
Submitted:
14 August 2023
Posted:
15 August 2023
You are already at the latest version
Abstract
The emergence of multidrug-resistant pathogens such as Listeria monocytogenes and Shiga toxin-producing Escherichia coli (STEC) poses public health challenges, and this has led to a continuous search for effective novel antimicrobial agents. This study aimed to isolate marine actinomycetes from South Africa, evaluate their in vitro antimicrobial activity against Listeria and STEC, and characterize their mechanisms of action. Marine actinoymcetes were isolated and identified by 16S rRNA sequencing. Gas chromatography-mass spectrometry (GC-MS) was used to identify the chemical constituents of bioactive actinomycetes secondary metabolites. Antibacterial activity of the secondary metabolites was assessed by the broth microdilution method and their mode of actions were predicted using computational docking. While five strains showed antibacterial activity during primary screening, only Nocardiopsis dassonvillei strain SOD(B)ST2SA2 exhibited activity during secondary screening for antibacterial activity. GC-MS identified five major bioactive compounds: 1-octadecene, diethyl phthalate, pentadecanoic acid, 6-octadecenoic acid, and tri-fluoroacetoxy hexadecane. SOD(B)ST2SA2`s extract demonstrated the minimum inhibitory con-centration and minimum bactericidal concentration ranging from 0.78–25 mg/ml and 3.13 to > 25 mg/ml, respectively. Diethyl phthalate displayed the lowest bacterial protein binding energies (kcal/mol): −7.2, dihydrofolate reductase; −6.0, DNA gyrase B, and −5.8, D-alanine:D-alanine lig-ase. Thus, marine N. dassonvillei SOD(B)ST2SA2 is a potentially good source of antibacterial compounds that can be used to control STEC and Listeria monocytogenes.
Keywords:
Nocardiopsis dassonvillei SOD(B)ST2SA2
; Antibacterial activity
; Molecular docking
; Listeria monocytogenes
1. Introduction
The 21st century is plagued by the emergence of multi-drug resistant bacteria such as foodborne pathogenic strains of Listeria and Shiga toxin-producing Escherichia coli (STEC) [1,2]. The major contributing factors to their antimicrobial resistance include the routine application of antimicrobials in domestic livestock for growth promotion, disease treatment and prophylaxis [3]. These pathogens acquire resistance towards antibiotic effects mainly through gene mutation, acquisition of resistant genes through horizontal gene transfer and biofilm formation [4]. Antimicrobial resistance (AMR) is anticipated to lead to approximately 10 million deaths annually by 2050, and the World Health Organization report estimates that globally 700,000 fatalities are linked to AMR each year [5]. Therefore, the increase and spread of antimicrobial resistance pose a danger to public health. AMR further threatens to push the health sector to a pre-antimicrobial era [6].
Listeria and STEC both cause diarrhoea in humans, which may progress to life-threatening conditions in the vulnerable groups [7]. Infection with Listeria monocytogenes could result in listeriosis, which may progress to meningitis and encephalitis in immune-compromised individuals [8]; and fetal malformations, stillbirths and spontaneous abortions in pregnant women [9]. Worldwide, it is estimated that each year, there are 23 150 cases and 5 463 deaths from listeriosis [10,11]. Moreover, listeriosis has the third highest case mortality rate (up to 30%) among the foodborne diseases [12]. Currently, Listeria contains 27 species [13], but only four of them are commonly isolated from food, namely: Listeria welshimeri, Listeria seeligeri, L. monocytogenes and Listeria ivanovii [14]. Among them, L. monocytogenes is the only species that is a major human pathogen of public health significance [15,16]. Rarely, L. ivanovii, L. innocua and L. seeligeri cause human infections [17]. L. ivanovii principally causes the disease in ruminants [18].
Infection with STEC may deteriorate into haemolytic uremic syndrome (HUS) in 5 to 10% of the cases, especially children and the elderly [19]. The syndrome is characterized by haemolytic anaemia, thrombocytopenia, and nephropathy. Worldwide, annual estimates of STEC induced acute illnesses, HUS cases and deaths are 2801000, 3890 and 230, respectively [20,21]. Based on the flagella (H) and somatic (O) antigens, STEC is divided into over 600 serogroups, and all of them have the ability to produce Shiga toxins. However, due to their low infectious dose (10 to 100 cells), the following serogroups are commonly linked to severe disease in humans: O26, O71, O103, O111, O121, O145 and O157 [1,22,23]. Among them, serotype O157 is the most frequently associated with serious food poisoning [24]. But recently, the incidence of serious disease due to non-O157 serotypes is increasing [22]. While a solution the treatment of listeriosis is antimicrobial chemotherapy [17]; for STEC infections, the use of antibiotics is a debatable matter and generally their use is not recommended. Therefore, the disease is usually managed by symptomatic treatment. Nonetheless antibiotic treatment with inhibitors of protein and cell wall synthesis can be an option when specific criteria regarding duration of disease, serotype, virulence profiles and patient group are satisfied [25].
As multidrug resistance of the implicated bacteria elevates and worsens morbidity and mortality rates, novel antimicrobials are needed to combat this crisis [26]. The actinomycetes, which are Gram-positive bacteria are prolific arsenal of antimicrobials and other bioactive compounds [27]. Over 45% of the total discovered biologically active natural metabolites are from actinomycetes [28]. Furthermore, over 80% of the therapeutically useful antibiotics are from these bacteria, with 50% of the antibiotics from the genus Streptomyces [29,30]. However, the possibility of finding novel metabolites with unique chemical structure from Streptomyces has significantly decreased, predominantly because of genetic exchange among species during evolution [31]. Therefore, bioprospecting of promising rare actinomycetes from less explored environments has been given primary attention recently. These actinomycetes have been described as strains other than commonly known Streptomyces or strains with less frequency of isolation under conventional cultivation techniques [31]. The rare actinomycetes genera include among others Frankia, Micromonospora, Micrococcus, Nocardia, Arthrobacter and Nocardiopsis [32].
Nocardiopsis species are biotechnologically important producers of different bioactive compounds (antimicrobials, anticancer agents, tumor inducers, toxins, immunomodulators, and others) and novel extracellular enzymes [33]. Ecologically, these array of metabolites aid the species to exist even in extreme habitats including marine, hypersaline habitats, desert regions and salterns [34]. Members of this genus are Gram-positive aerobes, catalase-positive and non-acid-fast actinomycetes. They possess nocardioform substrate mycelia and their aerial mycelia contain long chains of spores. They have a high content of guanine and cytosine in their DNA. Significant variations exist in properties of the strains isolated from different environments and their bioactive compounds [34]. In contrast, the terrestrial environments have been explored extensively [28], leaving marine environment underexploited. Thus, marine environment may harbour new actinomycetes with unique bioactive secondary metabolites [35]. This is because the harsh physicochemical conditions in the sea (such as high salinity and pressure and low temperature) tend to favour the microbial production of structurally and functionally unique molecules of industrial and pharmacological importance [36]. To our knowledge, there are no studies reported on the antimicrobial potential of marine actinomycetes fom Sodwana Beach, KwaZulu-Natal (KZN), South Africa against Listeria species and STEC.
Therefore, the aim of this study was to investigate the antimicrobial production of actinomycetes from the selected beaches in KwaZulu-Natal Province, South Africa for their antibacterial activity against L. monocytogenes and STEC isolates from beef. Moreover, the molecular interactions of the metabolites with the target receptor proteins were investigated to ascertain their mode of antibacterial activity.
2. Materials and Methods
2.1. Media formulations
2.1.1. Cross-streak agar
A medium comprising of yeast extract (3 g) (Biolab, Modderfontein, South Africa), peptone (3 g) (Biolab, Wadesville, South Africa), casein (3 g) (Sigma, Steinheim, United States of America), starch (8 g) (Merck Pty Ltd, Modderfontein, South Africa), K2HPO4 (0.5 g) (Minema chemicals, Johannesburg, South Africa), MgSO4.7H2O (0.5 g) (Minema chemicals, Durban, South Africa), agar (15 g) (Neogene, Heywood, United Kingdom), filtered marine water (1000 mL), and pH 7 was used.
2.1.2. Fermentation broth
A broth medium containing yeast extract (3 g), peptone (3 g), casein (3 g), starch (8 g), K2HPO4 (0.5 g), MgSO4.7H2O (0.5 g), glycerol (3 g) (Merck, Pty, Gauteng, South Africa), CaCO3 (0.75 g) (Minema chemicals, Johannesburg, South Africa); in 1000 mL of sterilized marine water, and pH 7 was used.
2.2. Test bacteria
The authors previously isolated and characterized the test bacteria used in this study (Table 1). L. monocytogenes ATCC 15313 and E. coli O157:H7 ATCC 43888 were procured from American Type Culture Collection (ATCC, Manassas, VA, USA) and used as controls.
2.3. Sample collection
In March 2021, marine water was aseptically collected for analysis from six beaches of KwaZulu-Natal Province: Alkantstrand, Blythedale, Salt Rock, Sodwana, Mthunzini, and Tinley Manor (Figure 1) using grab sampling method. In each beach, the water samples were collected every 100 meters from 3 different consecutive sites. In every site, the samples were obtained in triplicates (2 liters each) at a depth of 36 cm. The samples were then placed in cooler boxes and transported to the Microbiology laboratory of the University of Zululand.
2.4. Physicochemical parameters of the seawater
The following physicochemical parameters were measured at all beaches in triplicate in situ: dissolved oxygen (DO), pH, pressure, salinity, specific conductivity, temperature, and total dissolved solids (TDS) using a multi-parameter water meter (Hanna HI 98194, Romania).
2.5. Sample preparation
The water samples were immediately prepared and analysed upon arrival. One half of the samples was treated by heating at the water bath at 50 °C for 1 h [37,38]. The other half was left untreated. Serial dilutions were then conducted up to the 10-1. Subsequently, the water samples were concentrated through 0.45-micron nitrocellulose membrane filters using the EZ-streamTM pump filtration system (Merck Millipore, Molsheim, France) in preparation for culture-based analysis.
2.6. Isolation of actinomycetes
Following the filtration of the water samples, the filtration discs used were placed on three different culture media, namely actinomycetes isolation agar (Condalob, Madrid, Spain), yeast mold agar (Condalob, Madrid, Spain), and marine agar (Mast Group, Merseyside, U.K.), which were then incubated for 31 days at different temperatures based on the physicochemical properties of water (Table 2). Thereafter, the presumed colonies of actinomycetes were selected and sub-cultured on actinomycetes isolation agar. They were then preserved in 30% glycerol at -80 °C for long-term storage.
2.7. The screening for antibacterial compounds production in actinomycetes
The presumptive actinomycetes were screened for antibacterial activity against the test bacteria.
2.7.1. Primary Screening of antibacterial activity
The isolates were screened for antibiotic production by the cross-streak method against the test bacteria. Briefly, the presumptive actinomycetes were streaked as a straight line in the middle of a cross-streak agar medium. The inoculated plates were subsequently incubated for 7 days at different temperatures depending on the physicochemical parameters shown in Table 2. Thereafter, the test bacterial strains at exponential phase were adjusted to 1 × 106 colony-forming units per millilitre (CFU/mL) and streaked perpendicularly to the actinomycetes. The plates were then incubated at 37 °C for 24 h, and observed for the formation of the inhibition zones [39,40].
2.7.2. Secondary screening for antibacterial activity of actinomycete isolates
The active actinomycetes, based on primary screening, were inoculated onto 100 mL of the fermentation broth separately. The inoculated broths were then incubated for 7 days at 160 rpm and at different temperatures based on the physicochemical parameters shown in Table 2. The culture broths were centrifuged for 30 min at 5000 rpm. Thereafter, antimicrobial activity was evaluated by the agar well diffusion method. Briefly, the test bacterial inoculums, at the logarithm growth phase, were adjusted to 1 × 106 CFU/mL. Subsequently, the test bacterial lawns were prepared on Muller Hinton agar plates, followed by the boring of the wells (6 mm diameter). The cell-free supernatant (100 µL) was pipetted into the wells and incubated at 37 °C for 24 h. The agar plates with un-inoculated supernatant served as controls and zones of inhibition were recorded in millilitres [39,41].
2.8. Identification of actinomycetes
The molecular identification of the presumptive isolates that exhibited antibacterial activity during the primary and secondary screening of antibacterial activity was conducted by 16S rDNA gene sequencing. Briefly, genomic DNA was extracted from the cultures using the Quick-DNATM Fungal/Bacterial Miniprep Kit (Zymo Research, Catalogue No. D6005). The 16S target region was amplified using OneTaq(R) Quick-Load(R) 2X Master Mix (NEB, Catalogue No. M0486) with the primers 16S-27F (5’AGAGTTTGATCMTGGCTCAG’3) and 16S-1492R (5’CGGTTACCTTGTTACGACTT’3). The PCR cycling conditions used were: the initial denaturation step for 4 min at 94 °C, followed by denaturation (30 cycles at 94 °C for 30 s), annealing at 55 °C for 40 s and extension at 72 °C for 1 min, and a final extension for 7 min at 72 °C [42]. The PCR products were run on a gel and gel extracted with the ZymocleanTM Gel DNA Recovery Kit (Zymo Research, Catalogue No. D4001). The extracted fragments were sequenced in the forward and reverse direction (Nimagen, BrilliantDyeTM Terminator Cycle Sequencing Kit V3.1, BRD3-100/1000) and purified (Zymo Research, ZR-96 DNA Sequencing Clean-up KitTM, Catalogue No. D4050). The purified fragments were analyzed on the ABI 3500xl Genetic Analyzer (Applied Biosystems, ThermoFisher Scientific) for each reaction for every sample. CLC Bio Main Workbench v7.6 was used to analyze the .ab1 files generated by the ABI 3500XL Genetic Analyzer and results were obtained by BLAST search (NCBI).
2.9. Phylogenetic analysis
A phylogenetic tree of the strain showing antibacterial activity in both the primary and secondary screening, Nocardiopsis dassonvillei strain SOD(B)ST2SA2, and its phylogenetic neighbours was constructed based on 16S rRNA sequences. The tree was constructed using MEGA 11 software. And the evolutionary history of SOD(B)ST2SA2 was inferred using the neighbour-joining (NJ) method. The NJ bootstrapping was performed with 1000 replicates.
2.10. Extraction of the secondary metabolites
N. dassonvillei SOD(B)ST2SA2 was inoculated into the fermentation medium and cultured for up to 7 days at 23 °C in a shaking incubator at 160 rpm. Following the incubation, the broth was centrifuged at 10000 rpm for 15 min, and the supernatant was collected. To the collected supernatant, an equal volume of various solvents was added separately and kept in a rotary shaker (with a shaking speed of 160 rpm) at different times to extract the metabolites: 1 h for methanol, and 24 h for chloroform and ethyl acetate. After shaking, the mixtures were left to stand in separating funnels at a room temperature for up to 24 h to separate the organic phase from the aqueous phase. Thereafter, the organic phase was concentrated by rotary vacuum evaporator [32,39,43]. The concentrated extracts were each weighed and dissolved in 10% tween 80, and tested for their antibacterial activity against the field and ATCC strains of L. monocytogenes and STEC by agar-well diffusion method [44].
2.11. Chemical composition analysis of the N. dassonvillei SOD(B)ST2SA2’s extract
Antibacterial compounds in the bioactive chloroform extract were identified by gas chromatography-mass spectroscope (GC-MS). This was done by injecting 1 µL of sample into an RT x -5 column (30 × 0.32 nm) of the GC-MS model (Perkin Elmer, Clarus 500, USA). A carrier gas used was helium (3 mL/min). Identification of the chemical components of the extract was conducted using Perkin Elmer (Clarus 500, USA) gas chromatography coupled with (Clarus 500, USA) mass spectrometer (MS) [45].
2.12. Antibacterial assay of the chloroform extract
2.12.1. Minimum Inhibitory concentration (MIC) of the extract
To quantitatively estimate the minimum inhibitory concentration (MIC) of the extract, 96 well microdilution method was used [46,47]. Briefly, Mueller-Hinton broth (50 μL) was pipetted into each well. Thereafter, 50 μL of the extract (100 mg/mL in 10% DMSO) was poured into each well in the first row of the microplate and agitated thoroughly. By successive dilutions, 50 μL of the extract mixture per well in row A was transferred into wells located down the column. To ensure the volume in each well remains 50 μL, 50 μL of the mixture was emptied from each well of the last row of the microplate. Fifty microliters of the test bacteria, with a concentration diluted to 1 × 106 CFU/mL, were pipetted into each well. Ciprofloxacin (20 µg/mL) and 10% DMSO were positive and negative controls, respectively. The microplate was covered and placed in an incubator (MODEL) overnight at 37 °C. Afterwards, each well was pipetted with 40 μL of 0.2 mg/mL P-iodonitrotetrazolium violet (INT) dissolved in sterile distilled water. Subsequently, the mixture was incubated for 30 min at 37 °C. Thereafter, MIC was determined as the least concentration of the extract able to inhibit bacterial growth.
2.12.2. Minimum bactericidal concentration (MBC)
The agar dilution method was employed to determine the MBC of the SOD(B)ST2SA2 extract [47]. Briefly, from the wells that lacked bacterial growth after incubation a loop-full of culture medium was taken from each well and streaked on sterile nutrient agar plates and subsequently incubated for 12 h at 37 °C. The least concentration of the extract that completely killed Listeria or STEC was regarded as MBC of the extract.
2.12.3. Mechanism of antibacterial activity of the extract
Interactions of the ligands (pentadecanoic acid, diethyl phthalate, and 1-octadecene) against antibacterial target enzymes, D-alanine:D-alanine ligase (DDI), dihydrofolate reductase, and DNA gyrase B were simulated online by AutoDock Vina. Firstly, the 3D structures of DDI, dihydrofolate reductase, and DNA gyrase B were retrieved from the protein database (PDB ID: 2Zdg, 1DIS, and 1KZN, respectively). The structures were then optimized by deleting heteroatoms, water molecules, and other ligands before adding polar hydrogens by Discovery Studio [48]. Thereafter, chemical structures of the ligands (pentadecanoic acid, diethyl phthalate, and 1-octadecene) were obtained from the PubChem database and their energy was minimised by UCSF chimera [49]. Next, the docking simulations of the ligands and the proteins were executed by AutoDock Vina, and the best-docked conformations were chosen on the basis of their binding scores [50]. Finally, the docked complexes were visualized and analysed using the Discovery Studio visualizer [50].
3. Results
3.1. Physicochemical parameters of the marine water
The physiochemical parameters of water were measured, and the values obtained are shown in Table 2. The temperature ranged from 23–27 °C on the water surface, while the pH of the water was between 6.55 and 7.13. The water pressure ranged from 748.88 to 759.56 mmHg and salinity was 30.81–35.22 PSU. The concentration of the dissolved oxygen (DO) varied between 30.24% and 43.31%. The dissolved solid concentrations ranged from 23.55–26.72 g/L on the beaches.
3.2. Isolation and identification of actinomycetes
In total, 52 presumptive actinomycetes were isolated from the six beaches: the lowest number (1 isolate) was obtained at Alkantstrand beach, whereas the largest number (19 isolates) was found at Salt Rock beach. The majority of the isolates were obtained from the actinomycetes isolation agar and from the heated water samples.
3.3. Screening of antibacterial activity of actinomycetes
Out of the 52 obtained actinomycete isolates, only five strains (BLYST2SA3(i), SOD(A)ST2SA2, SOD(B)ST2SA2, SRST1SA3, and TBST2SA1) demonstrated antibacterial activity against the reference and field strains of STEC (Table 3) and L. monocytogenes (Table 4). SOD(B)ST2SA2 was the only strain that exhibited antibacterial activity during secondary screening (Table 3 and Table 4).
3.4. Molecular identification and phylogenetic analysis
Table 5 shows the five identified marine actinomycete isolates: BLYST2SA3(i), SOD(A)ST2SA2, SOD(B)ST2SA2, SRST1SA3, and TBST2SA1. The strain SOD(B)ST2SA2 that showed bioactivity against STEC and L. monocytogenes on primary and secondary screening, belonged to the genus Nocardiopsis (Table 5). The strain showed a percentage similarity of 99.93%–99.5% with species of Nocardiopsis, following the NCBI’s BLASTn analysis; it had a percentage similarity of 99.93% with Nocardiopsis dassonvillei XY236 (MT393632.1), Nocardiopsis dassonvillei A1W5 (MF321787.1), Nocardiopsis dassonvillei subsp. albirubida 0Act 405 (KC514117.1), Nocardiopsis dassonvillei HR10-5 (JN253591.1), Nocardiopsis dassonvillei HZNU_N_1 (CP022434.1), Nocardiopsis dassonvillei XY236 (MH432693.1) and Nocardiopsis dassonvillei subsp. albirubida 0Act920 (MG661744.1). The taxonomic position of SOD(B)ST2SA2 in relation to the other species in the genus Nocardiopsis is shown by the phylogenetic tree in Figure 2.
3.5. Extraction of the secondary metabolites
Chloroform, methanol, and ethyl acetate were used to extract secondary metabolites. Per 400 ml of the cell-free fermentation supernatant, chloroform extracted more secondary metabolites (3.1 g) compared to methanol and ethyl acetate, which produced 0.81 g and 0.1 g, respectively. The crude extracts had a brownish appearance.
3.6. Chemical composition analysis of the chloroform extract of N. dassonvillei SOD(B)ST2SA2
GC-MS spectrum of the chloroform extract showed 85 compounds (Table S1 and Table 6). The unsaturated fatty acid (6-octadecanoic acid, Z) was the main constituent (20.6%), followed by 1-octadecene (9.22%), trifluoroacetoxy hexadecane (7.7.82%), pentadecanoic acid (5.86%), and diethyl phthalate (3.43%) (Table 6).
3.7. Antibacterial activity of the crude extracts of N. dassonvillei SOD(B)ST2SA2
3.7.1. Agar well diffusion method
The findings of the in vitro antibacterial activity of the extracts from N. dassonvillei SOD(B)ST2SA2 are presented in Table 7 and Table 8. As clearly depicted in Table 7 and Table 8, only the chloroform extract showed inhibition of the growth of bacterial strains. The inhibition zone diameters varied from 0 to 28.67 ± 1.15 mm. The highest inhibition zone diameters, 28.67±1.15 mm (Table 7; Figure 3) and 16.33 ± 1.15 mm (Table 8), were observed against L. monocytogenes ILestanBR363 and the control strains E. coli O157:H7 ATCC 43888, respectively.
3.7.2. MIC, and MBC of the chloroform extract
The antibacterial performance of the chloroform extract was further assessed by determining its MIC and MBC, and the results are presented in Table 7 and Table 8. The MIC of the extract against all the L. monocytogenes strains was 6.25 mg/mL, whereas against STEC it was in a range of 0.78 to 25 mg/mL. The extract had the lowest MIC (0.78 mg/mL) against STEC KRbyGR83, while it possessed the highest MIC value of 25 mg/mL against STEC KEmpBS18 and STEC KEmpBP21 strains (Table 8). The MBC of the extract varied from 25 to > 25 mg/mL against the L. monocytogenes strains (Table 7), while against the STEC isolates, the extract had an MBC ranging from 3.13 to > 25 mg/mL (Table 8).
3.8. Antibacterial activity mechanism
3.8.1. Binding scores of the docked complexes
The molecular structures of the ligands’ interactions with the target bacterial proteins showed docking energy scores ranging from −7.2 to −5.0 kcal/mol (Table 9). The positive control (ciprofloxacin) possessed a relatively lower binding energy scores to the bacterial proteins than the test ligands. Its binding energies ranged from −9.6 to −7.5 kcal/mol. Among the test ligands, diethyl phthalate had lower binding energy scores to the proteins, varying from −7.2 to −5.8 kcal/mol. All the ligands displayed relatively lower binding energy scores to dihydrofolate reductase compared to the other bacterial proteins.
3.8.2. Molecular interactions
In the present study, the amino acid residue THR A:116 of dihydrofolate reductase (DHFR) interacted with pentadecanoic acid through a conventional hydrogen bond (Figure 4). Alkyl, pi-alkyl, and van der Waals bonds were the other bonds involved in the molecular interaction. The interactions of diethyl phthalate with the DHFR included the conventional hydrogen, carbon hydrogen, van der Waals, pi-pi stacked, alkyl, and pi-alkyl bonds (Figure 4). The molecular interactions of the active site of DHFR against 1-octadecene did not include any hydrogen bonds. However, other bonds participated, namely van der Waals, alkyl, pi-sigma, and pi-alkyl bonds (Figure 4). The control, ciprofloxacin, interacted with the enzyme DHFR through the following bonds: van der Waals, conventional hydrogen, carbon hydrogen, unfavourable acceptor-acceptor, pi-sigma, pi-pi stacked, alkyl, and pi-alkyl interactions (Figure S1).
Pentadecanoic acid displayed hydrogen bonds with the amino acid residues LYS A:153 and LYS A:116 of D-alanine:D-alanine ligase (DDl) (Figure S2). Van der Waals, pi-sigma, alkyl, and pi-alkyl bonds also participated in the interactions. LYS A:153 formed a hydrogen bond with a carbonyl group of diethyl phthalate (Figure S2). Van der Waals and pi-pi stacked bonds were also formed between diethyl phthalate and DDl (Figure S2). The interaction of DDI with 1-octadecene did not involve any hydrogen bonds, however, van der Waals, pi-sigma, alkyl, and pi-alkyl bonds participated (Figure S2). GLU A:189, LYS A:153, and LEU A:192 bonded ciprofloxacin to the enzyme DDl using the conventional hydrogen bonds, alkyl, pi-alkyl, pi-pi stacked, and van der Waals bonds (Figure S3).
There was a hydrogen bond between the carbonyl group of pentadecanoic acid and VAL A:167 of DNA gyrase B (Figure S4). Additionally, the ligand showed alkyl and van der Waals bond interactions with the protein. The ester oxygen of diethyl phthalate formed a hydrogen bond with ASN A:46 of DNA gyrase B (Figure S4). The other bonds formed were pi-anion, alkyl, pi-alkyl, and van der Waals bonds. Only van der Waals and alkyl bonds participated in the interaction interaction of 1-octadecene with the enzyme (Figure S4). The carbonyl group of ciprofloxacin formed hydrogen bonds with ARG A:76 and ARG A:136 of the enzyme (Figure S5). Moreover, van der Waals, halogen (fluorine), pi-anion, alkyl, and pi-alkyl interactions were also observed.
4. Discussion
The marine environments of KwaZulu-Natal Province, South Africa, are considered relatively underexploited, presenting an opportunity for the discovery of novel antibiotic-producing actinobacteria [61,62]. In the province, there is a lack of studies investigating the antimicrobial potential of marine actinomycetes against Listeria species and multi-drug resistant STEC. To address this research gap, the study focused on the isolation and identification of bioactive actinobacteria from the marine environments of KwaZulu-Natal Province.
The diversity and distribution of marine microorganisms are significantly influenced by the physicochemical parameters of seawater [63]. These parameters include temperature, salinity, pH, TDS, DO levels, specific conductivity and the presence of other chemical compounds. In the present study, the temperature of samples indicated a high probability of the microbial inhabitants of the studied sites being mesophiles. Our findings are similar to those of a previous study by Meena et al. [64] in the marine sediments from Port Blair Bay, India. The pH of the seawater samples was neutral, implying that the microbial inhabitants may be neutrophiles. In contrast, the sampling site (Mangrove forest) was found to be slightly basic (pH 7.68) [65]. In the current study, salinity was high (30.81–35.22 PSU) supporting the survival of halophilic microorganisms and archaea [66]. Comparable salinity ranges (32.3–34.4 PSU) were documented by Muduli et al. [67] from Port Blair Bay, Andaman and Nicobar Islands, India. The DO observed in this study implied the bacteria inhabiting the beaches to be aerobes and/or aero-tolerant. The concentration of TDS in our sampled sites was higher than the reported typical seawater concentration (3–4 g/l) [68]. The water pressure in some of our sampled sites was higher than the atmospheric pressure (750 mmHg), suggesting that most of the microbes occupying those environments to be barophiles [69]. Overall, there were statistically significant variations (p < 0.05) in water pressure and temperature among some of the beaches. This may cause differences in the microbial community structures of the respective beaches [70]. Furthermore, the physicochemical parameters measured in this study helped in the isolation and cultivation of the actinobacteria in culture conditions that were closest to their natural habitats.
In our present study, only N. dassonvillei strain SOD(B)ST2SA2 showed antibacterial potential against the test bacteria during the secondary screening of antibacterial activity. This suggests that this actinobacterial strain produces bioactive compounds that have antibacterial properties, which may be different from those produced by the other strains tested in the study. Similarly, in a different study by Salaria and Furhan [71], out of the 46 presumptive actinomycetes screened for antibacterial activity, only isolate A41 showed promising antibacterial effects during the secondary screening. The isolation of actinobacterial strains closely related to N. dassonvillei is significant because Nocardiopsis is considered a rare genus of actinomycetes. Nocardiopsis strains are ecologically versatile and biotechnologically important [34]. These bacteria are known to produce a wide range of bioactive compounds, including antimicrobial agents, anticancer substances, tumour inducers, toxins, and immunomodulators [34]. Additionally, they secrete various novel extracellular enzymes such as amylases, chitinases, cellulases, inulinases, xylanases, and proteases. The production of these bioactive compounds and enzymes holds potential for biotechnological applications in various fields [34].
The phylogenetic tree indicated that our isolate N. dassonvillei SOD(B)ST2SA2 formed a distinct lineage within the Nocardiopsis species. Similarly, Tang et al. [72] found their actinomycete isolate (Haloactinospora alba strain YIM 90648T [DQ923130]) to form a branch that was separate from that containing species of Streptomonospora, Nocardiopsis, and Thermobifida.
The organic solvent chloroform is relatively less frequently used to obtain anti-infective crude extracts [73]. However, it produced a higher yield of the crude extract than the common organic solvent(s) used both in this study and in another study by Siddharth and Rai [32]. Probably the nature of the target compounds in our study favours being extracted by chloroform. Additionally, in this study, the bioassay results established that the antibacterial agents were in the chloroform extract. In contrast, Okudoh [74] reported ethyl acetate fraction to possess antibacterial compounds in their study.
The antimicrobial potential of the chloroform crude extract was further affirmed by the presence of bioactive compounds with a wide array of reported antimicrobial and anti-cancer activities as revealed by the GC-MS analysis (Table 6). To our knowledge, the present study identifies trifluoroacetoxy hexadecane for the first time in the culture extracts of Nocardiopsis spp. and of actinomycetes. However, its isomer (4-trifluoroacetoxy hexadecane) is a natural product found in Streptomyces sparsus [75].
Crude extracts with MIC values up to 8 mg/mL are regarded as having at least some degree of inhibitory activity [76,77]. Therefore, in the present study the chloroform extract, with MIC values of 6.25 mg/mL against the L. moncytogenes strains, possesses some degree of antibacterial activity against the strains. In contrast, a strong antibacterial activity (MIC = 62.5 µg/mL) of a 4-bromophenol derived from Nocardipsis sp. SCA21 was reported by Siddharth and Rai [32] against L. monocytogenes ATCC 13932 in a previous study.
According to Aligiannis et al. [78], moderate microbial inhibitors are those crude extracts with MIC values ranging between 0.60 mg/mL and 1.50 mg/mL. Therefore, the chloroform extract in our study had moderate antimicrobial activity against some of the multi-drug resistant STEC strains. In a previous study, a crude extract from Pseudomonas aeruginosa exhibited a stronger antibacterial activity (MIC = 0.391 mg/mL) against an E. coli strain (ATCC 25925) [79].
AutoDock Vina software was used to simulate the interaction of the GC-MS-identified compounds (ligands) and ciprofloxacin against the antibacterial target enzymes (DDl, DNA gyrase B, and DHFR), which are common among bacterial strains. DDl is a bacterial enzyme involved in cell-wall biosynthesis. It catalyzes the formation of UDP-N-acetylmuramoyl pentapeptide, the peptidoglycan precursor [80]. DNA gyrase is a type II topoisomerase that is essential for bacterial DNA replication and transcription [81]. DHFR catalyzes the conversion of dihydrofolate to tetrahydrofolate (THF), which is required for the activity of folate-dependent enzymes and, as a result, is required for DNA synthesis and methylation [82]. These enzymes are therefore important targets for antibacterial drugs.
The affinity of biomolecular interactions and the efficacy of medications are frequently determined using binding free energy. It is defined as the free energy difference between the bound and completely unbound states [83]. Therefore, the lower the value, the more stable the complex formed between the ligand and target protein [84,85]. The substantially lower protein binding energy scores of diethyl phthalate in our study compared to the other test ligands indicated that it binds to the target proteins more stably. Generally, all the ligands showed potential to interact with the target bacterial proteins and induce antibacterial effects [85,86].
Ligand effectiveness to exert antibacterial action highly dependent on the type of interactions produced with receptors. Intermolecular interactions help to stabilize a ligand/receptor complex, consequently resulting in growth inhibition. Thus, the observed interactions in this study were perceived to have resulted in the inhibition and killing of the tested bacteria [76,85]. The findings were confirmed by Amer et al. [87], who concluded that the highest antimicrobial activity observed in one of their sulfadimidine analogues was likely due to the strong interaction with the binding site of DNA gyrase through the formation of H-bonds with important amino acids (Asp615 and two with Val634). Alkyl, pi-sigma as well as the halogen (fluorine) bonds observed in this study are categorised as covalent bonds. Ligands forming covalent bonds tend to bind permanently to their target sites [85]. Thus, it was concluded that the strong covalent bonds among the ligand-protein complexes resulted in the observed bacterial growth inhibition and/or death.
5. Conclusions
The present study revealed Sodwana Beach as a potential source of bioactive and rare actinomycetes. N. dassonvillei strain SOD(B)ST2SA2 was isolated for the first time in the marine waters of KwaZulu-Natal Province. Important bioactive compounds were identified in its crude extract. In addition, the compound trifluoroacetoxy hexadecane was identified for the first time in the extracts of Nocardiopsis spp. and actinomycetes. The bacterial strain showed some degree of antibacterial activity against beef isolates (L. monocytogenes strains and multi-drug-resistant strains of Shiga-toxigenic E. coli). The molecular docking results suggested the antimicrobial compounds within the crude extract to inhibit the bacterial growth through interacting with proteins such as DHFR, DDl and DNA gyrase B. Future studies should include further characterization of N. dassonvillei SOD(B)ST2SA2 and the purification and characterization of its bioactive compounds.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.M. and T.S.M.; methodology, S.S.N., S.H.K., E.M. and T.S.M.; software, E.M. and M.C.O.; validation, E.M. and T.S.M.; formal analysis, S.S.N., E.M. and T.S.M.; investigation, S.S.N. and S.H.K.; resources, E.M.; data curation, E.M.; writing—original draft preparation, S.S.N.; writing—review and editing, E.M., A.K.B. and T.S.M.; visualization, E.M.; supervision, E.M. and A.K.B.; project administration, E.M.; funding acquisition, E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Red Meat Research and Development South Africa (RMRD SA) and the Department of Trade, Industry, and Competition under grant number THRIP/22/30/11/2017. Mr. S.S. Ngema is the recipient of The Moses Kotane Institute (MKI) Research Fund (academic year 2022) and The FoodBev SETA Bursary (academic year 2023).
Institutional Review Board Statement
This study was approved by the University of Zululand’s Research Ethics Committee (UZREC) (Certification number UZREC 171110-030 PGD 2021/26) and date of approval 29 March 2021); and is part of an approved departmental project with ethics approval UZREC 171110-030.
Acknowledgments
The authors would like to thank Inqaba Biotechnical Company for its assistance in performing this research, as well as the University of KwaZulu-Natal and the University of Zululand for providing facilities for this project. The authors are grateful to Miss Nqobile Precious Zondo for her assistance in carrying out this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A. Virulence characteristics and antimicrobial resistance profiles of Shiga toxin-producing Escherichia coli isolates from humans in South Africa: 2006–2013. Toxins 2019, 11, 424. [Google Scholar] [CrossRef] [PubMed]
- Uludag, A.A.; Aydogdu, E.Ö.A.; Kimiran, A. The determination of presence of Listeria monocytogenes in ground meat sold in Istanbul. GU J Sci 2023, 36, 53–66. [Google Scholar] [CrossRef]
- Moreno, L.Z.; Paixão, R.; Gobbi, D.D.; Raimundo, D.C.; Ferreira, T.P.; Moreno, A.M.; Hofer, E.; Reis, C.M.; Matté, G.R.; Matté, M.H. Characterization of antibiotic resistance in Listeria spp. isolated from slaughterhouse environments, pork and human infections. J. Infect. Dev. Ctries. 2014, 8, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Courvalin, P. Antibiotic resistance in Listeria spp. Antimicrob. Agents Chemother. 1999, 43, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance. (accessed on 10 July 2023).
- Hamed, A.A.; Mohamed, O.G.; Aboutabl, E.A.; Fathy, F.I.; Fawzy, G.A.; El-Shiekh, R.A.; Al-Karmalawy, A.A.; Al-Taweel, A.M.; Tripathi, A.; Elsayed, T.R. Identification of antimicrobial metabolites from the Egyptian soil-derived Amycolatopsis keratiniphila revealed by untargeted metabolomics and molecular docking. Metabolites 2023, 13, 620. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.I.; Vázquez, S.; Cornejo, C.; D’Alessandro, B.; Braga, V.; Caetano, A.; Betancor, L.; Varela, G. Does Shiga toxin-producing Escherichia coli and Listeria monocytogenes contribute significantly to the burden of antimicrobial resistance in Uruguay? Front. Vet. Sci. 2020, 7, 583930. [Google Scholar] [CrossRef] [PubMed]
- Gómez, D.; Azón, E.; Marco, N.; Carramiñana, J.J.; Rota, C.; Ariño, A.; Yangüela, J. Antimicrobial resistance of Listeria monocytogenes and Listeria innocua from meat products and meat-processing environment. Food Microbiol. 2014, 42, 61–65. [Google Scholar] [CrossRef]
- Matle, I.; Mbatha, K.R.; Lentsoane, O.; Magwedere, K.; Morey, L.; Madoroba, E. Occurrence, serotypes, and characteristics of Listeria monocytogenes in meat and meat products in South Africa between 2014 and 2016. J. Food Saf. 2019, 39, e12629. [Google Scholar] [CrossRef]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: a systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef]
- Dufailu, O.A.; Yaqub, M.O.; Owusu-Kwarteng, J.; Addy, F. Prevalence and characteristics of Listeria species from selected African countries. Trop. Dis. Travel Med. Vaccines 2021, 7, 1–9. [Google Scholar] [CrossRef]
- Al-Brefkani, A.M.T. Study the antimicrobial resistance genes of Listeria monocytogenes isolated from industrial and clinical samples in Iraq. Iraqi J. Sci. 2023, 594–604. [Google Scholar] [CrossRef]
- Vishnuraj, M.; Ajay, G.; Aravind Kumar, N.; Renuka, J.; Pollumahanti, N.; Anusha Chauhan, H.; Vaithiyanathan, S.; Rawool, D.B.; Barbuddhe, S. Duplex real-time PCR assay with high-resolution melt analysis for the detection and quantification of Listeria species and Listeria monocytogenes in meat products. J. Food Sci. Technol. 2023, 60, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.; Duffy, G.; Sheridan, J.; Blair, I.; McDowell, D. Antibiotic resistance among Listeria, including Listeria monocytogenes, in retail foods. J. Appl. Microbiol. 2001, 90, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, L.S.; Gunathilaka, G.U.; Zhang, Y. Antimicrobial-resistant Listeria species from retail meat in metro Detroit. J. Food Prot. 2012, 75, 2136–2141. [Google Scholar] [CrossRef] [PubMed]
- Matle, I.; Mbatha, K.R.; Madoroba, E. A review of Listeria monocytogenes from meat and meat products: epidemiology, virulence factors, antimicrobial resistance and diagnosis. Onderstepoort J. Vet. Res. 2020, 87, 1–20. [Google Scholar] [CrossRef]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial resistance in Listeria species. Microbiol. Spectr. 2018, 6, 237–259. [Google Scholar] [CrossRef] [PubMed]
- Kayode, A.J.; Igbinosa, E.O.; Okoh, A.I. Overview of listeriosis in the Southern African Hemisphere. J. Food Saf. 2020, 40, e12732. [Google Scholar] [CrossRef]
- Iweriebor, B.C.; Iwu, C.J.; Obi, L.C.; Nwodo, U.U.; Okoh, A.I. Multiple antibiotic resistances among Shiga toxin-producing Escherichia coli O157 in feces of dairy cattle farms in Eastern Cape of South Africa. BMC Microbiol. 2015, 15, 1–9. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J.M.; Stapleton, J.; Angulo, F.J.; Yeung, D.H.; Kirk, M.D. Global incidence of human Shiga toxin–producing Escherichia coli infections and deaths: a systematic review and knowledge synthesis. Foodborne Pathog. Dis 2014, 11, 447–455. [Google Scholar] [CrossRef]
- Onyeka, L.O.; Adesiyun, A.A.; Keddy, K.H.; Manqele, A.; Madoroba, E.; Thompson, P.N. Prevalence and patterns of fecal shedding of Shiga toxin–producing Escherichia coli by cattle at a commercial feedlot in South Africa. J. Food Saf. 2022, 42, e12961. [Google Scholar] [CrossRef]
- Varcasia, B.M.; Tomassetti, F.; De Santis, L.; Di Giamberardino, F.; Lovari, S.; Bilei, S.; De Santis, P. Presence of Shiga toxin-producing Escherichia coli (STEC) in fresh beef marketed in 13 regions of Italy (2017). Microorganisms 2018, 6, 126. [Google Scholar] [CrossRef] [PubMed]
- Onyeka, L.O.; Adesiyun, A.A.; Keddy, K.H.; Madoroba, E.; Manqele, A.; Thompson, P.N. Shiga toxin–producing Escherichia coli contamination of raw beef and beef-based ready-to-eat products at retail outlets in Pretoria, South Africa. J. Food Prot. 2020, 83, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Berhilevych, O.; Kasianchuk, V.; Deriabin, O.; Kukhtyn, M. Isolation of Shiga toxin-producing strains of Escherichia coli from beef and swine carcasses and the characterization of their genes. Regul. Mech. Biosyst. 2018, 9, 275–280. [Google Scholar] [CrossRef]
- Agger, M.; Scheutz, F.; Villumsen, S.; Mølbak, K.; Petersen, A.M. Antibiotic treatment of verocytotoxin-producing Escherichia coli (VTEC) infection: a systematic review and a proposal. J. Antimicrob. Chemother. 2015, 70, 2440–2446. [Google Scholar] [CrossRef] [PubMed]
- de Kraker, M.E.; Lipsitch, M. Burden of antimicrobial resistance: compared to what? Epidemiol. Rev. 2021, 43, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, H.; Danesh, A.; Mohseni, M.; Khorasgani, M.R. Marine actinomycetes with probiotic potential and bioactivity against multidrug-resistant bacteria. Int. J. Mol. Cell. Med. 2018, 7, 44–52. [Google Scholar] [PubMed]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary metabolites and biodiversity of actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Jebakumar, S.R.D. Non-streptomycete actinomycetes nourish the current microbial antibiotic drug discovery. Front. Microbiol. 2013, 240. [Google Scholar] [CrossRef]
- Jagannathan, S.V.; Manemann, E.M.; Rowe, S.E.; Callender, M.C.; Soto, W. Marine actinomycetes, new sources of biotechnological products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef]
- Dhakal, D.; Pokhrel, A.R.; Shrestha, B.; Sohng, J.K. Marine rare actinobacteria: isolation, characterization, and strategies for harnessing bioactive compounds. Front. Microbiol. 2017, 8, 1106. [Google Scholar] [CrossRef]
- Siddharth, S.; Rai, V.R. Isolation and characterization of bioactive compounds with antibacterial, antioxidant and enzyme inhibitory activities from marine-derived rare actinobacteria, Nocardiopsis sp. SCA21. Microb. Pathog. 2019, 137, 103775. [Google Scholar] [CrossRef] [PubMed]
- Passari, A.K.; Mishra, V.K.; Singh, G.; Singh, P.; Kumar, B.; Gupta, V.K.; Sarma, R.K.; Saikia, R.; Donovan, A.O.; Singh, B.P. Insights into the functionality of endophytic actinobacteria with a focus on their biosynthetic potential and secondary metabolites production. Sci. Rep. 2017, 7, 11809. [Google Scholar] [CrossRef] [PubMed]
- Bennur, T.; Kumar, A.; Zinjarde, S.; Javdekar, V. Nocardiopsis species as potential sources of diverse and novel extracellular enzymes. Appl. Microbiol. Biotechnol. 2014, 98, 9173–9185. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine bacterial secondary metabolites: a treasure house for structurally unique and effective antimicrobial compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, M.; Colwell, R.R.; Hill, R.T. Isolation and diversity of actinomycetes in the Chesapeake Bay. Appl. Environ. Microbiol. 1993, 59, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Subramani, R.; Sipkema, D. Marine rare actinomycetes: a promising source of structurally diverse and unique novel natural products. Mar. Drugs 2019, 17, 249. [Google Scholar] [CrossRef]
- Valli, S.; Suvathi, S.S.; Aysha, O.S.; Nirmala, P.; Vinoth, K.P.; Reena, A. Antimicrobial potential of actinomycetes species isolated from marine environment. Asian Pac. J. Trop. Biomed. 2012, 2, 469–73. [Google Scholar] [CrossRef]
- Dudeja, S.; Chhokar, V.; Badgujjar, H.; Chauhan, R.; Soni, S.; Beniwal, V.; Kumar, A. Isolation and screening of antibiotic producing fungi from solid-state waste. Polymorphism 2020, 4, 59–71. [Google Scholar]
- Pandya, D.D.; Ram, D. Screening and characteristic study of antimicrobial actinomycetes from near-by soil of medicinal plants. Int. J. Pharm. Pharm. Sci. 2018, 10, 66–73. [Google Scholar]
- Gharechahi, J.; Zahiri, H.S.; Noghabi, K.A.; Salekdeh, G.H. In-depth diversity analysis of the bacterial community resident in the camel rumen. Syst. Appl. Microbiol. 2015, 38, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, S.; Jayashree, M.; Shivani, R.; Anwesha, S.; Rao, K.B. Characterisation and identification of antibacterial compound from marine actinobacteria: in vitro and in silico analysis. J. Infect. Public Health 2019, 12, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Shubha, M.; Ushashi, B.; Veena, S.; Bhaskara, R. Antimicrobial activity of Streptomyces variabilis strain-VITUMVB03 isolated from Kanyakumari marine sediments. Asian J. Pharm. Clin. Res. 2017, 10, 112–116. [Google Scholar]
- Ara, I.; Bukhari, N.A.; Aref, N.; Shinwari, M.M.; Bakir, M. Antiviral activities of streptomycetes against tobacco mosaic virus (TMV) in Datura plant: evaluation of different organic compounds in their metabolites. Afr. J. Biotechnol. 2012, 11, 2130–2138. [Google Scholar]
- Eloff, J.N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta Med. 1998, 64, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Kamazeri, T.S.A.T.; Abd Samah, O.; Taher, M.; Susanti, D.; Qaralleh, H. Antimicrobial activity and essential oils of Curcuma aeruginosa, Curcuma mangga, and Zingiber cassumunar from Malaysia. Asian Pac. J. Trop. Med. 2012, 5, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Adejoro, I.A.; Babatunde, D.D.; Tolufashe, G.F. Molecular docking and dynamic simulations of some medicinal plants compounds against SARS-CoV-2: an in silico study. J. Taibah Univ. Sci. 2020, 14, 1563–1570. [Google Scholar] [CrossRef]
- Gupta, S.; Singh, A.K.; Kushwaha, P.P.; Prajapati, K.S.; Shuaib, M.; Senapati, S.; Kumar, S. Identification of potential natural inhibitors of SARS-CoV2 main protease by molecular docking and simulation studies. J. Biomol. Struct. Dyn. 2021, 39, 4334–4345. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multi-threading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, U.; Das, M.; Datta, G. Identification of bioactive components in ethanolic and aqueous extracts of Amorphophallus campanulatus tuber by GC-MS analysis. Int. J. Phytomedicine 2013, 5, 243. [Google Scholar]
- Gheda, S.; Khalil, M.; Gheida, S. In vitro and in vivo preliminary results on Spirulina platensis for treatment of impetigo: topical cream application. Afr. J. Biotechnol. 2013, 12, 2498–2509. [Google Scholar]
- Ghalloo, B.A.; Khan, K.-u.-R.; Ahmad, S.; Aati, H.Y.; Al-Qahtani, J.H.; Ali, B.; Mukhtar, I.; Hussain, M.; Shahzad, M.N.; Ahmed, I. Phytochemical profiling, in vitro biological activities, and in silico molecular docking studies of Dracaena reflexa. Molecules 2022, 27, 913. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.; Osilesi, O.; Adebawo, O.; Onajobi, F.; Karigidi, K.; Muhammad, L. Nutrients compositions and phytochemical contents of edible parts of Chrysophyllum albidum fruit. J. Nutr. Food Sci. 2017, 7, 1–9. [Google Scholar]
- Tonisi, S.; Okaiyeto, K.; Mabinya, L.V.; Okoh, A.I. Evaluation of bioactive compounds, free radical scavenging and anticancer activities of bulb extracts of Boophone disticha from Eastern Cape Province, South Africa. Saudi J. Biol. Sci. 2020, 27, 3559–3569. [Google Scholar] [CrossRef] [PubMed]
- Ferdosi, M.; Ahmed, H.; Khan, I.; Javaid, A. Fungicidal potential of flower extract of Cassia fistula against Macrophomina phaseolina and Sclerotium rolfsii. J. Anim. Plant Sci. 2022, 32, 1028–1034. [Google Scholar]
- Ruiz-Torres, V.; Rodríguez-Pérez, C.; Herranz-López, M.; Martín-García, B.; Gómez-Caravaca, A.M.; Arráez-Román, D.; Segura-Carretero, A.; Barrajón-Catalán, E.; Micol, V. Marine invertebrate extracts induce colon cancer cell death via ROS-mediated DNA oxidative damage and mitochondrial impairment. Biomolecules 2019, 9, 771–808. [Google Scholar] [CrossRef] [PubMed]
- Selvin, J.; Shanmughapriya, S.; Gandhimathi, R.; Seghal Kiran, G.; Rajeetha Ravji, T.; Natarajaseenivasan, K.; Hema, T. Optimization and production of novel antimicrobial agents from sponge associated marine actinomycetes Nocardiopsis dassonvillei MAD08. Appl. Microbiol. Biotechnol. 2009, 83, 435–445. [Google Scholar] [CrossRef]
- Aruna, V. Antimicrobial activity of diethyl phthalate: an in silico approach. J. Adv. Sci. Res. 2020, 11, 158–165. [Google Scholar]
- Nafie, M.S.; Awad, N.M.; Tag, H.M.; Abd El-Salam, I.M.; Diab, M.K.; El-Shatoury, S.A. Micromonospora species from rarely-exploited Egyptian habitats: chemical profile, antimicrobial, and antitumor activities through antioxidant property. Appl. Microbiol. Biotechnol. 2021, 105, 2427–2439. [Google Scholar] [CrossRef]
- Okudoh, V.I.; Wallis, F.M. Antimicrobial activity of rare actinomycetes isolated from natural habitats in KwaZulu-Natal, South Africa. S. Afr. J. Sci. 2007, 103, 216–222. [Google Scholar]
- Ambati, M.; Kumar, M.S. Microbial diversity in the Indian Ocean sediments: an insight into the distribution and associated factors. Curr. Microbiol. 2022, 79, 115. [Google Scholar] [CrossRef] [PubMed]
- DeCastro, M.-E.; Escuder-Rodríguez, J.-J.; Becerra, M.; Rodríguez-Belmonte, E.; González-Siso, M.-I. Comparative metagenomic analysis of two hot springs from Ourense (northwestern Spain) and others worldwide. Front. Microbiol. 2021, 3534. [Google Scholar] [CrossRef] [PubMed]
- Meena, B.; Rajan, L.A.; Vinithkumar, N.V.; Kirubagaran, R. Novel marine actinobacteria from emerald Andaman & Nicobar Islands: a prospective source for industrial and pharmaceutical by-products. BMC Microbiol. 2013, 13, 145. [Google Scholar]
- Arumugam, T.; Kumar, P.S.; Kameshwar, R.; Prapanchana, K. Screening of novel actinobacteria and characterization of the potential isolates from mangrove sediment of south coastal India. Microb. Pathog. 2017, 107, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.A. Overview of extremophiles and their food and medical applications. In Physiological and Biotechnological Aspects of Extremophiles, Salwan, R.,Sharma, V., Eds.; Elsevier: Massachusetts, USA, 2020; pp. 65–87. [Google Scholar]
- Muduli, P.R.; Vinithkumar, N.; Begum, M.; Robin, R.; Vardhan, K.V.; Venkatesan, R.; Kirubagaran, R. Spatial variation of hydrochemical characteristics in and around port Blair Bay Andaman and Nicobar Islands, India. World Appl. Sci. J. 2011, 13, 564–571. [Google Scholar]
- Gupta, P.; Lal, B.; Nair, V. Solid CO2 storage by hydrate-based geo sequestration. In Nanotechnology for CO2 Utilization in Oilfield Applications, Sharma, T., Chaturvedi, K.R., Trivedi, J.J., Eds.; Elsevier: Massachusetts, USA, 2022; pp. 251–273. [Google Scholar]
- Yano, Y.; Nakayama, A.; Ishihara, K.; Saito, H. Adaptive changes in membrane lipids of barophilic bacteria in response to changes in growth pressure. Appl. Environ. Microbiol. 1998, 64, 479–85. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Tao, B.; Guo, J.; Li, J.; Chen, G. Changes in the microbial community structure and soil chemical properties of vertisols under different cropping systems in northern China. Front. Environ. Sci. 2018, 6, 132. [Google Scholar] [CrossRef]
- Salaria, N.; Furhan, J. Antibacterial potential of actinomycetes from north-western Himalayas against pathogenic bacteria. Biol. Bull. 2021, 48, 281–289. [Google Scholar] [CrossRef]
- Tang, S.-K.; Tian, X.-P.; Zhi, X.-Y.; Cai, M.; Wu, J.-Y.; Yang, L.-L.; Xu, L.-H.; Li, W.-J. Haloactinospora alba gen. nov., sp. nov., a halophilic filamentous actinomycete of the family Nocardiopsaceae. Int. J. Syst. Evol. Microbiol. 2008, 58, 2075–2080. [Google Scholar] [CrossRef]
- De La Hoz-Romo, M.C.; Díaz, L.; Villamil, L. Marine actinobacteria a new source of antibacterial metabolites to treat acne vulgaris disease-a systematic literature review. Antibiotics (Basel) 2022, 11, 965. [Google Scholar] [CrossRef]
- Okudoh, V.I. Isolation and Characterization of Antibiotic(s) Produced by Bacteria from KwaZulu-Natal Soils. PhD, University of KwaZulu-Natal, KwaZulu-Natal Province, 2010.
- Mangamuri, U.; Muvva, V.; Poda, S.; Naragani, K.; Munaganti, R.K.; Chitturi, B.; Yenamandra, V. Bioactive metabolites produced by Streptomyces cheonanensis VUK-A from Coringa mangrove sediments: isolation, structure elucidation and bioactivity. 3 Biotech. 2016, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Fabry, W.; Okemo, P.O.; Ansorg, R. Antibacterial activity of East African medicinal plants. J. Ethnopharmacol. 1998, 60, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Mogana, R.; Adhikari, A.; Tzar, M.; Ramliza, R.; Wiart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC complement. med. ther. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aligiannis, N.; Kalpoutzakis, E.; Mitaku, S.; Chinou, I.B. Composition and antimicrobial activity of the essential oils of two Origanum species. J. Agric. Food Chem. 2001, 49, 4168–4170. [Google Scholar] [CrossRef] [PubMed]
- Nxumalo, C.I.; Ngidi, L.S.; Shandu, J.S.E.; Maliehe, T.S. Isolation of endophytic bacteria from the leaves of Anredera cordifolia CIX1 for metabolites and their biological activities. BMC Complement. Med. Ther. 2020, 20, 300. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chang, J.S.; Herberg, J.T.; Horng, M.-M.; Tomich, P.K.; Lin, A.H.; Marotti, K.R. Allosteric inhibition of Staphylococcus aureus D-alanine:D-alanine ligase revealed by crystallographic studies. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 15178–15183. [Google Scholar] [CrossRef] [PubMed]
- Nastasă, C.; Vodnar, D.C.; Ionuţ, I.; Stana, A.; Benedec, D.; Tamaian, R.; Oniga, O.; Tiperciuc, B. Antibacterial evaluation and virtual screening of new thiazolyl-triazole Schiff bases as potential DNA-gyrase inhibitors. Int. J. Mol. Sci. 2018, 19, 222. [Google Scholar] [CrossRef]
- He, J.; Qiao, W.; An, Q.; Yang, T.; Luo, Y. Dihydrofolate reductase inhibitors for use as antimicrobial agents. Eur. J. Med. Chem. 2020, 195, 112268. [Google Scholar] [CrossRef]
- Hata, H.; Phuoc Tran, D.; Marzouk Sobeh, M.; Kitao, A. Binding free energy of protein/ligand complexes calculated using dissociation parallel cascade selection molecular dynamics and Markov state model. Biophys. Physicobiol. 2021, 18, 305–316. [Google Scholar] [CrossRef]
- Ashraf, Z.; Rafiq, M.; Seo, S.-Y.; Babar, M.M. Synthesis, kinetic mechanism and docking studies of vanillin derivatives as inhibitors of mushroom tyrosinase. Bioorg. Med. Chem. 2015, 23, 5870–5880. [Google Scholar] [CrossRef]
- Maliehe, T.S.; Selepe, T.N.; Mthembu, N.N.; Shandu, J.S. Antibacterial and anti-quorum sensing activities of Erianthemum dregeis leaf extract and molecular docking. Pharmacogn. J. 2023, 15, 279–285. [Google Scholar] [CrossRef]
- Chakraborty, C.; Mallick, B.; Sharma, A.R.; Sharma, G.; Jagga, S.; Doss, C.G.; Nam, J.S.; Lee, S.S. Micro-environmental signature of the interactions between druggable target protein, dipeptidyl peptidase-iv, and anti-diabetic drugs. Cell J. 2017, 19, 65–83. [Google Scholar]
- Amer, H.H.; Eldrehmy, E.H.; Abdel-Hafez, S.M.; Alghamdi, Y.S.; Hassan, M.Y.; Alotaibi, S.H. Antibacterial and molecular docking studies of newly synthesized nucleosides and Schiff bases derived from sulfadimidines. Sci. Rep. 2021, 11, 17953. [Google Scholar] [CrossRef]
Figure 1.
Map showing the six beaches of KZN Province, RSA, where water samples were obtained (Source: Ngema, unpublished).
Figure 1.
Map showing the six beaches of KZN Province, RSA, where water samples were obtained (Source: Ngema, unpublished).
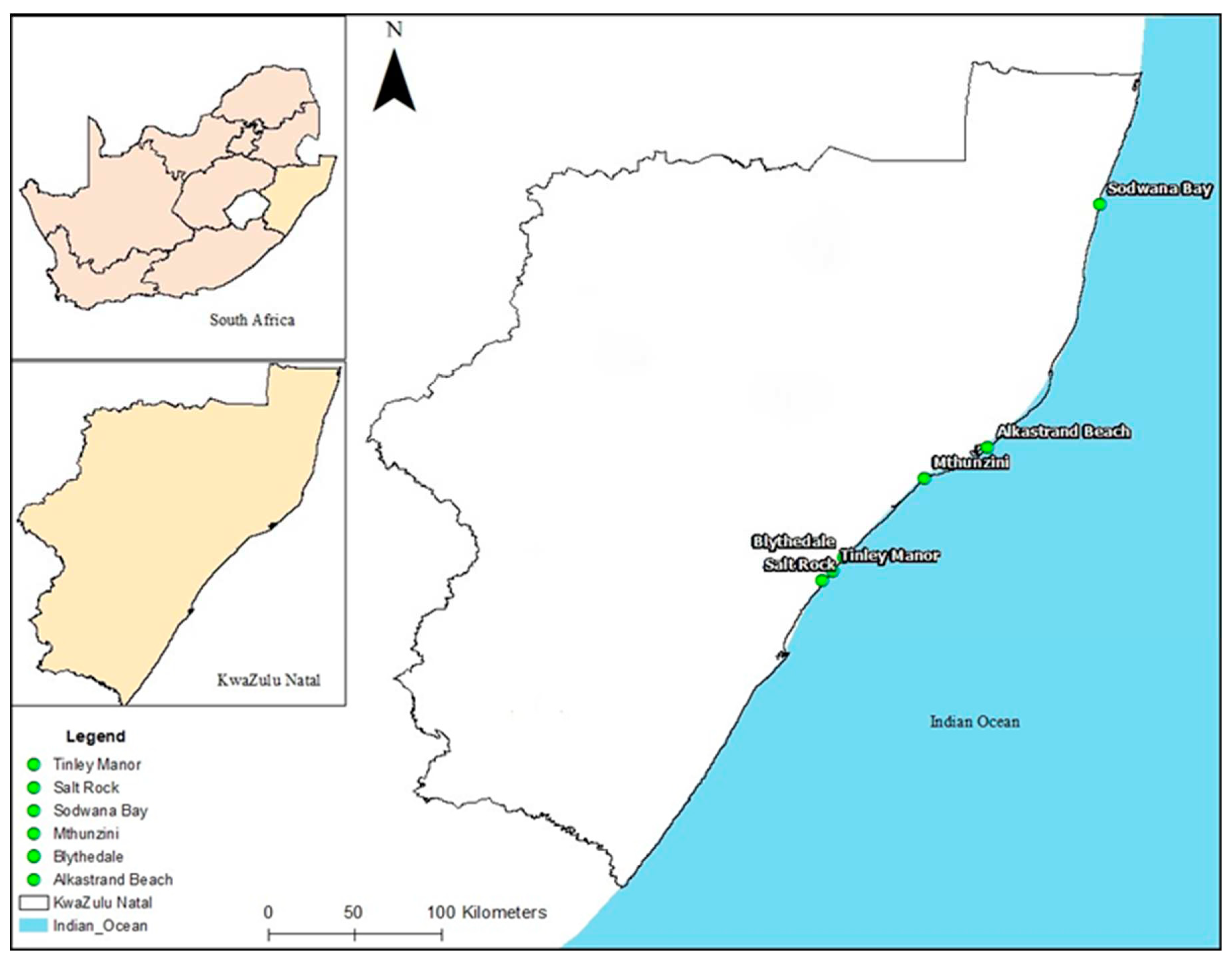
Figure 2.
Phylogenetic tree obtained by neighbour-joining analysis of 16S rRNA gene sequences showing the position of N. dassonvillei SOD(B)ST2SA2 and its phylogenetic neighbours. Numbers on branch nodes are bootstrap values (1000 re-samplings; only values over 50% are displayed). Bar, 10% sequence divergence. GenBank accession numbers are also given in parenthesis.
Figure 2.
Phylogenetic tree obtained by neighbour-joining analysis of 16S rRNA gene sequences showing the position of N. dassonvillei SOD(B)ST2SA2 and its phylogenetic neighbours. Numbers on branch nodes are bootstrap values (1000 re-samplings; only values over 50% are displayed). Bar, 10% sequence divergence. GenBank accession numbers are also given in parenthesis.

Figure 3.
Zones of inhibition (28.67±1.15 mm) of L. monocytogenes ILestanBR363 on Mueller-Hinton agar.
Figure 3.
Zones of inhibition (28.67±1.15 mm) of L. monocytogenes ILestanBR363 on Mueller-Hinton agar.
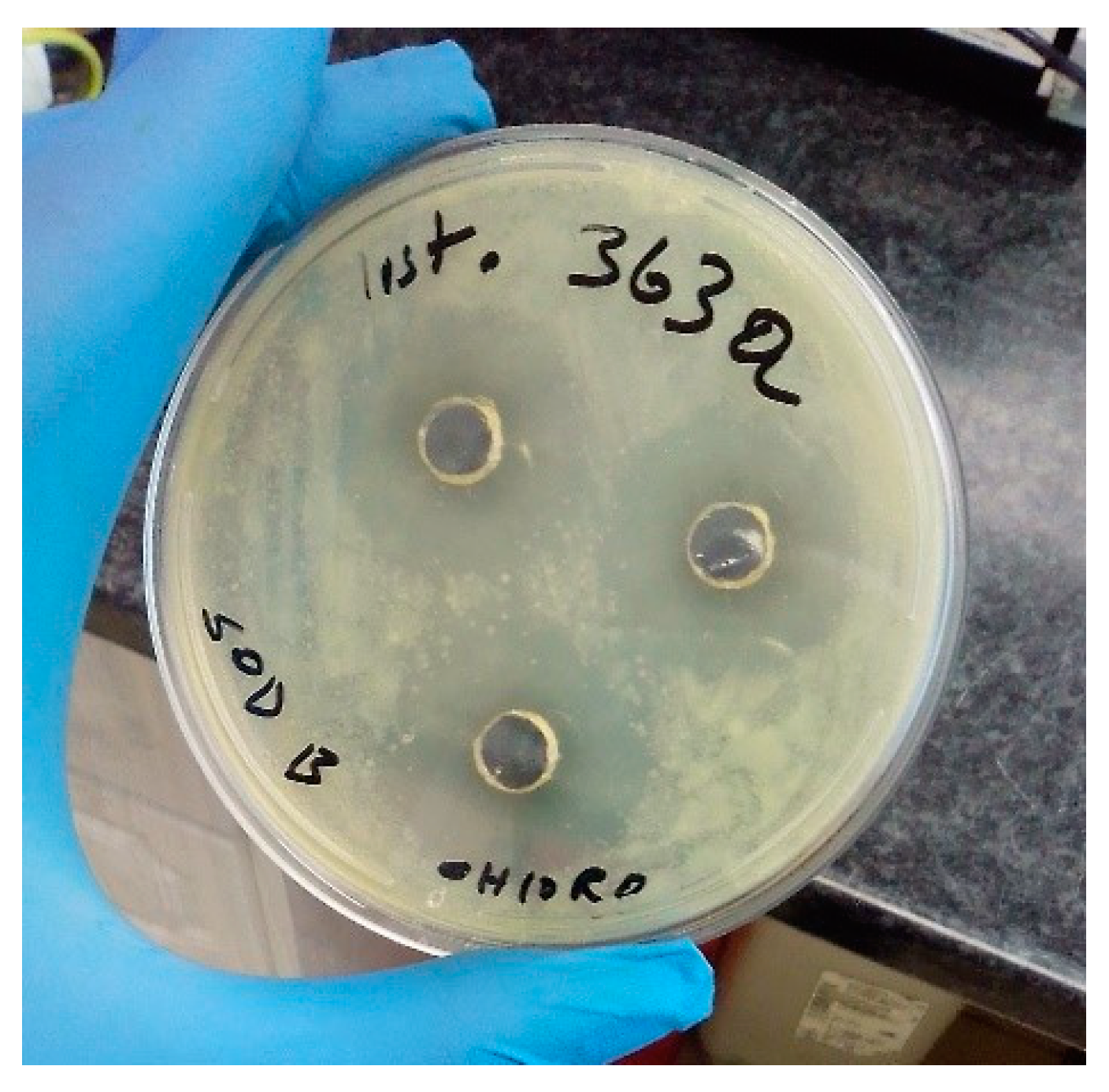
Figure 4.
2D and 3D binding interactions of the ligands against dihydrofolate reductase (PDB ID: 1DIS), respectively. The 3D interactions show the ligand in a binding pocket of the enzyme. Dashed lines indicate the interactions between the ligands and the amino acids of the enzyme.
Figure 4.
2D and 3D binding interactions of the ligands against dihydrofolate reductase (PDB ID: 1DIS), respectively. The 3D interactions show the ligand in a binding pocket of the enzyme. Dashed lines indicate the interactions between the ligands and the amino acids of the enzyme.
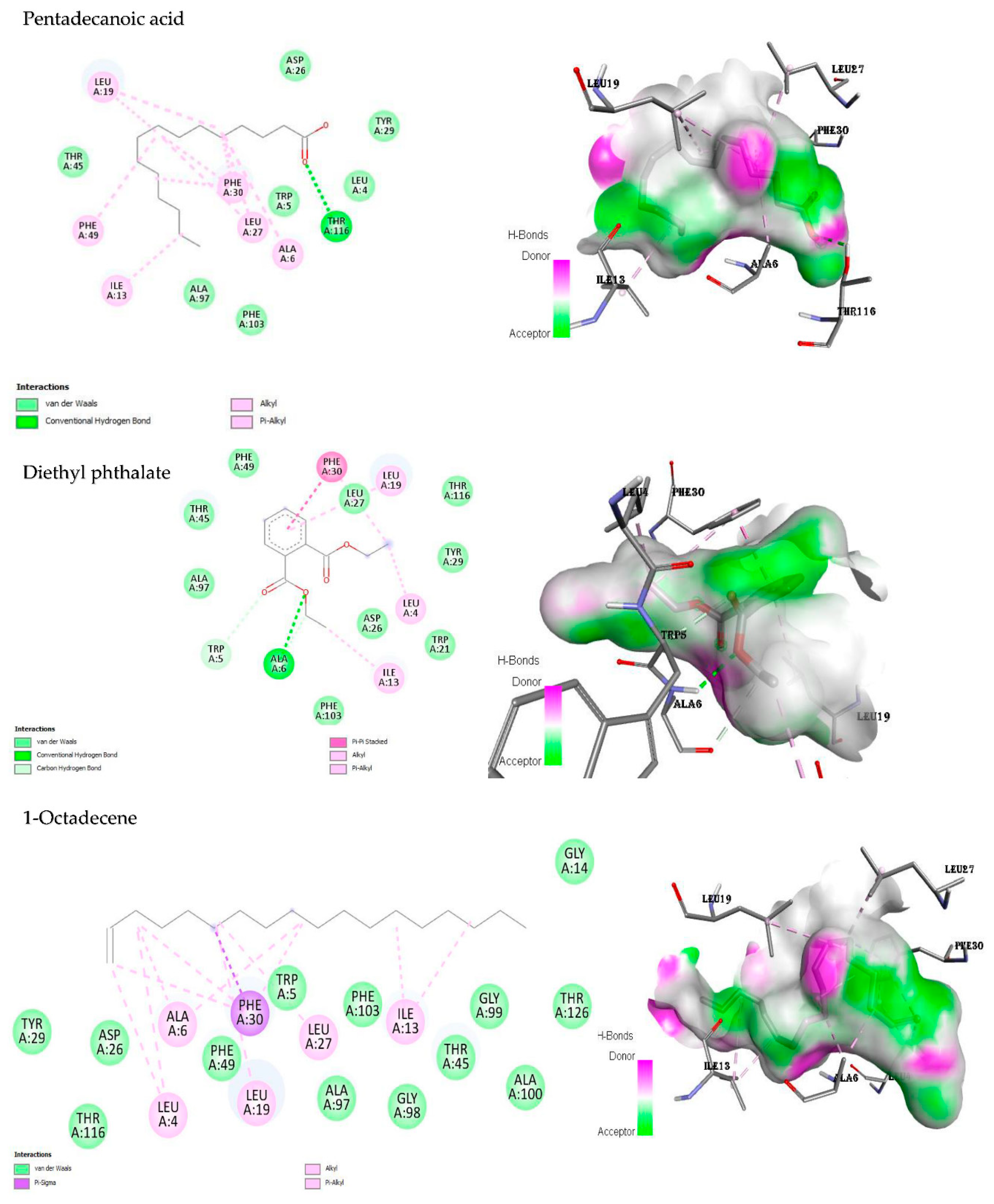

Table 1.
L. monocytogenes and STEC isolates from beef and beef products.
| Sample code | Sample description | L. monocytogenes strains | STEC strains | Resistance phenotype for STEC strains |
|---|---|---|---|---|
| KEmpBS18 | Chuck steak bone | — | KEmpBS18* | A25-AP10-TE30-TS25 |
| KEmpBP21 | Beef burger | — | KEmpBP21* | A25-AP10-TE30-TS25 |
| KVulFS71 | Brisket | — | KVulFS71* | A25-AP10-CIP5-C30-FOX30-TE30 |
| KRbyGR83 | Biltong powder | — | KRbyGR83* | A25-AP10-CIP5-C30-TS25 |
| KGEO151 | Ox intestines | — | KGEO151* | A25-AP10-TE30-TS25 |
| KGEO161 | Ox liver | KGEO161 | KGEO161 | A25-AP10 |
| KmelDO248 | Cow heels | — | KmelDO248* | A25-AP10-C30-TS25 |
| ILemanEO299 | Ox lungs | ILemanEO299 | ILemanEO299* | A25-AP10-C30-TS25 |
| ILemanER317 | Burger | ILemanER317 | ILemanER317 | A25-AP10-FOX30 |
| ILemanAP345 | Mince | ILemanAP345 | — | |
| ILestanBR361 | Droewors | ILestanBR361 | — | |
| ILestanBR363 | Droewors | ILestanBR363 | — | |
| ILestanGP395 | Mince | ILestanGP395 | — | |
| ILestanGP400 | Burger patties | ILestanGP400 | — |
Key: —, indicates that no strains were isolated; *, refers to multi-drug resistant strains; A25, AP10, CIP5, C30, TS25, and FOX30 refer to amoxicillin (25 µg), ampicillin (10 µg), ciprofloxacin (5 µg), chloramphenicol (3 0µg), co-trimoxazole (1.25-23.75 µg), and cefoxitin (30 µg), respectively.
Table 2.
Physicochemical parameters of the different beaches of KwaZulu-Natal.
| Beaches | Physicochemical parameters | ||||||
|---|---|---|---|---|---|---|---|
| pH ± SD | DO (%) ± SD | Specific conductivity (mS/cm) ± SD | Total dissolved solids (g/L) ± SD | Salinity (PSU) ± SD | Temperature (°C) ± SD | Pressure (mmHg) ± SD | |
| Alkantstrand | 6.66±0.22ab | 30.22±4.12a | 49.14±4.71a | 24.57±2.36a | 32.39±3.50a | 24.17±0.09a | 749.79±0.33a |
| Blythedale | 6.86±0.30ab | 38.70±8.15a | 51.85±0.39a | 25.93±0.19a | 34.11±0.28a | 24.94±0.44a | 748.88±0.10b |
| Mthunzini | 6.79±0.09ab | 41.28±5.53a | 53.43±0.22a | 26.72±0.11a | 35.22±0.16a | 26.96±0.09b | 754.35±0.16c |
| Salt Rock | 6.73±0.29ab | 40.67±1.72a | 48.96±5.25a | 24.49±2.63a | 32.04±3.70a | 26.03±0.01c | 754.51±0.09c |
| Sodwana | 7.13±0.05a | 42.41±5.50a | 47.09±10.09a | 23.55±5.04a | 30.81±7.14a | 23.24±0.49d | 759.56±0.49d |
| Tinley Manor | 6.55±0.13b | 43.31±4.31a | 50.88±2.16a | 25.45±1.08a | 33.41±1.60a | 24.94±0.27a | 754.53±0.09c |
Values with different letters (a, b, c and d) on the same column are significantly (p < 0.05) different. SD represents standard deviation.
Table 3.
Screening of antibacterial activity against multi-drug resistant STEC strains.
| STEC | Zones of inhibition (mm) | |||||
|---|---|---|---|---|---|---|
| Actinomycetes | ||||||
| Primary screening | Secondary screening | |||||
| BLYST2SA3(i) | SOD(A)ST2SA2 | SOD(B)ST2SA2 | SRST1SA3 | TBST2SA1 | SOD(B)ST2SA2 | |
| KEmpBS18 | + | +++ | ++ | + | − | + |
| KEmpBP21 | ++ | ++ | ++ | + | + | + |
| KVulFS71 | ++ | − | − | ++ | − | + |
| KRbyGR83 | ++ | ++ | ++ | ++ | − | − |
| KGEO151 | + | ++ | ++ | + | + | − |
| KmelDO248 | + | +++ | + | ++ | + | + |
| ILemanEO299 | + | +++ | ++ | ++ | - | + |
| O157:H7 ATCC 43888 | +++ | +++ | +++ | +++ | + | + |
Key: Inhibition zone diameter index: + (≤ 29 mm) weak activity, ++ (30 – 49 mm) moderate activity, +++ (≥ 50 mm) strong activity and – denotes no activity.
Table 4.
Screening of antibacterial activity against L. monocytogenes strains.
| L. monocytogenes | Zones of inhibition (mm) | |||||
|---|---|---|---|---|---|---|
| Actinomycetes | ||||||
| Primary screening | Secondary screening | |||||
| BLYST2SA3(i) | SOD(A)ST2SA2 | SOD(B)ST2SA2 | SRST1SA3 | TBST2SA1 | SOD(B)ST2SA2 | |
| KGEO161 | +++ | +++ | +++ | +++ | +++ | + |
| ILemanAP345 | +++ | +++ | +++ | +++ | +++ | + |
| ILemanEO299 | +++ | +++ | +++ | +++ | +++ | + |
| ILemanER317 | +++ | +++ | +++ | +++ | +++ | + |
| ILestanBR361 | +++ | +++ | +++ | +++ | +++ | + |
| ILestanBR363 | +++ | +++ | +++ | +++ | +++ | + |
| ILestanGP395 | +++ | +++ | +++ | +++ | +++ | + |
| ILestanGP400 | +++ | +++ | +++ | +++ | +++ | + |
Key: Inhibition zone diameter index: + (≤ 29 mm) weak activity, ++ (30 – 49 mm) moderate activity, +++ (≥ 50 mm) strong activity.
Table 5.
BLAST prediction names of actinomycete isolates.
| Beach name | Strain number | GenBank closest known Species (accession number) | Identification |
|---|---|---|---|
| Blythedale | BLYST2SA3(i) |
Streptomyces violaceoruber (CP0205701) Streptomyces californicus (CP070260.1) |
Streptomyces sp. |
| Sodwana | SOD(A)ST2SA2 |
Nocardiopsis dassonvillei (MF321787.1) Nocardiopsis dassonvillei (MH432693.1) Nocardiopsis dassonvillei subsp. albirubida (MG661744.1) Nocardiopsis dassonvillei (CP022434.1) Nocardiopsis dassonvillei (JN253591.1) |
Nocardiopsis dassonvillei |
| Sodwana | SOD(B)ST2SA2 |
Nocardiopsis dassonvillei (MT393632.1) Nocardiopsis dassonvillei (MH432693.1) Nocardiopsis dassonvillei subsp. albirubida (MG661744.1) Nocardiopsis dassonvillei (CP022434.1) Nocardiopsis dassonvillei subsp. albirubida (KC514117.1) Nocardiopsis dassonvillei (JN253591.1) Nocardiopsis dassonvillei (MF321787.1) |
Nocardiopsis dassonvillei |
| Salt Rock | SRST1SA3 | Streptomyces sp. (MH910227.1) | Streptomyces sp. |
| Tinley Manor | TBST2SA1 | Streptomyces sp. (CP047147.1) Streptomyces albidoflavus (MF663704.1) | Streptomyces sp. |
Table 6.
Major compounds identified in the chloroform extract of N. dassonvillei SOD(B)ST2SA2 and their potential biological role.
Table 6.
Major compounds identified in the chloroform extract of N. dassonvillei SOD(B)ST2SA2 and their potential biological role.
| No. | Molecular formula | Compound | Area % | Biological activity | Reference |
|---|---|---|---|---|---|
| 1. | C18H34O2 | 6-Octadecenoic acid, (Z) | 20.94 | Antimicrobial, anti-inflammatory, anti-androgenic, cancer preventative, dermatitigenic hypochlolesterolemic, 5-alpha reductase inhibitor, and anemiagenic insectifuge | [51,52] |
| 2. | C18H36 | 1-Octadecene | 9.22 | Antimicrobial and anticancer activities | [53] |
| 3. | C18H33F3O2 | Trifluoroacetoxy hexadecane | 7.82 | Antifungal, anti-oxidant and anticancer activity | [54,55,56] |
| 4. | C15H30O2 | Pentadecanoic acid | 5.86 | Anticancer, antibacterial and antifungal activity | [57] |
| 5. | C12H14O4 | Diethyl phthalate | 3.43 | Antibacterial, antifungal, and anticancer activity | [58,59,60] |
Table 7.
Antibacterial activity of N. dassonvillei SOD(B)ST2SA2’s crude extracts against L. monocytogenes strains.
Table 7.
Antibacterial activity of N. dassonvillei SOD(B)ST2SA2’s crude extracts against L. monocytogenes strains.
| L. monocytogenes strains | Zones of inhibition (mm) | Chloroform extract | Ciprofloxacin | |||||
|---|---|---|---|---|---|---|---|---|
| Crude extracts (100 mg/mL) | Control (10 µg/mL) | MIC (mg/mL) | MBC (mg/mL) | MIC (µg/mL) |
MBC (µg/mL) |
|||
| Chloroform | Methanol | Ethyl acetate | Ciprofloxacin | |||||
| KGEO161 | 15.33±0.58 | 0 | 0 | 24.7±1.2 | 6.25 | > 25 | 2.5 | 2.5 |
| ILemanAP345 | 0 | 0 | 0 | 27.0±2.7 | 6.25 | 25 | 0.16 | 1.25 |
| ILemanEO299 | 20.33±0.58 | 0 | 0 | 40.3±2.1 | 6.25 | 25 | 0.16 | 1.25 |
| ILemanER317 | 14.67±0.58 | 0 | 0 | 24.0±1.0 | 6.25 | 25 | 0.63 | 5 |
| ILestanBR361 | 25.67±1.15 | 0 | 0 | 36.3±1.2 | 6.25 | > 25 | 0.16 | 5 |
| ILestanBR363 | 28.67±1.15 | 0 | 0 | 36.3±1.5 | 6.25 | > 25 | 0.63 | 2,5 |
| ILestanGP395 | 21.33±0.58 | 0 | 0 | 36.0±5.3 | 6.25 | > 25 | 0.16 | 2,5 |
| ILestanGP400 | 23.00±1.00 | 0 | 0 | 27.3±1.2 | 6.25 | 25 | 0.63 | 1.25 |
| ATCC 15313 | 27.00±0.40 | 0 | 0 | 42.0±2.7 | 6.25 | 25 | 2.5 | 2.5 |
Table 8.
Antibacterial activity of N. dassonvellei SOD(B)ST2SA2’s crude extracts against STEC strains.
Table 8.
Antibacterial activity of N. dassonvellei SOD(B)ST2SA2’s crude extracts against STEC strains.
| STEC strains | Zones of inhibition (mm) | Chloroform extract | Ciprofloxacin | |||||
|---|---|---|---|---|---|---|---|---|
| Crude extracts (100 mg/mL) | Control (20 µg/mL) | MIC (mg/mL) | MBC (mg/mL) |
MIC (µg/mL) |
MBC (µg/mL) |
|||
| Chloroform | Methanol | Ethyl acetate | Ciprofloxacin | |||||
| KEmpBS18 | 10.67±0.58 | 0 | 0 | 46.33±2.31 | 25 | 25 | 0.039 | 0.313 |
| KEmpBP21 | 10.33±0.58 | 0 | 0 | 45.33±0.58 | 25 | 25 | 0.039 | 0.078 |
| KVulFS71 | 11.67±0.58 | 0 | 0 | 38±2.65 | 3.13 | 6.25 | 0.039 | 0.078 |
| KRbyGR83 | 15.67±1.15 | 0 | 0 | 47.33±4.16 | 0.78 | 3.13 | 0.039 | 0.039 |
| KGEO151 | 0 | 0 | 0 | 42.33±2.31 | − | − | 0.039 | 0.039 |
| KmelDO248 | 14.00±1.00 | 0 | 0 | 48±1.73 | 1.57 | 3.13 | 0.039 | 0.313 |
| ILemanEO299 | 14.33±0.58 | 0 | 0 | 50±5.57 | 25 | >25 | 0.039 | 0.078 |
| O157:H7 ATCC 43888 | 16.33±1.15 | 0 | 0 | 40.67±1.15 | 1.57 | 3.13 | 0.039 | 0.078 |
Table 9.
Binding energies of ligands to bacterial proteins (receptors).
| Ligands | Receptors | Binding energy scores (Kcal/mol) |
|---|---|---|
| Pentadecanoic acid | DDI | −5.0 |
| DNA gyrase B | −5.2 | |
| Dihydrofolate reductase | −6.3 | |
| Diethyl phthalate | DDI | −5.8 |
| DNA gyrase B | −6.0 | |
| Dihydrofolate reductase | −7.2 | |
| 1-Octadecene | DDI | −4.9 |
| DNA gyrase B | −4.8 | |
| Dihydrofolate reductase | −6.4 | |
| Ciprofloxacin (control) | DDI | −7.5 |
| DNA gyrase B | −7.6 | |
| Dihydrofolate reductase | −9.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



