredients used in typical dishes. A relevant data point is depicted in the graph below, based on the report of food consumption in Spain for the year 2022, conducted by the Ministry of Agriculture, Fisheries, and Food of the Spanish Government. It shows a predominance of fresh and seasonal food consumption, with Galicia being the last autonomous community in per capita consumption of ready-made dishes, including canned and frozen ready-made dishes, soups and creams, pizza, other ready-made pasta dishes, refrigerated tortillas, and other prepared dishes (Figure 1d).
The Galician coasts, known for their estuaries called “Galician rías”, have unique characteristics that influence the temperature and nutrients present in the water, favoring the cultivation of high-quality seafood (Figure 2a). The Galician estuaries are elongated and narrow, formed by the flooding of river valleys into the Atlantic Ocean. These estuaries are characterized by their mixing of freshwater and saltwater, creating an ecosystem rich in nutrients such as nitrates and phosphates. These topological peculiarities make the Galician estuaries a unique ecosystem that allows the growth of a wide variety of fish, and above all, shellfish, unparalleled in Europe.
In addition to marine products, the SEAD includes a wide variety of fresh seasonal vegetables, greens, and fruits. Notable vegetables such as turnip tops (Figure 2c), cabbage, collard greens, potatoes, onions, and garlic are used in the preparation of traditional dishes such as Galician or Portuguese broth (“caldo”) or stew (“cocido”) (Figure 2e), as well as in Portuguese traditional dishes like frying or white rice, along with a higher consumption of water and wine (Figure 1b). These foods provide fiber, vitamins, minerals, and phytonutrients that promote intestinal health and proper body functioning.
Figure 1.
(a) Territorial zone covered by the SEAD; (b) Nutritional pyramid of the South European Atlantic Diet; [modified and adapted from [8]; (c) Comparative consumption of the main beneficial elements of Iberian diets and the whole of Spain. Adapted from [9]; (d) Consumption of ready-made meals in Spain (by Regions): Own elaboration. Source: Ministry of Agriculture, Fisheries, and Food. Government of Spain. Food consumption report of Spain, 2022. Available at [14].
Figure 1.
(a) Territorial zone covered by the SEAD; (b) Nutritional pyramid of the South European Atlantic Diet; [modified and adapted from [8]; (c) Comparative consumption of the main beneficial elements of Iberian diets and the whole of Spain. Adapted from [9]; (d) Consumption of ready-made meals in Spain (by Regions): Own elaboration. Source: Ministry of Agriculture, Fisheries, and Food. Government of Spain. Food consumption report of Spain, 2022. Available at [14].
Regarding fats, SEAD primarily uses olive oil, although other vegetable oils, especially in the Portuguese region, such as sunflower or rapeseed oils, are also employed. The flavor and health properties of olive oil make it a key ingredient in Galician cuisine, adding a distinctive touch to the dishes.
We must highlight that Galicia has high-quality meats, resulting in two Protected Geographical Indications (PGI): “IGP Ternera Gallega” (PGI Galician Veal), recognized by the European Union with this quality seal since 1996 [15], and “IGP Vaca y Buey de Galicia” (Galician Cow and Ox) [16], with the most well-known native breed being the “Rubia Gallega” cow [16] (Figure 2d). This meat comes from cows and oxens of the main breeds in Galicia: Rubia Gallega, Morenas Gallegas (Cachena, Caldelana, Frieiresa, Limiana, and Vianesa), Asturiana de los Valles, Limousin, Rubia de Aquitaine, Frisona, and Pardo Alpina. In this excellent meat, the intramuscular fat percentage exceeds 5.6% on average, and it is characterized by being tender, juicy, with a marked persistent meaty flavor, and a slightly acidic aftertaste. Other quality meats with the seal in Galicia include “IGP Capón de Villalba” (poultry) and “IGP Lacón Gallego” from pork [17].
In Asturias, we find “IGP Ternera Asturiana” (Asturian veal) [18], and in northern Portugal, another quality meat can be found, “IGP Carne de Bovino Cruzado dos Lameiros do Barroso” [19] from the Barroso Agricultural Cooperative. In the Bierzo area of Spain, “IGP Botillo del Bierzo” is produced [20].
One of the most notable practices in shellfish aquaculture in Galicia is the use of “bateas.” Bateas are floating structures made of wood or metal installed in the estuaries for shellfish cultivation (Figure 2a). These bateas serve as platforms where ropes or nets are suspended in the water for mollusks to adhere to and grow (Figure 2b). Bateas allow for more efficient cultivation control, protecting shellfish from predators and facilitating their harvest. Different species of mollusks are cultivated in Galician bateas, with the most prominent being mussels, oysters, and scallops. Mussels are one of Galicia’s flagship products, cultivated in large quantities, making Galicia the top producer in Europe, with around 3000 bateas.
Figure 2.
Representative products of the South European Atlantic Diet (SEAD): a) Galician Estuary, where numerous “bateas” are visible, cultivating mussels, oysters, scallops, etc. Photo by Gabriel González (https://pxhere.com/es/photo/470608?utm_content=shareClip&utm_medium=referral&utm_source=pxhere). b) Underwater ropes of the “batea” showing the cultivation of mussels. c) Above: Detail of the plant and fruit known as “pimiento de Padrón” (Padrón pepper). Below: Detail of the most consumed vegetable in Galicia, turnip top. d) Example of native Galician Rubia Gallega cow. Photo by Luis Miguel Bugallo Sánchez (Creative Commons: (CC BY-SA 3.0). e) Galician broth (Photo by Juan Mejuto, Wikimedia Commons, cc-by-sa-2) and Portuguese green soup (Photo by Matheus Hidalgo, Creative Commons Attribution-Share Alike 2.5 Brazil). f) and g) Example of a typical table representing the Galician Atlantic diet, featuring native products such as octopus or barnacles, among other shellfish. Seafood and fish are also consumed in the traditional “empanada” (a type of pie). Red and white wines and water are the most common beverages. h) Bacalao (codfish), a symbol of Portuguese gastronomy. Photo by Liliana Fuchs (Creative Commons: (CC BY-NC-ND 2.0).
Figure 2.
Representative products of the South European Atlantic Diet (SEAD): a) Galician Estuary, where numerous “bateas” are visible, cultivating mussels, oysters, scallops, etc. Photo by Gabriel González (https://pxhere.com/es/photo/470608?utm_content=shareClip&utm_medium=referral&utm_source=pxhere). b) Underwater ropes of the “batea” showing the cultivation of mussels. c) Above: Detail of the plant and fruit known as “pimiento de Padrón” (Padrón pepper). Below: Detail of the most consumed vegetable in Galicia, turnip top. d) Example of native Galician Rubia Gallega cow. Photo by Luis Miguel Bugallo Sánchez (Creative Commons: (CC BY-SA 3.0). e) Galician broth (Photo by Juan Mejuto, Wikimedia Commons, cc-by-sa-2) and Portuguese green soup (Photo by Matheus Hidalgo, Creative Commons Attribution-Share Alike 2.5 Brazil). f) and g) Example of a typical table representing the Galician Atlantic diet, featuring native products such as octopus or barnacles, among other shellfish. Seafood and fish are also consumed in the traditional “empanada” (a type of pie). Red and white wines and water are the most common beverages. h) Bacalao (codfish), a symbol of Portuguese gastronomy. Photo by Liliana Fuchs (Creative Commons: (CC BY-NC-ND 2.0).

The cultivation of mussels is sustainable and is known for its high quality and exceptional taste. Oysters are also highly appreciated and cultivated in Galician estuaries. They are renowned for their delicate flavor and smooth texture. Scallops, another emblematic mollusk of the region, are also grown in bateas and highly valued in the local gastronomy. Similar practices for shellfish cultivation in bateas exist in other regions of Europe, though cultivated species and methods used may vary. For example, in France, similar structures called “parcs à huîtres” are used for oyster farming in the Brittany region. In Scotland, the “aquaculture” method is employed for cultivating mussels on ropes suspended in the water. While there are similar practices in other European regions, the diversity and quality of Galician shellfish are highly recognized and appreciated in the South European Atlantic Diet (SEAD) (Figure 2g). One culinary aspect that influences the quality of the diet is the way foods are cooked, with a predominance of boiling, grilling, or roasting in Galicia, while in Portugal, fried or grilled foods, such as bacalao (Figure 2h), are more commonly preferred. Excessive consumption of fried foods can increase the presence of saturated and trans fats in the food, which can have negative effects on health.
Many of the characteristics offered by the SEAD can be attributed to the trophic chain. A study conducted by CSIC determined the abundance and diversity of photosynthetic microorganisms present in the Vigo Estuary, making it an autotrophic ecosystem [21]. The marine strip in which Galicia is located makes its sea particularly fertile, rich in marine flora and fauna, thanks to the daily hours of sunlight it receives. When we add the water currents and temperature, generated by the confluence of the Cantabrian Sea and the Atlantic Ocean provides an unique and irreplaceable ecosystem. Thus, it forms a food chain of great nutritional value, starting from phytoplankton, algae, crustaceans, and small fish, all the way up to large fish [22].
In this ecosystem, algae play a fundamental role. Galician algae are a food with many antioxidant and anti-cancer properties, containing important bioactive molecules such as fucoxanthin and phlorotannins (Figure 3). Fucoxanthin is a type of carotenoid extracted from seaweed, specifically the pigment responsible for the brown color of brown algae. It is abundant in “Kombu” algae, particularly the Saccharina Latissima, a brown marine algae of the Laminariaceae family that grows in cold waters, along with the Undaria pinnatifida (Wakame), among others, characteristic of the Galician coasts [23]. At the molecular level, fucoxanthin is a xanthophyll [24], structurally similar as beta-carotene and vitamin A. Algae also contain alginic acid, which enriches this trophic chain. While not a traditional dish in this region, algae are becoming increasingly incorporated into the SEAD diet. The growing application of algae biotechnology is interesting [25] for other uses, such as the pharmaceutical and cosmetic industries, renewable energy, and as feed for fish, mollusks, and crustaceans. Therefore, algae are gaining increasing interest in nutrition due to their fiber content and low caloric value in weight loss diets, aligning with the recent framework of “Zero Obesity 2022-2030” in Galicia [26].
Scientific studies encompassed in the GALIAT6+7 project (driven by 6 companies and 7 scientific groups), aimed at conducting a randomized, family-focused dietary intervention to evaluate the Atlantic diet, have linked the quality of the SEAD with a series of health and longevity benefits [27,28]. It has been observed that people following this dietary pattern have a lower risk of cardiovascular diseases, type 2 diabetes, obesity, and certain types of cancer [28]. In relation to the latter, it has been reported that the SEAD may have positive effects at the epigenetic level. A diet rich in fresh foods, such as the SEAD, has been shown to modulate DNA methylation and histone modification, which can impact the regulation of genes related to health and longevity [13]. As a result of this epigenetics, Galicia is beginning to receive worldwide attention for its longevity, and there are several professionals from different disciplines, entrepreneurs and associations that study this phenomenon of centenarians in Ourense [29], where there are 384 centenarians, equivalent to 126 per 100,000 inhabitants, or Lugo with 333, which is equivalent to 96 per 100,000 inhabitants [30], comparable to longevity paradigms like Icaria in Greece or Okinawa in Japan [31,32].
An important study was conducted in recent years by Carballo-Casla and coworkers, related to the properties of the SEAD and all-cause mortality in older adults [33]. Another study confirmed that the SEAD is associated with lower concentrations of markers of coronary risk [34]. Adherence to the SEAD and occurrence of nonfatal acute myocardial infarction were analyzed by Oliveira and coworkers [35], and Tejera-Pérez reported a study about the SEAD and a recent proposal of the SEAD Index [7]. They concluded that SEAD should be considered an excellent dietary pattern and lifestyle. In addition, a study about the SEAD and depression risk in Europe was also reported [33]. Romero-Rodríguez and coworkers performed a thirteen-item questionnaire developed to assess the degree of adherence to the SEAD pattern of the Galician senior population. Another questionnaire was designed to determine the degree of knowledge about SEAD. The results confirmed that the adherence to this diet was medium to high [36].
In this review, which focuses more on Galicia and northern Portugal regions, our aim is to highlight the main components and biomolecules present in the SEAD which, according to data, contributes to longevity in the Galician-Portuguese population. The main objective of this work is, therefore, to compile and analyze for the first time, the bioactive compounds with anticancer properties found in significant amounts and consumption in the components of SEAD foods, from a chemical and pharmacological perspective, as well as their specificities in relation to other diets worldwide.
2. Materials and Methods
For the study of the gathered information, the following databases were consulted: Scifinder, PubMed, Protein Data Bank (PDB), and the Galiat6+7 [37]. Additionally, other government sources at the national or regional level were utilized.
2.1. Research Question and Research Strategy
The key research question was: “What are the frequently consumed components in the SEAD that have demonstrated antioxidant or anticancer properties supported by evidence from scientific studies?” A series of keywords (terms in pairs or single words) were used in the search: For instance, SEAD + antioxidant, Atlantic diet + anticancer, SEAD + anticancer, glucosinolates + anticancer, sulforaphane + anticancer, chemoprevention, nutrigenomic, nutraceutical, cancer, chronic diseases; phenolic compounds, etc. Additionally, a search was conducted for each relevant component (biomolecule or nutraceutical) present in the SEAD, food by food. In addition to the term search, the words “cancer” and “epigenetics” were also used. For each of the key relevant components of the SEAD, a search for crystallographic structures in the PDB related to some type of cancer-related process was performed by analyzing the target, introducing the corresponding component keyword into the PDB search engine. When co-crystals of biomolecules from the SEAD and tumor or antioxidant targets were found, they were included. The binding mode of ligand-protein was studied, and the corresponding figure showing the binding site was elaborated.
The search was conducted in various databases, including PubMed, Scifinder, PDB, and Galiat 6+7, covering publications from inception until July 2023 (Figure 4), as mentioned previously. Additionally, the search was extended to the gray literature. The search terms used, with a combination of terms whenever applicable in each database, included: Atlantic diet, Galician Seafood, and Epigenetics. Publications that were over 15 years old, written in languages other than Spanish or English, and those without full-text availability were initially discarded unless they were of crucial importance to the study.
3. Discussion
The analysis of the information, which encompasses different perspectives (dietary-nutritional, chemical, pharmacological), has been carried out considering each representative food of the diet that, due to its frequent consumption in the areas of Galicia, Asturias, and Northern Portugal mainly, can be considered intrinsic to the Atlantic diet. From each food, the main components that have shown scientific evidence of possessing antioxidant, antiproliferative, and overall anticancer activities have been selected.
3.1. Epigenetics and cancer. Longevity and benefits of the Atlantic diet. Previous studies.
In the last decades, there has been a significant increase in cancer cases worldwide. While various factors may contribute to this rise, it has been shown that changes in dietary habits play a significant role in cancer development and progression. Epigenetic events are accepted as the most important mechanisms in the development and progression of cancer [38,39]. The connection between diet and cancer is partly due to the effects of diet on epigenetics. However, it is important to note that the field of epigenetics and its relationship to diet and cancer is still evolving, and further research is needed to fully understand the mechanisms and clinical implications. Collectively, several epigenetic events can be triggered by diet, some of which include:
DNA methylation: Methylation is a process in which a methyl group is added to DNA, which can silence gene expression. Diet can influence DNA methylation, affecting gene regulation and contributing to cancer development [40]. Cancer cells are characterized by genomic instability associated with hypomethylation in Cytosine Phosphorus Guanine (CpG) in intergenic regions and repetitive elements, but also by marked hypermethylation of CpG islands in tumor suppressor genes. Several studies have indicated that some miRNA genes reside near CpG islands, so their expression can be regulated by epigenetic modifications at these sites [41,42].
DNA methylation or methylation of cytosine in the context of the CpG dinucleotide has important consequences on our genome. When hypermethylation occurs in a tumor suppressor gene promoter, it silences the gene and allows tumor cells to grow in a manner similar to deletions or mutations [43]. DNA methylation errors, in terms of hypermethylation or hypomethylation, are critically involved in tumorigenesis [44].
Histone modification: Histones are proteins that help package DNA into a compact structure. Histone modifications, such as acetylation and methylation, can regulate gene expression. Diet can affect histone modifications, which in turn can influence cancer risk [45].
MicroRNAs: MicroRNAs are small RNA molecules that regulate gene expression. Diet can affect the expression of microRNAs, which in turn can influence the onset and progression of cancer [46].
There are numerous studies that have investigated the relationship between changes in dietary habits and epigenetic events in humans. It has been observed that certain nutrients and bioactive compounds present in foods, such as polyphenols, specific vitamins, and omega-3 fatty acids, can have epigenetic effects and modify cancer risk [47,48]. A recent review analyzes the effect of diet components on cancer with epigenetic mechanisms. This study focused on the analysis of bioactive dietary components such as resveratrol, genistein, quercetin, curcumin, isothiocyanates (sulforaphane in broccoli sprouts), tea polyphenols, selenium, withaferin A, indole-3-carbinol, and their role in reducing cancer risk [49].
In cancer cells, as previously commented, increasing attention has been devoted to new molecular mechanisms linking the epigenetic scenario with miRNA deregulation. Currently, growing evidence shows that nutritional and natural agents, known as nutraceuticals, could modulate miRNA expression, thereby influencing cellular responses in health and disease conditions, including cancer. For instance, in relation to prostate cancer, among dietary components, polyphenols play a protective role either due to their anti-aging and antioxidant properties or their more general “cell protection” effects [46].
Cellular deterioration or disruption of homeostasis is often related to oxidative stress caused by free radicals or reactive oxygen species (ROS). Free radicals are unstable molecules that are produced in the body as a result of natural processes and external factors such as exposure to ultraviolet radiation, environmental pollution, tobacco consumption, dietary factors, among many others. These free radicals can cause damage to DNA and other cellular molecules leading to mutations and epigenetic alterations that contribute to cancer development [50].
Antioxidants act as defenders against free radicals, neutralizing and preventing them from causing damage to cells [51]. By protecting DNA and other cellular molecules, antioxidants help prevent the occurrence of mutations and epigenetic changes [52]. These epigenetic modifications play a crucial role in gene activation or deactivation, and antioxidants can help maintain a proper balance in these processes which can have important implications in cancer prevention and treatment [53]. There are non-enzymatic compounds with a more pronounced secondary action, among which are vitamin C and E, carotenoids, flavonoids, phenols, polyphenols, phytoestrogens, selenium, lipoic acid, sulforaphanes, glucosinolates, and coenzyme Q10. In the SEAD, many foods are sources of these antioxidants and are consumed with interesting frequency.
In 2016, a group of researchers from the Department of Laboratory Medicine and the Department of Pediatrics at the University Hospital of Santiago de Compostela, Spain, initiated the GALIAT6+7 project. The main goal of this project was to examine the scientific evidence that could support the promotion of the Atlantic diet as a healthy choice, allowing its incorporation into preventive family health strategies in line with the cultural and gastronomic heritage of Europe’s Atlantic regions. The conclusion of the study was that adherence to the Atlantic diet has been associated with a decrease in body mass index (BMI) and an improvement in lipid profile, as revealed by the data from the GALIAT6+7 clinical trial. The GALIAT study was the first clinical trial to investigate the effects of the Atlantic diet on metabolic and cardiovascular health as well as adiposity. If the study hypothesis of this work is confirmed, this dietary pattern could be included in health promotion strategies.
3.2. Nature of the components of the Southern European Atlantic Diet (SEAD)
Many fruits and vegetables contain natural compounds that act as blocking or neutralizing agents against carcinogens. These compounds can contribute to cancer protection by preventing the formation of cancer cells or inhibiting their growth.
Phenols: Phenols are antioxidant compounds found in various fruits and vegetables. Phenols neutralize free radicals and protect cells against oxidative damage, which can reduce the risk of cancer. Phenolic compounds comprise three groups: simple phenols (e.g., tyrosol, hydroxytyrosol, or 3,4-dihydroxyphenylethanol), phenolic acids (e.g., caffeic acid), and flavonoids (e.g., quercetin) [54].
Indoles: Indoles are compounds present in cruciferous vegetables such as broccoli, kale, and cauliflower. Indoles have demonstrated anticancer properties by modulating the activity of certain enzymes responsible for detoxifying carcinogens and interfering with cell growth-related signaling pathways. The biological role of I3C (indole-3-carbinol) and DIM (3,3’-diindolylmethane) is particularly noteworthy [55].
Flavones: Flavones are a type of flavonoid found in citrus fruits, such as oranges and lemons, as well as certain vegetables and herbs. These compounds have antioxidant and anti-inflammatory properties, and have been shown to inhibit the proliferation of cancer cells and promote apoptosis (programmed cell death) [56,57].
Isothiocyanates: Isothiocyanates are compounds formed from glucosinolates present in cruciferous vegetables, such as broccoli, horseradish, and Brussels sprouts. Isothiocyanates have demonstrated anticancer effects by inhibiting the proliferation of cancer cells, inducing apoptosis, and blocking tumor formation.
All these compounds can protect in various ways:
Antioxidant action: Many of these compounds have antioxidant properties, which means they can neutralize free radicals and other reactive oxygen compounds that can damage DNA and cells. By reducing oxidative stress, these compounds can prevent cellular damage and reduce the risk of mutations that could lead to cancer [58,59].
Anti-inflammatory activity: Some of these blocking agents have anti-inflammatory properties, which may be important in cancer prevention. Chronic inflammation has been associated with an increased risk of cancer, and compounds with anti-inflammatory properties can help reduce inflammation and, therefore, decrease the risk of cancer cell development [60].
Modulation of detoxifying enzymes: Some of these compounds can modulate the activity of detoxifying enzymes in the body, such as phase II enzymes, which are involved in the elimination of carcinogens and other toxic compounds from the body. By increasing the activity of these enzymes, blocking compounds can help eliminate carcinogens before they can damage cells [61].
Influence on gene expression: Some of these compounds can affect the expression of genes related to cell proliferation, apoptosis (programmed cell death), and cellular signaling. These effects can help prevent tumor formation and growth [62].
Regulation of hormonal metabolism: Some blocking agents can influence hormonal metabolism and help balance the levels of hormones related to cancer development, such as estrogens. This can be especially relevant in the case of certain hormone-related cancers, such as breast cancer [63].
3.3. Analysis of the foods and key components in the anticancer activity of the SEAD
Table 1 presents a summary of the foods and active compounds of the SEAD and their relationship with anticancer or antioxidant properties analyzed in this study.
3.3.1. Mostly Consumed Fish and Seafood in the SEAD: Mussels, octopus and cod.
Spain is the second-largest producer of mussels (Mytilus galloprovincialis) in the world, and almost all of it is produced in Galicia. The quality of Galician mussels is extraordinary, and it has been recognized by the European Commission with the “Protected Designation of Origin “DOP Mejillón de Galicia’“ [64] as a mark of quality.
The consumption of fresh fish is high, with Asturias ranking as the first and Galicia the third autonomous community in Spain in terms of per capita consumption. As for seafood consumption in Galicia, according to the Food Consumption Report of Spain 2022, it is also very high, placing the region in the second position (Figure 5). Among these highly consumed food groups in the SEAD, we will focus on three of them due to their high consumption and their content of anticancer substances.
Mussels are rich in various components that are currently being investigated, among which betaines and selenium stand out.
Betaines: They are chemical compounds formed by a methyl group and an amino group (Figure 6) found in a variety of foods, such as beets, spinach, whole grains, and seafood. Betaine exists widely in animals and plants. Among plants, Lycium barbarum and legumes contain betaine. Sugar beet molasses is the main source of betaine. Among animals, the liver, spleen, and amniotic fluid of mollusks such as octopus, cuttlefish, and shrimp, as well as vertebrates (including humans), contain betaine. These compounds have been studied in relation to cancer and their potential effects. Some studies have explored the role of betaines in the prevention and treatment of cancer. It has been observed that betaines can have anticancer properties because they are believed to act as methyl group donors [65]. This means that they can transfer methyl groups to different molecules in the body, which can have beneficial effects on various metabolic pathways. For example, it has been observed that betaines can participate in DNA methylation and other molecules, which can regulate gene expression and other cellular processes [66]. The effects of trimethylglycine (TMG) on colorectal cancer cell lines have been investigated. The results showed that TMG inhibited cell proliferation and promoted apoptosis in cancer cells, suggesting a potential antitumor effect. The effects of TMG on breast cancer models were also examined. The results indicated that TMG reduced cell migration and invasion, in addition to inhibiting tumor growth in mice. In the case of mussels, the presence of different types of betaines has been identified (Figure 6), with trimethylglycine (TMG) being the most common [60]. Other related compounds, such as dimethylglycine (DMG) and sarcosine, have also been detected.
The betaines present in mussels have antioxidant properties, anti-inflammatory effects, and promote healthy cardiovascular function. Anserine is a dipeptide that contains betaine and is found in the muscle tissue of mussels, along with other amino acids. It has antioxidant properties and has been investigated for its potential involvement in muscle health [67]. Carnosine is a similar dipeptide (formed by the combination of the amino acids beta-alanine and L-histidine) that is found in various tissues and foods, including muscle and some meat products like octopus. The anticancer effects of Carnosine have been described. It inhibits breast, ovarian, colon, and leukemic cancer cell proliferation; upregulates the expression of pro-inflammatory molecules; modulates cytokine secretion and alters U937 differentiation and phenotype. These effects may have implications for a role for carnosine in anti-cancer therapy [68].
Selenium: The optimal dietary level of selenium ensures its proper antioxidant and anticancer activity. Special attention is due to the antioxidant activities of selenium compounds, especially selenoproteins [69], and their importance in antioxidant defense. It is worth noting that data on selenium’s anticancer properties are still controversial. Moreover, selenium compounds as chemotherapeutic agents are usually used at supranutritional doses [70]. The main metabolic reactions of organic and inorganic selenium compounds in humans [71,72] are shown in Scheme 1.
Galician octopus (Octopus vulgaris) is the main ingredient of one of the prominent dishes of the SEAD, “pulpo a la gallega” or also known as “pulpo a la feria.” This festive dish is prepared with whole cooked octopus (usually in copper pots) and is a common presence at festivals, fairs, and pilgrimages in Galicia. It is served on a wooden plate and is often accompanied by the famous “cachelos,” skin-on boiled potatoes, and generously drizzled with olive oil and sweet or spicy paprika. A significant percentage of the octopus consumed in Galicia, which is part of the SEAD, is not native to the region, and the majority comes from Morocco [73], a country that exports various seafood products to Spain.
Octopus is a source of several bioactive compounds that may have beneficial health properties. Some of the main bioactive compounds present in octopus are as follows:
Omega-3 and polyunsaturated fatty acids: Octopus is a source of omega-3 fatty acids, which have been associated with various health benefits, including protection against cancer. These fatty acids have anti-inflammatory properties and may influence cellular growth and proliferation mechanisms.
Taurine: It is an amino acid found in high concentrations in octopus. Taurine has been suggested to have antioxidant properties and may play a role in cancer prevention [74]. Recently, taurine not only mitigates the side effects of chemotherapy in cancer but also possesses antitumor properties, including inhibiting cancer cell proliferation and inducing apoptosis in certain cancers by differentially regulating proapoptotic and antiapoptotic proteins. The underlying molecular mechanism also suggested that taurine can be a potential clinical application in tumor therapy [75].
Polysaccharides: Octopuses and other cephalopods contain various types of polysaccharides, such as chitin and glucans. Some studies have investigated the immunomodulatory and antitumor properties of marine-origin polysaccharides, including those found in octopuses. Particularly, Glycosaminoglycans (GAGs) are an important component of the tumor microenvironment (TME) [76]. It has been observed that certain marine polysaccharides can stimulate the immune system and enhance the activity of immune cells, such as macrophages and lymphocytes, which could have implications in the response against cancer. In in vitro and animal studies, some marine-origin polysaccharides have demonstrated antitumor activity by inhibiting the growth of cancer cells, inducing apoptosis, and reducing angiogenesis (formation of new blood vessels in the tumor). Some studies have confirmed interesting anti-proliferative effects of glycosaminoglycans present in the Norwegian lobster [77] and sea squid [78]. More research is needed in this field.
Bioactive peptides: Some octopus species produce toxic secretions as a predatory and defense mechanism in specialized organs called posterior secretory glands. The bioactive substances in these secretions can be small molecules like histidine or peptides like tachykinins. Octopus venoms contain tachykinin peptides that, despite being isolated from an invertebrate, exhibit characteristics similar as vertebrate peptides. The actions of tachykinin peptides are mediated by one or more tachykinin receptors, which, in the case of vertebrates, are neurokinin 1 receptor (NK1R), neurokinin 2 receptor (NK2R), and neurokinin 3 receptor (NK3R). It was shown that the most potent form of tachykinin, Oct-TK-III, was not only the most anionically charged but also the most structurally stable [79,80].
In Octopus vulgaris, the taquikinins Oct-TK-I and Oct-TK-II are found, which are also active in vertebrates. However, a comparison of their relative effects in vertebrate and invertebrate tissues has not been conducted. Taquikinins are involved in central nervous system pathways that mediate pain, anxiety, motor coordination, and cognition. Therefore, the taquikinins in the venom of Octopus vulgaris could be beneficial for the treatment of various disorders, such as irritable bowel syndrome, asthma, chronic pain, depression, Parkinson’s disease, breast cancer, and lung cancer [79,81]. In other recent studies, such as that conducted by Maria P. Ikonomopoulou et al., a peptide isolated from and modified in the Australian species Octopus Kaurna (Figure 7), known as Octpep-1, was investigated [82,83]. They studied its anticancer profile and its potential as a drug against melanoma with mutation in the BRAF gene, the dominant form of this disease, and with minimal effect on fibroblasts. Melanoma is the main skin cancer and causes about 57,000 deaths worldwide. The Octpep-1 peptide found in the ink reduced tumor progression in melanoma xenografts in mice and zebrafish. Therefore, it mediates selective cytotoxicity in BRAF-mutated melanoma in vitro and prevents tumor progression in vivo, providing a basis for melanoma therapy [83].
During the last decade, several studies related to octopus extracts and their potential antiproliferative activity have been conducted. A study conducted by Carolina Moreno-Félix et al. in 2012 examined fractions from an organic extract of fresh octopus. (Paraoctopus limaculatus) for biological activities such as antimutagenic and antiproliferative properties using Salmonella tester strains TA98 and TA100 with metabolic activation. This study indicated that within several compounds in the lipid fraction of the octopus, a group of saturated and unsaturated fatty acids are responsible for the bioactivity against AFB1 mutagenicity or proliferation of murine cancer cells, or both. The isolation and identification of the actual antimutagenic and antiproliferative compounds in octopus are the focus of ongoing research [84].
In another study by Finaia and coworkers, it was shown for the first time that lipid extracts from octopus by-products (Octopus vulgaris) have antiproliferative and apoptotic effects on human breast cancer cell lines [85].
Martín S. Hernandez Zazueta et al. investigated the antiproliferative and anti-inflammatory effects of Octopus vulgaris ink extracts (hexane, ethyl acetate, dichloromethane and water extracts) in human colorectal cancer cells (HT-29/HCT116) and breast cancer cells (MDA-MB-231), as well as RAW 264.7 murine cells treated with Lipopolysaccharide (LPS). All extracts, except ethyl acetate, exhibited antiproliferative effects without being cytotoxic to healthy ARPE-19 and RAW 264.7 cells [86]. Furthermore, from the Octopus vulgaris ink extracts, an antiproliferative and proapoptotic activity was observed for a metabolite called N-(2-ozoazepa-n3-yl)-pyrrolidine-2-carboxamide, known as Ozopromide (OPC). This novel metabolite derived from Octopus vulgaris ink demonstrates proapoptotic effects on A549 lung cancer cells and inhibits pro-inflammatory markers [87]. Specifically, antiproliferative activity has been also observed in cell lines of certain cancer types such as prostate (22Rv1), lung (A549), and cervical epithelioid adenocarcinoma (HeLa) [88]. Additionally, Octopus vulgaris extracts containing OPC have shown anti-inflammatory activity in LPS-stimulated RAW 264.7 cells, highlighting the potential of Octopus vulgaris ink as a nutraceutical product or supplement with antitumoral action [87].
The OPC also exhibited antioxidant and anti-inflammatory activity by reducing the production of reactive oxygen species (ROS) and affecting proinflammatory cytokines, as confirmed by in silico studies (Figure 8), demonstrating moderate affinity between OPC and these markers. In silico ADMET prediction studies suggested a lower potential for acute toxicity for OPC after oral administration, although further studies are needed to confirm this. Continued research is necessary to investigate the effects of OPC in in vitro and in vivo models of cancer and inflammation [87].
Cod (Gadus morhua) is a characteristic fish in the gastronomy of the SEAD, particularly in the northern region of Portugal. It is rich in nutrients and can provide several health-beneficial components. Although a direct relationship between cod components and anticancer activity has not been established, it is important to note that a diet rich in fish, such as cod, has been associated with overall health benefits, including the prevention of certain types of cancer.
Cod liver oil is also rich in vitamin D. One teaspoon (5 mL) of cod liver oil provides 15 μg (600 IU) of vitamin D3, a recommended daily amount. A recent Meta-Analysis confirms the anticancer properties of vitamin D, as numerous findings indicate that diet components, including vitamin D, may exert chemo-preventive effects through alterations in microRNA (miRNA) expression [89].
Some of the beneficial components present in cod include:
Omega-3 fatty acids: Cod is an excellent source of omega-3 fatty acids, especially eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). These fatty acids have demonstrated anti-inflammatory effects and may help reduce the risk of developing certain types of cancer.
High-quality proteins: Cod is a source of high-quality proteins that provide the essential amino acids necessary for proper cellular function and repair.
Vitamins and minerals: Cod contains a variety of essential vitamins and minerals, such as vitamin D, vitamin B12, vitamin A, iodine, selenium, and phosphorus. These nutrients are important for maintaining a healthy immune system and proper cellular function.
3.3.2. Genuine vegetables of the SEAD: Galician turnip top, garlic, potato, and Padrón pepper
The Brassicaceae family, also known as the cruciferous family, comprises the most consumed vegetables within the SEAD, with Galicia standing out for its consumption compared to the rest of Spain. Some of the well-known plants and vegetables belonging to this family include: Cabbage (Brassica oleracea var. capitata), Kale (Brassica oleracea var. sabellica), Broccoli (Brassica oleracea var. italica), Cauliflower (Brassica oleracea var. botrytis), Brussels Sprouts (Brassica oleracea var. gemmifera), Radish (Raphanus sativus), Arugula (Eruca sativa), Mustard (Brassica juncea, Brassica nigra), Turnip (Brassica rapa), Watercress (Nasturtium officinale), Horseradish (Armoracia rusticana), Chinese Cabbage (Brassica rapa var. pekinensis), and Red-Leaf Kale (Brassica oleracea var. acephala). Turnip tops from Galicia are a food with a quality seal, the “IGP Grelos de Galicia” [90]. In the Portuguese part of the SEAD, another important vegetable from the Brassicaceae family is green kale. It is widely used in one of Portugal’s most typical dishes, “caldo verde,” where it is cut into strips and also used as a garnish. In Galicia, specifically, three species of the Brassica genus are cultivated: B. oleracea, B. napus, and B. rapa, for horticultural and fodder consumption (Figure 9) [91].
Each of the Brassica varieties has unique characteristics and nutritional benefits, but they all share certain similarities in their flower and fruit structures.
Turnip top (Brassica rapa subsp. rapa), also known as Galician turnip top, is a leafy green vegetable that belongs to the species Brassica rapa, which is part of the Brassicaceae family [92]. Turnip top is a commonly cultivated and consumed plant in the Galicia region of Spain, closely integrated into the SEAD, where it is used in traditional dishes such as caldo gallego, cocido gallego, and lacón con grelos (Galician stewed pork with turnip tops). Turnip top leaves are rich in nutrients and have a bitter and spicy taste. As part of the Brassicaceae family, turnip top contains glucosinolates and sulforaphanes, phytochemical compounds with beneficial health properties. Glucosinolates are natural compounds found in plants of the Brassicaceae family and break down into biologically active compounds, such as sulforaphanes, when cut, chewed, or processed. Glucosinolates present in turnip top include sinigrin, glucoraphanin, glucobrassicin, and glucoiberin, among others (Figure 10).
Glucoraphanin: One of the most common glucosinolates found in vegetables like broccoli, Brussels sprouts, and kale. It is converted to sulforaphane, a bioactive compound with antioxidant properties and potential anticancer activity.
Sinigrin: Found in vegetables like horseradish, mustard, and mustard seeds. It can be converted to allyl isothiocyanate, which has been associated with cardiovascular health benefits and antimicrobial properties.
Glucobrassicin: Found in vegetables like cauliflower, cabbage, and Brussels sprouts. It can be converted to indole-3-carbinol, which has been studied for its potential to regulate hormonal metabolism and exhibit anticancer effects.
Gluconasturtiin: Found in vegetables like watercress and arugula. It can be converted to phenylethyl isothiocyanate, which has shown antimicrobial and anti-inflammatory activity.
Glucoiberin: It is found particularly in the seeds and leaves of these plants.
All these compounds are converted into sulforaphanes through the action of the enzyme “myrosinase” when the plant cells are ruptured. Sulforaphanes are known for their antioxidant, anti-inflammatory, and potentially anticancer properties. The conversion process from glucosinolate to sulforaphane involves a series of biochemical steps:
In the plant, glucosinolates are stored in separate cellular compartments from the “myrosinase.” When plant tissues are damaged, such as when vegetables are cut or chewed, the myrosinase enzyme reacts with the glucosinolates. This interaction triggers a biochemical reaction known as hydrolysis, where myrosinase breaks the glucosidic bond of the glucosinolate and releases different products. It is important to consider that the process of converting glucosinolate to sulforaphane can be influenced by various factors, such as the plant variety, processing, and cooking. Therefore, when damage occurs in the plant, glucosinolates are degraded into a variety of hydrolytic products (catalyzed by myrosinase) that are responsible for almost all the biological activities of this class of compounds (Scheme 2). The process begins with the hydrolysis of the thioglucoside bond, resulting in the formation of glucose and an unstable aglucone.
In particular, one of the products released by the hydrolysis of glucosinolate is a compound called thioglucosinolate, which quickly reacts with a water molecule and forms an isothiocyanate, which is an active form of sulfur compound. Sulforaphane is an example of an isothiocyanate formed from certain glucosinolates present in vegetables like broccoli, turnip tops and arugula.
The resulting sulforaphane from the conversion of glucosinolate to isothiocyanate has various beneficial biological activities. Sulforaphane has been shown to have antioxidant, anti-inflammatory, and anticancer properties [94], through the activity of Phenethyl isothiocyanate, inhibiting the Signal Transducer and Activator of Transcription 3 (STAT3) activation in prostate cancer cells [95]. Another study analyzed the antioxidant activity of turnip top [92]. Furthermore, it can influence the expression of genes related to detoxification and cellular protection. Therefore, due to its glucosinolate content, turnip top is a product comparable and equivalent to broccoli.
The most common sulforaphane in vegetables of the Brassicaceae family, such as broccoli, Brussels sprouts, and cabbages, is 1-isothiocyanate-4-(methylsulfinyl)butane, (4-MSOB). The precursor of sulforaphane is found in the form of a glucosinolate known as glucoraphanin. When Brassicaceae vegetables are cut, chewed, or processed, the myrosinase enzyme present in them converts glucoraphanin into sulforaphane, which is the active compound. Other source of sulforaphane is the glucoerucin from the rocket salad, which is converted in erucin by the myrosinase and finally reduce to give sulforaphane (Scheme 3).
The quantity and specific composition of glucosinolates and sulforaphanes can vary depending on factors such as the plant variety, cultivation, processing, and food preparation. The highest content of total glucosinolates is found in turnip tops and turnip greens [96]. Additionally, the content of glucosinolates and sulforaphanes may decrease with cooking time and temperature during cooking, as the myrosinase enzyme can be inactivated by heat, which is why it is recommended to cook them “al dente”.
Regarding the qualitative and quantitative analysis of these components in Galician cabbages and turnip tops, a study conducted by the Galician company Rosaleira [37], as part of the GALIAT6+7 project, focused on the study of functional biomolecules in Galician food products through agro-biotechnological research, and yielded the following conclusions:
Both fresh useful parts and fresh residue presented identical profiles of glucosinolates in both turnip tops and cabbages. In turnip tops, aliphatic glucosinolates were the major compounds, with gluconapin being the predominant one. In cabbages, indolic compounds were the majority, with glucobrassicin and neoglucobrassicin being the major compounds [37].
All three types of samples analyzed: fresh useful, fresh residue, and blanched, presented the same profile of phenolic compounds. The useful parts and the leaves processed after cooking showed the highest concentrations of flavonoids, hydroxycinnamic acids, and total phenolic compounds, while the residues presented the lowest concentrations of both flavonoids and hydroxycinnamic acids. In turnip tops, 10 phenolic compounds were identified, with sinapic acid being the most abundant, followed by derivatives of Kaempferol and Isorhamnetin. In cabbages, 15 phenolic compounds were identified, including flavonoids and hydroxycinnamic acids.
In the cooking waters of cabbages and turnip tops, the highest content of total glucosinolates was found after approximately 1 hour from the start of cooking. Aliphatic and indolic glucosinolates in the cooking waters presented different behavior, with aliphatic glucosinolates decreasing while indolic glucosinolates increasing their concentration.
The mentioned study by A Rosaleira company concluded that the residues of cabbages and turnip tops have a high content of glucosinolates, indicating that these waste products are interesting sources of these compounds to revalue their use and industrial utilization. On the other hand, the leaves of cabbages and turnip tops subjected to industrial blanching lose almost all the content of glucosinolates as these compounds degrade with heat. The loss of glucosinolates during blanching can be attributed to several reasons, such as the enzymatic action of myrosinase and high temperature cooking, which can degrade the compounds. Additionally, the duration and specific blanching conditions can also influence the loss of glucosinolates. The residues of cabbages and turnip tops have lower concentrations of phenolic compounds than the fresh useful leaves. The fresh leaves of cabbages and turnip tops have a higher antioxidant capacity than the vegetable residues from the stems and older leaves although the residues still retain high concentrations, with approximately 25% loss compared to the useful leaves. The leaves of cabbages and turnip tops after industrial blanching preserve high values of antioxidant capacity, with minimal losses compared to the fresh useful material. The vegetable residues of cabbages and turnip tops, considered waste products during industrial packaging, constitute an important source of vitamins and minerals, with potassium content higher than even the leaves used for consumption. During blanching, the glucosinolates lost in the leaves of cabbages and turnip tops are gradually transferred to the cooking waters, reaching a maximum content after 45 minutes from the start of the industrial process. This would be the optimal time to collect the cooking waters for use in the agri-food industry [37].
Isothiocyanates and, particularly, sulforaphane have shown to influence epigenetically through various targets. A recent review on the role of sulforaphane as an anticancer agent emphasizes that this type of molecule can simultaneously modulate multiple cellular targets involved in carcinogenesis, including (1) modulating carcinogen-metabolizing enzymes and blocking the action of mutagens; (2) inhibition of cell proliferation and induction of apoptosis; and (3) inhibition of neo-angiogenesis and metastasis. Suforaphane targets cancer stem cells through modulation of nuclear factor kappa B (NF-κB), Sonic hedgehog (SHH), epithelial-mesenchymal transition, and Wnt/β-catenin pathways [97].
Therefore, some of the main epigenetic targets of sulforaphane are:
Histone deacetylases (HDACs): Sulforaphanes have been shown to affect histone modifications, such as acetylation and methylation, which can also influence gene activity. Sulforaphane has demonstrated inhibitory effects on histone deacetylases (HDACs), enzymes responsible for removing acetyl groups from histones [98,99]. By inhibiting these enzymes, sulforaphane can increase histone acetylation, which is associated with enhanced gene accessibility and increased gene expression.
DNA methyltransferases (DNMTs): Sulforaphane can also modulate the activity of DNMTs, enzymes responsible for DNA methylation. DNA methylation is a key epigenetic modification that can silence gene expression. It has been observed that sulforaphane can reduce DNMT activity, leading to a decrease in DNA methylation and potential reactivation of silenced genes [100]. Sulforaphanes may indirectly act by increasing the expression of demethylating enzymes, such as ten-eleven translocase (TET) [101], which are involved in the oxidation of methyl groups in DNA, potentially leading to reduced methylation in certain regions of the genome.
Sonic hedgehog (SHH): Dysregulation of the sonic hedgehog (Shh) signaling pathway has been associated with cancer stem cells (CSC) and implicated in the initiation of pancreatic [102], leukemia [103], and lung [104] cancers. Rodova and coworkers proposed that pancreatic cancer preventative effects of sulforaphane may result from inhibition of the Shh pathway. Thus, sulforaphane potentially represents an inexpensive, safe, and effective alternative for the management of pancreatic cancer [102].
Wnt/β-Catenin: Colorectal cancer is most frequently driven by hyperactive Wnt/β-catenin signaling. Sulforaphane has been shown to inhibit cell growth and blocks Wnt/β-catenin signaling of colorectal cancer cells [105].
MicroRNAs (miRNAs): It has been observed that sulforaphane can modulate the expression of certain miRNAs, which may have epigenetic effects by regulating the expression of specific genes [106].
These epigenetic targets of sulforaphane suggest its ability to influence gene expression and modulate key biological processes. Additionally, at the genetic level, sulforaphanes interact with transcription factors, which bind to DNA and regulate gene transcription. Sulforaphane can influence the activity of epigenetic transcription factors, such as nuclear factor erythroid 2-related factor 2 (Nrf2) [107,108], and hypoxia-inducible factor 1 (HIF-1) [109]. These transcription factors can modulate the expression of genes involved in epigenetic processes, such as antioxidant response and cellular adaptation to stress. Nuclear factor erythroid 2-related factor (Nrf2) is an important regulator of cellular antioxidant defenses. Sulforaphane can activate the Nrf2-ARE (antioxidant response element related to nuclear factor erythroid 2-related factor) signaling pathway, which is involved in antioxidant response and cellular protection against oxidative stress. By activating this pathway, sulforaphane promote the expression of a variety of genes encoding antioxidant and detoxifying enzymes, thereby enhancing the body’s ability to neutralize free radicals and other toxic compounds.
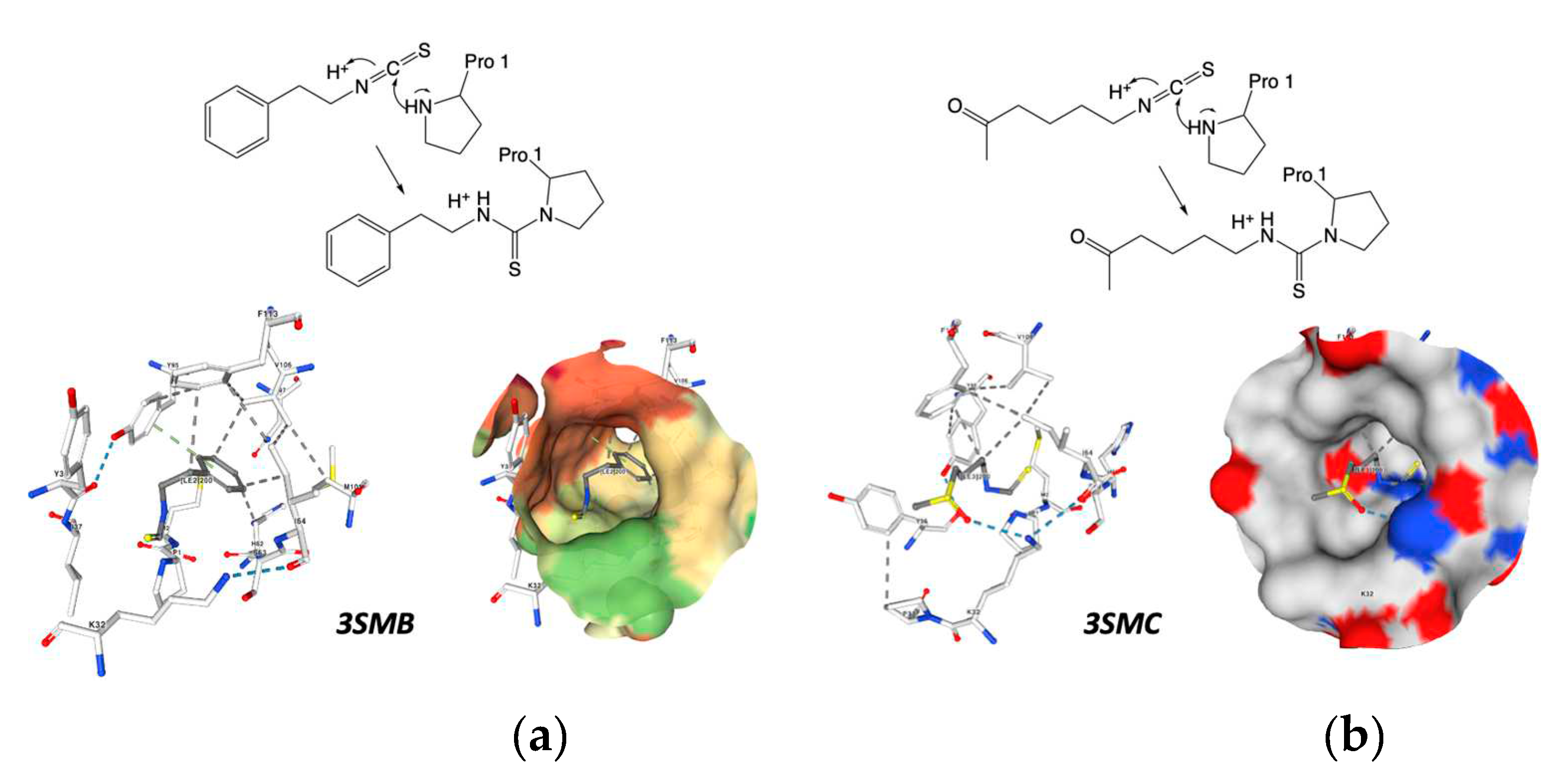
Isothiocyanates also target macrophage migration inhibitory factor (MIF), a widely distributed protein known for its inflammatory, pro-tumorigenic, pro-angiogenic, and anti-apoptotic properties. Isothiocyanates covalently inhibit MIF, providing insights into their cancer-preventive effects. Crichlow and coworkers present the crystallographic structures of human MIF bound to phenethylisothiocyanate and l-sulforaphane (Figure 11) [110].
Another predominant component in turnip tops is quercetin, a type of flavonoid (flavonol), with antioxidant activity found in a wide variety of plant-based foods (Figure 12). Quercetin has been shown to have antioxidant, anti-inflammatory, and antiproliferative properties [111,112], making it a bioactive compound with potential health benefits. Turnip tops are known to be a rich source of quercetin and other bioactive compounds. These compounds may contribute to the potential health benefits associated with turnip top consumption, such as protection against oxidative stress, reduction of inflammation, and support for the immune system.
Figure 11.
Crystallographic structures showing the interaction of: a) 3SMB crystal structure: Phenethylisothiocyanate Covalently Bound to Macrophage Migration Inhibitory Factor (MIF), pdb:. https://doi.org/10.2210/pdb3SMB/pdb b) 3SMC crystal structure: Macrophage Migration Inhibitory Factor (MIF) with Covalently Bound L-sulforaphane. PDB: https://doi.org/10.2210/pdb3SMC/pdb created with NGL [113].
Figure 11.
Crystallographic structures showing the interaction of: a) 3SMB crystal structure: Phenethylisothiocyanate Covalently Bound to Macrophage Migration Inhibitory Factor (MIF), pdb:. https://doi.org/10.2210/pdb3SMB/pdb b) 3SMC crystal structure: Macrophage Migration Inhibitory Factor (MIF) with Covalently Bound L-sulforaphane. PDB: https://doi.org/10.2210/pdb3SMC/pdb created with NGL [113].
Regarding quercetin’s binding substrates, this molecule has been found to interact with various proteins and enzymes involved in key biological processes. Some known binding substrates of quercetin include:
Protein kinases: Quercetin can inhibit the activity of certain protein kinases, which are enzymes involved in the regulation of cell growth and proliferation. This interference can affect cell signaling pathways and impact the growth of cancer cells.
ABC transporter proteins: Quercetin can interact with ATP-binding cassette (ABC) transporter proteins, which play a crucial role in drug resistance in cancer cells. By interfering with these proteins, quercetin may help overcome resistance to conventional treatments [114].
Inflammatory proteins: Quercetin has anti-inflammatory properties and can modulate the expression of inflammatory proteins, such as cytokines and transcription factors, which are involved in cancer progression [115].
Detoxification enzymes: Quercetin can induce the activity of detoxification enzymes, such as glutathione S-transferase (GST) enzymes, which play an important role in eliminating toxic compounds and carcinogens from the body [116].
Investigation into the utilization of anticancer agents that operate by enhancing the stability of telomeric G-quadruplex DNA has arisen as a fresh and captivating domain within the realm of discovering drugs for combating cancer. The G-quadruplex configuration plays a significant role in safeguarding the ends of chromosomes against undesired recombination and deterioration. Additionally, it hinders the operation of telomerase, the enzyme accountable for preserving telomere length. Protein Data Bank information about Quercetin shows that targeting human telomeric G-quadruplex DNA could be one of the mechanisms by which this flavonoid exert anticancer activity. Figure 12 shows a co-crystal of quercetin with Human Telomeric G-quadruplex DNA sequence (TTAGGGT)4 and how this molecule can interact with DNA. It interacts with the telomeric sequence through π-stacking at two sites: between T1pT2 and between G6pT7 [117].
On the other hand, other important bioactive flavonoid found in turnin tops is isorhamnetin. It is a methylated derivative of quercetin. It has antioxidant and anti-inflammatory properties similar to quercetin.
It is important to note that the exact levels of quercetin analogous in turnip tops may vary depending on various factors, such as the type of plant, cultivation methods, and maturity of the sprouts. However, overall, “turnip tops” are considered a good source of quercetins and other beneficial nutrients for health.
In addition to sulforaphanes and flavonoids, Diindolylmethane (DIM) and Indole-3-Carbinol (I3C) are two bioactive compounds found in brassicas such as broccoli, kale, cauliflower (widely consumed in the SEAD), and Brussels sprouts. These compounds are considered key components of brassicas due to their health-promoting properties and potential cancer-preventive effects [55,118,119]. I3C is a hydrolyzed glucosinolate product (Scheme 4). Cruciferous vegetables contain compounds like sulforaphane and I3C [120]. When exposed to stomach acid pH, I3C is transformed into various DIM products [121].
The interest in phytonutrients extracted from cruciferous vegetables dates back twenty years when it was observed that broccoli, DIM, and I3C prevented chemically induced breast cancer in animals. DIM is formed from I3C through a condensation reaction in the gastrointestinal tract during the digestion of brassicas. DIM has been extensively studied for its antioxidant, anti-inflammatory, and anticancer properties. They modulate different biological pathways involved in cell growth, proliferation, apoptosis, and angiogenesis, which may help prevent tumor formation and growth. On the other hand, I3C has also been the subject of numerous studies due to its anticancer properties. I3C can modulate the expression of genes related to detoxification and elimination of carcinogens, as well as inhibit the growth of cancer cells [123]. Additionally, I3C has shown anti-inflammatory and antioxidant effects that can contribute to its anticancer activity. Both DIM and I3C have demonstrated beneficial effects in the prevention and treatment of different types of cancer, such as breast cancer, prostate cancer, and colon cancer.
Garlic (Allium sativum) belongs to the Lily family (Liliaceae). It is cultivated for the bulb that forms at the base of the leaves and on the stem, which has a disc-like shape. Garlic is a widely used food in the SEAD, especially in the Portuguese region, where frying is a dominant culinary technique. It has numerous health-beneficial properties and has been extensively investigated for its potential health benefits, including the anticancer effects of some of its bioactive components (Scheme 5) [124,125]. Garlic contains several bioactive compounds, including:
Ajoene: An organosulfur compound associated with anticancer properties. Ajoene has been shown to inhibit the growth of cancer cells in vitro and in animal models [126,127].
Allicin: Another organosulfur compound that is formed when garlic is cut or crushed. Allicin has antioxidant properties and has demonstrated antitumor effects in preclinical studies [128].
Diallyl sulfide: A garlic-derived compound with antiproliferative and proapoptotic properties. It has been studied for its potential in preventing and treating cancer [129].
When garlic is mechanically crushed or cut, the enzyme alliinase stored in the vacuoles of the garlic cells is released. This enzyme breaks the carbon-sulfur bonds of alliin, transforming it into the thiosulfonate allicin, which is unstable. The instability of allicin leads to its rapid degradation into hundreds of sulfonated compounds.
The potato (Solanum tuberosum L.) is the tuber of the plant with the same name, an herbaceous plant belonging to the Solanaceae family. It is very typical in the cuisine of the SEAD, being a key ingredient in many well-known dishes worldwide. In Galicia, it is commonly used in dishes such as “cocido gallego,” “caldo gallego,” “carne o caldeiro,” and “caldeirada” of fishes. In the northern region of Portugal, it can be found in dishes like “Bacalhau à brás,” “Cozido à portuguesa,” or “caldo verde.” Potatoes are also featured in renowned dishes such as “Fabada” in Asturias or “Botillo” in El Bierzo.
Galicia is the Spanish autonomous community with the highest per capita consumption of potatoes, with individuals consuming 33.27 kilograms per year, compared to the national average of 28.89 kilograms. The cultivation of potatoes in Galicia is a Protected Geographical Indication (PGI), “IGP Patatas de Galicia” which bestows excellence on the potatoes produced in this region. Within this PGI, three main varieties are found: Kennebec, Agria, and Fina de Carballo [130].
Potatoes have been recognized by the Food and Agriculture Organization of the United Nations (FAO) as a staple and sustainable food for the growing global population [131]. Potatoes are rich in vitamin C, B-group vitamins, and heart-healthy minerals like potassium. The primary culinary technique in the SEAD is boiling, which is considered the healthiest and main method of preparing potatoes, leaving the skin on to retain its bioactive molecules.
Studies have also explored the antioxidant properties of potatoes [132]. In addition to vitamin C, potatoes are a source of other antioxidant molecules such as carotenoids and phenylpropanoids, and they contain a variety of secondary metabolites. Carotenoids are isoprenoid-based molecules synthesized in plastids with a polyene backbone consisting of conjugated C=C bonds.
The carotenoids content differs in each potato variety, which is evident in their color. Zeaxanthin is the carotenoid responsible for the orange color, while lutein is responsible for the yellow color (Figure 14). The Y locus encodes a β-carotene hydroxylase, which is a key determinant of the pulp color of the tuber [133,134]. Potatoes exhibit a great diversity of carotenoids, many of which are strong singlet oxygen quenchers and eliminators of other reactive oxygen species (ROS) [135].
In the northern Portuguese region, where dishes are often accompanied by white rice rather than potatoes, the more reddish-colored potato is preferred, with the Desiree variety being predominant, a result of mixed varieties. In general, potato cultivation in Portugal is characterized by the reddish-colored potato, a variety called Red Lady.
Galician Peppers (Capsicum annuum L.): In the Galician region of the SEAD, five native varieties of peppers have been awarded different quality seals: “IGP Pemento da Arnoia,” “IGP Pemento do Couto,” “IGP Pemento de Oímbra,” “IGP Pemento de Mougán,” and the famous Herbón peppers (Pimientos de Padrón or Padrón pepper) with the “IGP Pemento de Herbón” quality seal. The importance and relevance of these varieties are reflected in several quality seals, including four Protected Geographical Indications (IGP) and one Designation of Origin (DOP) for the Padrón peppers. They each have their respective regulatory boards that ensure the preservation of the quality of Galician peppers [136].
The Padrón pepper, of Mexican origin, arrived in Galicia through a pilgrimage carried out by Franciscan monks. It is known for its slightly spicy flavor, but in general, it is considered to have a mild taste. Capsaicin is a bioactive compound found in spicy peppers, including Padrón ones. It is responsible for their characteristic spicy flavor and has been studied for its potential beneficial health properties. Nevertheless, the capsaicin content is low compared to other spicier peppers. Capsaicin, dihydrocapsaicin, and some carotenoids are reported as the major active compounds with several pharmacological potentials, especially as anticancer and cardioprotectant [137] (Figure 15).
Padrón peppers also contain various antioxidants, such as carotenoids (like β-carotene), vitamin C, and vitamin E. These antioxidants help protect cells from oxidative stress and contribute to overall health. It is essential to note that the levels of components may vary depending on the Padrón pepper variety, growing conditions, and stage of maturity. In this variety of peppers, we can find capsaicin (close to 50%), dihydrocapsaicin (between 30-40%), nordihydrocapsaicin (less than 10%), homodihydrocapsaicin and homocapsaicin (residuals) (Figure 15).
Capsaicin has been the subject of numerous scientific studies, and it has been shown to possess antioxidant, anti-inflammatory, and analgesic properties. Additionally, there have been suggestions that capsaicin may have anticancer properties due to its ability to inhibit the growth of cancer cells and stimulate apoptosis (programmed cell death) in certain types of cancer [138,139,140,141,142], as mentioned above. Several in vitro and animal studies have investigated the effect of capsaicin on different types of cancer, including prostate [143], colon [144], lung, and breast cancers, and others [145]. These studies have shown promising results, but further research and clinical studies in humans are needed to confirm these findings. It has also been suggested that capsaicin could have inhibitory effects on angiogenesis, the process by which tumors develop new blood vessels to support their growth.
A study by Clark highlights that capsaicin exhibits strong anticancer activity by targeting multiple signaling pathways and cancer-associated genes in different tumor stages, including initiation, promotion, progression, and metastasis. The anticancer mechanisms of capsaicin include the activation of apoptosis, cell-growth arrest, and inhibition of angiogenesis and metastasis. Based on these results, the researchers concluded that capsaicin interacts synergistically with other cancer-preventive agents, providing the possibility for its potential use in cancer therapy along with other chemotherapeutic agents [146]. It is essential to note that the effects of capsaicin can vary depending on the dosage and the form of consumption. In the case of Padrón peppers, their content of capsaicin is generally low, which means that the potential health benefits associated with capsaicin may be limited in this specific case.
3.3.3. Native olive oil
Olea Europaea L arrived in Galicia during the Romanization of the Iberian Peninsula in the 1st century, finding in “Gallaecia” (a Roman province that encompassed the territories of the current autonomous community of Galicia, northern Portugal, and the territories of the current provinces of León, Zamora, and the autonomous community of Asturias) an ideal climate for olive cultivation. The native olive oil from Galicia, known as Galician olive oil, is a product of high quality and uniqueness produced in the northwest region of Spain. Although Galicia is not one of the main olive oil-producing regions in Spain, it has native olive varieties that adapt to its specific climate and soil, giving rise to olive oil with distinctive characteristics. Despite its limited production, olive oil consumption in the Atlantic northwest within the SEAD is high, even higher than in the Mediterranean diet, being a characteristic food of this diet. Currently, in Galicia, its consumption is 16 g per person per day [147].
In Galicia, a recent study carried out by the Biological Mission of Galicia belonging to the Spanish National Research Council (CSIC) has identified 20 olive varieties, making it an olive-growing region with a humid climate and characteristic soil, resulting in a variety of olives subject to international attention [148].
Two of the most prominent varieties are “Brava Gallega” and “Mansa Gallega.” Galician olive oil is produced in small quantities and is renowned for its quality, freshness, and unique organoleptic characteristics. Its cultivation is done on terraces and hillsides, taking advantage of the region’s topography. Additionally, an early harvest of the olives is carried out to preserve their properties and obtain higher-quality olive oil. This extra virgin olive oil, considered a gourmet product, has a high content of monounsaturated fatty acids and polyphenols, which confer antioxidant and anti-inflammatory properties. These bioactive compounds are mainly found in the unsaponifiable part and play a preventive role in cardiovascular diseases and reducing oxidative stress, proliferation of tumor cells, and cell cycle progression. Its consumption is recommended both raw, as a salad dressing, and to enhance the flavor of cooked dishes, such as fish, seafood, or vegetables.
In the Portuguese part of the SEAD, we find an extra virgin olive oil with a quality seal, the “DOP Trás-os-Montes.” In the region of Trás-os-Montes (Portugal), there is a high number of organic farmers, and the area also presents climatic, topographic, and edaphological differences that contribute to agricultural diversity. For cosmetic use, several studies have demonstrated potential antigenotoxicity in its ingredients.
Today, there is substantial evidence of the benefits of incorporating olive oil into the diet. Particularly, an excellent review on this topic has been published by Borzi and collaborators about olive oil effects on colorectal cancer. The interaction between gut microbiota and olive oil consumption could modulate colonic microbial composition or activity, with a possible role in cancer prevention [149]. Virgin olive oil (VOO) phenolic compounds, minor components of this fat, are known to be responsible for diverse health benefits when consumed in a regular diet. These benefits are mostly related to phenols such as tyrosol and hydroxytyrosol and secoiridoid derivatives such as ligstroside, oleuropein, oleocanthal, and oleacein, biomolecules found in extra virgin olive oil of the SEAD (Figure 16):
Hydroxytyrosol: A polyphenol present in extra virgin olive oil that has demonstrated antioxidant, anti-inflammatory, and anticancer properties [150,151,152]. It has been associated with the inhibition of cancer cell growth and induction of apoptosis (programmed cell death). A study showed that hydroxytyrosol has antiproliferative and proapoptotic properties in different tumor cells, and it suggests that extracellular production of hydrogen peroxide could be involved in these effects [153].
Oleocanthal: A phenolic compound found in extra virgin olive oil that has been associated with anti-inflammatory and anticancer properties. It has been shown that oleocanthal induces selective death of cancer cells without harming healthy cells. Experiments based on in vitro HT-29 human colon adenocarcinoma cells and in vivo chorioallantoic membrane assays showed an antitumor effect (due to increased AMPK) and apoptosis due to increased caspase-3 and poly-adenosine diphosphate-ribose polymerase, phosphorylation of p53 (Ser15)], and disruption of DNA [154].
Oleuropein: Another polyphenol present in extra virgin olive oil with antioxidant and anti-inflammatory properties. It has also been investigated for its anticancer potential due to its ability to inhibit the growth of tumor cells and stimulate apoptosis. In vitro studies using HT-29 human colon adenocarcinoma cells showed a decrease in cell proliferation after administration of this polyphenol to cells through activation of p53 pathway and #HIF-1 [155,156].
Ligstroside: A phenolic glucoside found in extra virgin olive oil. It has been shown that ligstroside has antioxidant properties and can help prevent the proliferation of cancer cells [157,158].
Squalene: A triterpene present in extra virgin olive oil that has demonstrated antioxidant and anticancer properties. Its potential for inhibiting the growth of cancer cells and preventing tumor formation has been investigated [159] confirming a chemopreventive behavior [160].
These molecules, along with other components of olive oil, work synergistically to provide health benefits and potential anticancer activity. It is essential to note that research in this field is still ongoing, and more studies are needed to fully understand the mechanisms of action and the effects of olive oil in cancer prevention and treatment [47,161,162].
3.3.4. Native Chestnut of the SEAD
The predominant chestnut variety in Galicia is known as “Castanea sativa,” commonly referred to as Spanish chestnut or Galician chestnut. The chestnut is one of the foods that have a quality seal, the “IGP Castaña de Galicia” (Protected Geographical Indication of Galician Chestnut). The preservation of traditional chestnut cultivation techniques in Galicia has allowed for the selection of a homogeneous set of native cultivars, contributing to the current fame and reputation of Galician chestnuts [163].
Chestnut consumption is a deeply rooted tradition in Galicia, especially during the autumn season. Chestnuts are consumed in various forms, whether roasted, boiled, mashed, or in sweet preparations such as desserts. Additionally, numerous festivals and fairs dedicated to chestnuts, such as the famous “Magosto,” are celebrated in Galicia, promoting their consumption and highlighting their cultural importance.
Chestnuts are a good source of complex carbohydrates, dietary fiber, proteins, vitamins (mainly vitamin C and some B-complex vitamins), and minerals such as potassium, selenium, magnesium, and phosphorus. They are also low in fat and cholesterol-free.
Chestnuts contain several bioactive components that have been of scientific interest (Figure 17). Many in vivo and in vitro studies have demonstrated how chestnut shell extract functions as an antioxidant and exhibits anticancer, anti-inflammatory, antidiabetic, and anti-obesity activities [164].
Some of the main bioactive components of chestnuts are:
Tannins: Chestnuts contain specific tannins known as hydrolysable tannins, which are a type of polyphenol. These tannins are mainly found in the shell or outer skin of the chestnut. Hydrolysable tannins have a complex chemical structure, as they are formed by glucose units linked to ellagic acid or its derivatives. These units can be joined in the form of dimers, trimers, or larger polymers. The hydrolysable tannins in chestnuts have antioxidant properties, allowing them to neutralize free radicals and protect cells from oxidative damage. Additionally, they have demonstrated potential as anticancer agents [165]. It is important to note that hydrolysable tannins in chestnuts are present in higher quantities in the outer shell, so direct consumption of chestnuts, such as roasted chestnuts, does not entail a significant intake of these compounds. Hydrolyzable tannins present in chestnuts are mainly composed of ellagic acid and its derivatives, such as gallic acid and hexahydroxydiphenic acid (HHDP). These tannins are responsible for the astringent and antioxidant properties of chestnuts.
Escin: It is a compound found in the seeds of the horse chestnut tree (Aesculus hippocastanum). It has been traditionally used in herbal medicine, and its properties and potential applications have been studied [166,167,168]. Escin has demonstrated anti-inflammatory and venotonic properties, that is, it can help strengthen blood vessels and reduce inflammation. Due to these properties, escin has been used in the treatment of venous disorders, such as varicose veins and chronic venous insufficiency. Escin is a group of steroidal saponins found in various plant species. Its potential anticancer properties have been investigated due to its anti-inflammatory, antioxidant, and antiproliferative properties [166,167]. Escin has also shown inhibitory effects on the growth of cancer cells in laboratory studies and animal models. It is believed to exert its anticancer action through different mechanisms, such as inducing apoptosis (programmed cell death), inhibiting angiogenesis (formation of new blood vessels in tumors), and suppressing inflammation [168].
Ellagic Acid: This is another phenolic compound present in chestnuts with antioxidant and anticancer properties [169]. Ellagic acid has been shown to have potential anticancer effects by inhibiting the growth of cancer cells and promoting apoptosis (programmed cell death) [170].
In addition to ellagic acid and hydrolyzable tannins, chestnuts contain other phenolic compounds and phytochemicals. Some of these compounds are:
Gallic Acid: It is a phenolic acid present in chestnuts and many other plants. It has antioxidant properties and has been associated with potential health benefits [171].
Quercetin: It is a flavonoid found in some varieties of chestnuts. As discussed earlier, quercetin is known by its antioxidant and anti-inflammatory activity, and it has been associated with various beneficial effects on health.
Rutin: Another flavonoid present in chestnuts. Rutin also possesses antioxidant properties and has been studied for its protective effects on blood vessels and its potential to improve circulation [172].
Vitamin C: Chestnuts also contain vitamin C, although in smaller amounts compared to other fruits. Vitamin C is an essential antioxidant that contributes to immune function and the maintenance of healthy tissues.
3.3.5. Autochthonous honey in the SEAD
The predominant autochthonous honey in the SEAD is known as “IGP Miel de Galicia” with its quality food seal. The peculiarities of Galicia’s climate, soil, and vegetation give rise to diverse honeys with very different characteristics and flavors [173]. It contains a variety of bioactive components, at least 180, although these vary depending on many factors. Some of these components have been studied for their potential anticancer properties (Figure 18). Honey contains several phenolic compounds, such as phenolic acid, flavonoids, and caffeic acid. These compounds have antioxidant and anti-inflammatory properties, which are believed to play a role in protection against cancer. Honey also contains bioactive peptides, some of which have shown anticancer activity, such as apamin [174]. Below are some of the most important biomolecules in honey with potential anticancer properties, some of which have already been co-crystallized with some of their anticancer targets:
Apamin: Although apamin is technically a neurotoxin produced by bees, it has been shown to have antitumor properties in in vitro studies and animal models [175]. However, more research is needed to fully understand its mechanism of action and its potential clinical application.
Among the flavonoids present in honey with anticancer activity, the following can be mentioned:
Quercetin: It is one of the most studied flavonoids in honey and has been associated with anticancer properties, as mentioned earlier.
Luteolin: Luteolin is a flavonoid identified in certain honeys. It is a polyphenolic compound belonging to the class of flavones. The amount and presence of luteolin in honey can vary depending on the botanical source of the honey, as different types of flowers produce honeys with distinct chemical profiles [176,177]. Recent data demonstrate that luteolin induces apoptotic cell death via antioxidant activity, acting as an anticancer agent against various types of human malignancies, including breast cancer [178]. Therefore, Luteolin can be considered a flavonoid with a multifaceted anticancer potential [179].
Apigenin: Another flavonoid found in honey and other plant sources is apigenin. It has been shown to possess anticancer activity by inhibiting angiogenesis (the formation of new blood vessels in tumors) and modulating different cell signaling pathways involved in the growth and proliferation of cancer cells, particularly protein kinase CK2 (‘casein kinase II’). The crystal structure of luteolin (Figure 19a) and apigenin (Figure 19b) in complex with the catalytic subunit of Zea mays CK2 has been solved. As seen, apigenin interacts with both the hinge region (Val116) and the positive area near Lys68 and the conserved water W1, the two main polar ligand anchoring points in the CK2 active site [180].
Kaempferol: Kaempferol-rich foods have been related to a decreased risk of developing certain types of cancers, including skin, liver, and colon cancers. Kaempferol is a dietary anticancer molecule with multiple mechanisms of action [181]. Some of these include apoptosis, cell cycle arrest at the G2/M phase, downregulation of epithelial-mesenchymal transition (EMT)-related markers, and inhibition of phosphoinositide 3-kinase/protein kinase B signaling pathways [182]. Wang reported on the mechanism of anticancer action and the potential clinical use of kaempferol in the treatment of breast cancer [183]. It has also been observed that kaempferol induces apoptosis in cancer cells and can inhibit their proliferation and migration. Death-associated protein kinase 1 (DAPK1) is a 160 kDa serine/threonine protein kinase that belongs to the Ca(2+)/calmodulin-dependent protein kinase subfamily [184] (Figure 20a).
Phenolic Acids: Honey contains several phenolic acids, such as caffeic acid and p-coumaric acid, which have also been investigated for their potential anticancer activity. These compounds have shown inhibitory effects on the growth of cancer cells and may possess antioxidant and anti-inflammatory properties. For example, caffeic acid significantly inhibits colony formation of human skin cancer cells. Topical application of caffeic acid to dorsal mouse skin significantly suppresses tumor incidence and volume in a solar UV (SUV)-induced skin carcinogenesis mouse model. Caffeic acid directly targets protein-serine/threonine kinase ERK1/2 inhibiting them in vitro. The cocrystal structure of ERK2 complexed with caffeic acid is shown in Figure 20b. These results suggest that caffeic acid exerts chemo-preventive activity against SUV-induced skin carcinogenesis [185].
3.3.6. Wines
Both Galicia and Northern Portugal are considered wine powerhouses in their respective regions. Both areas boast a long winemaking tradition, a wide variety of native grape varieties, and a climate and soils conducive to producing quality wines. This is why Galicia has five prestigious designations of origin, such as “DOP Rías Baixas”, “DOP Ribeiro”, “DOP Valdeorras”, “DOP Ribeira Sacra”, and “DOP Monterrey”, which have gained international recognition and great prestige [90]. In Northern Portugal, the most prominent designations of origin are Vinho Verde, Douro, and Dão. Vinho Verde is known for its light and refreshing white wines, while Douro is renowned for its production of Porto wines, as well as high-quality red wines. Dão is renowned for its elegant and structured red and white wines. Both regions have experienced significant growth in the quality and recognition of their wines in recent years. The diversity of microclimates and soils, combined with the passion of local winemakers and oenologists, has contributed to the success of these regions as wine powerhouses, attracting attention both nationally and internationally.
Within the framework of the SEAD, each designation of origin in Galicia has unique characteristics due to its Atlantic climate, ocean influence, and specific soils in each area. This offers a wide range of flavors and styles that reflect the diversity of Galicia as a wine region. Certain specific substances contribute to the mentioned profiles in Galician wines. These present a wide range of aromatic compounds, such as esters, aldehydes, terpenes, and phenolic compounds. These compounds come from the grape varieties used, such as Albariño, Treixadura, Caiño, Godello, Loureiro, or Mencía, among others [186], and they provide fruity, floral, and spicy notes to the wines. The acidity in Galician wines is primarily attributed to the presence of tartaric and malic acids. These natural acids contribute to the refreshing sensation in the mouth and the balance of the wine.
The granite and slate soils present in Galicia can influence the mineral profile of the wines. While there are no specific mineral substances directly responsible for this character, it is believed that the interaction of vine roots with these soils can influence mineral absorption and, ultimately, the expression of terroir in the wines. As a result, Galician wine contains a variety of bioactive components, including resveratrol (Figure 21), which is widely recognized for its beneficial health properties.
Native grape varieties, such as Albariño, Godello, and Mencía, have unique chemical compositions. For example, Albariño is characterized by a high content of aromatic compounds, such as geraniol and linalool (Figure 22), which contribute to distinctive floral notes. Godello is known for its good acidity and aromatic profile of white and citrus fruits. Mencía, used in red wines, presents phenolic compounds that provide color and structure to the wines.
Resveratrol is a polyphenol found in grape skins and has been associated with antioxidant, anti-inflammatory, and cardiovascular properties. The Mencía grape variety, one of the most cultivated red grapes in Galicia, is known to be particularly rich in resveratrol. Moderate consumption of wine rich in resveratrol has been suggested to contribute to reducing the risk of cardiovascular diseases and have positive effects on health, including anticancer therapy [187,188]. Particularly resveratrol has been the subject of numerous studies due to its potential anticancer activity [189]. Therefore, polyphenols of Galician wines have antioxidant properties and may contribute to cancer prevention through moderate wine consumption [190]. Some of the pharmacological targets identified for resveratrol include:
AMP-activated protein kinase (AMPK): Resveratrol can activate AMPK, an enzyme that regulates cellular metabolism and survival. AMPK activation can inhibit the growth of cancer cells and promote apoptosis (programmed cell death) [187].
Sirtuin family proteins: Resveratrol can activate sirtuins. In fact, a crystallographic structure, available at PDB, shows the Sirt5-trans-resveratrol interaction (Figure 21). Sirtuins are a class of enzymes involved in the regulation of aging and cellular metabolism [191]. Sirtuins can influence the expression of genes related to cancer and modulate different cellular processes. Sirtuin enzymes have been studied since then. This family of seven proteins known as the “longevity gene” plays an important role in the aging process, protecting cells and performing other functions in our body. Sirtuins protect against oxidative stress, repair our DNA making it more resistant to genetic toxicity, and activate metabolism to burn fat, i.e., they combat metabolic-energetic stress.
Nuclear factor kappa B (NF-κB): Resveratrol can inhibit the activation of NF-κB, a transcription factor that plays an important role in the regulation of the inflammatory response and cell survival [192]. Inhibition of NF-κB can reduce proliferation and promote apoptosis in cancer cells.
Estrogen receptor (ER): Resveratrol can interact with the estrogen receptor, which is involved in the growth and proliferation of certain hormone-sensitive cancers, such as breast cancer. Resveratrol can have estrogenic or antiestrogenic effects depending on the context and concentration [193].
In addition to resveratrol, Galician wine contains other polyphenols (Figure 22), such as flavonoids (such as catechins, proanthocyanidins, or quercetin), phenolic acids (ellagic acid), and stilbenes (such as caffeic and gallic acid), some of which have been mentioned previously.
In this regard, several studies have analyzed Galician wines. Figueiredo-González reported the evolution of anthocyanins and flavonols in red grapes of V. vinifera L. Mouratón, collected separately from the tip and shoulder positions of the bunch, and during over ripening [194]. Derivatives of five anthocyanins (malvidin, peonidin, petunidin, delphinidin, and cyanidin) and derivatives of six flavonols (quercetin, myricetin, kaempferol, laricitrin, isorhamnetin, and syringetin) were detected (Figure 22).
Another recent study by Sonia Sentellas and coworkers analyzed the combined extraction and purification process and demonstrated that malolactic fermentation lees of Albariño wines (white wine) are a valuable source of phenolic compounds, especially phenolic acids such as caftaric acid, cis and trans-coutaric acid (Figure 22) [195].
Mirás-Avalos reported the significance of amino acids in wine quality and their potential for classifying wines based on their variety. In this research, the amino acid compositions of Albariño, Godello, and Treixadura wines were analyzed. The study found that the most abundant amino acids present in these wines were proline, aspartic acid, glutamic acid, lysine, arginine, asparagine, alanine and histidine [196].
The grape seeds, along with the grape skins, are one of the parts that are discarded during the wine-making process, posing a challenge for many wineries. Scientific studies have shown that grape seeds contain bioactive compounds that could be beneficial for human health [197]. With the objective of utilizing these residues, scientists from the CSIC at the Biological Mission of Pontevedra have extracted oils from the grape seeds that remained as waste after the production of white and red wines from Galician grape varieties (vitis vinifera) such as Albariño, Caíño Blanco, Loureiro, and Mencía.
It has been found that each of these oils, depending on the grape variety they come from, has different concentrations of vitamin E, omega-3 fatty acids, and phenolic compounds called proanthocyanidins, with potential anticancer and antimetastatic effects [198], as well as favorable effects in diseases like diabetes. At the organoleptic level, some of these oils have been highly rated, and significant differences in aroma, taste, and color have been observed based on the variety of grape seeds they originate from.
Grape seed oil provides vitamin E and has a high concentration of linoleic acid (76%) and linolenic acid, essential fatty acids also known as Omega-6 and Omega-3, which are crucial for the synthesis of prostaglandins, substances necessary to reduce platelet aggregation and inflammation. It serves as an ally in maintaining cardiovascular health and helps in the prevention of hypertension, obesity, and diabetes [199].
3.3.7. Milk and dairy derivatives
The consumption of milk and its derivatives, especially cheese, is higher than that of the Mediterranean diet, with Galicia being one of the regions in Spain where per capita consumption is particularly elevated (Figure 23). Milk peptides are short sequences of amino acids naturally found in milk, which may have bioactive effects in the body. Some of these peptides have shown antitumor activity or are associated with anticancer properties:
Lactoferricin: Derived from lactoferrin, a protein present in milk, lactoferricin has demonstrated antitumor activity by inhibiting the growth of cancer cells and promoting apoptosis. It has also been observed to modulate the immune response and possess anti-inflammatory properties [200].
Casomorphins: These are opioid peptides found in casein, a protein present in milk. Some studies have suggested that casomorphins may have antiproliferative effects on cancer cells, especially in colon cancer [201]. However, further research is needed to fully understand their anticancer activity. The interaction of opioid peptides with cancer cells has been reported to exhibit anti-cancerous activity. Specifically, T47D human breast-cancer cells cultured with or without in culture express opioid receptors and opioid agonists. The effect of suppression of T47D cell proliferation by opioids is achieved via κ- and δ-opioid receptors. The mechanisms through which these opioid-like molecules stimulate the anti-cancer effect are probably via disassembling the actin microfilaments with the activation of focal adhesion kinase (FAK) phosphorylation and vinculin with substantial small GTPase Rac1 activation [202].
Alpha-lactalbumin: This peptide is present in the lactalbumin protein. Alpha-lactalbumin has been shown to possess antitumor properties by inhibiting the growth of cancer cells and promoting apoptosis [203]. It has also been suggested to modulate the immune response and exhibit anti-inflammatory properties [204]
3.3.8. Dietary Supplementation in the SEAD
The SEAD contains several bioactive molecules that provide great health benefits. Among them, we find hydroxytyrosol and oleic acid present in olive oil, a widely consumed food in this diet. A systematic review and meta-analysis evaluated the beneficial effects of dietary supplementation with olive oil, oleic acid, or hydroxytyrosol in metabolic syndrome [205]. There is also scientific evidence confirming the health benefits of supplementing the Mediterranean diet with extra virgin olive oil (EVOO). In a study with individuals at high cardiovascular risk, the incidence of major cardiovascular events was lower among those assigned to a diet supplemented with extra virgin olive oil or nuts compared to those on a low-fat diet [206]. In addition, a small crossover trial (Trial number: ISRCTN09220811) was conducted in the context of the “EUROLIVE” study with 200 participants who, after a two-week washout, assumed 25 mL of olive oil with different phenolic content three times a day for three weeks. The results indicated a linear decrease in the ratio of oxidative stress markers and total cholesterol/HDL-cholesterol matching the polyphenol content in olive oil [207].
The most characteristic condiment of the SEAD is red paprika, also known as paprika, which provides organoleptic and nutritional characteristics. It is present in many traditional dishes such as “pulpo a la gallega,” “carne o caldeiro,” or “caldeirada de pescado.” It is obtained from grinding dried red peppers, usually from the Capsicum annuum variety [208].
Red paprika contains various components that can provide health benefits due to their antioxidant properties, helping to neutralize free radicals and protect cells against oxidative damage, which may contribute to the development of diseases, including cancer. Some of these bioactive molecules include:
Capsaicinoids: These compounds are responsible for the spicy flavor of red paprika. Capsaicinoids have been shown to possess antioxidant, anti-inflammatory, and analgesic properties as already mentioned, and they can help improve cardiovascular health [209].
Carotenoids: Red peppers, from which red paprika is derived, are rich in carotenoids such as beta-carotene and capsanthin. These compounds act as antioxidants and are converted into vitamin A in the body, contributing to skin health, vision, and immune system function.
Vitamin C: Red paprika is a good source of vitamin C, an essential antioxidant that contributes to immune function, collagen formation, and protection against oxidative damage, as mentioned earlier.
In addition to paprika, some dishes in the SEAD are also supplemented with lard, which is consumed at higher rates in Galicia compared to the rest of Spain, giving a characteristic flavor to some of its dishes [210]. In northern Portugal, cinnamon or other herbs typical of the Mediterranean diet, such as cilantro, are also used.
Furthermore, the term “supplements” also encompasses dietary foods [211]. Dietary foods are those that have been modified in their original composition by adding, removing, or substituting some of their nutrients, such as carbohydrates, proteins, lipids, vitamins, and inorganic nutrients or minerals, and they are part of the usual diet.
Among the dietary foods modified by the food industry, many are tailored to address specific nutritional needs and requirements in certain conditions, such as gluten-free or low-gluten foods for celiac disease or gluten intolerance, lactose-free milk for lactose intolerance, and foods without added sugars for diabetes, or enriched with nutrients such as omega-3, vitamin D, calcium, or fiber.
There are also dietary supplements, which are products designed to complement the diet and provide additional nutrients. The population perceives them as safe, leading to increased self-prescription. These products come in various forms, such as tablets, capsules, powders, liquids, or bars, and they contain a variety of vitamins, minerals, herbs, amino acids, or other ingredients [212]. Different types of dietary supplements exist, with the most common being vitamins and minerals, which represent 50% of the supplements consumed in Europe [213]. They contain one or several essential vitamins and minerals, such as vitamin C, vitamin D, calcium, iron, among others. They are used to address specific nutritional deficiencies or to support certain life stages, such as pregnancy or lactation. It is advisable to be under medical supervision and to avoid self-prescription.
The consumption of probiotics has also increased. These are dietary supplements containing beneficial bacteria for the intestine. They help maintain a healthy balance of bacteria in the digestive tract and have been shown to have positive effects on intestinal health and immune function [121].
Regarding omega-3 fatty acids, it is important to remember that the SEAD is rich in fish and seafood, which are a source of this bioactive molecule.
Lastly, there are already antioxidant supplements on the market containing various bioactive molecules commonly found in the SEAD. For example, sulforaphane, found abundantly in vegetables of the Brassica oleracea genus, is marketed in doses of 20 mg, equivalent to almost three servings of broccoli (500 g). Hydroxytyrosol, highly present in extra virgin olive oil, is widely used in nutritional supplements in amounts ranging from 0.7 to 1.5 mg, prescribed for specific eye conditions such as macular degeneration or cataracts [214]. There are also supplements with a significant phytonutrient from brassicas, I3C, which acts as an anticancer molecule [215,216]. The most effective dose is 400 mg daily [217]. Finally, we find phenolic compounds extracted from seaweed that act as enzyme inhibitors and exhibit antimicrobial, antiviral, anticancer, antidiabetic, antioxidant, and anti-inflammatory activities [218].
5. Conclusions
Current dietary patterns, especially among young people in European countries, are undergoing significant changes. The influence of globalization, urbanization, modern lifestyles, and the availability of processed foods have led to an increase in the consumption of foods high in saturated fats, added sugars, ultra-processed foods, and a decrease in the consumption of fresh and healthy foods. This trend towards less healthy eating has concerning consequences for the health of young people and the population in general. The rise in obesity, cardiovascular diseases, diabetes, and other diet-related health issues are evidence of the need to take action to address this situation. In this context, there is a growing interest in returning to the Atlantic or Mediterranean dietary patterns, which have been associated with a range of health benefits.
These diets are characterized by high consumption of fruits, vegetables, legumes, whole grains, fish, dairy and dairy products, olive oil, and nuts, and moderate consumption of red meats and alcohol. The Atlantic diet has been extensively studied and has been shown to have positive effects in preventing chronic diseases, including cardiovascular diseases, type 2 diabetes, and some types of cancer. These eating patterns are rich in essential nutrients, antioxidants, healthy fatty acids, and bioactive compounds such as polyphenols and omega-3, which may have anti-inflammatory, antioxidant, and health-protective properties.
The review presented here gathers scientific evidence that demonstrates the connection between a healthy diet, exemplified by the SEAD, and the prevention of different types of cancer. Reverting to Atlantic dietary patterns involves promoting the consumption of fresh, minimally processed, and locally sourced foods while reducing the intake of processed and ultra-processed foods. This requires education and awareness about the health benefits of these eating patterns, as well as changes in the availability and accessibility of healthy foods.
Author Contributions
P.G.V, P.T and A.C.; methodology, P.G.V., A.C.; validation, P.T., A.C. and P.D.V.; formal analysis, P.G.V and A. C.; investigation, A.C.; resources, A.C.; data curation, A.C.; writing—original draft preparation, A.C. and P.G.V.; writing—review and editing, A. C and P.T.; visualization, P.T.; supervision, P.T. and A.C.; project administration, P.T. and A. C.; funding acquisition, P.T. and A. C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agencia Estatal de Investigación (AEI), project number PID2019–109517RB-I00 and the Xunta de Galicia ED431C 2022/18. ERDF funds are also acknowledged. This work was also financially supported by the Consellería de Cultura, Educación e Ordenación Universitaria of the Galician Government: EM2014/022 to A.C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization (WHO). Cancer, Https://Www.Who.Int/Health-Topics/Cancer#tab=tab_2, [Accessed 07/12/2023].
- International Agency for Research on Cancer; World Health Organization; Organización Panamericana de la Salud; Organización Mundial de la Salud Código Latinoamericano y Caribeño Contra El Cáncer, Https://Lacdev.Iarc.Who.Int, [Accessed 07/31/2023].
- Kliemann, N.; Al Nahas, A.; Vamos, E.P.; Touvier, M.; Kesse-Guyot, E.; Gunter, M.J.; Millett, C.; Huybrechts, I. Ultra-Processed Foods and Cancer Risk: From Global Food Systems to Individual Exposures and Mechanisms. Br J Cancer 2022, 127, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Levchenko, A. Epigenetics as a Mediator of Plasticity in Cancer. Science 2023, 379, eaaw3835. [Google Scholar] [CrossRef] [PubMed]
- Hardy, T.M.; Tollefsbol, T.O. Epigenetic Diet: Impact on the Epigenome and Cancer. Epigenomics 2011, 3, 503–518. [Google Scholar] [CrossRef]
- Aune, D. Plant Foods, Antioxidant Biomarkers, and the Risk of Cardiovascular Disease, Cancer, and Mortality: A Review of the Evidence. Advances in Nutrition 2019, 10, S404–S421. [Google Scholar] [CrossRef] [PubMed]
- Tejera-Pérez, C.; Sánchez-Bao, A.; Bellido-Guerrero, D.; Casanueva, F.F. The Southern European Atlantic Diet. Minerva Endocrinol 2021, 46. [Google Scholar] [CrossRef] [PubMed]
- The Atlantic Diet – Origin and Features. IJFS 2016, 5. [CrossRef]
- Leis Trabazo, R.; De Lamas Pérez, C.; Castro Pérez, X.; Solla, P. Dieta atlántica. Nutrición y gastronomía en Galicia. Nutr Hosp 2019. [Google Scholar] [CrossRef]
- Fundación Dieta Atlántica Bases Científicas de La Dieta Atlántica; Servizo de Publicacións e Intercambio Científico Campus Vida. Universidad de Santiago de Compostela, 2020; ISBN 978-94-17595-97-5.
- Carral, E.V.; Del Río, M.; López, Z. Gastronomy and Tourism: Socioeconomic and Territorial Implications in Santiago de Compostela-Galiza (NW Spain). IJERPH 2020, 17, 6173. [Google Scholar] [CrossRef]
- Alberto González-Garcés Santiso; Federico Vilas Martín, Xosé Antón Álvarez Salgado Ría de Vigo [Material Cartográfico] : Una Aproximación Integral al Ecosistema Marino de La Ría de Vigo; Instituto de Estudios Vigueses.; Vigo, 2008; ISBN 978-84-89599-34-5.
- Lorenzo, P.M.; Izquierdo, A.G.; Rodriguez-Carnero, G.; Fernández-Pombo, A.; Iglesias, A.; Carreira, M.C.; Tejera, C.; Bellido, D.; Martinez-Olmos, M.A.; Leis, R.; et al. Epigenetic Effects of Healthy Foods and Lifestyle Habits from the Southern European Atlantic Diet Pattern: A Narrative Review. Advances in Nutrition 2022, 13, 1725–1747. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación Informe Del Consumo Alimentario de España 2022. Available online: Https://Www.Mapa.Gob.Es/Eu/Alimentacion/Temas/Consumo-Tendencias/Informe-Consumo-2022-Baja-Res_tcm35-655390.Pdf (accessed on 31 July 2023).
- Axencia Galega da Calidade Alimentaria .Ministerio de Agricultura, Pesca y Alimentación Ternera Gallega. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp/Carnes/IGP_Ternera_Gallega.Aspx (accessed on 15 July 2023).
- Axencia Galega da Calidade Alimentaria .Ministerio de Agricultura, Pesca y Alimentación Vaca Gallega / Buey Gallego. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp/Carnes/IGP_Vaca_Gallega-Buey_Gallego.Aspx (accessed on 11 July 2023).
- Dirección General de Desarrollo Rural y Agroalimentación. Ministerio de Agricultura, Pesca y Alimentación. Calidad Diferenciada. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp, Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp (accessed on 31 July 2023).
- Dirección General de Desarrollo Rural y Agroalimentación. Ministerio de Agricultura, Pesca y Alimentación. Ternera Asturiana. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp/Carnes/IGP_Ternera_Asturiana.Aspx (accessed on 31 July 2023).
- Cooperativa Agrícola do Barroso, C. R. L. Direção-Geral de Agricultura e Desenvolvimento Rural Carne de Bovino Cruzado Dos Lameiros Do Barroso. Available online: Https://Tradicional.Dgadr.Gov.Pt/Pt/Cat/Carne/Carne-de-Bovino/552-Carne-de-Bovino-Cruzado-Dos-Lameiros-Do-Barroso-Igp (accessed on 31 July 2023).
- Ministerio de Agricultura, Pesca y Alimentación. Instituto Tecnológico Agrario (ITACYL) IGP Botillo Del Bierzo. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp/Embu_otros/IGP_botillo.Aspx (accessed on 31 July 2023).
- Cermeño, P. Marine Planktonic Microbes Survived Climatic Instabilities in the Past. Proc. R. Soc. B. 2012, 279, 474–479. [Google Scholar] [CrossRef]
- Rumin, J.; Nicolau, E.; Gonçalves De Oliveira Junior, R.; Fuentes-Grünewald, C.; Flynn, K.J.; Picot, L. A Bibliometric Analysis of Microalgae Research in the World, Europe, and the European Atlantic Area. Marine Drugs 2020, 18, 79. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Belik, A.A.; Kusaykin, M.I.; Malyarenko, O.S.; Zvyagintsevа, T.N.; Ermakova, S.P. Laminarans and 1,3-β-D-Glucanases. International Journal of Biological Macromolecules 2020, 163, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Martin, L. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Marine Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed]
- Rafael Tojo Sierra, Rosaura Leis Trabazo La Dieta Atlántica. El Pescado y Las Algas. Su Importancia En El Neurodesarrollo y La Función Cerebral; 2009th ed.; Sevizo de Publicacions e Intercambio Científico.Universidad de Santiago de Compostelade USC: Santiago de Compostela, 2009; ISBN 978-84-9887-033-6. [Google Scholar]
- Ha, A.W.; Na, S.J.; Kim, W.K. Antioxidant Effects of Fucoxanthin Rich Powder in Rats Fed with High Fat Diet. Nutr Res Pract 2013, 7, 475. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Malvar, M.; Benítez-Estévez, A.J.; Leis, R.; Sánchez-Castro, J.; Gude, F. Changes in Dietary Patterns through a Nutritional Intervention with a Traditional Atlantic Diet: The Galiat Randomized Controlled Trial. Nutrients 2021, 13, 4233. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Malvar, M.; Benítez-Estévez, A.J.; Sánchez-Castro, J.; Leis, R.; Gude, F. Effects of a Community-Based Behavioral Intervention with a Traditional Atlantic Diet on Cardiometabolic Risk Markers: A Cluster Randomized Controlled Trial (“The GALIAT Study”). Nutrients 2021, 13, 1211. [Google Scholar] [CrossRef]
- Asociación Ourensividad Asociación Ourensividad, Https://Ourensividad.Com, [Accessed 07/31/2023]; Ourense.
- Instituto Nacional de Estadística. Número de Personas Centenarias En Ourense y Lugo.Ministerio de Economía y Hacienda(2022). Https://Www.Ine.Es/Jaxi/Datos.Htm?Path=/T20/E245/P04/Provi/L0/&file=0ccaa003.Px.[Accessed 07/18/2023].
- Bojang, K.P.; Manchana, V. Nutrition and Healthy Aging: A Review. Curr Nutr Rep 2023. [Google Scholar] [CrossRef]
- Willcox, B.J.; Willcox, D.C.; Suzuki, M. Demographic, Phenotypic, and Genetic Characteristics of Centenarians in Okinawa and Japan: Part 1—Centenarians in Okinawa. Mechanisms of Ageing and Development 2017, 165, 75–79. [Google Scholar] [CrossRef]
- Carballo-Casla, A.; Ortolá, R.; García-Esquinas, E.; Oliveira, A.; Sotos-Prieto, M.; Lopes, C.; Lopez-Garcia, E.; Rodríguez-Artalejo, F. The Southern European Atlantic Diet and All-Cause Mortality in Older Adults. BMC Med 2021, 19, 36. [Google Scholar] [CrossRef]
- Guallar-Castillón, P.; Oliveira, A.; Lopes, C.; López-García, E.; Rodríguez-Artalejo, F. The Southern European Atlantic Diet Is Associated with Lower Concentrations of Markers of Coronary Risk. Atherosclerosis 2013, 226, 502–509. [Google Scholar] [CrossRef]
- Oliveira, A.; Lopes, C.; Rodríguez-Artalejo, F. Adherence to the Southern European Atlantic Diet and Occurrence of Nonfatal Acute Myocardial Infarction. The American Journal of Clinical Nutrition 2010, 92, 211–217. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, B.; Rivas-Casais, A.; Lorences-Touzón, R.; Piedrafita-Páez, N.; Muñoz-Ferreiro, N.; Vázquez-Odériz, L.; Romero-Rodríguez, Á. Adherence to and Knowledge about the Atlantic Diet Pattern in the Senior Population of the Galician Region (NW-Spain). Journal of Functional Foods 2022, 91, 105015. [Google Scholar] [CrossRef]
- GALIAT 6+7; Grupo de Genética, Mejora y Bioquímica de Brásicas - Empresa A Rosaleira Proyecto: Potenciación de Biomoléculas Funcionales En Productos Alimentarios de Origen Gallego a Través de La Investigación Agrobiotecnológica,2017. Available online: Https://Galiat6mas7.Com/Fotos/Biblioteca/2Objetivos.Resultados.Public.AR-Brasicas2017.Pdf (accessed on 31 July 2023).
- Sawan, C.; Herceg, Z. Histone Modifications and Cancer. In Advances in Genetics; Elsevier, 2010; Volume 70, pp. 57–85. ISBN 978-0-12-380866-0. [Google Scholar]
- Sapienza, C.; Issa, J.-P. Diet, Nutrition, and Cancer Epigenetics. Annu. Rev. Nutr. 2016, 36, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA Methylation: A Historical Perspective. Trends in Genetics 2022, 38, 676–707. [Google Scholar] [CrossRef]
- Riggs, A.D.; Jones, P.A. 5-Methylcytosine, Gene Regulation, and Cancer. In Advances in Cancer Research; Elsevier, 1983; Volume 40, pp. 1–30. ISBN 978-0-12-006640-7. [Google Scholar]
- Einav Nili, G.-Y.; Saito, Y.; Egger, G.; Jones, P.A. Cancer Epigenetics: Modifications, Screening, and Therapy. Annu. Rev. Med. 2008, 59, 267–280. [Google Scholar] [CrossRef]
- Robertson, K.D. DNA Methylation, Methyltransferases, and Cancer. Oncogene 2001, 20, 3139–3155. [Google Scholar] [CrossRef]
- Casalino, L.; Verde, P. Multifaceted Roles of DNA Methylation in Neoplastic Transformation, from Tumor Suppressors to EMT and Metastasis. Genes 2020, 11, 922. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Z.; Jia, J.; Du, T.; Zhang, N.; Tang, Y.; Fang, Y.; Fang, D. Overview of Histone Modification. In Histone Mutations and Cancer; Advances in Experimental Medicine and Biology; Fang, D., Han, J., Eds.; Springer Singapore: Singapore, 2021; Volume 1283, pp. 1–16. ISBN 9789811581038. [Google Scholar]
- Bosutti, A.; Zanconati, F.; Grassi, G.; Dapas, B.; Passamonti, S.; Scaggiante, B. Epigenetic and MiRNAs Dysregulation in Prostate Cancer: The Role of Nutraceuticals. ACAMC 2016, 16, 1385–1402. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 287–314. [Google Scholar] [CrossRef]
- Watier, T.; Mj Sanchez, A. Micro-RNAs, Exercise and Cellular Plasticity in Humans: The Impact of Dietary Factors and Hypoxia. MIRNA 2017, 6. [Google Scholar] [CrossRef]
- Özyalçin, B.; Sanlier, N. The Effect of Diet Components on Cancer with Epigenetic Mechanisms. Trends in Food Science & Technology 2020, 102, 138–145. [Google Scholar] [CrossRef]
- Álvarez Cruz, N. S.; Bague, A. J. El Cáncer. Orígenes, Prevención y Dieta Durante Su Tratamiento; Ediciones Madrid, 2012; ISBN 978-84-96709-94-2.
- Mladěnka, P. Special Issue Dietary (Poly)Phenols and Health. Nutrients 2022, 14, 1402. [Google Scholar] [CrossRef]
- Poljsak, B.; Milisav, I. The Role of Antioxidants in Cancer, Friends or Foes? CPD 2019, 24, 5234–5244. [Google Scholar] [CrossRef]
- Box, H.C.; Patrzyc, H.B.; Budzinski, E.E.; Dawidzik, J.B.; Freund, H.G.; Zeitouni, N.C.; Mahoney, M.C. Profiling Oxidative DNA Damage: Effects of Antioxidants. Cancer Sci 2012, 103, 2002–2006. [Google Scholar] [CrossRef] [PubMed]
- Rudzińska, A.; Juchaniuk, P.; Oberda, J.; Wiśniewska, J.; Wojdan, W.; Szklener, K.; Mańdziuk, S. Phytochemicals in Cancer Treatment and Cancer Prevention—Review on Epidemiological Data and Clinical Trials. Nutrients 2023, 15, 1896. [Google Scholar] [CrossRef]
- Williams, D.E. Indoles Derived From Glucobrassicin: Cancer Chemoprevention by Indole-3-Carbinol and 3,3’-Diindolylmethane. Front. Nutr. 2021, 8, 734334. [Google Scholar] [CrossRef] [PubMed]
- Pudenz, M.; Roth, K.; Gerhauser, C. Impact of Soy Isoflavones on the Epigenome in Cancer Prevention. Nutrients 2014, 6, 4218–4272. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-S.; Chiou, Y.-S.; Ho, C.-T.; Pan, M.-H. Chemoprevention by Resveratrol and Pterostilbene: Targeting on Epigenetic Regulation: Chemoprevention by Resveratrol and Pterostilbene. BioFactors 2018, 44, 26–35. [Google Scholar] [CrossRef]
- Nandini, D.; Rao, R.; Deepak, B.; Reddy, P. Sulforaphane in Broccoli: The Green Chemoprevention! ! Role in Cancer Prevention and Therapy. J Oral Maxillofac Pathol 2020, 24, 405. [Google Scholar] [CrossRef]
- Shimamoto, K.; Hayashi, H.; Taniai, E.; Morita, R.; Imaoka, M.; Ishii, Y.; Suzuki, K.; Shibutani, M.; Mitsumori, K. Antioxidant N-Acetyl-L-Cysteine (NAC) Supplementation Reduces Reactive Oxygen Species (ROS)-Mediated Hepatocellular Tumor Promotion of Indole-3-Carbinol (I3C) in Rats. J. Toxicol. Sci. 2011, 36, 775–786. [Google Scholar] [CrossRef]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in Inflammation: Mechanistic Aspects and Applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef] [PubMed]
- La Marca, M.; Beffy, P.; Della Croce, C.; Gervasi, P.G.; Iori, R.; Puccinelli, E.; Longo, V. Structural Influence of Isothiocyanates on Expression of Cytochrome P450, Phase II Enzymes, and Activation of Nrf2 in Primary Rat Hepatocytes. Food and Chemical Toxicology 2012, 50, 2822–2830. [Google Scholar] [CrossRef] [PubMed]
- Das, B.N.; Kim, Y.-W.; Keum, Y.-S. Mechanisms of Nrf2/Keap1-Dependent Phase II Cytoprotective and Detoxifying Gene Expression and Potential Cellular Targets of Chemopreventive Isothiocyanates. Oxidative Medicine and Cellular Longevity 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Wang, Z.-Y. Breast Cancer Cell Growth Inhibition by Phenethyl Isothiocyanate Is Associated with Down-Regulation of Oestrogen Receptor-A36. Journal of Cellular and Molecular Medicine 2009, 14, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Xunta de Galicia ORDEN de 20 de Marzo de 2019 Por La Que Se Aprueba El Reglamento de La Denominación de Origen Protegida Mexillón de Galicia-Mejillón de Galicia y Su Consejo Regulador. Available online: Https://Www.Xunta.Gal/Dog/Publicados/2019/20190408/AnuncioG0427-290319-0001_es.Html (accessed on 12 June 2023).
- Arumugam, M.K.; Paal, M.C.; Donohue, T.M.; Ganesan, M.; Osna, N.A.; Kharbanda, K.K. Beneficial Effects of Betaine: A Comprehensive Review. Biology 2021, 10, 456. [Google Scholar] [CrossRef]
- Van Puyvelde, H.; Dimou, N.; Katsikari, A.; Indave Ruiz, B.I.; Godderis, L.; Huybrechts, I.; De Bacquer, D. The Association between Dietary Intakes of Methionine, Choline and Betaine and Breast Cancer Risk: A Systematic Review and Meta-Analysis. Cancer Epidemiology 2023, 83, 102322. [Google Scholar] [CrossRef]
- Wu, G. Important Roles of Dietary Taurine, Creatine, Carnosine, Anserine and 4-Hydroxyproline in Human Nutrition and Health. Amino Acids 2020, 52, 329–360. [Google Scholar] [CrossRef]
- Prakash, M.D.; Fraser, S.; Boer, J.C.; Plebanski, M.; De Courten, B.; Apostolopoulos, V. Anti-Cancer Effects of Carnosine—A Dipeptide Molecule. Molecules 2021, 26, 1644. [Google Scholar] [CrossRef]
- Short, S.P.; Williams, C.S. Selenoproteins in Tumorigenesis and Cancer Progression. In Advances in Cancer Research; Elsevier, 2017; Volume 136, pp. 49–83. ISBN 978-0-12-812016-3. [Google Scholar]
- Kuršvietienė, L.; Mongirdienė, A.; Bernatonienė, J.; Šulinskienė, J.; Stanevičienė, I. Selenium Anticancer Properties and Impact on Cellular Redox Status. Antioxidants 2020, 9, 80. [Google Scholar] [CrossRef]
- Combs, G.F. Selenium in Global Food Systems. Br J Nutr 2001, 85, 517–547. [Google Scholar] [CrossRef]
- Rayman, M.P. The Use of High-Selenium Yeast to Raise Selenium Status: How Does It Measure Up? Br J Nutr 2004, 92, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Oficina Económica y Comercial de España en Rabat Informe Económico y Comercial En Marruecos. 2022. Available online: Https://Www.Icex.Es/Content/Dam/Es/Icex/Oficinas/097/Documentos/2022/09/Documentos-Anexos/DOC2022914490.Pdf (accessed on 31 July 2023).
- Choi, E.-J.; Tang, Y.; Lee, C.B.; Cheong, S.H.; Park, P.-J.; Moon, S.-H.; Kim, E.-K. Investigation of Antioxidant and Anticancer Potential of Taurine by Means of Multiple Chemical and Biological Assays. In Taurine 9; Advances in Experimental Medicine and Biology; Marcinkiewicz, J., Schaffer, S.W., Eds.; Springer International Publishing: Cham, 2015; Volume 803, pp. 179–189. ISBN 978-3-319-15125-0. [Google Scholar]
- Ma, N.; He, F.; Kawanokuchi, J.; Wang, G.; Yamashita, T. Taurine and Its Anticancer Functions: In Vivo and In Vitro Study. In Taurine 12; Advances in Experimental Medicine and Biology; Schaffer, S.W., El Idrissi, A., Murakami, S., Eds.; Springer International Publishing: Cham, 2022; Volume 1370, pp. 121–128. ISBN 978-3-030-93336-4. [Google Scholar]
- Wieboldt, R.; Läubli, H. Glycosaminoglycans in Cancer Therapy. American Journal of Physiology-Cell Physiology 2022, 322, C1187–C1200. [Google Scholar] [CrossRef] [PubMed]
- Sayari, N.; Balti, R.; Ben Mansour, M.; Ben Amor, I.; Graiet, I.; Gargouri, J.; Bougatef, A. Anticoagulant Properties and Cytotoxic Effect against HCT116 Human Colon Cell Line of Sulfated Glycosaminoglycans Isolated from the Norway Lobster ( Nephrops Norvegicus ) Shell. Biomedicine & Pharmacotherapy 2016, 80, 322–330. [Google Scholar] [CrossRef]
- Willis, C.M.; Klüppel, M. Chondroitin Sulfate-E Is a Negative Regulator of a Pro-Tumorigenic Wnt/Beta-Catenin-Collagen 1 Axis in Breast Cancer Cells. PLoS ONE 2014, 9, e103966. [Google Scholar] [CrossRef]
- Ruder, T.; Ali, S.A.; Ormerod, K.; Brust, A.; Roymanchadi, M.-L.; Ventura, S.; Undheim, E.A.B.; Jackson, T.N.W.; Mercier, A.J.; King, G.F.; et al. Functional Characterization on Invertebrate and Vertebrate Tissues of Tachykinin Peptides from Octopus Venoms. Peptides 2013, 47, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Maggi, C.A. The Mammalian Tachykinin Receptors. General Pharmacology: The Vascular System 1995, 26, 911–944. [Google Scholar] [CrossRef] [PubMed]
- Khawaja, A.M.; Rogers, D.F. Tachykinins: Receptor to Effector. The International Journal of Biochemistry & Cell Biology 1996, 28, 721–738. [Google Scholar] [CrossRef]
- Moral-Sanz, J.; Fernandez-Rojo, M.A.; Potriquet, J.; Mukhopadhyay, P.; Brust, A.; Wilhelm, P.; Smallwood, T.B.; Clark, R.J.; Fry, B.G.; Alewood, P.F.; et al. ERK and MTORC1 Inhibitors Enhance the Anti-Cancer Capacity of the Octpep-1 Venom-Derived Peptide in Melanoma BRAF(V600E) Mutations. Toxins 2021, 13, 146. [Google Scholar] [CrossRef]
- Moral-Sanz, J.; Fernandez-Rojo, M.A.; Colmenarejo, G.; Kurdyukov, S.; Brust, A.; Ragnarsson, L.; Andersson, Å.; Vila, S.F.; Cabezas-Sainz, P.; Wilhelm, P.; et al. The Structural Conformation of the Tachykinin Domain Drives the Anti-tumoural Activity of an Octopus Peptide in Melanoma BRAF V600E. British J Pharmacology 2022, 179, 4878–4896. [Google Scholar] [CrossRef]
- Moreno-Félix, C.; Wilson-Sánchez, G.; Cruz-Ramírez, S.-G.; Velázquez-Contreras, C.; Plascencia-Jatomea, M.; Acosta, A.; Machi-Lara, L.; Aldana-Madrid, M.-L.; Ezquerra-Brauer, J.-M.; Rocha-Alonzo, F.; et al. Bioactive Lipidic Extracts from Octopus ( Paraoctopus Limaculatus ): Antimutagenicity and Antiproliferative Studies. Evidence-Based Complementary and Alternative Medicine 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Fitahia, E.M.; Raheriniaina, C.E.; Bazin, M.A.; Huvelin, J.-M.; Logé, C.; Ranaivoson, E.; Nazih, H. Anti-Proliferative and Pro-Apoptotic Effect of Dichloromethane Extract of Octopus Vulgaris By-Products on Human Breast Cancer Cell Lines. Waste Biomass Valor 2015, 6, 237–242. [Google Scholar] [CrossRef]
- Hernández-Zazueta, M.S.; Luzardo-Ocampo, I.; García-Romo, J.S.; Noguera-Artiaga, L.; Carbonell-Barrachina, Á.A.; Taboada-Antelo, P.; Campos-Vega, R.; Rosas-Burgos, E.C.; Burboa-Zazueta, M.G.; Ezquerra-Brauer, J.M.; et al. Bioactive Compounds from Octopus Vulgaris Ink Extracts Exerted Anti-Proliferative and Anti-Inflammatory Effects in Vitro. Food and Chemical Toxicology 2021, 151, 112119. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Zazueta, M.S.; García-Romo, J.S.; Luzardo-Ocampo, I.; Carbonell-Barrachina, Á.A.; Taboada-Antelo, P.; Rosas-Burgos, E.C.; Ezquerra-Brauer, J.M.; Martínez-Soto, J.M.; Candia-Plata, M. del C.; Santacruz-Ortega, H. del C.; et al. N-(2-Ozoazepan-3-Yl)-Pyrrolidine-2-Carboxamide, a Novel Octopus Vulgaris Ink-Derived Metabolite, Exhibits a pro-Apoptotic Effect on A549 Cancer Cell Line and Inhibits pro-Inflammatory Markers. Food and Chemical Toxicology 2023, 177, 113829. [Google Scholar] [CrossRef]
- Hernández-Zazueta, M.S.; García-Romo, J.S.; Noguera-Artiaga, L.; Luzardo-Ocampo, I.; Carbonell-Barrachina, Á.A.; Taboada-Antelo, P.; Campos-Vega, R.; Rosas-Burgos, E.C.; Burboa-Zazueta, M.G.; Ezquerra-Brauer, J.M.; et al. Octopus Vulgaris Ink Extracts Exhibit Antioxidant, Antimutagenic, Cytoprotective, Antiproliferative, and Proapoptotic Effects in Selected Human Cancer Cell Lines. Journal of Food Science 2021, 86, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Zeljic, K.; Supic, G.; Magic, Z. New Insights into Vitamin D Anticancer Properties: Focus on MiRNA Modulation. Mol Genet Genomics 2017, 292, 511–524. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y alimentación Listado de Denominaciones de Origen Protegidas e Indicaciones Geográficas Protegidas de Vinos Registradas En La Unión Europea. Available online: Https://Www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/02_vinos_tcm30-426473.Pdf (accessed on 17 July 2023).
- Marta Francisco; Margarita Lema; Pablo Velasco Cultivo de Variedades Tradicionales de Brásicas En La Agricultura Ecológica. Available online: Https://Digital.Csic.Es/Bitstream/10261/87344/6/Cartea_Cultivo_variedades_tradicionales.pdf (accessed on 1 July 2023).
- Cao, Q.; Wang, G.; Peng, Y. A Critical Review on Phytochemical Profile and Biological Effects of Turnip (Brassica Rapa L.). Front. Nutr. 2021, 8, 721733. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrijver, R.; Hansen, M.; Gerhäuser, C.; Mithen, R.; et al. Glucosinolates in Brassica Vegetables: The Influence of the Food Supply Chain on Intake, Bioavailability and Human Health. Mol. Nutr. Food Res. 2009, 53, S219–S219. [Google Scholar] [CrossRef]
- Nuñez-Iglesias, M. j.; Novio, S.; García-Santiago, C.; Cartea, M. e.; Soengas, P.; Velasco, P.; Freire-Garabal, M. Effects of 3-Butenyl Isothiocyanate on Phenotypically Different Prostate Cancer Cells. Int J Oncol 2018. [Google Scholar] [CrossRef]
- Gong, A.; He, M.; Krishna Vanaja, D.; Yin, P.; Karnes, R.J.; Young, C.Y.F. Phenethyl Isothiocyanate Inhibits STAT3 Activation in Prostate Cancer Cells. Mol. Nutr. Food Res. 2009, 53, 878–886. [Google Scholar] [CrossRef]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of Glucosinolates in Cruciferous Vegetables Grown at the Same Site for Two Years under Different Climatic Conditions. J Agric Food Chem 2000, 48, 2862–2867. [Google Scholar] [CrossRef]
- Elkashty, O.A.; Tran, S.D. Sulforaphane as a Promising Natural Molecule for Cancer Prevention and Treatment. CURR MED SCI 2021, 41, 250–269. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, S.M.; Royce, S.G.; Licciardi, P.V.; Karagiannis, T.C. Dietary Sulforaphane in Cancer Chemoprevention: The Role of Epigenetic Regulation and HDAC Inhibition. Antioxidants & Redox Signaling 2015, 22, 1382–1424. [Google Scholar] [CrossRef]
- Jiang, L.-L.; Zhou, S.-J.; Zhang, X.-M.; Chen, H.-Q.; Liu, W. Sulforaphane Suppresses in Vitro and in Vivo Lung Tumorigenesis through Downregulation of HDAC Activity. Biomedicine & Pharmacotherapy 2016, 78, 74–80. [Google Scholar] [CrossRef]
- Su, X.; Jiang, X.; Meng, L.; Dong, X.; Shen, Y.; Xin, Y. Anticancer Activity of Sulforaphane: The Epigenetic Mechanisms and the Nrf2 Signaling Pathway. Oxidative Medicine and Cellular Longevity 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Dashwood, R.H. Epigenetic Regulation of NRF2/KEAP1 by Phytochemicals. Antioxidants 2020, 9, 865. [Google Scholar] [CrossRef]
- Rodova, M.; Fu, J.; Watkins, D.N.; Srivastava, R.K.; Shankar, S. Sonic Hedgehog Signaling Inhibition Provides Opportunities for Targeted Therapy by Sulforaphane in Regulating Pancreatic Cancer Stem Cell Self-Renewal. PLoS ONE 2012, 7, e46083. [Google Scholar] [CrossRef]
- Wang, F.; Huang, X.; Sun, Y.; Li, Z.; Sun, R.; Zhao, T.; Wang, M.; Yan, C.; Liu, P. Sulforaphane Regulates the Proliferation of Leukemia Stem-like Cells via Sonic Hedgehog Signaling Pathway. European Journal of Pharmacology 2022, 919, 174824. [Google Scholar] [CrossRef]
- Wang, F.; Sun, Y.; Huang, X.; Qiao, C.; Zhang, W.; Liu, P.; Wang, M. Sulforaphane Inhibits Self-Renewal of Lung Cancer Stem Cells through the Modulation of Sonic Hedgehog Signaling Pathway and Polyhomeotic Homolog 3. AMB Expr 2021, 11, 121. [Google Scholar] [CrossRef]
- Bernkopf, D.B.; Daum, G.; Brückner, M.; Behrens, J. Sulforaphane Inhibits Growth and Blocks Wnt/β-Catenin Signaling of Colorectal Cancer Cells. Oncotarget 2018, 9, 33982–33994. [Google Scholar] [CrossRef]
- Rafiei, H.; Ashrafizadeh, M.; Ahmadi, Z. MicroRNAs as Novel Targets of Sulforaphane in Cancer Therapy: The Beginning of a New Tale? Phytotherapy Research 2020, 34, 721–728. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka-Woźniak, K.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 Targeting by Sulforaphane: A Potential Therapy for Cancer Treatment. Critical Reviews in Food Science and Nutrition 2018, 58, 1391–1405. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Palliyaguru, D.L.; Kensler, T.W. Frugal Chemoprevention: Targeting Nrf2 with Foods Rich in Sulforaphane. Seminars in Oncology 2016, 43, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Sung, B.; Kang, Y.J.; Hwang, S.Y.; Kim, M.J.; Yoon, J.-H.; Im, E.; Kim, N.D. Sulforaphane Inhibits Hypoxia-Induced HIF-1α and VEGF Expression and Migration of Human Colon Cancer Cells. International Journal of Oncology 2015, 47, 2226–2232. [Google Scholar] [CrossRef] [PubMed]
- Crichlow, G.V.; Fan, C.; Keeler, C.; Hodsdon, M.; Lolis, E.J. Structural Interactions Dictate the Kinetics of Macrophage Migration Inhibitory Factor Inhibition by Different Cancer-Preventive Isothiocyanates. Biochemistry 2012, 51, 7506–7514. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. IJMS 2019, 20, 3177. [Google Scholar] [CrossRef]
- Tang, S.-M.; Deng, X.-T.; Zhou, J.; Li, Q.-P.; Ge, X.-X.; Miao, L. Pharmacological Basis and New Insights of Quercetin Action in Respect to Its Anti-Cancer Effects. Biomedicine & Pharmacotherapy 2020, 121, 109604. [Google Scholar] [CrossRef]
- Rose, A.S.; Bradley, A.R.; Valasatava, Y.; Duarte, J.M.; Prlic, A.; Rose, P.W. NGL Viewer: Web-Based Molecular Graphics for Large Complexes. Bioinformatics 2018, 34, 3755–3758. [Google Scholar] [CrossRef]
- Li, Y.; Revalde, J.; Paxton, J.W. The Effects of Dietary and Herbal Phytochemicals on Drug Transporters. Advanced Drug Delivery Reviews 2017, 116, 45–62. [Google Scholar] [CrossRef]
- Marefati, N.; Ghorani, V.; Shakeri, F.; Boskabady, M.; Kianian, F.; Rezaee, R.; Boskabady, M.H. A Review of Anti-Inflammatory, Antioxidant, and Immunomodulatory Effects of Allium Cepa and Its Main Constituents. Pharmaceutical Biology 2021, 59, 285–300. [Google Scholar] [CrossRef]
- Wiegand, H.; Boesch-Saadatmandi, C.; Regos, I.; Treutter, D.; Wolffram, S.; Rimbach, G. Effects of Quercetin and Catechin on Hepatic Glutathione-S Transferase (GST), NAD(P)H Quinone Oxidoreductase 1 (NQO1), and Antioxidant Enzyme Activity Levels in Rats. Nutrition and Cancer 2009, 61, 717–722. [Google Scholar] [CrossRef]
- Tawani, A.; Kumar, A. Structural Insight into the Interaction of Flavonoids with Human Telomeric Sequence. Sci Rep 2015, 5, 17574. [Google Scholar] [CrossRef] [PubMed]
- Megna, B.W.; Carney, P.R.; Nukaya, M.; Geiger, P.; Kennedy, G.D. Indole-3-Carbinol Induces Tumor Cell Death: Function Follows Form. Journal of Surgical Research 2016, 204, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, N.; Fritz, V.; Upadhyaya, P.; Kassie, F.; Hecht, S.S. Research on Cruciferous Vegetables, Indole-3-Carbinol, and Cancer Prevention: A Tribute to Lee W. Wattenberg. Mol Nutr Food Res 2016, 60, 1228–1238. [Google Scholar] [CrossRef]
- Royston, K.J.; Tollefsbol, T.O. The Epigenetic Impact of Cruciferous Vegetables on Cancer Prevention. Curr Pharmacol Rep 2015, 1, 46–51. [Google Scholar] [CrossRef]
- Banerjee, S.; Kong, D.; Wang, Z.; Bao, B.; Hillman, G.G.; Sarkar, F.H. Attenuation of Multi-Targeted Proliferation-Linked Signaling by 3,3′-Diindolylmethane (DIM): From Bench to Clinic. Mutation Research/Reviews in Mutation Research 2011, 728, 47–66. [Google Scholar] [CrossRef]
- Błoch-Mechkour, A.; Bally, T.; Sikora, A.; Michalski, R.; Marcinek, A.; Gȩbicki, J. Radicals and Radical Ions Derived from Indole, Indole-3-Carbinol and Diindolylmethane. J. Phys. Chem. A 2010, 114, 6787–6794. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, C.; Wu, D.; Zhang, Y.; Zhao, Y.; Li, W. Indole-3-Carbinol Stabilizes P53 to Induce MiR-34a, Which Targets LDHA to Block Aerobic Glycolysis in Liver Cancer Cells. Pharmaceuticals 2022, 15, 1257. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Ruan, J.; Zhuang, X.; Zhang, X.; Li, Z. Phytochemicals of Garlic: Promising Candidates for Cancer Therapy. Biomedicine & Pharmacotherapy 2020, 123, 109730. [Google Scholar] [CrossRef]
- Asemani, Y.; Zamani, N.; Bayat, M.; Amirghofran, Z. Allium Vegetables for Possible Future of Cancer Treatment. Phytotherapy Research 2019, 33, 3019–3039. [Google Scholar] [CrossRef]
- Li, H.; Jeong, J.H.; Kwon, S.W.; Lee, S.K.; Lee, H.J.; Ryu, J.-H. Z-Ajoene Inhibits Growth of Colon Cancer by Promotion of CK1α Dependent β-Catenin Phosphorylation. Molecules 2020, 25, 703. [Google Scholar] [CrossRef]
- Kaschula, C.H.; Tuveri, R.; Ngarande, E.; Dzobo, K.; Barnett, C.; Kusza, D.A.; Graham, L.M.; Katz, A.A.; Rafudeen, M.S.; Parker, M.I.; et al. The Garlic Compound Ajoene Covalently Binds Vimentin, Disrupts the Vimentin Network and Exerts Anti-Metastatic Activity in Cancer Cells. BMC Cancer 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, X.; Luo, W.; Zhu, J.; Zhao, J.; Wang, M.; Sang, L.; Chang, B.; Wang, B. Allicin in Digestive System Cancer: From Biological Effects to Clinical Treatment. Front. Pharmacol. 2022, 13, 903259. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Su, Q. Molecular Mechanisms for the Anti-Cancer Effects of Diallyl Disulfide. Food and Chemical Toxicology 2013, 57, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Consellería do Medio Rural Diario Oficial de Galicia (DOGA). 2016. Available online: Https://Www.Xunta.Gal/Dog/Publicados/2016/20160627/AnuncioG0426-140616-0003_es.Pdf (accessed on 31 July 2023).
- Devaux, A.; Kromann, P.; Ortiz, O. Potatoes for Sustainable Global Food Security. Potato Res. 2014, 57, 185–199. [Google Scholar] [CrossRef]
- Hellmann, H.; Goyer, A.; Navarre, D.A. Antioxidants in Potatoes: A Functional View on One of the Major Food Crops Worldwide. Molecules 2021, 26, 2446. [Google Scholar] [CrossRef]
- Wolters, A.-M.A.; Uitdewilligen, J.G.A.M.L.; Kloosterman, B.A.; Hutten, R.C.B.; Visser, R.G.F.; Van Eck, H.J. Identification of Alleles of Carotenoid Pathway Genes Important for Zeaxanthin Accumulation in Potato Tubers. Plant Mol Biol 2010, 73, 659–671. [Google Scholar] [CrossRef]
- Brown, C.R.; Kim, T.S.; Ganga, Z.; Haynes, K.; De Jong, D.; Jahn, M.; Paran, I.; De Jong, W. Segregation of Total Carotenoid in High Level Potato Germplasm and Its Relationship to Beta-Carotene Hydroxylase Polymorphism. Am. J. Pot Res 2006, 83, 365–372. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Ministerio de agricultura, Pesca y Alimentación. Pliego de Condiciones IGP Pementos de Galicia. Available online: Https://www.Mapa.Gob.Es/Es/Alimentacion/Temas/Calidad-Diferenciada/Dop-Igp/ (accessed on 22 June 2023).
- Mandal, S.K.; Rath, S.K.; Logesh, R.; Mishra, S.K.; Devkota, H.P.; Das, N. Capsicum annuum L. and Its Bioactive Constituents: A Critical Review of a Traditional Culinary Spice in Terms of Its Modern Pharmacological Potentials with Toxicological Issues. Phytotherapy Research 2023, 37, 965–1002. [Google Scholar] [CrossRef]
- Chapa-Oliver, A.; Mejía-Teniente, L. Capsaicin: From Plants to a Cancer-Suppressing Agent. Molecules 2016, 21, 931. [Google Scholar] [CrossRef]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and Clinical Uses of Capsaicin. European Journal of Pharmacology 2013, 720, 55–62. [Google Scholar] [CrossRef]
- Chen, M.; Xiao, C.; Jiang, W.; Yang, W.; Qin, Q.; Tan, Q.; Lian, B.; Liang, Z.; Wei, C. Capsaicin Inhibits Proliferation and Induces Apoptosis in Breast Cancer by Down-Regulating FBI-1-Mediated NF-ΚB Pathway. DDDT 2021, Volume 15, 125–140. [Google Scholar] [CrossRef]
- Que, T.; Ren, B.; Fan, Y.; Liu, T.; Hou, T.; Dan, W.; Liu, B.; Wei, Y.; Lei, Y.; Zeng, J.; et al. Capsaicin Inhibits the Migration, Invasion and EMT of Renal Cancer Cells by Inducing AMPK/MTOR-Mediated Autophagy. Chemico-Biological Interactions 2022, 366, 110043. [Google Scholar] [CrossRef]
- Wang, F.; Xue, Y.; Fu, L.; Wang, Y.; He, M.; Zhao, L.; Liao, X. Extraction, Purification, Bioactivity and Pharmacological Effects of Capsaicin: A Review. Critical Reviews in Food Science and Nutrition 2022, 62, 5322–5348. [Google Scholar] [CrossRef]
- Díaz-Laviada, I. Effect of Capsaicin on Prostate Cancer Cells. Future Oncology 2010, 6, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Popescu, G.D.A.; Scheau, C.; Badarau, I.A.; Dumitrache, M.-D.; Caruntu, A.; Scheau, A.-E.; Costache, D.O.; Costache, R.S.; Constantin, C.; Neagu, M.; et al. The Effects of Capsaicin on Gastrointestinal Cancers. Molecules 2020, 26, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, D.; Huang, J.; Hu, Y.; Xu, Y. Application of Capsaicin as a Potential New Therapeutic Drug in Human Cancers. J Clin Pharm Ther 2020, 45, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Lee, S.-H. Anticancer Properties of Capsaicin Against Human Cancer. Anticancer Res 2016, 36, 837–843. [Google Scholar]
- Xunta de Galicia Encuesta Sobre Los Hábitos Alimentarios de La Población Adulta Gallega. 2007. Available online: Https://www.Sergas.Es/Cas/Publicaciones/Docs/SaludPublica/PDF-2153-Es.Pdf (accessed on 12 July 2023).
- Consejo Superior de Investigaciones Científicas (CSIC) Identificadas Veinte Variedades de Olivo Autoctonas de Galicia Únicas En El Mundo. 2022. Available online: Https://Novedades-Ptis.Csic.Es/Digitalizacion-y-Sociedad-Futura/Identificadas-Veinte-Variedades-de-Olivo-Autoctonas-de-Galicia-Unicas-En-El-Mundo/ (accessed on 31 July 2023).
- Borzì, A.; Biondi, A.; Basile, F.; Luca, S.; Vicari, E.; Vacante, M. Olive Oil Effects on Colorectal Cancer. Nutrients 2018, 11, 32. [Google Scholar] [CrossRef]
- De Pablos, R.M.; Espinosa-Oliva, A.M.; Hornedo-Ortega, R.; Cano, M.; Arguelles, S. Hydroxytyrosol Protects from Aging Process via AMPK and Autophagy; a Review of Its Effects on Cancer, Metabolic Syndrome, Osteoporosis, Immune-Mediated and Neurodegenerative Diseases. Pharmacological Research 2019, 143, 58–72. [Google Scholar] [CrossRef]
- De Las Hazas, M.C.L.; Rubio, L.; Macia, A.; Motilva, M.J. Hydroxytyrosol: Emerging Trends in Potential Therapeutic Applications. CPD 2018, 24, 2157–2179. [Google Scholar] [CrossRef] [PubMed]
- Vilaplana-Pérez, C.; García-Flores, L.A.; Gil-Izquierdo, A. Hydroxytyrosol and Potential Uses in Cardiovascular Diseases, Cancer, and AIDS. Front. Nutr. 2014, 1. [Google Scholar] [CrossRef]
- Fabiani, R.; Fuccelli, R.; Pieravanti, F.; De Bartolomeo, A.; Morozzi, G. Production of Hydrogen Peroxide Is Responsible for the Induction of Apoptosis by Hydroxytyrosol on HL60 Cells. Mol. Nutr. Food Res. 2009, 53, 887–896. [Google Scholar] [CrossRef]
- Parkinson, L.; Keast, R. Oleocanthal, a Phenolic Derived from Virgin Olive Oil: A Review of the Beneficial Effects on Inflammatory Disease. IJMS 2014, 15, 12323–12334. [Google Scholar] [CrossRef]
- Cárdeno, A.; Sánchez-Hidalgo, M.; Rosillo, M.A.; De La Lastra, C.A. Oleuropein, a Secoiridoid Derived from Olive Tree, Inhibits the Proliferation of Human Colorectal Cancer Cell Through Downregulation of HIF-1α. Nutrition and Cancer 2013, 65, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Boss, A.; Bishop, K.; Marlow, G.; Barnett, M.; Ferguson, L. Evidence to Support the Anti-Cancer Effect of Olive Leaf Extract and Future Directions. Nutrients 2016, 8, 513. [Google Scholar] [CrossRef]
- De Stefanis; Scimè; Accomazzo; Catti; Occhipinti; Bertea; Costelli Anti-Proliferative Effects of an Extra-Virgin Olive Oil Extract Enriched in Ligstroside Aglycone and Oleocanthal on Human Liver Cancer Cell Lines. Cancers 2019, 11, 1640. [CrossRef]
- Emma, M.R.; Augello, G.; Di Stefano, V.; Azzolina, A.; Giannitrapani, L.; Montalto, G.; Cervello, M.; Cusimano, A. Potential Uses of Olive Oil Secoiridoids for the Prevention and Treatment of Cancer: A Narrative Review of Preclinical Studies. IJMS 2021, 22, 1234. [Google Scholar] [CrossRef]
- Newmark, H.L. Squalene, Olive Oil, and Cancer Risk: Review and Hypothesis. Annals NY Acad Sci 1999, 889, 193–203. [Google Scholar] [CrossRef]
- Smith, T.J. Squalene: Potential Chemopreventive Agent. Expert Opinion on Investigational Drugs 2000, 9, 1841–1848. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Marino Gammazza, A.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential Health Benefits of Olive Oil and Plant Polyphenols. IJMS 2018, 19, 686. [Google Scholar] [CrossRef]
- Jacob, C. Inspired by Nature: Antioxidants and Nanotechnology. Antioxidants 2018, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Axencia Galega de Calidade Alimentaria. Consellería del Medio Rural Pliego de Condiciones IGP Castaña de Galicia. 2019. Available online: Https://Mediorural.Xunta.Gal/Sites/Default/Files/Produtos/En-Tramitacion/IGP_Castana_de_Galicia_Pliego_de_condiciones_septiembre_2019_CCC.Pdf (accessed on 22 July 2023).
- Hu, M.; Yang, X.; Chang, X. Bioactive Phenolic Components and Potential Health Effects of Chestnut Shell: A Review. J. Food Biochem. 2021, 45. [Google Scholar] [CrossRef] [PubMed]
- Sorice, A.; Siano, F.; Capone, F.; Guerriero, E.; Picariello, G.; Budillon, A.; Ciliberto, G.; Paolucci, M.; Costantini, S.; Volpe, M. Potential Anticancer Effects of Polyphenols from Chestnut Shell Extracts: Modulation of Cell Growth, and Cytokinomic and Metabolomic Profiles. Molecules 2016, 21, 1411. [Google Scholar] [CrossRef] [PubMed]
- Fazliev, S.; Tursunov, K.; Razzokov, J.; Sharipov, A. Escin’s Multifaceted Therapeutic Profile in Treatment and Post-Treatment of Various Cancers: A Comprehensive Review. Biomolecules 2023, 13, 315. [Google Scholar] [CrossRef]
- Liu, Z.-R.; Sun, L.-Z.; Jia, T.-H.; Jia, D.-F. β-Aescin Shows Potent Antiproliferative Activity in Osteosarcoma Cells by Inducing Autophagy, ROS Generation and Mitochondrial Membrane Potential Loss. J BUON 2017, 22, 1582–1586. [Google Scholar]
- Cheong, D.H.J.; Arfuso, F.; Sethi, G.; Wang, L.; Hui, K.M.; Kumar, A.P.; Tran, T. Molecular Targets and Anti-Cancer Potential of Escin. Cancer Letters 2018, 422, 1–8. [Google Scholar] [CrossRef]
- Jung, B.S.; Lee, N.-K.; Na, D.S.; Yu, H.H.; Paik, H.-D. Comparative Analysis of the Antioxidant and Anticancer Activities of Chestnut Inner Shell Extracts Prepared with Various Solvents: Chestnut Inner Shell Extracts for Functional Foods. J. Sci. Food Agric. 2016, 96, 2097–2102. [Google Scholar] [CrossRef]
- Park, W.Y.; Song, G.; Park, J.Y.; Ahn, K.S.; Kwak, H.J.; Park, J.; Lee, J.H.; Um, J.-Y. Ellagic Acid Improves Benign Prostate Hyperplasia by Regulating Androgen Signaling and STAT3. Cell Death Dis 2022, 13, 554. [Google Scholar] [CrossRef]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing Gallic Acid for Its Broad Spectrum Applications. MRMC 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxidative Medicine and Cellular Longevity 2018, 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Axencia galega da calidade alimentaria. Consellería del Medio Rural Pliego de Condiciones y Características IGP Miel de Galicia. 2020. Available online: Https://Mediorural.Xunta.Gal/Sites/Default/Files/Produtos/Entramitacion/Pliego_de_condiciones_Miel_de_Galicia_julio_2020_final.Pdf (accessed on 23 July 2023).
- Chan-Zapata, I.; Segura-Campos, M.R. Honey and Its Protein Components: Effects in the Cancer Immunology. J Food Biochem 2021, 45. [Google Scholar] [CrossRef] [PubMed]
- Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; Md, S. Apamin-Conjugated Alendronate Sodium Nanocomplex for Management of Pancreatic Cancer. Pharmaceuticals 2021, 14, 729. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomedicine & Pharmacotherapy 2019, 112, 108612. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Fasoulakis, Z.; Koutras, A.; Syllaios, A.; Schizas, D.; Garmpis, N.; Diakosavvas, M.; Angelou, K.; Tsatsaris, G.; Pagkalos, A.; Ntounis, T.; et al. Breast Cancer Apoptosis and the Therapeutic Role of Luteolin. chr 2021, 116, 170. [Google Scholar] [CrossRef]
- Prasher, P.; Sharma, M.; Singh, S.K.; Gulati, M.; Chellappan, D.K.; Zacconi, F.; De Rubis, G.; Gupta, G.; Sharifi-Rad, J.; Cho, W.C.; et al. Luteolin: A Flavonoid with a Multifaceted Anticancer Potential. Cancer Cell Int 2022, 22, 386. [Google Scholar] [CrossRef]
- Lolli, G.; Cozza, G.; Mazzorana, M.; Tibaldi, E.; Cesaro, L.; Donella-Deana, A.; Meggio, F.; Venerando, A.; Franchin, C.; Sarno, S.; et al. Inhibition of Protein Kinase CK2 by Flavonoids and Tyrphostins. A Structural Insight. Biochemistry 2012, 51, 6097–6107. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol – A Dietary Anticancer Molecule with Multiple Mechanisms of Action: Recent Trends and Advancements. Journal of Functional Foods 2017, 30, 203–219. [Google Scholar] [CrossRef]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H.; et al. Kaempferol: A Key Emphasis to Its Anticancer Potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The Mechanism of Anticancer Action and Potential Clinical Use of Kaempferol in the Treatment of Breast Cancer. Biomedicine & Pharmacotherapy 2019, 117, 109086. [Google Scholar] [CrossRef]
- Yokoyama, T.; Kosaka, Y.; Mizuguchi, M. Structural Insight into the Interactions between Death-Associated Protein Kinase 1 and Natural Flavonoids. J. Med. Chem. 2015, 58, 7400–7408. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Fu, Y.; Malakhova, M.; Kurinov, I.; Zhu, F.; Yao, K.; Li, H.; Chen, H.; Li, W.; Lim, D.Y.; et al. Caffeic Acid Directly Targets ERK1/2 to Attenuate Solar UV-Induced Skin Carcinogenesis. Cancer Prevention Research 2014, 7, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Museo do Viño Uvas y Vinos de Galicia. Available online: Https://Museovinogalicia.Xunta.Gal/Es/El-Vino/Uvas-y-Vinos-de-Galicia (accessed on 31 July 2023).
- Ren, B.; Kwah, M.X.-Y.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.-L.; Wang, L.; Ong, P.S.; et al. Resveratrol for Cancer Therapy: Challenges and Future Perspectives. Cancer Letters 2021, 515, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an Anti-Cancer Agent: A Review. Critical Reviews in Food Science and Nutrition 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, M.; Chen, Y.; Wang, X.J.; Tang, X. Resveratrol: An Overview of Its Anti-Cancer Mechanisms. Life Sciences 2018, 207, 340–349. [Google Scholar] [CrossRef]
- Mach, N. Efecto de los polifenoles del vino sobre la prevención del cáncer. NUTRICION HOSPITALARIA 2015, 535–551. [Google Scholar] [CrossRef]
- Deus, C.M.; Serafim, T.L.; Magalhães-Novais, S.; Vilaça, A.; Moreira, A.C.; Sardão, V.A.; Cardoso, S.M.; Oliveira, P.J. Sirtuin 1-Dependent Resveratrol Cytotoxicity and pro-Differentiation Activity on Breast Cancer Cells. Arch Toxicol 2017, 91, 1261–1278. [Google Scholar] [CrossRef]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial Action of Resveratrol: How and Why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef]
- Qasem, R.J. The Estrogenic Activity of Resveratrol: A Comprehensive Review of in Vitro and in Vivo Evidence and the Potential for Endocrine Disruption. Critical Reviews in Toxicology 2020, 50, 439–462. [Google Scholar] [CrossRef]
- Figueiredo-González, M.; Cancho-Grande, B.; Boso, S.; Santiago, J.L.; Martínez, M.C.; Simal-Gándara, J. Evolution of Flavonoids in Mouratón Berries Taken from Both Bunch Halves. Food Chemistry 2013, 138, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Mir-Cerdà, A.; Carretero, I.; Coves, J.R.; Pedrouso, A.; Castro-Barros, C.M.; Alvarino, T.; Cortina, J.L.; Saurina, J.; Granados, M.; Sentellas, S. Recovery of Phenolic Compounds from Wine Lees Using Green Processing: Identifying Target Molecules and Assessing Membrane Ultrafiltration Performance. Science of The Total Environment 2023, 857, 159623. [Google Scholar] [CrossRef] [PubMed]
- Mirás-Avalos, J.M.; Bouzas-Cid, Y.; Trigo-Córdoba, E.; Orriols, I.; Falqué, E. Amino Acid Profiles to Differentiate White Wines from Three Autochtonous Galician Varieties. Foods 2020, 9, 114. [Google Scholar] [CrossRef]
- Sochorova, L.; Prusova, B.; Jurikova, T.; Mlcek, J.; Adamkova, A.; Baron, M.; Sochor, J. The Study of Antioxidant Components in Grape Seeds. Molecules 2020, 25, 3736. [Google Scholar] [CrossRef]
- Ravindranathan, P.; Pasham, D.; Balaji, U.; Cardenas, J.; Gu, J.; Toden, S.; Goel, A. Mechanistic Insights into Anticancer Properties of Oligomeric Proanthocyanidins from Grape Seeds in Colorectal Cancer. Carcinogenesis 2018, 39, 767–777. [Google Scholar] [CrossRef]
- Garavaglia, J.; Markoski, M.M.; Oliveira, A.; Marcadenti, A. Grape Seed Oil Compounds: Biological and Chemical Actions for Health. Nutr Metab Insights 2016, 9, NMI.S32910. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Moreira, A.C.; Nazmi, K.; Moniz, T.; Vale, N.; Rangel, M.; Gomes, P.; Bolscher, J.G.M.; Rodrigues, P.N.; Bastos, M.; et al. Lactoferricin Peptides Increase Macrophages’ Capacity To Kill Mycobacterium Avium. mSphere 2017, 2, e00301–17. [Google Scholar] [CrossRef]
- Mori, S.; Fujiwara-Tani, R.; Kishi, S.; Sasaki, T.; Ohmori, H.; Goto, K.; Nakashima, C.; Nishiguchi, Y.; Kawahara, I.; Luo, Y.; et al. Enhancement of Anti-Tumoral Immunity by β-Casomorphin-7 Inhibits Cancer Development and Metastasis of Colorectal Cancer. IJMS 2021, 22, 8232. [Google Scholar] [CrossRef]
- Kallergi, G. Distinct Signaling Pathways Regulate Differential Opioid Effects on Actin Cytoskeleton in Malignant MCF7 and Nonmalignant MCF12A Human Breast Epithelial Cells. Experimental Cell Research 2003, 288, 94–109. [Google Scholar] [CrossRef]
- Mahanta, S.; Paul, S. Stable Self-Assembly of Bovine α-Lactalbumin Exhibits Target-Specific Antiproliferative Activity in Multiple Cancer Cells. ACS Appl. Mater. Interfaces 2015, 7, 28177–28187. [Google Scholar] [CrossRef]
- Teixeira, F.J.; Santos, H.O.; Howell, S.L.; Pimentel, G.D. Whey Protein in Cancer Therapy: A Narrative Review. Pharmacological Research 2019, 144, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Pastor, R.; Bouzas, C.; Tur, J.A. Beneficial Effects of Dietary Supplementation with Olive Oil, Oleic Acid, or Hydroxytyrosol in Metabolic Syndrome: Systematic Review and Meta-Analysis. Free Radical Biology and Medicine 2021, 172, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N Engl J Med 2018, 378, e34. [Google Scholar] [CrossRef]
- Covas, M.-I.; Nyyssönen, K.; Poulsen, H.E.; Kaikkonen, J.; Zunft, H.-J.F.; Kiesewetter, H.; Gaddi, A.; De La Torre, R.; Mursu, J.; Bäumler, H.; et al. The Effect of Polyphenols in Olive Oil on Heart Disease Risk Factors: A Randomized Trial. Ann Intern Med 2006, 145, 333. [Google Scholar] [CrossRef]
- Sanati, S.; Razavi, B.M. hosseinzadeh, hossein A Review of the Effects of Capsicum Annuum L. and Its Constituent, Capsaicin, in Metabolic Syndrome. Iranian Journal of Basic Medical Sciences 2018, 21. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Biological Properties, Bioactive Constituents, and Pharmacokinetics of Some Capsicum Spp. and Capsaicinoids. IJMS 2020, 21, 5179. [Google Scholar] [CrossRef] [PubMed]
- Varela Mosquera, G.; O. Moreiras Tuní; ansón Oliart R., Ávila Torres JM, Cuadrado Vives C., Estalrich Rodríguez Patricia, Del Pozo de la Calle S.; Rodríguez Sangrador M. Consumo de Alimentos En Galicia: La Dieta Atlántica 2004.
- Baladia, E.; Moñino, M.; Martínez-Rodríguez, R.; Miserachs, M.; Russolillo, G.; Picazo, Ó.; Fernández, T.; Morte, V. Uso de Suplementos Nutricionales y Productos a Base de Extractos de Plantas En Población Española: Un Estudio Transversal. Rev Esp Nutr Hum Diet 2022, 26, 217–229. [Google Scholar] [CrossRef]
- Coppens, P.; Delmulle, L.; Gulati, O.; Richardson, D.; Ruthsatz, M.; Sievers, H.; Sidani, S. Use of Botanicals in Food Supplements. Ann Nutr Metab 2006, 50, 538–554. [Google Scholar] [CrossRef]
- Skeie, G.; Braaten, T.; Hjartåker, A.; Lentjes, M.; Amiano, P.; Jakszyn, P.; Pala, V.; Palanca, A.; Niekerk, E.M.; Verhagen, H.; et al. Use of Dietary Supplements in the European Prospective Investigation into Cancer and Nutrition Calibration Study. Eur J Clin Nutr 2009, 63, S226–S238. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, Z.; Feng, Z.; Hao, J.; Shen, W.; Li, X.; Sun, L.; Sharman, E.; Wang, Y.; Wertz, K.; et al. Hydroxytyrosol Protects against Oxidative Damage by Simultaneous Activation of Mitochondrial Biogenesis and Phase II Detoxifying Enzyme Systems in Retinal Pigment Epithelial Cells. The Journal of Nutritional Biochemistry 2010, 21, 1089–1098. [Google Scholar] [CrossRef]
- Wu, Y.; Li, R.; Huang, H.; Fletcher, A.; Yu, L.; Pham, Q.; Yu, L.; He, Q.; Wang, T. Inhibition of Tumor Growth by Dietary Indole-3-Carbinol in a Prostate Cancer Xenograft Model May Be Associated with Disrupted Gut Microbial Interactions. Nutrients 2019, 11, 467. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.; Nisani, S.; Chamovitz, D.A. Indole-3-Carbinol: A Plant Hormone Combatting Cancer. F1000Res 2018, 7, 689. [Google Scholar] [CrossRef] [PubMed]
- A Patil, S.; Addo, J.K.; Deokar, H.; Sun, S.; Wang, J.; Li, W.; Suttle, D.P.; Wang, W.; Zhang, R.; Buolamwini, J.K. Synthesis, Biological Evaluation and Modeling Studies of New Pyrido[3,4-b] Indole Derivatives as Broad-Spectrum Potent Anticancer Agents. Drug Des 2017, 06. [Google Scholar] [CrossRef]
- Mateos, R.; Pérez-Correa, J.R.; Domínguez, H. Bioactive Properties of Marine Phenolics. Marine Drugs 2020, 18, 501. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
Active compounds found in brown algae native to the SEAD.
Figure 4.
Main flow of information and search strategy used in this study.
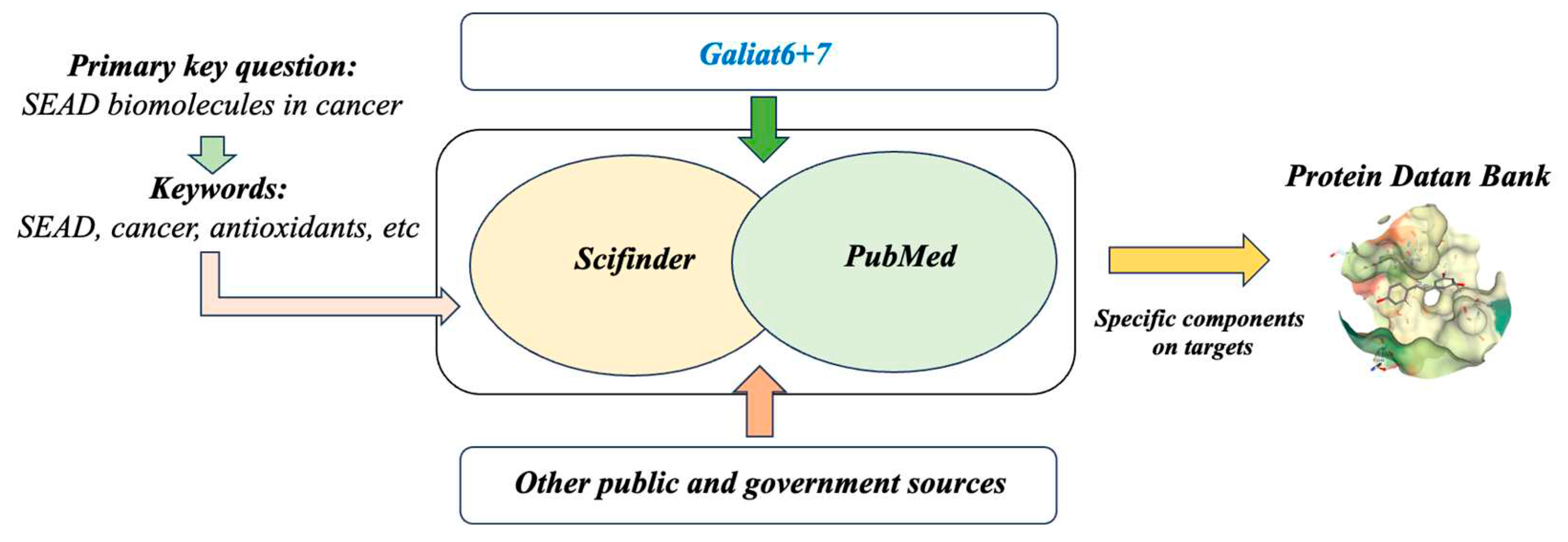
Figure 5.
Fresh fish, shellfish, crustaceans and mollusks consumption in Spain (2022). Self-made. Source: Ministry of Agriculture, Fisheries and Food. Government of Spain [14].
Figure 5.
Fresh fish, shellfish, crustaceans and mollusks consumption in Spain (2022). Self-made. Source: Ministry of Agriculture, Fisheries and Food. Government of Spain [14].
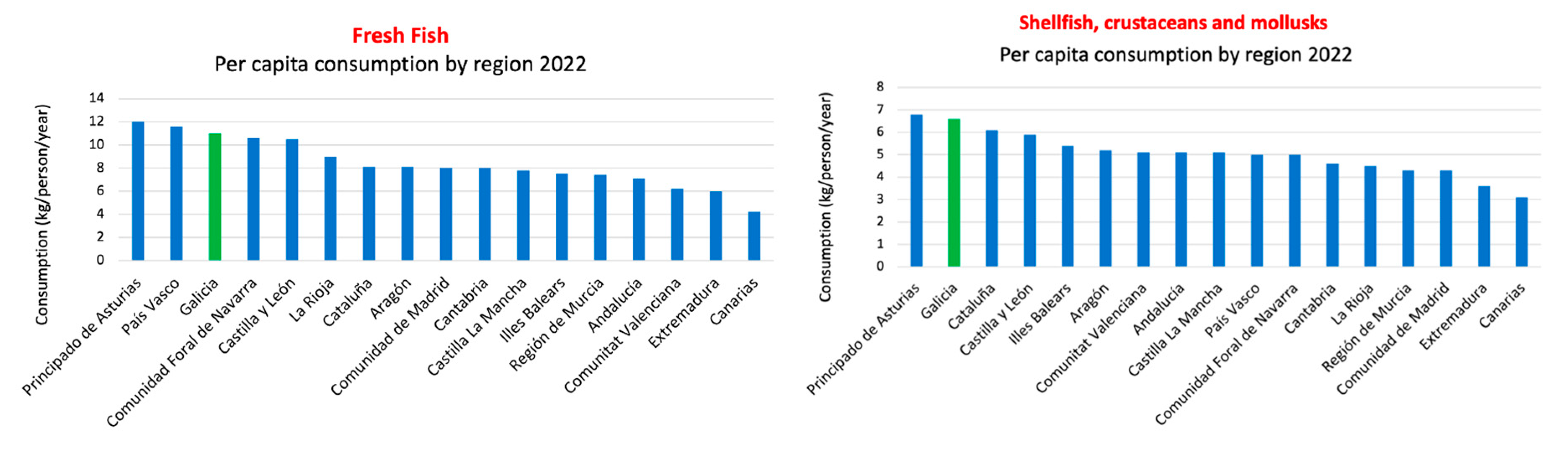
Figure 6.
Main betaines in the diet and other biomolecules present in mussels and octopus.
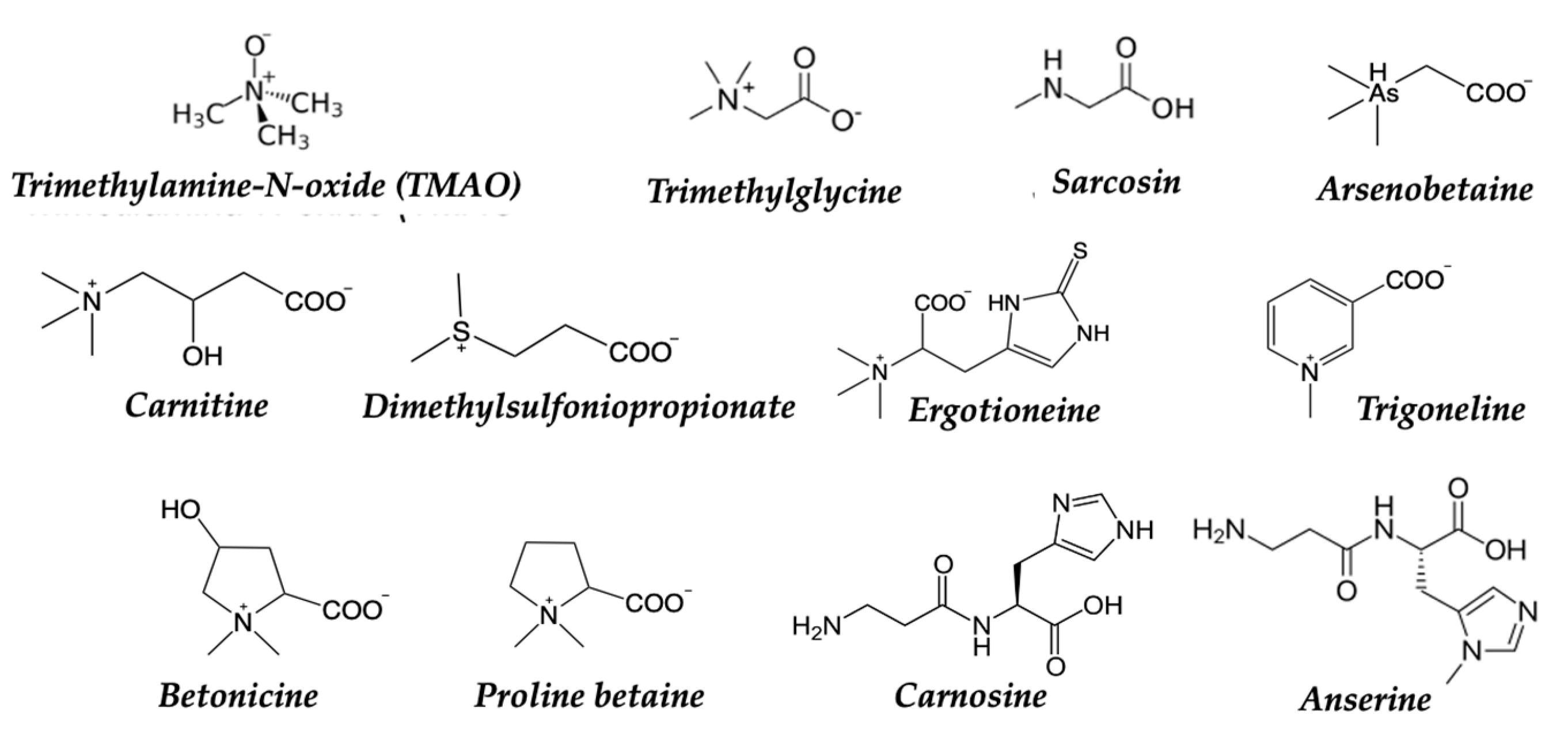
Scheme 1.
Main metabolic reactions of organic and inorganic Se compounds in humans. Adapted and modified from [71]. Briefly, H2Se is formed during reduction both inorganic (selenate) and organic (SeMet), species. Both SeMet and CH3SeCys are enzymatically converted to CH3SeH. Glutathione (GSH) or other thiols undergo these reactions when reduction of inorganic Se species occurs. Reduction-Oxidation Species are generated by two presented reactions. SeMet—L-selenomethionine, SeCys—selenocysteine, CH3SeH—monomethylselenol, H2Se—hydrogen selenide, HSePO3—selenophosphate, SeO2—Se dioxide.
Scheme 1.
Main metabolic reactions of organic and inorganic Se compounds in humans. Adapted and modified from [71]. Briefly, H2Se is formed during reduction both inorganic (selenate) and organic (SeMet), species. Both SeMet and CH3SeCys are enzymatically converted to CH3SeH. Glutathione (GSH) or other thiols undergo these reactions when reduction of inorganic Se species occurs. Reduction-Oxidation Species are generated by two presented reactions. SeMet—L-selenomethionine, SeCys—selenocysteine, CH3SeH—monomethylselenol, H2Se—hydrogen selenide, HSePO3—selenophosphate, SeO2—Se dioxide.
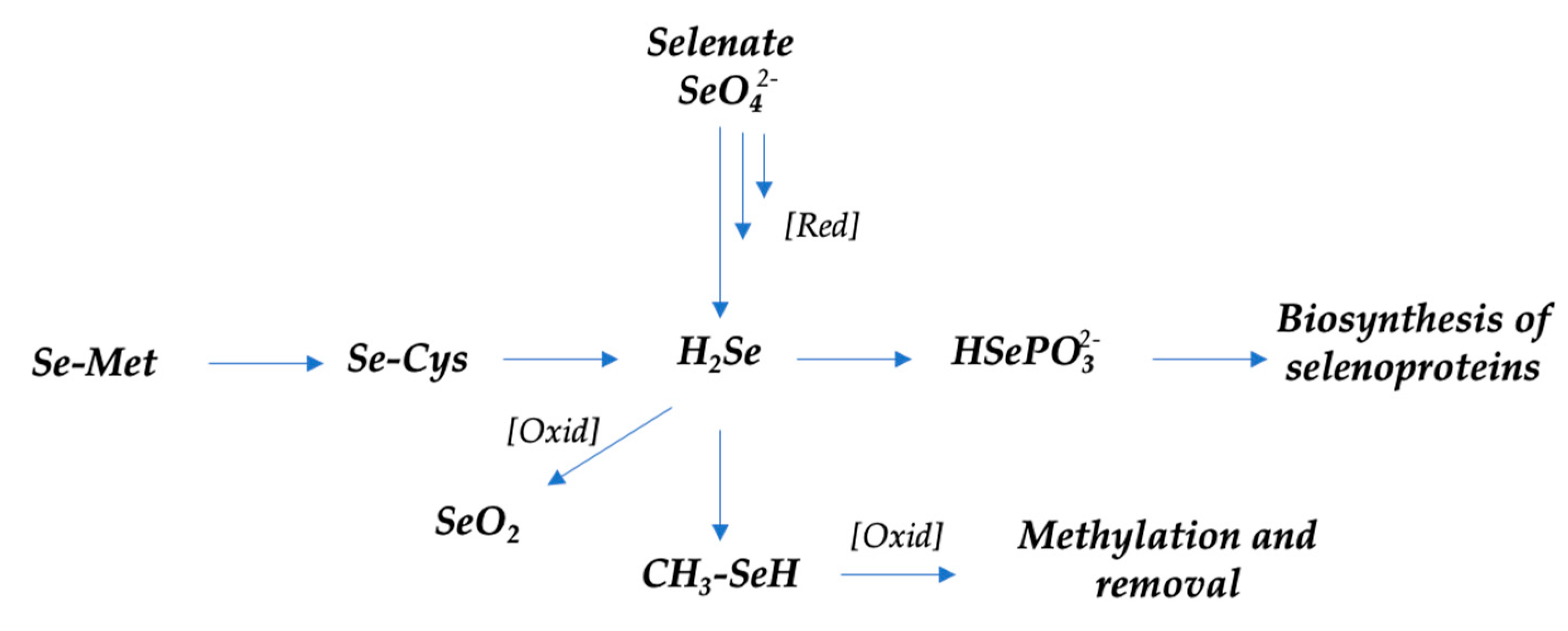
Figure 7.
Chemical structure of OCTPEP-1 peptide.
Figure 8.
In silico analysis was performed using the AutoDock program to explore the most promising interactions between N-(2-oxoazepan-3-yl)-pyrrolidine-2-carboxamide (OPC) and selected cancer targets in 22Rv1 cells. The molecule of ozopromide was docked into the active sites of: a) Cyclin D1, b) ERK1, c) BAD d) c-SRC e) ERK f) ACE2. Adapted and modified from [88].
Figure 8.
In silico analysis was performed using the AutoDock program to explore the most promising interactions between N-(2-oxoazepan-3-yl)-pyrrolidine-2-carboxamide (OPC) and selected cancer targets in 22Rv1 cells. The molecule of ozopromide was docked into the active sites of: a) Cyclin D1, b) ERK1, c) BAD d) c-SRC e) ERK f) ACE2. Adapted and modified from [88].
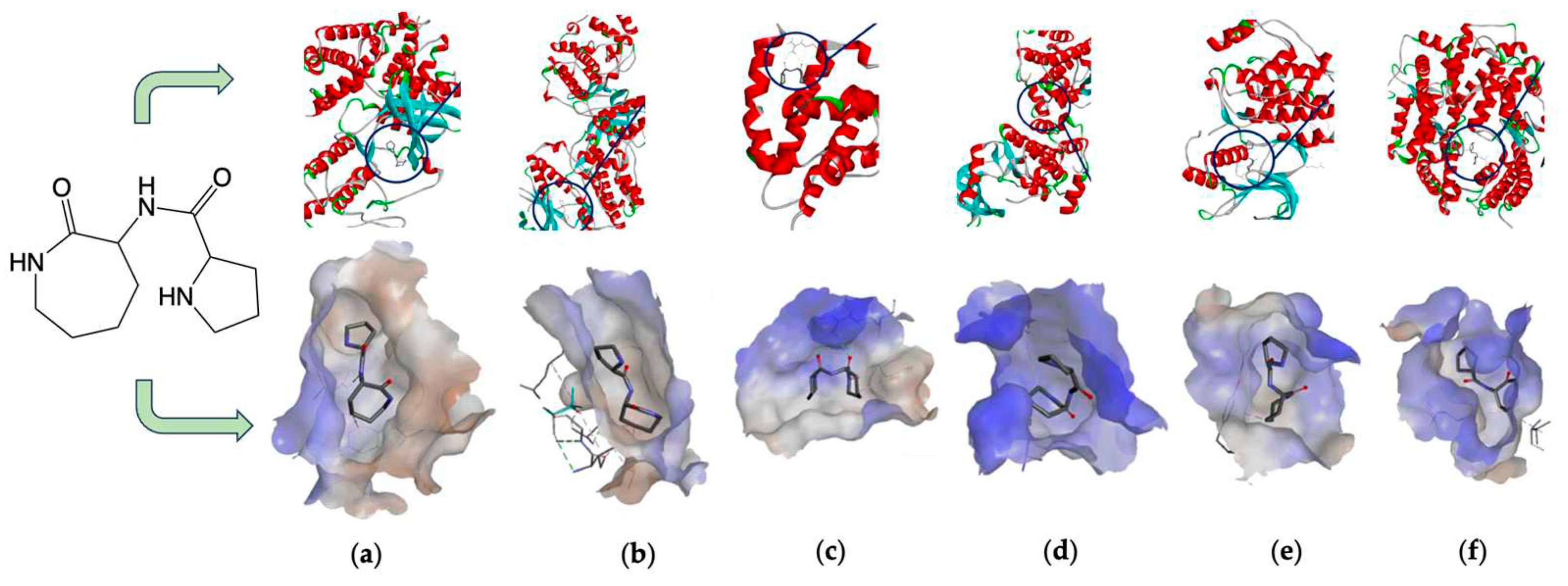
Figure 9.
Distribution of Brassica varieties by provinces in Galicia (up). b) Brassica Oleracea (cabbage) in Spain (down): Adapted and modified from [17].
Figure 9.
Distribution of Brassica varieties by provinces in Galicia (up). b) Brassica Oleracea (cabbage) in Spain (down): Adapted and modified from [17].
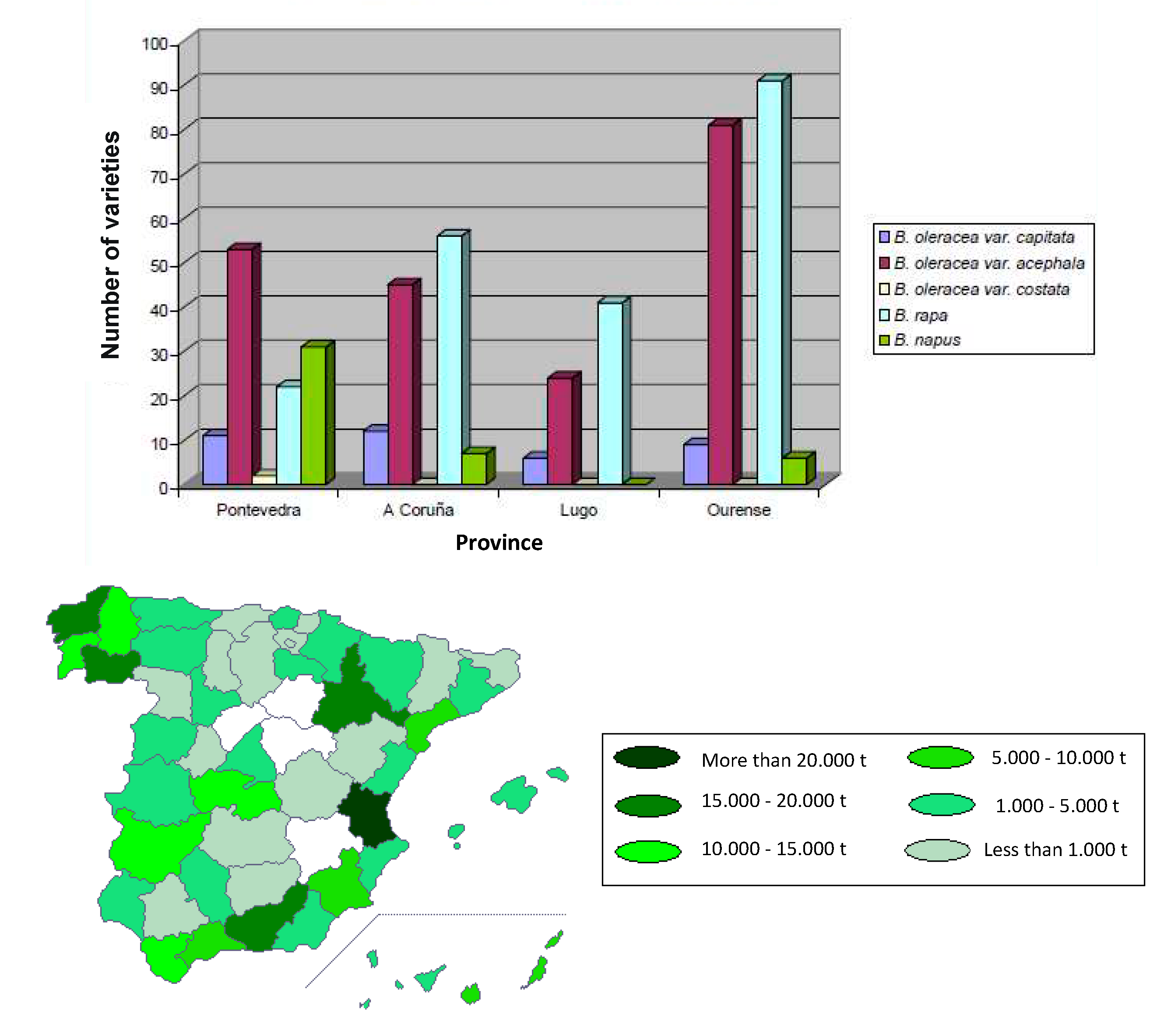
Figure 10.
General structure of glucosinolates and main glucosinolates present in the SEAD.
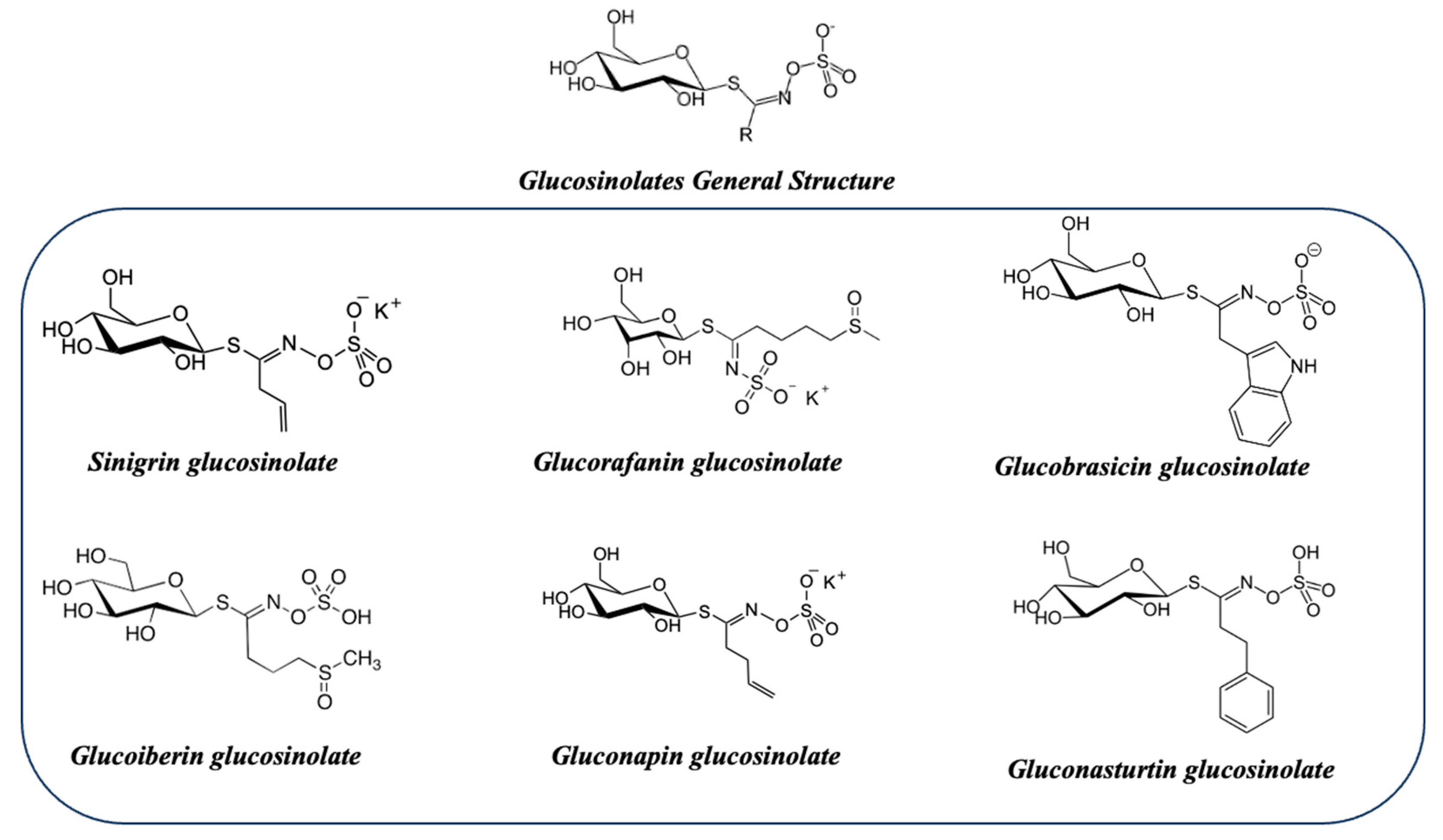
Scheme 2.
Hydrolysis products from glucosinolates.
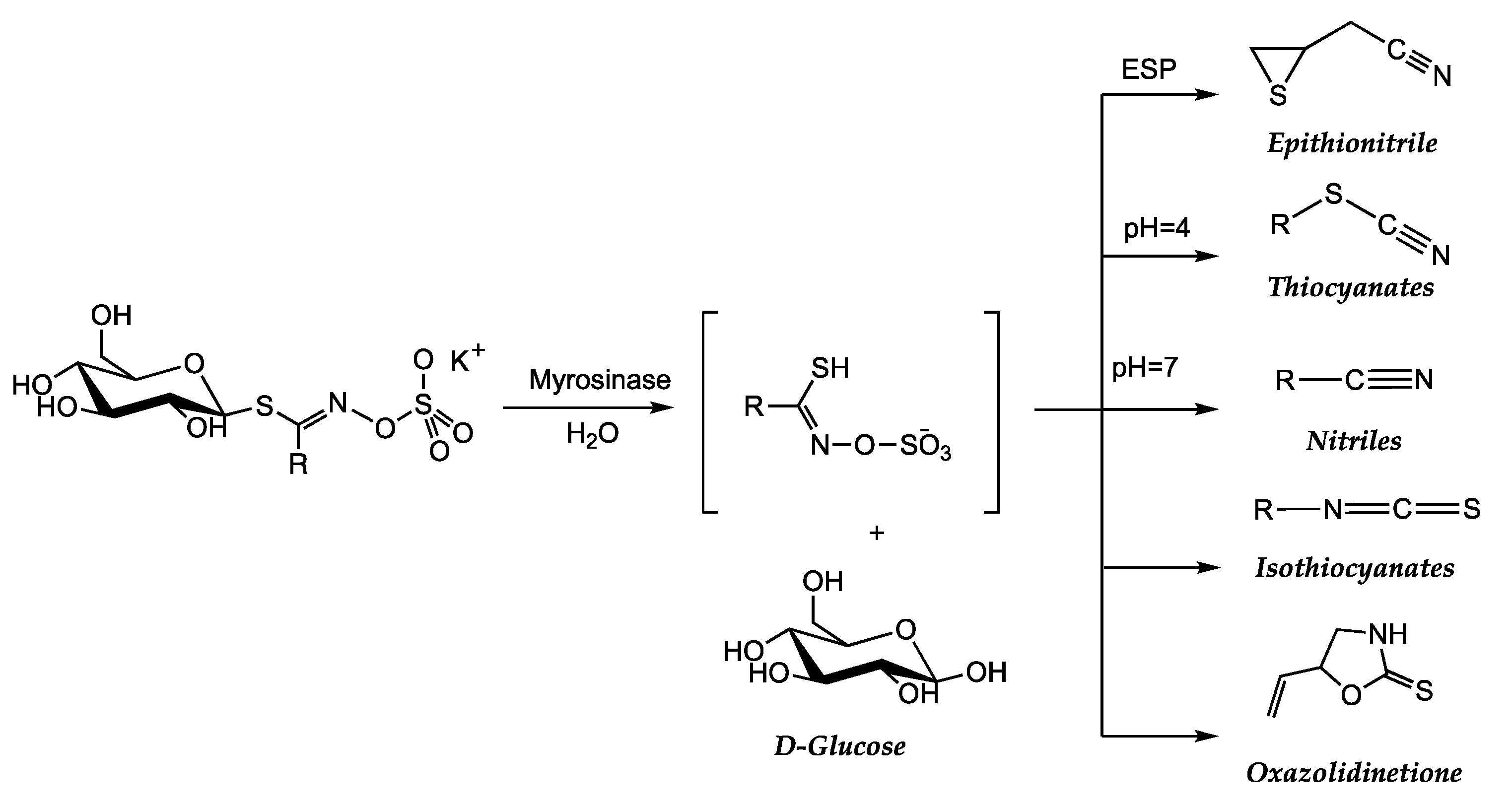
Scheme 3.
Conversion of glucosinolates found in Rocket salad, turnip greens, or cruciferous vegetables into sulforaphane.
Scheme 3.
Conversion of glucosinolates found in Rocket salad, turnip greens, or cruciferous vegetables into sulforaphane.
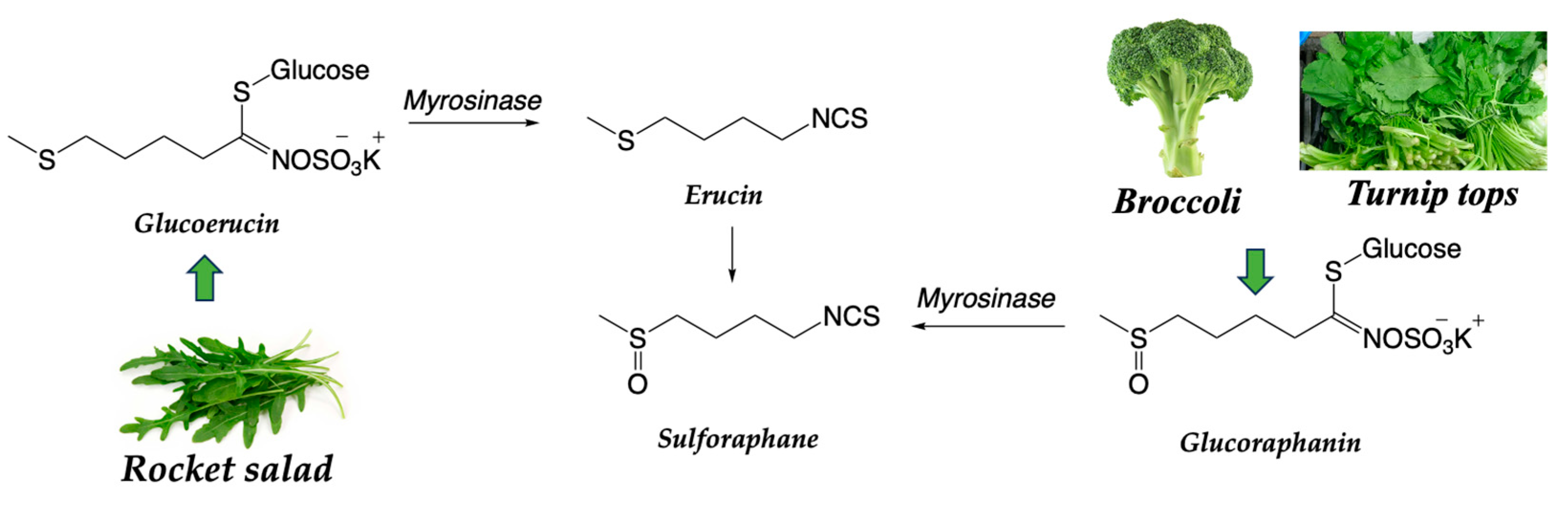
Figure 12.
2MS6 crystal structure: Human Telomeric G-quadruplex DNA sequence (TTAGGGT)4 complexed with Flavonoid Quercetin. PDB: https://doi.org/10.2210/pdb2MS6/pdb created with NGL [113].
Figure 12.
2MS6 crystal structure: Human Telomeric G-quadruplex DNA sequence (TTAGGGT)4 complexed with Flavonoid Quercetin. PDB: https://doi.org/10.2210/pdb2MS6/pdb created with NGL [113].

Scheme 4.
Hydrolysis of glucobrassicin glucosinolate, assisted by the enzyme myrosinase, yields I3C (indole-3-carbinol), followed by the subsequent biosynthesis of DIM (3,3’-diindolylmethane) [122].
Scheme 4.
Hydrolysis of glucobrassicin glucosinolate, assisted by the enzyme myrosinase, yields I3C (indole-3-carbinol), followed by the subsequent biosynthesis of DIM (3,3’-diindolylmethane) [122].
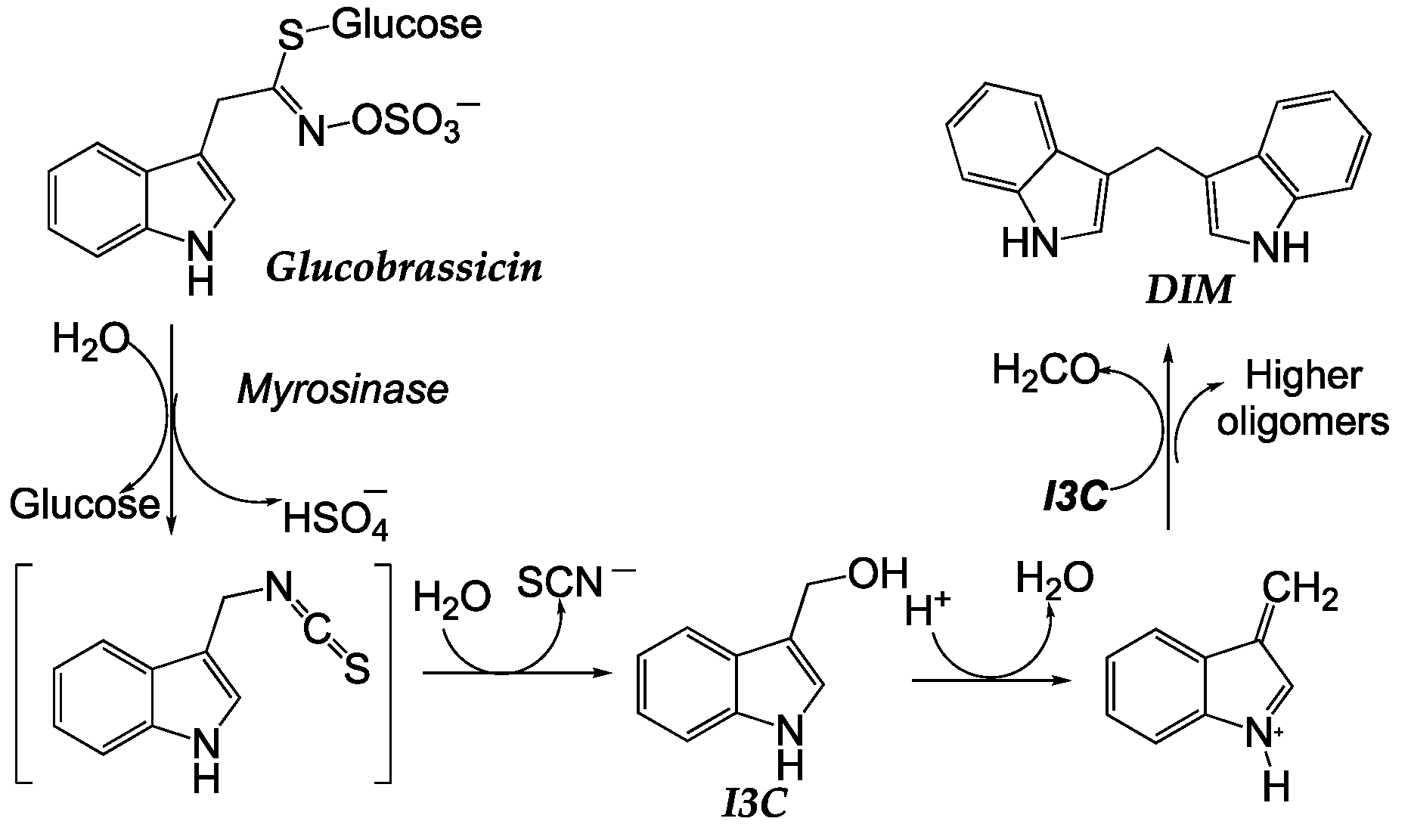
Scheme 5.
Biosynthetic pathway for the conversion of components in garlic from alliin.
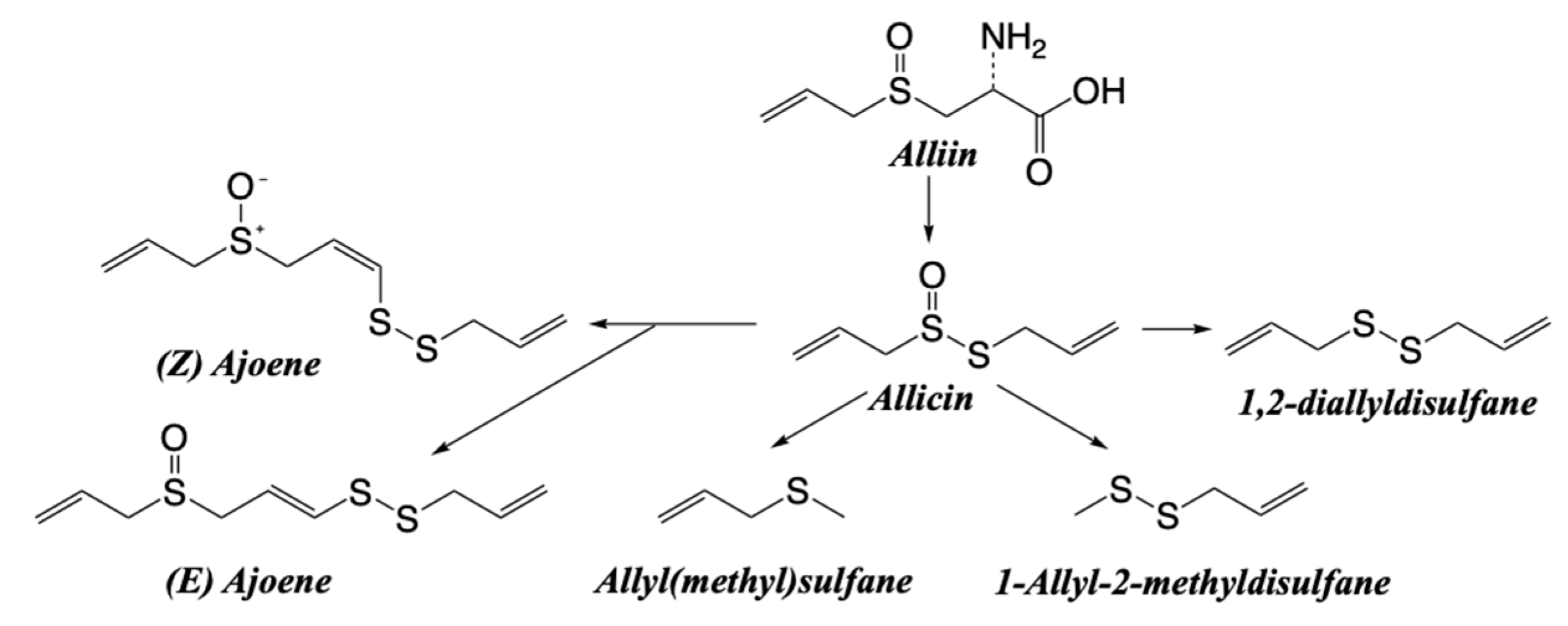
Figure 13.
Own elaboration. Source: Ministry of Agriculture, Fisheries, and Food. Government of Spain. Report on food consumption in Spain 2022 [14].
Figure 13.
Own elaboration. Source: Ministry of Agriculture, Fisheries, and Food. Government of Spain. Report on food consumption in Spain 2022 [14].
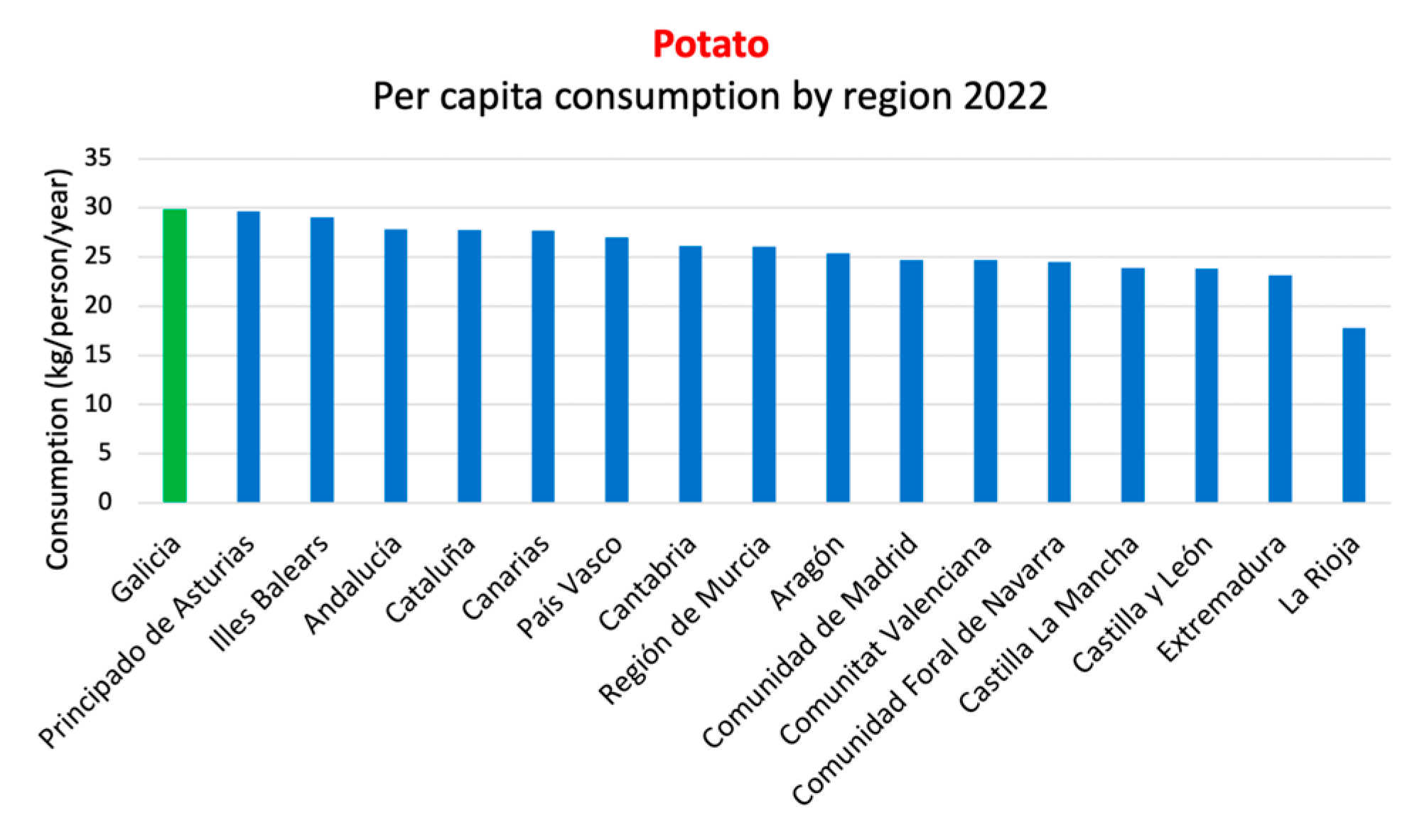
Figure 14.
Chemical structures of Lutein and Zeaxanthin.
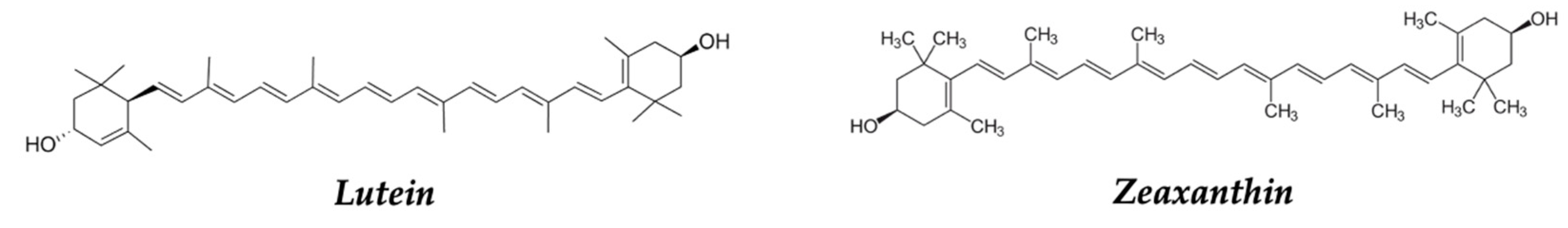
Figure 15.
Chemical structure of capsaicin and other derivatives in Capsicum annuum L.
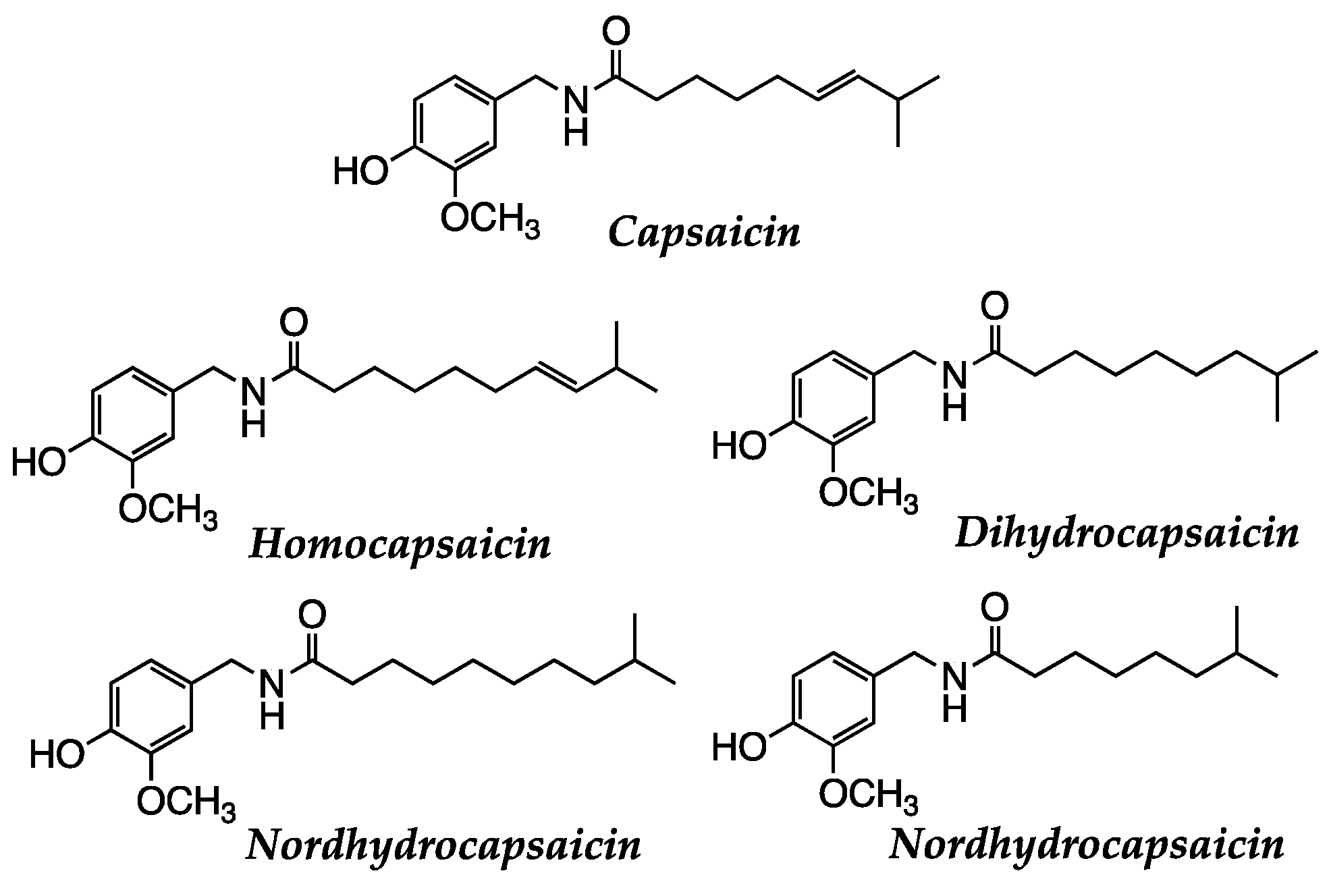
Figure 16.
Some of the main active principles of olive oil.
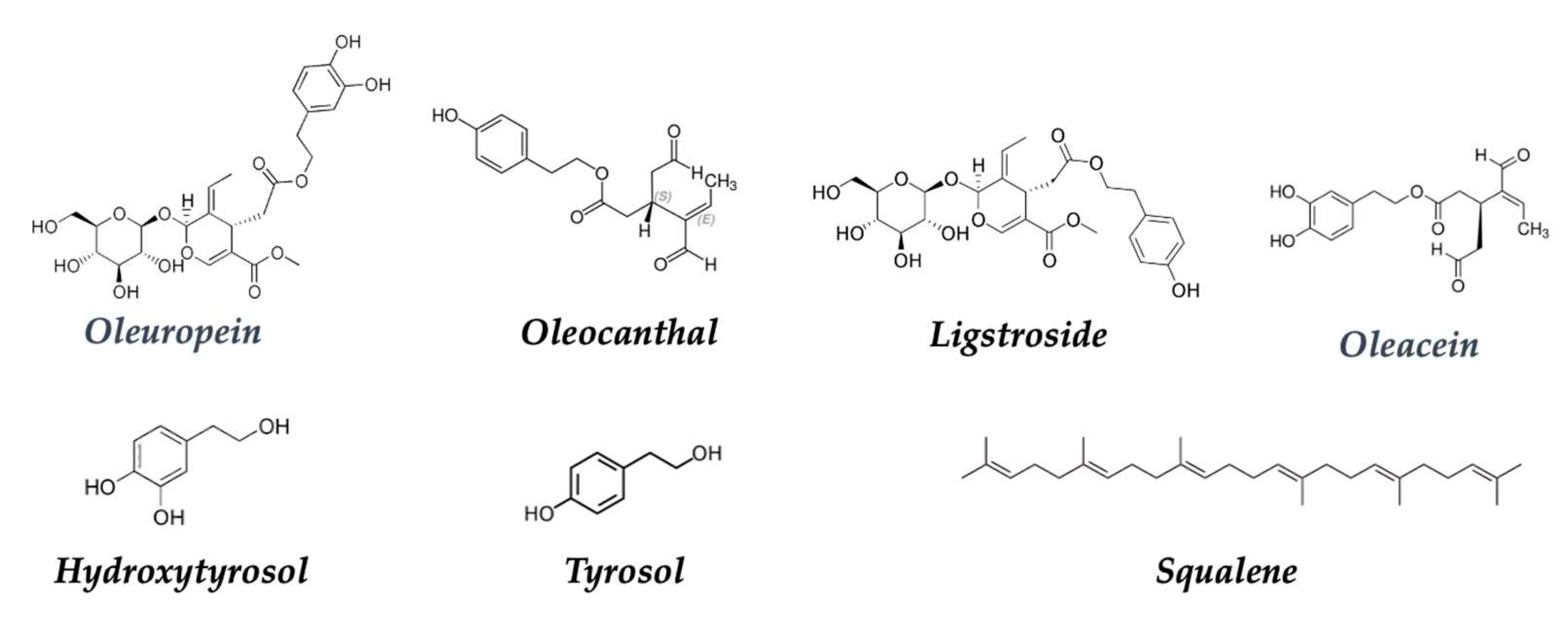
Figure 17.
Main biomolecules with anticancer and antioxidant profile of chestnuts.
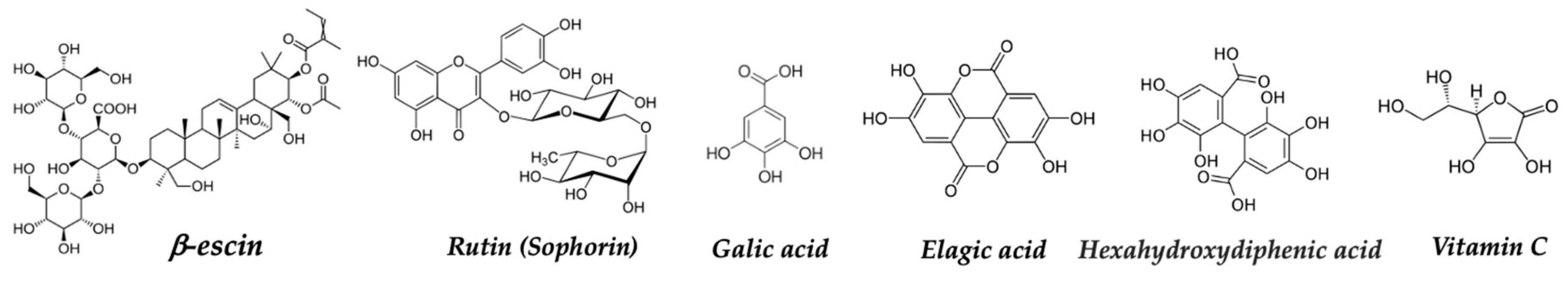
Figure 18.
Most significant biomolecules found in honey.
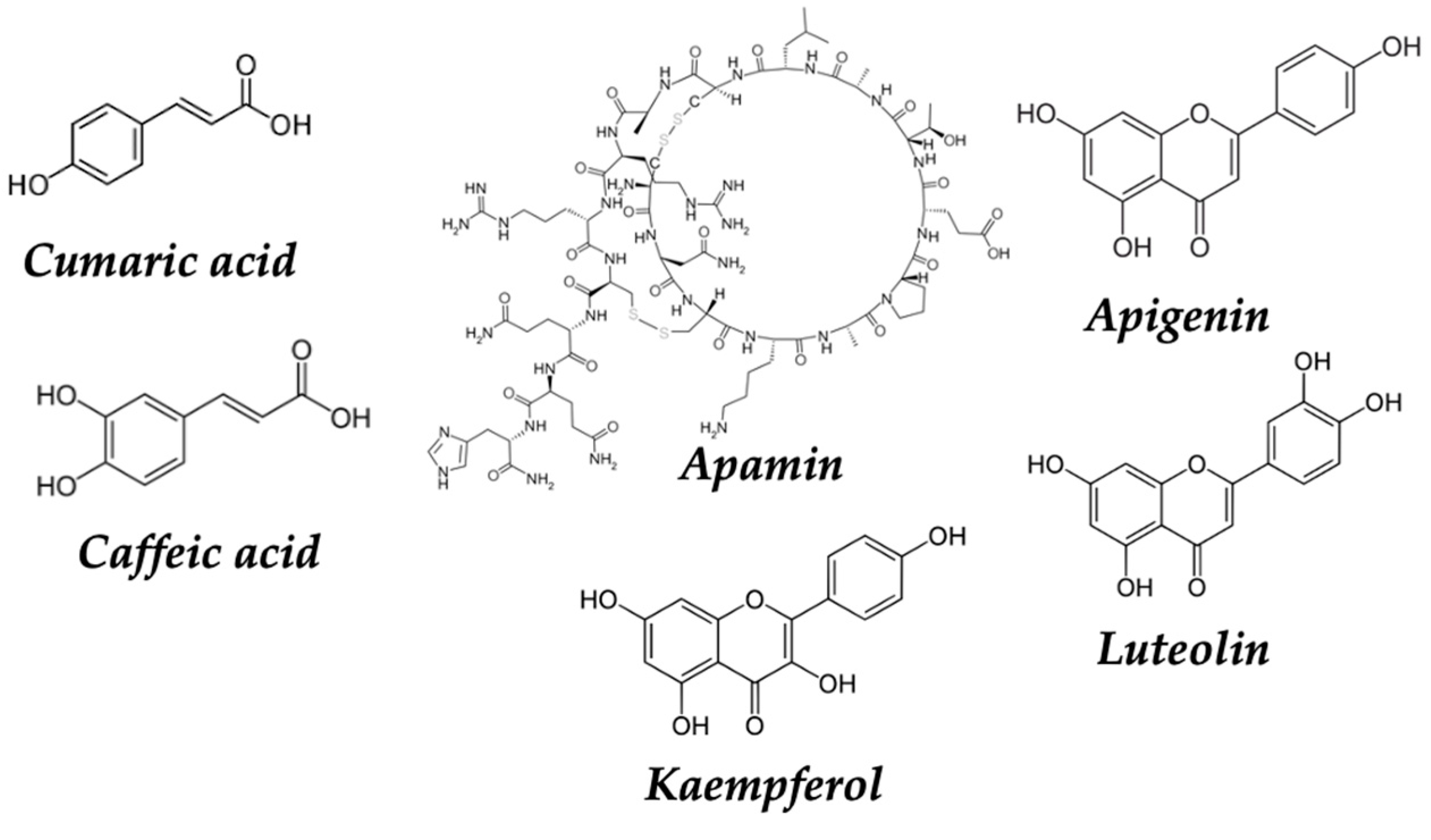
Figure 19.
a) 4DGN crystal structure: Crystal Structure of maize CK2 in complex with the inhibitor Luteolinin, PDB: https://doi.org/10.2210/pdb4DGN/pdbb) 4DGM. crystal structure: Crystal Structure of maize CK2 in complex with the inhibitor apigenin. PDB: https://doi.org/10.2210/pdb4DGM/pdb, created with NGL [113].
Figure 19.
a) 4DGN crystal structure: Crystal Structure of maize CK2 in complex with the inhibitor Luteolinin, PDB: https://doi.org/10.2210/pdb4DGN/pdbb) 4DGM. crystal structure: Crystal Structure of maize CK2 in complex with the inhibitor apigenin. PDB: https://doi.org/10.2210/pdb4DGM/pdb, created with NGL [113].

Figure 20.
a) 5AUX crystal structure: Crystal structure of DAPK1 in complex with kaempferol, PDB: https://doi.org/10.2210/pdb5AUX/pdb. b) 4N0S crystal structure: Crystal Structure showing the complex of ERK2 with caffeic acid. PDB: https://doi.org/10.2210/pdb4N0S/pdb, created with NGL [113].
Figure 20.
a) 5AUX crystal structure: Crystal structure of DAPK1 in complex with kaempferol, PDB: https://doi.org/10.2210/pdb5AUX/pdb. b) 4N0S crystal structure: Crystal Structure showing the complex of ERK2 with caffeic acid. PDB: https://doi.org/10.2210/pdb4N0S/pdb, created with NGL [113].
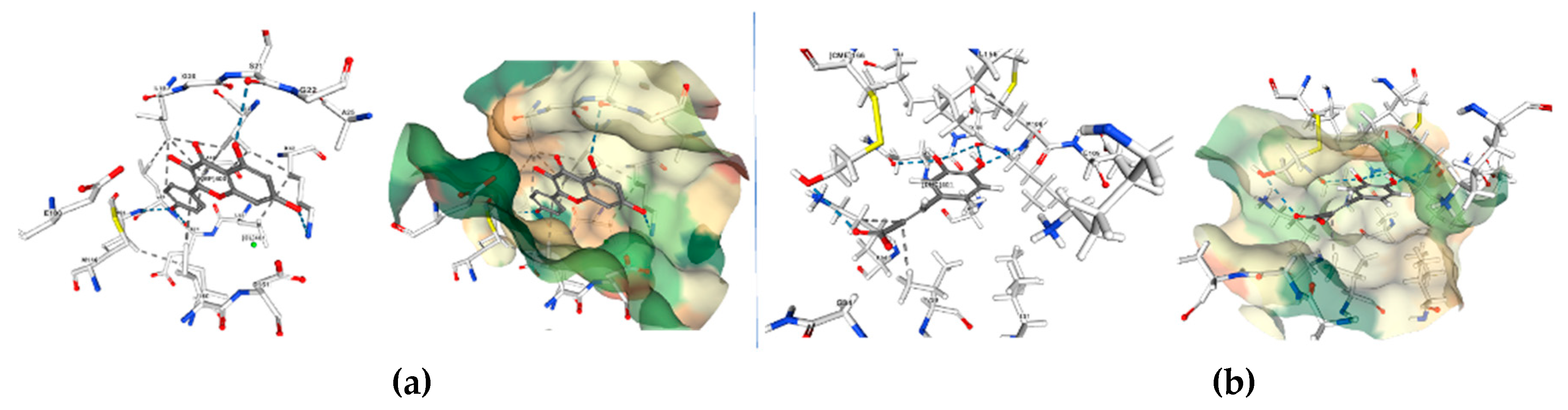
Figure 21.
4HDA crystal structure: Crystal structure of human Sirt5 in complex with Fluor-de-Lys peptide and resveratrol, PDB: https://doi.org/10.2210/pdb4HDA/pdb, created with NGL [113].
Figure 21.
4HDA crystal structure: Crystal structure of human Sirt5 in complex with Fluor-de-Lys peptide and resveratrol, PDB: https://doi.org/10.2210/pdb4HDA/pdb, created with NGL [113].
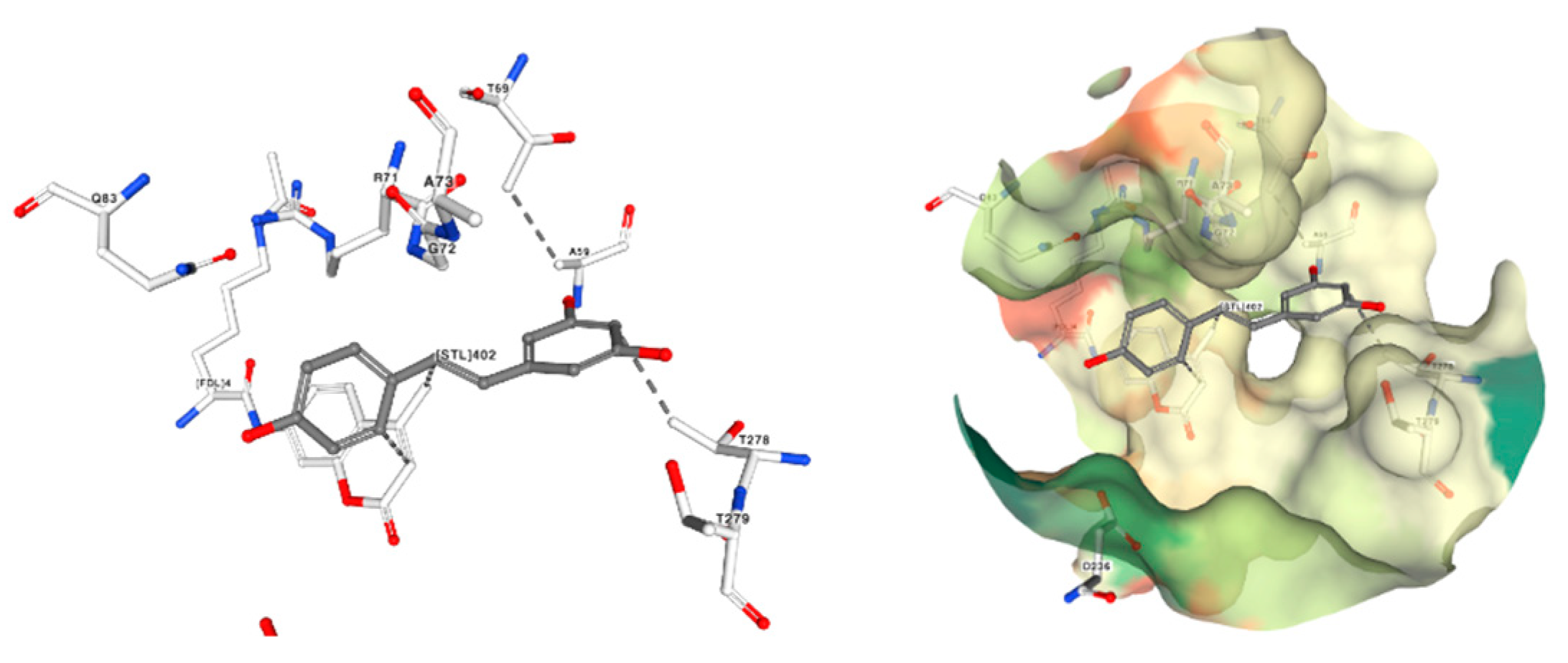
Figure 22.
Some of the main bioactive components of Galician wines.
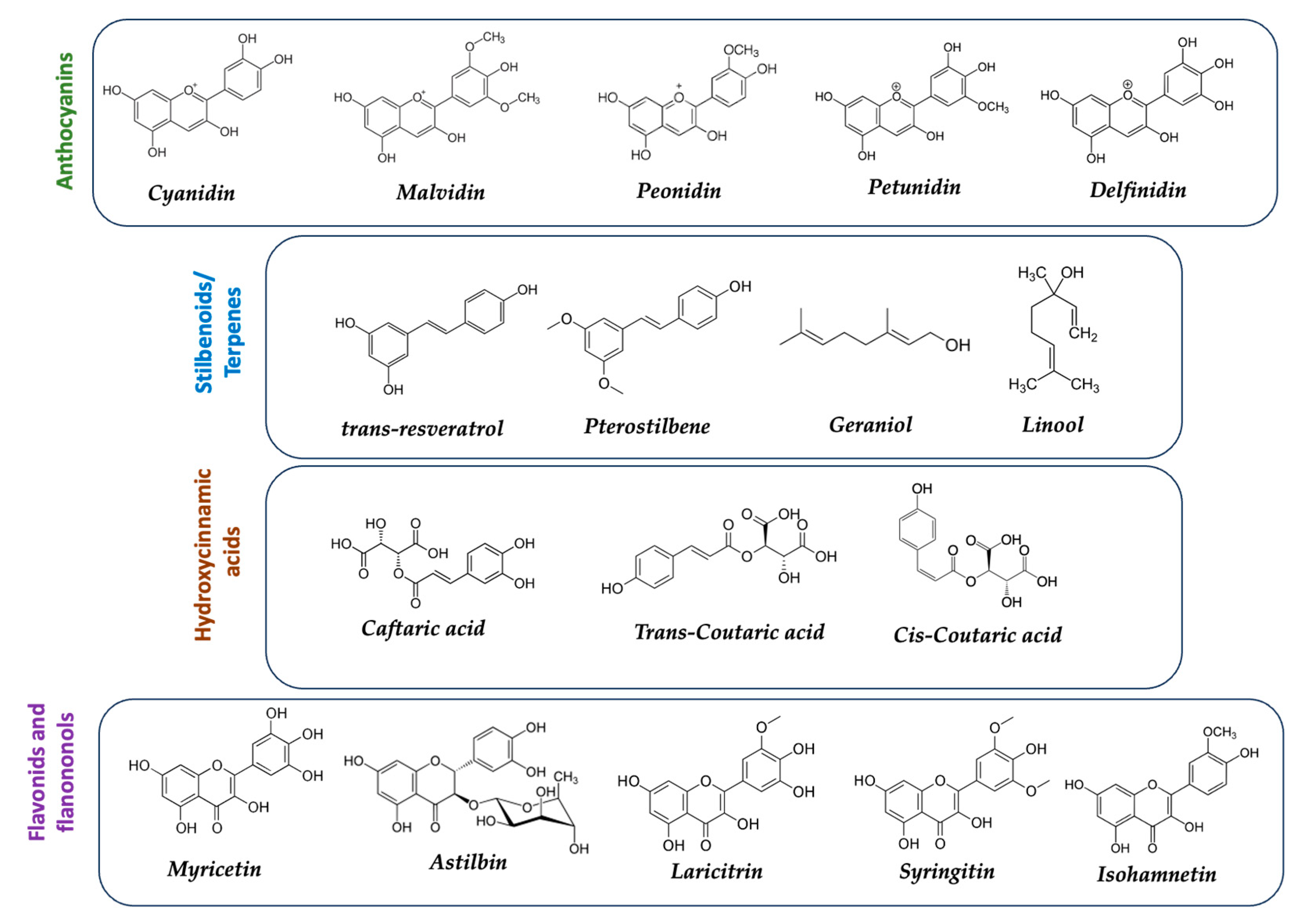
Figure 23.
a) Chemical structure of β-Casomorphin. b) Milk and dairy derivatives consumption in Spain. Self-made. Source: Ministry of Agriculture, Fisheries and Food. Food consumption report of Spain, 2022. Government of Spain. Source: [14].
Figure 23.
a) Chemical structure of β-Casomorphin. b) Milk and dairy derivatives consumption in Spain. Self-made. Source: Ministry of Agriculture, Fisheries and Food. Food consumption report of Spain, 2022. Government of Spain. Source: [14].
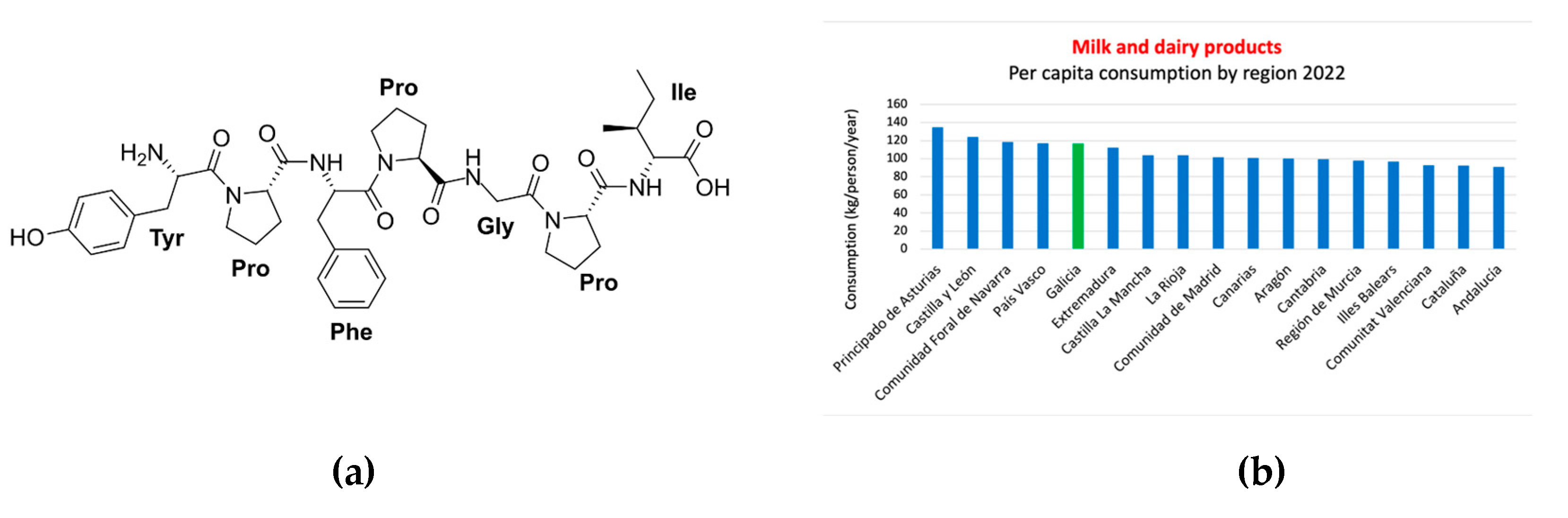

Table 1.
Relationship between the main foods of the SEAD, their key components, and the reported pharmacological activity.
Table 1.
Relationship between the main foods of the SEAD, their key components, and the reported pharmacological activity.
| Dietary source | Main bioactive component | Activity | Reference |
|---|---|---|---|
| Fish, shellfish, seaweed | Omega-3 fatty acids | Antioxidant, Anti-inflammatory, Hypocholesterolemic | [25] |
| Betaine (TMG, Anserine, Carnosine) | Anticancer, antioxidant, Anti-inflammatory | [67,68] | |
| Selenium | Antioxidant | [59,60] | |
| Vitamin D | Epigenetic modifications | [89] | |
| Fucoxanthin | Antioxidant | [24] | |
| Octopus | OCTPEP-1 | Anticancer | [83] |
| Selenium | Antioxidant | [59] | |
| Taurin | Antiproliferation | [74] | |
| Ozopromide | Antioxidant, anti-inflammatory | [87,88] | |
| Brassicaceae family | Glucosinolate | Antioxidant, anti-inflammatory, anticancer | [37,92] |
| Sulphorafane | Antioxidant, anti-inflammatory, anticancer | [37,96,97,98,99] | |
| Quercetin | Antioxidant, anti-inflammatory, anticancer | [111,112,115,117] | |
| Indole-3-carbinol | Anticancer | [55,117,123] | |
| 3,3-Diindolylmethane | Anticancer | [120,121] | |
| Onion, garlic, leek | Ajoene | Anticancer | [126] |
| Allicin | Antioxidant, anticancer | [127] | |
| Diallyl sulfide | Antiproliferative, Anticancer | [128,129] | |
| Potatoes | Carotenoids | Antioxidant | [131,132,133] |
| Vitamin C | Antioxidant | [131,132,133] | |
| Peppers of Padrón | Capsaicine | Anticancer, Anti-inflammatory | [138,139,140,141,142,143,144,145] |
| b-Caroten | Antioxidant, Cardioprotective | [134] | |
| Olive oil | Hydroxytyrosol | Anticancer | [149,150,151] |
| Oleuropein | Anticancer | [155,156] | |
| Ligstroside | Antioxidant | [157,158] | |
| Oleocanthal | Anti-inflammatory | [154] | |
| Squalene | Antioxidant | [158] | |
| Chestnuts | Tannins | Antioxidant, Anticancer | [164,165] |
| Escin | Anticancer | [165,166,167] | |
| Ellagic acid | Antioxidant, Anticancer | [169] | |
| Rutin | Antioxidant | [171] | |
| Honey | Apamin | Anticancer | [173,174] |
| Apigenin | Anticancer | [179] | |
| Kaempferol | Anticancer | [180,181,182,183,184] | |
| Wine | Resveratrol | Anticancer | [187,188,189,190,191,192] |
| Tannins | Antioxidant, anticancer | ||
| Milk and dairy derivatives | Bioactive peptides | Antioxidant, Anticancer | [200] |
| Lactoferricin | Antioxidant, Anticancer | [200] | |
| Casomorphins | Antioxidant, Anticancer | [201] | |
| Alpha-lactalbumin | Antioxidant, Anticancer | [203] | |
| Paprika, red pepper | Zeaxanthin, lutein | Antioxidant | [132,133,207,208,209] |
| Supplements | Sulphorafane | Anticancer | [37,96,97,98,99] |
| Caspsaicinoids | Antioxidant, Anti-inflammatory, Anticanalgesic | [208] | |
| Hidroxytyrosol | Antioxidant, Anticancer | [213] | |
| Zeaxanthin, lutein | Antioxidant | [132,133] | |
| Indole-3-carbinol | Anticancer | [214,215] | |
| Phlorotannins | Antioxidant, Anticáncer | [217] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



