
Submitted:
16 August 2023
Posted:
17 August 2023
You are already at the latest version
Abstract
Basil (Ocimum basilicum L.) is one of the important medicinal and aromatic plants. Light intensity is an indispensable factor for plants due to its effect on photosynthesis and physiological processes. Here, we investigated the impact of light intensities and harvesting times on the photosynthesis of green and purple basil. The experiment involved subjecting the plants to three different sunlight levels for 12 days: complete – 100%, 50%, and 30% sunlight. In addition, we scrutinize the harvesting time. The highest levels of photosynthetic and protective pigments were detected under full sunlight conditions in the purple basil harvested at noon. The highest levels of soluble and storage carbohydrates were recorded in the purple basil grown under full sunlight and harvested in the early morning. In contrast, their lowest levels were obtained in plants grown under 30% sunlight and harvested at noon time. Under all light treatments, the maximum quantum yield of photosystem II (FV/FM) was detected at 4 a.m. in both basil varieties; it decreased at noon and increased again at 5 p.m. Non-Photochemical Quenching (NPQ) got most elevated in the green variety under all light intensities at noon. However the highest NPQ was detected in the purple variety at 8 a.m. The NPQ was lowest in both basil varieties during the early morning and afternoon. Full sunlight at noon caused temporary photoinhibition and reduced car-bohydrates while enhancing pigment concentration and photo-protective mechanisms in basil plants.
Keywords:
Chlorophyll fluorescence
; harvest time
; shading
; Controlled Environment Agriculture (CEA)
; photoinhibition
1. Introduction
Due to the adverse effects of chemical medicine and the growing trend in herbal remedies, significant agricultural lands have been devoted to the cultivation of medicinal plants in recent decades. About 150 species are known for the genus Ocimum, of which the basil (Ocimum basilicum L.) is the most important and is cultivated in many parts of the world because of its commercial value [1]. Basil has many applications in the food, pharmaceutical, and cosmetics industries. Basil oil is used as a spice in pastries, salads, soft drinks, ice cream, perfume factories and oral and dental products such as toothpaste [2]. Furthermore, essential oils repel insects and have anti-parasitic, anti-bacterial, antifungal, and antioxidant properties [3].
Light is recognized as an essential factor for plants due to its effect on photosynthesis, physiological processes, plant structure and morphology, and the production of phytochemicals [4]. During the growing season, the quantity of solar energy plants receive determines their development and function. High and low light intensities can limit photosynthesis and carbon fixation in plants. In full sunlight, light inhibition occurs, leading to reduced photosynthesis, especially when other stresses are present [5]. Low light intensities also stress the plants, as they caused reduced carbon concentration and photosynthetic net stabilization and hamper plant growth [6]. In addition to its effects on photosynthesis, light intensity also affects plant temperature and morphological responses, which in unfavorable levels, also cause stress [7]. Due to the critical role of light in photosynthesis (source of energy absorbed by pigments, induction of pigment synthesis, stomatal opening), when the light is lower than a species-dependent levels, plants often strive to expose themselves to higher light levels during their growth [6,8,9].
High light intensities impose problems for the crops especially during the summer time [10]. Therefore, decreasing the light intensity during the seasons with high light intensity is a standard practice in the greenhouses. One way to adjust the light intensity is to use shading techniques. Shading grids are used to protect horticultural crops from excessive sunlight, environmental hazards (wind and hail), or flying pests (birds and insects) [11]. Shading reduces the quantity of light reaching the plant surface, which reduces plant growth and development, especially under conditions of other stresses [12]. The cons and pros of high and low light intensities have been widely studied before. For instance, in Salvia, photosynthetic pigments increased at a low light intensity, and overall biomass weight and plant height declined with decreasing light intensity [13]. In sunflowers, plant growth and soluble protein content were higher at higher light intensity (350 µmol m-2 s-1), leading to higher growth due to increase in CO2 fixation. Chlorophyll content increased with decreasing light intensity (125 µmol m-2 s-1). Furthermore, carbohydrate levels were higher in plants grown at higher light intensities. Nitrate reductase activity in both light treatments decreased with increasing age of sunflower plants [14]. In Brassica campestris ssp., malondialdehyde content, which indicates damage to plant cells, increased under low light. Exposure to the shade caused a decrease in chlorophyll a, soluble proteins, net photosynthesis, transpiration, stomatal conductance and antioxidant activity. Moreover, low light intensity reduced anthocyanin biosynthetic enzymes and caused B. campestris plants to change their color from purple to green [15].
Plants are exposed to different light levels during the course of the day. This would result in different plant metabolite profiles during different times of a day. Therefore, harvesting plant materials at different time during the day may result in achieving different metabolites and biochemicals in the harvested plant products [16]. Although the application of shading at different levels is a common practice for greenhouse crop production in different parts of the world (especially in the mid-range latitudes) on the seasons characterized by high light intensities, determination of photosynthetic functionality under different light levels during the course of a day has not been addressed so far. The present study aimed to investigate the photosynthetic functionality and the level of photoprotective machinery under different shade levels at different time-point of the day on basil plants with other pigmentations. We hypothesized that, there would be higher pressure on the photosynthesis system at noon due to higher light intensities. Furthermore, due to different pigmentations in green and purple, we hypothesized that they respond differently to the light levels. Basil was used in this investigation since it is one of the most essential herbs/medicinal plants, widely used in mid-range latitude countries and the availability of varieties with different pigmentations (green and purple).
2. Materials and Methods
2.1. Plant material and growth conditions
Seeds from two commercial basil plants, green 'Mobarake' and purple 'Ardestan', were sown in a mixture of coir-pith and perlite in a 1:1 ratio (v:v) in a greenhouse. After one month, in the six-leaf stage, basil seedlings were transferred to pots containing 1: 1: 1 soil, sand, and leaf soil, and regularly irrigated. To investigate the effect of different light intensities at different times of day, a factorial split-plot experiment was conducted in a completely randomized design with two factors including basil variety (green and purple), three light intensities (full light, 50% and 30% of full sunlight) and four harvesting times (4 a.m., 8 a.m., 12 and 5 p.m.). For decreasing light intensity, shade nets with the ability to reduce light intensity by 50% and 70% were installed in the greenhouse at a distance of 1.5 m above the plants. Plants were positioned under these shades, along with the regular greenhouse lighting. Twelve days after the light treatments, plants were sampled.

Table 1.
Light intensity (µmol m-2 s-1) under different shade levels and at different time course of a day.
Table 1.
Light intensity (µmol m-2 s-1) under different shade levels and at different time course of a day.
| Light Intensity (µmol m−2 s−1) | |||
|---|---|---|---|
| Time-point | 100% of sunlight | 50% of sunlight | 30% of sunlight |
| 8 a.m. | 470 | 235 | 141 |
| 12 | 1230 | 615 | 369 |
| 5 p.m. | 650 | 325 | 195 |
2.2. Determination of chlorophyll and carotenoid contents
Chlorophyll (Chl) and carotenoid contents in plant leaves were determined based on the method of Lichtenthaler and Wellburn (1983). The samples (500 mg) were extracted in 10 ml acetone (80%), and the extract was separated by centrifugation (SIGMA-3K30) at 7000 g for 5 min. Then the absorbance of the supernatant was spectrophotometrically (Lambda 25-UV/VIS spectrometer) measured at 645, 663, and 470 nm and the content of Chl a, b, total Chl, and carotenoids were calculated based on the Lichtenthaler and Wellburn (1983).
2.3. Determination of total anthocyanins
Anthocyanin content was measured based on the Tang et al. (2005) method. For this purpose, 500 mg of leaf powder was mixed with 10 ml of acidified ethanol with hydrochloric acid (1%) and then incubated in an incubator shaker at 4 °C for 24 h. After centrifugation (10 min at 7000 g), the absorbance of the supernatant was measured at 530 and 657 nm [17].
2.4. Evaluation of soluble and storage carbohydrate contents
To obtain the plant extract for measuring carbohydrate contents, 500 mg of the plant powder was mixed in 5 mL of 96% ethanol and placed on an incubator shaker at 4 °C for 24 h. After centrifugation (10 min at 4000 g), the supernatant was separated, and 5 ml of 70% ethanol was poured onto the remaining sediments and placed in an incubator shaker at 4 °C for one h. After centrifugation, the top solution was mixed with the previous solution. Plant extract (50 μl) was mixed with 3 ml of anthrone (150 mg of anthrone powder per 100 ml of 72% sulfuric acid) to measure soluble carbohydrates. The samples were then placed in a bain-marie at 100 °C for ten min. Finally, the sample absorbance was measured at 625 nm. Concentrations of 0, 25, 75, and 100 mg ml-1 of monohydrate glucose were used to draw the standard curve [18]. The sediments that remained after the extraction were used to measure starch carbohydrates. To do so, 1.25 ml of distilled water was added to completely dried sediments, and then 1.265 ml of perchloric acid was added to it. After centrifugation (10 min at 4000 g), the supernatant was removed, and all steps were re-applied to the residue. Then, the samples were placed on ice for 30 min. They were smoothed with filter paper and finally reached a volume of 2.5 ml. To prepare the sample solution, 0.625 ml of the extract was mixed with 2.5 ml of 0.002 Antron solution in 98% sulfuric acid, then placed in a bain-marie of 100 °C for 8 min. It was placed on ice immediately. Finally, the absorption of the samples was read at a wavelength of 630 nm [19].
2.5. Chl a fluorescence
The youngest fully developed leaves were used for measuring the maximum quantum efficiency of PSII (FV/FM) with the Handy Fluorcam FC 1000-H (Photon Systems Instruments, PSI, Czech Republic). Intact leaves attached to the plants were dark-adapted for 20 min. After dark adaptation, intact plants were immediately used to measure FV/FM. The Fluorcam consisting of a CCD camera and four fixed LED panels, one pair supplying the measuring pulses and the second pair providing actinic illumination and saturating flash, were used. FV/FM was calculated using a custom-made protocol [20,21,22]. Images were recorded during short measuring flashes in darkness. At the end of the short flashes, the samples were exposed to a saturating light pulse (3900 μmol m−2 s−1) that resulted in a transitory saturation of photochemistry and reduction of the primary quinone acceptor of PSII [22]. After reaching steady-state fluorescence, two successive series of fluorescence data were digitized and averaged, one during short measuring flashes in darkness (Fo) and the other during the saturating light flash (Fm). Fv was calculated by the expression Fv= Fm - Fo from these two parameters. The FV/FM was calculated using the ratio (Fm - Fo)/Fm. The average values and standard deviation of FV/FM were calculated by using version 7 FluorCam software.
Following overnight dark adaption, the polyphasic Chl a fluorescence (OJIP) transient on young fully formed basil leaves were evaluated using a FluorPen FP 100-MAX (photon system instruments, Drasov, Czech Republic). A saturating light of approximately 3900 µmol m-2 s-1 initiated the fluorescence measurement. The JIP test was used to assess the OJIP transients (Strasser et al. 2000). The following data from the original size was used after extraction by FluorPen software: fluorescence intensities at 50 µs (F 50 µs, considered as the minimum fluorescence F0), 2 ms (J-step, FJ), 60 ms (I-step, FI), and maximum fluorescence (Fm). At time 0 and time to obtain maximum fluorescence, the performance index was determined using the absorption basis (PIABS) and densities of QA- reducing PSII reaction centers. The parameters related to the OJIP fluorescence transients include the probability that a trapped exciton promotes an electron in ETC beyond QA- (Ψ0), the quantum yield of electron transport (φE0), the quantum yield of energy dissipation (φD0), quantum yield for primary photochemistry (φPAV), the maximum quantum yield of primary photochemistry (φP0), specific energy fluxes per reaction center (RC) for energy absorption (ABS/RC), trapped energy flux (TR0/RC), electron transport flux (ET0/RC) and dissipated energy flux (DI0/RC) were calculated according to Kalhor et al. (2018).
2.6. Statistical analysis
Statistical analysis was performed using SAS statistical analysis software (SAS Institute Inc., Cary, N.C.). The experiment was arranged on a factorial basis with a completely randomized design with 10 replications for photosynthetic measurements and three replications for physiological evaluations. All data are presented as the mean ± standard deviation (SD). Mean separations were conducted using a Duncan test protected by ANOVA at P ≤ 0.05.
3. Results
3.1. Effects of shading and harvest time on pigment concentrations of basil plants
The light intensities and harvest time significantly influenced concentrations of all photosynthetic pigments in basil leaves. In general, the purple variety contained higher levels of chlorophyll pigments under 100% and 50% light levels, while there was no considerable difference between the two basil varieties when plants were exposed to a 30% light level. At full sunlight, the highest chlorophyll content was detected at noon, while at 50% light levels, the highest chlorophyll content was performed in the morning. The purple variety had the highest levels of chlorophyll a, b, and their total amount when harvested at noon under full sunlight conditions. On the other hand, the green variety had the lowest levels of these components when harvested in the morning under the same light intensity (Figure 1A–C).
The purple variety showed the highest carotenoid content when harvested at noon and in full sunlight, as well as at 50% light in the morning and noon. Conversely, the green variety had the lowest carotenoids when harvested at the morning and noon under full sunlight treatment, and at noon with 50% light level. The purple variety also had a lower carotenoid when harvested in the morning under full sunlight treatment (Figure 1D).
The purple variety had the highest level of anthocyanin when harvested at noon under full sunlight, while the green variety had the lowest level under 30% light level during both morning and noon harvest, as well as 50% of full sunlight during morning harvest. Overall, the purple variety had enormously higher levels of anthocyanin in all treatments than the green variety. In general, the levels of anthocyanin decreased in both varieties as the light intensity decreased (Figure 1E).
3.2. Effects of shading and harvest time on leaf carbohydrate contents of basil plants
The light intensities and harvest time significantly influenced both soluble and storage carbohydrate contents in basil leaves. Both basil varieties had the highest storage and soluble carbohydrates when exposed to full sunlight and harvested in the morning. On the other hand, the lowest amount of carbohydrates was detected in the green variety when exposed to 30% sunlight and harvested at noon. The purple variety also had lower carbohydrate levels in the same light intensity when harvested at noon. In general, carbohydrate levels were higher in the morning than at noon. As light intensity decreased, the amount of carbohydrates in both varieties also reduced (Figure 2 and Figure 3).
3.3. Chlorophyll a fluorescence influenced by light intensity and day-time in basil varieties
In this study, to assess the photosynthetic functionality under different light intensities and different times of day, a fast OJIP transient was used. Due to a large number of measurements and for better understanding, the diagrams of OJIP transients were drawn separately for each variety and light intensity, while the measurement during different time courses of a day is presented in each graph (Figure 3). The most normal OJIP is seen in both varieties and at all intensities of light at 4 a.m., and with the passage of time and the onset of daylight, disturbances in different stages of OJIP, especially in the final stages occurred (Figure 3). Under different shade treatments in both basil varieties, there were declines in the final steps of the OJIP transient when the measurements were done during the daytime (8 a.m., 12, 5 p.m.).
In general, under all light intensities, the maximum quantum yield of the green variety's photosystem II (FV/FM) was higher than its ratio in the purple variety (Figure 4). Furthermore, under all light treatments in both basil varieties, the FV/FM was at its highest value at 4 a.m., then with exposure to sunlight, FV/FM was decreased at 8 a.m. and 12 and increased again at 5 p.m. The highest FV/FM was detected in green basil plants at 4 a.m. under all light treatments, while the lowest FV/FM was obtained in purple basil plants at 12 when exposed to full sunlight or 50% of sunlight conditions (Figure 4).
The highest performance index in absorbance basis (PIABS) was detected in the green variety under 50% and 30% of full sunlight at 5 p.m. belonged the lowest PIABS belonged to purple variety under full sunlight and 50% at 12. The lowest PIABS in the green variety was obtained under all light intensities at 8 a.m. The highest PIABS in the purple variety was detected under full sunlight at 4 a.m. (Figure 5).
For the green variety the highest variation in OJIP transient-derived parameters was detected for mainly DI0/RC and PIABS (Figure 6). Compared to the 4 a.m. measurement, there was an increase in DI0/RC when the measurements were done during the day (8 a.m., 12, and 5 p.m.). In the case of PIABS, there was a decrease when plants were exposed to full sunlight during the daytime measurements. The same reduction was obtained in plants exposed to 50% and 30% of full sunlight when measurements were done at 8 a.m. and 12, while at 5 p.m. there was an increase in the value of PIABS. Purple variety showed considerable variations in OJIP transient-derived parameters especially for ɸD0, DI0/RC, Vj and PIABS (Figure 6). DI0/RC, Vj and ɸD0 were sharply increased, while PIABS was dramatically decreased when the measurements were done during the day-time (8 a.m., 12, and 5 p.m.).
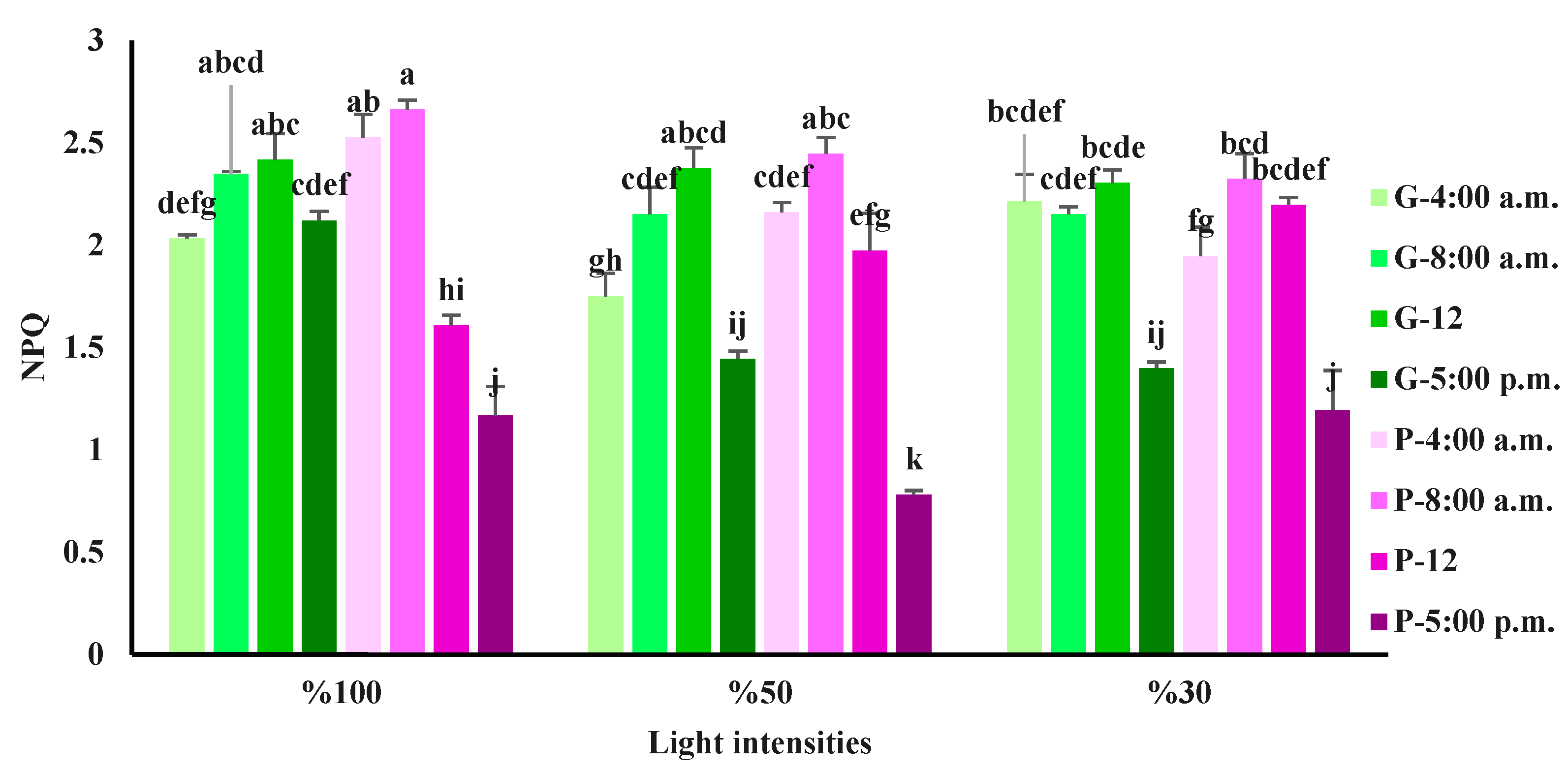
The highest NPQ was detected in the purple variety exposed to full light at 8 a.m. The lowest NPQ was also detected in the purple variety exposed to 50% of full sunlight at 5 p.m. (Figure 7). In both basil varieties; there was no difference or a slight increase in NPQ of the plants at 8 a.m. compared to its values at 4 a.m. In general, both basil varieties showed the lowest NPQ at 5 p.m.
4. Discussion
Photosynthetic pigment increased in green basil, while decreased in purple basil by reducing light intensity
Photosynthetic pigments play a critical role in facilitating the absorption and transfer of light energy in plants, thereby directly influencing photosynthetic efficiency. Among these pigments, chlorophyll is important and is a significant indicator of photosynthetic capacity (Hai-He et al., 2007; Ma et al., 2010). In general, chlorophyll levels were increased in the green variety and decreased in purple variety by reducing the light intensity (Figure 1). Many factors including light's quantity and quality affect the chlorophyll level in plants [23]. Similar to the green variety of the present study, it was found that the amount of chlorophyll in the sage increased with decreasing light intensity [13]. However, it has been shown that plants that are grown under the shadow receive lower levels of light per unit of leaf area compared to those grown under full sunlight. Consequently, it is common for shade-exposed plants to have elevated levels of chlorophyll, particularly chlorophyll b, to compensate for the decrease in received light intensity [24]. Similar to our result, higher chlorophyll content in the purple basil variety than their level in the green basil variety has been reported by Hosseini et al (2019) where plants were grown under different light spectra. It seems that the high anthocyanin level of the purple variety which is considered as a shade screen for the leaf to protect the leaf from high light intensities induces chlorophyll accumulation to compensate the decreased light level received by the mesophyll [25]. In the present study, chlorophyll levels increased at noon time when plants exposed to full sunlight condition, while at the same time, they were decreased when shadings were applied (Figure 1). The level of chlorophyll in plant leaves depends on many factors including genotype, location, light level and etc. An increase in chlorophyll levels as the result of exposure to high light intensity has been reported in green leaf plants [6,8,9]. On the other hand, a decrease in chlorophyll content due to high light intensity exposure has been reported [26,27,28].
Anthocyanins are stable pigments with the antioxidant role of protect photosynthetic system against photooxidation under stress conditions [29]. Accumulation of anthocyanins as an important pigment in leaves is affected by various factors such as temperature, access to water and light. Accumulation of anthocyanins in the leaves can act as a protective surface that reduces light harmful effects [30]. Under intense light conditions, plants undergo changes in their metabolism, structure and pigment composition in order to compete and survive in the new environment. Many plants in their leaves and fruits under high light and other stress conditions induce the synthesis of additional pigments such as anthocyanins [31]. Anthocyanin levels decreased in both basil varieties with decreasing light intensity (Figure 1). A positive correlation between anthocyanin levels and tolerance to high light stress has been proposed [32]. According to Phippen and Simon (1998), the purple basil variety has a special type of anthocyanin that is derived from cyanidin. In petunia plants, the reduction of light intensity caused a decrease in the production of anthocyanin [33]. As expected, the purple basil contained more anthocyanins levels than the green variety [34,35]. The purple variety has a higher amount of anthocyanin (0.6 to 1 mg / g fresh weight) than the green variety (less than 0.5 mg / g fresh weight), and the cause of the purple color in this variety is attributed to the accumulation of anthocyanins [36]. Due to the accumulation of anthocyanins in vacuoles [37], they increase plant tolerance by protecting the photosynthetic system against stresses such as high light radiation by increasing the amount of fluorescence [38,39].
Carotenoids serve as both pigments that receive light and protect chloroplasts when they are under stress. Under high light intensity, carotenoids primarily function to protect, while in environments with low light radiation, they absorb light and transmit its energy to chlorophyll (Ayalew, 2018). In the present study, carotenoid levels increased as light intensity decreased in green variety, with higher levels detected in the morning, while in the purple variety, carotenoid levels mainly accumulated in the noon time.
Photosynthetic functionality decreased by high light stress mainly in purple basil variety
During the process of photosynthesis, light energy is converted into chemical energy. This chemical energy is stored as carbohydrates for consumption during plant growth [40]. In our study, the amount of carbohydrates in the plant decreased by decreasing the light intensity by shading. Furthermore, the highest amount of carbohydrates was detected in the morning harvest. The light intensity can affect plant growth and stress tolerance in plants due to its effect on the synthesis of carbohydrate products [41]. Decreased and weak growth due to exposure to low light intensities is mainly due to reduced energy inputs, obtained from light reactions (ATP and NADPH), for carbon reactions resulting in a decrease in CO2 fixation and as a consequence reduced sugar synthesis as the cytoskeleton of plant structure [40]. Similar results were obtained in a survey of the effect of different light intensities on sunflowers [14]. In another study on roses, it was found that plants contained less carbohydrates when exposed to a high and stressful light intensity which was consistent with the results of the present study at noon. Decreased carbohydrate production when exposed to high light intensity is known to reduce the photosystem II functionality and thus reduce the production of sugars in the plant [32].
Photosynthetic reactions in plants depend on environmental conditions. The quantity and quality of light are among the factors that can affect photosynthetic reactions [42]. Environmental stresses usually increase Fo. The increase in FO indicates the degradation of the electron transfer chain of photosystem II due to the reduction of QA capacity and its complete lack of oxidation due to the slow flow of electrons along the path of photosystem II and the total inactivation of photosystem II [43]. The increase in FO was also attributed to damage to the D1-bound protein of photosystem II [44,45]. Maximum fluorescence (FP) is related to the maximum emission of fluorescence due to the reduction of all electron carriers [46]. In this study, FP levels decreased in both varieties when light intensity increased during the day. In the experiment on roses, the amount of FP decreased when the rose was exposed to high light intensity [32]. A decrease in FP is likely associated with reduced activity of the water-degrading enzyme complex as well as electron transfer cycles in or around photosystem II [47]. FV/FM is calculated from the ratio of variable fluorescence to maximum fluorescence. This index indicates the maximum quantum efficiency of photosystem II for converting absorbed light into chemical energy [48]. This parameter decreases during stress conditions. The decrease of this index is the result of damage to the reaction centers of photosystem II. It indicates the occurrence of photoinhibition due to environmental stresses[49]. The FV/FM index is used to diagnose photosynthetic system disorders and its reduction indicates a decrease in photochemical productivity in photosystem II and damage to the photosynthetic system [50]. FV/FM represents the ratio of light used in the photosynthesis process to the total light absorbed by chlorophyll; its decrease indicates a decrease in the photosynthesis capacity [51]. Decreased photochemical performance is due to the occurrence of inhibition due to the inactivation of photosystem II reaction centers and damage to the D1 protein [49]. In the present study conducted at 4 a.m. in both varieties, FV/FM was at its highest value, but when the light intensity was increased (noon), the FV/FM was decreased. It then rose again with decreasing light intensity at 5 p.m. An increase in FV/FM reduces energy loss in the form of heat in the plant [52]. The results of this study were consistent with the research of Hazrati et al. (2016), which reported that decreasing FV/FM was associated with increased light intensity and increased heat dissipation in the form of heat [53]. The decrease in FV/FM is due to photosystem II damage, which reduces photosynthesis efficiency [54]. The green variety showed a higher FV/FM compared to the purple variety, which is in line with the study of Hosseini et al. (2019).
PIABS is a parameter that is defined as an indicator of system performance per absorbed light [55]. This index is one of the most useful biophysical parameters in showing the difference between the response of photosystem II to normal conditions and conditions in which the plant is under stress [5]. In other words, PIABS is an indicator of photon absorption performance. This parameter is very sensitive to environmental changes and stresses and is successfully used to track photosynthetic performance when the plant is exposed to many abiotic stresses, including high light stress [5], elevated temperatures [56], salinity stress [57], drought [58], and nutrient deficiency [59]. The decrease in PIABS may be due to electron inhibition due to the decline in the normal performance of photosystem II [60]. In the present study, PIABS decreased with increasing light intensity during the day. In wheat, the PIABS decreased from 12 to 5 p.m. compared to its values in the morning [61].
NPQ is an indicator of the dissipation received energy in the form of heat. Elevation in the NPQ indicates the high capacity of the xanthophyll cycle and the plant's ability to withstand stress through energy loss in the form of heat [62]. The NPQ is highly related to the functioning of the xanthophyll cycle, which facilitates energy dissipation in the form of heat under high light intensities [63]. In the present study, NPQ was highest in the green variety at noon time, which may be related to the high level of carotenoids at noon time in the green variety (Figure 1). It has been shown that plants that are exposed to high light have higher levels of NPQ than plants exposed to low light [8,54]. In the purple variety, NPQ reached its highest level at 8 a.m and decreased at noon with the highest light intensity during the measurement. Studies have shown that the accumulation of anthocyanin in plants reduces NPQ under conditions of high light intensity. Indeed, anthocyanin act as a light shield and protect the plant's photosynthetic system from high light intensity [64]. However, in the present study, higher anthocyanin level in the purple variety did not provide more protection on the photosynthetic apparatus. This can be more elucidated by lower FV/FM and NPQ during the noon time in the purple variety, while in the green variety, which contained lower anthocyanin levels, showed higher FV/FM and NPQ during the noon time. It has been reported that although anthocyanin protects plants from high light stress by having antioxidant properties, it reduces photosynthetic functionality [25]. Agati et al. (2022) challenged the known role of anthocyanin in the photoprotection of photosynthesis by showing a shortage of proper investigation using red and green leaves plants or the discrepancy between the finding of controlled environments with the field [65]. Therefore, it is still early to make a firm conclusion related to the role of anthocyanin on protection over photosynthesis machinery..
5. Conclusions
In the present study, photosynthetic and protective pigments of green and purple basil varieties were influenced by shade level and measurement time during the day. Although the purple variety showed higher levels of chlorophyll and anthocyanin pigments than the green variety, these pigments decreased considerably when shade treatments were applied. The green variety showed a lower photoinhibition level during the day than the purple variety. As a result, there were lower variations on the fate of excited energy in the green variety compared to the purple one. The higher photoinhibition during day time in the purple variety can be related to their lower capacity to dissipate the extra energy on the photosynthetic system as the heat (NPQ). In conclusion, higher anthocyanin levels in purple variety did not necessarily prevent photoinhibition in purple variety.
References
- Ioannidis, D., L. Bonner, and C.B. Johnson, UV-B is required for normal development of oil glands in Ocimum basilicum L.(Sweet Basil). Annals of Botany, 2002, 90, 453–460. [CrossRef] [PubMed]
- Labra, M., et al., Morphological characterization, essential oil composition and DNA genotyping of Ocimum basilicum L. cultivars. Plant Science 2004, 167, 725–731. [CrossRef]
- Sajjadi, S.E. Analysis of the essential oils of two cultivated basil (Ocimum basilicum L.) from Iran. DARU Journal of Pharmaceutical Sciences, 2006, 14, 128–130. [Google Scholar]
- Paez, A., et al., Growth, soluble carbohydrates, and aloin concentration of Aloevera plants exposed to three irradiance levels. Environmental and Experimental Botany, 2000, 44, 133–139. [CrossRef]
- Ashrostaghi, T., et al., Light intensity: The role player in cucumber response to cold stress. Agronomy, 2022, 12, 201. [CrossRef]
- Esmaeili, S., et al., Elevated light intensity compensates for nitrogen deficiency during chrysanthemum growth by improving water and nitrogen use efficiency. Scientific Reports, 2022, 12, 10002. [CrossRef]
- Long, S.P., S. Humphries, and P.G. Falkowski, Photoinhibition of photosynthesis in nature. Annual review of plant biology, 1994, 45, 633–662. [CrossRef]
- Ghorbanzadeh, P., et al., Dependency of growth, water use efficiency, chlorophyll fluorescence, and stomatal characteristics of lettuce plants to light intensity. Journal of Plant Growth Regulation, 2021, 40, 2191–2207. [CrossRef]
- Vatankhah, A., et al., Plants exposed to titanium dioxide nanoparticles acquired contrasting photosynthetic and morphological strategies depending on the growing light intensity: a case study in radish. Scientific Reports, 2023, 13, 5873. [CrossRef]
- Zhang, S., K. Ma, and L. Chen, Response of photosynthetic plasticity of Paeonia suffruticosa to changed light environments. Environmental and Experimental Botany, 2003, 49, 121–133. [CrossRef]
- Whitelam, G.C. and K.J. Halliday, Light and plant development. 2007: Blackwell Pub.
- Tesfaye, K., S. Walker, and M. Tsubo, Radiation interception and radiation use efficiency of three grain legumes under water deficit conditions in a semi-arid environment. European journal of Agronomy, 2006, 25, 60–70. [CrossRef]
- Zervoudakis, G., et al., Influence of light intensity on growth and physiological characteristics of common sage (Salvia officinalis L.). Brazilian archives of biology and technology, 2012, 55, 89–95. [CrossRef]
- De la Mata, L., et al., Study of the senescence process in primary leaves of sunflower (Helianthus annuus L.) plants under two different light intensities. Photosynthetica, 2013, 51, 85–94. [CrossRef]
- Zhu, H., et al., Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (Brassica campestris ssp. Chinensis Makino). PloS one, 2017, 12, e0179305.
- Santana, A.C., et al., Effect of harvest at different times of day on the physical and chemical characteristics of vegetable-type soybean. Food Science and Technology, 2012, 32, 351–356. [CrossRef]
- Teng, S., et al., Sucrose-specific induction of anthocyanin biosynthesis in Arabidopsis requires the MYB75/PAP1 gene. Plant physiology, 2005, 139, 1840–1852. [CrossRef]
- Irigoyen, J., D. Einerich, and M. Sánchez-Díaz, Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiologia plantarum, 1992, 84, 55–60. [CrossRef]
- McCready, R., et al., Determination of starch and amylose in vegetables. Analytical chemistry, 1950, 22, 1156–1158. [CrossRef]
- Aliniaeifard, S., P. Malcolm Matamoros, and U. van Meeteren, Stomatal malfunctioning under low VPD conditions: induced by alterations in stomatal morphology and leaf anatomy or in the ABA signaling? Physiologia plantarum, 2014, 152, 688–699. [CrossRef]
- Aliniaeifard, S. and U. van Meeteren, Natural variation in stomatal response to closing stimuli among Arabidopsis thaliana accessions after exposure to low VPD as a tool to recognize the mechanism of disturbed stomatal functioning. Journal of Experimental Botany, 2014, 65, 6529–6542. [CrossRef]
- Genty, B., J.-M. Briantais, and N.R. Baker, The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta (BBA)-General Subjects, 1989, 990, 87–92. [CrossRef]
- Abreu, P.P., et al., Photosynthetic responses of ornamental passion flower hybrids to varying light intensities. Acta physiologiae plantarum, 2014, 36, 1993–2004. [CrossRef]
- Wadhwa, R., N. Kumari, and V. Sharma, Varying light regimes in naturally growing Jatropha curcus: pigment, proline and photosynthetic performance. Journal of Stress Physiology & Biochemistry, 2010, 6.
- Zhao, S., et al., Anthocyanin accumulation provides protection against high light stress while reducing photosynthesis in apple leaves. International Journal of Molecular Sciences, 2022, 23, 12616. [CrossRef]
- Biswal, A.K., et al., Light intensity-dependent modulation of chlorophyll b biosynthesis and photosynthesis by overexpression of chlorophyllide a oxygenase in tobacco. Plant physiology, 2012, 159, 433–449. [CrossRef] [PubMed]
- Muhidin, et al. The effect of shade on chlorophyll and anthocyanin content of upland red rice. in IOP Conference Series: Earth and Environmental Science. 2018. IOP Publishing.
- Zhang, H., et al., Adaptive changes in chlorophyll content and photosynthetic features to low light in Physocarpus amurensis Maxim and Physocarpus opulifolius “Diabolo”. PeerJ, 2016, 4, e2125. [CrossRef]
- He, F., et al., Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules, 2010, 15, 9057–9091. [CrossRef]
- Oren-Shamir, M. and A. Levi-Nissim, UV-light effect on the leaf pigmentation of Cotinus coggygria ‘Royal Purple’. Scientia horticulturae, 1997, 71, 59–66. [CrossRef]
- Merzlyak, M.N. and O.B. Chivkunova, Light-stress-induced pigment changes and evidence for anthocyanin photoprotection in apples. Journal of Photochemistry and Photobiology B: Biology, 2000, 55, 155–163. [CrossRef]
- Bayat, L., et al., Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants, 2018, 10.
- Albert, N.W., et al., Light-induced vegetative anthocyanin pigmentation in Petunia. Journal of experimental botany, 2009, 60, 2191–2202. [CrossRef] [PubMed]
- Hosseini, A., et al., Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiology and Molecular Biology of Plants, 2019, 25, 741–752. [CrossRef] [PubMed]
- Tenore, G.C., et al., Antioxidant and antimicrobial properties of traditional green and purple “Napoletano” basil cultivars (Ocimum basilicum L.) from Campania region (Italy). Natural product research, 2017, 31, 2067–2071. [CrossRef]
- Landi, M., et al., Antioxidant and photosynthetic response of a purple-leaved and a green-leaved cultivar of sweet basil (Ocimum basilicum) to boron excess. Environmental and experimental botany, 2013, 85, 64–75. [CrossRef]
- Chanoca, A., et al., Anthocyanin vacuolar inclusions form by a microautophagy mechanism. The plant cell, 2015, 27, 2545–2559. [CrossRef] [PubMed]
- Gould, K.S., D.A. Dudle, and H.S. Neufeld, Why some stems are red: cauline anthocyanins shield photosystem II against high light stress. Journal of Experimental Botany, 2010, 61, 2707–2717. [CrossRef]
- Hosseini, A., et al., Photosynthetic and growth responses of green and purple basil plants under different spectral compositions. Physiology and Molecular Biology of Plants, 2019, 25, 741–752. [CrossRef] [PubMed]
- Bryant, D.A. and N.-U. Frigaard, Prokaryotic photosynthesis and phototrophy illuminated. Trends in microbiology, 2006, 14, 488–496. [CrossRef]
- Cakmak, I. and V. Römheld, Boron deficiency-induced impairments of cellular functions in plants. Plant and Soil, 1997, 193, 71–83. [CrossRef]
- Chen, M., J. Chory, and C. Fankhauser, Light signal transduction in higher plants. Annu. Rev. Genet., 2004, 38, 87–117. [CrossRef]
- Zlatev, Z.S. and I.T. Yordanov, Effects of soil drought on photosynthesis and chlorophyll fluorescence in bean plants. Bulg. J. Plant Physiol, 2004, 30, 3–18.
- Shomali, A., et al., Artificial neural network (ANN)-based algorithms for high light stress phenotyping of tomato genotypes using chlorophyll fluorescence features. Plant Physiology and Biochemistry, 2023, 2023, 107893.
- Strasser, R.J., A. Srivastava, and M. Tsimilli-Michael, The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing photosynthesis: mechanisms, regulation and adaptation, 2000, 25, 445–483.
- Mehta, P., et al., Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiology and biochemistry, 2010, 48, 16–20. [CrossRef] [PubMed]
- Chaves, M.M., J. Flexas, and C. Pinheiro, Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of botany, 2009, 103, 551–560. [CrossRef]
- Kocheva, K., et al., Evaluation of chlorophyll fluorescence and membrane injury in the leaves of barley cultivars under osmotic stress. Bioelectrochemistry, 2004, 63, 121–124. [CrossRef]
- Shomali, A., et al., Photoinhibition in horticultural crops: An overview of the effect of light quality and signaling in the underlying photoprotection mechanisms. International Journal of Horticultural Science and Technology, 2023, 10, 39–50.
- Willadino, L., et al., Estresse salino em duas variedades de cana-de-açúcar: enzimas do sistema antioxidativo e fluorescência da clorofila. Revista Ciência Agronômica, 2011, 42, 417–422. [CrossRef]
- Li, J., et al., Influence of drought stress on photosynthetic characteristics and protective enzymes of potato at seedling stage. Journal of The Saudi Society of Agricultural Sciences, 2017, 16, 82–88. [CrossRef]
- Broetto, F., H.M. Duarte, and U. Lüttge, Responses of chlorophyll fluorescence parameters of the facultative halophyte and C3–CAM intermediate species Mesembryanthemum crystallinum to salinity and high irradiance stress. Journal of plant physiology, 2007, 164, 904–912. [CrossRef]
- Hazrati, S., et al., Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiology and Biochemistry, 2016, 106, 141–148. [CrossRef]
- Shomali, A., et al., Synergistic effects of melatonin and gamma-aminobutyric acid on protection of photosynthesis system in response to multiple abiotic stressors. Cells, 2021, 10, 1631.
- Kalhor, M.S., et al., Enhanced salt tolerance and photosynthetic performance: Implication of ɤ-amino butyric acid application in salt-exposed lettuce (Lactuca sativa L.) plants. Plant physiology and biochemistry, 2018, 130, 157–172. [CrossRef] [PubMed]
- Martinazzo, E.G., A. Ramm, and M.A. Bacarin, The chlorophyll a fluorescence as an indicator of the temperature stress in the leaves of Prunus persica. Brazilian journal of plant physiology, 2012, 24, 237–246.
- Mathur, S., P. Mehta, and A. Jajoo, Effects of dual stress (high salt and high temperature) on the photochemical efficiency of wheat leaves (Triticum aestivum). Physiology and Molecular Biology of Plants, 2013, 19, 179–188. [CrossRef] [PubMed]
- Sousaraei, N., et al., Screening of tomato landraces for drought tolerance based on growth and chlorophyll fluorescence analyses. Horticulture, environment, and biotechnology, 2021, 62, 521–535. [CrossRef]
- Kalaji, H.M., et al., Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant physiology and biochemistry, 2014, 81, 16–25. [CrossRef]
- Liang, Y., et al., Responses of Jatropha curcas seedlings to cold stress: photosynthesis-related proteins and chlorophyll fluorescence characteristics. Physiologia Plantarum, 2007, 131, 508–517. [CrossRef]
- Zivcak, M., et al., Measurements of chlorophyll fluorescence in different leaf positions may detect nitrogen deficiency in wheat. 2014.
- Porcar-Castell, A., et al., Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: mechanisms and challenges. Journal of experimental botany, 2014, 65, 4065–4095. [CrossRef]
- Shomali, A., et al., Diverse physiological roles of flavonoids in plant environmental stress responses and tolerance. Plants, 2022, 11, 3158. [CrossRef]
- Zhu, H., et al., Anthocyanins function as a light attenuator to compensate for insufficient photoprotection mediated by nonphotochemical quenching in young leaves of Acmena acuminatissima in winter. Photosynthetica 2018, 56, 445–454. [CrossRef]
- Agati, G., et al., Anthocyanins in photoprotection: knowing the actors in play to solve this complex ecophysiological issue. The New Phytologist, 2021, 232, 2228. [CrossRef] [PubMed]
Figure 1.
Chlorophyll a (A), chlorophyll b (B), total chlorophyll (C), total carotenoids (D), and anthocyanin (E) of green (G) and purple (P) varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at the different time course of a day (G-M: Green basil harvested in the early morning, G-N: Green basil harvested at noon, P-M: Purple basil harvested in the early morning and P-N: Purple basil harvested at noon). Bars are means ± SE.
Figure 1.
Chlorophyll a (A), chlorophyll b (B), total chlorophyll (C), total carotenoids (D), and anthocyanin (E) of green (G) and purple (P) varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at the different time course of a day (G-M: Green basil harvested in the early morning, G-N: Green basil harvested at noon, P-M: Purple basil harvested in the early morning and P-N: Purple basil harvested at noon). Bars are means ± SE.
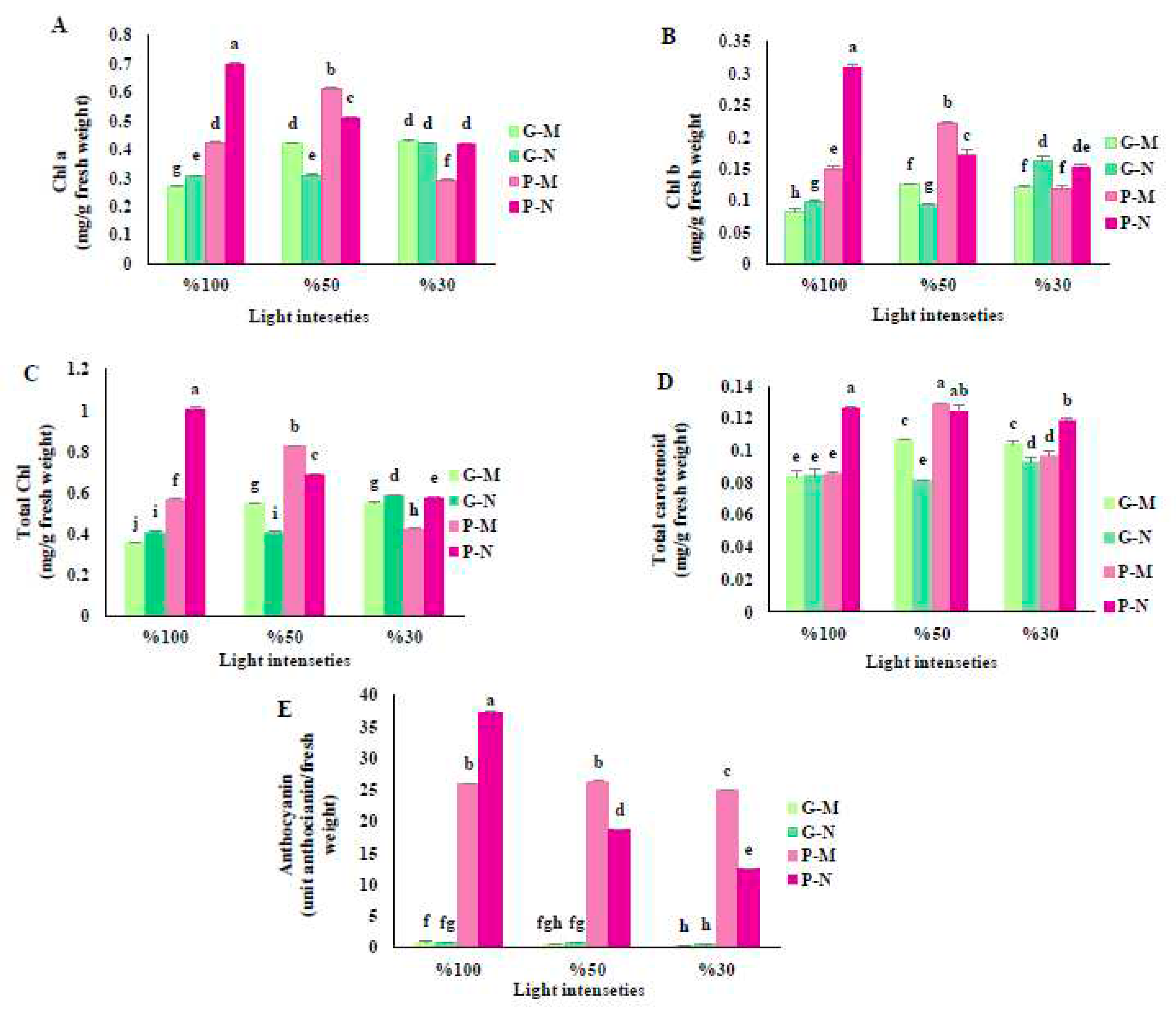
Figure 2.
Storage carbohydrate (A) and soluble carbohydrate (B) of green (G) and purple (P) varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at the different time course of a day (G-M: Green basil harvested in the early morning, G-N: Green basil harvested at noon, P-M: Purple basil harvested in the early morning and P-N: Purple basil harvested at noon). Bars are means ± SE.
Figure 2.
Storage carbohydrate (A) and soluble carbohydrate (B) of green (G) and purple (P) varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at the different time course of a day (G-M: Green basil harvested in the early morning, G-N: Green basil harvested at noon, P-M: Purple basil harvested in the early morning and P-N: Purple basil harvested at noon). Bars are means ± SE.
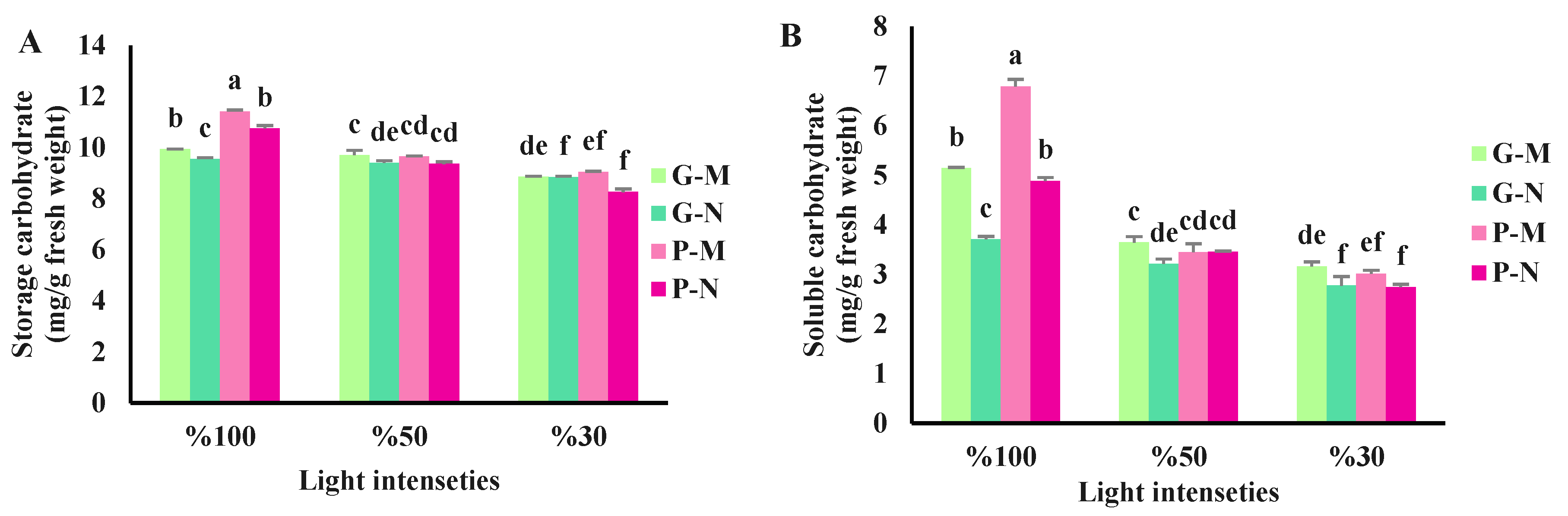
Figure 3.
OJIP transient trend in green and purple varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at various time courses of a day (full sunlight conditions (A), 50% of full sunlight (B), 30% of full sunlight (C)) and Purple (full sunlight conditions (D), 50% of full sunlight (E), 30% of full sunlight (F)) basil at different measurement time-points. Bars are means ± SE.
Figure 3.
OJIP transient trend in green and purple varieties of basil exposed to different light intensities (100: 100% sunlight, 50: 50% of sunlight and 30: 30% of sunlight) and harvested at various time courses of a day (full sunlight conditions (A), 50% of full sunlight (B), 30% of full sunlight (C)) and Purple (full sunlight conditions (D), 50% of full sunlight (E), 30% of full sunlight (F)) basil at different measurement time-points. Bars are means ± SE.
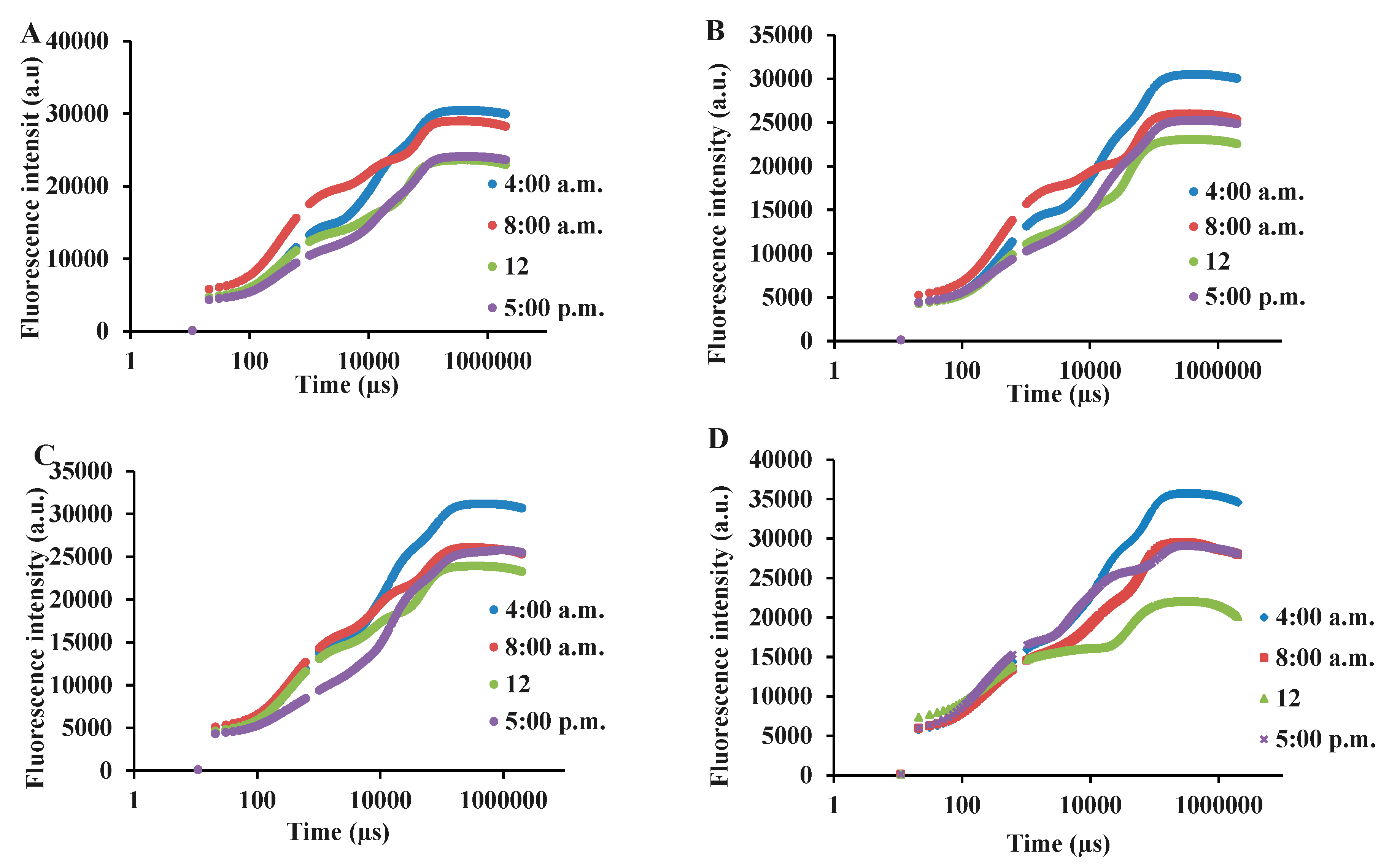
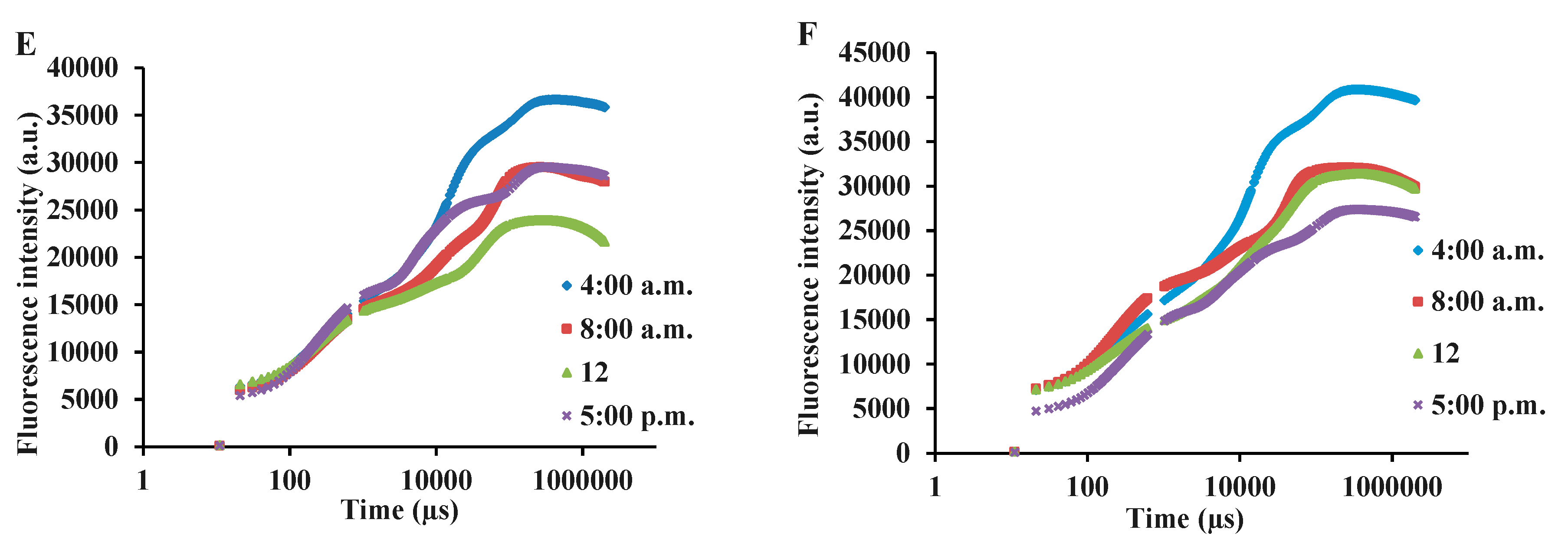
Figure 4.
Maximum quantum yield of photosystem II (FV/FM) from the fluorescence transient exhibited by leaves of green (G) and purple (p) basil grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at an additional time of a day. Bars represent means ± SD.
Figure 4.
Maximum quantum yield of photosystem II (FV/FM) from the fluorescence transient exhibited by leaves of green (G) and purple (p) basil grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at an additional time of a day. Bars represent means ± SD.
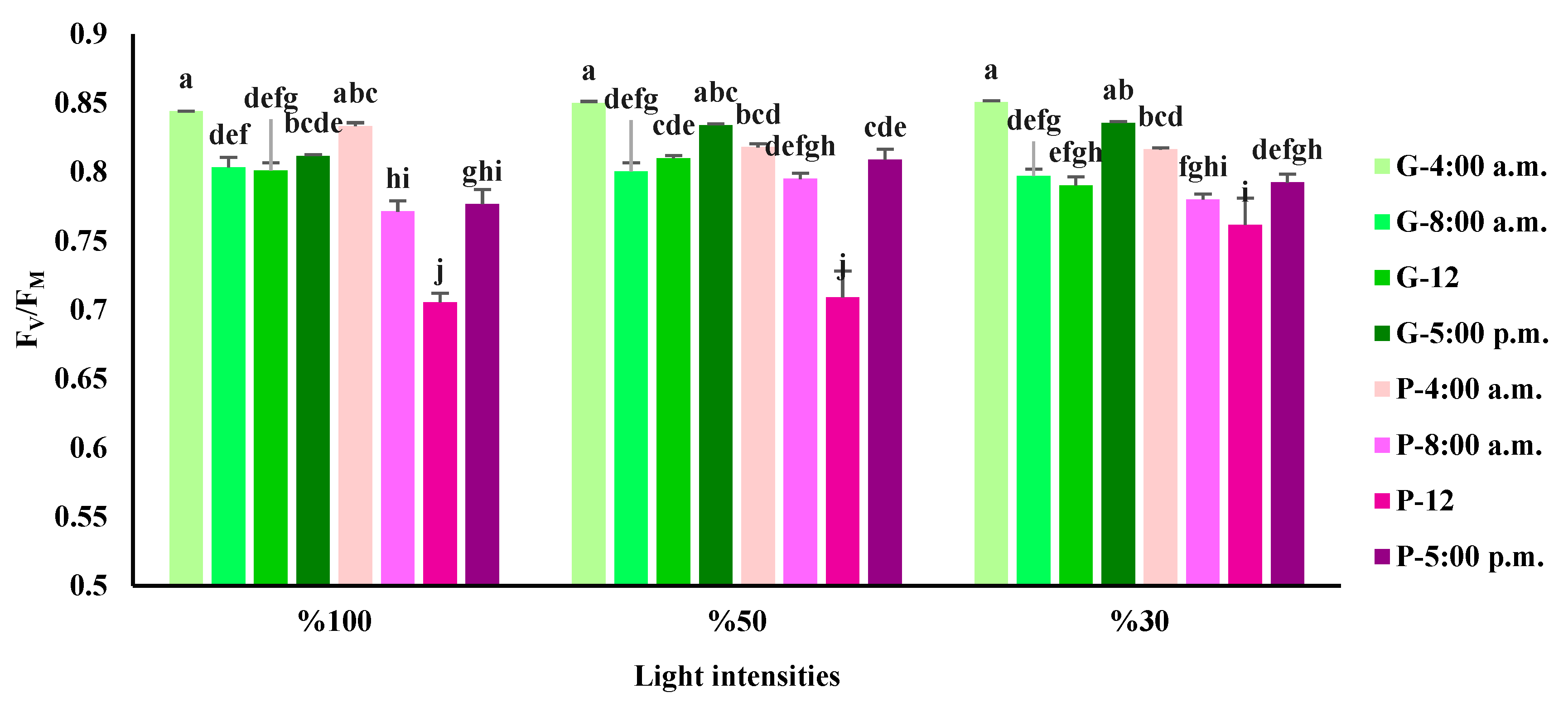
Figure 5.
Performance index in absorbance basis (PIABS) obtained from the fluorescence transient exhibited by leaves of green (G) and purple (P) basil plants grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at various time courses of a day. Bars represent means ± SD.
Figure 5.
Performance index in absorbance basis (PIABS) obtained from the fluorescence transient exhibited by leaves of green (G) and purple (P) basil plants grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at various time courses of a day. Bars represent means ± SD.
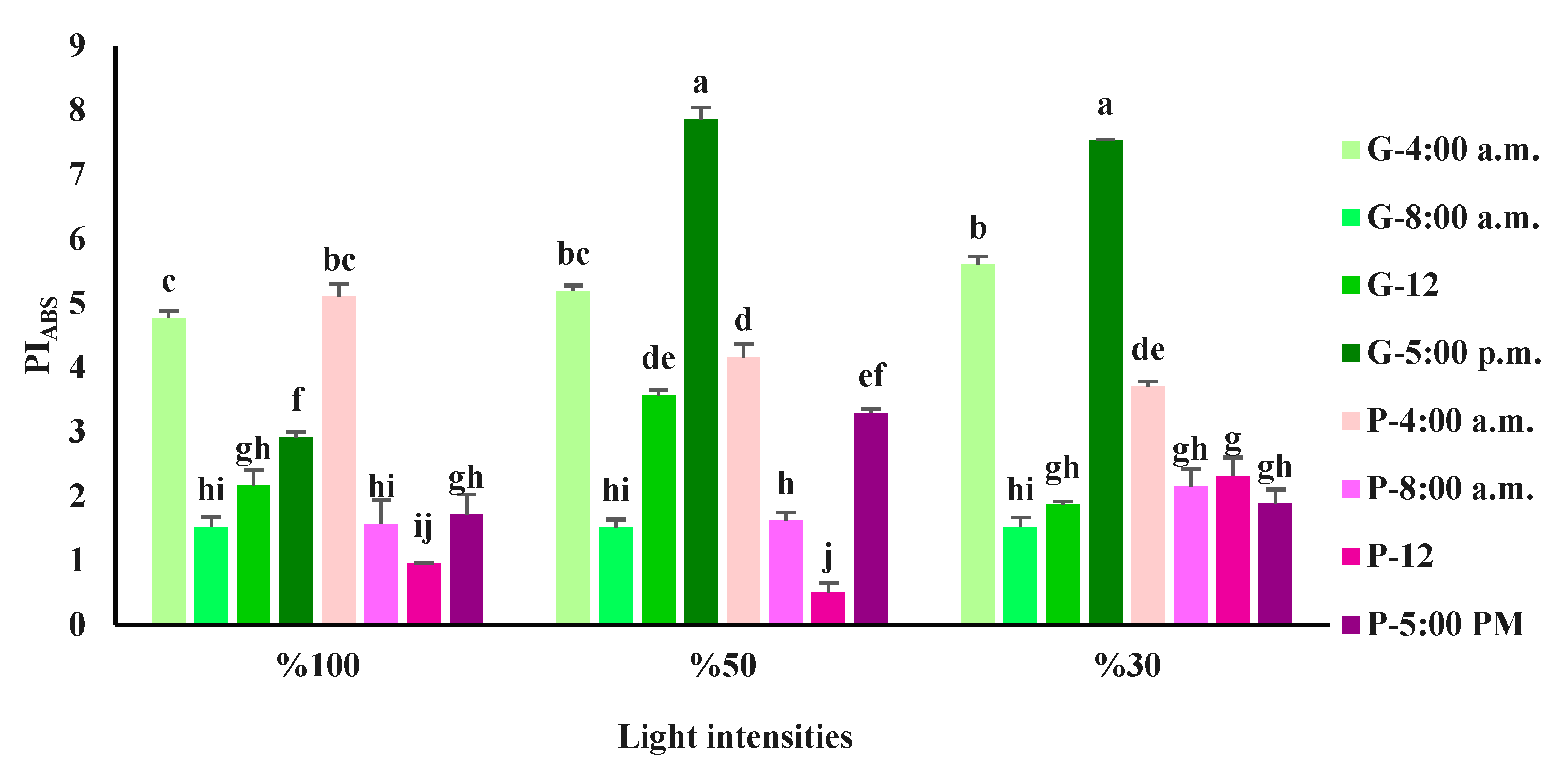
Figure 6.
Spider plot of OJIP test parameters derived from the fluorescence transient displayed by leaves of purple and green basil plants cultivated under various light intensities [full sunlight, 50%, and 30% light intensity of full sunlight] at different time courses of a day. The derived parameter values were displayed in comparison to the plants measured at 4 a.m.
Figure 6.
Spider plot of OJIP test parameters derived from the fluorescence transient displayed by leaves of purple and green basil plants cultivated under various light intensities [full sunlight, 50%, and 30% light intensity of full sunlight] at different time courses of a day. The derived parameter values were displayed in comparison to the plants measured at 4 a.m.
Figure 7.
Non-Photochemical Quenching (NPQ) from the fluorescence transient exhibited by leaves of green (G) and purple (P) basil plants grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at a different time of a day. Bars represent means ± SD.
Figure 7.
Non-Photochemical Quenching (NPQ) from the fluorescence transient exhibited by leaves of green (G) and purple (P) basil plants grown under different light intensities (100: 100% sunlight, 50: 50% of sunlight, and 30: 30% of sunlight) and harvested at a different time of a day. Bars represent means ± SD.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



