Submitted:
15 August 2023
Posted:
16 August 2023
You are already at the latest version
Abstract
The aims of the review were to elucidate the spatial variation in sea ice algae primary production rates and biomasses (Chl a) in the Canadian Arctic – Greenland region, characterized by comparable physical settings. A database was compiled from 30 studies of sea ice algae primary production rates, biomasses (Chl a), snow and ice thicknesses, ice types, nutrient (Si(OH)4, PO4, (NO3+NO2), and (NH4) concentrations in ice and below ice from the region. Production rates were significantly high (463 mg C m-2 d-1) in Resolute Bay and in Northern Baffin Bay (317 mg C m-2 d-1) both in Canadian Arctic, as compared to a rate of 0.2 mg C m-2 d-1 in northeast Greenland. The biomasses reached 340 mg Chl a m-2 in Resolute Bay in comparison to a 0.02 mg Chl a m-2 in southwest Greenland. Primary production at other Canadian and Greenland sites were comparable but sea ice Chl a was higher (15.0 ± 13.4 mg Chl a m-2) at Canadian sites as compared to Greenland (0.8 ± 0.5 mg Chl a m-2). Resolute and Northern Baffin Bays production rates were significantly higher when compared to other Arctic Ocean sites outside the studied region. The review concludes that high production rates and biomasses in Resolute and Northern Baffin Bay’s were related to inflow and mixing of nutrient rich waters of Pacific origin. A conceptual model with drivers and inhibitors of sea ice algae primary production is projected, and the database compiles a dataset of published data for further studies.
Keywords:
sea ice algae primary production
; Chl a
; snow and sea ice thickness
; nutrients
; oceanography
; Canadian Arctic Archipelago
; Greenland
1. Introduction
Winter sea ice covers the entire Arctic Ocean, Canadian Archipelago, Hudson and Baffin Bays (Wratten et al., 2022), east and west Greenland (Karami et al., 2021) with a 15.9∙106 km2 in median maximum during March (Francis and Wu, 2022). In contrast, the aerial extent of the summer (June) sea ice in the Arctic Ocean has been decreasing since late 1970 from 12.5∙106 km2 in 1980 to 10.6∙106 km2 in 2022, equal to a loss of summer sea ice of 2.0∙106 km2 (Parkinson et al., 2014; Francis and Wu, 2022; Jeong et al., 2022). The decrease in summer sea ice extent is mainly driven by increasing temperatures in the Arctic (Steiner et al., 2021), and it is foreseen, that the Arctic Ocean will reach an ice-free state in summer within the next two decades (Jahn et al., 2018). From a biological point of view, sea ice is an ecosystem with defined pathways of matter and energy (Lund-Hansen et al., 2020) with microorganisms, zooplankton, fish, seals and polar bears, which rely and depend on sea ice for maintaining their life cycles (Kohlbach et al., 2017). Microorganisms living in sea ice comprise bacteria (Kaartokallio et al., 2013), viruses (Luthanen et al., 2017), fungi (Hassett et al., 2017) heterotrophic and autotrophic protists (Bluhm et al., 2017). Most of the autotrophic organisms as sea ice algae are located at the ice-seawater interface (Horner and Schrader, 1982; Lund-Hansen et al., 2020a) where biomasses can reach concentrations 50-100 times higher compared to concentrations of pelagic phytoplankton (Arrigo, 2017). It is estimated that sea ice algae contribute about 10% of total marine-produced carbon in the Arctic Ocean (Arrigo, 2017). The important ecological functioning of sea ice algae in polar marine ecosystems is to establish an early carbon source for planktonic grazers during the ice-covered winter and early spring (Michel et al., 2002; 2006). Sea ice algae are later released from the melting ice and can seed the water column below the sea ice and ignite the pelagic primary production (Michel et al., 1993; Olsen et al., 2017), or sink to the bottom of the ocean where benthic living organisms can decompose the organic material (Boetius et al., 2013). Nevertheless, an initial literature-based survey of sea ice algae primary production revealed some significant differences in production rates between sites in the Canadian Arctic-Greenland region. This is exemplified by high rates of 463 mg C m-2 d-1 in Resolute Bay (Smith et al., 1988), and 317 mg C m-2 d-1 in Northern Baffin Bay (Nozais et al., 2001) both Canadian Arctic, as compared to the 0.2 mg C m-2 d-1 in Young Sound, northeast Greenland (Rysgaard et al., 1999). Differences in production rates are also mirrored in biomasses produced as for 340 mg Chl a m-2 in Resolute Bay (Smith and Hermann, 1991) and 0.02 mg Chl a m-2 in Kapisillit, west Greenland (Lund-Hansen et al., 2021). Nonetheless, morphology, climatology, light and oceanographic conditions in the region are comparable and similar from a general point of view. This raises the following questions addressed in the present review: 1) Why are there significant spatial differences in the sea ice algae primary production rates and biomasses in the Canadian Artic-Greenland region in spite of comparable physical settings, and 2) What are the driving forces of the differences? 3) Are the differences explained by differences in sea ice conditions, snow cover, nutrients in ice and below ice? The extended spatial variation in primary production is a conundrum considering the comparable physical settings in the region whereby other drivers than light (Leu et al., 2015; Lund-Hansen et al. 2020a) could be parts of the explanation. Addressed questions are responded to by means of a literature study, where data were extracted from a total of 30 studies from study sites in Hudson Bay, the Canadian Arctic Archipelago, and around Greenland (Figure 1). Data on study sites as position, time of sampling, methods applied for measuring primary production, ice types, ice and snow thickness, rates of primary production, Chl a, with references are given in Table S1 (S1 in Supplementary data). Data on nutrients Si(OH)4, PO4, (NO3+NO2), and (NH4) concentrations in ice and in water below, N:P ratios, are given in Table S2 (S2 in Supplementary data). All production rates and concentrations are given for the lower most section (5-10 cm) of the ice. Primary production is abbreviated as PP, Canadian Arctic as CA, and Greenland as GL In the following.
2. Physical and biological parameters of the sea ice
2.1. Ice, snow, production rates and Chl a
All CA samples were collected from land fast first year ice (FYI) except for two samplings in a mixture of pack and land fast ice in northern BB, drift ice in FB, whereas all GL samples were collected in land fast FYI. Sea ice thickness was high (1.44 ± 0.28 m) at CA sites but more than half (0.63 ± 0.41 m) at GL sites except for YS (1.31 ± 0.42 m) (Figure 2a). Snow thickness was comparable at all sites except for the 38 cm (FO) and the 42 cm (YS) at CA and GL sites, respectively (Figure 2b). Highest average PP rates were found at the two CA sites RB (147 ± 19.1 mg C m-2 d-1) and Northern BB (97.1 ± 75.2 mg C m-2 d-1). The rates at other CA sites as HB (3.05 ± 4.1mg C m-2 d-1) and FB (4.5 ± 0.7 mg C m-2 d-1), were quite similar to the average PP at GL sites (4.5 ± 0.7 mg C m-2 d-1) (Figure 2c). The average sea ice Chl a concentration reached a significant maximum at the RB (79.9 mg m-2) CA site with an average at CA sites - except RB site - of 13.5 ± 12.7 mg m-2, but significantly higher than the average (0.8 ± 0.5 mg m-2) at GL sites (Figure 2d)
2.2. Nutrients in sea ice and in water below
Si(OH)4 in the ice reached between 2.0 and 4.0 µM L-1 at CA sites, and similar at GL sites except for the high (8.9 µM L-1) KL concentrations (Figure 2e). The below sea ice water average concentrations of Si(OH)4 were about 3 times higher (9.6 ± 3.1 µM L-1) compared to in ice concentrations (2.8 ± 1.0 µM L-1) at CA sites. Below ice Si(OH)4 average concentrations at GL sites (6.4 ± 1.3 µM L-1) were just slightly higher compared to Si(OH)4 in ice (4.9 ± 2.9 µM L-1) at same sites (Figure 2e-f). PO4 concentrations in water below ice were similar at all locations except for the low (0.01 µM L-1) PO4 concentration in KL (Figure 2g-h). The GL in ice (NO3+NO2) concentrations (2.9 ± 0.5 µM L-1) were comparable to the CA sites (2.5 ± 1.6 µM L-1) which were slightly higher in both GL water below ice (5.0 ± 3.1 µM L-1) and at CA sites (3.9 ± 3.3 µM L-1) (Figure 2i-j). The average N:P ratio was two times higher in CA sea ice (6.4 ± 6.3) compared to GL (3.0 ± 1.3), but comparable in water in CA (5.0 ± 2.5) and GL (6.3 ± 5.0). There was a statistically significant (p < 0.01) correlation between under ice Si(OH)4 concentrations and PP rates.
2.3. Primary production at other Arctic sites
Present average RB, northern BB PP rates were also higher than PP rates measured at other Arctic locations, as for central Arctic Ocean, the Beaufort Sea, Chukchi Sea, Beaufort Shelf, and the Baltic Sea (Table 1). Exception is a very high PP rate of 310 mg C m-2 d-1 measured in the central Arctic Ocean (Gosselin et al., 1997), but exceeded by a rate of 463 mg C m-2 d-1 measured in RB (Smith et al., 1988). Studies outside the CA-GL region were not included in the review as they were obtained at shelf seas and more open water sites.
3. Discussion
There is a general Arctic seasonal signal in sea ice PP rates and Chl a with low winter-early spring rates and Chl a concentrations which both reach a maximum in the transition from winter to spring (Horner et al., 1982; Lund-Hansen et al., 2020). All CA studies were carried out between April and June where ice algae spring bloom occurs during early and late May at 70 °N (Horner et al., 1982; Mundy et al., 2014), whereas spring blooms occur earlier around March-April in southwest GL (Mikkelsen et al., 2008; Lund-Hansen et al., 2014). All samplings were carried out as time-series studies covering several weeks, which ensures that the seasonal variability in PP rates and biomasses (Chl a) were included in the studies. Applying core barrels, either SIPRE Corer or Mark II Kovacs corer for sampling of ice cores in all studies further ensured the comparability. Majority of the 30 independent studies were conducted on land fast first year ice (FYI) as for all GL, and 2 CA FYI/pack ice and 1 land fast/drifting ice, which emphasizes further the comparability between studies and sites. There were no significant differences in biomass or PP rates between land fast and pack ice locations, which is in line with Leu et al. (2015), who found that maximum Chl a did not significantly differ between land fast and pack-ice stations. The 14C method (Nielsen, 1952) was applied in most of the studies with laboratory incubations of melted ice samples (Lund-Hansen et al., 2018; Matthes et al., 2021) in separate bottles inoculated with 14C in a light gradient and 2-4h of incubation in the majority of studies. The 2-4h incubation time provides a PP rate between net and gross PP (Sanz-Martin et al., 2019). A study of in situ and laboratory based 14C PP incubations showed some 10 times lower PP rates measured in situ compared to laboratory-based incubations (Bergmann et al., 1991). Reasons for the significant differences are unclear and accordingly in situ data were excluded and only laboratory-based PP rates included, as also for Frobisher Bay in CA where PP rates were measured in situ (Hsiao, 1988). The in-situ Diving-PAM fluorometer (Rysgaard et al., 2001), the laboratory-based biomass accumulation rates (Riedel et al., 2008), and the O2 concentration changes (Smith et al., 1988; Campbell et al., 2016) have been applied in the studies. These are all considered robust methods for estimating PP (Rysgaard et al., 2001; Glud et al., 2002), and accordingly included in the review. The methodological variations and differences in analytical procedures between studies are accordingly considered of minor importance.
Light intensities at the bottom of the ice, where the majority of algae are located (Lund-Hansen et al., 2020a; Leu et al., 2015), are, apart from any seasonal variation in light, generally regulated by the sea ice and the snow cover with the high albedo (Perovich et al., 1993) and light attenuation coefficients (Lund-Hansen et al., 2018). It was demonstrated that a decrease in snow cover thickness increased PP and photosynthetic performance (Søgaard et al., 2010; Campbell et al., 2016), but other studies were not able to establish any a clear relation as for Nozais et al. (2001) in northern BB. A halted and reduced ice algae spring bloom followed a sudden increase in under ice light induced by manual snow clearing (Lund-Hansen et al., 2014; 2020a). Present review showed no general correlations between thickness of sea ice and snow relative to PP rates and biomasses. Glud et al., (2007) discussed that the low PP rates in YS could be related to a thick (45.0 cm), which might be the case here, though PP rates are still low at other GL sites with much lower (10. 0 cm) snow depths.
There were no specific patterns or relations between study locations and concentrations of nutrients, except for relatively high concentrations in water below ice both of Si(OH)4 and (NO3+NO2), whereas PO4 concentrations were on average higher in the ice compared to the water. Accordingly, linkages between PP rates and nutrient concentrations can be complex. This is here illustrated by the significant high (147 mg C m-2 d-1) average PP rates in RB and northern BB (97.1 mg C m-2 d-1), where low (0.05 µM L-1) and high (10.0 µM L-1) NO3 concentrations were measured in the water, respectively. In RB it was noted that “...NO3 concentrations were highly variable week to week, and obviously reflected the movement of different water masses…” (Welch and Bergmann, 1989). A negative relationship between below ice NO3 concentrations and bottom ice Chl a biomass was found by Rózańska et al. (2009), addressing this to uptake of nutrients by the algae. Stratification caused by melting of sea ice can further reduce the flux of nutrients to the ice-ocean interface and reduce PP, as demonstrated for the central Arctic Ocean (Lund-Hansen et. al, 2020b). Due to stratification, it was suggested that nitrogen limitation was the main factor to explain the low nutrient concentrations in Dease Strait (Campbell et al., 2016). Nitrogen was also pointed out as the limiting nutrient in a study of algal growth in FB (Riedel et al., 2008). For the entire data-set we found a significant (p < 0.01) correlation between average Si(OH)4 concentrations in the water below and PP rates. This points towards Si(OH)4 as an important driver, and it has been shown to limit sea ice algae photosynthesis and blooms especially at a later stage in a diatom dominated bloom in FB (Riedel et al., 2008).
There is a net flow of water from west to east through the CA Archipelago driven by the higher sea levels in the Pacific Ocean, and waters contain higher nutrient concentrations compared to Atlantic waters and especially Si(OH)4 (Michel et al., 2006). Colombo et al. (2020) showed further a clear iron enrichment of the water column with the flow of water from the Pacific through the Barrow Strait passing Resolute Bay towards Baffin Bay. Iron was not measured in any of the present studies but as an important marine phytoplankton micronutrient (Kanna et al., 2014; Yoshida et al., 2021), and iron might have added to the high PP rates in RB and northern BB. Nonetheless, our compiled data showed a tendency that nutrient concentrations as for Si(OH)4 and (NO3+NO2) were higher in the water below the ice at CA sites compared to GL, and especially for (NO3+NO2) and PO4 in BB, although differences were not statistically significant. A recent study in DS showed statistically significant high Chl a concentrations above a sill exposed to increased current speeds and mixing in a tidal strait (Dalman et al., 2019). The term “invisible polynyas’’ was applied to this phenomenon with a reference to large-scale areas of open waters in the ice (Hannah et al., 2009), and often being highly productive (Smith et al., 2007). Accordingly, it is supposed that the high RB PP rates are the combined results of inflow of nutrient rich water of Pacific origin (Michel et al., 2006) and tidal generated mixing of the water masses above a sill in the Barrow Strait adjacent to Resolute Bay (Michel et al., 2006). In comparison, northern BB locations were all located within the North Water Polynya, being one of the most productive of the Canadian polynyas (Stirling, 1997; Klein et al., 2002). This high production relies on the general circulation of nutrient rich water masses of Pacific origin which flows into the northern Baffin Bay through the Nares Strait (Tremblay et al., 2002). The water masses flow at relatively high current speeds of 0.20-0.35 m s-1 (Ingram et al., 2002) and are also rich in Si(OH)4 and PO4 (Tremblay et al., 2002). This strongly indicates that the very high northern BB PP rates are related to the flow of these nutrient rich waters. The relatively high current speeds impede the development of a strong stratification below the sea ice, which inhibits the vertical flux of nutrients (Lund-Hansen et al., 2020b). Sills are present at the entrances to NF and YS in GL, but apparently no tide generated mixing of water and nutrients, where YS is in addition governed by the nutrient depleted East Greenland Current (Rysgaard et al., 1991; 2001). The YS, the Kobberfjord and Kapisillit locations in NF are relatively small fjords with no through flow of nutrient rich waters although nutrient concentrations below water are relatively high in NF (Søgaard et al., 2010).
A conceptual model with drivers of ice algae PP at the ice-water interface in the CA-GL Arctic region where most of the ice algae are located at the sea ice-water interface (Leu et al., 2015) (Figure 3). The amount of light that reaches the bottom of the sea ice for photosynthesis is controlled mainly by the thickness of the snow cover, its water content and age (Hancke et al., 2018), and comparatively less by the physical properties of the ice itself (Perovich, 2017). Nutrients are taken up from the water column, where the supply of these are controlled by advection of water masses below the sea ice. The review pointed out that the high ice algae PP rates in parts of CA were related to inflow of nutrient rich water of Pacific origin, as compared to the very low production rates in East Greenland rates influenced by the nutrient low East Greenland arriving from the Arctic Ocean (Rysgaard et al., 2001). A second parameter is the vertical flux of nutrients to the bottom of the ice when a stratification of the water column is established by melting of the ice, or inflow of water with a lower density, which strongly inhibits the vertical mixing and thereby the flux of the nutrients towards the bottom of the sea ice (Randelhoff et al., 2016). Specifically, the weaker stratification enhanced the vertical transport of nutrients towards ice algae as observed in RB, where the mixing was promoted by sills (Michel et al., 2006). The grazing by copepods and amphipods is a third parameter that can severely affect and reduce ice algae biomasses (Sampei et al., 2021) though not the photosynthesis.
4. Conclusions
There were some significant differences in spatial distribution of sea ice algae PP rates and biomasses as in RB and northern BB in CA with very high PP rates and biomasses. PP rates outside RB and BB were similar in CA and GL, but average Chl a outside was higher in CA compared to GL. The RB and BB are the exceptions, and the review pointed out that the high ice algae PP rates and biomasses were related to large-scale driven inflow and mixing of nutrient rich waters of Pacific origin. Waters were mixed by stronger currents above sills and in straits that generated a high flux of nutrients towards the bottom of the sea ice, which sustained the high PP rates. There has been an extended focus on light as the main driver in ice algae PP, but we conclude that advection and mixing of nutrient rich water masses are also important drivers.
Author Contributions
LC.L.-H., and L.M.C.: conceptualization; writing - original draft preparation; B.S. and D.H.S.: writing – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
The paper was conceived and developed in the frame of the project FACE-IT (The Future of Arctic Coastal Ecosystems – Identifying Transitions in Fjords and Adjacent Coastal Areas). FACE-IT has received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement No. 869154. This is a contribution to the Arctic Science Partnership (ASP) asp-net.org.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data availability statement
No new data were created in this study, and data sharing is not applicable to this article.
Conflicts Interests
All authors declare to have no conflicts of interest.
References
- Andersen, O.G.N. Primary production associated with sea ice at Godhavn, Disko, West Greenland. Ophelia 1977, 16, 205–220. [Google Scholar] [CrossRef]
- Arrigo, K., 2017. Sea ice as a habitat for primary producers. In “Sea Ice”, 3rd ed.; Thomas, D.N., Ed., Wiley Blackwell, Oxford, UK, 2017; 652p.
- Bergmann, M.; Welch, H.; Butler-Walker, J.; Siferd, T. Ice algal photosynthesis at resolute and saqvaqjuac in the Canadian arctic. J. Mar. Syst. 1991, 2, 43–52. [Google Scholar] [CrossRef]
- Boetius, A.; Albrecht, S.; Bakker, K.; Bienhold, C.; Felden, J.; Fernández-Méndez, M.; Hendricks, S.; Katlein, C.; Lalande, C.; Krumpen, T.; et al. Export of Algal Biomass from the Melting Arctic Sea Ice. Science 2013, 339, 1430–1432. [Google Scholar] [CrossRef] [PubMed]
- Bluhm, B.A., Swadling, K.M., Gradinger, R., 2017. In “Sea Ice”, 3rd ed.; Thomas, D.N., Ed., Wiley Blackwell, Oxford, UK, 2017; 652p.
- Booth, J. The Epontic Algal Community of the Ice Edge Zone and Its Significance to the Davis Strait Ecosystem. ARCTIC 1984, 37, 234–243. [Google Scholar] [CrossRef]
- Campbell, K. , Mundy, C., Landy, J., Delaforge, A., Michel, C., Rysgaard, S. Community dynamics of bottom-ice algae in Dease Strait of the Canadian Arctic. Prog. Oceanogr. 2016, 149, 27–39. Biochemistry 1991, 30, 7586–7597. [Google Scholar] [CrossRef]
- Colombo, M.; Jackson, S.L.; Cullen, J.T.; Orians, K.J. Dissolved iron and manganese in the Canadian Arctic Ocean: On the biogeochemical processes controlling their distributions. Geochim. et Cosmochim. Acta 2020, 277, 150–174. [Google Scholar] [CrossRef]
- Dalman, L.A.; Else, B.G.T.; Barber, D.; Carmack, E.; Williams, W.J.; Campbell, K.; Duke, P.J.; Kirillov, S.; Mundy, C.J. Enhanced bottom-ice algal biomass across a tidal strait in the Kitikmeot Sea of the Canadian Arctic. Elementa: Sci. Anthr. 2019, 7. [Google Scholar] [CrossRef]
- Dunbar, M.; Acreman, J. Standing crops and species composition of diatoms in sea ice From Robeson Channel to the Gulf of St. Lawrence. Ophelia 1980, 19, 61–72. [Google Scholar] [CrossRef]
- Dupont, F. Impact of sea-ice biology on overall primary production in a biophysical model of the pan-Arctic Ocean. J. Geophys. Res. Oceans 2012, 117. [Google Scholar] [CrossRef]
- Fernández-Méndez, M.; Katlein, C.; Rabe, B.; Nicolaus, M.; Peeken, I.; Bakker, K.; Flores, H.; Boetius, A. Photosynthetic production in the central Arctic Ocean during the record sea-ice minimum in 2012. Biogeosciences 2015, 12, 3525–3549. [Google Scholar] [CrossRef]
- Francis, J.A. , Wu, B. Why has no new record-minimum Arctic sea-ice extent occurred since September 2012? Environ. Res. Lett. 2020, 15, 114034. [Google Scholar] [CrossRef]
- Glud, R.N.; Rysgaard, S.; Kühl, M. A laboratory study on O2 dynamics and photosynthesis in ice algal communities: quantification by microsensors, O2 exchange rates, 14C incubations and a PAM fluorometer. Aquat. Microb. Ecol. 2002, 27, 301–311. [Google Scholar] [CrossRef]
- Glud, R.N., Rysgaard, S., Kühl, M., Hansen, J.W., 2007. The sea ice in Young Sound: Implications for carbon cycling. Carbon cycling in Arctic marine ecosystems: case study Young Sound. Meddr. Grønland 58, 62-85.
- Gosselin, M.; Legendre, L.; Demers, S.; Ingram, R.G. Responses of Sea-Ice Microalgae to Climatic and Fortnightly Tidal Energy Inputs (Manitounuk Sound, Hudson Bay). Can. J. Fish. Aquat. Sci. 1985, 42, 999–1006. [Google Scholar] [CrossRef]
- Gosselin, M.; Legendre, L.; Therriault, J.-C.; Demers, S. Light and nutrient limitation of sea-ice microalgae (Hudson Bay, Canadian Arctic. J. Phycol. 1990, 26, 220–232. [Google Scholar] [CrossRef]
- Gosselin, M.; Levasseur, M.; Wheeler, P.A.; Horner, R.A.; Booth, B.C. New measurements of phytoplankton and ice algal production in the Arctic Ocean. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 1997, 44, 1623–1644. [Google Scholar] [CrossRef]
- Gradinger, R. Sea-ice algae: Major contributors to primary production and algal biomass in the Chukchi and Beaufort Seas during May/June 2002. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2009, 56, 1201–1212. [Google Scholar] [CrossRef]
- Haecky, P.; Andersson, A. Primary and bacterial production in sea ice in the northern Baltic Sea. Aquat. Microb. Ecol. 1999, 20, 107–118. [Google Scholar] [CrossRef]
- Hannah, C.G.; Dupont, F.; Dunphy, M. Polynyas and Tidal Currents in the Canadian Arctic Archipelago. ARCTIC 2009, 62, 83–95. [Google Scholar] [CrossRef]
- Hassett, B.T.; Ducluzeau, A.L.L.; Collins, R.E.; Gradinger, R. Spatial distribution of aquatic marine fungi across the western Arctic and sub-arctic. Environ. Microbiol. 2017, 19, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.; Schrader, G. Relative Contributions of Ice Algae, Phytoplankton, and Benthic Microalgae to Primary Production in Nearshore Regions of the Beaufort Sea. ARCTIC 1982, 35, 485–503. [Google Scholar] [CrossRef]
- Hsiao, S. Spatial and seasonal variations in primary production of sea ice microalgae and phytoplankton in Frobisher Bay, Arctic Canada. Mar. Ecol. Prog. Ser. 1988, 44, 275–285. [Google Scholar] [CrossRef]
- Ingram, R.G., Bâcle, J., Barber, D.G., Gratton, Y., Melling, H., 2002. An overview of physical processes in the North Water. Deep Sea Research Part II: Topical Studies in Oceanography 49, 4893-4906.
- Jahn, A.; Kay, J.E.; Holland, M.M.; Hall, D.M. How predictable is the timing of a summer ice-free Arctic? Geophys. Res. Lett. 2016, 43, 9113–9120. [Google Scholar] [CrossRef]
- Jeong, H.; Park, H.; Stuecker, M.F.; Yeh, S. Record Low Arctic Sea Ice Extent in 2012 Linked to Two-Year La Niña-Driven Sea Surface Temperature Pattern. Geophys. Res. Lett. 2022, 49. [Google Scholar] [CrossRef]
- Kaartokallio, H.; Søgaard, D.; Norman, L.; Rysgaard, S.; J, L.T.; Delille, B.; Thomas, D. Short-term variability in bacterial abundance, cell properties, and incorporation of leucine and thymidine in subarctic sea ice. Aquat. Microb. Ecol. 2013, 71, 57–73. [Google Scholar] [CrossRef]
- Kanna, N.; Toyota, T.; Nishioka, J. Iron and macro-nutrient concentrations in sea ice and their impact on the nutritional status of surface waters in the southern Okhotsk Sea. Prog. Oceanogr. 2014, 126, 44–57. [Google Scholar] [CrossRef]
- Karami, M.P.; Myers, P.G.; de Vernal, A.; Tremblay, L.B.; Hu, X. The role of Arctic gateways on sea ice and circulation in the Arctic and North Atlantic Oceans: a sensitivity study with an ocean-sea-ice model. Clim. Dyn. 2021, 57, 2129–2151. [Google Scholar] [CrossRef]
- Klein, B.; LeBlanc, B.; Mei, Z.-P.; Beret, R.; Michaud, J.; Mundy, C.-J.; von Quillfeldt, C.H.; Garneau, M.; Roy, S.; Gratton, Y.; et al. Phytoplankton biomass, production and potential export in the North Water. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2002, 49, 4983–5002. [Google Scholar] [CrossRef]
- Kohlbach, D.; Smik, L.; Belt, S.T.; Hop, H.; Wold, A.; Graeve, M.; Assmy, P. A multi-trophic marker approach reveals high feeding plasticity in Barents Sea under-ice fauna. Prog. Oceanogr. 2022, 208. [Google Scholar] [CrossRef]
- Lehmann, N.; Kienast, M.; Granger, J.; Bourbonnais, A.; Altabet, M.A.; Tremblay, J. Remote Western Arctic Nutrients Fuel Remineralization in Deep Baffin Bay. Glob. Biogeochem. Cycles 2019, 33, 649–667. [Google Scholar] [CrossRef]
- Lewis, E.L.; Ponton, D.; Legendre, L.; Leblanc, B. Springtime sensible heat, nutrients and phytoplankton in the Northwater Polynya, Canadian Arctic. Cont. Shelf Res. 1996, 16, 1775–1792. [Google Scholar] [CrossRef]
- Leu, E.; Mundy, C.; Assmy, P.; Campbell, K.; Gabrielsen, T.; Gosselin, M.; Juul-Pedersen, T.; Gradinger, R. Arctic spring awakening – Steering principles behind the phenology of vernal ice algal blooms. Prog. Oceanogr. 2015, 139, 151–170. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Hawes, I.; Sorrell, B.K.; Nielsen, M.H. Removal of snow cover inhibits spring growth of Arctic ice algae through physiological and behavioral effects. Polar Biol. 2013, 37, 471–481. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Hawes, I.; Nielsen, M.H.; Dahllöf, I.; Sorrell, B.K. Summer meltwater and spring sea ice primary production, light climate and nutrients in an Arctic estuary, Kangerlussuaq, west Greenland. Arctic, Antarct. Alp. Res. 2018, 50. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Søgaard, D.H.; Sorrell, B.K.; Gradinger, R.; Meiners, K.M. Arctic Sea Ice Ecology–Seasonal Dynamics in Algal and Bacterial Productivity; Springer: Berlin, Germany, 2020; 178p. [Google Scholar]
- Lund-Hansen, L.; Hawes, I.; Hancke, K.; Salmansen, N.; Nielsen, J.; Balslev, L.; Sorrell, B. Effects of increased irradiance on biomass, photobiology, nutritional quality, and pigment composition of Arctic sea ice algae. Mar. Ecol. Prog. Ser. 2020, 648, 95–110. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Bendtsen, J.; Stratmann, T.; Tonboe, R.; Olsen, S.M.; Markager, S.; Sorrell, B.K. Will low primary production rates in the Amundsen Basin (Arctic Ocean) remain low in a future ice-free setting, and what governs this production? J. Mar. Syst. 2020, 205, 103287. [Google Scholar] [CrossRef]
- Lund-Hansen, L.C.; Petersen, C.M.; Søgaard, D.H.; Sorrell, B.K. A Comparison of Decimeter Scale Variations of Physical and Photobiological Parameters in a Late Winter First-Year Sea Ice in Southwest Greenland. J. Mar. Sci. Eng. 2021, 9, 60. [Google Scholar] [CrossRef]
- Luhtanen, A.-M.; Eronen-Rasimus, E.; Oksanen, H.M.; Tison, J.-L.; Delille, B.; Dieckmann, G.S.; Rintala, J.-M.; Bamford, D.H. The first known virus isolates from Antarctic sea ice have complex infection patterns. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Matthes, L.C.; Ehn, J.K.; Dalman, L.A.; Babb, D.G.; Peeken, I.; Harasyn, M.; Kirillov, S.; Lee, J.; Bélanger, S.; Tremblay, J.; et al. Environmental drivers of spring primary production in Hudson Bay. Elementa: Sci. Anthr. 2021, 9. [Google Scholar] [CrossRef]
- Michel, C.; Legendre, L.; Therriault, J.-C.; Demers, S.; Vandevelde, T. Springtime coupling between ice algal and phytoplankton assemblages in southeastern Hudson Bay, Canadian Arctic. Polar Biol. 1993, 13. [Google Scholar] [CrossRef]
- Michel, C.; Nielsen, T.G.; Nozais, C.; Gosselin, M. Significance of sedimentation and grazing by ice micro- and meiofauna for carbon cycling in annual sea ice (northern Baffin Bay). Aquat. Microb. Ecol. 2002, 30, 57–68. [Google Scholar] [CrossRef]
- Michel, C.; Ingram, R.; Harris, L. Variability in oceanographic and ecological processes in the Canadian Arctic Archipelago. Prog. Oceanogr. 2006, 71, 379–401. [Google Scholar] [CrossRef]
- Mikkelsen, D.M.; Rysgaard, S.; Glud, R.N. Microalgal composition and primary production in Arctic sea ice: a seasonal study from Kobbefjord (Kangerluarsunnguaq), West Greenland. Mar. Ecol. Prog. Ser. 2008, 368, 65–74. [Google Scholar] [CrossRef]
- Mock, T.; Gradinger, R. Determination of Arctic ice algal production with a new in situ incubation technique. Mar. Ecol. Prog. Ser. 1999, 177, 15–26. [Google Scholar] [CrossRef]
- Mundy, C.J.; Gosselin, M.; Gratton, Y.; Brown, K.; Galindo, V.; Campbell, K.; Levasseur, M.; Barber, D.; Papakyriakou, T.; Bélanger, S. Role of environmental factors on phytoplankton bloom initiation under landfast sea ice in Resolute Passage, Canada. Mar. Ecol. Prog. Ser. 2014, 497, 39–49. [Google Scholar] [CrossRef]
- Nielsen, E.S. The Use of Radio-active Carbon (C14) for Measuring Organic Production in the Sea. ICES J. Mar. Sci. 1952, 18, 117–140. [Google Scholar] [CrossRef]
- Rózanska, M.; Gosselin, M.; Poulin, M.; Wiktor, J.M.; Michel, C. Influence of environmental factors on the development of bottom ice protist communities during the winter–spring transition. Mar. Ecol. Prog. Ser. 2009, 386, 43–59. [Google Scholar] [CrossRef]
- Olsen, L.M.; Laney, S.R.; Duarte, P.; Kauko, H.M.; Fernández-Méndez, M.; Mundy, C.J.; Rösel, A.; Meyer, A.; Itkin, P.; Cohen, L.; et al. The seeding of ice algal blooms in Arctic pack ice: The multiyear ice seed repository hypothesis. J. Geophys. Res. Biogeosciences 2017, 122, 1529–1548. [Google Scholar] [CrossRef]
- Parkinson, C.L. Spatially mapped reductions in the length of the Arctic sea ice season. Geophys. Res. Lett. 2014, 41, 4316–4322. [Google Scholar] [CrossRef] [PubMed]
- Pineault, S.; Tremblay, J.; Gosselin, M.; Thomas, H.; Shadwick, E. The isotopic signature of particulate organic C and N in bottom ice: Key influencing factors and applications for tracing the fate of ice-algae in the Arctic Ocean. J. Geophys. Res. Oceans 2013, 118, 287–300. [Google Scholar] [CrossRef]
- Perovich, D.K.; Roesler, C.S.; Pegau, W.S. Variability in Arctic sea ice optical properties. J. Geophys. Res. Oceans 1998, 103, 1193–1208. [Google Scholar] [CrossRef]
- Perovich, D. K. (2017). Sea ice and sunlight. In D. N. Thomas (Ed.), Sea ice (3rd ed., pp. 110–137, 652 pp). Oxford: Wiley Blackwell.
- Riedel, A.; Michel, C.; Gosselin, M.; LeBlanc, B. Winter–spring dynamics in sea-ice carbon cycling in the coastal Arctic Ocean. J. Mar. Syst. 2008, 74, 918–932. [Google Scholar] [CrossRef]
- Randelhoff, A.; Fer, I.; Sundfjord, A.; Tremblay, J.; Reigstad, M. Vertical fluxes of nitrate in the seasonal nitracline of the Atlantic sector of the Arctic Ocean. J. Geophys. Res. Oceans 2016, 121, 5282–5295. [Google Scholar] [CrossRef]
- Rózanska, M.; Gosselin, M.; Poulin, M.; Wiktor, J.M.; Michel, C. Influence of environmental factors on the development of bottom ice protist communities during the winter–spring transition. Mar. Ecol. Prog. Ser. 2009, 386, 43–59. [Google Scholar] [CrossRef]
- Rysgaard, S.; Nielsen, T.; Hansen, B. Seasonal variation in nutrients, pelagic primary production and grazing in a high-Arctic coastal marine ecosystem, Young Sound, Northeast Greenland. Mar. Ecol. Prog. Ser. 1999, 179, 13–25. [Google Scholar] [CrossRef]
- Rysgaard, S.; Kühl, M.; Glud, R.N.; Hansen, J.W. Biomass, production and horizontal patchiness of sea ice algae in a high-Arctic fjord (Young Sound, NE Greenland). Mar. Ecol. Prog. Ser. 2001, 223, 15–26. [Google Scholar] [CrossRef]
- Sampei, M., Fortier, L., Raimbault, P., Matsuno, K., Vet, Y., Quéguiner, LaFond, A., Babin, M., Hirawake, T., (2021). An estimation of the quantitative impacts of copepod grazing on an under sea-ice spring phytoplabnkton bloom in western Baffin Bay, Canadian Arctic. Elementa: Sceince of the Anthropocene, 9. [CrossRef]
- Sanz-Martín, M.; Vernet, M.; Cape, M.R.; Mesa, E.; Delgado-Huertas, A.; Reigstad, M.; Wassmann, P.; Duarte, C.M. Relationship Between Carbon- and Oxygen-Based Primary Productivity in the Arctic Ocean, Svalbard Archipelago. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Smith, R.E.H., Anning, J., Pierre Clement, Cota, G., 1988. Abundance and production of ice algae in Resolute Passage, Canadian Arctic. Mar. Ecol. Prog. Ser 48, 251-263.
- Smith, R.E.; Herman, A.W. Productivity of sea ice algae: In situ vs. incubator methods. J. Mar. Syst. 1991, 2, 97–110. [Google Scholar] [CrossRef]
- Smith Jr., W.O., Barber, D., (Eds.). 2007. Polynyas: Windows to the world. Elsevier Oceanography Series, Vol. 74. pp 474.
- Steiner, N.S.; Bowman, J.; Campbell, K.; Chierici, M.; Eronen-Rasimus, E.; Falardeau, M.; Flores, H.; Fransson, A.; Herr, H.; Insley, S.J.; et al. Climate change impacts on sea-ice ecosystems and associated ecosystem services. Elementa: Sci. Anthr. 2021, 9. [Google Scholar] [CrossRef]
- Stirling, I. The importance of polynyas, ice edges, and leads to marine mammals and birds. J. Mar. Syst. 1997, 10, 9–21. [Google Scholar] [CrossRef]
- Søgaard, D.H.; Kristensen, M.; Rysgaard, S.; Glud, R.N.; Hansen, P.J.; Hilligsøe, K.M. Autotrophic and heterotrophic activity in Arctic first-year sea ice: seasonal study from Malene Bight, SW Greenland. Mar. Ecol. Prog. Ser. 2010, 419, 31–45. [Google Scholar] [CrossRef]
- Søgaard, D.H.; Thomas, D.N.; Rysgaard, S.; Glud, R.N.; Norman, L.; Kaartokallio, H.; Juul-Pedersen, T.; Geilfus, N.-X. The relative contributions of biological and abiotic processes to carbon dynamics in subarctic sea ice. Polar Biol. 2013, 36, 1761–1777. [Google Scholar] [CrossRef]
- Søgaard, D.H.; Sorrell, B.K.; Sejr, M.K.; Andersen, P.; Rysgaard, S.; Hansen, P.J.; Skyttä, A.; Lemcke, S.; Lund-Hansen, L.C. An under-ice bloom of mixotrophic haptophytes in low nutrient and freshwater-influenced Arctic waters. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Tremblay, J.; Gratton, Y.; Carmack, E.C.; Payne, C.D.; Price, N.M. Impact of the large-scale Arctic circulation and the North Water Polynya on nutrient inventories in Baffin Bay. J. Geophys. Res. Atmos. 2002, 107, 26–1. [Google Scholar] [CrossRef]
- Tremblay, J.É., Bélanger, S., Barber, D., Asplin, M., Martin, J., Darnis, G., Fortier, L., Gratton, Y., Link, H., Archambault, P., 2011. Climate forcing multiplies biological productivity in the coastal Arctic Ocean. Geophysical Research Letters 38.
- Welch, H.E.; Bergmann, M.A. Seasonal Development of Ice Algae and its Prediction from Environmental Factors near Resolute, N.W.T., Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1793–1804. [Google Scholar] [CrossRef]
- Wratten, E.E.; Cooley, S.W.; Mann, P.J.; Whalen, D.; Fraser, P.; Lim, M. Physiographic Controls on Landfast Ice Variability from 20 Years of Maximum Extents across the Northwest Canadian Arctic. Remote. Sens. 2022, 14, 2175. [Google Scholar] [CrossRef]
- Yoshida, K.; Seger, A.; Corkill, M.; Heil, P.; Karsh, K.; McMinn, A.; Suzuki, K. Low Fe Availability for Photosynthesis of Sea-Ice Algae: Ex situ Incubation of the Ice Diatom Fragilariopsis cylindrus in Low-Fe Sea Ice Using an Ice Tank. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
Figure 1.
Study sites (red circles) and names of locations, which are abbreviated in the text as: Franklin Bay (FB), Dease Strait (DS), Resolute Bay (RB), Northern Baffin Bay (BB), Robeson Channel (RC), Hudson Bay (HB), Frobisher Bay (FO), Disko Island (Di), Kangerlussuaq (KL), and Young Sound (YS), Nuuk Fjord (NF) comprises Malene Bight, Kapisillit, and Kobberfjord.
Figure 1.
Study sites (red circles) and names of locations, which are abbreviated in the text as: Franklin Bay (FB), Dease Strait (DS), Resolute Bay (RB), Northern Baffin Bay (BB), Robeson Channel (RC), Hudson Bay (HB), Frobisher Bay (FO), Disko Island (Di), Kangerlussuaq (KL), and Young Sound (YS), Nuuk Fjord (NF) comprises Malene Bight, Kapisillit, and Kobberfjord.
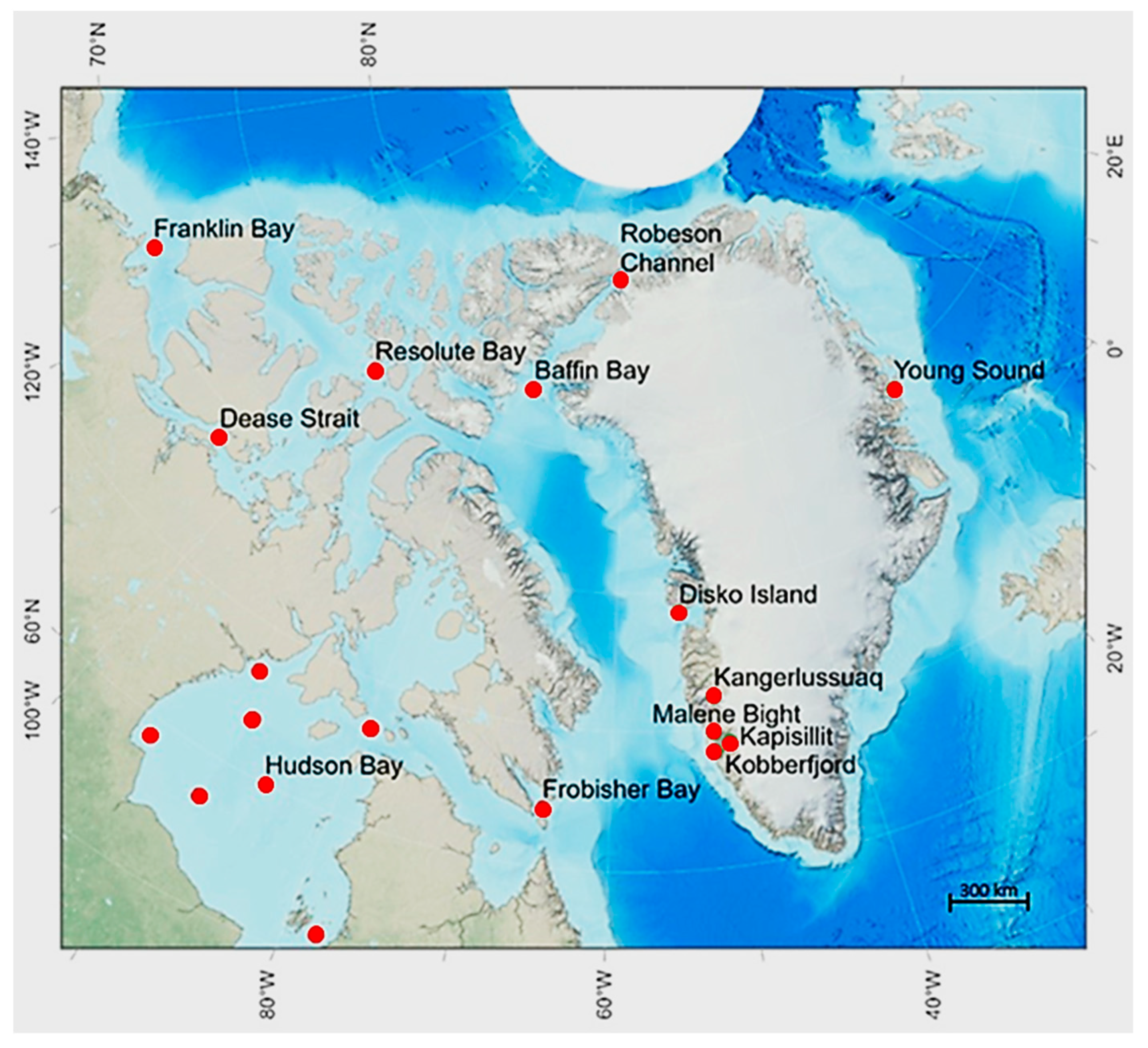
Figure 2.
Average ± s.d. sea ice thickness (a), snow thickness (b), primary production (c), Chl a (d), Si(OH)4 in ice (e), Si(OH)4 in water (f), PO4 in ice (g), PO4 in water (h), (NO3+NO2) in ice (i), and (NO3+NO2) in water (j) at locations with abbreviations as in Figure 1. In water means, that water was sampled 1-2 m below the sea ice. Note that numbers for Nuuk Fjord (NF) are averages comprising the three locations – Kapisillit, Malene Bight, and Kobberfjord. Numbers from each of these three locations are given in Tables S1 and S2. The * signifies that the parameter or component was not measured at that specific location.
Figure 2.
Average ± s.d. sea ice thickness (a), snow thickness (b), primary production (c), Chl a (d), Si(OH)4 in ice (e), Si(OH)4 in water (f), PO4 in ice (g), PO4 in water (h), (NO3+NO2) in ice (i), and (NO3+NO2) in water (j) at locations with abbreviations as in Figure 1. In water means, that water was sampled 1-2 m below the sea ice. Note that numbers for Nuuk Fjord (NF) are averages comprising the three locations – Kapisillit, Malene Bight, and Kobberfjord. Numbers from each of these three locations are given in Tables S1 and S2. The * signifies that the parameter or component was not measured at that specific location.
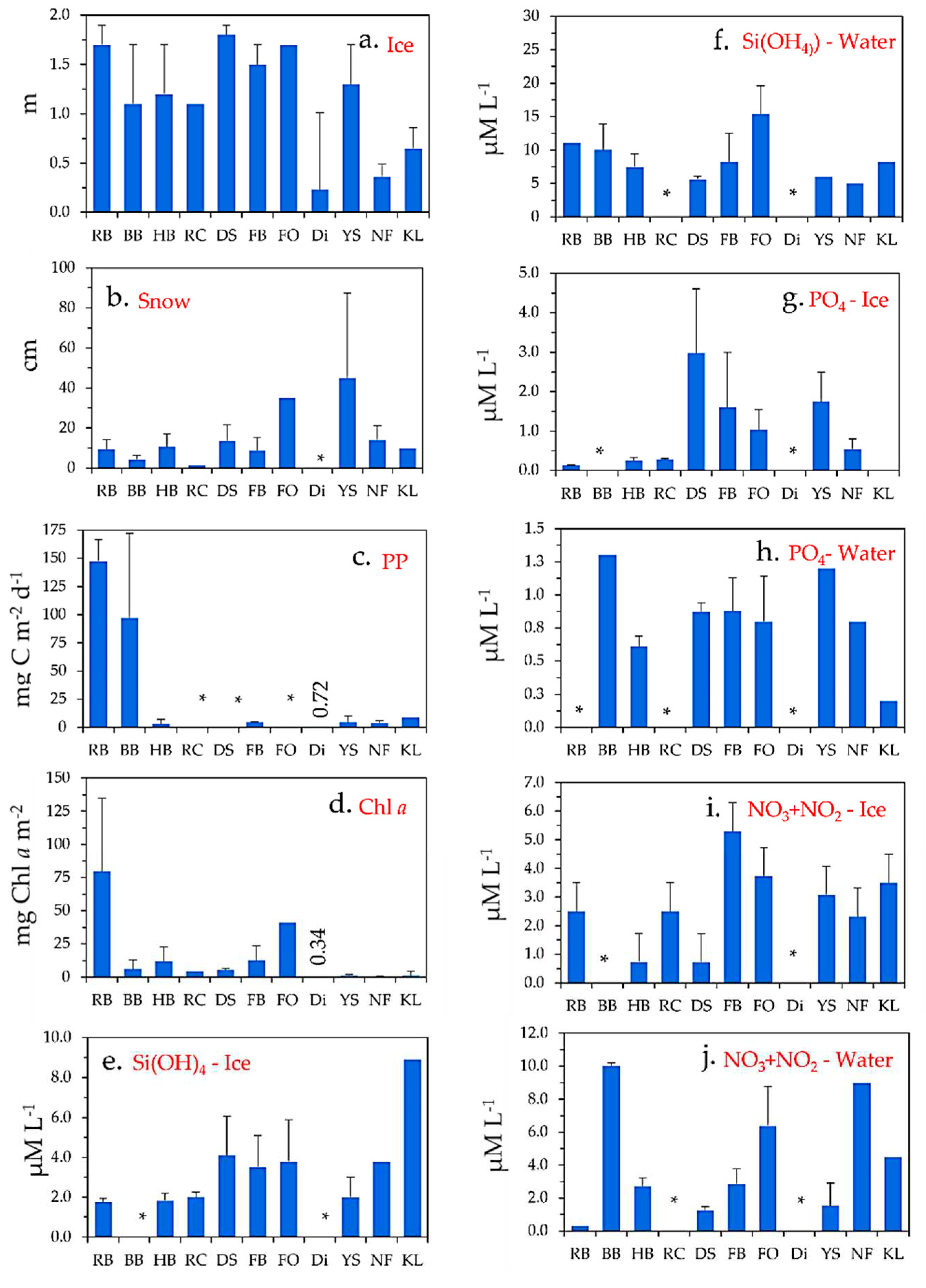
Figure 3.
Conceptual model with drivers of ice algae primary production at the sea ice-water interface.
Figure 3.
Conceptual model with drivers of ice algae primary production at the sea ice-water interface.

Table 1.
Ranges of sea ice algae primary production from RB (Resolute Bay) and northern BB (Baffin Bay), and other Arctic locations.
Table 1.
Ranges of sea ice algae primary production from RB (Resolute Bay) and northern BB (Baffin Bay), and other Arctic locations.
| Location. | Production Rate | Reference |
|---|---|---|
| Resolute Bay | 20.6 – 469 mg C m-2 d-1 | (Smith & Hermann, 1991) |
| Northern Baffin Bay | 26.3 - 317 mg C m-2 d-1 | (Nozais et al. 2001) |
| Arctic Ocean | 0.5-310 mg C m-2 d-1 | (Gosselin et al., 1997) |
| Arctic Ocean | 5.8 mg C m-2 d-1 | (Fernández-Méndez et al., 2015) |
| Beaufort Sea | 2.8-11.2 mg C m-2 d-1 | (Horner, 1982) |
| Chukchi Sea | 20-30 mg C m-2 d-1 | (Gradinger et al., 2009) |
| Beaufort Sea | 4-9 mg C m-2 d-1 | (Gradinger et al., 2009) |
| Pan Arctic PP model | 60-100 mg C m-2 d-1 | (Dupont, 2012) |
| Greenland Sea | 0.25-1.71 mg C m-2 d-1 | (Mock & Gradinger, 1999) |
| Davis Strait | 0.003-2.36 mg C m-2 d-1 | (Booth, 1984) |
| Baltic Sea | 1.0-2.4 mg C m-2 d-1 | (Haecky & Andersson, 1999) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



