
Submitted:
15 August 2023
Posted:
16 August 2023
You are already at the latest version
Abstract
In Argentina there are urban areas of intensive cultivation around most of the big cities, where the soils may be unsuitable for some of the horticultural species and the use of chemical fertilizers is common. Bioinoculants based on plant growth promoting rhizobacteria (PGPR) could be an al-ternative for crop improvement, without affecting the environment. These formulations are gen-erally liquid, and their main disadvantage is the low viability of microorganisms. The objectives of this work were to evaluate a different inoculation method, the application of the biofilm pro-duced by Bacillus subtilis as a growth promoter on seeds of three varieties of Lactuca sativa. Biofilm was obtained under static culture conditions, the liquid (planktonic) inoculum was produced at 150 rpm. The major effects were observed with Bacillus subtilis subsp. spizizenii, that showed an-tifungal activity against phytopathogens, synthesize plant growth regulators (IAA, cytokinin and ABA) and solubilize phosphates. The best results were obtained with the Grand Rapid variety inoculated with biofilm, its positive effect was throughout the plant development until the har-vest time, when the bacterium was recovered from the interior of the roots. The biofilm of B. sub-tilis subsp. spizizenii would be an innovative and promising seed inoculant for horticultural spe-cies.
Keywords:
bacterial biofilm
; Bacillus subtilis
; Lactuca sativa
; PGPR
1. Introduction
Agriculture is considered one of the essential activities for humans, not only because of its primary function in food supply but also because of its economic, social, and environmental importance. In Argentina, because of its extensive territory and the asymmetry in the distribution of the population, around most of the large cities there are areas dedicated to the intensive cultivation of vegetables, the so-called urban horticultural strips or green belts, generally made up of small or medium farms (1 to 40 hectares), which main advantage is the proximity to consumer markets. Various horticultural species are produced in them, in some cases, in unsuitable climatic conditions.
Urban and peri-urban horticulture demand soils of high and sustained fertility, with a low incidence of pathogens and very low concentrations of pollutants to guarantee high production rates and optimal final quality of the product to be commercialized. However, urban soils often do not constitute the ideal substrate for horticultural activities. These is because they present poor structure, are formed by layers of different origins, and in some cases have high concentrations of heavy metals [1,2].
Another aspect to consider in these intensive crops is the use of inorganic fertilizers. Of the total greenhouse gases (GHG) generated worldwide, the agricultural sector contributes 65-80% annually, mainly due to the excessive use of nitrogenous fertilizers [3,4]. Thus, the implementation of agronomic techniques that avoid or reduce pollution and gaseous emissions, improve the soil physical and chemical conditions, and the nutrition, yield, and harmlessness of crops, are essential to achieve sustained development.
In this sense, the incorporation of microorganisms into the soil, using bioinoculants, as a promising ecological and friendly alternative to improve the stability of ecosystems and the physical and chemical conditions of soils, is becoming increasingly relevant [5]. Among these microorganisms, a group of beneficial bacteria for plants called Plant Growth Promoting Rhizobacteria (PGPR) [6], are capable of surviving and proliferating in the presence of the autochthonous soil microbiota. These bacteria develop their PGPR activity through different mechanisms, which include their action as biofertilizers, phytostimulators, biocontrolers, rhizoremediators and stress controllers. The action as biofertilizers favors the acquisition of nutrients and improves the structure of degraded soils through biological nitrogen fixation [7], phosphates solubilization and mineralization [8], potassium mobilization or iron sequestration [9]. The phytostimulatory action occurs through the production of phytohormones, such as Indole Acetic Acid (IAA), gibberellins, cytokinins, among others, which modify morphogenesis and cell proliferation in plant tissues [10,11]. These bacteria also act as biocontrolers by releasing antibiotics, lytic enzymes, and other metabolites capable of controlling the proliferation of pathogenic microorganisms in the soil, favoring the development of plants [12,13].
Bacillus is a genus of interest as PGPR, since it presents a wide physiological diversity that allows it to be present in different habitats, both terrestrial and aquatic. Bacteria belonging to this genus are recognized for their action as biofertilizers, phytostimulants, and biological control agents since they produce more than two dozen antibiotics, predominantly protein [14,15]. There are several species of Bacillus, including B. subtilis subs. spizizenii and B. subtilis var. natto.
Another important feature of this genus is its ability to produce biofilm. In the laboratory, depending on the culture conditions, Bacillus subtilis can develop biofilms at the air-liquid interface or grow as the free-living planktonic form [16]. Bacterial biofilms consist mainly of bacteria in a three-dimensional exopolysaccharide matrix, with lesser amounts of protein, DNA, and lysate products [17,18]. During its formation, lipopeptides are induced and accumulate, some with antibacterial and antifungal properties [19]. In nature, biofilm represents a protected mode of growth that spares cells from environmental fluctuations in humidity, temperature, pH, nutrients concentrations, and facilitates cell waste removal [20].
On the other hand, the genus Bacillus presents versatility for its application, making it an excellent candidate for the development of bioinoculants [6]. On the market, bioinoculants exist mainly as liquid and solid supported formulations. In Argentina, most of the formulations with bacteria are in liquid form, with the bacteria in planktonic form. The main problems with this type of formulations are the low viability of the bacteria and the need to store them at low temperatures.
One common crop in the horticultural belts is Lactuca sativa. Worldwide, the main producing countries of Lactuca sativa are China, the USA, Spain, Italy, and India [21]. It is one of the most important vegetables in the leafy group, and the third largest crop after potatoes and tomatoes. There are several varieties of Lactuca sativa, its characteristics and its widespread use make it a good vegetable in which to test the capacities of an inoculant.
The objective of this work was to evaluate the biofilm of Bacillus subtilis as a growth promoter in different varieties of Lactuca sativa, comparing it with the effect of the traditional liquid inoculation (planktonic cells) of the bacterium.
2. Materials and Methods
2.1. Microorganisms and Culture Media for Strain Activation
The bacteria Bacillus subtilis subsp. spizizenii and Bacillus subtilis var. natto were obtained from the collection of Faculty of Agronomy of Buenos Aires University and, the phytopathogens Fusarium solani and Pythium ultimum were provided by San Pedro INTA Experimental Station (Plant Pathology Laboratory). The bacterial strains were activated in nutritive agar media (Merck) at 30 °C for 24 h and, fungi were seeded in Potato Dextrose Agar (PDA) (Merck) and incubated at 25 °C for 10 days.
2.2. Antifungal Activity of Bacillus
The bacteria Bacillus subtilis subsp. spizizenii and Bacillus subtilis var. natto, previously activated, were grown on PDA at 30 °C and 150 rpm. Aliquots were taken at 24, 48, 72 and, 96 h and were centrifugated at 3500 g, separating the cell package and supernatant.
Spore suspension of Fusarium solani and Phytium ultimum were spread uniformly on Petri dishes with PGA (Merck). Then, sterile filter paper dishes of 0.5 cm were deposited and instilled twice with 10µl of bacterial supernatants. The plates were incubated at 25 °C for 7 days. The inhibition of fungal growth was evaluated by measuring the diameter of inhibition halo. The procedure was performed in triplicate.
2.3. Growth Curves of Bacillus Strains in Different Carbon Sources
Bacillus subtilis subsp. spizizenii and Bacillus subtilis var. natto were cultivated in Minimal Salts Medium (MSM), containing 1 g/L K2HPO4, 0.3 g/L KH2PO4, 0.5 g/L NH4Cl, 0.1 g/L NH4NO3, 0.1 g/L Na2SO4, 0.01 g/L MgSO4.7H2O, 1 mg/L MnSO4.4H2O, 1mg/L FeSO4.7H2O, 0.5 g/l CaCl2, 0.01 g/L EDTA, 1L deionized water pH=7±0.4 [22] with 35mM L-glutamic acid and, 1% glucose or 1% glycerol as carbon source. The bacteria were incubated in rotary shaker incubator (New Brunswick Scientific Edison) at 150 rpm and 30 °C. Aliquots of 5 ml were taken at 0, 24, 48, 72 and, 96 h to measure the optical density of the culture using spectrophotometer (Mettler Toledo) at a wavelength of 610 nm. The assay was performed by triplicate.
2.4. Co-Cultures of Bacillus and Phytopathogen Fungi
The bacterium Bacillus subtilis subsp. spizizenii was cultivated during 24 h in MSM with 1% glycerol and 35 mM L-glutamic acid at 30 °C and 150 rpm. Then, 5 buttons of 1-cm in diameter of PGA with fungi growth corresponding to Fusarium solani or Phytium ultimum were added to each Erlenmeyer. These co-cultures of were incubated at 30 °C during 96 h. From these co-cultures, aliquots of 5 mL were extracted every 12 hours to measure optical density using spectrophotometer (Mettler Toledo) at a wavelength of 610 nm.
In stationary phase of bacteria, aliquots of 5 mL of co-cultures were taken every 10 hours to measure antifungal activity and pH.
2.5. Quantification of Plant Growth Regulators Produced by Bacillus subtilis subsp. spizizenii
The bacterium was grown in MSM with 1% glycerol and 35 mM L-glutamic acid at 30 °C and 150 rpm for 96 hours, reaching a concentration of 1.108 CFU/mL. Vials containing 5 mL of this culture were lyophilized and used to determine of plant hormones by liquid chromatography. For extraction process, the modified Bieleski solvent was used: MeOH-HCO2H-H2O 15:1:4 (v/v/v). The lyophilized material was resuspended with cold extraction solvent and homogenized for one hour in the cold. It was centrifugated at 13,000 g during 20 minutes at 4 °C. In order to remove pigments and lipids the extracts were filtrated in Sep-Pak Plus C18 columns. Subsequently, it was evaporated under vacuum at 40 °C near dryness. For the purification, the method of Dobrev [23] was used. Extracts were diluted with 5 mL of 1 M formic acid and passed through OASIS MCX columns. The column was washed with 5 mL of 1 M formic acid. Abscisic acid and indole-acetic acid were eluted with 5 mL of methanol. The phosphate riboside cytokinins were then eluted with 5 mL of 0.35 M ammonia in water. Subsequently, the basic cytokinins, ribosides and glycosides were eluted with 5 mL of 0.35 M ammonium in 60% v/v of methanol. Finally, the solvents were evaporated in rotovap at 40 °C. The samples were dissolved in 100 µL of acetonitrile: water (50:50) (v/v). Then, 5 µL were injected in Agilent 1100 Series HPLC (High Performance Liquid Chromatography) through an Eclipse XDB-C18 column at a flow rate of 0.5 mL/min using a linear gradient of acetonitrile (B) in 0.0005% v/v acidified water with acetic acid (A): 10% B for 5 minutes, 17% B for 10 minutes, then 50% B for 11 minutes, finally increased to 90% B and maintained for 5 minutes. The areas were read at a wavelength of 270 nm. Standards purchased from Sigma were used for calibration, and the retention times were ABA 16.8 minutes, IAA 16.4 minutes, trans zeatin (tZ) 5.5 minutes and trans zeatin riboside (tZR) 13.9 minutes.
2.6. Phosphate Solubilization in Liquid Medium
A culture of Bacillus subtilis subsp. spizizenii in broth NBRIP (National Botanical Research Institute’s phosphate growth medium) [24] was used. Samples of 1.5 mL were taken at 24, 48, 72 and, 96 h of culture, they were centrifuged in a microcentrifuge at 12,000 rpm and the supernatants were used for phosphate determination through vanadate-molybdate method [25] by spectrophotometer at 460 nm.
2.7. Growth Promotion Assays
Bacillus subtilis subsp. spizizenii was cultivated in liquid MSM with 1% glycerol and 35 mM L-glutamic acid. For the production of bacteria in a planktonic state, they were cultivated at 30 °C with agitation at 150 rpm for 96 h, in a rotatory agitation incubator (New Brunswick Scientific Edison), obtaining a count of 1.108 CFU/mL. For the production of biofilm, the culture was maintained under static conditions at 30 °C for 96 h, obtaining a count of 1.109 CFU/g. In both cases, the bacterial counts were carried out by technique of dilution in physiological solution and spread on a plate with nutritive agar [26].
2.8. Seed Germination Assay
Seeds of Lactuca sativa (lettuce) of three varieties were used: Waldman´s Green, Crimor and Grand Rapid. The seeds were disinfected by washing with 70% alcohol and then three times with sterile distilled water. A layer of cotton covered with sterile filter paper with the pore size equivalent to Whatman Grade 3 was placed in sterile Petri dishes and moistened with 5 mL of sterile distilled water. Ten seeds were placed in each Petri dish. Each seed was inoculated with 0.1 mL of planktonic culture and maintained under dark conditions at 22 °C. The seeds that received the treatment with bacterial biofilm were mixed with it due to its great adherence on the seeds. Simultaneously, a control with seeds that received 0.1 mL of distilled water was performed.
A completely randomized design with three replicates per treatment were used. Observations were made 4 and 7 days after inoculation without uncovering the boxes, and a visible radicle length of at least 2 mm was the criterion for germination [27]. The Germination Percentage (G%) was determined according to Araya [28].
At 15 days, the length of hypocotyl and the radicle were measured in each seedling.
2.9. Greenhouse Assays
Seeds of L. sativa of Crimor and Grand Rapid varieties were soaked for 15 minutes with planktonic inoculum or biofilm. Subsequently, they were sown in seedling trays with cells of 5-cm in diameter and 10-cm in depth with commercial substrate and compost (3:1). Twenty-five seeds were placed at each treatment. The assay was carried out at an average temperature of 20 °C. After 25 days, the seedlings were harvested and separated in aerial biomass and root, cutting each seedling at the height of the neck. Each part of seedling was weighed (using an analytical balance with 0.0001g precision) as fresh biomass due to its small size.
On the other hand, seeds of L. sativa of Grand Rapid variety were inoculated with planktonic inoculum or biofilm and sown in seedling trays as describe above. After 20 days, the seedlings were transplanted into 2 L pots, placing one seedling per pot and filling with the same substrate (commercial substrate and compost, 3:1). The assay was carried out at greenhouse at the Faculty of Agronomy (University of Buenos Aires) (34º45´S, 60º31´W) at an average temperature of 24 °C. Twenty-five plants were used per treatment. After 60 days, the plants were harvested and dried in an oven at 70 °C until constant weight. Then, the plants were separated in aerial biomass and root, and each part was weighed.
2.10. Statistical Analysis
The simple ANOVA test was used. Means were compared using Tukey's test at a significance level of 0.05.
3. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.1. Bacillus As a Biocontrol Agent
3.1.1. Antifungal Activity of Liquid Culture Supernatants of Bacillus subtilis subsp. spizizenii and Bacillus subtilis var. natto
For both Bacillus strains, the synthesis of water-soluble metabolites with the capacity to inhibit the growth of the fungi F. solani and P. ultimum was observed after 48 hours of culture (Table 1). The maximum antifungal activity of B. subtilis subsp spizizenii was at 96 h of culture, being similar for both fungi (15, 9 ± 2,1 mm for F solani and 16,2 ± 1,4 mm for P. ultimum). In the case of B. subtilis var natto, there were not significant increase of the antifungal activity after 48 h of culture. The antifungal activity of B. subtilis subsp. spizizenii was always higher than that of B. subtilis var. natto, at 96 h of culture B. subtilis subsp. spizizenii was 274% higher for P solani and more than 352% for P. ultimum, with respect to B. subtilis var. natto in both cases.
3.1.2. Growth and Antifungal Activity of Bacillus subtilis subsp. spizizenii and Bacillus subtilis var. natto in Simple Liquid Media
Both bacteria were also capable of synthesizing antifungal metabolites in simple culture media, using glucose or glycerol as carbon source (Table 2). In those media, the antifungal activity was detected after 72 h of culture, which correspond to late stationary phase (Figure 1). Bacillus subtilis subsp. spizizenii showed its maximum antifungal activity at 96 h of culture, while it remained constant for Bacillus natto. Like in complex medium, B. subtilis antifungal activities were always superior to B. natto. For both bacteria, the fungal growth inhibition with glucose or glycerol was similar, with a halo of approximately 15 mm, although the growth in glucose was the double than in glycerol (Figure 1). Glycerol is a promising carbon source for a biofertilizer production, being glycerol a disposal of biodiesel industry its use may reduce biofertilizer manufacturing costs. For these reasons, following assays were made with Bacillus subtilis subsp. spizizenii in simple media with glycerol as carbon source.
3.1.3. Effect of Co-Culture of Bacillus subtilis subsp. spizizenii with Phytopathogenic Fungi
The presence of phytopathogens fungal buttons (P. ultimum or F. solani) in the culture medium of B. subtilis subsp. spizizenii did not affect the bacterial growth (Figure 2a) but caused an increase of pH media (Figure 2b); after 60 h of culture the pH of bacterium alone was 6.5 ± 0.3 mm while with P. ultimum button was 8.8 ± 0.3 mm and 8.6 ± 0.2 mm with F. solani button.
Antifungal activities of the cell free supernatants of Bacillus subtilis subsp. Spizizenii cultures were also not affected by the presence of the fungi in the culture; being the inhibitory effect of cell-free supernatants of the bacteria cultured alone similar to that of the cocultures with F. solani or P. ultimum. The antifungal activities were detected at 72 h of culture, being maximum at 96 h (Table 3).
3.2. Biofertilization Mechanisms
3.2.1. Quantitative Evaluation of Inorganic Phosphorus Solubilization by B. subtilis subsp. Spizizenii
3.2.2. Synthesis of Plant Growth Regulators by Bacillus subtilis subsp. Spizizenii
Indole acetic acid (IAA), the cytokinins trans zeatin T (tZ) and trans zeatin riboside (tZR) and abscisic acid (ABA) production in a culture of B. subtilis subsp. Spizizenii in its stationary phase was quantified. The mean IAA concentration was 0.38 µg/mL. Among the cytokinins, only the synthesis of tZ was detected (0.14 µg/mL), and the average concentration of ABA was 0.29 µg/mL (Table 4).
3.3. Effects of Bacillus subtilis subsp. Spizizenii Inoculation As Planktonic Form or Biofilm on Three Varieties of Lactuca sativa
3.3.1. Effects on Seed Germination
The effect of the bacterium on three varieties of Lactuca sativa was tested. For all varieties, the non-inoculated seeds reached their maximum germination after four days (Table 5). The Crimor variety presented the lowest germination capacity; for this variety, the planktonic inoculum caused a 12% increase in germination. For Waldman´s Green and Grand Rapid varieties planktonic inoculum had no effect. The inoculation with biofilm caused a delay in the germination of all lettuce varieties, after 4 days none of the seeds had germinated, however, after 7 days Waldman´s Green and Grand Rapid reached the control values, while for Crimor variety germination was similar to that of the planktonic inoculum.
3.3.2. Effects on Seedlings
Lactuca sativa seed inoculation with B. subtilis subsp. Spizizenii had a positive effect on seedlings for Crimor and Grand Rapid varieties, the positive effect was depended on the mode of application. The greatest effect was on the seeds of the Grand Rapid variety. Compared with the control, the inoculation of plankton or biofilm produced an increase in radicle length of 30% and 53%, respectively (p<0.05). Hypocotyl development was also higher with both types of inoculation (p<0.05), showing increases of 19% with respect to the control for planktonic inoculation and 31% with biofilm. The biofilm application was significantly more effective than the planktonic one, showing an increase of 17% in the radicle and 10% in the hypocotyl (Figure 4).
In the Crimor variety as in Grand Rapid varieties, the effect was also dependent on the form of inoculation. Compared to control, inoculation with plankton or biofilm produced a 30% and 40% increase in radicle length, respectively (p<0.05), while hypocotyl length increases were 14% and 26% respectively (p<0.05). As in Grand Rapid, the application of the biofilm turned out to be more effective than the planktonic inoculation, with increases of 8% in the growth of the radicle and 10% in the development of the hypocotyl (Figure 4).
In the case Waldman's Green, although the differences with the control were not significant, it showed an increase of 20% in the radicle for both inoculation methods and increases of hypocotyl of 10% for planktonic inoculum and 17% for biofilm inoculum respect to control (Figure 4).
3.3.3. Effect on the Growth of 25-Day-Old Plants of the Crimor and Grand Rapid Varieties
Due to the higher PGPR activities observed with the Crimor and Grand Rapid, the trials continued with these varieties. For both, root development was favored with seed inoculation (Figure 5a). For the Crimor variety, there was an increase of 42% with planktonic inoculation and 57% with biofilm application; there were no significant differences between the inoculation methods. In the case of the Grand Rapid variety, the mode of inoculum application induced differences in root development, with greater growth for biofilm (82%) than for planktonic inoculum (39%); being the former 33% more effective than planktonic inoculum.
Inoculation of seeds with Bacillus. subtilis subsp. spizizenii also exerted a positive effect on the aerial biomass of L. sativa (Figure 5b). In the Crimor variety, an increase of 37% was observed with the planktonic inoculum and 47% with biofilm, there was no significant difference between the inoculation methods. In the Grand Rapid variety, increases of 30% were found with the planktonic inoculum and 77% with the application of the biofilm. As in the case of the roots, the effect of biofilm was 36% higher than with the planktonic inoculum.
3.3.4. Effect on the Growth of Plants of the Grand Rapid Variety at Harvest Time
To determine if the positive effect of the inoculation was prolonged over time, the growth achieved by the Grand Rapid variety at the time of harvest (60 days) was studied. At that time, the positive effects of the inoculation on roots and aerial parts continued (Figure 6). In root development, the application of the bacteria in a planktonic state or as a biofilm showed increases of 58% and 95%, respectively. In the case of the aerial part, the application of the bacteria in a planktonic state or as a biofilm compared to control showed increases of 56% and 86%, respectively. Significant differences between inoculation treatments also persist, with biofilm being the most effective method, showing an increase of 24% in root development and 20% in the aerial part compared to planktonic inoculation.
3.4. Bacterial Endophytism
The presence of B. subtilis subsp. spizizenii in the roots of L. sativa var. Grand Rapid was evaluated at 60 days of seed inoculation. It was detected 4. 104 CFU/g of fresh root weight in roots from seeds inoculated with biofilm, while no bacteria were recovered in control roots.
4. Discussion
Sustainable soil use motivates the development of technologies that allow the production of quality crops with the least possible impact on the ecosystem. To fulfill this advance, application of bioinoculants based on free living forms of plant growth promoting rhizobacteria (PGPR) have been developed. In this work, an alternative form of bioinoculant for L. sativa, based on a bacterial biofilm is proposed, in which a PGPR bacterium is protected from the environment.
PGPR bacteria are known to have a positive interaction with plant roots, directly by influencing plant growth or indirectly by modifying the rhizosphere environment [29]. This last effect could be mediated by the release of substances that can act as bio controls, for example, antibiotics, lithic enzymes, and other metabolites able to control the proliferation of soil pathogenic microorganisms, resulting in the improvement of plant development and growth [12,13]. In this study, both, B. subtilis subsp. spizizenii and B. subtilis var. natto showed antifungal activity (measured as inhibition halos) against P. ultimum and F. solani in complex and simple media. Moreover, these metabolites were present in cell free supernatants, implying that they were not attached to the bacterial cell. Walker [30] had similar results with aqueous supernatants of Bacillus subtilis, strains J7, B3 and C1, active against Botrytis cinerea. Also, the antifungal activity was similar in complex medium o in simple media using glucose or glycerol as carbon source. The similar bacterial behavior in different media suggests that this could also happen in natural environments. According to Sullivan [31], in nature the antifungal metabolites would be synthesized when bacterial density is high enough to act against pathogens. Besides, Hultberg [32] showed that the application of these kind of metabolites does not affect the indigenous flora at the rhizosphere level. This fact positions them as an alternative to fungicides of synthetic origin [33].
For both, B. subtilis subsp. spizizenii and B. subtilis var. natto, in all media assayed, antifungal activity was detected in the late phase of bacterial growth, being maximum in the late stationary phase. Other bacterium of the genus Bacillus, Bacillus sp. B209, also synthesize antifungal metabolites in the stationary phase [34]. In all cases, B. subtilis subsp. spizizenii always showed higher fungal activity than B. subtilis var. natto, which made it a better prospect to a bioinoculant. The growth of B. subtilis subsp. spizizenii was not affected by the fungus in the culture, although the pH media were higher in fungus presence. The antifungal activity of B. subtilis subsp. spizizenii were not induced by the present of the fungi in the culture, which suggest that the synthesis of the antifungal metabolites would be innate.
Other types of PGPR effects are associated with the capacity of the microorganisms to enhance nutrient availability for plant growth, for example, phosphates solubilization and mineralization [8], B. subtilis subsp. spizizenii was able to solubilize inorganic phosphate, what is fundamental for plant nutrition. The main phosphate dissolving activity was in the exponential growth, diminishing when the bacterial enters the stationary phase, suggesting that this activity could be regulated in the bacterium. The development processes in plants are controlled by internal signals that depend on the adequate supply of minerals through the roots; because of that, the action of PGPRs by increasing the levels of different minerals in the tissues of various plant species contributes to growth of plants [35,36].
Another way in which a microorganism can act as a PGPR is through the production of phytohormones, such as indole acetic acid (AIA), gibberellins, cytokines, which modify the plant morphogenesis and cellular proliferation [10,11]. In this work, metabolites with plant growth activity (IAA and cytokinin) were detected in the stationary phase of growth of B. subtilis subsp. spizizenii in minimum medium, with glycerol and L-glutamic acid as carbon and nitrogen source respectively. These plant growth factors could be easily assimilated by the plant, being phytostimulation one of the possible mechanisms of this bacterium to promote plant development. In agreement, other authors attribute the PGPR effect exerted by certain bacteria on different plant species to the production of IAA, as indicated by Valero [37] for Oryza sativa L. plants inoculated with Pseudomonas sp. Likewise, Ahmed [38] worked with IAA-producing Bacillus strains and found increases of 40% in Solanum tuberosum seedlings inoculated with the bacteria. Moreover, auxin and cytokinin-type phytohormones favor root development [39,40].
Its various PGPR actions (synthesis of antifungal metabolites and plant growth hormones, and its ability to dissolve phosphates), make B subtilis subsp. spizizenii a good candidate for the production of bioinoculants. Bioinoculants are formulations based on one or more PGPRs microorganisms. Currently, most of these products are marketed in liquid form (the microorganisms are in their free-living state, also called planktonic), so they require contact times with the seeds, expressed as imbibition time, which vary from 15 minutes to several hours [36,41]. The main challenge of this type of formulation is that they must have a high number of viable cells that must be preserve over time to hold its effectiveness throughout the entire marketing chain. Besides, when the bioinoculant is used in seeds there must be a close PGPR microorganism-seed interaction, which is generally achieved by adding substances, making it more expensive.
Considering these two aspects, cell viability and close interaction between microorganism and seed, the bacterial biofilm would be an innovative and promising proposal. In this sense, B. subtilis subsp. spizizenii, can form a biofilm [42], a characteristic that differentiates it from other microorganisms used as a bioinoculant. The cells of the biofilm are held together by an extracellular matrix composed of exopolysaccharides, proteins, and nucleic acids [43]. The exopolysaccharide network would provide an anchoring site that would protect the bacterial cells. In addition, the bacteria would be in permanent contact with the seed, the physicochemical characteristics of the biofilm allow it to function as a mucilage, enabling greater adherence to the seed, with a longer seed-microorganism contact time. Gonzalez [44] found that the presence of a mucilage in seeds can favor the adhesion of microorganisms and the assimilation of the products synthesized by them, as is the case of gibberellins or other hormones that facilitate germination. Furthermore, this matrix can be degraded by the biofilm microorganisms using it as a nutrient source, which would increase their survival.
Linked to the use of biofilm as bioinoculant, another aspect to consider is that the growth-promoting effect exerted by bacteria depends on the relationship between the bacterial strain and the plant species; a bacterium may show excellent effects on one plant species but not on another. This plant-microorganism interaction is so particular that different responses have been observed with the application of the same microorganism between two varieties of the same plant species [45]. Each plant species produces its own chemical molecules that attract certain microorganisms (chemotaxis) and generate the appropriate conditions for their establishment and multiplication [46]. Bacillus mycoides showed a marked PGPR effect on papaya, rice, cassava and sunflower seeds, with an increase in the germination in all cases, however, it did not have a beneficial effect on L. sativa seeds. Even more, other microorganisms, such as Trichoderma harziarum, Enterobacter aerogenes and Microbacterium sp., showed deleterious effects on L. sativa, although were effective PGPR on other plant species [44]. These facts emphasize the great specificity that exists between the microbial strain and the plant species. In the case of L. sativa, literature cites that bacterium belonging to the genera Hafnia and Beijerinckia as PGPRs. Díaz-Vargas [47] indicated that the strains HP-3, HP-27 corresponding to Hafnia alvei and the strain S4BE of the genus Beijerinckia showed increases greater than 50% in germination tests.
The International Seed Testing Association (ISTA) established that the germination process is a metabolically active state, physiologically manifested by cell division and differentiation, being the emergence of the radicle considered a sign of seed germination. According to this, it was studied the effect of B. subtilis subsp. spizizenii inoculation, in its planktonic or biofilm form, on the radicle emergency of three varieties of L. sativa: Crimor, Waldman´s Green and Grand Rapid. The effect of both types of inoculation form on germination was dependent on the L. sativa variety, with a positive effect only for Crimor, which is probably due to its low germination power, unlike the high power of the other varieties. This delay may be due to the biofilm could function initially as a barrier between the seed and the environmental signals (humidity, temperature, gaseous environment) that induce germination [48]. Once this barrier is overcome, the exopolysaccharide matrix of the biofilm would allow closer contact between the bacteria It should be noted that although the application of the biofilm delayed the germination of all seed varieties, it had no negative effect on any of them, reaching similar germination percentages in all cases after 7 days. and the seed, favoring their interaction.
The effect of seed inoculation on plant development was also dependent on lettuce variety. The major effect was on Grand Rapid, lesser for Crimor variety and none for Waldman`s Green. In the case of Grand Rapid, the positive effect of seed inoculation was prolonged in time throughout the vegetable development: from the formation of the radicle and the hypocotyl, through the plant growth until the harvest time; always having the biofilm a superior effect than the planktonic inoculum. Both, roots and aerial part development, were positively affected, thus, roots development favored by bacteria inoculation increased the ability of the root system to absorb nutrient, allowing a greater growth of the aerial part of the crop. These results agree with those reported by Pereira [49] and Kloepper [29], growth-promoting bacteria such as P. fluorescens are characterized by increasing root development, that directly affects crop yield. Díaz-Vargas [47] informed that the liquid inoculant of Pseudomonas aeruginosa SP5 and Azospirillum brasilense T2P010 in Lactuca sativa, showed stimulation both, over germination and over the vegetative development of the crop. Furthermore, these plant growth effects could also be related to the production of a metabolite with plant growth activity (IAA and cytokinin) and phosphate solubilization as previously indicated. The development process in plants is controlled by internal signals that depend on the adequate supply of minerals through the roots; because of that, the action of PGPRs by increasing the levels of different minerals in the tissues of various plant species contributes to growth. of plants [35,36]. Therefore, considering that auxin and cytokinin-type phytohormones favor root development [39,40] and allow the plant to access a greater amount of minerals, we could assume that these would be other mechanisms of action that B. subtilis subsp. spizizenii to exert its growth-promoting activity on L. sativa.
In this work, B. subtilis subsp. spizizenii was recovered from the interior of the roots of Grand Rapid inoculated with biofilm, which indicates that the bacterium was able to leave the biofilm, get into and proliferate in the interior of L. sativa roots, persisting until the harvest time. Initial bacterial infection and colonization does not mean that it will continue over time. Wulff [50] observed in cabbage plants after 10 days of inoculation with B. subtilis, the presence of the bacterium in root tissues (abundantly) and in its aerial part. However, after 35 days they did not detect the bacterium in any plant tissue. Other authors also detected a decrease in the bacterial population of the inoculated microorganism, Kloepper [51], when examining the ability of 6 strains with PGPR activity to internally colonize cucumber roots, found that bacterial populations fell after 21 days of inoculation. Lamb [52] found that Pseudomonas aureofaciens did not remain viable inside maize plants grown in hydroponics, however, when the plant grew in soil, the bacterium was able to persist inside the plant. According to Jacobs [53], the environment within host cells may be repressive enough to restrict the growth rate of endophytic bacteria.
To summarize, in this work, we were able to verify that biofilm of B. subtilis subsp. spizizenii can act as a biofertilizer and phyto-stimulator of L. sativa, as well as being a producer of metabolites with proven antifungal activity against common soil phytopathogens that attack this vegetable.
5. Conclusions
Seed inoculation with biofilm of Bacillus subtilis subsp. spizizenii was superior to the traditional inoculation (planktonic form) for Lactuca sativa. The positive effect of biofilm inoculation was observed throughout the lettuce development, from the formation of the radicle and the hypocotyl until the moment of harvest, persisting the bacterium inside the roots. These results suggest that Bacillus subtilis subsp. spizizenii biofilms would be a promising proposal as seed inoculants for horticultural species.
Author Contributions
Conceptualization, Gabriela Cristina Sarti, Mirta Esther Galelli, Antonio Paz-González; methodology, Gabriela Cristina Sarti, Silvana Arreguini; Josefina Ana Eva Cristóbal-Miguez, Antonio Paz-González; formal analysis, Gabriela Cristina Sarti, Mirta Esther Galelli, Josefina Ana Eva Cristóbal-Miguez; Antonio Paz-González; investigation, Gabriela Cristina Sarti, Mirta Esther Galelli, Josefina Ana Eva Cristóbal-Miguez; resources, Gabriela Cristina Sarti, Antonio Paz-González; writing—original draft preparation, Gabriela Cristina Sarti, Mirta Esther Galelli, Silvana Arreguini, Antonio Paz-González; writing—review and editing, Gabriela Cristina Sarti, Mirta Esther Galelli, José Alfredo Curá, Antonio Paz-González; supervision, Gabriela Cristina Sarti, Mirta Esther Galelli, José Alfredo Curá, Antonio Paz-González; project administration, Gabriela Cristina Sarti; funding acquisition, Gabriela Cristina Sarti. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Universidad de Buenos Aires, Proyectos de Ciencia y Técnica, grant number (UBACyT) N° 20020170100080BA”.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giuffré, L.; Ratto, S.; Marbán, L.; Schonwald, J.; Romaniuk R. Riesgo por metales pesados en horticultura urbana. Cienc. suelo, Argentina. 2005, 23, 1, 101-106.
- Crispo, M.; Dobson, M.C.; Blevins, R.S; Meredith, W.; Lake, J.A. Heavy metals and metalloids concentrations across UK urban horticultural soils and factors influencing their bioavailability to food crops. Environ. pollut. 2001, 288, 117960. [CrossRef]
- Smith, P.; Bustamante, M.; Ahammad, H.; Clark, H.; Dong, E.A.; Elsiddig; H.; Haberl, R.; Harper, J.; House, M.; Jafari, O.; Masera, C.; Mbow, N.H.; Ravindranath, C.W.; Rice, C.; Robledo Abad, A.; Romanovskaya, F.; Sperling, F.; Tubiello, J. Agriculture, Forestry and Other Land Use (AFOLU). In: Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Edenhofer, O., R. Pichs-Madruga, Y. Sokona, E. Farahani, S. Kadner, K. Seyboth, A. Adler, I. Baum, S. Brunner, P. Eickemeier, B. Kriemann, J. Savolainen, S. Schlömer, C. von Stechow, T. Zwickel and J.C. Minx (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA. 2014.
- Lima, L.; Ongaratto, F.; Fernandes, M.; Cardoso, A.; Lage, J.; Silva, L.; Reis, R.; Malheiros, E. Response of pasture nitrogen fertiliation on greenhouse gas emission and net protein contribution of nellore Young Bulls. Animals. 2022, 12, 3173. [CrossRef]
- Sammauria, S.; Kumawat, S.; Kumawat, P.; Singh, J.; Jatwa, T.K. Microbial inoculants: potential tool for sustainability of agricultural production systems. Arch. Microbiol. 2020, 202, 677–693. [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 6, 1583–1594. [CrossRef]
- Pérez, J.; Carmona, S.; Zamudio, E.; Rivera, N.; Calva G. Bioremediation of soils from oil spill impacted sites using biosurfactants producing, native, free-living nitrogen fixing bacteria. Rev. Int. de Contam. Amb. 2017, 33, 105-114. [CrossRef]
- Gomez Ramirez, L.F.; Uribe Velez, D. Phosphorus solubilizing and mineralizing Bacillus spp. Contribute to rice growth promotion using soil amended with rice Straw. Current Microbiol. 2021, 78, 932-943. [CrossRef]
- Das, P.P.; Singh, K.R.B.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.H.; Kumar, A.; Singh, J. Plant-soil-microbes: A tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environ. Res. 2022, 214, 1, 113821. [CrossRef]
- Suzuki, H.; Park, S. Okubo, K.; Kitamura, J.; Ueguchi-Tanaka, M.; Iuchi, S,; Katoh, E.; Kobayashi, M.; Yamaguchi, I.; Matsuoka, M.; Asami, T.; Nakajima, M. Differential expression and affinities of Arabidopsis giberellins receptors can explain variation in phenotypes of multiple knock-out mutants. The Plant Journal. 2009, 60, 48-55. [CrossRef]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR Mediated Alterations in Root Traits: Way Toward Sustainable Crop Production. Frontiers of Sustainable Food Systems. 2021, 4. [CrossRef]
- Sarti, G.; Miyazaki, S. Actividad antifúngica de extractos crudos de Bacillus subtilis contra fitopatógenos de soja (Glycine max) y efecto de su coinoculación con Bradyrhizobium japonicum. Agrociencia. 2013, 47, 373-383.
- Mekonnen, H.; Kibret, M. The roles of plant growth promoting rhizobacteria in sustainable vegetable production in Ethiopia. Chem. Biol. Technol. Agric. 2021, 8, 15. [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: structures, syntheses and specific functions. Molecular Microbiology. 2005, 56, 4, 845-857. [CrossRef]
- Miao, S.; Liang, J.; Xu, Y.; Yu, G.; Shao, M. Bacillaene, Sharp objects consist in the arsenal of antibiotics produced by Bacillus. Cell. Physio. 2022. [CrossRef]
- Sarti, G.; Miguez Cristóbal, J.; Curá, A. Optimización de las condiciones de cultivo para el desarrollo de una biopelícula bacteriana y su aplicación como biofertilizante en Solanum lycopersicum L.var. Río grande. Rev. Protección Veg. Cuba. 2019, 34, 2224-4697.
- Reichard, C.; Parsek, M. Confocal laser scanning microscopy for analysis of Pseudomonas aeruginosa biofilm architecture and matrix localization. Front. Microbiol. 2019, 10, 677. [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thumheer, T. Biofilm matrixome: Extracellular components in structured microbial communities. Trends in Microbiol. 2020, 28, 8, 668-681. [CrossRef]
- Billings, N.; Ramirez Millan, M.; Caldara, M.; Rusconi, R.; Tarasova, Y.; Stocker, R.; Ribbeck, K. The Extracellular Matrix Component Psl Provides Fast-Acting Antibiotic Defense in Pseudomonas aeruginosa Biofilms. PLoS Pathog. 2013, 9, 8, e1003526. [CrossRef]
- Daboor, S.; John, R.; Rohde, A.; Zhenyu, C. Disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate lyase enhances pathogen eradication by antibiotics. J. Cyst. Fibros. 2020, 30, 4-11. [CrossRef]
- Viteri, M.L.; Ghezán, G.; Iglesias, D. Tomate y lechuga: producción comercialización y consumo. Estudio socioeconómico de los sistemas: INTA, Balcarce, Argentina. 2013, 14, 185-190.
- Gerhardt, P.; Murray, R.; Wood, W.; Krieg, N. Methods of general and molecular Bacteriology. American Society for Microbiology. Washington DC. 1994.
- Dobrev, P.; Kaminek, M. Fast and Efficient Separation of Cytokinins from Auxin and Abscisic Acid and Their Purification. J. Chromatogr. A. 2002, 950, 21-29. [CrossRef]
- Nautiyal, S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms C. FEMS Microbiol. Lett. 1999, 170, 265-270. [CrossRef]
- Kitson, R.; Mellon, M.Colorimetric Determination of Phosphorus as molybdivanadophosphoric acid. Industrial and Engineering. Chem. Anal. Edit. 1994, 16, 6, 379-383. [CrossRef]
- Frioni, L. Ecología Microbiana del Suelo. Dpto. de Publ. y Ed. de la Univ. de la Rep., Montevideo Uruguay. 1990, pp 90-94.
- Marti, L. Efecto de la salinidad y de la temperatura en la germinación de semillas de Linonium mansaltrarum. Tesis doctoral, Universidad Politécnica Superior de Valencia. España, Gandía. 2010.
- Araya, E.; Gómez, L.; Hidalgo, N.; Valverde, R. Efecto de la luz y del ácido giberélico sobre la germinación in vitro de (Alunus Acuminata). Agron. Costarricense. 2000, 24, 1, 75-80.
- Kloepper, J.; Zablotowicz, R. Tipping, E.; Lifshitz, R. Plant growth promotion mediated by bacterial rhizosphere colonizer. D.L. Keister y P.B. Cregan (eds.) The rhizosphere and plant growth. Kluwer. Dordrecht. The Netherlands. 1991, 315-326.
- Walker, R.; Powell, A.; Seddon, B. Bacillus isolates from the spermosphere of peas and dwarf French beans with antifungal activity against Botrytis cinerea and Pythium species. J. Applied Microbiol. 1998, 84,791-801. [CrossRef]
- Sullivan, E. Molecular genetics of biosurfactants production. Curr. Opin. Biotech. 1998, 9, 263-269. [CrossRef]
- Hultberg, M.; Alsberg, T.; Khalil, S.; Alsanius, B. Suppression of disease in tomato infected by Pythium ultimum with a biosurfactant produced by Pseudomonas koreensis. Biocontrol. 2010, 55, 3, 435-444. [CrossRef]
- Ambrico, A.; Trupo, M. Efficacy of cell free supernatant from Bacillus subtilis ET-1, an Iturin a producer strain, on biocontrol of green and gray mold. Postharvest Biol. Technology. 2017, 134, 5-10. [CrossRef]
- Cornea, C.; Grebenisan, I.; Mateescu, R.; Campeanu, E. Isolation and characterization of new Bacillus spp. Strains useful as biocontrol agents of plants pathogens. Roumanian Biotechnol. Letters.2003, 8, 1, 1115-1122.
- Mayak, S.; Tirosh, T.; Glick, B. Plant growth-promoting bacteria that confer resistance to water stress in tomatoes and pepper. Plant Science. 2004, 166, 525-530. [CrossRef]
- Turan, M.; Melek, E.; Ertan, Y.; Adem, G.; Kenan, K.; Recep, K.; Atilla, D. Plant growthpromoting rhizobacteria improved growth, nutrient, and hormone content of cabbage (Brassica oleracea) seedlings. Turk. J. Agricul. For. 2014, 38, 327-333. [CrossRef]
- Valero, N. Potencial biofertilizante de bacterias diazotróficas y solubilizadoras de fosfatos asociadas al cultivo de arroz (Oryza sativa L.) Tesis de Maestría. Universidad Nacional de Colombia. 2003.
- Ahmed, A.; Hasnain, S. Auxin-producing Bacillus sp.: Auxin quantification and effect on the growth of Solanum tuberosum. Pure Appl Chem. 2010, 82, 1, 313-319. [CrossRef]
- Vikram, A. Efficacy of phosphate solubilizing bacteria isolated from vertisols on growth and yield parameters of sorghum. Res. J. Microbiol. 2007, 2, 550-559.
- Ortíz-Castro, R.; Contreras-Cornejo, H. Macías-Rodríguez, L.; López-Bucio, J. The role of microbial signals in plant growth and development. Plant Signal. Behav. 2009. [CrossRef]
- Barreto, D.; Valero, N.; Muñoz, A.; Peralta, A. Efecto de microorganismos rizosféricos sobre germinación y crecimiento temprano de Anacardium excelsum. Zonas Áridas 2007 11, 1, 240-250.
- Galelli, M.; Sarti, G.; Miyazaki, S. Lactuca sativa biofertilization using biofilm from Bacillus with PGPR activity. J. Appl. Hortic. 2015, 17,2, 186-191.
- Hobley, L. Giving structure to the biofilm matrix: an overview of individual strategies and emerging common themes. FEMS Microbiol. Rev. 2015, 39, 649-669. [CrossRef]
- González, F.; Fuentes, N. Mecanismo de acción de cinco microorganismos promotores de crecimiento vegetal. Rev. Col. de Ciencias Agr. 2016, 34,1, 17-22.
- Ogata, K.; Arellano, C.; Zúñiga, D. Efecto de diferentes bacterias aisladas de rizósfera de Caesalpina spinosa en la germinación de diferentes especies vegetales culivados. Zonas Áridas 2008, 12, 1, 137-153.
- Dakora, F.; Phillips, D. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil. 2002, 245, 35–47. [Google Scholar] [CrossRef]
- Díaz Vargas, P.; Ferrera-Cerrato, R.; Almaraz-Suárez, J.; Alcántar, G. Inoculación de bacterias promotoras de crecimiento en lechuga. Terra. 2001, 4, 231–238. [Google Scholar]
- Varela, S.; Arana, V. Latencia y germinación de semillas. Tratamientos pregerminativos. INTA Bariloche, Silvicultura en Vivero. Cuadernillo Nº 3. 2011, 3-10.
- Pereira, J.; Cavalcante, V.; Baldani, J.; Dobereiner, J. Sorghum and rice inoculation with Azospirillum sp. and Herbaspirillum seropedicae in field. Plant Soil. 1988, 110, 269-274. [CrossRef]
- Wulff, E.; van Vuurde, L.; Hockenhull, J. The ability of the biological control agent Bacillus subtilis, strain BB, to colonise vegetable brassicas endophytically following seed inoculation. Plant and Soil. 2003, 255, 463-474. [CrossRef]
- Kloepper, W.; Wei, G.; Tuzun, S. Rhizosphere population dynamics and internal colonisation of cucumber by plant growth-promoting rhizobacteria which induce systemic resistance to Colletotrichum orbiculare. In Biological Control of Plant Diseases. Ed. E C Tjamos, G C Papavizas and R J Cook. 1992; pp. 185-191. [CrossRef]
- Lamb, T.; Tonkyn, D.; Kluepfel, D. Movement of Pseudomonas aerofaciens from the rhizosphere to aerial plant tissue. Can. J. Microbiol. 1996, 42, 112-1120.
- Jacobs, M.; Bugbee, W.; Gabrielson, D. Enumeration, location, and characterization of endophytic bacteria within sugar beet roots. Can. J. Bot. 1985, 63, 1262–1265. [Google Scholar] [CrossRef]
Figure 1.
Growth curves of B. subtilis strains grown in MSM, 35mM L-glutamic acid and 1% glycerol or 1% glucose. Data are the mean of three experiments.
Figure 1.
Growth curves of B. subtilis strains grown in MSM, 35mM L-glutamic acid and 1% glycerol or 1% glucose. Data are the mean of three experiments.
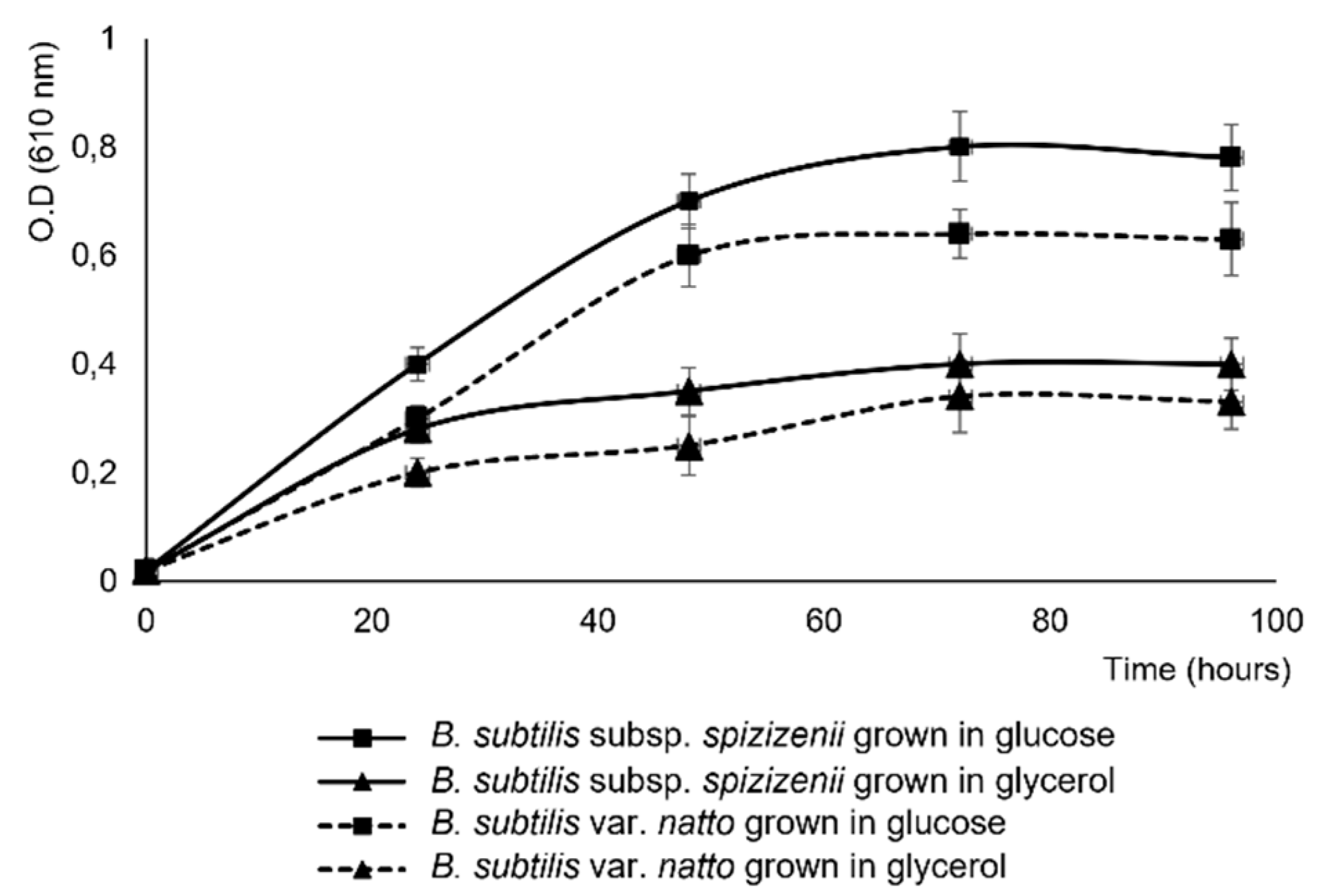
Figure 2.
(A) Growth curve and (B) pH of the media of B. subtilis subsp. Spizizenii cocultured with buttons of P. ultimum or F. solani in MSM, 1%, 35mM L-glutamic acid, 1% glycerol. Data are the mean of three experiments.
Figure 2.
(A) Growth curve and (B) pH of the media of B. subtilis subsp. Spizizenii cocultured with buttons of P. ultimum or F. solani in MSM, 1%, 35mM L-glutamic acid, 1% glycerol. Data are the mean of three experiments.
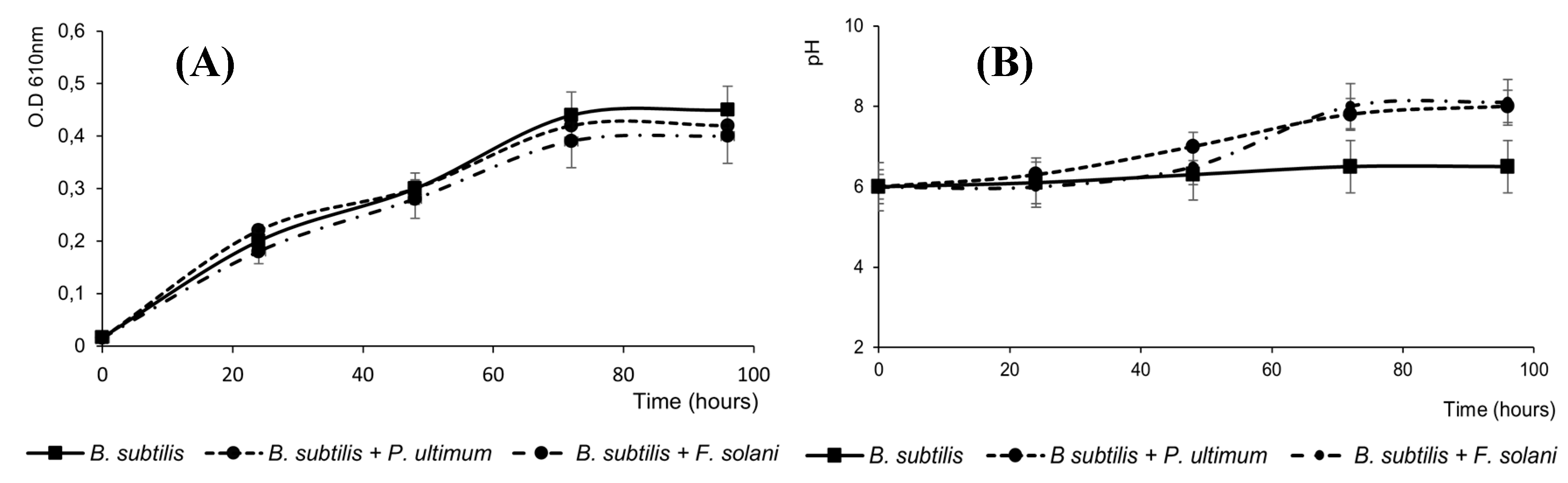
Figure 3.
Solubilization of phosphate by B. subtilis subsp. Spizizenii grown in NBRIP broth. Data are the mean of three experiments.
Figure 3.
Solubilization of phosphate by B. subtilis subsp. Spizizenii grown in NBRIP broth. Data are the mean of three experiments.
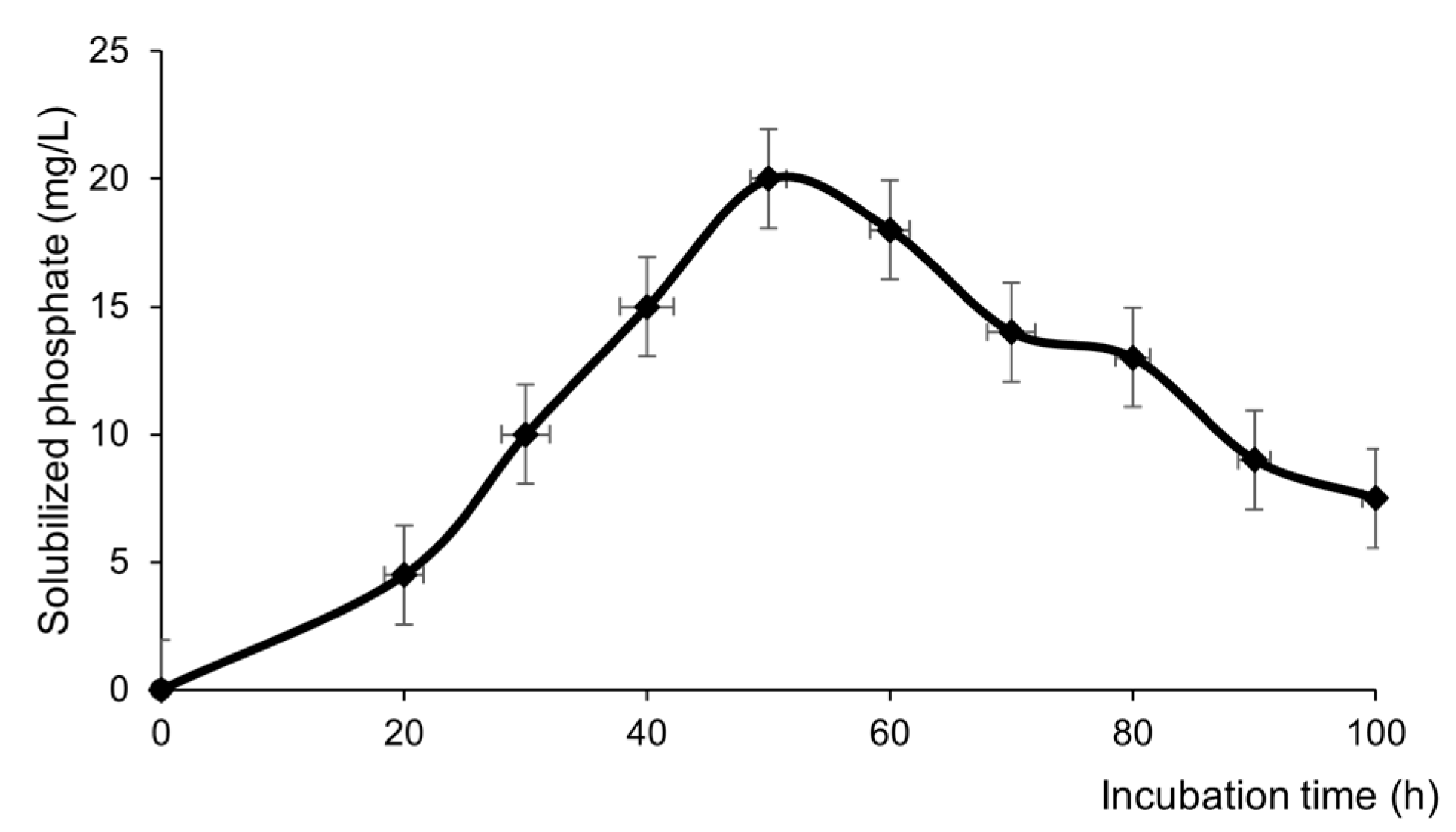
Figure 4.
Morphological parameters of seedlings of different varieties of L. sativa grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. (A) Radicle length. (B) Hypocotyl length. Different letters correspond to significant differences between treatments for each L. sativa variety (p<0.05).
Figure 4.
Morphological parameters of seedlings of different varieties of L. sativa grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. (A) Radicle length. (B) Hypocotyl length. Different letters correspond to significant differences between treatments for each L. sativa variety (p<0.05).
Figure 5.
Biomass of different varieties of L. sativa grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. (A) Root biomass. (B) Aerial biomass. Different letters correspond to significant differences between treatments for each L. sativa variety (p<0.05).
Figure 5.
Biomass of different varieties of L. sativa grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. (A) Root biomass. (B) Aerial biomass. Different letters correspond to significant differences between treatments for each L. sativa variety (p<0.05).
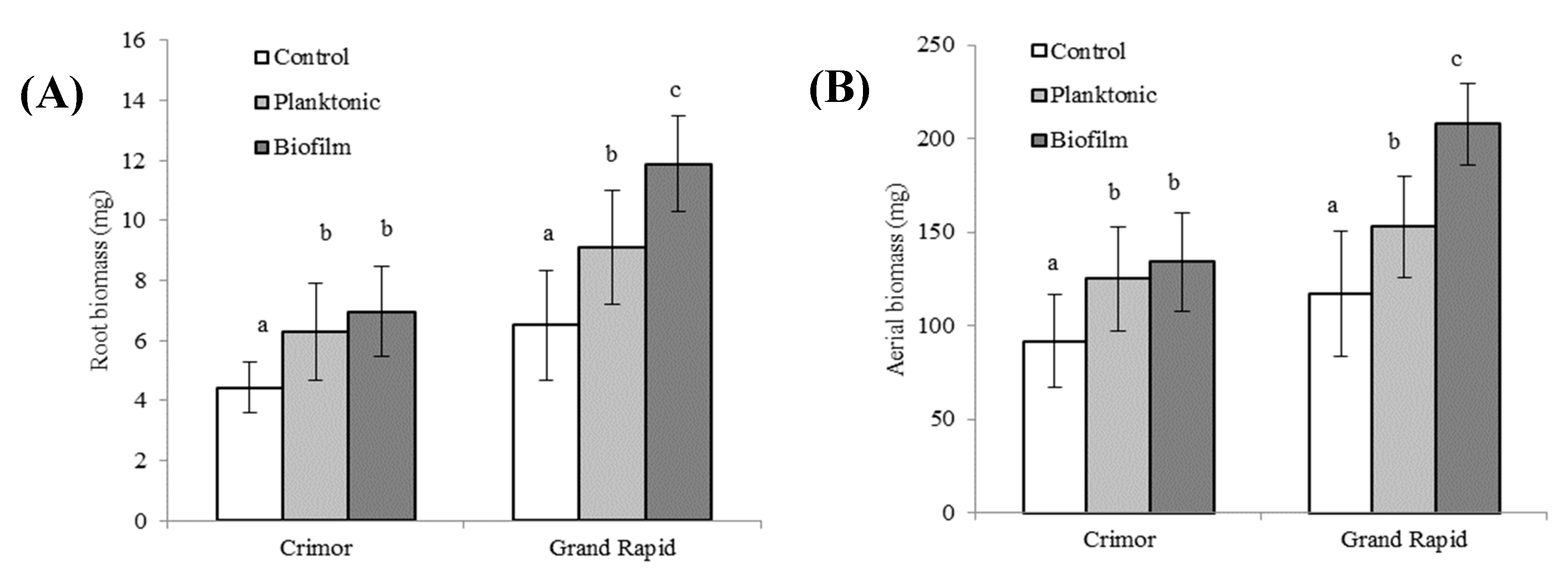
Figure 6.
Biomass of Grand Rapid L. sativa variety grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. Different letters correspond to significant differences between treatments (p<0.05).
Figure 6.
Biomass of Grand Rapid L. sativa variety grown from seeds inoculated with Bacillus subtilis subsp. spizizenii in its planktonic or biofilm form. Different letters correspond to significant differences between treatments (p<0.05).
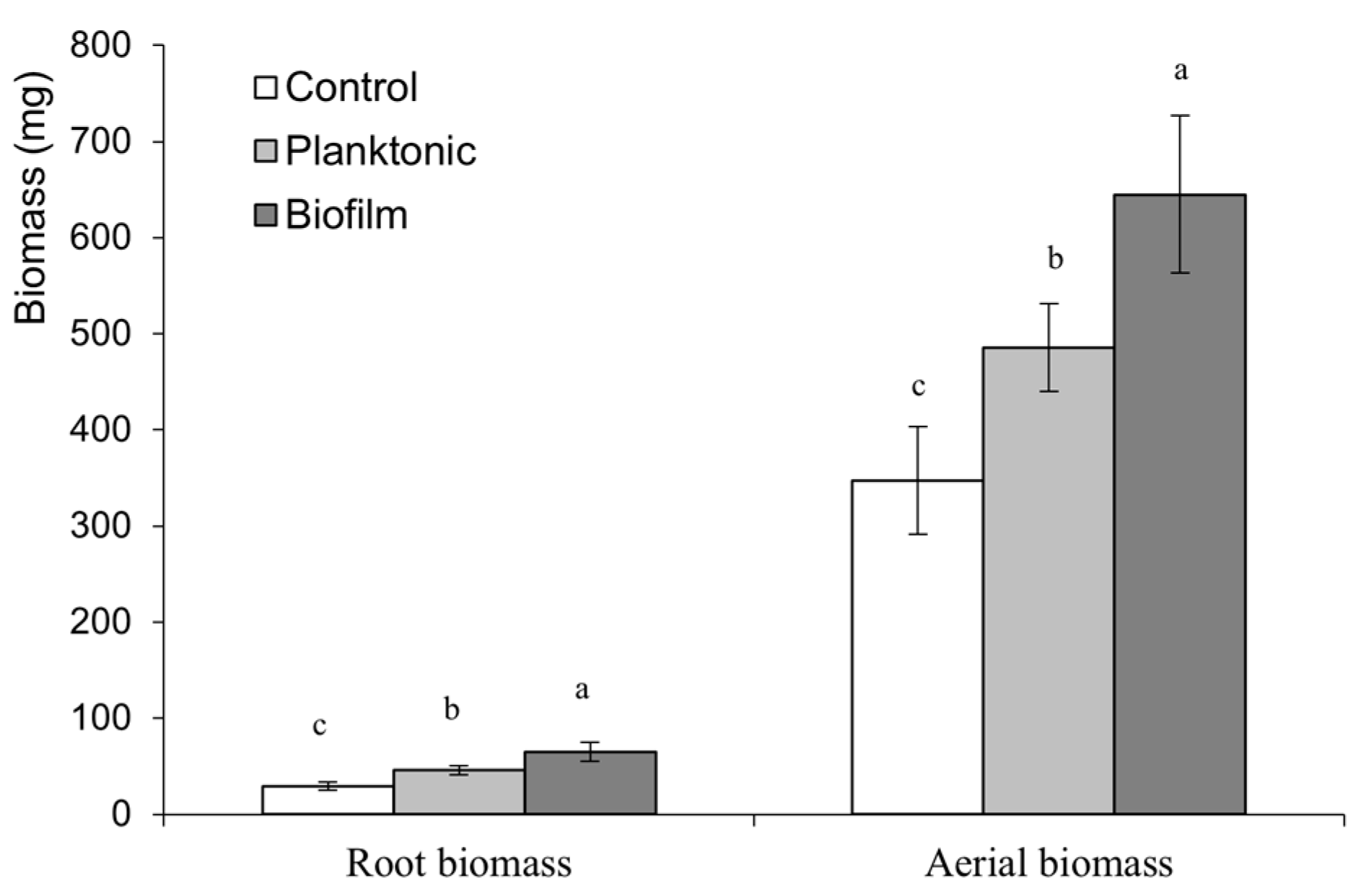

Table 1.
Antifungal activities of cell free supernatants of two B. subtilis strains grown in liquid potato dextrose.
Table 1.
Antifungal activities of cell free supernatants of two B. subtilis strains grown in liquid potato dextrose.
| Inhibition halo diameter (mm) | |||||
|---|---|---|---|---|---|
| Bacteria | Fungus | Incubation time (h) | |||
| 24 | 48 | 72 | 96 | ||
| B. subtilis subsp. spizizenii | F. solani | ND | 6,7 ± 1,8 b | 9,3 ± 1,5 b | 15,9 ± 2,1 a |
| P. ultimum | ND | 6,8± 2,2 b | 8,4 ± 1,8 b | 16,2 ± 1,4 a | |
| B. subtilis var. natto | F. solani | ND | 3,1± 1,8 a | 4,2 ± 2,3 a | 5,8 ± 2,1 a |
| P. ultimum | ND | 2,9± 1,1 a | 3,8 ± 1,0 a | 4,6 ± 1,3 a | |
The antifungal activity was determined as the inhibition halo diameter (mean (mm) ± standard deviation). Different letters in the same row represent significant differences (p<0.05). ND: not detected.
Table 2.
Antifungal activities of cell free supernatants of two B. subtilis strains grown in glucose or glycerol media.
Table 2.
Antifungal activities of cell free supernatants of two B. subtilis strains grown in glucose or glycerol media.
| Inhibition halo diameter (mm) | |||||||
|---|---|---|---|---|---|---|---|
| Culture medium | Bacteria | Fungus | Incubation time (h) | ||||
| 24 | 48 | 72 | 96 | ||||
| Glycerol 1% | B. subtilis subsp. spizizenii | F. solani | ND | ND | 10,2 ± 0,7 a | 15,6 ± 1,2 b | |
| P. ultimum | ND | ND | 12,3 ± 1,1 a | 16,0 ± 0,8 b | |||
| B. subtilis var. natto | F. solani | ND | ND | 5,1 ± 1,4 a | 5,3 ± 1,7 a | ||
| P. ultimum | ND | ND | 6,2 ± 0,9 a | 5,9 ± 1,2 a | |||
| Glucose 1% | B. subtilis subsp. spizizenii | F. solani | ND | ND | 11,2 ± 0,8 a | 14,1 ± 1,1 b | |
| P. ultimum | ND | ND | 13,3 ± 1,1 a | 13,9 ± 2,1 a | |||
| B. subtilis var. natto | F. solani | ND | ND | 4,3 ± 1,8 a | 3,5 ± 1,3 a | ||
| P. ultimum | ND | ND | 5,2 ± 2,3 a | 4,3 ± 1,2 a | |||
The antifungal activity was determined as the diameter inhibition halo (mean (mm) ± standard deviation). Different letters in the same row represent significant differences (p<0.05). ND: not detected.
Table 3.
Antifungal activity of cell-free supernatants of B. subtilis subsp. Spizizenii cultured alone or with P. ultimum or F. solani.
Table 3.
Antifungal activity of cell-free supernatants of B. subtilis subsp. Spizizenii cultured alone or with P. ultimum or F. solani.
| Inhibition halo diameter (mm) | |||||
|---|---|---|---|---|---|
| Culture | Fungus | Incubation time (h) | |||
| 24 | 48 | 72 | 96 | ||
| B. subtilis subsp. Spizizenii | F. solani | ND | ND | 12,2 ± 0,9 a | 15,2 ± 1,2 b |
| P. ultimum | ND | ND | 10,0 ± 1,1 a | 15,4 ±1,7 b | |
| Coculture of B. subtilis subsp. Spizizenii + F. solani | F. solani | ND | ND | 11,2 ± 1,5 a | 11,3 ± 1,1 a |
| P. ultimum | ND | ND | 13,8 ± 1,2 a | 14,1 ± 0,8 a | |
| Coculture of B. subtilis subsp. Spizizenii + P. ultimum | F. solani | ND | ND | 10,9 ± 1,8 a | 14,9 ± 2,1 b |
| P. ultimum | ND | ND | 9,8 ± 1,9 a | 14,7 ± 1,5 b | |
Bacillus were grown in MSM, 35 mM L-glutamic acid, 1% glycerol, with or without the presence of button of the fungi P. ultimum or F. solani. Antifungal activity of the cell free extracts was determined as the diameter of the inhibition halo (mean (mm) ± standard deviation). Different letters in the same row represent significant differences (p<0.05).
Table 4.
Quantification of growth hormones in a culture of B. subtilis subsp. Spizizenii.
| IAA | tZ | tZR | ABA |
|---|---|---|---|
| 0.38 ± 0.14 | 0.14 ± 0.05 | ND | 0.29 ± 0.05 |
Plant growth regulators (µg/mL) synthetized by B. subtilis subsp. Spizizenii grown in MSM, 35 mM L-glutamic acid and 1% glycerol. IAA: indole acetic acid, tZ: trans zeatin T; tZR: trans zeatin riboside; ABA; abscisic acid. ND: not detected.
Table 5.
Germination percentages of L. sativa seeds inoculated with planktonic form or biofilm of B. subtilis subsp. Spizizenii.
Table 5.
Germination percentages of L. sativa seeds inoculated with planktonic form or biofilm of B. subtilis subsp. Spizizenii.
| Germination (%) at 4 days | Germination (%) at 7 days | |||||
|---|---|---|---|---|---|---|
| L. sativa varieties | Control | Planktonic inoculum | Biofilm | Control | Planktonic inoculum | Biofilm |
| Waldman´s Green | 97,3 ± 0,8 a | 96,2 ± 1,4 a | ND | 97,3 ± 0,9 a | 96,2 ± 1,4 a | 96,4 ± 1,1 a |
| Crimor | 80,2 ± 1,3 a | 90,1 ± 2,0 b | ND | 80,2 ± 1,3 a | 90,1 ± 2,0 b | 94,2 ± 2,3 b |
| Grand Rapid | 96,3 ± 1,1 a | 97,1 ± 1,5 a | ND | 96,3 ± 1,4 a | 97,1 ± 0,8 a | 97,3 ± 0,9 a |
Controls correspond to uninoculated seeds. Different letters within each row and at times of 4 days and 7 days correspond to significant differences between treatments (p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



