
Submitted:
11 August 2023
Posted:
14 August 2023
You are already at the latest version
Abstract
The dogs with canine visceral leishmaniasis (CVL) play a key role in the transmission cycle of Leishmania infantum to humans in the urban environment. There is a consensus regarding the importance of developing a vaccine to control this disease. Despite many efforts to develop a protective vaccine against CVL, the ones currently available, CaniLeish®, Leish-tec® and LetiFend® have limited effectiveness. This is due, in part, to the complexity of the immune response of the naturally infected dogs against the parasite and the complexity of the parasite transmission cycle. Thus, strategies such as the development of a transmission-blocking vaccines (TBVs), already being applied to other vector-borne diseases like malaria and dengue, would be an attractive alternative to control leishmaniasis. TBVs induce the production of antibodies in the vertebrate host, which can inhibit parasite development in the vector and/or interfere with aspects of vector biology, leading to an interruption of parasite transmission. To date, there are few TBVs studies for CVL and other leishmaniasis forms. However, the few studies that exist show promising results justifying further development of this approach.
Keywords:
Canine visceral leishmaniasis
; vaccines
; transmission-blocking vaccines.
1. Introduction
Dogs with visceral leishmaniasis (VL) play a key role as reservoirs of Leishmania infantum protozoan parasites in urban areas. Canines are considered natural reservoirs because the parasites can survive in skin macrophages, facilitating the transmission to vectors [1]. Indeed, a substantial overlap between locations where human cases are detected and high canine seroprevalence have been reported, underscoring the close relationship between canine and human infections [1,2,3,4]. The vector Lutzomyia longipalpis, described by Lutz and Neiva (1912), accounts for 90% of VL transmissions in Latin America [5,6].
The recommended control measures by Health Ministry in Brazil include treatment of patients with the disease, usage of insecticides, and euthanasia of infected dogs [7]. Despite some studies showing that eliminating seropositive dogs reduces the incidence of VL in both dogs and humans [8,9,10], the current scenario shows that the euthanasia of dogs is far from solving the problem of parasite transmission [7,11,12,13,14]. In addition to being controversial among researchers, euthanasia of infected dogs has been harshly criticized from an ethical point of view, in addition to not being well accepted by tutors.
Upon all the limitations that control programs face, vaccination is considered the most cost-effective control tool for human and canine diseases [15,16]. Thus, the development of vaccines against CVL remains a priority. To achieve this goal, the development of new strategies to control parasite transmission is a pressing need. The present review aims to briefly describe the available CVL vaccines as well as their drawbacks, emphasizing the potential of transmission-blocking strategies as innovative tools to ultimately prevent disease.
2. Canine visceral leishmaniasis immunology and currently available commercial vaccines
The outcome of the CVL is influenced by the parasite species and the host's immune response [17,18,19]. Despite the complexity of the disease and the different clinical signs that dogs can exhibit, it has already been shown that the resistance profile is associated with a strong induction of a Th1 response, with the production of IL-12, IFN-γ, IL-2 and TNF-α. Conversely, a Th2 profile, including the cytokines IL-4, IL-5, IL-10, IL-13, and TGF-β, is related to susceptibility [19,20,21,22,23,24,25]. The compartmentalized organ-specific immune response has been associated with both granuloma maturation in liver and antileishmanial activity in L. donovani infection (Stäger et al., 2003).
In L. infantum naturally-infected dogs, the immunophenotypic profile of peripheral blood cells showed a prominent reduction in the absolute numbers of total CD5+ T-cells and their T-cells subsets (CD4+ and CD8+) [19,26]. Both oligosymptomatic and symptomatic dogs usually have a predominantly Th2 immune response, which are correlated with disease progression and high parasitism [19,25,27,28,29,30]. Notably, a characteristic of disease susceptibility is the proliferation of non-immunoprotective B-cells, due to the depletion of T-cells in dogs with high parasite loads [31]. An increase in total IgG levels has been observed in oligosymptomatic and symptomatic dogs, when compared to healthy and asymptomatic dogs [32,33,34].
It has currently been established that a vaccine against CVL represents a critical tool to controlling both human and canine cases [15,19] and a better understanding of the immunology behind the resistance and susceptibility profile is essential for the development of vaccines against CVL [35]. Studies focusing on the development of vaccine candidates have increased in recent years and, to date, there are only three commercially available vaccines for CVL: CaniLeish® (Virbac, France), Leish-Tec® (Ceva, Brazil), and LetiFend® (LETIPharma, Spain). However, more recently, Brazilian Ministry of Agriculture and Livestock determined the suspension of the manufacture and sale of Leish-Tec® and, so far, new recommendations are being awaited [36].
CaniLeish® was first placed on the market in 2011 in Europe and consists of purified excreted-secreted proteins from L. infantum (LiESP) associated with the adjuvant Quil-A saponin. This vaccine induces an immune response characterized by an increased T-cell producing IFN-γ and decreased in vitro parasite load in macrophages, correlated with NO production, characteristics of a Th1 response [37,38]. The leishmanicidal activity, NO, and IFN-γ production were observed in vitro after incubation using lymphocytes from vaccinated dogs with L. infantum-infected macrophages [39]. Later, Lemesre and colleagues (2007) observed, in a double-blind randomized efficacy field trial with dogs, an increase of IFN-γ and NO production and the efficacy of vaccination was 92% [40]. In 2014, Martin et al. demonstrated that, after L. infantum experimental infection in LiESP/QA-21 vaccinated dogs, there was an increase in IFN-γ, iNOS and NO production, accompanied by a low parasite burden [41]. Lastly, Oliva et al. (2014) observed an efficacy of 68.4% after exposure of vaccinated dogs to natural infection during two consecutive VL transmission seasons [42].
Leish-Tec® was licensed in 2014 and is the only vaccine available authorized for use in Brazil. It consists of recombinant A2 (rA2) protein, which was the first to be identified as the amastigote-specific virulence factor in Leishmania [43], with the adjuvant saponin. Fernandes et al. (2008) observed that the vaccinated dogs produced higher IFN-γ levels, however, parasites were detected in 57.14% and 28.5% bone marrow and blood of vaccinated animals, respectively. The authors concluded that immunization with rA2 was immunogenic and able to provide partial protection in dogs [44]. Afterward, Regina-Silva et al. (2016) demonstrated that vaccinated dogs presented higher levels of total IgG, IgG2, and IgG1 anti-A2 when compared to the control group. In addition, analysis of parasitological exams and xenodiagnosis demonstrated an efficacy of 58.1% [43]. Subsequently, Grimaldi et al. (2017) showed the same antibody production than the aforementioned study in vaccinated animals in a field trial; however, 26.49% of the dogs converted to a seropositive status and 43% of the vaccinated dogs developed the disease over time. The authors concluded that Leish-Tec® offers promising results, however, it needs to be optimized to ensure efficacy in dogs under field conditions [45]. Finally, Aguiar-Soares et al. (2020) reaffirmed the vaccine's ability to induce increased IFN-γ production by CD8+ T-cells [46].
The vaccine LetiFend® was authorized for use in Europe in 2016 [46]. This formulation consists of the Protein Q, which is a genetic fusion of five antigenic fragments from four L. infantum proteins, namely acidic ribosomal proteins Lip2a, Lip2b, LiP0, and the histone H2A, without an adjuvant [47]. Molano et al. (2003) were the first to test the Protein Q in association with Bacillus Calmette-Guérin (BCG) as adjuvant and demonstrated long-lasting cellular and humoral responses and activation in macrophages to produce NO [48]. Moreover, the vaccine triggered a delayed-type hypersensitivity (DTH) and the vaccinated and infected dogs were asymptomatic [49]. Lastly, Cotrina et al. (2018) showed that Protein Q induced 72% efficacy in preventing clinical cases of CVL. The authors concluded that LetiFend® is safe and lowers the risk of developing CVL clinical signs [50]. The main characteristics of the commercial vaccines for CVL discussed in this section are summarized in Table 1.
The studies briefly discussed above indicate that the commercially available vaccines have limitations and none of them are 100% effective, meaning that vaccinated dogs could still be infected with L. infantum. Some researchers consider their efficacy limited and may interfere with the interpretation of serological tests for dog disease diagnosis [13]. In addition, there are still few studies that analyze xenodiagnosis and, due to this, it is not known for sure whether these vaccines are capable of impacting the transmission of the parasite. Developing an ideal vaccine is far from an easy task due to parasite antigenic and biological complexity and its ability to evade the host's immune response [51]. The ideal vaccine, in addition to protecting the vaccinated animal, should be capable of interrupting the parasite's transmission cycle. To circumvent the problems related to the transmission of the parasite, TBV have been gaining ground and have shown promising results in interfering with the biological cycle of vectors [52].
3. Development of transmission-blocking vaccines (TBVs): a strategy to interrupt pathogens transmission
The development of TBVs stands out as an approach that provides collective protection, since it aims to disrupt the pathogen’s transmission chain by vectors [52]. The principle of this type of vaccine is to use vector or parasite antigens to induce the production of antibodies against the vector or against the parasite. These antibodies will likely interfere with the pathogen’s survival or virulence in the vector, and with the biological aspects of the vector. Thus, feeding on a vaccinated and infected host would reduce vector competence parasite transmission [53].
The first attempts to develop TBVs were related to malaria control, aiming to interrupt or reduce the transmission cycle by targeting the reproductive initial stages of malarial parasites [54,55]. Membrane proteins, such as Pvs25 and Pvs28, expressed on the surface of parasite’s zygotes and ookinetes have been extensively studied for Plasmodium vivax’s TBV, effectively suppressing the development of ookinetes in mosquitoes [56,57]. The use of polyclonal anti-midgut antibodies blocked the development of both Plasmodium falciparum and Plasmodium vivax parasites in five different species of mosquitoes, reducing mosquito survival and fecundity. These data revealed the potential use of antibodies for the development of vaccines against such midgut receptors [58].
Other candidates for TBVs, such as the monoclonal antibodies (MG96) binding to the midgut glycoproteins of Anopheles stephensi, resulted in a 100% dose-dependent blockade against Plasmodium yoelii development in the vector midgut [59]. The glycoprotein from Anopheles gambiae aminopeptidase N glycoprotein (AgAPN1), which is a target for Jacalin (lectin), plays an important role in inhibiting ookinete attachment by masking glycan ligands on midgut epithelial surface glycoproteins. The α-AgAPN1 anti-IgG strongly inhibited both Plasmodium berghei and P. falciparum development in different mosquito species, implying that the glycoprotein has a conserved role in ookinete invasion of the midgut and, therefore, may be a target for the development of TBVs for malaria control [60]. Another important molecule involved in the establishment of Plasmodium falciparum infection in Anopheles gambiae midgut is Carboxypeptidase B (CPB). The addition of antibodies directed against the carboxypeptidase gene (CPBAg1) to a P. falciparum-containing blood meal inhibited CPB activity and blocked parasite development in the midgut [61]. Although TBV candidates have shown good results when it comes to controlling the transmission of Plasmodium sp. and the survival of the vectors, there are still no studies in humans that attest to the safety and efficacy of this approach in controlling malaria.
In addition to malaria, TBVs have already been tested for other vector-borne diseases. In the 1990s, Ramasamy et al. showed that feeding Aedes aegypti with blood from vaccinated animals resulted in a significant reduction in susceptibility to infection by the Ross River virus and Murray Valley encephalitis virus [62]. Similarly, Ramasamy and Ramasamy (1990) demonstrated that antibodies generated by mice immunized with Anopheles farauti midgut antigens significantly reduce the number of P. berghei oocytes developing in the vector [63]. Notably, antigens from A. aegypti demonstrated a noteworthy performance against the mosquito cycle by up to 90% [64]. Finally, studies of TBVs are not restricted to only these diseases [65].
4. Development of TBVs for the control of leishmaniasis
The use of sandfly antigens to develop vaccines for leishmaniasis control started in 1996. In the first study, hamsters were immunized with different concentrations of sandfly gut antigens. After three doses, Phlebotomus duboscqi fed directly on these vaccinated animals. The authors showed an increase in P. duboscqi-specific IgG antibodies production in the vaccinated animals, in addition to increased sandfly mortality and reduction in egg production [66]. Later, Tonui et al. (2001) immunized mice with crude whole parasites, rgp63, LPG or a cocktail containing rgp63 and LPG, all derived from Leishmania major. After immunization, these vaccinated animals were infected with L. major and, then, P. duboscqi fed directly on these animals. They observed the lowest infection rates as well as impairment in parasite development, when compared to the sandflies fed on the control group [67]. Kamhawi et al. (2004) immunized mice with PpGalec, a galectin reported in the midgut of the Phlebotomus papatasi. After five immunizations the sandflies were submitted to artificial feeding containing L. major parasites and antibodies triggered by immunization. A reduction of L. major promastigotes in the midgut of these vectors was observed [68].
Using a different approach, in the study conducted by Vilela et al. (2006) rabbits were immunized through repeated sandfly bites and L. longipalpis females fed directly on these vaccinated animals. The authors observed a decline in the fecundity of these sandflies, in addition to the increased mortality of females [69]. In that same year, Saraiva et al. (2006) conducted a study to evaluate the performance of the Leishmune® vaccine as TBV in dogs. The animals were immunized with the vaccine and the infection of L. longipalpis females with L. chagasi was accessed by feeding through a chick-skin membrane, in the presence of immunized dog’s immunized sera plus L. chagasi parasites. The authors observed a decrease of 20.7% in infection index, demonstrated by the ability to block the attachment of parasites in the midgut and by the lower rate of parasite infection [70]. Even though Leishmune® displayed some interference in Leishmania infection in sandflies, although it does not contain vector antigens in its formulation and was not developed as a TBV, in 2014 its commercialization was suspended in Brazil for not meeting the requirements of the phase III study [71].
Coutinho-Abreu et al. (2010) identified a possible target for TBV. In that study, the authors induced knockdown of PpChit1 transcripts through injection of dsRNA into the P. papatasi thorax. Mice were immunized with PpChit1 and the serum was collected. After the blood meal containing L. major and the serum of immunized animals a significant reduction in promastigotes present in the midgut of infected P. papatasi was observed [72].
Bongiorno et al. (2013) proposed to evaluate the ability of the vaccine CaniLeish® to act as a TBV. In this study, vaccinated and naturally infected dogs were exposed directly to Phlebotomus perniciosus. The sandflies that fed on vaccinated animals had a lower rate of infection, in addition to having a lower parasite load in the midgut, when compared to those sandflies that fed on the control group [73]. Although this vaccine was not developed as TBV and does not contain vector antigens in its formulation, these results are promising, however, further studies are needed to assess the real ability of CaniLeish® to act as a TBV. The main findings of TBVs discussed in this section are summarized in Table 2.
In addition to the studies mentioned above, several groups have been studying the use of vector antigens incorporated into vaccines, especially for VL, associated or not with parasite antigens. However, these vaccines were intended to evaluate only the protection against the development of infection. Despite several studies demonstrating promising efficacy, these vaccines have not been tested as TBV. Thus, the field of TBV for leishmaniasis is still little explored, despite the promising results obtained in the few existing studies.
5. Conclusion: Future trends and perspectives of TBVs for visceral leishmaniasis control
Although commercially available vaccines for use in dogs lead to protective immune responses, there is still an important bottleneck in demonstrating the effectiveness in preventing parasitic transmission to the vector. Therefore, new strategies are required to control the vector and the transmission of the parasite, obtaining an effective reduction in the number of cases of canine and human disease. In this sense, the development of TBVs for dogs using antigens from the vector could be an important strategy for controlling the spread of VL, given that the antibodies produced in the vaccinated dog could prevent parasite development in the vector.
In fact, for achievieving the expected effects of TBV, it is first necessary to carry out the rational selection of antigens. Notably, important physiological events occur after sandfly blood meals interfering with the host immune system effector mechanisms such as complement activation [74]. The result is the establishment of Leishmania infection in the host demonstrating its importance for parasite survival. In this context, the induction of specific antibody production in the host could contribute to reduce the parasite persistence in the sandfly [52]. Furthermore, several studies have used salivary antigens in association with Leishmania antigens to evaluate vaccine protection in the vertebrate host, with promising results (Table 2). However, new studies are required to verify their ability to interrupt the parasite transmission specially in dogs, which have an important role as reservoirs in L. infantum transmission. Regardless of the type of antigen used, it is possible to hypothesize that those related to physiology of sandflies resulting in a successful blood meal could be more promising antigen candidates to compose a TBV formulation. Although the studies available so far have demonstrated the effect on the vector biological cycle and on infection, the field of TBVs lacks studies that demonstrate how antibody-vector interaction occurs and how these antibodies exert their effect. One hypothesis is that antibodies induced by TBV would damage epithelial cells in sandfly midgut. This hypothesis was supported by the observed degeneration of sandfly midgut epithelium, accompanied by a decrease in the L. major infection [67]. However, further studies are needed to demonstrate such interactions and their effects at the cellular level.
The selection of parasite antigens to act as TBV should take their biological effect under consideration. In fact, some molecules have been described as important role in the parasite-vector interaction, such as lipophosphoglycan (LPG). It is known that the LPG is extremely important for the attachment of the parasite to the intestinal epithelium of the vector and, therefore, escape from the peritrophic matrix and thus avoid elimination with the faecal bolus [75]. However, new studies are required to evaluate how such antibodies could exert their function as TBV. Even though TBVs are different from current commercially available vaccines, they must also fit within certain criteria, such as safety, reproducibility and allow for large-scale and cost-effective production [76]. Furthermore, there are other criteria that TBVs candidates must follow, such as (i) inducing high antibody titers in the vaccinated animal, so that it can exert the effect on the vector and (ii) display low levels of polymorphisms [54]. Moreover, one of the greatest challenges in developing TBVs is the ability to maintain high antibody titers in the vaccinated animal, especially due to the nature of the immunizer, since most of the proposed antigens are not normally found in the host [65]. Despite the progress, there are still important points to be clarified, such as (i) if the proteases activated by the blood meal, especially trypsin, can compromise the biological action of the ingested antibody and how this can present an obstacle to the development of TBVs; (ii) how long the activity of the antibodies can be maintained in the vector and (iii) what is the minimum time required for the antibody to trigger its biological action in the vector.
Regardless of its promising effects, the field of TBVs in leishmaniasis is still little explored. This could be due to some reasons, such as (i) the difficulty to maintain the vector under laboratory conditions, due to need of proper infrastructure and trained professionals [77]; (ii) the complexity of finding an antigen that is capable of inducing high titers of antibodies in the host and being able, at the same time, to interfere with biological aspects of the vector and also in blocking the binding of the parasite to the vector; or even (iii) that efforts are more focused on the development of vaccines that protect the host against infection, taking the focus away from the TBVs strategy. In fact, studies of vaccines that contain vector antigens are increasing, since many of the studies demonstrate protection against infection in the vaccinated animal. Lastly, most TBVs studies were performed under laboratory conditions, which may mask the real effectiveness of the antigens tested. In this sense, it is necessary to identify promising antigens to compose TBVs to act in the control of the transmission of leishmaniasis, in addition to the need to carry out field tests to verify the real effectiveness of this strategy.
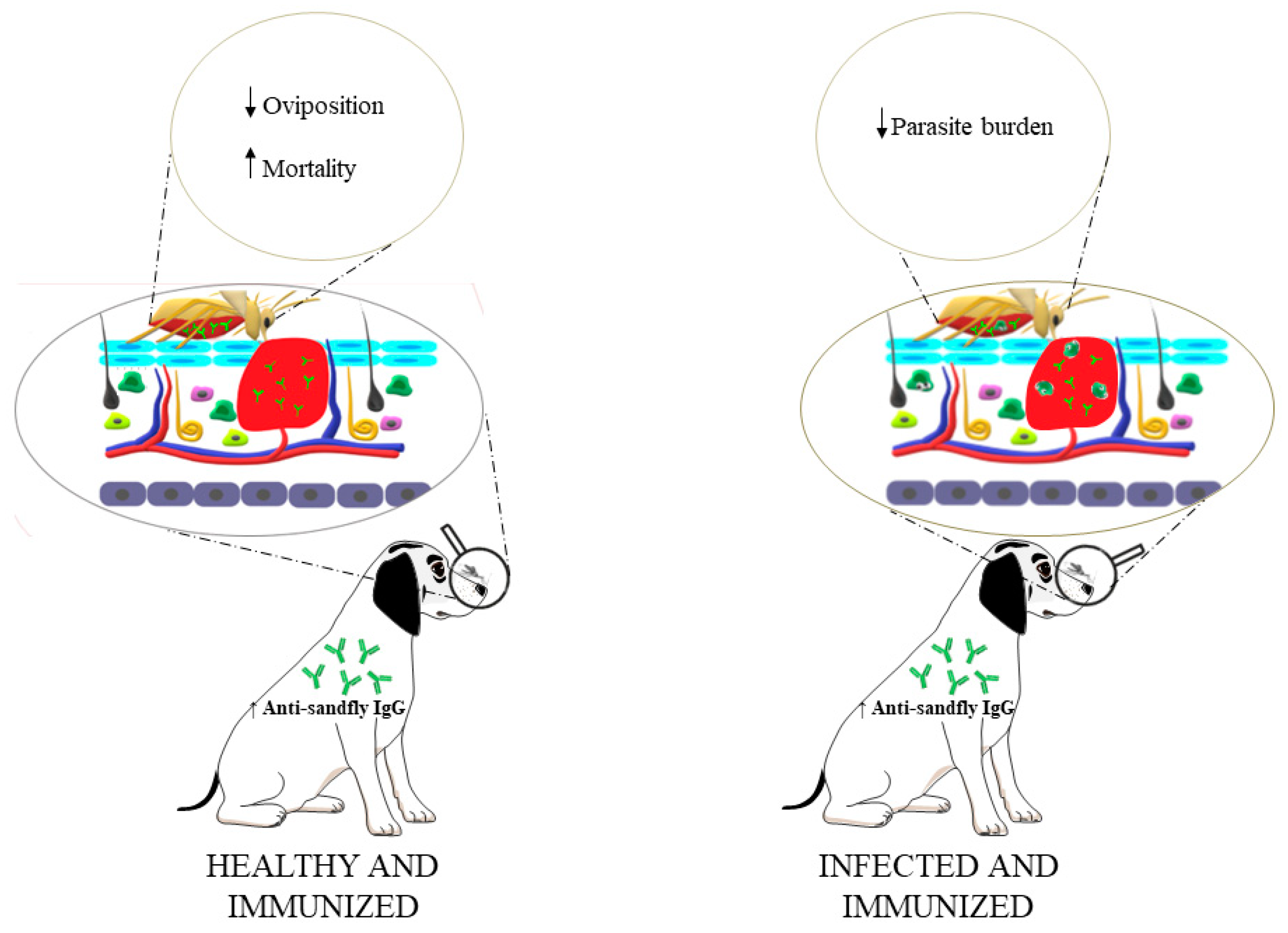
Our research group has already identified different L. longipalpis antigens that could be used in this promising strategy. These antigens showed the ability to exert the two main goals of a TBV: (i) alter the insect's homeostasis leading to death or reduced oviposition; and/or (ii) reduce the parasite load in the vector's intestine (Figure 1) [25,52,64,78,79]. Although these antigens showed excellent results, more studies are needed to evaluate their performance under field conditions. In addition, these sandfly antigens could be incorporated with new Leishmania antigens and/or in association with commercially available vaccines to improve the control of parasite transmission in VL endemic areas. Since the antibodies induced by vaccination prevent parasite development in the insect and its subsequent transmission, interrupting the epidemiological cycle becomes possible and preventing human and canine VL cases would then be feasible. Another TBV advantage would be to allow the safe treatment of infected dogs, without compromising human’s and other animal’s health. The field of TBVs should gain more focus in the coming years, as it presents a promising new strategy to effectively control transmission.
Figure 1.
Transmission-blocking vaccines (TBVs) for canine leishmaniasis control. The arrows ↑ or ↓ indicate the increase or decrease in the related parameters evaluated in the sandfly, respectively.
Figure 1.
Transmission-blocking vaccines (TBVs) for canine leishmaniasis control. The arrows ↑ or ↓ indicate the increase or decrease in the related parameters evaluated in the sandfly, respectively.
Author Contributions
JCL, AAMG, DSO, LAR, RMSM, ENN, JCF-C and RCG wrote the manuscript. JCL, AAMG, and HR assisted in drawing the figures. DFSP, AVS, DFVB, MACF and PCCR reviewed the manuscript. JCL, AAMG, ASG, PROP, MMM, ESD, WOD, DS-L and RCG drafted and critically evaluated the manuscript.
Funding
This research was financially supported through grants from CAPES (Coordination for the Improvement of Higher Education Personnel, Brazil), CNPq (National Council for Scientific and Technological Development), Fundação de Amparo à Pesquisa de Minas Gerais (FAPEMIG); Departamento de Ciência e Tecnologia do Ministério da Saúde do Brasil; and QUATREE (GranvitaPet).
Acknowledgments
ASG: MMM, ESD, WOD and RCG would like to thank CNPq for their research fellowships. We thank Randall Johnson for reviewing the language of the revised version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fraga,B.M.; Solcá, M.S.; Silva, V.M.; Borja, L.S.; Nascimento, E.G.; Oliveira, G.G.S.; Pontes-de-Carvalho, L.C.; Veras, P.S.T. S.; dos Santos, W.L.C. Temporal distribution of positive results of tests for detecting Leishmania infection in stray dogs of an endemic area of visceral leishmaniasis in the Brazilian tropics: A 13 years survey and association with human disease. Vet Parasitol 2012, 21, 591–594. [CrossRef]
- Cunha, A.M.; Chagas, E. Estudos sobre o parasito. In: Leishmaniose visceral americana, nova entidade mórbida do homem na América do Sul. Mem. Inst. Oswaldo Cruz 1937, 32, 329–337. [Google Scholar] [CrossRef]
- Maurício, I.L.; Stothard, J.R.; Miles, M.A. The strange case of Leishmania chagasi. Parasitol Today 2000, 16, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.M.; Vieira, C.P.; Dibo, M.R.; Guirado, M.M.; Rodas, L.A.C.; Chiaravalloti-Neto, F. Dispersal of Lutzomyia longipalpis and expansion of canine and human visceral leishmaniasis in São Paulo State, Brazil. Acta Trop 2016, 164, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Lutz, A.; Neiva, A. Contribuição para o conhecimento das espécies do genero Phlebotomus existentes no Brazil. Memórias do Instituto Oswaldo Cruz 1912, 4, 84–95. [Google Scholar] [CrossRef]
- Soares, R.P.P.; Turco, S.J. Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae): A review. An Acad Bras Ciênc 2003, 75, 301–330. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Miró, G.; Bowman, D.D.; Gradoni, L.; Otranto, D. Culling Dogs for Zoonotic Visceral Leishmaniasis Control: The Wind of Change. Trends Parasitol 2019, 35, 97–101. [Google Scholar] [CrossRef]
- Ashford, D.A.; David, J.R.; Freire, M.S.; David, R.A.; Sherlock, Í.R. de A.; Eulálio, M. da C.; Sampaio, D.B.P.; Badaró, R.J. da S. Studies on Control of Visceral Leishmaniasis: Impact of Dog Control on Canine and Human Visceral Leishmaniasis in Jacobina, Bahia, Brazil. American Journal of Tropical Medicine and Hygiene 1998, 59, 53–57. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B.; Batista-de-Melo, L.M.; Borja-Cabrera, G.P.; Palatnik, M.; Lavor, C.C. Improving methods for epidemiological control of canine visceral leishmaniasis based on a mathematical model. Impact on the incidence of the canine and human disease. An Acad Bras Cienc 2004, 76, 583–593. [Google Scholar] [CrossRef]
- Nunes, C.M.; Pires, M.M.; da Silva, K.M.; Assis, F.D.; Filho,J. G.; Perri, S.H.V. Relationship between dog culling and incidence of human visceral leishmaniasis in an endemic area. Veterinary Parasitology 2010, 170, 131–133. [Google Scholar] [CrossRef]
- Romero, G.A.; Boelaert, M. Control of visceral leishmaniasis in latin america-a systematic review. PLoS Negl Trop Dis 2010, 19, e584. [Google Scholar] [CrossRef]
- Costa, C.H. How effective is dog culling in controlling zoonotic visceral leishmaniasis? A critical evaluation of the science, politics and ethics behind this public health policy. Rev Soc Bras Med Trop 2011, 44, 232–242. [Google Scholar] [CrossRef]
- Marcondes, M. , Day, M.J. (2019). Current status and management of canine leishmaniasis in Latin America. Res Vet Sci 2019, 123, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Vaz, T.P.; Gama-Melo, M.O.; Quaresma, P.F.; Gontijo, C.M.F.; Santos, G.; Barbosa, F.S.; Fontes, G. Evaluation of the Euthanasia of Seropositive Dogs for Canine Visceral Leishmaniasis as the Only Method of Controling the Disease in the Enzootic Area in the Midwestern Minas Gerais. Pesquisa Veterinaria Brasileira 2020, 40, 107–112. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B. Vaccines for Canine Leishmaniasis. Front Immunol 2012, 17, 3:69. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Jain, N.K. Vaccines for visceral leishmaniasis: A review. J. of Immunological Methods 2015, 422, 1–12. [Google Scholar] [CrossRef]
- McMahon-Pratt, D.; Alexander, J. (2004). Does the Leishmania major paradigm of pathogenesis and protection hold for new world cutaneous leishmaniases or the visceral disease? Immunol Rev 2004, 201, 206–224. [Google Scholar] [CrossRef] [PubMed]
- Roatt, B.; Aguiar-Soares, R.D.O.; Coura-Vital, W.; Ker, H.G.; Moreira, N.D.; Vitoriano-Souza, J.; Giunchetti, R.C.; Carneiro, C.M.; Reis, A.B. Immunotherapy and immunochemotherapy in visceral leishmaniasis: Promising treatments for this neglected disease. Frontiers Immunology 13;5:272. 2014. [Google Scholar] [CrossRef]
- Giunchetti, R.C.; Silveira, P.; Resende, L.A.; Leite, J.C.; de Oliveira Melo-Júnior, O.A.; Rodrigues Alves, M.L.; Costa, L.M.; Lair, D.F.; Chaves, V.R.; Soares, I.S.; Mendonça, L.Z.; Lanna, M.F.; Ribeiro, H.S.; Maia-Gonçalves, A.A.; Santos, T.A.P.; Roatt, B.R.; Aguiar-Soares, R.D.O.; Vitoriano-Souza, J.; Moreira, N.D.; Mathias, F.A.S.; Cardoso, J.M.O.; Coura-Vital, W.; Galdino, A.S.; Viana, K.F.; Martins-Filho, O.A.; Silveira-Lemos, D.; Dutra, W.O.; Reis, A.B. Canine visceral leishmaniasis biomarkers and their employment in vaccines. Vet Parasitol 2019, 271, 87–97. [Google Scholar] [CrossRef]
- Pinelli, E.; Rutten, V.P.M.; Bruysters, M.; Moore, P.F.; Ruitemberg, J. Compensation for decreased expression of B7 molecules on Leishmania infantum-infected canine macrophages results in restoration of parasite-specific T-cell proliferation and gamma interferon production. Infect Immun 1999, 67, 237–243. [Google Scholar] [CrossRef]
- Pinelli, E.; Gonzalo, R.M.; Boog, C.J.; Rutten, V.P.; Gebhard, D.; del Real, G.; Ruitenberg, J. Leishmania infantum-specific T cell lines derived from asymptomatic dogs that lyse infected macrophages in a major histocompatibility complex-restricted manner. Eur J Immunol 1995, 25, 1594–1600. [Google Scholar] [CrossRef]
- Lage, R.S.; Oliveira, G.C.; Buzek, S.C.U.; Guerra, L.L.; Giunchetti, R.C.; Corrêa-Oliveira, R.; Reis, A.B. Analysis of the cytokine profile in spleen cells from dogs naturally infected by Leishmania chagasi. Vet Immunol Immunopathol 2007, 115, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.B.; Giunchetti, R.C.; Carrillo, E.; Martins-Filho, O.A.; Moreno, J. Immunity to Leishmania and the rational search for vaccines against canine leishmaniasis. Immunoparasitology series 2010, 26, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.A.; Alexandre, P.G.; Soares, C.M.; Marques, C.; Rodrigues, O.R.; Brito, T.V.; Fonseca, I.P.; Alves, L.C.; Santos-Gomes, G.M. Cytokine gene expression in the tissues of dogs infected by Leishmania infantum. J Comp Pathol 2011, 145, 336–344. [Google Scholar] [CrossRef]
- Gonçalves, A.A.M.; Leite, J.C.; Resende, L.A.; Mariano, R.M.S.; Silveira, P.; Melo-Júnior, O.A.O.; Ribeiro, H.S.; de Oliveira, D.S.; Soares, D.F.; Santos, T.A.P.; Marques, A.F.; Galdino, A.S.; Martins-Filho, O.A.; Dutra, W.O.; da Silveira-Lemos, D. , Giunchetti, R.C. An Overview of Immunotherapeutic Approaches Against Canine Visceral Leishmaniasis: What Has Been Tested on Dogs and a New Perspective on Improving Treatment Efficacy. Front Cell Infect Microbiol 2019, 18, 9:427. [Google Scholar] [CrossRef]
- Leal, G.G.D.A.; Roatt, B.M.; Aguiar-Soares, R.D.O.; Carneiro, C.M.; Giunchetti, R.C.; Teixeira-Carvalho, A.; Martins-Filho,O. A.; Francisco, A.F.; Cardoso, J.M.; Mathias, F.A.S.; Correa-Oliveira, R.; Carneiro, M.; Coura-Vital, W.; Reis, A.B. Immunological profile of resistance and susceptibility in naturally infected dogs by Leishmania infantum. Vet Parasitol 2014, 205, 472–482. [Google Scholar] [CrossRef]
- Barbieri, C.L. Immunology of canine leishmaniasis. Parasite Immunol 2006, 28, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.F.; Moura, E.P.; Ribeiro, R.R.; Michalick, M.S.; Kalapothakis, E.; Bruna-Romero, O.; Tafuri, W.L.; Teixeira, M.M.; Melo, M.N. Expression of IFN-gamma, TNF-alpha, IL-10 and TGF-beta in lymph nodes associates with parasite load and clinical form of disease in dogs naturally infected with Leishmania (Leishmania) chagasi. Vet Immunol Immunopathol 2009, 128, 349–358. [Google Scholar] [CrossRef]
- Do Nascimento,P. R.; Martins, D.R.; Monteiro,G.R.; Queiroz, P.V.; Freire-Neto, F.P.; Queiroz, J.W.; Lima, A.L.M.; Jeroimo, S.M.B. Association of pro-inflammatory cytokines and iron regulatory protein 2 (IRP2) with Leishmania burden in canine visceral leishmaniasis. PLoS ONE 2013, 11, e73873. [Google Scholar] [CrossRef]
- Moreira, M.L.; Costa-Pereira, C.; Alves, M.L.; Marteleto, B.H.; Ribeiro,V. M.; Peruhype-Magalhães, V.; Giunchetti, R.C.; Martins-Filho, O.A.; Araújo, M.S. Vaccination against canine leishmaniosis increases the phagocytic activity, nitric oxide production and expression of cell activation/migration molecules in neutrophils and monocytes. Vet Parasitol 2016, 15, 233–245. [Google Scholar] [CrossRef]
- Koutinas, A.F.; Koutinas, C.K. Pathologic Mechanisms Underlying the Clinical Findings in Canine Leishmaniosis due to Leishmania infantum/chagasi. Vet Pathol 2014, 51, 527–538. [Google Scholar] [CrossRef]
- Carson, C.; Antoniou, M.; Ruiz-Argüello, M.B.; Alcami, A.; Christodoulou, V.; Messaritakis, I.; Blackwell, J.M.; Courtenay, O. A prime/boost DNA/ Modified vaccinia virus Ankara vaccine expressing recombinant Leishmania DNA encoding TRYP is safe and immunogenic in outbred dogs, the reservoir of zoonotic visceral leishmaniasis. Vaccine 2009, 27, 1080–1086. [Google Scholar] [CrossRef]
- Chaabouni, A.; Boubaker, E.R.; Mhadhbi, M.; Gharbi, M.; Sassi, A. Comparative analysis of the Leishmania infantum-specific antibody repertoires and the autoantibody repertoires between asymptomatic and symptomatic dogs. Vet Parasitol 2018, 15, 9–17. [Google Scholar] [CrossRef]
- De Freitas, J.C.; Lopes-Neto, B.E.; de Abreu, C.R.; Coura-Vital, W.; Braga, S.L.; Reis, A.B.; Nunes-Pinheiro, D.C. Profile of anti-Leishmania antibodies related to clinical picture in canine visceral leishmaniasis. Res Vet Sci 2012, 93, 705–709. [Google Scholar] [CrossRef]
- Srivastava, S.; Shankar, P.; Mishra, J.; Singh, S. Possibilities and challenges for developing a successful vaccine for leishmaniasis. Parasit Vectors 2016, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Brasil. Ministério da Agricultura e Pecuária. Mapa suspende fabricação e venda e determina o recolhimento de lotes de vacina contra Leishmaniose. Available at: https://www.gov.br/agricultura/pt-br/assuntos/noticias/mapa-suspende-fabricacao-e-venda-e-determina-o-recolhimento-de-lotes-de-vacina-contra-leishmaniose-apos-fiscalizacao. Access in: 0731/2023.
- Lemesre, J.L.; Holzmuller, P.; Cavaleyra, M.; Gonçalves, R.B.; Hottin, G.; Papierok, G. Protection against experimental visceral leishmaniasis infection in dogs immunized with purified excreted secreted antigens of Leishmania infantum promastigotes. Vaccine 2005, 23, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.; Vouldoukis, I.; Martin, V.; McGahie, D.; Cuisinier, A.M.; Gueguen, S. Use of a liesp/qa-21 vaccine (canileish) stimulates an appropriate th1-dominated cell-mediated immune response in dogs. PLoS Negl Trop Dis 2012, 6, e1683. [Google Scholar] [CrossRef]
- Holzmuller, P.; Cavaleyra, M.; Moreaux, J.; Kovacic, R.; Vincendeau, P.; Papierok, G.; Lemesre, J.L. Lymphocytes of dogs immunized with purified excreted-secreted antigens of Leishmania infantum co-incubated with Leishmania infected macrophages produce IFN gamma resulting in nitric oxide-mediated amastigote apoptosis. Vet Immunol Immunopathol 2005, 106, 247–257. [Google Scholar] [CrossRef]
- Lemesre, J.L.; Holzmuller, P.; Gonçalves, R.B.; Bourdoiseau, G.; Hugnet, C.; Cavaleyra, M.; Papierok, G. Long-lasting protection against canine visceral leishmaniasis using the LiESAp-MDP vaccine in endemic areas of France: Double-blind randomised efficacy field trial. Vaccine 2007, 25, 4223–4234. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Vouldoukis, I.; Moreno, J.; McGahie, D.; Gueguen, S.; Cuisinier, A.M. The protective immune response produced in dogs after primary vaccination with the LiESP/QA-21 vaccine (CaniLeish®) remains effective against an experimental challenge one year later. Vet Res 2014, 25, 45:69. [Google Scholar] [CrossRef]
- Oliva, G.; Nieto, J.; Foglia Manzillo, V.; Cappiello, S.; Fiorentino, E.; Di Muccio, T.; Scalone, A.; Moreno, J.; Chicharro, C.; Carrillo, E.; Butaud, T.; Guegand, L.; Martin, V.; Cuisinier, A.M.; McGahie, D.; Gueguen, S.; Cañavate, C.; Gradoni, L. (2014). A Randomised, Double-Blind, Controlled Efficacy Trial of the LiESP/QA-21 Vaccine in Naïve Dogs Exposed to Two Leishmania infantum Transmission Seasons. PLoS Negl Trop Dis Oct 2014, 9, e3213. [Google Scholar] [CrossRef]
- Regina-Silva, S.; Feres, A.M.L.T.; França-Silva, J.C.; Dias, E.S.; Michalsky, É.M.; de Andrade, H.M.; Coelho, E.A.F.; Ribeiro, G.M.; Fernandes, A.P.; Machado-Coelho, G.L.L. Field randomized trial to evaluate the efficacy of the Leish-Tec® vaccine against canine visceral leishmaniasis in an endemic area of Brazil. Vaccine 2016, 34, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.P.; Costa, M.M.S.; Coelho, E.A.F.; Michalick, M.S.M.; de Freitas, E.; Melo, M.N.; Tafuri, W.L.; Resende, D.M.; Hermont, V.; Abrantes, C.F.; Gazzinelli, R.T. Protective immunity against challenge with Leishmania (Leishmania) chagasi in beagle dogs vaccinated with recombinant A2 protein. Vaccine 2008, 26, 5888–5895. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, G.; Teva, A.; Dos-Santos, C.B.; Santos, F.N.; Pinto, I.D.S.; Fux, B.; Leite, G.R.; Falqueto, A. Field trial of efficacy of the Leish-tec® vaccine against canine leishmaniasis caused by Leishmania infantum in an endemic area with high transmission rates. PLoS ONE 2017, 12, e0185438. [Google Scholar] [CrossRef]
- Aguiar-Soares, R.D. de O.; Roatt, B.M.; Mathias, F.A.S.; Reis, L.E.S.; Cardoso, J.M. de O.; Brito, R.C.F.; Ker, H.G.; Corrêa-Oliveira, R.; Giunchetti, R.C.; Reis, A.B. Phase I and II Clinical Trial Comparing the LBSap, Leishmune®, and Leish-Tec® Vaccines against Canine Visceral Leishmaniasis. Vaccines 2020, 8, 690. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency, 2016. LetiFend_: Summary of Product Characteristics. http://www.ema.europa.eu/docs/en_GB/document_library/EPAR_-_Summary_for_the_public/veterinary/003865/WC500207742.pdf (acessed ,2022). 18 May.
- Molano, L.; Alonso, G.M.; Mirón, C.; Redondo, E.; Requena, J.M.; Soto, M.; Cieto, C.G.; Alonso, C. A Leishmania infantum multi-component antigenic protein mixed with live BCG confers protection to dogs experimentally infected with L. infantum. Vet Immunol Immunopathol 2003, 92, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Carcelén, J.; Iniesta, V.; Fernández-Cotrinaa, J.; Serrano, F.; Parejob, J.C.; Corralizac, I.; Gallardo-Solerc, A.; Marañnón, F.; Soto, M.; Alonsoe, C.; Gómez-Nietoa, C. The Chimerical Multi-Component Q protein from Leishmania in the absence of adjuvant protects dogs against an experimental Leishmania infantum infection. Vaccine 2009, 27, 5964–5973. [Google Scholar] [CrossRef]
- Cotrina, J.F.; Iniesta, V.; Monroy, I.; Baz, V.; Hugnet, C.; Marañon, F.; Fabra, M.; Gómez-Nieto, L.C.; Alonso, C. A large-scale field randomized trial demonstrates safety and efficacy of the vaccine LetiFend® against canine leishmaniosis. Vaccine 2018, 36, 1972–1982. [Google Scholar] [CrossRef]
- Kaye, P.M.; Cruz, I.; Picado, A.; Van Bocxlaer, K.; Croft, S.L. Leishmaniasis immunopathology—Impact on design and use of vaccines, diagnostics and drugs. Semin Immunopathol 2020, 42, 247–264. [Google Scholar] [CrossRef]
- Graciano, R.C.D.; Ribeiro, J.A.T.; Macedo, A.K.S.; de S Lavareda, J.P.; de Oliveira, P.R.; Netto, J.B.; Nogueira, L.M.; Machado, J.M.; Camposda-Paz, M.; Giunchetti, R.C.; Galdino, A.S. Recent Patents applications in red biotechnology: A mini-review. Recent Pat Biotechnol 2019, 13, 170–186. [Google Scholar] [CrossRef]
- Coutinho-Abreu, I.V.; Ramalho-Ortigao, M. Transmission blocking vaccines to control insect-borne diseases - A review. Mem Inst Oswaldo Cruz 2010, 105, 1–12. [Google Scholar] [CrossRef]
- Coelho, C.H.; Rappuoli, R.; Hotez, P.J.; Duffy, P.E. Transmission-Blocking Vaccines for Malaria: Time to Talk about Vaccine Introduction. Trends Parasitol 2019, 35, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Lê, H.G.; Kang, J.M.; Jun, H.; Lee, J.; Moe, M.; Thái, T.L.; Lin, K.; Myint, M.K.; Yoo, W.G.; Sohn, W.-M.; Kim, T.-S.; Na, B.K. Genetic diversity and natural selection of transmission-blocking vaccine candidate antigens Pvs25 and Pvs28 in Plasmodium vivax Myanmar isolates. Acta Trop 2019, 198, 105104. [Google Scholar] [CrossRef]
- Grotendorst, C.A.; Kumar, N.; Carter, R.; Kaushal, D.C. A surface protein expressed during the transformation of zygotes of Plasmodium gallinaceum is a target of transmission-blocking antibodies. Infect Immun 1984, 45, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Hisaeda, H.; Collins, W.E.; Saul, A.; Stowers, A.W. Antibodies to Plasmodium vivax transmission-blocking vaccine candidate antigens Pvs25 and Pvs28 do not show synergism. Vaccine 2001, 20, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Lal, A.A.; Patterson, P.S.; Sacci, J.B.; Vaughan, J.A.; Paul, C.; Collins, W.E.; Wirtz, R.A.; Azad, A.F. Anti-mosquito midgut antibodies block development of Plasmodium falciparum and Plasmodium vivax in multiple species of Anopheles mosquitoes and reduce vector fecundity and survivorship. Proc Natl Acad Sci 2002, 98, 5228–5233. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, R.R.; Fields, I.; Shahabuddin, M.; Azad, A.F.; Sacci, J.B.Jr. Monoclonal antibody MG96 completely blocks Plasmodium yoelii development in Anopheles stephensi. Infect Immun 2003, 71, 6995–7001. [Google Scholar] [CrossRef]
- Dinglasan, R.R.; Kalume, D.E.; Kanzok, S.M.; Ghosh, A.K.; Muratova, O.; Pandey, A.; Jacobs-Lorena, M. Disruption of Plasmodium falciparum development by antibodies against a conserved mosquito midgut antigen. Proc Natl Acad Sci 2007, 104, 13461–13466. [Google Scholar] [CrossRef]
- Lavazec, C.; Boudin, C.; Lacroix, R.; Bonnet, S.; Diop, A.; Thiberge, S.; Boisson, B.; Tahar, R.; Bourgouin, C. Carboxypeptidases B of Anopheles gambiae as targets for a Plasmodium falciparum transmission-blocking vaccine. Infect Immun 2007, 75, 1635–1642. [Google Scholar] [CrossRef]
- Ramasamy, M.S.; Sands, M.; Kay, B.H.; Fanning, I.D.; Lawrence, G.W.; Ramasamy, R. Anti-mosquito antibodies reduce the susceptibility of Aedes aegypti to arbovirus infection. Med Vet Entomol 1990, 4, 49–55. [Google Scholar] [CrossRef]
- Ramasamy, M.S.; Ramasamy, R. Effect of anti-mosquito antibodies on the infectivity of the rodent malaria parasite Plasmodium berghei to Anopheles farauti. Med Vet Entomol 1990, 4, 161–166. [Google Scholar] [CrossRef]
- Rodrigues-Alves, M.L.; Melo-Júnior, O.A.O.; Silveira, P.; Mariano, R.M.D. S.; Leite, J.C.; Santos, T.A.P.; Soares, I.S.; Lair, D.F.; Melo, M.M.; Resende, L.A.; da Silveira-Lemos, D.; Dutra, W.O.; Gontijo, N.F.; Araujo, R.N.; Sant'Anna, M.R.V.; Andrade, L.A.F.; da Fonseca, F.G.; Moreira, L.A.; Giunchetti, R.C. Historical Perspective and Biotechnological Trends to Block Arboviruses Transmission by Controlling Aedes aegypti Mosquitos Using Different Approaches. Front Med 2020, 23, 275. [Google Scholar] [CrossRef]
- Londono-Renteria, B.; Troupin, A.; Colpitts, T.M. Arbovirosis and potential transmission blocking vaccines. In Parasites and Vectors 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Ingonga, P.; Mbati, P.A.; Anjili, C.O.; Mutani, A.; Wishitemi, B.; Odongo, S.; Robert, L.L.; Githure, J.I. The effect of immune sera from hamsters immunized with sandfly gut and whole body extract antigens on the fecundity and mortality of Phlebotomus duboscqi (Diptera: Psychodidae). Acta Trop 1996, 60, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Tonui, W.K.; Mbati, P.A.; Anjili, C.O.; Orago, A.S.; Turco, S.J.; Githure, J.I.; Koech, D.K. Transmission Blocking Vaccine Studies in Leishmaniasis: I. Lipophosphoglycan Is a Promising Transmission Blocking Vaccine Molecule against Cutaneous Leishmaniasis. East Afr Med J 2001, 78, 84–89. [Google Scholar] [CrossRef]
- Kamhawi, S.; Ramalho-Ortigao, M.; Pham, V.M.; Kumar, S.; Lawyer, P.G.; Turco, S.J.; Barillas-Mury, C.; Sacks, D.L.; Valenzuela, J.G. A Role for Insect Galectins in Parasite Survival. Cell 2004, 119, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Vilela, M.L.; Souza, N.A.; Oliveira, S.M.P.; Costa-Pinto, D.; Cabello, P.H.; Rangel, E.F.; Traub-Cseko, Y.M. Considerations on the effect of anti-sandfly antibodies on biological parameters of Lutzomyia longipalpis (Lutz & Neiva, 1912) (Diptera: Psychodidae: Phlebotominae). Braz J Biol. 2006, 66, 175–183. [Google Scholar] [CrossRef]
- Saraiva, E.M.; Barbosa, A.D.F.; Santos, F.N.; Borja-Cabrera, G.P.; Nico, D.; Souza, L.O.P.; Mendes-Aguiar, C.D.O.; De Souza, E.P.; Fampa, P.; Parra, L.E.; Menz, I.; Dias, J.G.; De Oliveira, S.M.; Palatnik-De-Sousa, C.B. The FML-vaccine (Leishmune®) against canine visceral leishmaniasis: A transmission blocking vaccine. Vaccine 2006, 24, 2423–2431. [Google Scholar] [CrossRef]
- MAPA, 2014. Ministério da Agricultura, Pecuária e Abastecimento. Nota técnica n°038/2014/DFIP/DAS. Suspensão da licença de fabricação e comercialização do produto Leishmune vacina contra leishmanioses visceral canina. Brasília. www.agricultura.gov.br/assuntos/politica-agricola/arquivos/notatecnica-dfip-38-14- leishmune.pdf/view (acessed ,2022). 14 May.
- Coutinho-Abreu, I.V.; Sharma, N.K.; Robles-Murguia, M.; Ramalho-Ortigao, M. Targeting the Midgut Secreted Ppchit1 Reduces Leishmania Major Development in Its Natural Vector, the Sand Fly Phlebotomus Papatasi. PLoS Neglected Tropical Diseases 2010, 4. [Google Scholar] [CrossRef]
- Bongiorno, G.; Paparcone, R.; Manzillo, V.F.; Oliva, G.; Cuisinier, A.M.; Gradoni, L. Vaccination with LiESP/QA-21 (CaniLeish®) Reduces the Intensity of Infection in Phlebotomus Perniciosus Fed on Leishmania Infantum Infected Dogs-A Preliminary Xenodiagnosis Study. Veterinary Parasitology 2013, 197, 691–695. [Google Scholar] [CrossRef]
- Rios-Barros, L.V.; Silva-Moreira, A.L.; Horta, M.F.; Gontijo, N.F.; Castro-Gomes, T. How to get away with murder: The multiple strategies employed by pathogenic protozoa to avoid complement killing. Mol Immunol 2022, 149, 27–38. [Google Scholar] [CrossRef]
- Secundino, N.; Kimblin, N.; Peters, N. C.; Lawyer, P.; Capul, A. A.; Beverley, S. M.; Turco, S. J.; Sacks, D. Proteophosphoglycan confers resistance of Leishmania major to midgut digestive enzymes induced by blood feeding in vector sand flies. Cellular Microbiology, 2010, 12, 7, 906–918, Petitdidier, E.; Pagniez, J.; Pissarra, J.; Holzmuller, P.; Papierok, G.; Vincendeau, P.; Lemesre, J.L.; Bras-Gonçalves, R. Peptide-based vaccine successfully induces protective immunity against canine visceral leishmaniasis. npj Vaccines 2019, 4, 1–9. [CrossRef]
- Petitdidier, E.; Pagniez, J.; Pissarra, J.; Holzmuller, P.; Papierok, G.; Vincendeau, P.; Lemesre, J.L.; Bras-Gonçalves, R. Peptide-based vaccine successfully induces protective immunity against canine visceral leishmaniasis. npj Vaccines 2019, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lawyer, P.; Killick-Kendrick, M.; Rowland, T.; Rowton, E.; Volf, P. Laboratory colonization and mass rearing of phlebotomine sand flies (Diptera, Psychodidae). Parasite 2017, 24, 42. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, H.S.; Pereira, D.F.S.; Melo-Junior, O.; Mariano, R.M. da S.; Leite, J.C.; Silva, A.V. da, Oliveira, D.S. de, Gonçalves, A.A.M.; Lair, D.F.; Soares, I. dos S.; Santos, T.A.P.; Galdino, A.S.; Silveira-Lemos, D. da, Paes, P.R. de O.; Melo, M.M.; Dutra, W.O.; Araujo, R.N.; Giunchetti, R.C. Vaccine approaches applied to controlling dog ticks. Ticks and Tick-borne Diseases 2021, 12. [Google Scholar] [CrossRef]
- Giunchetti, R.C.; Filho, O. A. M.; Mendonça, L.Z.; Gontijo, N.F.; Bartholomeu, D.C.; Borges, W.C. Imunobiológico para controle do vetor da Leishmaniose, processos de obtenção e uso. Brazil Patent BR 10 2013 02 2805 2, 2013.

Table 1.
The main results of commercial vaccines for CVL.
| Continent/ Country |
Product | Company | Vaccine composition | Efficacy biomarkers | References |
|---|---|---|---|---|---|
| Europe | CaniLeish® | Virbac (France) | Purified excreted-secreted proteins of L. infantum (LiESP) associated with the adjuvant muramyl dipeptide (MDP) | ↑IFN-γ and nitrite; 75% of vaccinated dogs showed in vitro leishmanicidal effect | Holzmuller et al., 2005 [39] |
| ↑IFN-γ and nitrite; 100% efficacy (bone marrow culture) | Lemesre et al., 2005 [37] | ||||
| ↑IFN-γ and nitrite; 92% efficacy (bone marrow PCR) | Lemesre et al., 2007 [40] | ||||
| ↑IFN-γ, iNOS and nitrite; ↑ in vitro leishmanicidal effect | Moreno et al., 2012 [38] | ||||
| ↑IFN-γ, iNOS and nitrite; parasites were detected in bone marrow in 3/10 vaccinated dogs (qPCR) | Martin et al., 2014 [41] | ||||
| ↑IgG2; 68.4% efficacy (PCR and culture, serological and clinical examinations) | Oliva et al., 2014 [42] | ||||
| Brazil | Leish-Tec® | CEVA (France) | Recombinant A2 (rA2) protein with the adjuvant saponin | ↑IgG, IgG2, IFN-γ and IL-10; parasites detected in 57.14% (bone marrow culture) and 28.5% (blood PCR) in vaccinated dogs | Fernandes et al., 2008 [44] |
| ↑IgG, IgG2 and IgG1; 58.1% efficacy (bone marrow culture + xenodiagnoses) | Regina-Silva et al., 2016 [43] | ||||
| ↑IgG and ↑IgG2; 43% of vaccinated dogs developed the disease |
Grimaldi et al., 2017 [45] | ||||
| ↑ CD8+IFN-γ+ | Aguiar-Soares et al., 2020 [46] | ||||
| Europe | LetiFend® | LETI Laboratories (Spain) | Protein Q – a genetic fusion of five antigenic fragments from four L. infantum proteins, named acidic ribosomal proteins Lip2a, Lip2b, LiP0, and the histone H2A | ↑ DTH (9/10 vaccinated dogs); 90% of vaccinated dogs remain healthy (lymph nodes culture, clinical and anatomic-pathologic analysis) | Molano et al., 2003 [48] |
| ↑ NO production and DTH; parasites detected in vaccinated dogs (single dose): 1/7, 1/7 and 0/ 7 (PCR of skin, lymph node and spleen, respectively); parasites detected in vaccinated dogs (two doses): 4/7, 1/7 and 2/ 7 (PCR of skin, lymph node and spleen, respectively) | Carcelén et al., 2009 [49] | ||||
| ↑IgG2 anti-Protein Q; 72% efficacy (lymph nodes or bone marrow PCR and smear) | Cotrina et al., 2018 [50] |
The arrows (↑ and ↓) indicate the increase and decrease in biomarker levels, respectively, when compared to control groups. iNOS: inducible nitric oxide synthase; DTH: delayed hypersensitivity; NO: nitric oxide.
Table 2.
The main results of TBVs for leishmaniasis.
| TBV composition | Vector / Parasite | Vaccination schedule/ Animals/ Artificial or in vivo feeding | Evaluated parameters | Main findings | Reference |
|---|---|---|---|---|---|
| Sandfly gut antigens | Phlebotomus duboscqi | 1 IM dose, followed by 2 SC doses (14th and 21th day) / 24 hamsters / in vivo feeding | Humoral response; Survival and fecundity of sandflies | ↑ P. duboscqi-specific IgG; ↓ survival; egg production and egg hatching | Ingonga et al., 1996 [66] |
| Crude whole Leishmania major parasites or rgp63 or LPG or rgp63/LPG | Phlebotomus duboscqi / Leishmania major | 4 IV doses, at 7-day interval or 3 IV doses, at 14-day interval / BALB/c mice, posteriorly infected with L. major/ in vivo feeding | Humoral response; Infection rate in sandfly; Promastigote forms presented after blood meal; Histopathology of midgut | ↑ IgG anti-soluble L. major antigen; ↓ infection rate; ↓ infective metacyclic forms | Tonui et al., 2001 [67] |
| PpGalec | Phlebotomus papatasi / Leishmania major | 5 doses / BALB/c / Artificial feeding | Infection rate by ex vivo and in vivo analyses in sandflies | ↓ infection rate | Kamhawi et al., 2004 [68] |
| Repeated sandfly bites | Lutzomyia longipalpis | Repeated bites of 100-120 females / Rabits / in vivo feeding | Humoral response; sandfly survival and oviposition analysis | ↑ IgG anti-sandfly; ↑ mortality; ↓ oviposition | Vilela et al., 2006 [69] |
| Leishmune® | Lutzomyia longipalpis / Leishmania chagasi | 3 SC doses at 20-day interval / mongrel dogs / Artificial feeding | Infection rate (in vitro and in vivo analysis) in sandflies | ↑ L. chagasi binding to sandfly midguts; ↓ infection rate | Saraiva et al., 2006 [70] |
| PpChit1 | Phlebotomus papatasi /Leishmania major | 3 SC doses at 14-day interval / BALB/c / Artificial feeding | Infection rate in sandflies | ↓ infection rate | Coutinho-Abreu et al., 2010 [72] |
| CaniLeish® | Phlebotomus perniciosus / Leishmania infantum | 3 doses / beagle dogs, natural infected after vaccination / in vivo infection | Infection rate in sandflies | ↓ infection rate | Bongiorno et al., 2013 [73] |
The arrows (↑ and ↓) indicate the increase and decrease in biomarker levels, respectively, when compared to control groups. IgG: immunoglobulin; IM: intramuscular; IV: intravenous; SC: subcutaneous.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



