
Submitted:
14 August 2023
Posted:
15 August 2023
You are already at the latest version
Abstract
Paratuberculosis (PTB) is a chronic granulomatous enteritis caused by Mycobacterium avium subsp. paratuberculosis (MAP) that affects a wide variety of domestic and wild animals. It is considered as one of the diseases with the highest economic impact in the ruminant industry. Despite many efforts and intensive research, PTB control is still controversially discussed and diagnostic and immunoprophylactic tools lack great limitations. Thus, models play a crucial role in understanding the pathogenesis of infection and disease, and in testing novel vaccine candidates. Here, we review the potential and limitations of different experimental approaches currently used in PTB research, focusing on laboratory animals and cell based models. The aim of this review is to offer a vision of the models that have been used and what has been achieved or discovered with each one so that the reader can choose the best model to answer their scientific questions and prove their hypotheses. Also, we bring forward new approaches that we consider worth exploring in the near future.
Keywords:
paratuberculosis
; in vivo
; in vitro
; ex vivo
; infection model
; macrophage
; neutrophil
; epithelial cell
; rabbit
; experimental infection
1. Introduction
Johne’s disease or paratuberculosis (PTB), caused by Mycobacterium avium subspecies paratuberculosis (MAP), is a chronic granulomatous enteritis which affects large populations of ruminants globally. Prevention strategies to combat the spread of PTB among herds involve adhering to strict calving practices to ensure young susceptible animals do not come in contact with MAP contaminated colostrum, milk, or feces. Achieving PTB control requires critical advances in both diagnosis and vaccination areas. It is also well accepted that progress in these areas requires a deeper knowledge of host—pathogen interactions during infection. Although PTB was first described in the late 1800s its pathogenic mechanisms have not been clearly defined. This is probably due to the lack of economic and adequate laboratory animals that faithfully resemble ruminant PTB and also to the absence of harmonization in the use of models among PTB research groups. It has been said that the optimization of an animal model to study MAP infection is stymied by the complexity of this disease [1]. For this reason, some groups have focused on large animal models as these are the target species for which control is desired. However, these animal models are many times overlooked by other research groups, in part because of the prohibitive costs of maintaining these animals in a research environment. In this context, better experimental models are needed and these should be clearly defined and shared throughout the PTB research community.
Extensive reviews have been previously published elsewhere describing both ruminant and laboratory animal models [2,3]. Here, we review the potential and limitations of different experimental approaches used in PTB research, focusing on non-ruminant animals and cell based models. Figure 1 summarizes this information and illustrates the relative contribution of each model to this field. In addition, we complete the review bringing forward new approaches that we consider worth exploring in the near future.
2. In vivo animal models in PTB research
As previously stated, ruminant species are the first choice considering their role as natural hosts of this bacterium and because the disease causes economic losses mostly in ruminant livestock, meaning that a vaccine for these species would be highly valued. However, the use of ruminant species as models, even small ones, may be restricted by several reasons related to space requirements, costs of the animals and the maintenance of the facilities, volume of daily excreta produced and therefore the need of permanent cleaning, complexity in restraint maneuvers, among others. In addition, when the observation of clinical features are pursued, ruminant MAP infection experiments get lengthy, adding to cost and slowing down scientific progress. Laboratory animal models are normally the most near to reality choice when evaluation of new therapies, vaccine or pathogenesis studies are the goal. Although it is impossible to reflect the complete image of PTB in laboratory animals, these non-ruminant models are advantageous because they are less expensive and, in some cases, PTB compatible signs can be achieved in a shorter period of time. They can be used in the first steps of vaccine screening or when large animal modeling is not possible. In this review, we will focus on other alternative in vivo models, based on laboratory species. Also, we will refer to the amoebae, as the simplest eukaryotic model of infection.
2.1. Mouse model
Mice have been extensively used in PTB research for pathogenesis and vaccine prototype testing. Actually, the use of this model has been recommended for initial steps of vaccine screening due to similarities with the granulomatous inflammation and immunological responses between mice and ruminants [2]. An in depth review of studies including this animal model has been performed recently [3]. Therefore this section will only highlight the main features of this mode.
Mice strains with different genetic backgrounds have been assayed, although BALB/c and C57BL/6 have been the most widely used strains [3]. MAP has been administered through the intraperitoneal route (IP) in most cases (67% of the studies reviewed in [3]) followed by the oral route (20% of the studies reviewed in [3]). The oral route has shown to be less reproducible and it does not recreate all the classical features of intestinal PTB [3,4] only producing granulomatous lesions in mesenteric lymph nodes [5] and disseminated infection in high dose challenges [4]. However, the immune profile achieved in this model is similar to that observed in cattle [4]. Intraperitoneal challenge produces changes by 6-12 weeks post-infection (w.p.i.), mainly lymphocytic inflammatory response, whereas granulomatous inflammation is detected by 12 w.p.i. [6]. Shao et al. [7] reported differences in CFU load in liver from 2-8 w.p.i by the intraperitoneal route, with no bacterial recovery after that. They also detected differences in weight gain among groups from 2-12 w.p.i. Also, a recent study compared the early stage of MAP infection through different administration routes in the murine model, detecting increases in the weight of the spleen and the liver and more severe histopathological changes through the IP route compared to the oral route [8].
The infection outcome in the murine model is normally evaluated by assessing MAP burden in tissues and histopathology of liver, spleen and intestine, since gross pathological lesions are normally not achieved. The drawback of this model regarding PTB is that it fails to reproduce diarrhea or severe intestinal lesions. Blood extraction in mice can be difficult impeding in vivo follow-up of many parameters at the same time that are quantifiable in blood or its components (humoral or cellular). Performing ex vivo functional cell assays can be afforded by increasing the number of animals, dedicating each individual to a specific time-point and by isolating bone marrow at the experiment end-points when immune cell analysis is required [9,10,11].
2.2. Rabbit model
Rabbits are susceptible to the development of MAP infection in both experimental [12,13,14,15,16] and natural [17,18] conditions. Diarrhoea, fecal shedding, weight loss and intestinal colonization signs can be studied in the rabbit model.
First studies on experimental MAP challenge in rabbits were mostly carried out to prove infection and attempt to develop an appropriate model to study pathogenesis and screen vaccine candidates. Therefore, different animal ages, challenge doses and routes including intraperitoneal, intravenous and oral were tested [13,15,19,20,21]. The oral route mimicking the natural route of MAP infection was the most pursued. Further studies have used this challenge route to evaluate the impact of: diet [16] or a probiotic [22] on MAP infection; MAP infection on microbiome [23]; vaccination sequence on the progression of MAP infection and immunological response [24]; and passively transferred antibodies on MAP colonization (personal communication Fernández and Jolly [25]). This oral challenge model has also been complemented with ex vivo assays that permit functional analysis of immune cells and the study of trained immunity exerted by vaccination [26]. When using this model it should be considered that reliable reproduction of typical clinical, pathological and microbiological signs seems to depend greatly on strain origin and passage, obtaining higher degrees of infection when low passage cattle field strains are used. Key aspects of the most relevant studies involving experimental oral MAP infection in rabbits are summarized in Table 1.
The rabbit digestive tract can closely resemble that of ruminants because of the similarities between the sacculus rotundus in rabbits and the ileocaecal valve in ruminants. Also relevant to some extent is the fact that the physiological body temperature of the rabbit (38.3 - 39.4 ºC) is closer to that of the cow (37.8 - 39.2 ºC) compared to that of the mouse (36.5 - 38 ºC). This could somehow impact the results since it seems that MAP infection is enhanced at 39 ºC compared to 37 ºC [28]. Finally, the rabbit’s biological cycle allows short term challenge experiments permitting the development of mild PTB signs in less than 5 months. These characteristics, together with their size, handling and blood sampling easiness, as well as the volume of blood possible to be collected in periodical sampling, make rabbits a convenient experimental species to model PTB, offering important advantages for studying key aspects of paratuberculosis that cannot be investigated in other laboratory animals because many symptoms such as diarrhoea or faecal shedding cannot be reproduced in those models. Regarding their immunological system, two particular features would place the rabbit as a model closer to ruminants than the mouse: on one hand, the presence of a primary intestine-associated lymphoid structure, which is determinant for the generation of diversity in the B cell repertoire (ileal Peyer's patch in ruminants and cecum in rabbits); on the other hand, the fact that both rabbits and ruminants have been classified as γδ high species according to the much higher proportion of γδ T lymphocytes (up to 60%) in their T cell circulating pool than the γδ low species, such as the mouse, in which the proportion of γδ T cells is around 5% [29]. Although the role of γδ T lymphocytes in PTB is not fully understood, bovine γδ T lymphocytes are known to be early responders to MAP infection [30,31] and evidence for a key role of this subpopulation in the clinical phase of the disease has also been found from observations in naturally infected animals [32,33,34].
2.3. Ferret model
The ferret has been used as a model of infectious diseases [35], mainly for tuberculosis (TB), for which this species has been postulated as an environmental reservoir [36,37]. In 1997, Lugton and coworkers [38] reported a chronic low-level natural infection of ferrets with M. avium subsp. avium. More recently, Bannantine et al. [39] have published the first report using ferrets as an experimental model of MAP infection. They stated that 3-4 month old female ferrets challenged orally with 108 CFU MAP lost body weight after 16 w.p.i., developed an specific antibody response by week 13, while IFN-γ response remained undetectable until 20 w.p.i., the final endpoint of their experiment. Taking into account those results, they proposed ferrets as a valid small animal model for testing virulence of MAP strains. As ferrets have a very short intestinal tract, as an adaptation of their strict carnivore diet [40], Bannatine et al. [39] have suggested that this anatomical feature could be an advantage for using this species as an experimental model of PTB, as the site of infection after oral inoculation would be well circumscribed. The usefulness of this carnivorous animal as a model in the study of PTB should be further confirmed.
2.4. Protozoan model
Eukaryotic organisms like amoeba have been noted to interact with mycobacterial species in the environment. Based on the ability for some mycobacteria to grow within environmental amoebae [41,42,43] and data suggesting increased virulence of mycobacteria upon replication within amoebae [44,45] Phillips et al. [46] have evaluated the influence of MAP infection on amoeba metabolic activity in the Acanthamoeba castellanii infection model, finding that this amoeba can be used as a quick initial screening tool for selection of virulence factors with relevance in macrophage infection.
3. From in vivo animal models to ex vivo and in vitro systems
Apart from in vivo models, most of the contributions to the knowledge of this pathology have been made working on ex vivo or in vitro models, focused on the use of cells of epithelial and/or of phagocytic origin; since the target organ of this infection in ruminants is the intestine and the associated lymphoid tissue, and macrophages are the main cellular niche in which MAP can survive and replicate within its host.
Ex vivo and in vitro cellular infection assays can be fast and cost effective alternatives to in vivo experimentation for the evaluation of the MAP-host cell interactions. These assays can contribute with information regarding pathogenesis and initial notions on vaccine efficacy. In any case, the most relevant models found in the literature are further described in the following sections.
3.1. Ex vivo models
Ex vivo means “outside of a living body” implying that cells or tissues have been previously isolated from an animal and maintained in culture conditions for the study. Therefore an in vitro assay involves an established cell system such as a cell line, whereas an ex vivo assay is based on cells or tissues from a living organism, this is, primary cells. In the same way, the level of cellular complexity will be superior in an ex vivo assay being closer to the organism’s natural conditions and therefore offering advantages. Although higher variability will be observed due to the use of more than one individual, it will be closer to reality, specially regarding PTB, a disease in which many animals are not susceptible, not developing clinical signs despite being MAP carriers at some point of their lives.
These models can be based on cells isolated from tissues and/or whole blood, or on tissue slices from different organs, conserving the original cell structures and intercellular connections. In all cases, treated or untreated, healthy or diseased animals can be used as donors, increasing the complexity of the resulting information from in vivo experiments. In PTB research different ex vivo models have been used.
3.1.1. Intestinal models
Ex vivo models using intact intestinal tissue have been developed, such as the culture of intestinal explants obtained by biopsy, first described in 1959 [47], and the everted sleeve method [48]. The use of both has been explored in the context of PTB research: Schleig and colleagues [49] showed that explant cultures of different intestinal sections are suitable models to study early events of MAP invasion, such as the cellular attachment process. In the same year, Sigurdardottir and colleagues demonstrated that MAP invasion occurs in small intestinal areas with and without Peyer's patches using the everted sleeve method [50]. More recently it has been described that intestinal explants can be stimulated ex vivo inducing immune responses to a wide range of stimuli, detecting changes in expressed cellular phenotypes as well as in the levels of biomarkers of inflammation [51]. We believe that these models could be useful to study differences in MAP pathogenesis on primary cultures obtained from animals with PTB and different degrees/types of intestinal pathology versus healthy control animals.
The simplest intestinal model could be considered the primary culture of intestinal epithelial cells. Primary cultures of small or large bovine intestinal epithelial cells (BIEC) have been used to evaluate the effects of virulence factors of other microbes, toxic compounds and the innate immune responses through Pattern Recognition Receptor (PRR) signaling. Li et al. [52] used BIEC obtained from neonatal bovine ileum to evaluate the effect of Saccharomyces cerevisiae components on MAP adherence. Nevertheless, these cell suspensions might be contaminated with non-epithelial cells (mostly fibroblasts) and therefore, a series of purification steps would be required to obtain relatively pure BIEC [53]. The main constraints to overcome primary cell culturing are that they normally enter into a non-dividing state after a few passages and also the possible variability between donors. However this system would be valuable to evaluate MAP intestinal invasion and early pathogenesis.
3.1.2. Immune cell models
Macrophages are considered key players in PTB since these immune cells act as reservoirs providing a favorable environment for MAP replication. Once phagocytosed by macrophages, MAP is able to arrest phagosome-lysosome fusion, therefore inhibiting acidification of the cellular compartment and hampering pathogen destruction [54]. MAP can control monocyte and macrophage apoptosis feeding the infection cycle [55]. Other phagocytes that have been less studied, probably because they have not been considered as important cell types in MAP pathogenesis, are neutrophils.
Macrophages and monocytes
Macrophages play a critical role in the host-pathogen interaction of PTB. Actually, they have a dual role, being effector cells that can mediate both the destruction of MAP and its survival, proliferation and dissemination after the pathogen inhibits phagolysosome maturation. Monocyte-derived macrophages (MDMs) generated from peripheral blood mononuclear cells (PBMCs) have been widely used to model macrophages for ex vivo studies. As for PTB, MDMs have been extensively used to study MAP pathogenesis, screen for MAP mutants, study the immune response (phagocytosis, growth and cytokine production) against MAP infection and in functional assays to aid in vaccine prototype evaluation or to complement specific resistance/susceptibility related host trait findings.
MDMs have been infected with MAP alone or in co-culture with other cell types. In this section we will discuss studies involving MDMs cultured alone, whereas MDMs co-cultured with other primary cells or cell lines are discussed in the co-culture section.
Regarding methodology for monocyte isolation and enrichment, the majority of studies include protocols that isolate PBMCs first and select for adherent cells on plastic culture plates for subsequent differentiation into macrophages. A few works include protocols where MDMs have been obtained after monocyte isolation from PBMCs using an anti-CD14 antibody coupled to magnetic microbeads [56,57,58]. Arteche-Villasol and collaborators [58] found that the magnetic separation yielded a higher proportion of monocytes from the PBMC fraction than the plate method, although the latter achieved higher yields of MDMs than those isolated through the former, in sheep and goats. It is worth mentioning that the monocyte isolation techniques not only affect yield and purity, but also impact the resulting phenotype of cultured cells [59]. Temperature is another important factor to consider as it was demonstrated that physiological temperatures greatly influence MAP gene expression profiles and speed of macrophage invasion [28]. Many groups have further contributed with methodology that can aid the evaluation and quantitation of the bacterial intracellular growth and that can be complementary to MAP infection assays in MDMs. Mitchell et al. [60] showed that real-time qPCR assays provide a more accurate and precise method for evaluating MAP intracellular growth dynamics in MDMs to study strain differences compared to fluorescent quantitation. Also, confocal microscopy has been suggested as a valuable tool to quantify MAP in MDM infection assays [61].
The first studies performed with electron microscopy showed that MAP was not degraded four weeks after infection of MDMs [62] indicating that MAP could actually survive inside macrophages. Also, demonstrated was the fact that both monocytes and MDMs required the presence of serum in order to efficiently phagocytose MAP and that MDMs showed higher phagocytosis levels than freshly adherent monocytes [63]. Further on, Khalifeh and collaborators found that IL-10 and TGF-β inhibited the destruction of intracellular MAP in the presence of IFN-γ in MDMs from naturally infected cows compared to MDMs from healthy animals, suggesting important immune regulatory roles for these cytokines during infection with MAP [33]. Afterwards, Weiss and collaborators, using a combination of gene expression studies and functional assays, suggested that the inhibition of phagolysosomal function and apoptosis and the expression of inhibitors of macrophage activation were probably important factors behind the survival of MAP in bovine macrophages [64]. The same group later studied the role of IL-10 showing that neutralization of this cytokine enabled macrophages to kill 57% of MAP organisms within 96 hours along with an increase in acidification of phagosomes, apoptosis of macrophages, and production of nitric oxide (NO); suggesting that induction of IL-10 expression by MAP could be considered a virulence factor [65]. All these studies were important to gain knowledge on MAP-macrophage interaction and the role of IL-10 in PTB and ultimately set the conditions for future assay
Also regarding pathogenesis elucidation, Khalifeh and coworkers evaluated the role of NO production in MAP viability within macrophages, by culturing MDMs from subclinically or clinically infected cows and healthy controls, observing that NO production was highest in the subclinically infected cows and that it peaked when cells were co-stimulated with MAP and IFN-γ [66]. Authors hypothesized that in the transition from subclinical to clinical stage an increase in the bacterial load of the cells would cease IFN-γ upregulation and lead to an impairment of NO generation.
MDM ex vivo assays have also been used to generate knowledge for novel vaccine and therapy design, either by gaining knowledge of cell-MAP interaction at the molecular level or by treating animals and performing ex vivo functional assays for evaluation afterwards. Among many studies, it has been shown that inhibitors of mitogen-activated protein kinases (MAPK) such as MAPKp38 increase production of nitric oxide after MAP infection [67], that vitamin D3 analogs promote phagocytosis of MAP by MDMs [68], and that antibodies directed against lipoarabinomannan (LAM) used to opsonize MAP prior to MDMs contact increase macrophage apoptosis and TNF-α secretion [69]. Vaccination has shown to enhance microbicidal activity of macrophages against MAP, as seen by a significant reduction of MAP viability as well as upregulation of iNOS and IL-10 in infected MDMs from vaccinated animals compared to non-vaccinated animals [70]. All these studies have shown that different therapies can enhance macrophage functions at least in ex vivo conditions.
Studies focused on host genetics and MAP strain differences have also been performed using MDMs aiming at identifying markers that can be exploited for therapy or diagnosis. Actually, the possibility of finding strain differences in MDMs has been exploited to screen attenuated [57,71] and transposon mutants [72] in bovine MDMs looking for novel vaccine candidates. Some studies have failed to find differences in functionality between MDMs of PTB positive and control animals but have found important strain differences [73], whereas other studies have shown that MDMs with certain SNPs (TLR2-1903 TT genotype) produce higher levels of IL-12p40 and IL-1β when stimulated with MAP compared to cells derived from other genotype (TLR2-1903 CT and CC), suggesting these SNPs for marker-assisted breeding strategies as an additional tool in PTB control strategies [74]. The importance of host genetics has been further demonstrated in studies that showed that the ability of MDMs to limit MAP viability is associated with specific genetic profiles [75]. Many studies have identified genetic variants in regulatory pathways of the macrophages that may affect the susceptibility of cows that are healthy/resistant to MAP infection [76,77]. All these studies support the use of genomic and transcriptomic approaches to enable the identification of markers associated particularly with susceptibility to MAP infection.
In summary, MDM models have been and are very useful to study pathogenesis focused on the MAP-macrophage interplay. However, we must bear in mind that they are insufficient in providing accurate information on the events that take place during natural infection, as shown by the analysis of MAP transcriptome from tissues of naturally infected cows or from in vitro infected macrophages [78].
Granulomas
The granuloma has been considered as a hallmark of mycobacterial infection and the evaluation of factors that affect its development and evolution is complex. A model for the in vivo dynamics of bovine paratuberculosis granulomas has been proposed by Koets et al. [79]. In an effort to simulate the earliest events in the immune response leading to tuberculous granuloma formation, a number of in vitro models have been developed (reviewed in [80]). In the field of PTB, early interaction and coordination of macrophages upon MAP infection has been studied starting from MDM in vitro cultures that were kept for 10 days in order to form granulomas [81,82]. In the first study, Multiplicity of Infection (MOI) of 1:8, 1:16, and 1:33 were assayed, finding that the highest number of microgranulomas was achieved at the lowest MOI, 1:8 [81]. In the second study, authors found that at MOI 1:2 and below, macrophages displayed increased longevity compared to uninfected cells and formed clusters that secreted pro-inflammatory cytokines necessary for a cell-mediated immune response [82]. While, at higher MOIs, the viability of host MDMs was negatively impacted and intracellular MAP reproduced over the first five days of infection.
Since maintaining granuloma structures in vitro over an extended period of time is still a limitant of this model [83], only features regarding the early stage of granuloma formation can be studied. Although it has been proposed that granulomas are marked by the interaction of various immune cell types, which acquire different phenotypes in it, as is the case of M1/M2 polarization of macrophages, to date, only small and simple granuloma-like structures based on MDMs have been evaluated as a model of PTB. Such a model, although still lacking the degree of realism and cellular complexity achieved in vivo, could be refined and provide in the near future information related to relevant aspects of PTB pathogenesis: mechanisms involved in granuloma induction and maintenance, activation of MAP latency genes [84], role of different leukocyte subpopulations, characteristics of the granuloma that favor bactericidal functions versus bacterial persistence [85].
Neutrophils
Several studies have shown that neutrophils can play an important role in the innate immune protection against mycobacterial infections [86,87]. Although neutrophils are absent in lesions of advanced stages of MAP infection, their presence at infection sites in early phases has been reported [88]. Furthermore, neutrophils seem to assist in the induction of specific Th1 and Th17 cells in response to a tuberculosis vaccine [89]. In any case, their possible role in defense against MAP has been evidenced in transcriptomic studies that describe the impairment of neutrophil recruitment and activation during PTB [90,91,92]. These findings have recently directed the focus on neutrophils leading to studies that have looked into the behavior of these immune cells against MAP in ex vivo assays.
Neutrophils kill microbes through a set of mechanisms that include phagocytosis, reactive oxygen species (ROS) production, degranulation and extracellular trap (ET) release [93]. Neutrophils isolated from healthy cattle have shown ET liberation against MAP and effective killing in vitro [94]. Caprine neutrophils have also demonstrated a strong innate response against MAP, using their entire repertoire of effector functions, ET release, degranulation, chemotaxis and phagocytosis [95]. Vaccination against PTB in rabbits increased the phagocytic activity of these immune cells along with their ability to release ET against mycobacterial and non-mycobacterial agents [26]. In this study, the degree of protection of the different vaccine prototypes was correlated to the ex vivo antimicrobial activity of neutrophils, indicating that this cell type may play a role in protection.
Neutrophils are tricky cell types to work with and functional assays must be performed with freshly isolated cells making this model less attractive than monocytes or macrophages. The number of protocols for isolation, culture and evaluation of functionality is continuously growing and are largely dependent on the host species [93]. Improvement of the methodology will probably nurture further research on this phagocytic cell and its role in PTB.
3.1.3. Co-cultures of primary cells alone or combined with cell lines
Co-cultures (a culture of more than one cell type) can be an option to study interactions between different cell types. These kinds of models can give information on crosstalk between the cell types evaluated and also on host-pathogen interactions. Therefore, most ex vivo assays described in the literature have focused on macrophages, alone or in co-culture with other cell types, including lymphocytes [96,97], PBMCs [98], neutrophils [94] or even epithelial cell lines [99,100]. The objective of each of these co-culture models varies depending on the cell type and on the origin of the cells (vaccinated, healthy or MAP infected animals).
Hostetter et al. [101] found that mycobactericidal functions of MDMs were enhanced in MAP infected cattle and that the addition of autologous CD4+ T cells did not increase bacterial killing. However, CD4+ T cells from non-infected animals did increase bacterial killing in autologous macrophages. The same group tested the influence of γδ T cells on MDMs in vitro finding that these cells from MAP sensitized animals are not able to produce enough IFN-γ in order to enhance mycobacterial killing or nitrite production by infected macrophages [102].
Co-culturing MDMs with WC1+ and WC1- γδ T lymphocytes has shown that these lymphocyte subsets modulate effector functions of MDMs such as MAP killing ability [96]. Furthermore, Pooley and co-workers [97] have developed a mycobacterial growth inhibition assay based on a monocyte-lymphocyte co-culture model and have used it to evaluate vaccination on sheep. They conclude that this system can quantify the ability of PBMCs to kill MAP, serving as a predictor of vaccine failure or non-response, being potentially useful for novel vaccine screening. Similarly, Davis and co-workers have developed ex vivo platforms in cattle to study the functional activity of CD4+, CD8+ and γδ T and NK cells stimulated with MAP antigen-primed antigen presenting cells (dendritic cells (DC) present in blood, monocyte-derived DC, and monocyte-derived macrophages) [57,103,104,105]. Authors highlight the usefulness of these platforms to assess bacterial viability, cells’ phenotype, cytotoxicity mechanisms and, in consequence, to examine the potential role of the cell-mediated immune response in preventing establishment of a persistent infection of MAP [106].
In the same way, another interesting ex vivo assay that complements an in vivo model is that based on co-culture of MDMs with monocyte deprived-PBMCs that have been previously primed or not with MAP-stimulated DC. This assay showed that macrophages from animals vaccinated with prototype vaccines, cultured alone or in co-culture with unprimed PBMCs showed increased MAP killing ability. This was in agreement with a more effective clearance of MAP infection from calf tissues in vivo [98].
The co-culture of MDMs with autologous neutrophils isolated from healthy cattle was performed to study the interaction of both cell types with MAP and Mycobacterium bovis alone or in co-culture showing cooperation and synergistic effects against mycobacteria [94]. In this work, co-cultures were performed putting both cell types in contact in the same culture well or separately in a transwell system, sharing nutrients and metabolites but avoiding physical contact. ET release and bacterial killing were evaluated along with cytokine secretion offering information on the mechanisms that operate in the defense of these cell types against MAP.
Finally, using a permeable support system, Lamont and coworkers [99] were able to artificially construct an epithelial cell layer with the bovine mammary epithelial cells (MAC-T cell line), along with bovine MDMs immediately underneath to represent subepithelial macrophages. Thus, this system aims at mimicking the MAP epithelial invasion step prior to uptake by macrophages and could answer questions about the interaction between the bacteria and MAC-T cells and the impact of this one on macrophages [99,100]. This co-culture system has shown that MAP leads to phagosome acidification in the epithelial cells and IL-β release that helps for an efficient epithelial transverse ending in macrophage uptake [99]. Further studies using this model have permitted a deep characterization of early MAP infection identifying metabolic, DNA repair and virulence genes that can be considered as novel drug targets for pathogenesis studies [100].
3.1.4. Organoids
Organoids are three-dimensional organ-like structures composed of functional, stem cell-derived multicellular aggregates that can be indefinitely propagated to resemble a specific organ when cultivated with essential niche factors. Intestinal organoid culture was first developed in mice and humans following the identification of signaling pathways involved in the maintenance and proliferation of fast-cycling intestinal stem cells that expressed high levels of leucine-rich repeat-containing G protein-coupled receptor 5 (Lgr5) expresses the cell surface marker [107]. In these species, organoids can be obtained from adult intestinal epithelial stem cells (Lgr5+ IESC), from adult reprogrammed stromal cells or induced pluripotent stem (iPSC) cells, or from embryonic stem cells (ESC). In the case of ruminants and other farm animals, although advances produced recently in the field of development of methodologies for obtaining ESC and iPSC [108,109], to the best of our knowledge, only methods for obtaining organoids from IESC have been described. Since the organoids are generated from stem cells, they have the capacity to generate crypt-like domains, with the potential to differentiate into many of the heterogeneous epithelial cell types, contrary to transformed or immortalized cancer-derived cell lines, and spontaneously organize into tissue-like structures that reflect characteristics of the digestive segment of origin [110]. The methodology for obtaining and working with enteroids is beyond the scope of this review and can be consulted in recent literature [111].
Protocols for culturing ruminant organoids from different digestive segments have been set out [112,113,114,115,116,117,118,119,120]. Since the first report in 2009, intestinal organoids have evolved as a potential alternative to in vivo models for various experimental purposes, such as imaging, molecular analysis and gene editing, but also as reductionist approaches to study the interaction of epithelial cells with other relevant actors in the physiological context, such as immune cells [121,122] or microbiota [123,124] or even to evaluate the early host-pathogen interaction in the context of many infectious diseases (protozoa: [125,126]; virus: reviewed in [127]; bacterias: [128,129,130,131,132,133]).
Studies on animal organoids have gained more attention in various fields, including veterinary medicine, given recent advances in organoid technology (reviewed in [134]). In the field of PTB studies, the first steps in this direction are just beginning to be taken [119]. In that recently published work, a 3D bovine enteroid model is developed, with the apical side exposed on the exterior surface, enabling infection of the organoids without need of microinjection and concluding that this model was permissive to infection with MAP and therefore useful in investigating early stages of MAP pathogenesis. In the near future, this model could certainly be used to shed light on other aspects of pathogenesis such as, cell tropism, viral factors, biological differences among MAP genotypes, exploring new signaling pathways targetable in disease prevention, thus facilitating the discovery of interactions between mycobacteria and host cells in a more physiological environment. Even so, these experimental systems still bear some limitations, namely the lack of immune cells and other types of cells normally present in the architecture of the tissue. So a next generation of these organoids, thats includes co-culture with different cells of the immune system [122,135] could be very useful in the context of research on the immunopathogenesis of the infection and new vaccine candidate evaluation, by studying the fate of MAP infection in organoids co-cultured with immune cells, both obtained from vaccinated versus unvaccinated animals. Organoids co-cultured with macrophages could provide new evidence on the role of the M1/M2 dichotomy in the context of this infection. On the other hand, genetic markers associated with phenotypes of susceptibility or resistance to PTB have been described (reviewed in [136]) as well as an additional intermediate category of “tolerant hosts” [137]. Taking into account that organoids maintain the individual genetic fingerprint, the usefulness of this model for gaining knowledge on the internal differences between those phenotypes is raised.
Among the early events of MAP infection, the bacterial interaction of MAP enterocytes, M cells [138,139,140] and goblet cells [141] on the lining epithelium has been identified. Similarly, organoids obtained from different intestinal segments have been used to study the regional differences detected in invasion and response to MAP infection in ileum versus jejunum [142,143,144].The use of intestinal organoids could be further used to deepen the knowledge on these MAP-cell interactions.
3.2. In vitro models
Immortalized cell lines grown as monolayers are often used in research instead of primary cells. They offer several advantages, such as eliminating donor to donor variation, cost effectiveness, easiness in use, provision of an unlimited supply of material and offering a bypass to ethical concerns associated with the use of animal tissues. As pure cell populations, cell lines allow for less experimental variability and greater reproducibility of results. Since their advent in the 1950s, cell lines have revolutionized scientific research and have become a central workhorse of biomedical research. However, despite being a powerful tool, one must be careful when using cell lines. Cell lines should display and maintain functional features as close to primary cells as possible. This may be particularly difficult to determine as often the functions of the primary cells are not entirely understood. Since cell lines are genetically manipulated this may alter their phenotype, native functions and their responsiveness to stimuli. Serial passage of cell lines can further cause genotypic and phenotypic variation over an extended period of time and genetic drift can also cause heterogeneity in cultures at a single point in time.
3.2.1. Macrophage cell lines
Ruminant macrophage cell lines have been widely used in order to evaluate the interaction of MAP with its target cell. BOMAC, derived from peritoneal macrophages transfected with SV40 plasmid DNA [145], is the most reported one. Although it has been proposed that BOMAC cells are inherently dysfunctional, lack several receptors, dissimilar to bovine primary culture macrophages and posses an insufficient capability to phagocytose MAP [146], different groups have reported comparable results regarding MAP interaction of this cell line with respect to bovine MDM [70,147] and valuable data were obtained from this model. Even more, less assay-to-assay variation in the level of MAP infection in BOMAC compared to MDM cells has been reported [148,149]. Studies of metabolism and virulence of MAP have been done on BOMAC [149,150]. The modulation by bovine antibodies directed to mycobacterial antigens or by Saccharomyces cerevisiae components on BOMAC cells in response to MAP infection were analyzed by evaluating translocation, phagocytic activity, ROS production, immune-related gene expression, cytokine secretion and apoptosis [69,151,152,153,154,[69,151–154,]. The use of another ruminant macrophage cell line, MOCL-4, that had been established from spontaneously-proliferating adherent mononuclear cells from sheep blood has also been evaluated as a model for MAP cellular infection [150].
Murine macrophage cell lines have long been used to elucidate host-mycobacteria interactions and intracellular growth characteristics of MAP [54,155]. RAW 264.7 is a murine macrophage cell line which is extremely sensitive to stimulants [156]. Despite this drawback, several studies have utilized this cell line for studying nitric oxide production in response to mycobacterial infections [157,158] and have reported differences in the immune responses and gene expression elicited between non pathogenic and pathogenic mycobacteria [159,160,161,162]. In addition, Everman et al. [153] have used this model to evaluate the effect of antibodies in opsonize MAP. Thirunavukkarasu et al. [163] have also utilized RAW 264.7 cells as a model to examine a series of macrophage activation parameters in response to MAP, a non-pathogenic mycobacterium, as well as mycobacterial antigens. This research demonstrates the usefulness of this model in the study of mycobacterial immunopathogenesis. Even more, this system has been featured for selection of attenuated vaccine candidates [164].
The phagocytic J774.A1 is another murine macrophage cell line that was used to evaluate macrophage activation, phagosome maturation, cytokine production, stimulation of CD4+ T cells and intracellular survival of MAP [55,155,165,166], and to study the antimicrobial or immunomodulatory effect of molecules on the ability of macrophages to clear MAP infection [167,168].
THP-1 is a human leukemia monocytic cell line that can be differentiated into a macrophage-like phenotype by stimuli such as phorbol-12-myristate-13-acetate (PMA) [169]. This cell line has been evaluated by several authors with the aim of shedding light on molecular mechanisms in host-pathogen interactions, mainly focused on the relation of MAP with human Crohn's disease [170,171,172,173,174]. The use of the THP-1 model has also provided new information in the context of PTB pathogenesis [175]. Motamedi Boroojeni et al. [176] have evaluated THP-1 derived macrophages as a model to evaluate the immunogenicity of constructed Salmonella expressing MAP-candidate vaccine genes.
Another cell line, U-937, exhibiting monocyte morphology from malignant cells of human origin, was differentiated into macrophage-like cells with PMA and used to evaluate MAP infection [177].
In sum, macrophage cell lines from different origins (bovine, ovine, murine, human) have been used over time to provide valuable studies on different aspects of MAP-host interaction at the cellular and receptor-ligand level, biological differences between MAP genotypes, the importance of different bacterial virulence factors and also from a therapeutic perspective [154,168].
3.2.2. Epithelial cell lines
The intestinal epithelium is the largest surface that acts as a primary barrier against pathogens. Mycobacterial invasion of intestinal epithelial cells is a complex event, requiring participation of several bacterial and host factors. Initially, the role of microfold epithelial cells (M cells) in MAP intestinal uptake was highlighted [178] and afterwards, it was learned that invasion also occurs through other cell types, in areas with or without Peyer's patches [139] and the use of epithelial cell lines has greatly contributed to this knowledge [179,180].
Most of the studies on MAP and in vitro epithelial cell models have been conducted using intestinal epithelial lines of murine origin or bovine epithelial cell lines of organs other than intestine. Different groups have been working on obtaining bovine intestinal cell lines models [181,182,183,184,185]. More recently Katwal et al. [186] have developed and characterized an early passage and immortalized BIEC line obtained from the ileum of a 2 day old calf. These cells expressed TLR 1–10 as well as cell surface sugars relevant to the host-pathogen interaction [186,187]. The use of these epithelial models could surely improve the knowledge about the early interface of MAP intestinal invasion.
Additionally, some epithelial models could clarify the role of the interaction of MAP with different epithelial cell types at the intestinal mucosa. The M cell differentiation from BIECs has been succeeded by co-culture with murine Peyer’s patch lymphocytes [188,189] or treatment with supernatant from bovine PBMC cultured with IL-2 [181]. In the field of mycobacterial infections, the model proposed by Kerneis et al. [188], has been used concomitantly with other mouse models to prove that the translocation by M cell is a vital entry mechanism that contributes to the pathogenesis of Mycobacterium tuberculosis [190]. This model would also be valuable for studying early MAP pathogenesis. Unexpectedly, despite all these reports, there is no bovine intestinal epithelial line available in Culture Collections. Nevertheless, other non-intestinal cell lines have been used as models for early events in MAP invasion: Bannantine et al. [191] have first used Madin-Darby bovine kidney epithelial cells (MDBK) as a model of bovine intestinal mucosa. In addition, Patel et al. [192] have described that MAP was capable of infecting confluent monolayers of MDBK cells. Since epithelial cells are not phagocytic in nature, MAP invasion of MDBK cells indicates that the bacteria might trigger their own uptake, probably by inducing cytoskeleton reorganization [192]. This model was used to evaluate the relevance of MAP lipids and proteins on pathogenesis [193,194] and the role of specific antibodies in MAP invasion, translocation and cell-mediated killing [153]. Likewise, Everman et al. [195] have developed an interesting cell culture model that simulates the passage of bacteria from their uptake by the intestinal epithelium (modeled by a primary infection of MDBK monolayer infection), spread to the tissue phagocytes (mimicked by the infection of RAW 264.7 cell line with MAP obtained from MDBK lysates), and ultimate return to the intestinal epithelium during the later stages of infection (modeled by a secondary infection of MDBK with MAP recovered of the intracellular compartment of RAW 264.7). This model offers an alternative to study both the host and bacterial mechanisms used during the invasion and infection process of the ruminant intestine, and discover unidentified changes or interactions that occur during the disease. For example, this research group has performed a gene expression analysis and detected different profiles of immune signals as well as bacterial phenotypes between the primary and secondary infected MDBK.
MAP has been reported to infect mammary tissue [196,197]. Various studies have demonstrated that MAP can infect the MAC-T cell line from both the apical and basolateral surfaces with comparable efficiency- and survive inside these cells, causing an impact on their gene expression [192,193,198]. Moreover, Patel et al. [192] have described that prior incubation of MAP in MAC-T cells enhanced the efficiency of invasion of MDBK cells, hypothesizing that MAP present in milk could have an invasive phenotype enhancing infection of the suckling calves. This model was also used to evaluate co-infection processes between MAP and other pathogens as Escherichia coli, Staphylococcus aureus or Streptococcus agalactiae and has demonstrated rapid baso-apical translocation of MAP in epithelial cells [199] and fast internalization of S. aureus in MAC-T cells previously infected by MAP [200]. Knockout variants for TLR 4 or IL-10Rα of this model have been recently used to demonstrate the role of those receptors in the regulation of the innate response to MAP [201,202].
3.2.3. Co-culture of cell lines
The co-culture of different cell-lines constitutes the simplest manner to include some elements of the microenvironment in the model as an over-simplification of the complex in vivo crosstalk of different cell types in a tissue context. Even so, such in vitro cell culture systems still offer more controllable, versatile and reproducible setups compared with in vivo and ex vivo systems. To our knowledge only two research studies using cell lines in co-culture related to PTB have been published. These studies allowed to increase the understanding about the modulation of the CD4+ T cell-stimulatory capacity of MAP infected macrophages [165] and the ability of MAP to stimulate the synthesis of integrins in macrophages, permitting efficient attachment and translocation through the endothelial layer [203].
Thus far we have reviewed and discussed different experimental approaches used in PTB research. Table 2 summarizes the analysis carried out in this work.
4. Modeling PTB: future perspectives
The pressure to avoid animal experimentation and search for alternatives is growing. Added to this, the in vitro alternatives that are based on cell lines present limitations that have been mentioned throughout this work. In this context, other approaches are possible. Following are models that we think are worth considering in the future for PTB research.
4.1. Nematode model
The nematode Caenorhabditis elegans can be infected by a wide variety of bacterial species, including mycobacteria, and has therefore been used for the last years as a model for studying some aspects of host-pathogen interaction. Even when the absence of a clear macrophage cell lineage is posed in C. elegans [204], evidences from studies of different intracellular pathogens, such as Legionella pneumophila [205], indicate C. elegans as a host allowing analysis of specific virulence mechanisms, of relevance in the pathogen-macrophage interplay. Although no study has been performed to date using this model for MAP studies, several authors have reported positive experiences using this model to evaluate other mycobacteria of pathological interest, such as Mycobacterium avium subsp. hominissuis (MAH) [206,207] and Mycobacterium marinum [208]. Interestingly, host defense molecules such as ROS and lysozymes, produced in response to bacteria, are delivered to the lumen of the worm intestine [209,210], rather than to an endosomal compartment, as would occur within macrophages. It should also be noted that the intestine of these worms is covered with an epithelium quite similar to that of the mammalian intestine, sharing comparable morphology, structure, and function. In those experiments evaluating C. elegans as a model in the pathogenesis of mycobacteria, the worms are fed with them and the infection occurs at the intestinal level.
Galbadage et al. [211] have demonstrated that C. elegans has particular value for studying the role of MAPK pathways in mycobacterial pathogenesis. Bermudez et al. [207], tested the pathogenicity of different adherence-deficient clones of MAH in C. elegans and found that all of them had altered ability to colonize this host. In the light of these results, C. elegans appears as a promising genetically tractable pathogenicity model of both, the host and the pathogen. It would be interesting to test the capacity of MAP to infect this nematode causing pathology, as a first step toward considering this model in pathogenesis studies as well as in the preliminary selection of attenuated vaccine candidates for PTB.
4.2. Organs-on-a-chip
Other advanced culture systems such as organoid-derived 2D monolayer [212] and organ-on-a-chip systems have been emerging to overcome the weakness in current organoid systems, such as their static nature, without media flow or application of mechanical strain. These advanced culture platforms offer great potential to widen the scope of research through dissociation of 3D organoids into single cells and their integration into advanced culture systems [213,214]. Organs-on-a-chip are self-contained and modular in vitro models with easily controllable features, involving microfluidic approaches coupled with cell culture.
Regarding PTB, both gut-on-a-chip and immune system-on-a-chip can be attractive models. Gut-on-a-chip is fairly advanced in human medicine and has been used to study the Inflammatory Bowel Disease [215], a gastrointestinal inflammatory disorder that shows similarity to PTB. Recent work has described the study of host-pathogens interactions in the context of intestine-on-a-chip models [216]. Studies based on human primary intestinal cells have shown that these systems have a closer transcriptomic profile and functionality to the intestine in vivo, when compared with other in vitro models. So, intestine-on-a-chip models appear as a promising tool for further research in the PTB field [134,217]. As for immune cells or immune organs on-a-chip, these have been recently reviewed [218] and regarding the potential application of these models in PTB, it seems that the lymph-node-on-a-chip and the inflammation-on-a-chip can be options that should be explored.
4.3. Precision cut intestinal slices (PCIS)
Ruminant animal model experiments require huge budgets and appropriate facilities, as mentioned before. Although laboratory animal models can support PTB research, sometimes the differences in results between these models and the target species can hamper translation. Added to this, there is an increasing pressure to avoid animal experimentation in any format and search for alternatives. The in vitro alternatives based on cell lines present limitations that have been pointed out such as lacking in vivo characteristics: like fluidic flow or periodic peristalsis of limited crosstalk between host and microorganism and between different cell types. In this context, there is a clear need for alternative models with higher complexity that can better resemble the disease in the natural host. As for PTB, the source of the intestine can be animals that go to slaughter, taking advantage of tissues that would otherwise be residues. Precision-cut tissue slices (PCTS) should be explored as it has been described as an invaluable model providing the cellular and structural complexity of the host tissue [219].
Precision-cut intestinal slices (PCIS) has been described as a novel ex vivo model to study transport, metabolism and toxicology of drugs [220]. This technique was first reported in 2005 by de Kanter et al. [221] using rat small intestine and colon. The intestinal slices are obtained by embedding excised intestinal tissue in agarose and cutting very thin sheets (200-400 μm) while submerged in an oxygenated buffer that guarantees cell viability using a specialized microtome. PCIS can be incubated and remain viable for up to 24 hours permitting the study of solute transport and metabolism studies.
Although PCIS has been used for decades to study pharmacological and toxicological effects, the use of this technique to assess host-pathogen interactions has not been fully developed and literature shows far less studies than precision-cut slices of tissues of other origins such as lung. Actually, precision-cut lung slices (PCLS) have been used to evaluate antimycobacterial agents against Mycobacterium abscessus in mice [222] and to study the early lung response to infection by M. bovis and Mycobacterium tuberculosis in cattle [223]. Up to date PCIS has been used to study viral infections such as avian influenza [224], coronavirus [225] and hepatitis C [226] but bacterial infections have not been assessed. This is probably due to technical issues that must be resolved and that can include the optimization of the culture medium and the development of preservation techniques that require more research as pointed out by Li and collaborators [220].
In the near future, PCTS will be an invaluable model to assess host-pathogen interaction and response to vaccines in a “whole organ” mode [219] and hopefully PCIS can be used to study PTB pathogenesis and evaluate vaccine candidates and/or therapies, as well.
5. Conclusions
PTB control has been difficult due to the long incubation period of the disease and failures both in diagnosis and in immunoprophylaxis. This review has been intended to give a view of the wide content of experimental models used to study PTB. Having standardized experimental infection models will aid in a deeper understanding of this pathology and benefit the development of control tools. So far, laboratory animal and cell based models have contributed to increase knowledge on MAP pathogenesis, host-MAP interaction and associated genetic traits. Furthermore, these models have contributed to initial phases of therapy evaluation as well as in the testing of novel vaccine candidates. In years to come, advances in omics approaches and their combination with the models described herein or even with novel tissue engineering techniques will certainly contribute to the development of new tools in the context of PTB research.
Author Contributions
Conceptualization, A.J., B.F., S.L.M. and N.E.; methodology, A.J., B.F., S.L.M. and N.E; writing—original draft preparation, A.J., B.F., S.L.M. and N.E; writing—review and editing, A.J., B.F., S.L.M. and N.E; funding acquisition, N.E., S.L.M. and B.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agencia Estatal de Investigación, Ministerio de Ciencia e Innovación of Spain with additional funding by FEDER of the European Union, grant number PID2021-125807OB-C22, by Agencia Nacional de Promoción Científica y Técnica of Argentina BID-PICT-2018-03595), and by Universidad de Buenos Aires of Argentina (UBACyT 20020190200158BA).
Data Availability Statement
No new data were created. Results of research engines run to obtain the material that has been reviewed in this work can be shared upon request to the authors.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Stabel, J.R.; Bannantine, J.P.; Hostetter, J.M. Comparison of sheep, goats, and calves as infection models for Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2020, 225, 110060. [Google Scholar] [CrossRef]
- Hines, M.E., 2nd; Stabel, J.R.; Sweeney, R.W.; Griffin, F.; Talaat, A.M.; Bakker, D.; Benedictus, G.; Davis, W.C.; de Lisle, G.W.; Gardner, I.A.; Juste, R.A.; Kapur, V.; Koets, A.; McNair, J.; Pruitt, G.; Whitlock, R.H. Experimental challenge models for Johne's disease: a review and proposed international guidelines. Vet. Microbiol. 2007, 122, 197–222. [Google Scholar] [CrossRef]
- Talaat, AM. ; Wu; C, Hines, ME2nd. Chapter 16: Experimental animal models of paratuberculosis. In Paratuberculosis: Organism, disease and control, 2nd ed; Marcel A Behr, Karen Stevenson, Vivek Kapur. CABI, Boston, MA, 2020; pp. 213–247. [Google Scholar]
- Cooney, M.A.; Stelle, J.L.; Steinberg, H.; Talaat, A.M. A murine oral model for Mycobacterium avium subsp. paratuberculosis infection and immunomodulation with Lactobacillus casei ATCC 334. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef]
- Veazey, R.S.; Horohov, D.W.; Krahenbuhl, J.L.; Taylor, H.W.; Oliver, J.L. 3rd; Snider, T.G. 3rd.Comparison of the resistance of C57BL/6 and C3H/He mice to infection with Mycobacterium paratuberculosis. Vet. Microbiol. 1995, 47, 79–87. [Google Scholar] [CrossRef]
- Shin, S.J.; Wu, C.; Steinberg, H.; Talaat, A.M. Identification of novel virulence determinants in Mycobacterium paratuberculosis by screening a library of insertional mutants. Infect. Immun. 2006, 74, 3825–33. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Cui, N.; Tang, Y.; Chen, F.; Cui, Y.; Dang, G.; Liu, S. A candidate subunit vaccine induces protective immunity against Mycobacterium avium subspecies paratuberculosis in mice. NPJ Vaccines 2023, 8, 72. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.T.; Shim, S.; Kim, S.; Woo, S.H.; Kim, D.Y.; Yoo, H.S. Immunopathological mechanisms in the early stage of Mycobacterium avium subsp. paratuberculosis infection via different administration routes in a murine model. PLoS One 2023, 18, e0281880. [Google Scholar] [CrossRef]
- Hussain, T.; Zhao, D.; Ali Shah, S.Z.; Wang, J.; Yue, R.; Liao, Y.; Sabir, N.; Yang, L.; Zhou, X. Responses of Murine Macrophages Infected by Mycobacterium avium subspecies paratuberculosis by Targeting Interleukin-10 and TGF-β-Activated Protein Kinase 1 Binding Protein 2. Front. Immunol. 2018, 8, 915. [Google Scholar] [CrossRef]
- Hussain, T.; Zhao, D.; Shah, S.Z.A.; Sabir, N.; Wang, J.; Liao, Y.; Song, Y.; Dong, H.; Hussain, M.M.; Ni, J.; Yang, L.; Zhou, X. Nilotinib: A Tyrosine Kinase Inhibitor Mediates Resistance to Intracellular Mycobacterium Via Regulating Autophagy. Cells 2019, 8, 506. [Google Scholar] [CrossRef]
- Abdissa, K.; Ruangkiattikul, N.; Ahrend, W.; Nerlich, A.; Beineke, A.; Laarmann, K.; Janze, N.; Lobermeyer, U.; Suwandi, A.; Falk, C.; Schleicher, U.; Weiss, S.; Bogdan, C.; Goethe, R. Relevance of inducible nitric oxide synthase for immune control of Mycobacterium avium subspecies paratuberculosis infection in mice. Virulence 2020, 11, 465–481. [Google Scholar] [CrossRef]
- Harding, H.P. Experimental infection with Mycobacterium johneei. J. Comp. Pathol. Therap. 1957, 67, 37–IN11. [Google Scholar] [CrossRef] [PubMed]
- Mokresh, A.H.; Czuprynski, C.J.; Butler, D.G. A rabbit model for study of Mycobacterium paratuberculosis infection. Infect. Immun. 1989, 57, 3798–3807. [Google Scholar] [CrossRef] [PubMed]
- Mokresh, A.H.; Butler, D.G. Granulomatous enteritis following oral inoculation of newborn rabbits with Mycobacterium paratuberculosis of bovine origin. Can. J. Vet. Res. 1990, 54, 313–319. [Google Scholar]
- Vaughan, J.A.; Lenghaus, C.; Stewart, D.J.; Tizard, M.L.; Michalski, W.P. Development of a Johne’s disease infection model in laboratory rabbits following oral administration of Mycobacterium avium subspecies paratuberculosis. Vet. Microbiol. 2005, 105, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Arrazuria, R.; Molina, E.; Mateo-Adad, M.; Arostegui, I.; Garrido, J.; Juste, R.A.; Elguezabal, N. Effect of various dietary regimens infection on oral challenge with Mycobacterium avium subsp. paratuberculosis in a rabbit model. Res.Vet. Sci. 2015, 101, 80–83. [Google Scholar]
- Beard, P.M.; Rhind, S.M.; Buxton, D.; Daniels, M.J.; Henderson, D.; Pirie, A.; Rudge, K.; Greig, A.; Hutchings, M.R.; Stevenson, K.; Sharp, J.M. Natural paratuberculosis infection in rabbits in Scotland. J. Comp. Pathol. 2001, 124, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Greig, A.; Stevenson, K.; Henderson, D.; Perez, V.; Hughes, V.; Pavlik, I.; Hines, M.E.; McKendrick, I.; Sharp, J.M. Epidemiological study of paratuberculosis in wild rabbits in Scotland. J. Clin. Microbiol. 1999, 37, 1746–1751. [Google Scholar] [CrossRef]
- Francis, J. Infection of laboratory animals with Mycobacterium johnei. J. Comp. Pathol. 1943, 53, 140–150. [Google Scholar] [CrossRef]
- Hirch, A. Infection of hamsters and rabbits with Mycobacterium johnei. J. Comp. Pathol. 1956, 66, 260–269. [Google Scholar] [CrossRef]
- Collins, P.; Matthews, P.R.; McDiarmid, A.; Brown, A. The pathogenicity of Mycobacterium avium and related mycobacteria for experimental animals. J. Med. Microbiol., 1983, 16, 27–35. [Google Scholar] [CrossRef]
- Oyanguren, M.; Molina, E.; Mugica, M.; Badiola, A.; Ladero-Auñon, I.; Elguezabal, N. Probiotic bacteria modulate immune responses to paratuberculosis vaccination. In Proceedings of the 15th International Association for Paratuberculosis Colloquium. Dublin, Ireland, 2022., 12-16 June.
- Arrazuria, R.; Elguezabal, N.; Juste, R.A.; Derakhshani, H.; Khafipour, E. Mycobacterium avium Subspecies paratuberculosis Infection Modifies Gut Microbiota under Different Dietary Conditions in a Rabbit Model. Front Microbiol. 2016, 7, 446. [Google Scholar] [CrossRef] [PubMed]
- Arrazuria, R.; Molina, E.; Garrido, J.M.; Pérez, V.; Juste, R.A.; Elguezabal, N. Vaccination sequence effects on immunological response and tissue bacterial burden in paratuberculosis infection in a rabbit model. Vet Res. 2016, 47, 77. [Google Scholar] [CrossRef]
- Fernández, B. (CONICET-Universidad de Buenos Aires, Ciudad de Buenos Aires, Argentina); Jolly, A. (Universidad de Buenos Aires, Ciudad de Buenos Aires, Argentina). Personal communication, 2023.
- Ladero-Auñon, I.; Molina, E.; Oyanguren, M.; Barriales, D.; Fuertes, M.; Sevilla, I.A.; Luo, L.; Arrazuria, R.; De Buck, J.; Anguita, J.; Elguezabal, N. Oral vaccination stimulates neutrophil functionality and exerts protection in a Mycobacterium avium subsp. paratuberculosis infection model. NPJ Vaccines 2021, 6, 102. [Google Scholar] [CrossRef]
- Arrazuria, R.; Ladero, I.; Molina, E.; Fuertes, M.; Juste, R.; Fernández, M.; Pérez, V.; Garrido, J.; Elguezabal, N. Alternative Vaccination Routes against Paratuberculosis Modulate Local Immune Response and Interference with Tuberculosis Diagnosis in Laboratory Animal Models. Vet. Sci. 2020, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Lamont, E.A.; Sreevatsan, S. Paradigm redux–Mycobacterium avium subspecies paratuberculosis-macrophage interactions show clear variations between bovine and human physiological body temperatures. Microb. Pathog. 2010, 48, 143–9. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Jakobsen, I.; Gu, X.; Nei, M. Diversity and evolution of T-cell receptor variable region genes in mammals and birds. Immunogenetics 1999, 50, 301–8. [Google Scholar] [CrossRef]
- Charavaryamath, C.; Gonzalez-Cano, P.; Fries, P.; Gomis, S.; Doig, K.; Scruten, E.; Potter, A.; Napper, S.; Griebel, P.J. Host responses to persistent Mycobacterium avium subspecies paratuberculosis infection in surgically isolated bovine ileal segments. Clin. Vaccine Immunol. 2013, 20, 156–65. [Google Scholar] [CrossRef]
- Krueger, L.A.; Beitz, D.C.; Humphrey, S.B.; Stabel, J.R. Gamma delta T cells are early responders to Mycobacterium avium ssp. paratuberculosis in colostrum-replete Holstein calves. J. Dairy Sci. 2016, 99, 9040–9050. [Google Scholar] [CrossRef]
- Khalifeh, M.S.; Stabel, J.R. Effects of gamma interferon, interleukin-10, and transforming growth factor beta on the survival of Mycobacterium avium subsp. paratuberculosis in monocyte-derived macrophages from naturally infected cattle. Infect. Immun. 2004, 72, 1974–1782. [Google Scholar] [CrossRef] [PubMed]
- Albarrak, S.M.; Waters, W.R.; Stabel, J.R.; Hostetter, J.M. Evaluating the cytokine profile of the WC1+ γδ T cell subset in the ileum of cattle with the subclinical and clinical forms of MAP infection. Vet. Immunol. Immunopathol. 2018, 201, 26–31. [Google Scholar] [CrossRef]
- Ludwig, L.; Egan, R.; Baquero, M.; Mansz, A.; Plattner, B.L. WC1+ and WC1neg γδ T lymphocytes in intestinal mucosa of healthy and Mycobacterium avium subspecies paratuberculosis-infected calves. Vet. Immunol. Immunopathol. 2019, 216, 109919. [Google Scholar] [CrossRef]
- Albrecht, R.A.; Liu, W.C.; Sant, A.J.; Tompkins, S.M.; Pekosz, A.; Meliopoulos, V.; Cherry, S.; Thomas, P.G.; Schultz-Cherry, S. Moving Forward: Recent Developments for the Ferret Biomedical Research Model. mBio 2018, 9, e01113–18. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.L.; Labes, R.E.; Mackintosh, C.G. Oral infection of ferrets with virulent Mycobacterium bovis or Mycobacterium avium: susceptibility, pathogenesis and immune response. J. Comp. Pathol. 2000, 123, 15–21. [Google Scholar] [CrossRef] [PubMed]
- McCallan, L.; Corbett, D.; Andersen, P.L.; Aagaard, C.; McMurray, D.; Barry, C.; Thompson, S.; Strain, S.; McNair, J. A New Experimental Infection Model in Ferrets Based on Aerosolised Mycobacterium bovis. Vet. Med. Int. 2011, 2011, 981410. [Google Scholar] [CrossRef] [PubMed]
- Lugton, I.W.; Wobeser, G.; Morris, R.S.; Caley, P. Epidemiology of Mycobacterium bovis infection in feral ferrets (Mustela furo) in New Zealand: I. Pathology and diagnosis. N. Z. Vet. J. 1997, 45, 140–50. [Google Scholar] [CrossRef]
- Bannantine, J.P.; Gupta, T.; Zinniel, D.K.; Hikal, A.; Quinn, F.D.; Barletta, R.G. Use of a Ferret Model to Test Efficacy and Immunogenicity of Live Attenuated Mycobacterium avium Subspecies paratuberculosis Vaccines. Methods Mol. Biol. 2022, 2411, 95–104. [Google Scholar] [CrossRef]
- Mayer, J.; Marinim, R.P.; Fox, J.G. Biology and Diseases of Ferrets. Lab. Anim. Med. 2015, 2015, 577–622. [Google Scholar] [CrossRef]
- Mura, M.; Bull, T.J.; Evans, H.; Sidi-Boumedine, K.; McMinn, L.; Rhodes, G.; Pickup, R.; Hermon-Taylor, J. Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp. paratuberculosis within Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2006, 72, 854–9. [Google Scholar] [CrossRef] [PubMed]
- Drancourt, M. Looking at amoebae as a source of mycobacteria. Microb. Pathog. 2014, 77, 119–24. [Google Scholar] [CrossRef]
- Samba-Louaka, A.; Robino, E.; Cochard, T.; Branger, M.; Delafont, V.; Aucher, W.; Wambeke, W.; Bannantine, J.P.; Biet, F.; Héchard, Y. Environmental Mycobacterium avium subsp. paratuberculosis Hosted by Free-Living Amoebae. Front. Cell. Infect. Microbiol. 2018, 8, 28. [Google Scholar] [CrossRef]
- Tenant, R.; Bermudez, L.E. Mycobacterium avium genes upregulated upon infection of Acanthamoeba castellanii demonstrate a common response to the intracellular environment. Curr. Microbiol. 2006, 52, 128–33. [Google Scholar] [CrossRef] [PubMed]
- Dubois, V.; Pawlik, A.; Bories, A.; Le Moigne, V.; Sismeiro, O.; Legendre, R.; Varet, H.; Rodríguez-Ordóñez, M.D.P.; Gaillard, J.L.; Coppée, J.Y.; Brosch, R.; Herrmann, J.L.; Girard-Misguich, F. Mycobacterium abscessus virulence traits unraveled by transcriptomic profiling in amoeba and macrophages. PLoS Pathog. 2019, 15, e1008069. [Google Scholar] [CrossRef]
- Phillips, I.L.; Everman, J.L.; Bermudez, L.E.; Danelishvili, L. Acanthamoeba castellanii as a Screening Tool for Mycobacterium avium Subspecies paratuberculosis Virulence Factors with Relevance in Macrophage Infection. Microorganisms 2020, 8, 1571. [Google Scholar] [CrossRef] [PubMed]
- Randall, K.J.; Turton, J.; Foster, J.R. Explant culture of gastrointestinal tissue: a review of methods and applications. Cell. Biol. Toxicol. 2011, 27, 267–84. [Google Scholar] [CrossRef] [PubMed]
- Karasov, W.H.; Diamond, J.M. A simple method for measuring intestinal nutrient uptake in vitro. J. Comp. Physiol. B. 1983, 152, 105–116. [Google Scholar] [CrossRef]
- Schleig, P.M.; Buergelt, C.D.; Davis, J.K.; Williams, E.; Monif, G.R.; Davidson, M.K. Attachment of Mycobacterium avium subspecies paratuberculosis to bovine intestinal organ cultures: method development and strain differences. Vet. Microbiol. 2005, 108, 271–9. [Google Scholar] [CrossRef] [PubMed]
- Sigurdardóttir, O.G.; Bakke-McKellep, A.M.; Djønne, B.; Evensen, O. Mycobacterium avium subsp. paratuberculosis enters the small intestinal mucosa of goat kids in areas with and without Peyer's patches as demonstrated with the everted sleeve method. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 223–30. [Google Scholar] [CrossRef] [PubMed]
- Brand, R.M.; Biswas, N.; Siegel, A.; Myerski, A.; Engstrom, J.; Jeffrey Metter, E.; Brand, R.E.; Cranston, R.D.; McGowan, I. Immunological responsiveness of intestinal tissue explants and mucosal mononuclear cells to ex vivo stimulation. J. Immunol. Methods. 2018, 463, 39–46. [Google Scholar] [CrossRef]
- Li, Z.; You, Q.; Ossa, F.; Mead, P.; Quinton, M.; Karrow, N.A. Assessment of yeast Saccharomyces cerevisiae component binding to Mycobacterium avium subspecies paratuberculosis using bovine epithelial cells. BMC Vet. Res. 2016, 12, 1–10. [Google Scholar] [CrossRef]
- Villena, J.; Aso, H.; Rutten, V.P.M.G.; Takahashi, H.; van Eden, W.; Kitazawa, H. Immunobiotics for the Bovine Host: Their Interaction with Intestinal Epithelial Cells and Their Effect on Antiviral Immunity. Front Immunol. 2018, 9, 326. [Google Scholar] [CrossRef]
- Hostetter, J.; Steadham, E.; Haynes, J.; Bailey, T.; Cheville, N. Phagosomal maturation and intracellular survival of Mycobacterium avium subspecies paratuberculosis in J774 cells. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 269–83. [Google Scholar] [CrossRef]
- Kabara, E.; Coussens, P.M. Infection of Primary Bovine Macrophages with Mycobacterium avium Subspecies paratuberculosis Suppresses Host Cell Apoptosis. Front. Microbiol. 2012, 3, 215. [Google Scholar] [CrossRef]
- Langelaar, M.F.; Weber, C.N.; Overdijk, M.B.; Müller, K.E.; Koets, A.P.; Rutten, V.P. Cytokine gene expression profiles of bovine dendritic cells after interaction with Mycobacterium avium ssp. paratuberculosis (M.a.p.), Escherichia coli (E. coli) or recombinant M.a.p. heat shock protein 70. Vet. Immunol. Immunopathol. 2005, 107, 153–61. [Google Scholar] [CrossRef]
- Abdellrazeq, G.S.; Fry, L.M.; Elnaggar, M.M.; Bannantine, J.P.; Schneider, D.A.; Chamberlin, W.M.; Mahmoud, A.H.A.; Park, K.T.; Hulubei, V.; Davis, W.C. Simultaneous cognate epitope recognition by bovine CD4 and CD8 T cells is essential for primary expansion of antigen-specific cytotoxic T-cells following ex vivo stimulation with a candidate Mycobacterium avium subsp. paratuberculosis peptide vaccine. Vaccine 2020, 38, 2016–2025. [Google Scholar] [CrossRef]
- Arteche-Villasol, N.; Benavides, J.; Espinosa, J.; Vallejo, R.; Royo, M.; Ferreras, M.D.C.; Pérez, V.; Gutiérrez-Expósito, D. Optimized in vitro isolation of different subpopulation of immune cells from peripheral blood and comparative techniques for generation of monocyte-derived macrophages in small ruminants. Vet. Immunol. Immunopathol. 2020, 230, 110131. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Andersen, M.N.; Møller, H.J. Monocyte isolation techniques significantly impact the phenotype of both isolated monocytes and derived macrophages in vitro. Immunology 2020, 159, 63–74. [Google Scholar] [CrossRef]
- Mitchell, R.M.; Gollnick, N.S.; Sreevatsan, S.; Russell, D.G.; Schukken, Y.H. Quantification of Mycobacterium avium subsp. paratuberculosis (MAP) survival in monocyte-derived macrophages. Vet. Immunol. Immunopathol. 2011, 139, 73–8. [Google Scholar] [CrossRef]
- Mathie, H.A.; Jensen, K.; Stevens, J.M.; Glass, E.J.; Hope, J.C. Quantifying Mycobacterium avium subspecies paratuberculosis infection of bovine monocyte derived macrophages by confocal microscopy. J. Microbiol. Methods. 2020, 168, 105779. [Google Scholar] [CrossRef]
- Bendixen, P.H.; Bloch, B.; Jorgensen, J.B. Lack of intracellular degradation of Mycobacterium paratuberculosis by bovine macrophages infected in vitro and in vivo: light microscopic and electron microscopic observations. Am. J. Vet. Res. 1981, 42, 109–13. [Google Scholar]
- Zurbrick, B.G.; Czuprynski, C.J. Ingestion and intracellular growth of Mycobacterium paratuberculosis within bovine blood monocytes and monocyte-derived macrophages. Infect. Immun. 1987, 55, 1588–93. [Google Scholar] [CrossRef]
- Weiss, D.J.; Evanson, O.A.; Deng, M.; Abrahamsen, M.S. Sequential patterns of gene expression by bovine monocyte-derived macrophages associated with ingestion of mycobacterial organisms. Microb. Pathog. 2004, 37, 215–24. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Evanson, O.A.; de Souza, C.; Abrahamsen, M.S. A critical role of interleukin-10 in the response of bovine macrophages to infection by Mycobacterium avium subsp paratuberculosis. Am. J. Vet. Res. 2005, 66, 721–6. [Google Scholar] [CrossRef] [PubMed]
- Khalifeh, M.S.; Al-Majali, A.M.; Stabel, J.R. Role of nitric oxide production in dairy cows naturally infected with Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2009, 131, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.D. Blocking the mitogen activated protein kinase-p38 pathway is associated with increase expression of nitric oxide synthase and higher production of nitric oxide by bovine macrophages infected with Mycobacterium avium subsp paratuberculosis. Vet. Immunol. Immunopathol. 2015, 164, 1–9. [Google Scholar] [CrossRef]
- Wherry, T.L.T.; Dassanayake, R.P.; Casas, E.; Mooyottu, S.; Bannantine, J.P.; Stabel, J.R. Exogenous Vitamin D3 Modulates Response of Bovine Macrophages to Mycobacterium avium subsp. paratuberculosis Infection and Is Dependent Upon Stage of Johne's Disease. Front. Cell. Infect. Microbiol. 2022, 11, 773938. [Google Scholar] [CrossRef]
- Jolly, A.; Lompardía, S.; Hajos, S.E.; Mundo, S.L. Evidence of a pro-apoptotic effect of specific antibodies in a bovine macrophage model of infection with Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2016, 169, 47–53. [Google Scholar] [CrossRef]
- Arteche-Villasol, N.; Gutiérrez-Expósito, D.; Vallejo, R.; Espinosa, J.; Elguezabal, N.; Ladero-Auñon, I.; Royo, M.; Del Carmen Ferreras, M.; Benavides, J.; Pérez, V. Early response of monocyte-derived macrophages from vaccinated and non-vaccinated goats against in vitro infection with Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2021, 52, 69. [Google Scholar] [CrossRef]
- Lamont, E.A.; Talaat, A.M.; Coussens, P.M.; Bannantine, J.P.; Grohn, Y.T.; Katani, R.; Li, L.L.; Kapur, V.; Sreevatsan, S. Screening of Mycobacterium avium subsp. paratuberculosis mutants for attenuation in a bovine monocyte-derived macrophage model. Front. Cell. Infect. Microbiol. 2014, 4, 87. [Google Scholar] [CrossRef]
- Rathnaiah, G.; Lamont, E.A.; Harris, N.B.; Fenton, R.J.; Zinniel, D.K.; Liu, X.; Sotos, J.; Feng, Z.; Livneh-Kol, A.; Shpigel, N.Y.; Czuprynski, C.J.; Sreevatsan, S.; Barletta, R.G. Generation and screening of a comprehensive Mycobacterium avium subsp. paratuberculosis transposon mutant bank. Front. Cell. Infect. Microbiol. 2014, 4, 144. [Google Scholar] [CrossRef]
- Gollnick, N.S.; Mitchell, R.M.; Baumgart, M.; Janagama, H.K.; Sreevatsan, S.; Schukken, Y.H. Survival of Mycobacterium avium subsp. paratuberculosis in bovine monocyte-derived macrophages is not affected by host infection status but depends on the infecting bacterial genotype. Vet. Immunol. Immunopathol. 2007, 120, 93–105. [Google Scholar] [CrossRef]
- Koets, A.; Santema, W.; Mertens, H.; Oostenrijk, D.; Keestra, M.; Overdijk, M.; Labouriau, R.; Franken, P.; Frijters, A.; Nielen, M.; Rutten, V. Susceptibility to paratuberculosis infection in cattle is associated with single nucleotide polymorphisms in Toll-like receptor 2 which modulate immune responses against Mycobacterium avium subspecies paratuberculosis. Prev. Vet. Med. 2010, 93, 305–15. [Google Scholar] [CrossRef]
- Badia-Bringué, G.; Canive, M.; Alonso-Hearn, M. Control of Mycobacterium avium subsp. paratuberculosis load within infected bovine monocyte-derived macrophages is associated with host genetics. Front. Immunol. 2023, 14, 1042638. [Google Scholar] [CrossRef] [PubMed]
- Ariel, O.; Brouard, J.S.; Marete, A.; Miglior, F.; Ibeagha-Awemu, E.; Bissonnette, N. Genome-wide association analysis identified both RNA-seq and DNA variants associated to paratuberculosis in Canadian Holstein cattle in vitro experimentally infected macrophages. BMC Genomics 2021, 22, 162. [Google Scholar] [CrossRef]
- Marete, A.; Ariel, O.; Ibeagha-Awemu, E.; Bissonnette, N. Identification of Long Non-coding RNA Isolated From Naturally Infected Macrophages and Associated With Bovine Johne's Disease in Canadian Holstein Using a Combination of Neural Networks and Logistic Regression. Front. Vet. Sci. 2021, 8, 639053. [Google Scholar] [CrossRef]
- Janagama, H.K.; Lamont, E.A.; George, S.; Bannantine, J.P.; Xu, W.W.; Tu, Z.J.; Wells, S.J.; Schefers, J.; Sreevatsan, S. Primary transcriptomes of Mycobacterium avium subsp. paratuberculosis reveal proprietary pathways in tissue and macrophages. BMC Genomics 2010, 11, 561. [Google Scholar] [CrossRef]
- Koets, A.P.; Eda, S.; Sreevatsan, S. The within host dynamics of Mycobacterium avium ssp. paratuberculosis infection in cattle: where time and place matter. Vet. Res., 2015, 46, 61. [Google Scholar] [CrossRef] [PubMed]
- Elkington, P.; Lerm, M.; Kapoor, N.; Mahon, R.; Pienaar, E.; Huh, D.; Kaushal, D.; Schlesinger, L.S. In Vitro Granuloma Models of Tuberculosis: Potential and Challenges. J. Infect. Dis. 2019, 219, 1858–1866. [Google Scholar] [CrossRef]
- Abendaño, N.; Tyukalova, L.; Barandika, J.F.; Balseiro, A.; Sevilla, I.A.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Mycobacterium avium subsp. paratuberculosis isolates induce in vitro granuloma formation and show successful survival phenotype, common anti-inflammatory and antiapoptotic responses within ovine macrophages regardless of genotype or host of origin. PLoS One 2014, 9, e104238. [Google Scholar] [CrossRef]
- Rice, H.J.; McDaniel, M.M.; Holland, A.; Eda, S. Modelling Bovine Granuloma Formation In Vitro upon Infection with Mycobacterium avium Subspecies paratuberculosis. Vet. Sci. 2019, 6, 80. [Google Scholar] [CrossRef]
- Ganesan, N.; Ronsmans, S.; Vanoirbeek, J.; Hoet, P.H.M. Assessment of Experimental Techniques That Facilitate Human Granuloma Formation in an In Vitro System: A Systematic Review. Cells 2022, 11, 864. [Google Scholar] [CrossRef]
- Lamont, E.A.; Bannantine, J.P.; Armién, A.; Ariyakumar, D.S.; Sreevatsan, S. Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp. paratuberculosis cultures. PLoS One 2012, 7, e30648. [Google Scholar] [CrossRef]
- Lugo-Villarino, G.; Hudrisier, D.; Benard, A.; Neyrolles, O. Emerging trends in the formation and function of tuberculosis granulomas. Front. Immunol. 2013, 3, 405. [Google Scholar] [CrossRef]
- Kroon, E.E. Neutrophils: Innate effectors of TB resistance? Front. Immunol. 2018, 9, 2637. [Google Scholar] [CrossRef]
- Hilda, J.N.; Das, S.; Tripathy, S.P.; Hanna, L.E. Role of neutrophils in tuberculosis: a bird’s eye view. Innate Immun. 2020, 26, 240–247. [Google Scholar] [CrossRef]
- Khare, S.; Nunes, J.S.; Figueiredo, J.F.; Lawhon, S.D.; Rossetti, C.A.; Gull, T.; Rice-Ficht, A.C.; Adams, L.G. Early phase morphological lesions and transcriptional responses of bovine ileum infected with Mycobacterium avium subsp. paratuberculosis. Vet. Pathol., 2009, 46, 717–28. [Google Scholar] [CrossRef] [PubMed]
- Trentini, M.M.; de Oliveira, F.M.; Kipnis, A.; Junqueira-Kipnis, A.P. The role of neutrophils in the induction of specific Th1 and Th17 during vaccination against tuberculosis. Front. Microbiol. 2016, 7, 898. [Google Scholar] [CrossRef] [PubMed]
- David, J.; Barkema, H.W.; Guan, L.L.; De Buck, J. Gene-expression profiling of calves 6 and 9 months after inoculation with Mycobacterium avium subspecies paratuberculosis. Vet. Res. 2014, 45, 96. [Google Scholar] [CrossRef]
- Gossner, A.; Watkins, C.; Chianini, F.; Hopkins, J. Pathways and genes associated with immune dysfunction in sheep Paratuberculosis. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Alonso-Hearn, M.; Canive, M.; Blanco-Vazquez, C.; Torremocha, R.; Balseiro, A.; Amado, J.; Varela-Martinez, E.; Ramos, R.; Jugo, BM.; Casais, R. RNA-Seq analysis of ileocecal valve and peripheral blood from Holstein cattle infected with Mycobacterium avium subsp. paratuberculosis revealed dysregulation of the CXCL8/IL8 signaling pathway. Sci. Rep. 2019, 9, 14845. [Google Scholar]
- Ladero-Auñon, I.; Elguezabal, N. Ladero-Auñon, I.; Elguezabal, N. Neutrophil functional assays. In: Encyclopedia of Infection and Immunity. 1st ed; Nima Razei. Elsevier. 2022, 4 pp 141-152.
- Ladero-Auñon, I.; Molina, E.; Holder, A.; Kolakowski, J.; Harris, H.; Urkitza, A.; Anguita, J.; Werling, D.; Elguezabal, N. Bovine Neutrophils Release Extracellular Traps and Cooperate With Macrophages in Mycobacterium avium subsp. paratuberculosis clearance In Vitro. Front. Immunol. 2021, 12, 645304. [Google Scholar] [CrossRef]
- Criado, M.; Pérez, V.; Arteche-Villasol, N.; Elguezabal, N.; Molina, E.; Benavides, J.; Gutiérrez-Expósito, D. Evaluation of the innate immune response of caprine neutrophils against Mycobacterium avium subspecies paratuberculosis in vitro. Vet. Res. 2023, 54, 61. [Google Scholar] [CrossRef] [PubMed]
- Baquero, M.M.; Plattner, B.L. Bovine peripheral blood WC1+ and WC1neg γδ T lymphocytes modulate monocyte-derived macrophage effector functions during in vitro Mycobacterium avium subspecies paratuberculosis infection. Cell. Immunol., 2017, 315, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Pooley, H.B.; Plain, K.M.; Purdie, A.C.; Begg, D.J.; Whittington, R.J.; de Silva, K. Integrated vaccine screening system: using cellular functional capacity in vitro to assess genuine vaccine protectiveness in ruminants. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef]
- Eshraghisamani, R.; Arrazuria, R.; Luo, L.; De Buck, J. Evaluation of Mycobacterium avium subsp. paratuberculosis isocitrate lyase (IcL) and ABC transporter (BacA) knockout mutants as vaccine candidates. Front. Cell. Infect. Microbiol. 2023, 30, 1149419. [Google Scholar] [CrossRef]
- Lamont, E.A.; O'Grady, S.M.; Davis, W.C.; Eckstein, T.; Sreevatsan, S. Infection with Mycobacterium avium subsp. paratuberculosis results in rapid interleukin-1β release and macrophage transepithelial migration. Infect. Immun. 2012, 80, 3225–35. [Google Scholar] [CrossRef]
- Lamont, E.A.; Xu WW, X.; Sreevatsan, S. Host-Mycobacterium avium subsp. paratuberculosis interactome reveals a novel iron assimilation mechanism linked to nitric oxide stress during early infection. BMC Genomics 2013, 14, 694. [Google Scholar] [CrossRef]
- Hostetter, J.; Zhang, W.; Simutis, F. Mycobacterium avium subspecies paratuberculosis infection of cattle does not diminish peripheral blood-derived macrophage mycobactericidal activity. Immunol. Lett. 2006, 107, 76–9. [Google Scholar] [CrossRef]
- Simutis, F.J.; Jones, D.E.; Hostetter, J.M. Failure of antigen-stimulated gammadelta T cells and CD4+ T cells from sensitized cattle to upregulate nitric oxide and mycobactericidal activity of autologous Mycobacterium avium subsp. paratuberculosis-infected macrophages. Vet. Immunol. Immunopathol. 2007, 116, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Park, K.T.; Allen, A.J.; Bannantine, J.P.; Seo, K.S.; Hamilton, M.J.; Abdellrazeq, G.S.; Rihan, H.M.; Grimm, A.; Davis, W.C. Evaluation of two mutants of Mycobacterium avium subsp. paratuberculosis as candidates for a live attenuated vaccine for Johne's disease. Vaccine 2011, 29, 4709–19. [Google Scholar] [CrossRef]
- Park, K.T.; ElNaggar, M.M.; Abdellrazeq, G.S.; Bannantine, J.P.; Mack, V.; Fry, L.M.; Davis, W.C. Phenotype and Function of CD209+ Bovine Blood Dendritic Cells, Monocyte-Derived-Dendritic Cells and Monocyte-Derived Macrophages. PLoS One 2016, 11, e0165247. [Google Scholar] [CrossRef]
- Abdellrazeq, G.S.; Elnaggar, M.M.; Bannantine, J.P.; Park, K.T.; Souza, C.D.; Backer, B.; Hulubei, V.; Fry, L.M.; Khaliel, S.A.; Torky, H.A.; Schneider, D.A.; Davis, W.C. A Mycobacterium avium subsp. paratuberculosis relA deletion mutant and a 35 kDa major membrane protein elicit development of cytotoxic T lymphocytes with ability to kill intracellular bacteria. Vet. Res. 2018, 49, 53. [Google Scholar] [CrossRef]
- Davis, W.C.; Mahmoud, A.H.; Abdellrazeq, G.S.; Elnaggar, M.M.; Dahl, J.L.; Hulubei, V.; Fry, L.M. Ex vivo Platforms to Study the Primary and Recall Immune Responses to Intracellular Mycobacterial Pathogens and Peptide-Based Vaccines. Front. Vet. Sci. 2022, 9, 878347. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; Clevers, H. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature, 2009, 459, 262–5. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Nowak-Imialek, M.; Chen, X.; Chen, D.; Herrmann, D.; Ruan, D.; Chen, A.C.H.; Eckersley-Maslin, M.A.; Ahmad, S.; Lee, Y.L.; Kobayashi, T.; Ryan, D.; Zhong, J.; Zhu, J.; Wu, J.; Lan, G.; Petkov, S.; Yang, J.; Antunes, L.; Campos, L.S.; Fu, B.; Wang, S.; Yong, Y.; Wang, X.; Xue, S.G.; Ge, L.; Liu, Z.; Huang, Y.; Nie, T.; Li, P.; Wu, D.; Pei, D.; Zhang, Y.; Lu, L.; Yang, F.; Kimber, S.J.; Reik, W.; Zou, X.; Shang, Z.; Lai, L.; Surani, A.; Tam, P.P.L.; Ahmed, A.; Yeung, W.S.B.; Teichmann, S.A.; Niemann, H.; Liu, P. Establishment of porcine and human expanded potential stem cells. Nat. Cell. Biol. 2019, 21, 687–699. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, X.; Zheng, Y.; Wang, Z.; Zhao, G.; Ren, J.; Zhang, J.; Wu, J.; Wu, B.; Chen, Y.; Sun, W.; Li, Y.; Su, J.; Ding, Y.; Gao, Y.; Liu, M.; Bai, X.; Sun, L.; Cao, G.; Tang, F.; Bao, S.; Liu, P.; Li, X. Establishment of bovine expanded potential stem cells. Proc. Natl. Acad. Sci. USA. 2021, 118, e2018505118. [Google Scholar] [CrossRef]
- Beumer, J.; Clevers, H. Cell fate specification and differentiation in the adult mammalian intestine. Nat. Rev. Mol. Cell Biol. 2020, 22, 39–53. [Google Scholar] [CrossRef]
- Beaumont, M.; Blanc, F.; Cherbuy, C.; Egidy, G.; Giuffra, E.; Lacroix-Lamandé, S.; Wiedemann, A. Intestinal organoids in farm animals. Vet. Res. 2021, 52, 33. [Google Scholar] [CrossRef]
- Powell, R.H.; Behnke, M.S. WRN conditioned media is sufficient for in vitro propagation of intestinal organoids from large farm and small companion animals. Biol. Open. 2017, 6, 698–705. [Google Scholar] [CrossRef]
- Hamilton, C.A.; Young, R.; Jayaraman, S.; Sehgal, A.; Paxton, E.; Thomson, S.; Katzer, F.; Hope, J.; Innes, E.; Morrison, L.J.; Mabbott, N.A. Development of in vitro enteroids derived from bovine small intestinal crypts. Vet. Res. 2018, 49, 54. [Google Scholar] [CrossRef]
- Alfajaro, M.M.; Kim, J.Y.; Barbé, L.; Cho, E.H.; Park, J.G.; Soliman, M.; Baek, Y.B.; Kang, M.I.; Kim, S.H.; Kim, G.J.; Park, S.I.; Pendu, J.L.; Cho, K.O. Dual Recognition of Sialic Acid and αGal Epitopes by the VP8* Domains of the Bovine Rotavirus G6P[5] WC3 and of Its Mono-reassortant G4P[5] RotaTeq Vaccine Strains. J Virol. 2019, 93, e00941–19. [Google Scholar] [CrossRef]
- Derricott, H.; Luu, L.; Fong, W.Y.; Hartley, C.S.; Johnston, L.J.; Armstrong, S.D.; Randle, N.; Duckworth, C.A.; Campbell, B.J.; Wastling, J.M.; Coombes, J.L. Developing a 3D intestinal epithelium model for livestock species. Cell. Tissue Res. 2019, 375, 409–424. [Google Scholar] [CrossRef]
- Töpfer, E.; Pasotti, A.; Telopoulou, A.; Italiani, P.; Boraschi, D.; Ewart, M.A.; Wilde, C. Bovine colon organoids: From 3D bioprinting to cryopreserved multi-well screening platforms. Toxicol. In Vitro 2019, 61, 104606. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.R.; Yang, H.; Lee, S.I.; Haq, I.; Ock, S.A.; Wi, H.; Lee, H.C.; Lee, P.; Yoo, J.G. Robust Three-Dimensional (3D) Expansion of Bovine Intestinal Organoids: An In Vitro Model as a Potential Alternative to an In Vivo System. Animals (Basel) 2021, 11, 2115. [Google Scholar] [CrossRef]
- Smith, D.; Price, D.R.G.; Burrells, A.; Faber, M.N.; Hildersley, K.A.; Chintoan-Uta, C.; Chapuis, A.F.; Stevens, M.; Stevenson, K.; Burgess, S.T.G.; Innes, E.A.; Nisbet, A.J.; McNeilly, T.N. The Development of Ovine Gastric and Intestinal Organoids for Studying Ruminant Host-Pathogen Interactions. Front. Cell. Infect. Microbiol. 2021, 11, 733811. [Google Scholar] [CrossRef]
- Blake, R.; Jensen, K.; Mabbott, N.; Hope, J.; Stevens, J. The Development of 3D Bovine Intestinal Organoid Derived Models to Investigate Mycobacterium avium ssp paratuberculosis Pathogenesis. Front. Vet. Sci. 2022, 9, 921160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, J.; Yan, P.; He, L.; Zhang, X.; Wang, X.; Shi, Y.; Deng, L.; Zhang, Z.; Zhao, B. In-depth analysis of the relationship between bovine intestinal organoids and enteroids based on morphology and transcriptome. J. Tissue Eng. Regen. Med. 2022, 16, 1032–1046. [Google Scholar] [CrossRef]
- Noel, G.; Baetz, N.W.; Staab, J.F.; Donowitz, M.; Kovbasnjuk, O.; Pasetti, M.F.; Zachos, N.C. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Sci. Rep. 2017, 7, 45270. [Google Scholar] [CrossRef] [PubMed]
- Bar-Ephraim, Y.E.; Kretzschmar, K.; Clevers, H. Organoids in immunological research. Nat. Rev. Immunol. 2020, 20, 279–293. [Google Scholar] [CrossRef]
- Son, Y.S.; Ki, S.J.; Thanavel, R.; Kim, J.J.; Lee, M.O.; Kim, J.; Jung, C.R.; Han, T.S.; Cho, H.S.; Ryu, C.M.; Kim, S.H.; Park, D.S.; Son, M.Y. Maturation of Human Intestinal Organoids in Vitro Facilitates Colonization by Commensal Lactobacilli by Reinforcing the Mucus Layer. FASEB J. 2020, 34, 9899–9910. [Google Scholar] [CrossRef]
- Puschhof, J.; Pleguezuelos-Manzano, C.; Martinez-Silgado, A.; Akkerman, N.; Saftien, A.; Boot, C.; de Waal, A.; Beumer, J.; Dutta, D.; Heo, I.; Clevers, H. Intestinal organoid cocultures with microbes. Nat. Protoc. 2021, 16, 4633–4649. [Google Scholar] [CrossRef]
- Heo, I.; Dutta, D.; Schaefer, D.A.; Iakobachvili, N.; Artegiani, B.; Sachs, N.; Boonekamp, K.E.; Bowden, G.; Hendrickx, A.P.A.; Willems, R.J.L.; Peters, P.J.; Riggs, M.W.; O'Connor, R.; Clevers, H. Modelling Cryptosporidium infection in human small intestinal and lung organoids. Nat. Microbiol. 2018, 3, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Delgado, N.E.; Carranza-Torres, I.E.; García-Davis, S.; Rivera, G.; Morán-Martínez, J.; Betancourt-Martínez, N.D.; Groothuis, G.M.M.; de Graaf, I.A.M.; Carranza-Rosales, P. Development of a Novel Ex-vivo 3D Model to Screen Amoebicidal Activity on Infected Tissue. Sci. Rep. 2019, 9, 8396. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.E.; Ramani, S.; Blutt, S.E.; Estes, M.K. Organoids to Dissect Gastrointestinal Virus-Host Interactions: What Have We Learned? Viruses 2021, 13, 999. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; Faruque, A.S.; Zaidi, A.K.; Saha, D.; Alonso, P.L.; Tamboura, B.; Sanogo, D.; Onwuchekwa, U.; Manna, B.; Ramamurthy, T.; Kanungo, S.; Ochieng, J.B.; Omore, R.; Oundo, J.O.; Hossain, A.; Das, S.K.; Ahmed, S.; Qureshi, S.; Quadri, F.; Adegbola, R.A.; Antonio, M.; Hossain, M.J.; Akinsola, A.; Mandomando, I.; Nhampossa, T.; Acácio, S.; Biswas, K.; O'Reill, C.E.; Mintz, E.D.; Berkeley, L.Y.; Muhsen, K.; Sommerfelt, H.; Robins-Browne, R.M.; Levine, M.M. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 2013, 382, 209–22. [Google Scholar] [CrossRef]
- Hill, D.R.; Huang, S.; Nagy, M.S.; Yadagiri, V.K.; Fields, C.; Mukherjee, D.; Bons, B.; Dedhia, P.H.; Chin, A.M.; Tsai, Y.H.; Thodla, S.; Schmidt, T.M.; Walk, S.; Young, V.B.; Spence, J.R. Bacterial colonization stimulates a complex physiological response in the immature human intestinal epithelium. Elife 2017, 6, e29132. [Google Scholar] [CrossRef]
- Roodsant, T.; Navis, M.; Aknouch, I.; Renes, I.B.; van Elburg, R.M.; Pajkrt, D.; Wolthers, K.C.; Schultsz, C.; van der Ark, K.C.H.; Sridhar, A.; Muncan, V. A Human 2D Primary Organoid-Derived Epithelial Monolayer Model to Study Host-Pathogen Interaction in the Small Intestine. Front. Cell. Infect. Microbiol. 2020, 10, 272. [Google Scholar] [CrossRef]
- Abuaita, B.H.; Lawrence, A.L.E.; Berger, R.P.; Hill, D.R.; Huang, S.; Yadagiri, V.K.; Bons, B.; Fields, C.; Wobus, C.E.; Spence, J.R.; Young, V.B.; O'Riordan, M.X. Comparative transcriptional profiling of the early host response to infection by typhoidal and non-typhoidal Salmonella serovars in human intestinal organoids. PLoS Pathog. 2021, 17, e100998. [Google Scholar] [CrossRef]
- Co, J.Y.; Margalef-Català, M.; Monack, D.M.; Amieva, M.R. Controlling the polarity of human gastrointestinal organoids to investigate epithelial biology and infectious diseases. Nat Protoc. 2021, 16, 5171–5192. [Google Scholar] [CrossRef]
- Aguirre Garcia, M.; Hillion, K.; Cappelier, J.M.; Neunlist, M.; Mahe, M.M.; Haddad, N. Intestinal Organoids: New Tools to Comprehend the Virulence of Bacterial Foodborne Pathogens. Foods 2022, 11, 108. [Google Scholar] [CrossRef]
- Kawasaki, M.; Goyama, T.; Tachibana, Y.; Nagao, I.; Ambrosini, Y.M. Farm and Companion Animal Organoid Models in Translational Research: A Powerful Tool to Bridge the Gap Between Mice and Humans. Front. Med. Technol. 2022, 4, 895379. [Google Scholar] [CrossRef]
- Hentschel, V.; Seufferlein, T.; Armacki, M. Intestinal organoids in coculture: redefining the boundaries of gut mucosa ex vivo modeling. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G693–G704. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, A.; Pelzer, K.; Sriranganathan, N. The Paratuberculosis Paradigm Examined: A Review of Host Genetic Resistance and Innate Immune Fitness in Mycobacterium avium subsp. paratuberculosis Infection. Front. Vet. Sci. 2021, 8, 721706. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hearn, M.; Badia-Bringué, G.; Canive, M. Genome-wide association studies for the identification of cattle susceptible and resilient to paratuberculosis. Front. Vet. Sci. 2022, 9, 935133. [Google Scholar] [CrossRef] [PubMed]
- Sigurdardóttir, O.G.; Valheim, M.; Press, C.M. Establishment of Mycobacterium avium subsp. paratuberculosis infection in the intestine of ruminants. Adv. Drug Deliv. Rev. 2004, 56, 819–34. [Google Scholar] [CrossRef]
- Bermudez, L.E.; Petrofsky, M.; Sommer, S.; Barletta, R.G. Peyer's patch-deficient mice demonstrate that Mycobacterium avium subsp. paratuberculosis translocates across the mucosal barrier via both M cells and enterocytes but has inefficient dissemination. Infect. Immun. 2010, 78, 3570–7. [Google Scholar] [CrossRef]
- Ponnusamy, D.; Periasamy, S.; Tripathi, B.N.; Pal, A. Mycobacterium avium subsp. paratuberculosis invades through M cells and enterocytes across ileal and jejunal mucosa of lambs. Res. Vet. Sci. 2013, 94(2), 306–312. [Google Scholar] [CrossRef]
- Golan, L.; Livneh-Kol, A.; Gonen, E.; Yagel, S.; Rosenshine, I.; Shpigel, N.Y. Mycobacterium avium paratuberculosis invades human small-intestinal goblet cells and elicits inflammation. J. Infect. Dis. 2009, 199, 350–4. [Google Scholar] [CrossRef]
- Facciuolo, A.; Gonzalez-Cano, P.; Napper, S.; Griebel, P.J.; Mutharia, L.M. Marked Differences in Mucosal Immune Responses Induced in Ileal versus Jejunal Peyer's Patches to Mycobacterium avium subsp. paratuberculosis Secreted Proteins following Targeted Enteric Infection in Young Calves. PLoS One 2016, 11, e0158747. [Google Scholar] [CrossRef]
- Facciuolo, A.; Lee, A.H.; Gonzalez Cano, P.; Townsend, H.G.G.; Falsafi, R.; Gerdts, V.; Potter, A.; Napper, S.; Hancock, R.E.W.; Mutharia, L.M.; Griebel, P.J. Regional Dichotomy in Enteric Mucosal Immune Responses to a Persistent Mycobacterium avium ssp. paratuberculosis Infection. Front. Immunol. 2020, 11, 1020. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Bissonnette, N.; Do, D.N.; Dudemaine, P.L.; Wang, M.; Facciuolo, A.; Griebel, P. Regionally Distinct Immune and Metabolic Transcriptional Responses in the Bovine Small Intestine and Draining Lymph Nodes During a Subclinical Mycobacterium avium subsp. paratuberculosis Infection. Front. Immunol. 2021, 12, 760931. [Google Scholar] [CrossRef]
- Stabel, J.R.; Stabel, T.J. Immortalization and characterization of bovine peritoneal macrophages transfected with SV40 plasmid DNA. Vet. Immunol. Immunopathol. 1995, 45, 211–20. [Google Scholar] [CrossRef]
- Woo, S.R.; Sotos, J.; Hart, A.P.; Barletta, R.G.; Czuprynski, C.J. Bovine monocytes and a macrophage cell line differ in their ability to phagocytose and support the intracellular survival of Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2006, 110, 109–20. [Google Scholar] [CrossRef]
- Alonso-Hearn, M.; Magombedze, G.; Abendaño, N.; Landin, M.; Juste, R.A. Deciphering the virulence of Mycobacterium avium subsp. paratuberculosis isolates in animal macrophages using mathematical models. J. Theor. Biol. 2019, 468, 82–91. [Google Scholar] [CrossRef]
- Tooker, B.C.; Coussens, P.M. Phagocytosis of M. paratuberculosis fails to activate expression of NADH dehydrogenase and nucleolin-related protein in bovine macrophages. Immunol Lett. 2004, 93, 137–42. [Google Scholar] [CrossRef]
- Abendaño, N.; Sevilla, I.A.; Prieto, J.M.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Mycobacterium avium subspecies paratuberculosis isolates from sheep and goats show reduced persistence in bovine macrophages than cattle, bison, deer and wild boar strains regardless of genotype. Vet. Microbiol. 2013, 163, 325–34. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hearn, M.; Abendaño, N.; Ruvira, M.A.; Aznar, R.; Landin, M.; Juste, R.A. Mycobacterium avium subsp. paratuberculosis (Map) Fatty Acids Profile Is Strain-Dependent and Changes Upon Host Macrophages Infection. Front. Cell. Infect. Microbiol. 2017, 7, 89. [Google Scholar] [CrossRef]
- Mundo, S.L.; Fontanals, A.M.; García, M.; Durrieu, M.; Alvarez, E.; Gentilini, E.R.; Hajos, S.E. Bovine IgG1 antibodies against Mycobacterium avium subsp. paratuberculosis protein p34-cx improve association of bacteria and macrophages. Vet Res. 2008, 39, 6. [Google Scholar] [CrossRef]
- Jolly, A.; Colavecchia, S.B.; Fernández, B.; Fernández, E.; Mundo, S.L. Antibodies Induced by Lipoarabinomannan in Bovines: Characterization and Effects on the Interaction between Mycobacterium avium subsp. paratuberculosis and Macrophages In Vitro. Vet. Med. Int. 2011, 2011, 258479. [Google Scholar] [CrossRef] [PubMed]
- Everman, J.L.; Bermudez, L.E. Antibodies against invasive phenotype-specific antigens increase Mycobacterium avium subspecies paratuberculosis translocation across a polarized epithelial cell model and enhance killing by bovine macrophages. Front. Cell. Infect. Microbiol. 2015, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kang, H.; You, Q.; Ossa, F.; Mead, P.; Quinton, M.; Karrow, N.A. In vitro bioassessment of the immunomodulatory activity of Saccharomyces cerevisiae components using bovine macrophages and Mycobacterium avium ssp. paratuberculosis. J. Dairy Sci. 2018, 101, 6271–6286. [Google Scholar] [CrossRef] [PubMed]
- Kuehnel, M.P.; Goethe, R.; Habermann, A.; Mueller, E.; Rohde, M.; Griffiths, G.; Valentin-Weigand, P. Characterization of the intracellular survival of Mycobacterium avium ssp. paratuberculosis: phagosomal pH and fusogenicity in J774 macrophages compared with other mycobacteria. Cell. Microbiol. 2001, 3, 551–66. [Google Scholar] [CrossRef]
- Raschke, W.C.; Baird, S.; Ralph, P.; Nakoinz, I. Functional macrophage cell lines transformed by Abelson leukemia virus. Cell. 1978, 15, 261–7. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Levis, W.R.; Quinn, M.R.; Park, S.Y.; Schuller-Levis, G.B. Regulation of nitric oxide induced by mycobacterial lipoarabinomannan in murine macrophages: effects of interferon-beta and taurine-chloramine. Int. J. Lepr. Other Mycobact. Dis. 2000, 68, 444–51. [Google Scholar]
- Means, T.K.; Jones, B.W.; Schromm, A.B.; Shurtleff, B.A.; Smith, J.A.; Keane, J.; Golenbock, D.T.; Vogel, S.N.; Fenton, M.J. Differential effects of a Toll-like receptor antagonist on Mycobacterium tuberculosis-induced macrophage responses. J. Immunol. 2001, 166, 4074–82. [Google Scholar] [CrossRef]
- Bohsali, A.; Abdalla, H.; Velmurugan, K.; Briken, V. The non-pathogenic mycobacteria M. smegmatis and M. fortuitum induce rapid host cell apoptosis via a caspase-3 and TNF dependent pathway. BMC Microbiol. 2010, 10, 237. [Google Scholar] [CrossRef]
- Helguera-Repetto, A.C.; Chacon-Salinas, R.; Cerna-Cortes, J.F.; Rivera-Gutierrez, S.; Ortiz-Navarrete, V.; Estrada-Garcia, I.; Gonzalez-y-Merchand, J.A. Differential macrophage response to slow- and fast-growing pathogenic mycobacteria. Biomed. Res. Int. 2014, 2014, 916521. [Google Scholar] [CrossRef]
- Fu, Y.R.; Gao, K.S.; Ji, R.; Yi, Z.J. Differential transcriptional response in macrophages infected with cell wall deficient versus normal Mycobacterium tuberculosis. Int. J. Biol. Sci. 2015, 11, 22–30. [Google Scholar] [CrossRef]
- Taka, S.; Gazouli, M.; Sotirakoglou, K.; Liandris, E.; Andreadou, M.; Triantaphyllopoulos, K.; Ikonomopoulos, J. Functional analysis of 3'UTR polymorphisms in the caprine SLC11A1 gene and its association with the Mycobacterium avium subsp. paratuberculosis infection. Vet. Immunol. Immunopathol. 2015, 167, 75–9. [Google Scholar] [CrossRef]
- Thirunavukkarasu, S.; Plain, K.M.; Purdie, A.C.; Whittington, R.J.; de Silva, K. IFN-γ fails to overcome inhibition of selected macrophage activation events in response to pathogenic mycobacteria. PLoS One. 2017, 12, e0176400. [Google Scholar] [CrossRef] [PubMed]
- Barletta, R.G.; Bannantine, J.P.; Stabel, J.R.; Muthukrishnan, E.; Anderson, D.K.; Dutta, E.; Manthena, V.; Hanafy, M.; Zinniel, D.K. Mycobacterium avium subsp. paratuberculosis Candidate Vaccine Strains Are Pro-apoptotic in RAW 264.7 Murine Macrophages. Vaccines (Basel). 2023, 11, 1085. [Google Scholar] [CrossRef] [PubMed]
- Zur Lage, S.; Goethe, R.; Darji, A.; Valentin-Weigand, P.; Weiss, S. Activation of macrophages and interference with CD4+ T-cell stimulation by Mycobacterium avium subspecies paratuberculosis and Mycobacterium avium subspecies avium. Immunology 2003, 108, 62–9. [Google Scholar] [CrossRef]
- Ghosh, P.; Wu, C.W.; Talaat, A.M. Key role for the alternative sigma factor, SigH, in the intracellular life of Mycobacterium avium subsp. paratuberculosis during macrophage stress. Infect. Immun. 2013, 81, 2242–57. [Google Scholar] [CrossRef]
- Hostetter, J.M.; Steadham, E.M.; Haynes, J.S.; Bailey, T.B.; Cheville, N.F. Cytokine effects on maturation of the phagosomes containing Mycobacterium avium subspecies paratuberculosis in J774 cells. FEMS Immunol. Med. Microbiol. 2002, 34, 127–34. [Google Scholar] [CrossRef]
- Cirone, K.M.; Lahiri, P.; Holani, R.; Tan, Y.L.; Arrazuria, R.; De Buck, J.; Barkema, H.W.; Cobo, E.R. Synthetic cathelicidin LL-37 reduces Mycobacterium avium subsp. paratuberculosis internalization and pro-inflammatory cytokines in macrophages. Cell. Tissue Res. 2020, 379, 207–217. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: an in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Cossu, A.; Sechi, L.A.; Zanetti, S.; Rosu, V. Gene expression profiling of Mycobacterium avium subsp. paratuberculosis in simulated multi-stress conditions and within THP-1 cells reveals a new kind of interactive intramacrophage behaviour. BMC Microbiol. 2012, 12, 87. [Google Scholar] [CrossRef]
- Shin, M.; Shin, S.W.; Jung, M.; Park, H.; Park, H.; Yoo, H.S. Host gene expression for Mycobacterium avium subsp. paratuberculosis infection in human THP-1 macrophages. Pathog. Dis. 2015, 73. [Google Scholar] [CrossRef]
- Mostoufi-Afshar, S.; Tabatabaei, M.; Ghahramani Seno, M.M. Mycobacterium avium subsp. paratuberculosis induces differential cytosine methylation at miR-21 transcription start site region. Iran. J. Vet. Res. (IVJR) 2018, 19, 262–269. [Google Scholar]
- Park, H.T.; Park, W.B.; Kim, S.; Lim, J.S.; Nah, G.; Yoo, H.S. Revealing immune responses in the Mycobacterium avium subsp. paratuberculosis-infected THP-1 cells using single cell RNA-sequencing. PLoS One. 2021, 16, e0254194. [Google Scholar] [CrossRef]
- Park, H.T.; Lee, S.M.; Ko, S.; Kim, S.; Park, H.E.; Shin, M.K.; Kim, D.; Yoo, H.S. Delineating transcriptional crosstalk between Mycobacterium avium subsp. paratuberculosis and human THP-1 cells at the early stage of infection via dual RNA-seq analysis. Vet Res. 2022, 53, 71. [Google Scholar] [CrossRef]
- Borrmann, E.; Möbius, P.; Diller, R.; Köhler, H. Divergent cytokine responses of macrophages to Mycobacterium avium subsp. paratuberculosis strains of Types II and III in a standardized in vitro model. Vet. Microbiol. 2011, 152, 101–11. [Google Scholar] [CrossRef]
- Motamedi Boroojeni, A.; Derakhshandeh, A.; Haghkhah, M.; Jalali, S.A.H.; Eraghi, V.A. A Novel Approach to Deliver a Mycobacterium avium subsp. paratuberculosis Antigen in Eukaryotic Cells. Mol. Biotechnol. 2019, 61, 506–512. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Yao, Y.; Wu, J.; Tang, X.; Gu, T.; Li, G. The fibroblast growth factor-2 arrests Mycobacterium avium sp. paratuberculosis growth and immunomodulates host response in macrophages. Tuberculosis (Edinb). 2015, 95, 505–14. [Google Scholar] [CrossRef]
- Momotani, E.; Whipple, D.L.; Thiermann, A.B.; Cheville, N.F. Role of M cells and macrophages in the entrance of Mycobacterium paratuberculosis into domes of ileal Peyer's patches in calves. Vet. Pathol. 1988, 25, 131–7. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Hearn, M.; Patel, D.; Danelishvili, L.; Meunier-Goddik, L.; Bermudez, L.E. The Mycobacterium avium subsp. paratuberculosis MAP3464 gene encodes an oxidoreductase involved in invasion of bovine epithelial cells through the activation of host cell Cdc42. Infect. Immun. 2008, 76, 170–8. [Google Scholar] [CrossRef]
- Pott, J.; Basler, T.; Duerr, C.U.; Rohde, M.; Goethe, R.; Hornef, M.W. Internalization-dependent recognition of Mycobacterium avium ssp. paratuberculosis by intestinal epithelial cells. Cell. Microbiol. 2009, 11, 1802–15. [Google Scholar] [CrossRef]
- Miyazawa, K.; Hondo, T.; Kanaya, T.; Tanaka, S.; Takakura, I.; Itani, W.; Rose, M.T.; Kitazawa, H.; Yamaguchi, T.; Aso, H. Characterization of newly established bovine intestinal epithelial cell line. Histochem. Cell. Biol. 2010, 133, 125–34. [Google Scholar] [CrossRef]
- Chiba, E.; Villena, J.; Hosoya, S.; Takanashi, N.; Shimazu, T.; Aso, H.; Tohno, M.; Suda, Y.; Kawai, Y.; Saito, T.; Miyazawa, K.; He, F.; Kitazawa, H. A newly established bovine intestinal epithelial cell line is effective for in vitro screening of potential antiviral immunobiotic microorganisms for cattle. Res Vet Sci. 2012, 93, 688–94. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, N.; Tomosada, Y.; Villena, J.; Murata, K.; Takahashi, T.; Chiba, E.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; Ikegami, S.; Itoh, H.; Kawai, Y.; Saito, T.; Alvarez, S.; Kitazawa, H. Advanced application of bovine intestinal epithelial cell line for evaluating regulatory effect of lactobacilli against heat-killed enterotoxigenic Escherichia coli-mediated inflammation. BMC Microbiol. 2013, 13, 54. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kanmani, P.; Ishizuka, T.; Miyazaki, A.; Soma, J.; Albarracin, L.; Suda, Y.; Nochi, T.; Aso, H.; Iwabuchi, N.; Xiao, J.Z.; Saito, T.; Villena, J.; Kitazawa, H. Development of an in vitro immunobiotic evaluation system against rotavirus infection in bovine intestinal epitheliocytes. Benef. Microbes. 2017, 8, 309–321. [Google Scholar] [CrossRef]
- Zhan, K.; Lin, M.; Liu, M.M.; Sui, Y.N.; Zhao, G.Q. Establishment of primary bovine intestinal epithelial cell culture and clone method. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 54–57. [Google Scholar] [CrossRef]
- Katwal, P.; Thomas, M.; Uprety, T.; Hildreth, M.B.; Kaushik, R.S. Development and biochemical and immunological characterization of early passage and immortalized bovine intestinal epithelial cell lines from the ileum of a young calf. Cytotechnology 2019, 71, 127–148. [Google Scholar] [CrossRef]
- Katwal, P.; Uprety, T.; Okda, F.; Antony, L.; Thomas, M.; Chase, C.; Diel, D.G.; Nelson, E.; Young, A.; Li, F.; Scaria, J.; Kaushik, R.S. Characterization of bovine ileal epithelial cell line for lectin binding, susceptibility to enteric pathogens, and TLR mediated immune responses. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101581. [Google Scholar] [CrossRef]
- Kernéis, S.; Bogdanova, A.; Kraehenbuhl, J.P.; Pringault, E. Conversion by Peyer's patch lymphocytes of human enterocytes into M cells that transport bacteria. Science 1997, 277, 949–52. [Google Scholar] [CrossRef]
- El Bahi, S.; Caliot, E.; Bens, M.; Bogdanova, A.; Kernéis, S.; Kahn, A.; Vandewalle, A.; Pringault, E. Lymphoepithelial interactions trigger specific regulation of gene expression in the M cell-containing follicle-associated epithelium of Peyer's patches. J. Immunol. 2002, 168, 3713–20. [Google Scholar] [CrossRef]
- Nair, V.R.; Franco, L.H.; Zacharia, V.M.; Khan, H.S.; Stamm, C.E.; You, W.; Marciano, D.K.; Yagita, H.; Levine, B.; Shiloh, M.U. Microfold Cells Actively Translocate Mycobacterium tuberculosis to Initiate Infection. Cell. Rep. 2016, 6, 1253–1258. [Google Scholar] [CrossRef]
- Bannantine, J.P.; Huntley, J.F.J.; Miltner, E.; Stabel, J.R.; Bermudez, L.E. The Mycobacterium avium subsp. paratuberculosis 35 kDa protein plays a role in invasion of bovine epithelial cells. Microbiology 2003, 149, 2061–2069. [Google Scholar] [CrossRef]
- Patel, D.; Danelishvili, L.; Yamazaki, Y.; Alonso, M.; Paustian, M.L.; Bannantine, J.P.; Meunier-Goddik, L.; Bermudez, L.E. The ability of Mycobacterium avium subsp. paratuberculosis to enter bovine epithelial cells is influenced by preexposure to a hyperosmolar environment and intracellular passage in bovine mammary epithelial cells. Infect. Immun. 2006, 74, 2849–55. [Google Scholar] [CrossRef]
- Alonso-Hearn, M.; Eckstein, T.M.; Sommer, S.; Bermudez, L.E. A Mycobacterium avium subsp. paratuberculosis LuxR regulates cell envelope and virulence. Innate Immun. 2010, 16, 235–47. [Google Scholar] [CrossRef]
- Everman, J.L.; Danelishvili, L.; Flores, L.G.; Bermudez, L.E. MAP1203 Promotes Mycobacterium avium Subspecies paratuberculosis Binding and Invasion to Bovine Epithelial Cells. Front. Cell. Infect. Microbiol. 2018, 8, 217. [Google Scholar] [CrossRef]
- Everman, J.L.; Eckstein, T.M.; Roussey, J.; Coussens, P.; Bannantine, J.P.; Bermudez, L.E. Characterization of the inflammatory phenotype of Mycobacterium avium subspecies paratuberculosis using a novel cell culture passage model. Microbiology (Reading). 2015, 161, 1420–1434. [Google Scholar] [CrossRef]
- Sweeney, R.W.; Whitlock, R.H.; Rosenberger, A.E. Mycobacterium paratuberculosis isolated from fetuses of infected cows not manifesting signs of the disease. Am. J. Vet. Res. 1992, 53, 477–80. [Google Scholar] [PubMed]
- Thompson, B.R.; Clark, R.G.; Mackintosh, C.G. Intra-uterine transmission of Mycobacterium avium subsp paratuberculosis in subclinically affected red deer (Cervus elaphus). N. Z. Vet. J. 2007, 55, 308–13. [Google Scholar] [CrossRef]
- Kugadas, A.; Lamont, E.A.; Bannantine, J.P.; Shoyama, F.M.; Brenner, E.; Janagama, H.K.; Sreevatsan, S. A Mycobacterium avium subsp. paratuberculosis Predicted Serine Protease Is Associated with Acid Stress and Intraphagosomal Survival. Front. Cell. Infect. Microbiol. 2016, 6, 85. [Google Scholar] [CrossRef]
- Schwarz, D.G.G.; Shoyama, F.M.; Oliveira, L.L.; Sreevatsan, S.; Moreira, M.A.S. Rapid baso-apical translocation of Mycobacterium avium ssp. paratuberculosis in mammary epithelial cells in the presence of Escherichia coli. J. Dairy Sci. 2018, 101, 6287–6295. [Google Scholar] [CrossRef] [PubMed]
- Pena, J.L.; Gonçalves Schwarz, D.G.; Willian de Lima Brasil, A.; Licursi de Oliveira, L.; Albuquerque Caldeira, J.L.; Scatamburlo Moreira, M.A. Differences in the coinfective process of Staphylococcus aureus and Streptococcus agalactiae in bovine mammary epithelial cells infected by Mycobacterium avium subsp. paratuberculosis. Microb Pathog. 2020, 149, 104476. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, U.K.; Wu, X.; McAllister, C.; Mutharia, L.; Karrow, N.A. Role of Toll-Like Receptor 4 in Mycobacterium avium subsp. paratuberculosis Infection of Bovine Mammary Epithelial (MAC-T) Cells In Vitro. Microbiol Spectr. 2023, 11, e0439322. [Google Scholar] [CrossRef]
- Shandilya, U.K.; Wu, X.; McAllister, C.; Mutharia, L.; Karrow, N.A. Impact of Mycobacterium avium subsp. paratuberculosis infection on bovine IL10RA knockout mammary epithelial (MAC-T) cells. In Vitro Cell Dev Biol Anim. 2023, 59, 214–223. [Google Scholar] [CrossRef]
- Phillips, I.L.; Danelishvili, L.; Bermudez, L.E. Macrophage Proteome Analysis at Different Stages of Mycobacterium avium Subspecies paratuberculosis Infection Reveals a Mechanism of Pathogen Dissemination. Proteomes 2021, 9, 20. [Google Scholar] [CrossRef]
- Engelmann, I.; Pujol, N. Innate immunity in C. elegans. Adv. Exp. Med. Biol. 2010, 708, 105–21. [Google Scholar] [CrossRef]
- Komura, T.; Yasui, C.; Miyamoto, H.; Nishikawa, Y. Caenorhabditis elegans as an alternative model host for legionella pneumophila, and protective effects of Bifidobacterium infantis. Appl. Environ. Microbiol. 2010, 76, 4105–8. [Google Scholar] [CrossRef] [PubMed]
- Everman, J.L.; Ziaie, N.R.; Bechler, J.; Bermudez, L.E. Establishing Caenorhabditis elegans as a model for Mycobacterium avium subspecies hominissuis infection and intestinal colonization. Biol. Open. 2015, 4, 1330–5. [Google Scholar] [CrossRef]
- Bermudez, L.E.; Rose, S.J.; Everman, J.L.; Ziaie, N.R. Establishment of a Host-to-Host Transmission Model for Mycobacterium avium subsp. hominissuis Using Caenorhabditis elegans and Identification of Colonization-Associated Genes. Front. Cell. Infect. Microbiol. 2018, 8, 123. [Google Scholar] [CrossRef] [PubMed]
- Viri, V.; Arveiler, M.; Lehnert, T.; Gijs, M.A.M. An In Vivo Microfluidic Study of Bacterial Load Dynamics and Absorption in the C. elegans Intestine. Micromachines (Basel) 2021, 12, 832. [Google Scholar] [CrossRef]
- Mallo, G.V.; Kurz, C.L.; Couillault, C.; Pujol, N.; Granjeaud, S.; Kohara, Y.; Ewbank, J.J. Inducible antibacterial defense system in C. elegans. Curr. Biol. 2002, 12, 1209–14. [Google Scholar] [CrossRef]
- Chávez, V.; Mohri-Shiomi, A.; Maadani, A.; Vega, L.A.; Garsin, D.A. Oxidative stress enzymes are required for DAF-16-mediated immunity due to generation of reactive oxygen species by Caenorhabditis elegans. Genetics 2007, 176, 1567–77. [Google Scholar] [CrossRef]
- Galbadage, T.; Shepherd, T.F.; Cirillo, S.L.; Gumienny, T.L.; Cirillo, J.D. The Caenorhabditis elegans p,8 MAPK Gene plays a key role in protection from mycobacteria. Microbiologyopen 2016, 5, 436–52. [Google Scholar] [CrossRef]
- Sutton, K.M.; Orr, B.; Hope, J.; Jensen, S.R.; Vervelde, L. Establishment of bovine 3D enteroid-derived 2D monolayers. Vet. Res. 2022, 53, 15. [Google Scholar] [CrossRef] [PubMed]
- Candarlioglu, P.L.; Dal Negro, G.; Hughes, D.; Balkwill, F.; Harris, K.; Screen, H.; Morgan, H.; David, R.; Beken, S.; Guenat, O.; Rowan, W.; Amour, A. Organ-on-a-chip: current gaps and future directions. Biochem. Soc. Trans. 2022, 50, 665–673. [Google Scholar] [CrossRef]
- Csukovich, G.; Pratscher, B.; Burgener, I.A. The World of Organoids: Gastrointestinal Disease Modelling in the Age of 3R and One Health with Specific Relevance to Dogs and Cats. Animals (Basel) 2022, 2, 2461. [Google Scholar] [CrossRef]
- Xian, C.; Zhang, J.; Zhao, S.; Guang Li, X. Gut-on-a-chip for disease models. J. Tissue. Engineer. 2023, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Baddal, B.; Marrazzo, P. Refining Host-Pathogen Interactions: Organ-on-Chip Side of the Coin. Pathogens. 2021, 10, 203. [Google Scholar] [CrossRef]
- Pimenta, J.; Ribeiro, R.; Almeida, R.; Costa, P.F.; da Silva, M.A.; Pereira, B. Organ-on-Chip Approaches for Intestinal 3D In Vitro Modeling. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Morsink, MAJ. ; Willemen, NGA.; Leijten, J.; Bansal, R.; Shin, S.R. Immune organs and immune cells on a chip: an overview of biomedical applications. Micromachines, 2020, 11, 849. [Google Scholar] [CrossRef]
- Majorova, D.; Atkins, E.; Martineau, H.; Vokral, I.; Oosterhuis, D.; Olinga, P.; Wren, B.; Cuccui, J.; Dirk Werling, D. Use of Precision-Cut Tissue Slices as a Translational Model to Study Host-Pathogen Interaction. Front. Vet. Sci. 2021, 8, 686088. [Google Scholar] [CrossRef]
- Li, M.; de Graaf, I.A.M.; Groothuis, G.M.M. Precision-cut intestinal slices: alternative model for drug transport, metabolism, and toxicology research. Expert. Opin. Drug. Metab. Toxicol. 2016, 12, 175–90. [Google Scholar] [CrossRef] [PubMed]
- de Kanter, R.; Tuin, A.; van de Kerkhof, E.; Martignoni, M.; Draaisma, A.L.; de Jager, M.H.; de Graaf, I.A.; Meijer, D.K.; Groothuis, G.M. A new technique for preparing precision-cut slices from small intestine and colon for drug biotransformation studies. J. Pharmacol. Toxicol. Methods. 2005, 51, 65–72. [Google Scholar] [CrossRef]
- Molina-Torres, C.A.; Flores-Castillo, O.N.; Carranza-Torres, I.E.; Guzmán-Delgado, N.E.; Viveros-Valdez, E.; Vera-Cabrera, L.; Ocampo-Candiani, J.; Verde-Star, J.; Castro-Garza, J.; Carranza-Rosales, P. Ex vivo infection of murine precision-cut lung tissue slices with Mycobacterium abscessus: a model to study antimycobacterial agents. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 52. [Google Scholar] [CrossRef]
- Remot, A.; Carreras, F.; Coupé, A.; Doz-Deblauwe, É.; Boschiroli, M.L.; Browne, J.A.; Marquant, Q.; Descamps, D.; Archer, F.; Aseffa, A.; Germon, P.; Gordon, S.V.; Winter, N. Mycobacterial Infection of Precision-Cut Lung Slices Reveals Type 1 Interferon Pathway Is Locally Induced by Mycobacterium bovis but Not M. tuberculosis in a Cattle Breed. Front. Vet. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Punyadarsaniya, D.; Winter, C.; Mork, A.K.; Amiri, M.; Naim, H.Y.; Rautenschlein, S.; Herrler, G. Precision-cut intestinal slices as a culture system to analyze the infection of differentiated intestinal epithelial cells by avian influenza viruses. J. Virol. Methods. 2015, 212, 71–75. [Google Scholar] [CrossRef]
- Krimmling, T.; Beineke, A.; Schwegmann-Weßels, C. Infection of porcine precision cut intestinal slices by transmissible gastroenteritis coronavirus demonstrates the importance of the spike protein for enterotropism of different virus strains. Vet. Microbiol. 2017, 205, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martinec, O.; Huliciak, M.; Staud, F.; Cecka, F.; Vokral, I.; Cerveny, L. Anti-HIV and Anti-Hepatitis C Virus Drugs Inhibit P-Glycoprotein Efflux Activity in Caco-2 Cells and Precision-Cut Rat and Human Intestinal Slices. Antimicrob. Agents. Chemother. 2019, 63, e00910–19. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Modeling PTB. Relative contribution of the different models discussed in this paper. Data are expressed as the percentage of assays found for each model in PTB research. The information used to calculate these percentages was obtained by the search performed the 7th August 2023 in PubMed using the following key words: “(mouse OR murine) AND paratuberculosis”, “rabbit AND paratuberculosis”, “(protists OR amoeba) AND paratuberculosis”, “ferret AND paratuberculosis”, “macrophage cell line AND paratuberculosis”, “macrophage cell model AND paratuberculosis”, “epithelial cells AND paratuberculosis”, “neutrophil AND paratuberculosis”, “cells co-culture AND paratuberculosis”, and ”granuloma in vitro AND paratuberculosis”. The inclusion criteria was considered to be a research study (not a review) on the use of the experimental model of infection to evaluate some aspects of PTB. In total, 217 assays were considered that met this criterion. It is important to note that these experiments were published in slightly fewer papers, since in some cases the authors published results obtained in different models in the same paper. Those works referring to MAP but in the context of Crohn's disease were excluded. (Created with BioRender.com. Biorender Publication and License Rights agreement number: OB25PTVZ5Q).
Figure 1.
Modeling PTB. Relative contribution of the different models discussed in this paper. Data are expressed as the percentage of assays found for each model in PTB research. The information used to calculate these percentages was obtained by the search performed the 7th August 2023 in PubMed using the following key words: “(mouse OR murine) AND paratuberculosis”, “rabbit AND paratuberculosis”, “(protists OR amoeba) AND paratuberculosis”, “ferret AND paratuberculosis”, “macrophage cell line AND paratuberculosis”, “macrophage cell model AND paratuberculosis”, “epithelial cells AND paratuberculosis”, “neutrophil AND paratuberculosis”, “cells co-culture AND paratuberculosis”, and ”granuloma in vitro AND paratuberculosis”. The inclusion criteria was considered to be a research study (not a review) on the use of the experimental model of infection to evaluate some aspects of PTB. In total, 217 assays were considered that met this criterion. It is important to note that these experiments were published in slightly fewer papers, since in some cases the authors published results obtained in different models in the same paper. Those works referring to MAP but in the context of Crohn's disease were excluded. (Created with BioRender.com. Biorender Publication and License Rights agreement number: OB25PTVZ5Q).
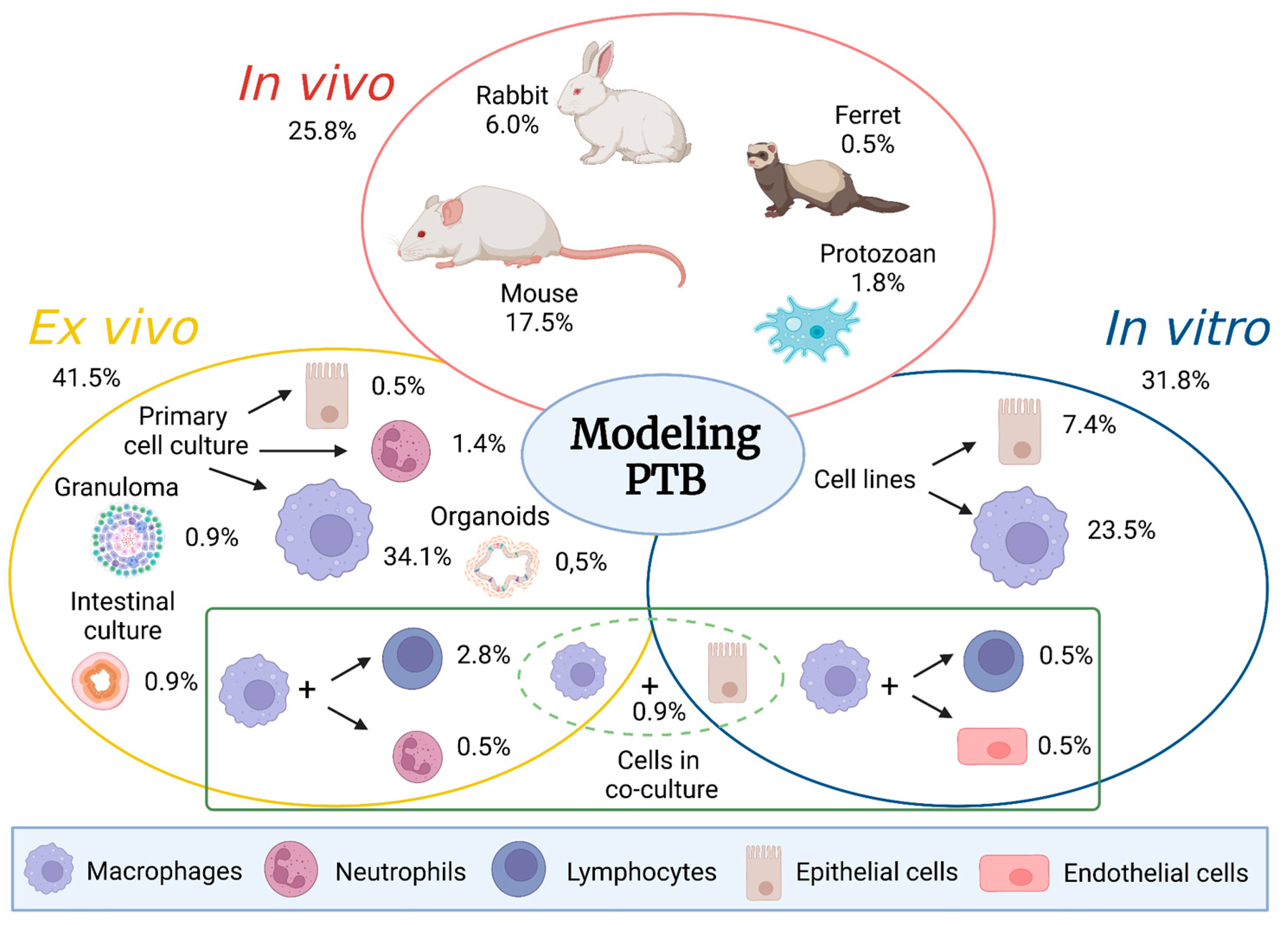

Table 1.
Experimental MAP oral challenge studies in rabbits.
| Age at challenge | Infective strain source | Infective dose (dose number) |
Study end-point w.p.i. |
Evidence of infection %* | Experiment objective |
Reference |
|---|---|---|---|---|---|---|
| 4-5 w | Rabbit /hamster |
1.6-5.6 mg (7) | 4-40 | 6-50 | Pathogenesis | [20] |
| 1-2 d | Cattle | 7x106 CFU (5-10) | 32-40 | 43 | Pathogenesis | [13] |
| 1-2 d | Cattle | 3.6x108 CFU (1) | 2-36 | 38 | Pathogenesis | [14] |
| 1-2 d | Cattle | 2.6x108 CFU (1) | 2-36 | 100 | Pathogenesis | [14] |
| 3 m | Cattle | 5x108 CFU (3) | 104-128 | 50 | Pathogenesis | [15] |
| 2 w | Cattle | 1x108 CFU (3) | 8-84 | 19 | Pathogenesis | [15] |
| 8 w | Cattle K10 | 1x109 CFU (3) | 16-20 | 40-87 | Diet evaluation | [16] |
| 16 w | Cattle K10 | 4x108 CFU (3) | 20 | 80 | MAP infection and diet effect on microbiota | [23] |
| 16 w/ 30 w |
Cattle K10 | 4x108 CFU (6) | 20 | 100 | MAP infection and diet effect on microbiota | [23] |
| 12 w | Cattle K10 | 4x108 CFU (3) | 25 | 80 | Vaccination sequence efficacy | [24] |
| 13 w | Cattle K10 | 1x109 CFU (3) | 24-25 | 80 | Vaccination routes | [27] |
| 13-14 w | Cattle NK-764 | 3x108 CFU (3) | 19 | 60 | Vaccination efficacy and trained immunity | [26] |
| 15 w | Cattle NK-832 | 3x108 CFU (3) | 12 | 100 | Effect of vaccination and probiotics and trained immunity | [22] |
| 8-11 w | Cattle | 1x109 CFU (3) | 21 | 100 | Effect of passively transferred antibodies | [25] |
w: weeks, d: days, CFU: colony forming units, w.p.i.: weeks post-infection, * Evidence of infection was determined by post-mortem tissue culture if data were available, otherwise MAP PCR in tissues, histology, fecal culture or fecal PCR were considered.
Table 2.
Summary of the main experimental models for the study of PTB considered in this review...
| Models | Reported application* P D V T |
References | Main limitations | Scientific potential | |
|---|---|---|---|---|---|
|
In vivo non-ruminant animal models |
Mouse | ✓ ✓ ✓ | [4,5,6,7,8,9,10,11] | Lack of development of clinical signs | Great amount of immunological reagents Well defined model Initial vaccine screening platform |
| Rabbit | ✓ ✓ ✓ ✓ | [13,14,15,16,20,22,23,24,25,26,27] | Scarcity of immunological reagents | Clinical outcome closely resembling ruminant PTB | |
| Ferret | ✓ ✓ | [39] | Scarcity of immunological reagents Carnivore diet Outbred nature of the model |
Strain virulence factor research | |
| Protozoa | ✓ | [41,43,46] | Unicellular organism | Strain virulence factor research | |
| Ex vivo models | Intestinal | ✓ ✓ | [49,50,52] | Difficult to manage No standardized protocols available |
Early stage pathogenesis |
| Immune cells: macrophages/ monocytes |
✓ ✓ ✓ | [33,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,78] | High variability between donors and lower reproducibility | Strain virulence factor research Host genetics MAP-host interaction at receptor-ligand level |
|
| Granulomas | ✓ ✓ | [81,82] | Limited growth potential Not all granuloma cell types included |
Early stage pathogenesis Initial vaccine screening platform |
|
| Immune cells: neutrophils | ✓ ✓ | [26,93,94,95] | Role in PTB not fully understood Short life span cells Inability to cryopreserve or expand them in vitro |
Early stage host-pathogen interaction | |
| Co-cultures of primary cells | ✓ ✓ | [57,94,96,97,98,99,100,101,102,103,104,105,106] | Limited growth potential High variability between donors |
Dissection of cross-talk between key cell types in upon MAP infection Initial vaccine screening platform |
|
| Organoids | ✓ | [119] | No standardized protocols available | Early stage pathogenesis | |
| In vitro models | Macrophage cell lines | ✓ ✓ ✓ | [54,55,69,145,146,147,148,149,150,151,152,153,154,155,163,165,166] | Genetically transformed cells Lower or different response in relation to primary cells |
Strain virulence factor research MAP-host interaction at receptor-ligand level |
| Epithelial cell lines | ✓ ✓ ✓ | [139,179,180,191,192,193,194,195,198,199,200,201,202] | Only to study initial interactions as they are not the target cells | MAP intestinal invasion (cellular and molecular level), early pathogenesis and therapy | |
| Co-culture of cell lines | ✓ | [165,203] | Simple and controllable set up | Early stage pathogenesis |
* P: pathogenesis, D: diagnosis, V: vaccination, T: therapy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



