
Submitted:
11 August 2023
Posted:
15 August 2023
You are already at the latest version
Abstract
Stevia rebaudiana (Bert.) is a natural sweetener with diterpenic glycosides, offering 300 times more sweetness than sucrose, and boasting low-calorie content, along with medicinal and therapeutic properties. To address a limited genetic diversity, a germplasm bank comprising 115 segregated lines of stevia was established in Colombia. Notwithstanding, the fungus Athelia rolfsii adversely impacted key stevia lines, leading to wilting and plant mortality. For this reason our study aimed to explore the biocontrol potential of native strains of Trichoderma spp., isolated from productive lots in Colombia's Cordoba department, as a solution to manage A. rolfsii. Soil samples underwent serial dilution technique (103 conidia.mL-1) to obtain Trichoderma spp. isolates, which were thoroughly morphologically and molecularly characterized. A total five native strains were identified through sequencing the PCR products using universal ITS primers, and comparisons with the GENBANK database confirmed their identity (OK310695.1, ON237703.1, ON238106.1, ON237737.1, and ON237923.1). Their biocontrol efficacy against A. rolfsii was evaluated using the dual culture technique. Remarkably, all strains exhibited significant biocontrol capacity against the A. rolfsii isolate INVEPAR-05 (OK271308.1), except for Trichoderma viride INVEPAR-T10 (ON237923.1). These findings suggest that Trichoderma spp. could be a potential option to manage the fungus in stevia.
Keywords:
Antagonistic capacity
; Biocontrol agents
; ITS1-5.8s-ITS2 region
; Dual Cultures
1. Introduction
Stevia (Stevia rebaudiana Bert.) is a perennial herbaceous plant belonging to the Asteraceae family, which comprises approximately 230 species, of which about 20 are extensively cultivated, primarly as herbs or shrubs [1]. Among these 20 species, it was found that S. rebaudiana possesses the sweetest essence [2]. It produces diterpene glycosides that are 300 times sweeter than sucrose, making it a powerful, natural low-calorie sweetener with medicinal and therapeutic properties beneficial to human health.
Different varieties of S. rebaudiana were introduced to Colombia in the late 1980s. With the increase in its cultivation in the Colombia, a collaboration agreement was signed in 1994 between the Antioquia Department in Colombia and the Japan International Development Organization Ltda (JAIDO) to introduce of the Morita I, II, and III varieties. These varieties were developed in Japan by the plant breeder Toyosigue Morita, and many of the existing biological materials in the country stem from this introduction.
Due to the limited genetic diversity and the lack of profitable varieties adapted to different sub-regions of the country, challenges arise in the efficient utilization of the crop. To address this issue, the University of Córdoba in Colombia and the Institute of Plant Biotechnology (IBP) in Cuba have worked on generating segregated lines for a germplasm bank consisting of 115 genotypes [3]. Some important lines in this germplasm bank possess specific characteristics, such as late flowering, erect architecture, and significant contents of diterpene glycosides; however, these promising lines have been continuously affected by the fungus Athelia rolfsii (Curzi) C.C. Tu & Kimbr. (anamorph Sclerotium rolfsii Sacc.), as described by Mordue [4], leading to the wilting and death of these plants, causing a significant limitation in our research programs.
In this context, the stem rot of Stevia sp. caused by A. rolfsii is the most common disease affecting seedlings during multiplication in nursery rooting beds or propagation areas, resulting in losses of up to 40% of the seedlings. Damage to established plants in the field can vary from 10% to 40%, possibly due to latent infections acquired while in the nursery. Symptoms in seedlings affected by A. rolfsii include wilting and loss of turgor in leaves, sometimes accompanied by dark lesions at the base of the stem, leading to the gradual death of the plant. The fungus primarily affects the main stem, producing dark lesions and small brown sclerotia at its base and surroundings, while leaves touching the infected soil exhibit circular lesions with concentric rings [5]. In the field, the fungus causes yellowing and a partial or total wilting of the branches and leaves, with whitish mycelial growth covering the soil near the plant. In advanced stages, small white or brown sclerotia can be observed adhering to or surrounding the base of the stem, producing a large number of sclerotia, allowing the fungus to survive for long periods in the soil [6]. Therefore, preventive management, such as substrate treatment in rooting beds or acclimatization of micropropagated plants, is essential.
The first global report of stem rot in Stevia was by Chang, et al. [7] in Alberta, Canada. Subsequently, this disease has been reported worldwide. Its distribution, includes different countries such as Egypt, India, the United States, Italy, Turkey, France, and Ecuador, among others [8,9,10,11,12,13,14].
Although research has been conducted on the identification of A. rolfsii, the efficient management of this disease remains a challenge. Some studies on the chemical and biological control of A. rolfsii, the causative agent of stem rot in S. rebaudiana, have shown promising results. Hilal and Baiuomy [8] evaluated two commercial fungicides, topsin M (thiophanate-methyl) and vitavax (carboxin), along with two biocides, plant ward (Trichoderma harzianum) and Rizo N (Bacillus subtilis), to control fungal diseases in Stevia, including that caused by A. rolfsii, finding that the commercial fungicide topsin M was the most effective. Another study conducted by Shwetha and Hegde [15] performed an in vitro assessment of biorationals against A. rolfsii, discovering that cow urine completely inhibited the mycelial growth of the pathogen. Meanwhile, Koehler and Shew [6] carried out efficacy trials of fungicides for the management of A. rolfsii, evaluating azoxystrobin, flutolanil, and tebuconazole, and found that azoxystrobin was effective in disease control. In more recent studies, Sanabria-Velazquez, et al. [16] evaluated anaerobic soil disinfestation (ASD) and fungicide application for the management of stevia stem rot (SSR) and Septoria leaf spot (SLS) in laboratory and field experiments. The results revealed that the combination of ASD with organic fungicides, such as pyroligneous acid (PA), proved effective in reducing the viability of A. rolfsii sclerotia.
Given the recognized nature of stevia as a plant with medicinal and therapeutic properties beneficial to health, it is imperative to consider alternatives that avoid the use of agrochemicals and pesticides. In this context, it is necessary to identify and adopt agronomic practices that are both human health and the environmentally safe, without compromising crop productivity and profitability.
For this purpose we aimed to investigate the biocontrol capacity of native strains of Trichoderma spp. isolated from productive plots in the department of Córdoba, Colombia, against the A. rolfsii isolate INVEPAR-05. It is noteworthy, that understanding the biocontrol potential of native strains of Trichoderma spp. plays a fundamental role in achieving a sustainable agronomic approach, which in turn, would strengthen the comercial viability of stevia, thus improving the economic well-being of farmers in the context of building a new and long-awaited period of peace after a century of armed conflict in Colombia. Additionally, this strategy represents a sustainable and efficient option to mitigate the impact of diseases and increase stevia production in tropical regions like Colombia.
2. Materials and Methods
2.1. Location and Field Sampling
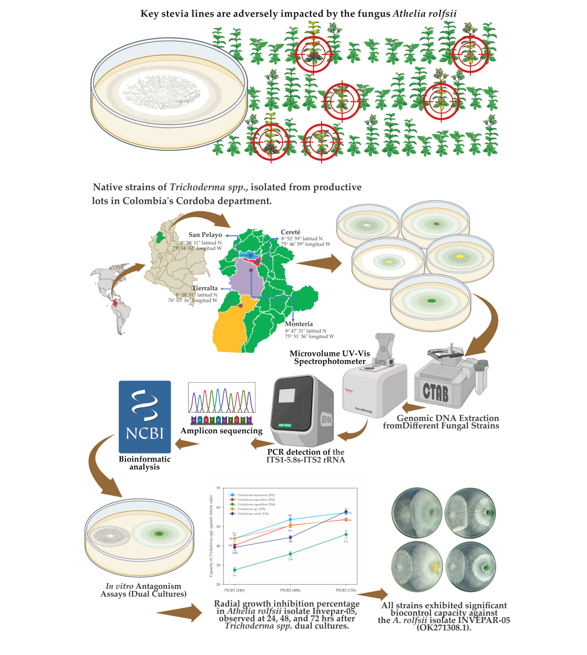
The research was carried out at the experimental campus, the main headquarters of the University of Córdoba, located in Montería, Colombia. The geographical coordinates of the site are 8° 47' 31" N latitude and 75° 51' 36" W longitude, with an altitude of 18 meters above sea level. The region experiences an average annual precipitation of 1,274 mm and an average temperature of 28°C. Total solar irradiance is approximately 1,800 light hours per year, while the relative humidity remains at 84% [17].
Soil samples were collected from four different cities: Montería, Cereté, San Pelayo, and Tierralta (Figure 1). Two fields under vegetable cultivation were carefully selected within each municipality for the sampling process. In each chosen field, a total of five soil sub-samples were collected randomly, situated 20 cm away from the base of the stem and at a depth of 20 cm. The sampling methodology adhered to the guidelines provided by Martyniuk and Martyniuk [18] and Tejera, et al. [19]. Subsequently, the collected sub-samples from each field were merged to create a composite sample, weighing approximately 500 g per lot.
2.2. Laboratory Processing
The isolation of Trichoderma spp. from soil samples was conducted following the serial dilution technique as described by Nelson, et al. [20]. A soil-water solution with a 1:1 ratio was prepared, and 1 mL of the solution was diluted to a concentration of 1x 10-3. From this dilution, 100 μL was then plated on Potato Dextrose Agar (PDA) culture medium. The resulting Trichoderma spp. colonies were transferred to Petri dishes containing the same medium to obtain pure cultures. On the other hand, the isolation of A. rolfsii was obtained from plants within the germplasm bank of stevia segregating lines that were affected by the fungus. Sclerotia were extracted from the infected tissues and cultured on PDA medium. The same methodology used for Trichoderma spp. purification was applied to obtain pure cultures of A. rolfsii. Subsequently, the pathogen was identified using the 16S marker, and the data was uploaded to the NCBI GenBank.
2.3. Molecular identification of Trichoderma spp and A. rolfsii
2.3.1. Genomic DNA Extraction from Different Fungal Strains
Genomic DNA was extracted using the modified Cetyltrimethylammonium Bromide (CTAB 2X) method [21]. A portion of fungal filaments from each strain was finely ground using liquid nitrogen in a crucible. Subsequently, 540 µL of CTAB 2X solution (as detailed in Table 1) and 60 µL of 10% PVP (Merck KGaA, Darmstadt, Germany, part number: P5288) were added to the samples. The mixtures were then incubated in a water bath at 65°C with vortex agitation every 15 minutes for a total of 60 minutes. Afterward, the samples were allowed to rest at room temperature for 5 minutes. Next, 900 µL of chloroform-isoamyl alcohol solution (24:1) (Merck KGaA, part number: 25666) was added, and the samples were gently inverted for 5 minutes. Following centrifugation at 15,000 x g for 5 minutes, the supernatant was carefully transferred to new 1.5 mL reaction tubes containing 600 µL of isopropanol (Merck KGaA, part number: I9516), which were mixed gently by inversion. The samples were then cooled at -20°C for 10 minutes, followed by a 3-minute centrifugation step to pellet the DNA. The supernatant was discarded, and the DNA pellets were washed with 600 µL of 70% ethanol (Merck KGaA, part number: 65350-M), centrifuged again, and the ethanol was carefully removed. Finally, 100 µL of DNA rehydration solution was added to each pellet. After DNA extraction, the quantity and purity were meticulously evaluated using 2 µL of each sample on a NanoDrop OneC spectrophotometer (Thermo Scientific, Missouri, TX, USA). The DNA quantity was measured at 260 nm, and purity was assessed by calculating the absorbance ratios at 260/280 nm and 260/230 nm [22].
2.3.2. Amplification of the fungal ITS fragment
Samples with higher concentrations and better quality of DNA were amplified using the PCR technique in a thermocycler (Bio-Rad T100™ thermocycler, Hercules, CA, USA), using the universal primers ITS4: 5'- TCCTCCGCTTATTGATATGC-3' and ITS3: 5′-GCATCGATGAAGAACGCAGC-3′ (Sigma-Aldrich, Darmstadt, Germany, part number 200-00485), which amplify the ITS1-5.8s-ITS2 region of the rRNA gene, [23]. The reaction solution was prepared in a final volume of 50 µL, which contained 25 µL of DreamTaq Hot Start PCR Master Mix (2X) (DreamTaq™ Hot Start PCR Master Mix, Thermo Fisher Scientific, part number K9011), 1 µL of each 10 mM primer, 2 µL of DNA (~157 ng/µL) and sufficient molecular grade water to reach the desired volume. The PCR products were evaluated by electrophoresis on 1% agarose gels in a horizontal electrophoresis chamber and then visually analyzed in a photodocumenter (Enduro GDS Labnet, Tewksbury, MA, USA). The size of the fragments was determined by comparison with a 1 Kb molecular weight marker (Sigma-Aldrich, Darmstadt, Germany, part number D0428).
2.3.3. Sequencing of PCR Products and Sequence Analysis
The PCR products were sequenced using the Sanger technique [14]. The reading of the sequences in AB1 format was performed with the 4Peaks software v 1.8 (Nucleobytes B.V. Gerberastraat 117 1431 RA Aalsmeer, The Netherlands. KvK Amsterdam 56362757), which is linked to the bioinformatic tool Blast, to compare each nucleotide problem se- quence (Query Sequence) with those in the GenBank database of the NCBI (United States National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov—accessed on Aug 03 2023).
2.4. In vitro Antagonism Assays (Dual Cultures)
To evaluate the mechanisms of antagonistic action of different Trichoderma spp. strains against Sclerotium sp, in vitro antagonism assays were performed using the method described by Bell [24]. The experiments followed a completely randomized (DCA), consisting of five treatments, with six replicates per treatment. Petri dishes with a diameter of 9 cm containing PDA medium were used. In each dish, 1 cm diameter PDA discs were prepared, each hosting pure growths of either A. rolfsii or Trichoderma spp. Each Trichoderma spp. strain constituted a separate treatment, with six replicates per treatment. Additionally, PDA discs with pure A. rolfsii growths were included as controls. The experiments were replicated three times to ensure the robustness of the results.
2.4.1. Evaluation of the Antagonistic Capacity of Trichoderma spp. vs. A. rolfsii isolate INVEPAR-05
The evaluation involved observing mechanisms of competition, mycoparasitism, and antibiosis, following the scale described by Bell [24] (Table 2). Radial growth of the pathogen (RGP) and antagonists (RGA) was measured using a graduated ruler. The percentage of radial growth inhibition (PIGR) for each Trichoderma spp. isolate was calculated using the formula given by Skidmore and Dickinson [25]:
where R1 represents the radius of A. rolfsii growth used as control, and R2 is the radius of the pathogen in confrontation with the Trichoderma spp. isolate. Growth readings were taken at 24, 48, and 72 hours after initiating the dual cultures.
2.5. Statistical Analysis
The data was analyzed using analysis of variance (ANOVA), followed by Tukey's mean separation (P ≤ 0.01) for post-hoc comparisons. All data were processed by a two way ANOVA in SigmaPlot for Windows v. 14.0 (Systat Software, Inc., San Jose, CA, USA).
3. Results
3.1. Molecular identification of Trichoderma spp and A. rolfsii
Five strains of Trichoderma spp. were isolated from the horticultural zone of Córdoba department. Three isolates were found in Cereté (T02, T04, and T05), one in San Pelayo (T10), and one in Tierralta (T01). No strains of the fungus were detected in Montería. These isolates were identified as follows: Trichoderma harzianum INVEPAR T01, Trichoderma asperellum isolate INVEPAR-T02, Trichoderma asperellum INVEPAR-T4.6, Trichoderma sp. INVEPAR-T05, and Trichoderma viride INVEPAR-T10 (GenBank: OK310695.1, ON237703.1, ON238106.1, ON237737.1, and ON237923.1, respectively). The fungal pathogen affecting the plants within the germplasm bank of segregated Stevia sp. lines was identified as A. rolfsii isolate INVEPAR-05 (GenBank: OK271308.1) Table 3, Figure 2 and Figure 3.
This fungus was also found in commercial stevia plots in the department of Cordoba, along with Athelia rolfsii isolate UCLV-IBP [26].
3.2. In vitro Antagonistic Capacity of Trichoderma spp. Strains against Athelia rolfsii
Figure 4 presents the results for the three evaluations of the percentage of radial growth inhibition (PIGR) of A. rolfsii caused by the five species of Trichoderma isolated from commercial vegetable crops in productive areas of the Cordoba department. The statistical analysis reveals significance (0.01 and 0.05) in all three measurements (PIGR 1, PIGR 2, and PIGR 3), indicating that the PIGR results in all three measurements were influenced by the effect of different Trichoderma spp. strains.
The in vitro antagonistic capacity of Trichoderma spp. against A. rolfsii is a critical aspect to assess their potential as biocontrol agents. The results from the analysis of variance demonstrates that the different Trichoderma sp. strains exerted varying degrees of growth inhibition on A. rolfsii, indicating their potential effectiveness in suppressing the growth of this pathogenic fungus.
The results of the study show the percentage of radial growth inhibition (PIGR) of various Trichoderma spp. strains against the pathogen A. rolfsii. The experiment was conducted at three different time points (PIGR1, PIGR2, and PIGR3) to assess the bio-control potential of each Trichoderma spp. strain.
Among the Trichoderma spp. strains evaluated, Trichoderma harzianum (T01) exhibited the highest inhibitory effect on Athelia rolfsii isolate INVEPAR 05, with PIGR values of 50.67%, 47.33%, and 52.34% at PIGR1, PIGR2, and PIGR3, respectively. This suggests that Trichoderma sp. (T05) has strong biocontrol capabilities against the pathogen, making it a promising candidate for managing A. rolfsii in Stevia sp.
The significance observed in the three measurements of PIGR suggests that the antagonistic effects of Trichoderma spp. were consistently impactful across different time points, reinforcing their biocontrol potential. These findings are promising for the development of sustainable and eco-friendly strategies to manage A. rolfsii infections in Stevia sp.
Trichoderma asperellum (T02) and Trichoderma sp. (T05) also showed considerable in-hibitory activity against A. rolfsii, with PIGR values ranging from 43.95% to 50.53%. These results indicate that both Trichoderma asperellum (T02) and Trichoderma sp. (T05) have the potential to suppress the growth of the pathogen and can be further explored for their biocontrol efficacy.
On the other hand, Trichoderma viride (T10) exhibited moderate inhibitory effects, with PIGR values of approximately 48.63% at all time points. While it displayed some biocontrol activity against A. rolfsii, further investigation is needed to understand the underlying mechanisms and optimize its effectiveness.
Interestingly, Trichoderma asperellum (T04) demonstrated the least inhibitory activity among the tested strains, with PIGR values ranging from 30.91% to 38.56%. While its biocontrol capacity against A. rolfsii was lower compared to other strains, it still showed some level of suppression. Future studies could focus on identifying factors that may enhance its antagonistic potential.
4. Discussion
The findings of this study provide crucial insights into the in vitro antagonistic capacity of Trichoderma spp. strains against the pathogenic fungus A. rolfsii isolate INVEPAR-05, which is prevalent in greenhouses and commercial Stevia fields in Córdoba department. Trichoderma species are well-documented biocontrol agents, extensively studied for their potential in managing diverse phytopathogens [27,28,29,30]. In this investigation, we evaluated five Trichoderma strains for their ability to inhibit the radial growth of the A. rolfsii isolate INVEPAR-05 at a time-course study (PIGR1, PIGR2, and PIGR3).
Our findings demonstrate the significant biocontrol potential of Trichoderma harzianum (T01), Trichoderma asperellum (T02), and Trichoderma sp. (T05) against the A. rolfsii isolate INVEPAR-05, with PIGR values ranging from 43.95% to 52.34%. These results are consistent with previous studies reporting the antagonistic activity of these Trichoderma sp. against different phytopathogens. Notably, T. virens, T. harzianum, T. asperellum, and T. viride species have been identified as the most effective biological control agents, significantly impacting root and stem rot caused by Sclerotium rolfsii in both greenhouse and field conditions [30]. Moreover, Shrestha, et al. [31] suggest T. harzianum and T. asperellum as the most capable species to parasitize S. rolfsii sclerotia. In a study by Raja, et al. [32], the strains T. harzianum Thar23, T. asperellum Tasp49, T. longibrachiatum Tlongi5, and T. citrinoviride Tcitri2 demonstrated high efficacy in controlling S. rolfsii Sacc.
The variation in antagonistic capacity among Trichoderma spp. strains may be attributed to differences in their mechanisms of action and production of secondary metabolites [33,34].
Trichoderma spp have shown antifungal activity against phytopathogenic fungi, employing different groups of secondary metabolites such as terpenes, pyrones, gliotoxin, gliovirin, and peptaibols [35]. This author highlighted the remarkable bioactivity of gliotoxin isolated from Trichoderma virens against Sclerotium rolfsii, with an effective dose 50% of 2.11 g mL-1. Gliotoxin (GT), a fungal secondary metabolite containing sulfur and belonging to the epidithiodioxopiperazine (ETP) class, exhibits a redox cycle between its oxidized and reduced forms, generating reactive oxygen species (ROS) and disrupting intracellular redox homeostasis by interacting with thiol groups of antioxidants such as glutathione [36].
Peptaibols are another essential group of secondary metabolites produced by Trichoderma sp, contributing to the antibiotic activity against various plant pathogens [37]. For instance, in Trichoderma harzianum, three peptaibols named trichorzins HA II, HA V, and HA VI reduced cucumber mosaic virus (CMV) lesions in Vigna sesquipedalis plants by up to 90% [38].
Proteases, produced by T. harzianum, significantly affect the growth of various phytopathogenic fungi, including Botrytis cinerea, Mucor circinelloides, Aspergillus flavus, Aspergillus fumigatus, and Rhizoctonia solani. The aspartic protease P6281 of T. harzianum alters the integrity of B. cinerea cell walls, preventing lesions in fruits like oranges and apples and inhibiting spore formation in B. cinerea, M. circinelloides, A. flavus, and A. fumigatus [39].
Additionally, some secondary metabolites produced by T. harzianum can induce plant defensive mechanisms. For instance, the flavoenzyme ThLAAO (L-amino acid oxidase) of T. harzianum ETS323 exhibits antibacterial activity and, when expressed in tobacco plants, induces the expression of genes related to defense pathways mediated by SA, JA, and ET, as well as the accumulation of ROS, conferring resistance to Sclerotinia sclerotiorum and Botrytis cinerea and resistance to B. cinerea in cabbage plants [40].
Ayyandurai, et al. [33] evaluated the inhibitory capacity of Trichoderma longibrachiatum and Trichoderma asperellum against Sclerotium rolfsii growth. They observed that T. longibrachiatum exhibited the highest inhibition of the pathogen's mycelial growth, reaching 84.44%, followed by T. asperellum with 75.55% inhibition. GC-MS analysis identified the major compounds present in both species. T. longibrachiatum was characterized by significant presence of 2-tricosenoic acid (3.29%), hexadecane (3.12%), and phenol (27.18%), while T. asperellum contained 2,6,10-trimethyltridecane (3.44%) as one of its main compounds.
Prajapati, et al. [41] found that culture filtrates of T. harzianum and T. viride completely inhibited the mycelial growth of S. rolfsii at 4, 6, and 8 days after incubation, while T. asperellum inhibited S. rolfsii with an inhibition percentage of 56.67%, 49.26%, and 47.41% at 4, 6, and 8 days after incubation, respectively. Sab, et al. [42] reported that mycelial growth of S. rolfsii was highly inhibited by the culture filtrate of T. harzianum and T. viride, with a reduction of 70% and 59%, respectively, after 8 days of incubation. Thus, it is speculated that the metabolites in the culture filtrates inhibited the mycelial growth of S. rolfsii.
Trichoderma harzianum, in particular, is known for its ability to produce several antifungal compounds, such as chitinases, glucanases, and proteases, crucial in suppressing fungal growth [43,44]. On the other hand, Trichoderma asperellum has been reported to induce systemic resistance in plants, enhancing their defense mechanisms against pathogenic attacks [45,46].
The observed significance of PIGR at all three time points indicates that Trichoderma spp antagonistic effects were consistently impactful over time, reinforcing their potential as effective biocontrol agents. These results align with the findings of studies by Schirmböck, et al. [47], Sab, et al. [42], Matas-Baca, et al. [48], Vinale, et al. [49], and Prajapati, Patel and Patil [41]highlighting the temporal stability of Trichoderma sp. antagonistic activity.
Moreover, molecular identification of Trichoderma strains using gene sequencing and comparison with the GeneBank database allowed precise species identification. Molecular methods are essential for accurate taxonomic classification of Trichoderma spp., as morphology alone may not distinguish closely related species [34,50,51,52]. However, for initial species confirmation, the internal transcribed spacer (ITS) region of ribosomal DNA is commonly used, but it may not provide sufficient resolution for closely related species due to its high conservation. In the case of Trichoderma sp. INVEPAR-T05, ITS failed to identify the species, suggesting the future use of the more variable TEF1 sequence for closely related species and groups [53,54].
The combination of morphological and molecular approaches enhances the validity and reliability of rapid field selection, as recommended by different authors [55,56,57]. Although this study focuses on the in vitro antagonistic capacity of Trichoderma spp., future research should extend to field trials to evaluate the efficacy of these strains under real agricultural conditions. Recent studies suggest that factors such as soil types, climatic conditions, and population densities can influence the biocontrol potential of Trichoderma spp. strains [58,59,60].
Our results highlight the promising biocontrol potential of Trichoderma harzianum (T01), Trichoderma asperellum (T02), and Trichoderma sp. (T05) against the pathogenic fungus A. rolfsii isolate INVEPAR-05 in stevia cultivation. These findings contribute to the growing knowledge of sustainable and environmentally friendly strategies for plant disease management. Future research on the underlying mechanisms of Trichoderma sp. antagonistic activity and its interaction with A. rolfsii will pave the way for the development of effective and environmentally friendly biofungicides.
5. Conclusions
This study sheds light on the significant in vitro antagonistic capacity of Trichoderma spp. strains against the pathogenic fungus A. rolfsii isolate INVEPAR-05, commonly found in greenhouses and commercial stevia fields in Córdoba department. Trichoderma species have long been recognized as effective biocontrol agents, and our evaluation of five different strains reaffirms their potential in managing diverse phytopathogens.
Among the Trichoderma spp strains tested, Trichoderma harzianum (T01), Trichoderma asperellum (T02), and Trichoderma sp. (T05) exhibited remarkable biocontrol potential against A. rolfsii, with PIGR values indicating significant inhibition of the pathogen's radial growth. These findings are consistent with previous research identifying T. virens, T. harzianum, T. asperellum, and T. viride as effective biological control agents against root and stem rot caused by Sclerotium rolfsii.
The variation in antagonistic capacity among Trichoderma spp. strains can be attributed to differences in their mechanisms of action and production of secondary metabolites. Trichoderma species employ various secondary metabolites, such as gliotoxin, peptaibols, and proteases, to combat phytopathogenic fungi effectively.
Our study underscores the temporal stability of Trichoderma spp. antagonistic activities, making them reliable and consistent biocontrol agents over time. The use of molecular methods, in addition to morphological characterization, enhances an accurate species identification and aids in rapid field selection.
Future research should include field trials to evaluate the efficacy of Trichoderma spp. strains under real agricultural conditions, considering various factors that may influence their biocontrol potential. Continued investigations into the underlying mechanisms of Trichoderma's antagonistic activity and its interaction with A. rolfsii will pave the way for the development of highly effective and eco-friendly biofungicides.
In summary, Trichoderma spp. strains show great promise as biocontrol agents in stevia cultivation, offering sustainable and environmentally friendly alternatives to traditional chemical-based disease management strategies. This study contributes to the advancement of knowledge in agricultural science, with potential implications for enhancing crop productivity and sustainability in the future.
Author Contributions
Conceptualization, LAR-P, JDJ-N, MIO-O and NVR; methodology, LAR-P, MBH, AGR and JDJ-N; formal analysis, LAR-P and MFP.; investigation, LAR-P, MFP, AGR, AMJ-R and YYP-R; writing—original draft preparation, LAR-P, JDJ-N, AJ, KAP and MFP; writing—review and editing, LAR-P, JDJ-N, KAP and N.V.R.; supervision, MIO-O and NVR; funding acquisition, LAR-P, AJ-O, JDJ-N, AJ and EC-C All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by Facultad de Ciencias Agrícolas, Universidad de Córdoba, CO, Colombia, through the program: Production systems and food security. Research line: Agri-food production systems and hydrobiological resources. Grants: FCA-04-19. K.A.P is a current holder of a fellowship from CONACyT Mexico (CVU:227919).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors thank Facultad de Ciencias Agrícolas, Universidad de Córdoba, CO, Colombia, and Instituto de Biotecnología de las Plantas, Universidad Central “Marta Abreu” de Las Villas, Cuba, for assistance with the conditions necessary for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yadav, A.K.; Singh, S.; Dhyani, D.; P.S., A. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can J Plant Sci 2011, 91, 1-27. [CrossRef]
- Soejarto, D.D.; Compadre, C.M.; Kinghorn, D. Ethnobotanical notes in Stevia. Bot Mus Leafl 1983, 29, 1-25.
- Rodriguez-Paez, L.A.; Jimenez-Ramirez, A.M.; Pompelli, M.F.; ineda-Rodriguez, Y.Y.; Jarma-Orozco, A.; Jaraba-Navas, J.D.; Aramendiz-Tatis, H.; Combatt-Caballero, E.; Oloriz-Ortega, M.I.; Rodríguez, N.V. Physiological and enzymatic evaluation of selected genotypes of Stevia rebaudiana Bertoni. Agronomy 2023, 13, 403. [CrossRef]
- Mordue, J.E.M. Descriptions of pathogenic fungi and bacteria. No. 410; CMI: Kew, U.K, 1974.
- Tamayo-Molano, P.J. Tecnologia para el cultivo de estevia. Rionegro, Antioquia: Corporación colombiana de investigación agropecuaria. Agrosavia 2006, Availabe in https://repository.agrosavia.co/handle/20.500.12324/17960. Accessed in Aug 01, 2023.
- Koehler, A.M.; Shew, H.D. Enhanced overwintering survival of Stevia by QoI gungicides used for management of Sclerotium rolfsii. Plant Dis 2017, 101, 1417-1421. [CrossRef]
- Chang, K.F.; Howard, R.J.; Gaudiel, R.G.; Hwang, S.F. First report of Stevia as a host of Sclerotinia sclerotiorum. Plant Dis 1997, 81, 311-311. [CrossRef]
- Hilal, A.A.; Baiuomy, M.A. First record of fungal diseases of stevia (Stevia rebaudiana Bertoni) in Egypt. Egypt J Agric Res 2000, 78, 1435-1448. [CrossRef]
- Kamalakannan, A.; Valluvaparidasan, V.; Chitra, K.; Rajeswari, E.; Salaheddin, K.; Ladhalakshmi, D.; Chandrasekaran, A. First report of root rot of stevia caused by Sclerotium rolfsii in India. Plant Pathol 2007, 56, 350-350. [CrossRef]
- Koehler, A.; Shew, H. First report of stem and root rot of Stevia caused by Sclerotium rolfsii in North Carolina. Plant Dis 2014, 98, 1005. [CrossRef]
- Erper, I.; Ozer, G.; Yildirim, E.; Ozgen, T.; Turkkan, M. First report of southern blight caused by Athelia rolfsii on candyleaf in Turkey. J Plant Pathol 2020, 102, 245-246. [CrossRef]
- Le Bihan, Z.; Gaudin, J.; Robledo-Garcia, F.; Cosson, P.; Hastoy, C.; Rolin, D.; Schurdi-Levraud, V. First report of Sclerotium stem rot caused by Athelia rolfsii on Stevia rebaudiana in southwestern France. Plant Dis 2020, 104, 584. [CrossRef]
- Vélez-Olmedo, J.B.; Vélez-Zambrano, S.; Bonfim, B.A.; Cuenca, E.C.; García, S.; Cedeño, A.G.; Pinho, D.B. First report of Athelia rolfsii (Curzi) causing stem and root rot on stevia (Stevia rebaudiana Bertoni) in Ecuador. J Plant Pathol 2021, 103, 743. [CrossRef]
- Carrieri, R.; Cozzolino, E.; Tarantino, P.; Cerrato, D.; Lahoz, E. First report of southern blight on candyleaf (Stevia rebaudiana) caused by Sclerotium rolfsii in Italy. Plant Dis 2016, 100, 220. [CrossRef]
- Shwetha, G.S.; Hegde, Y.R. Management of Sclerotium wilt of Stevia rebaudiana through biorationals. Int J Plant Prot 2012, 5, 248-251.
- Sanabria-Velazquez, A.D.; Enciso-Maldonado, G.A.; Maidana-Ojeda, M.; Diaz-Najera, J.F.; Ayvar-Serna, S.; Thiessen, L.D.; Shew, H.D. Integrated pathogen management in Stevia using anaerobic soil disinfestation combined with different fungicide programs in USA, Mexico, and Paraguay. Agronomy 2023, 13, 1358. [CrossRef]
- Schludecker, M. Tiempo Montería. Metroblue 2023, Available on https://www.meteoblue.com/es/tiempo/semana/monter%C3%ADa_colombia_3674453. Acessed on Aug 01 2023.
- Martyniuk, S.; Martyniuk, M. Occurrence of Azotobacter spp. in some Polish soils. Pol J Environ Stud 2003, 12, 371-374.
- Tejera, N.; Lluch, C.; Martìnez-Toledo, M.V.; Gonzàlez-López, J. Isolation and characterization of Azotobacter and Azospirillum strains from the sugarcane rhizosphere. Plant Soil 2005, 270, 223-232. [CrossRef]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium species: an illustrated manual for identification; Pennsylvania State University Press: Harrisburg, Pennsylvania, USA, 1983.
- Pineda-Rodriguez, Y.Y.; Pompelli, M.F.; Jarma-Orozco, A.; Rodríguez, N.V.; Rodriguez-Paez, L.A. A new and profitable protocol to DNA extraction in Limnospira maxima. Methods and Protocols 2023, 6, 62. [CrossRef]
- Wilfinger, W.W.; Mackey, K.; Chomczynski, P. Effect of pH and ionic strength on the spectrophotometric assessment of nucleic acid purity. BioTechniques 1997, 22, 474-481. [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; J.W., T. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, Innis, M.A., Gelfand, D.H., Sninsky, J.J., T.J., W., Eds.; Academic Press, Inc.: New York, 1990.
- Bell, D.K. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 1982, 72, 379. [CrossRef]
- Skidmore, A.M.; Dickinson, C.H. Colony interactions and hyphal interference between Septoria nodorum and Phylloplane Fungi. Trans Br Mycol Soc 1976, 66, 57-64. [CrossRef]
- Rodriguez-Paez, L.A.; Pineda-Rodriguez, Y.Y.; Jaraba-Navas, J.D.D.; Jarma-Orozco, A.; Combatt-Caballero, E.M.; Jimenez-Ramirez, A.M.; Oloriz-Ortega, M.I.; Veitia-Rodriguez, N. Athelia rolfsii isolate UCLV-IBP 5.8S ribosomal RNA gene, partial sequence; internal transcribed spacer 2, complete sequence; and large subunit ribosomal RNA gene, Partial Sequence. Available on https://www.ncbi.nlm.nih.gov/nuccore/OM345235. Accessed on August 03 2023. NCBI 2022.
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl Microbiol Biotechnol 2010, 87, 787-799. [CrossRef]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.M.; El-Meihy, R. Phylogenetic diversity of Trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology 2020, 9, 189. [CrossRef]
- Benttoumi, N.; Colagiero, M.; Sellami, S.; Boureghda, H.; Keddad, A.; Ciancio, A. Diversity of nematode microbial antagonists from Algeria shows occurrence of nematotoxic Trichoderma spp. Plants 2020, 9, 941. [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of Its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int J Mol Sci 2022, 23 2329. [CrossRef]
- Shrestha, U.; Dee, M.E.; Piya, S.; Ownley, B.H.; Butler, D.M. Soil inoculation with Trichoderma asperellum, T. harzianum or Streptomyces griseoviridis prior to anaerobic soil disinfestation (ASD) does not increase ASD efficacy against Sclerotium rolfsii germination. Appl Soil Ecol 2020, 147, 103383,. [CrossRef]
- Raja, M.; Sharma, R.K.; Jambhulkar, P.P.; Pandian, R.T.P.; Sharma, P. Comparative evaluation of native Trichoderma species from groundnut rhizosphere against stem rot caused by Sclerotium rolfsii Sacc. . Indian Phytopathol 2023, 76, 459-471. [CrossRef]
- Ayyandurai, M.; Akila, R.; Manonmani, K.; Mini, M.L.; Vellaikumar, S.; Brindhadevi, S.; Theradimani, M. Combined application of Trichoderma longibrachiatum T(SP)-20 and Trichoderma asperellum T(AR)-10 in the management of stem rot of groundnut. Legume Res 2022, 46, 215-221. [CrossRef]
- Guzmán-Guzmán, P.; Kumar, A.; de los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.d.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma species: our best fungal allies in the biocontrol of plant diseases - a review. Plants 2023, 12, 432. [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms 2020, 8, 401. [CrossRef]
- Bulgari, D.; Fiorini, L.; Gianoncelli, A.; Bertuzzi, M.; E., G. Enlightening gliotoxin biological system in agriculturally relevant Trichoderma spp. Front Microbiol 2020, 11, 200. [CrossRef]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother Res 2020, 34, 2835-2842. [CrossRef]
- Kai, K.; Mine, K.; Akiyama, K.; Ohki, S.; Hayashi, H. Anti-plant viral activity of peptaibols, trichorzins HA II, HA V, and HA VI, isolated from Trichoderma harzianum HK-61. J Pest Sci 2018, 43, 283-286. [CrossRef]
- Deng, J.J.; Huang, W.Q.; Li, Z.W.; Lu, D.L.; Zhang, Y.; Luo, X.C. Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme Microb Technol 2018, 112, 35-42. [CrossRef]
- Peng, K.-C.; Lin, C.-H.; Liao, C.-F.; Yu, H.-C.; Lo, C.-T.; Yang, H.-H.; Lin, K.-C. Expression of L-amino acid oxidase of Trichoderma harzianum in tobacco confers resistance to Sclerotinia sclerotiorum and Botrytis cinerea. Plant Sci 2021, 303, 110772. [CrossRef]
- Prajapati, B.K.; Patel, J.K.; Patil, R.K. Bioeficacy of Trichoderma spp. against Sclerotium rolfsii Sacc., an incitant of collar rot of chick pea in vitro. Bioscan 2015, 10, 1745-1748.
- Sab, J.; Nagaraja, A.; Nagamma, G. Efficacy of bio-pesticides against Sclerotium rolfsii Sacc. causing collar rot of chickpea (Cicer arietinum L.). Availabe on https://api.semanticscholar.org/CorpusID:86373077. Accessed August 03, 2023 2014.
- Xiao, Z.; Zhao, Q.; Li, W.; Gao, L.; Liu, G. Strain improvement of Trichoderma harzianum for enhanced biocontrol capacity: Strategies and prospects. Front Microbiol 2023, 14, 1146210. [CrossRef]
- Pasquoto-Stigliani, T.; Guilger-Casagrande, M.; Campos, E.V.R.; Germano-Costa, T.; Bilesky-José, N.; Migliorini, B.B.; Feitosa, L.O.; Sousa, B.T.; de Oliveira, H.C.; Fraceto, L.F.; et al. Titanium biogenic nanoparticles to help the growth of Trichoderma harzianum to be used in biological control. J Nanobiotechnology 2023, 21, 166. [CrossRef]
- Hossain, M.A.; Swarna, F.T.; Arabi, R.A.; Hamim, I. Trichoderma asperellum suppresses viral diseases and promotes the growth and yield of country bean. Front Agron 2023, 5, 1150359. [CrossRef]
- Zhang, Y.; Xiao, J.; Yang, K.; Wang, Y.; Tian, Y.; Liang, Z. Transcriptomic and metabonomic insights into the biocontrol mechanism of Trichoderma asperellum M45a against watermelon Fusarium wilt. PLoS ONE 2022, 17, e0272702. [CrossRef]
- Schirmböck, M.; Lorito, M.; Wang, Y.L.; Hayes, C.K.; Arisan-Atac, I.; Scala, F.; Harman, G.E.; Kubicek, C.P. Parallel formation and synergism of hydrolytic enzymes and peptaibol antibiotics, molecular mechanisms involved in the antagonistic action of Trichoderma harzianum against phytopathogenic fungi. Appl Environ Microbiol 1994, 60, 4364-4370. [CrossRef]
- Matas-Baca, M.A.; García, C.U.; Pérez-Álvarez, S.; Flores-Córdova, M.A.; Escobedo-Bonilla, C.M.; Magallanes-Tapia, M.A.; Chávez, E.S. Morphological and molecular characterization of a new autochthonous Trichoderma sp. isolate and its biocontrol efficacy against Alternaria sp. Saudi J Biol Sci 2022, 29, 2620-2625. [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol Biochem 2008, 40, 1-10. [CrossRef]
- Cao, Z.-J.; Qin, W.-T.; Zhao, J.; Liu, Y.; Wang, S.-X.; Zheng, S.-Y. Three new Trichoderma species in Harzianum clade associated with the contaminated substrates of edible fungi. J Fungi 2022, 8, 1154. [CrossRef]
- Tamizi, A.A.; Mat-Amin, N.; Weaver, J.A.; Olumakaiye, R.T.; Akbar, M.A.; Jin, S.; Bunawan, H.; Alberti, F. Genome sequencing and analysis of Trichoderma (Hypocreaceae) isolates exhibiting antagonistic activity against the papaya dieback pathogen, Erwinia mallotivora. J Fungi 2022, 8, 246. [CrossRef]
- Šašić Zorić, L.; Janjušević, L.; Djisalov, M.; Knežić, T.; Vunduk, J.; Milenković, I.; Gadjanski, I. Molecular approaches for detection of Trichoderma green mold disease in edible mushroom production. Biology 2023, 12, 299. [CrossRef]
- Sarria, G.; Garcia, A.; Mestizo, Y.; Medina, C.; Varón, F.; Mesa, E.; Hernandez, S. Antagonistic interactions between Trichoderma Spp. and Phytophthora palmivora (Butler) from oil palm in Colombia. Eur J Plant Pathol 2021, 161, 751-768. [CrossRef]
- Guerra-Mateo, D.; Gené, J.; Baulin, V.; Cano-Lira, J.F. Phylogeny and taxonomy of the genus Amphichorda (Bionectriaceae): an update on beauveria-like strains and description of a novel species from marine sediments. Diversity 2023, 15, 795. [CrossRef]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: authoritative guidelines on molecular identification of Trichoderma. Fungal Divers 2021, 107, 1-69. [CrossRef]
- Zhang, S.; Xiang, D.; Sun, C.; Han, K.; Li, T.; Zhou, J.; Xu, B. Morphological and molecular identification of peach brown rot disease in Tibet and exploration of the biocontrol efficiency of Trichoderma. J Fungi 2022, 8, 1174. [CrossRef]
- Noman, E.A.; Al-Gheethi, A.A.; Talip, B.A.; Mohamed, R.M.S.R.; Almoheer, R.; Al-Wrafy, F.A.; Al-Shorgani, N.; El Enshasy, H.A. New fungal strains from peat soil in Malaysia: morphological and molecular characteristics. Sustainability 2023, 15, 5902. [CrossRef]
- Zehra, A.; Dubey, M.K.; Meena, M.; Upadhyay, R. Effect of different environmental conditions on growth and sporulation of some Trichoderma species. J Environ Biol 2017, 38, 197-203. [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Front Plant Sci 2017, 8, 102. [CrossRef]
- Rimkus, A.; Namina, A.; Dzierkale, M.T.; Grigs, O.; Senkovs, M.; Larsson, S. Impact of growth conditions on the viability of Trichoderma asperellum during storage. Microorganisms 2023, 11, 1084. [CrossRef]
Figure 1.
Origins of Trichoderma spp. strains isolated from horticultural crops in Córdoba, Colombia.
Figure 1.
Origins of Trichoderma spp. strains isolated from horticultural crops in Córdoba, Colombia.
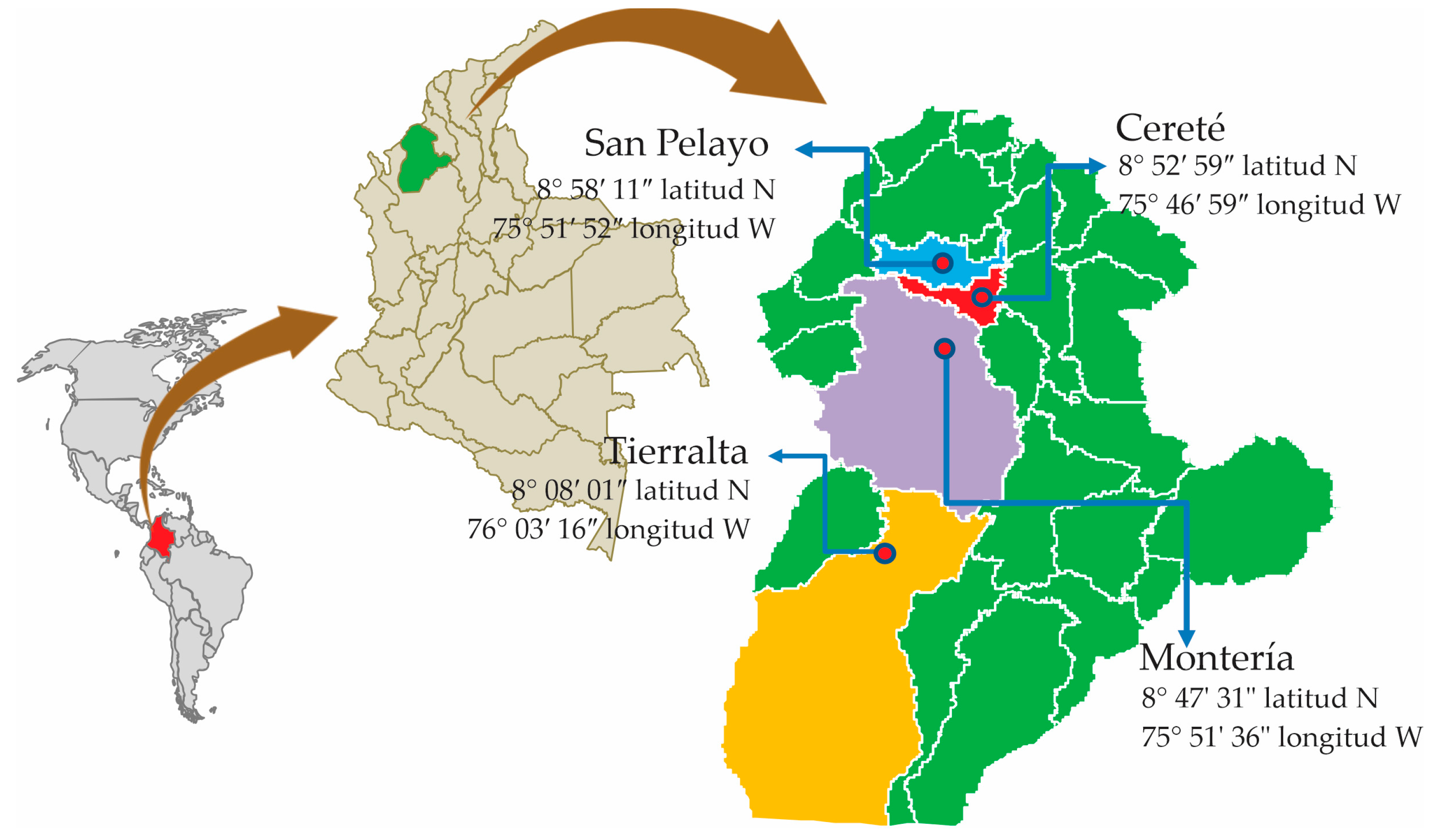
Figure 2.
Results of a 1% agarose gel electrophoresis used to analyze the PCR products of the ITS gene in various isolates. A molecular weight marker (MWM) was employed as a size reference, and the following abbreviations were used for each isolate: T01 for Trichoderma harzianum INVEPAR T01, T02 for Trichoderma asperellum isolate INVEPAR-T02, T04 for Trichoderma asperellum INVEPAR-T4.6, T05 for Trichoderma sp. INVEPAR-T05, Trichoderma viride INVEPAR-T10, and I05 for Athelia rolfsii isolate INVEPAR 05.
Figure 2.
Results of a 1% agarose gel electrophoresis used to analyze the PCR products of the ITS gene in various isolates. A molecular weight marker (MWM) was employed as a size reference, and the following abbreviations were used for each isolate: T01 for Trichoderma harzianum INVEPAR T01, T02 for Trichoderma asperellum isolate INVEPAR-T02, T04 for Trichoderma asperellum INVEPAR-T4.6, T05 for Trichoderma sp. INVEPAR-T05, Trichoderma viride INVEPAR-T10, and I05 for Athelia rolfsii isolate INVEPAR 05.
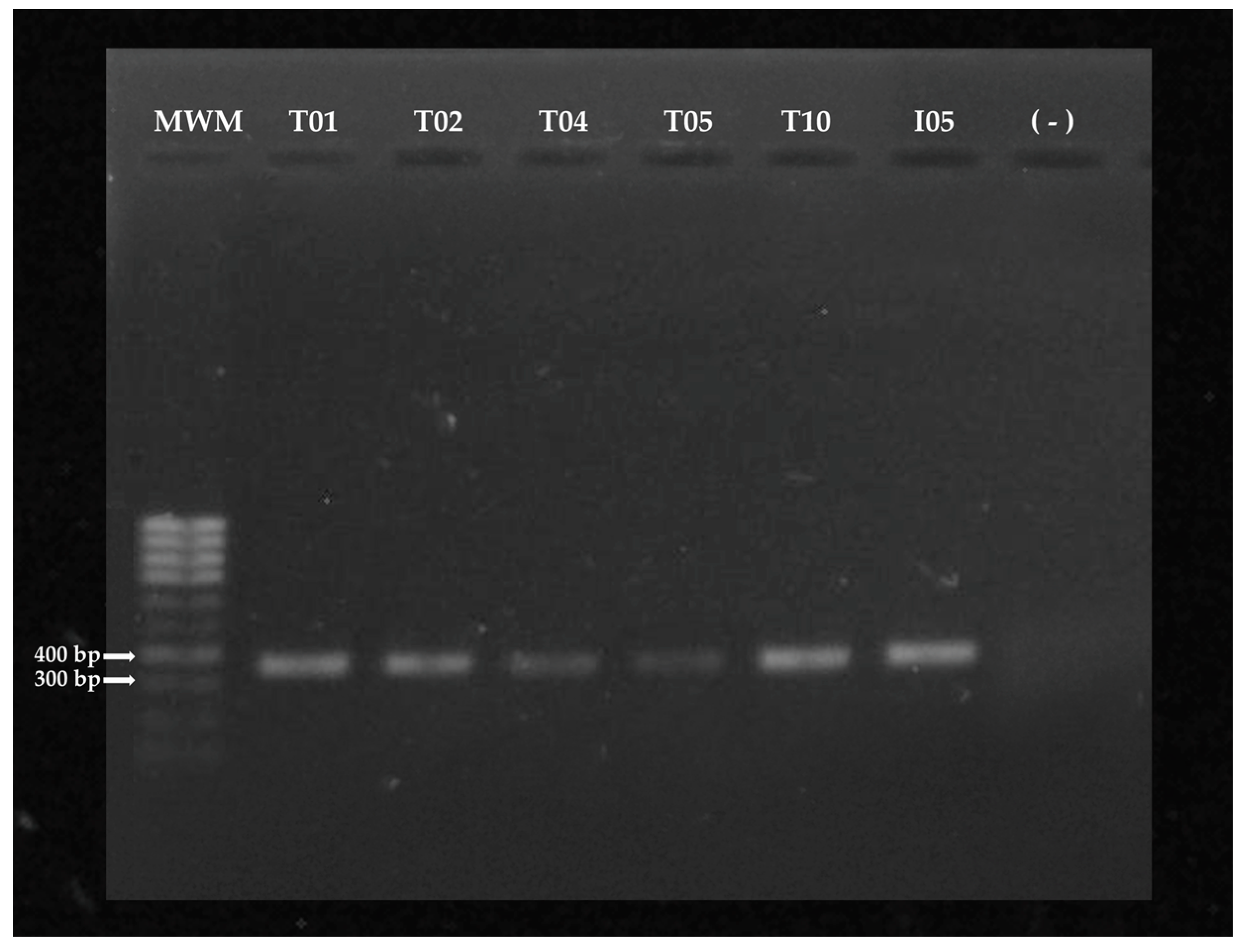
Figure 3.
Neighbor joining phylogenetic tree (max. seq. difference of 0.75) of the isolated Trichoderma strains reported in the GenBank DB. For data visualization MEGA11 Software (https://www.megasoftware.net) for macOS was employed.
Figure 3.
Neighbor joining phylogenetic tree (max. seq. difference of 0.75) of the isolated Trichoderma strains reported in the GenBank DB. For data visualization MEGA11 Software (https://www.megasoftware.net) for macOS was employed.
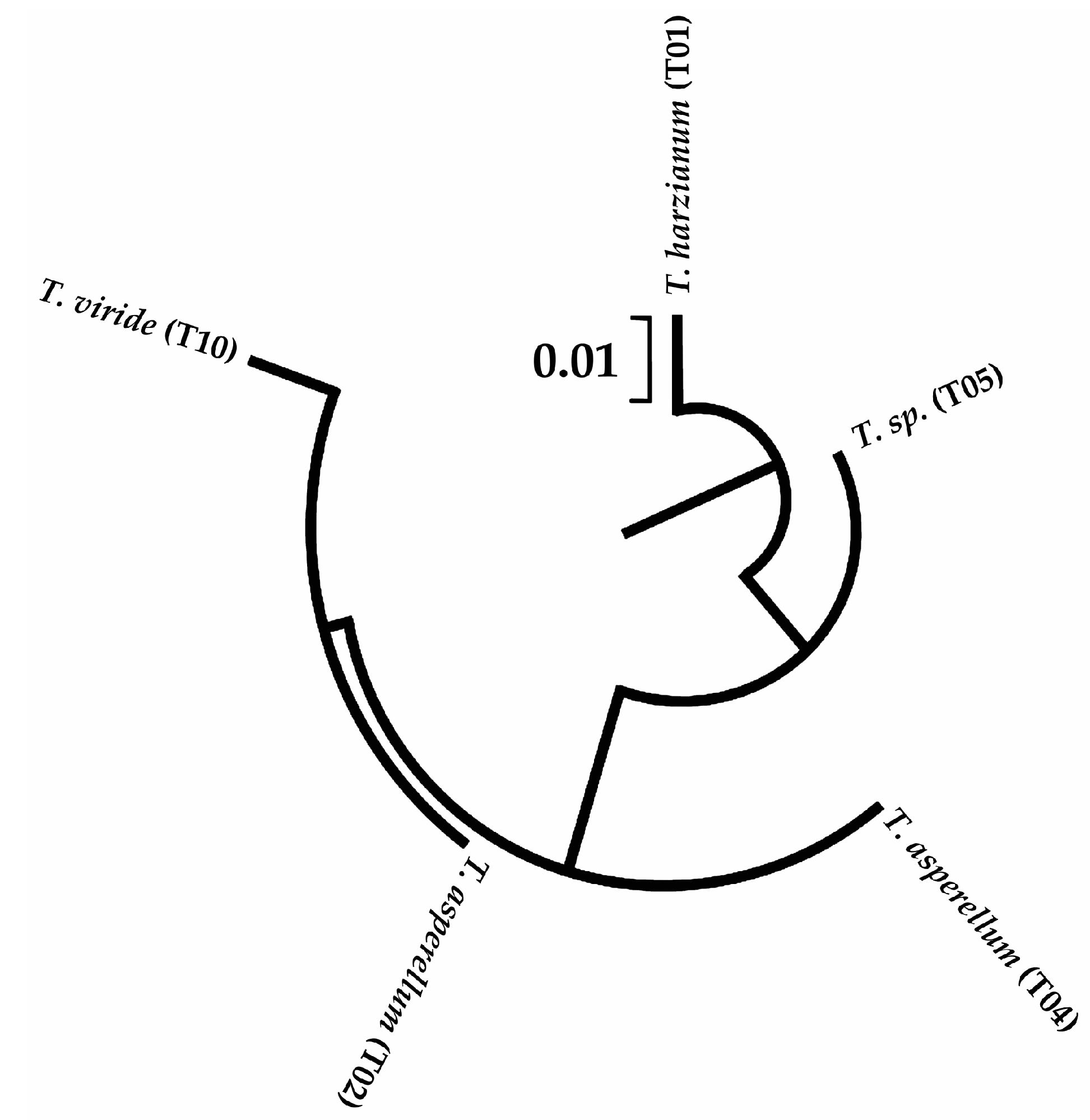
Figure 4.
Analysis of radial growth inhibition percentage averages in Athelia rolfsii isolate Invepar-05. Observations at 24, 48, and 72 hours following dual cultures with Trichoderma spp strains.
Figure 4.
Analysis of radial growth inhibition percentage averages in Athelia rolfsii isolate Invepar-05. Observations at 24, 48, and 72 hours following dual cultures with Trichoderma spp strains.
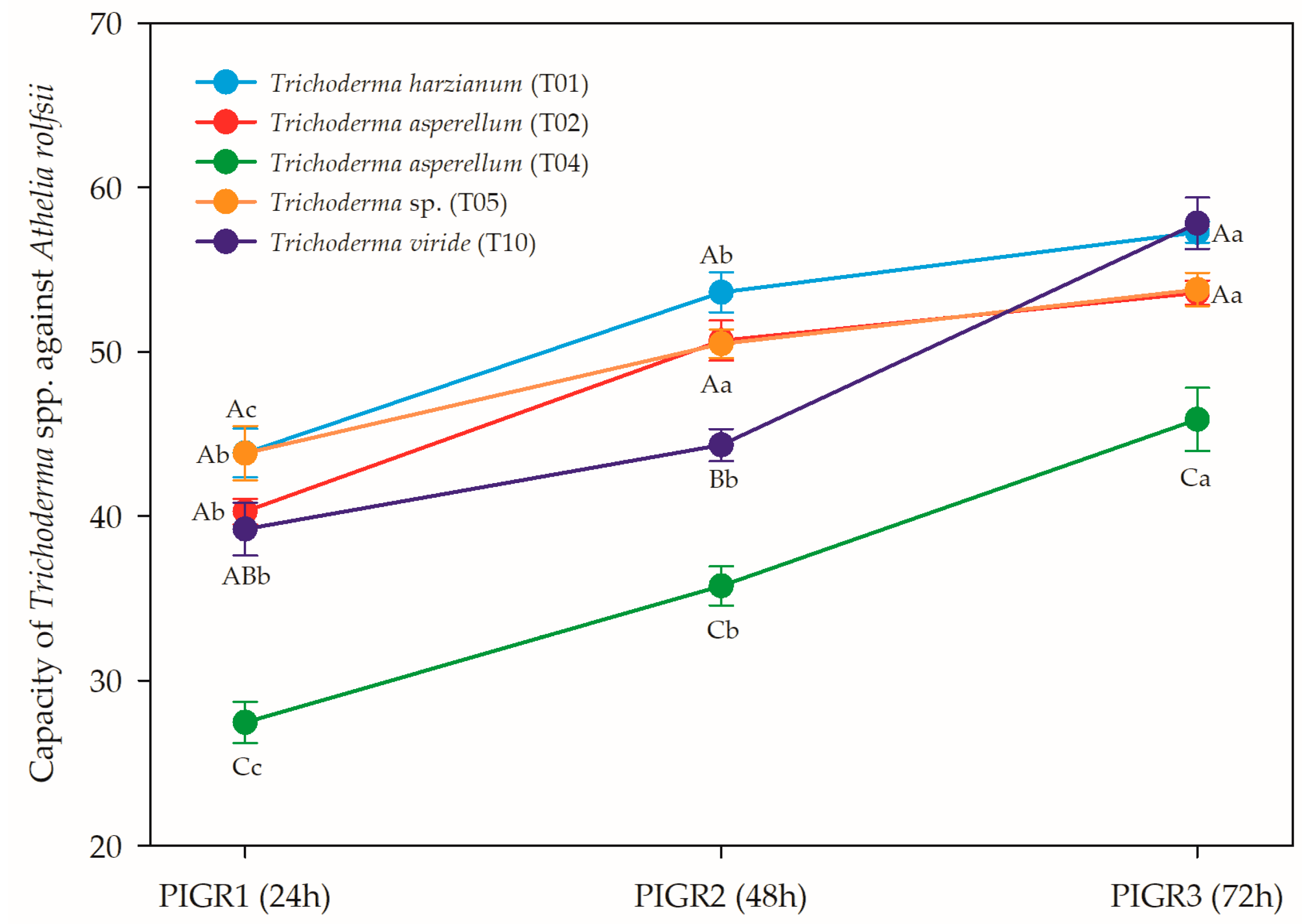

Table 1.
Preparation of the solution (CTAB 2X).
| Solution | Final Concentration | 10 mL |
|---|---|---|
| dH2O (Merck KGaA, part number: 38796) | 5.6 mL | |
| 1 M Tris-HCl- 7.5 (Merck KGaA, part number: T3253) | 100 mM | 1.0 mL |
| 5 M NaCl (Merck KGaA, part number: 1064041000) | 1400 mM | 2.8 mL |
| 0.5 M EDTA-8.0 (Merck KGaA, part number: 324506) | 20 mM | 0.4 mL |
| CTAB (Merck KGaA, part number: H6269-500G) | 2% (w/v) | 0.2 g |
| PVP 10% (w/v) (Merck KGaA, part number: T3253-500G) | 1% (v/v) | 0.1 mL |
Table 2.
Antagonistic capacity of Trichoderma according to the scale of Bell [24].
Table 2.
Antagonistic capacity of Trichoderma according to the scale of Bell [24].
| Degree | antagonistic capacity |
|---|---|
| 0 | No invasion of the antagonist on the colony of A. rolfsii |
| 1 | Invasion of 25% of the colony of the fungus A. rolfsii |
| 2 | Invasion of 50% of the fungal colony A. rolfsii |
| 3 | Invasion of 100% of the colony of the fungus A. rolfsii |
| 4 | Invasion of 100% of the colony of the fungus A. rolfsii, sporulating on it. |
Table 3.
"GeneBank reports of the strains under study.
| Strain | URL GenBank |
|---|---|
| Trichoderma asperellum (T02) | https://www.ncbi.nlm.nih.gov/nuccore/ON237703 |
| Trichoderma harzianum (T01) | https://www.ncbi.nlm.nih.gov/nuccore/OK310695 |
| Trichoderma sp. (T05) | https://www.ncbi.nlm.nih.gov/nuccore/ON237737 |
| Trichoderma viride (T10) | https://www.ncbi.nlm.nih.gov/nuccore/ON237923 |
| Trichoderma asperellum (T04) | https://www.ncbi.nlm.nih.gov/nuccore/ON238106 |
| Athelia rolfsii isolate INVEPAR-05 | https://www.ncbi.nlm.nih.gov/nuccore/OK271308 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



