
Submitted:
17 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Antibiotics were once called miracle drugs which hit the market with astounding impact as it was considered the ultimate cure for infectious diseases in the mid-20th century. However, today nearly all bacteria that afflict humankind have become resistant to these wonder drugs once developed to stop them, imperilling the foundation of modern medicine. During the COVID pandemic, there was a surge in macrolide use to treat secondary infections and this persistent use of macrolide antibiotics has provoked the emergence of macrolide resistance. In view of the current dearth of new antibiotics in the pipeline, it is essential to find an alternative way to combat drug resistance. Antibiotic potentiators or adjuvants are non-antibacterial active molecules that, when combined with antibiotics, increase their activity. Thus, potentiating the existing antibiotics is one of the promising approaches to tackle and minimize the impact of antimicrobial resistance. Several natural and synthetic compounds have demonstrated effectiveness in potentiating macrolide antibiotics against MDR pathogens. The present review summarizes the different resistance mechanisms adapted by bacteria to resist macrolides and further emphasizes the major macrolide potentiators identified which could serve to revive the antibiotic and can be used for the reversal of macrolide resistance.
Keywords:
antibiotic
; antimicrobial resistance
; azithromycin
; macrolide resistance
; multidrug resistance
; potentiation
; adjuvants
; macrolide potentiator
1. Introduction
Macrolides are one of the most clinically significant and widely prescribed drugs worldwide after β-lactams, used to treat both Gram-positive and Gram-negative bacterial infections [1]. During the COVID-19 pandemic, the use of macrolide antibiotics was greatly increased due to their efficacy in treating community-acquired respiratory tract infections, anti-inflammatory and immunomodulatory properties though there were no sufficient studies evidencing treatment with macrolides, alone or in combination was effective in treating COVID patients [2,3,4]. Overall, this extensive use acted as a strong selective pressure contributing to the development and expansion of macrolide-resistant determinants amongst both Gram-positive and Gram-negative bacteria [5]. Recently, the WHO classified antibiotic-resistant priority pathogens as critical, high, and medium priority based on the significance and severity of the infection caused by these organisms and the urgency of the need for new antibiotics against these pathogens. Unsurprisingly, this list included macrolide-resistant organisms as high-priority pathogens [6]. In times of increasing antibiotic resistance, it is of critical importance to study and understands the bacterial resistance mechanisms, which could help in the development of new drugs or find other alternative approaches that can help to reduce the emergence of resistant pathogens. Antibiotic potentiator is one such approach which can restore the activity of antibiotics against MDR pathogens. Potentiating the activity of an existing antibiotic significantly reinstitutes the therapeutic option and uncovering such molecules that increase the activity of the antibiotic has recently gained much attention [7]. Several reviews have emphasized antibiotic resistance and the significance of potentiators, but there is a notable absence of focus on macrolide antibiotics, despite their broad clinical use [8,9]. In this review, the authors summarise the different mechanisms which Gram-negative bacteria adopt for macrolide resistance as well as the most recent discoveries and developments in the field of macrolide potentiators.
2. Resistance mechanisms to macrolide: Special emphasis on Gram-negative pathogens
Macrolides are one of the most frequently used antibiotics with azithromycin being at the top in many countries around the world [10,11]. Macrolides are classified based on their macrolactone skeleton and mainly fall under 14-, 15- and 16-membered rings (Figure 1). Macrolides are especially known for their enhanced activity against Gram-positive bacterial infections, however, this group of antibiotics has also been used to treat multiple Gram-negative infections caused by Bordetella pertussis, Neisseria gonorrhoeae, Chlamydia spp., and Campylobacter spp. (Table 1). Further, azithromycin has demonstrated a potential role in disaggregating biofilms produced by non-fermenter Gram-negative pathogen Pseudomonas spp. [12,13]. This has opened the door for researchers and clinicians to research the synergistic effect of azithromycin with antipseudomonal antibiotics. Also, azithromycin has shown potential efficacy in the treatment of enterobacterial infections and has been widely used in clinical settings to treat diarrheogenic and systemic infections caused by different enterobacterial species such as Shigella spp. and Salmonella typhi. Apart from azithromycin, other macrolides such as fluoroketolides viz. solithromycin have also shown potential in-vitro activity against clinically relevant Enterobacteriaceae [14]. The enhanced activity of azithromycin against Enterobacteriaceae and its inappropriate and indiscriminate use has led to the emergence of resistance in different members of this family worldwide [15]. Bacterial resistance to macrolide antibiotics is mainly observed due to two major reasons such as the narrow binding affinity of the drug to its target or the efflux out of the macrolides from the bacterial cells. The binding affinity of the drug alters due to the modification of either the bacterial ribosomes or the target modification in the antibiotic, whereas the efflux mechanism appears due to high efflux activity within the cell or alteration of membrane permeability. Apart from these, drug-inactivating mechanisms are also significant for macrolide resistance and are chiefly observed in Gram-negative pathogens. The different mechanism of macrolide resistance and their association with mobile genetic elements are presented in Table 2.
2.1. Target modification
The primary reason for macrolide resistance in bacterial pathogens is due to the modifications in the ribosomal target site. The most prominent target site modification is the methylation of 23S rRNA wherein it is modified by adenine-N6 methyltransferases post-transcriptionally through the addition of one or two methyl groups to an adenine, mainly at position A2058, of 23S rRNA. This methylation is mainly done by adenine-N6 methyltransferases coded by the erythromycin ribosomal methylase (erm) family genes [16]. Apart from the methylation of rRNA, mutation in the ribosomal RNA have also been reported to confer resistance to various macrolides and amongst them, the mutation at position A2058 or A2059 in the ribosomal RNA is considered the significant one which leads to alteration of the ribosomal target site and eventually inhibits the binding of macrolide antibiotics [17].
The substitution of 23s rRNA in the A2058 or A2059 positions has been well-established for causing macrolide resistance in Enterobacteriaceae and other Gram-positive isolates alike [18]. Apart from these two, alterations at A745, A752, U754, G2057, A2032, A2062, A2503, U2609, C2610 or C2611 have also been described in Enterobacteriaceae which can affect the activity of macrolides [15]. Ribosomal protein alterations such as alterations in L4 encoded by the rplD gene and L22 encoded by the rplV gene has also been identified to play a major role in the development of macrolide resistance in E. coli [19].
2.2. Bacterial efflux mechanism towards macrolide resistance
The efflux pump mechanism for macrolide resistance in bacteria is one another significant mechanism to evade antibiotic action by minimizing the intracellular concentration of the drug. Several efflux proteins have been documented in the bacterial genome which can transport specific molecules or a range of substrates from within the cells to outside. These efflux pumps can be encoded either in plasmids or on chromosomes and provide resistance to multiple classes of antibiotics [20]. Among them, the AcrAB-TolC-like RND-type efflux pumps are known to be the most significant ones and are present in a majority of the enterobacterial isolates. The effect of AcrAB-TolC in the extrusion of macrolides is due to the development of mutants for the expression of genes encoding this efflux pump. Apart from AcrAB-TolC, other pumps like MdtEF-TolC (RND-type) or MacAB-TolC (ABC transporter) have also been identified to play an important role in the extrusion of macrolides from the bacterial cells [21]. Other than these chromosomally encoded efflux pumps, several transferable pumps have also been described which are known to be more substrate-specific and result in increasing antimicrobial resistance. Mef and Msr subfamilies belonging to MFS & ABC efflux families respectively, are encoded on plasmids and are mainly responsible for efflux pump-mediated macrolide resistance in bacteria [22]. Till now, five transferable efflux pumps associated with macrolide resistance have been described in enterobacterial isolates, and among them, three belong to the ABC efflux family viz. msr(A), msr(D), and msr(E) and the rest belong to MF family mef(A) and mef(B). Currently, mef(A) is the most frequently reported efflux pump among Enterobacteriaceae [23].
2.3. Enzymatic macrolide inactivation
The enzymatic degradation of macrolide antibiotics is another mode of resistance mechanism carried out by two different classes of enzymes viz. macrolide phosphotransferases and macrolide esterases (Figure 2). Phosphotransferases are macrolide-inactivating enzymes commonly reported in bacterial pathogens (Table 2). To date, six macrolide resistance phosphotransferase encoding genes have been described and four of them viz. mphA, mphB, mphD, & mphE have been described in Enterobacteriaceae of which mphA being the most prevalent one [24]. This enzyme alters the structure of 14-, 15-, and 16-membered lactone rings of macrolide by introducing phosphate to the 2/-hydroxyl group of the macrolide amino sugar, which eventually inhibits the interaction of the antibiotic with A2058. The hydrolytic inactivation of macrolide can also be mediated by the presence of esterases enzymes encoded by ere gene which is known to be widespread in a variety of enterobacterial isolates such as Klebsiella, Enterobacter or Citrobacter (Table 2). Because of its esterase activity, this enzyme encoded by ere(A) and ere(B) hydrolyzes the ester bond at the macrolactonic ring. Interestingly, the substrate profile of both enzymes differs from each other.
3. Approaches to tackle macrolide resistance
This section of the review discusses the various therapeutic modalities that have been devised to combat macrolide resistance, like the development of vaccines or immunotherapeutics, combination medication therapy, nano-antibiotics, and macrolide potentiation. Clarithromycin-resistant H. pylori have become a global concern considering the unavailability of any effective medication against it. The H. pylori infection is aggravated by its resistance to even second-generation macrolides. Presently, two candidate vaccines viz. Helicobacter pylori surfome antigens & Gastric Cancer vaccine are active against this high-priority clarithromycin-resistant H. pylori and are in pre-clinical development. These vaccines are likely to combat macrolide-resistant H. pylori infection and may aid in the improvement of patient outcomes and prevention of hospitalizations (who. int) [25]. In a recent study, the efficiency of clarithromycin nanocrystals towards H. pylori has also been investigated and was found to be effective [26]. The nanocrystals were identified to increase the bioavailability of the drug at the specific site of action compared to other forms of antibiotics like powder [27]. Another study revealed enhanced activity of azithromycin nanoparticles (antibiotic-loaded with poly lactide-co-glycolides) against several clinical pathogens such as E. coli, H. influenza, and S. aureus [28]. Colistin, the peptidal antibiotic has been identified to have synergy with macrolide antibiotics and this combination has been identified to be effective against pathogenic Gram-negative rods like A. baumannii, P. aeruginosa and K. pneumoniae [29]. However, the potentiation approach to augment the efficiency of antibiotics seems to be the most promising approach to combat AMR because of the broader coverage, cost-effectiveness and versatility of the approach.
3.1. Antibiotic potentiators and exploration of their diverse mechanisms of action
Antibiotic potentiators, also known as adjuvants, are active compounds that have no or little antibacterial activity, but when combined with antibiotics, they can expand the spectrum and enhance the activity of the existing antibiotic against pathogenic bacteria. This renders the resistant bacteria vulnerable to antibiotics again. The high frequency of MDR bacterial infections and the low pace of the discovery of new and effective antimicrobials threaten the future of our healthcare system. So, the best strategy is to increase the efficiency and potentiate the effectiveness of the existing obsolete drugs and make them more useful, which is also economical. Recently there has been a surge of studies focussing on screening and development of compounds that have antibiotic potentiation ability. Many existing drugs which increase the permeability of the outer membrane and reduce the frequency of the spontaneous resistance for the partner antibiotic has been repurposed to check their potentiation activity [30,31,32].The major general mechanisms by which an antibiotic potentiator acts are i) inhibiting antibiotic efflux pump; efflux pump inhibitor (EPI) ii) by inhibiting the drug modifying enzyme, and iii) by increasing the membrane permeability thereby allowing the antibiotic penetration into the bacterial cells.
3.1.1. Efflux pump inhibitors
As efflux pumps are considered to be an expeditious and efficient resistant strategy in bacteria, the identification of efflux pump inhibitors (EPI) is a popular approach to inhibit drug expulsion from bacterial cells [33]. The efflux pumps present in the bacterial outer membrane expel the antibiotics limiting the concentration of antibiotics in the bacterial foci preventing its action at the target site. The EPI molecule can be used in combination with antibiotics to enhance their activity against bacterial efflux system and these inhibitors work by blocking the function of the drug efflux transporters which can be achieved by inhibiting the driving forces of the transporters or strong binding to efflux transporters themselves. The success rate of EPIs in potentiating macrolide antibiotics are limited, however, an earlier study has reported, the use of peptide nucleic acid (PNA) antisense agents to reduce the expression of RND efflux pumps in Campylobacter jejuni, sensitizing the strain to clinically important antibiotics ciprofloxacin and erythromycin [34]. Similarly, several pump inhibitors viz. capsaicin, homoisoflavonoid, peptide nucleic acids or phenylalanine-arginine β-naphthylamide targets different pumps like NorA, RND and CmeABC and promote the antimicrobial activity against resistant bacteria [35].
3.1.2. Modifying enzyme inhibitors
Multi-drug resistant bacteria produce a diverse range of enzymes which can degrade the antibiotics that were supposed to kill the pathogens. These cellular enzymes can change a drug by transferring the chemical moiety or hydrolyzing it, rendering it ineffective. Modifying enzyme inhibitors comprise a wide variety of chemical compounds that specifically target the bacterial enzymes responsible for the hydrolysis of antibiotics and hence increase the effectiveness of the co-administered antibiotic. β-lactamase inhibitors are the most significant and successful clinically used antibiotic adjuvants which prevent the bacterial degradation of β-lactam antibiotics by inhibiting β-lactamase enzymes [36]. They are also referred to as suicide inactivators of β-lactamase due to their irreversible mechanism of action by forming an irreversible acyl-enzyme complex by a covalent bond throughout the catalysis reaction with β-lactam. The most common β-lactamase inhibitors includes clavulanic acid (combined with amoxicillin; Co-amoxiclav), sulbactam (combined with ampicillin; ampicillin-sulbactam, tazobactam (combined with piperacillin; piperacillin-tazobactam), avibactam (ceftazidime-avibactam) and relebactam (imipenem-cilastatin-relebactam) [37]. Though there have been no effective macrolide phosphotransferase or macrolide esterases identified so far, screening of compounds that can inhibit these enzymes could aid in potentiating macrolide antibiotics.
3.1.3. Membrane permeabilizer
The use of membrane permeabilizers to increase antibiotic uptake has proven to be a successful strategy. Different kinds of compounds like peptides, nanoparticles and small molecules can act as outer membrane permeabilizer [29,38]. Due to the powerful activity of nanoparticles, they are emerging as warheads to counter and combat bacterial drug resistance. Some nanoparticles possess antibacterial activity, however, few interact with the antibiotic and enhance the antibacterial activity of the partner drugs thereby acting as a potentiator. Metal nanoparticles such as silver nanoparticles, and gold nanoparticles as well as several non-antibiotic nanomaterials like graphene oxide nanosheet (GO), graphene oxide-zinc oxide nanocomposite (GN/ZnO) and zinc oxide nanoparticles (ZnO) have been identified to have potentiating activity towards different antibiotics including macrolides [38,39].
4. Macrolide Potentiators & their current status
Antibiotic potentiation is the most promising of the numerous options outlined above to address the increasing problem of AMR. Re-purposing compounds having antibiotic potentiation activity is a less expensive and time-consuming alternative compared to developing new antibiotic scaffolds. Certain macrolide antibiotics such as clarithromycin and azithromycin are traditionally not used against Gram-negative pathogens due to their minimised outer-membrane lipopolysaccharide penetration, hence the efficacy and spectrum of such drugs can be improved by using these antibiotics with a molecule that can facilitate the passage of these antibiotics across the bacterial cell by permeabilizing the outer membrane. The quest for macrolide potentiators dates back to the early 1990s. The potentiating effect of normal human serum with macrolide antibiotics was examined in an investigation by Pruul & McDonald in 1992 [40]. The study revealed that the presence of 40% serum decreased the MIC of azithromycin by 26-fold for serum-resistant E. coli and 15-fold for S. aureus [40]. There have been several reviews in the past highlighting the usefulness of antibiotic adjuvants, different antibiotic adjuvants of antibiotics particularly of the β-lactam antibiotics [41]. However, there have been no or limited reviews on macrolide potentiators. In the following sections, the present review enlists the natural, synthetic, and peptide-based potentiators studied to date. The chemical structures of few important macrolide potentiators discussed in this review are presented in Figure 3, 1-10.
4.1. Natural potentiators
Plants have traditionally been a rich source of compounds, and several investigations have been done in search of natural compounds having antibiotic potentiation activity [42]. The continuous search for natural products led to the discovery of active extracts of Lycopus europaeus that can affect potentiate macrolide antibiotics. This plant is commonly known as Gipsywort, a perennial plant found on the banks of river and canals of the United Kingdom. The plant is also known to have anti-gonadotropic and anti-thyreotropic properties, due to the presence of phenolic chemicals in it. At 512µg/ml, two of its isopimarane diterpenes extracts, methyl-1a-acetoxy-7a-14a-dihydroxy-8,15-isopimaradien-18-oate (Figure 3: 1a) and methyl-1a,14a-diacetoxy-7a-hydroxy-8,15-isopimaradien-18-oate (Figure 3: 1b), showed potentiation activity of erythromycin by two-fold against macrolide-resistant S. aureus isolates expressing msr(A) multidrug efflux pump. At this concentration, these extracts showed no antibacterial activity [43]. Similarly, the chloroform extract of Rosmarinus officinalis L., generally known as rosemary, has also been identified to potentiate macrolide antibiotics against macrolide-resistant S. aureus strains decreasing its MIC to 16 µg/ml from 64µg/ml. Carnosic acid (Figure 3: 2) and Carnosol (Figure 3: 3), the two primary components of rosemary were examined for their potential to boost antibiotic action against resistant strains of S. aureus. Carnosic acid (2) potentiated the activity of erythromycin by eight-fold against msr(A) expressing S. aureus strains and the possible reason for its potentiating ability could be the inhibition of efflux pumps [44]. Natural EPI’s ability to block EmrD-3, a member of the Major facilitator superfamily (MFS) transporter family, has been studied in Vibrio cholerae. The EmrD-3 efflux inhibitory action of allyl sulphide, a bioactive component of Allium sativum, was determined and both A. sativum extract and allyl sulfide were discovered to reduce the MICs of several antimicrobials, including erythromycin, by fourfold in bacterial cells expressing EmrD-3 [45]. Further, Holarrhena antidysenterica, an ethnobotanical plant mainly used to treat bacterial infections, diarrhoea, dysentery, and fever, has also been assessed as a potential EPI. The potentiating ability of Conessine, its principal compound, has been determined with different antibiotics including erythromycin against wild-type P. aeruginosa PAO1 strain K767, MexAB-OprM overexpressed strain K1455, and MexB deleted strain K1523. The overexpression of the MexAB-OprM efflux pump is known to confer resistance to erythromycin and deletion of mexB resulting in loss of MexAB-OprM affected the susceptibility to antibiotic erythromycin. Conessine significantly reduced the MIC of erythromycin by 4- to 8-fold in contrast to the wild-type strain [46].
In another study including 29 plant species, Cytisus striatus showed potential potentiating activity of macrolide antibiotics. NMR-based metabolomics investigation was done to further analyze the chemicals that potentially function as antibiotic potentiators and isoflavonoids present in Cytisus striatus was found to enhance the antimicrobial effects of erythromycin against MRSA strains. The 22 isoflavonoids identified were evaluated for their potential as antibiotic adjuvants using the structure-activity relationship (SAR) out of which Genistein (Figure 3: 4a), Biochanin A (Figure 3: 4b), and Tectorigenin (Figure 3: 4c) was identified to potentiate erythromycin activity by reducing MIC by 2-to 8-fold against MRSA strains. This research indicates a clear synergy between isoflavonoids and erythromycin, indicating their considerable capacity for being used in the treatment of antibiotic-resistant bacterial infections like MRSA, however, the mechanistic approach is lacking [47].
A study examined the potentiation ability of a wide variety of small molecules from the library based on nitrogen-dense marine alkaloid scaffolds combined with macrolide antibiotics azithromycin, erythromycin, and clarithromycin against A. baumannii AB5075 isolate. The research led to the discovery of two compounds of marine alkaloid scaffolds, (Fig. 5a and 5b), that had previously shown potentiation of β-lactam antibiotics against drug-resistant A. baumannii and P. aeruginosa (Figure 3). The MIC of most of the macrolide antibiotics in combination with these compounds individually were found to be drastically reduced. The MIC of erythromycin reduced from 32μg/mL to 4μg/mL, the effective concentration of azithromycin reduced from 64μg/mL to 8μg/mL, and clarithromycin from 32 to 0.25μg/mL. The efficiency of this potentiation strategy for boosting clarithromycin activity has also been investigated in vivo using an A. baumannii 5 infection model of Galleria mellonella and a high survival rate of the host was observed after a single dosage of compound 5a and clarithromycin combination. On analyzing the mode of action, it was identified that the compounds 5a and 5b do not disrupt efflux pumps or increase cell membrane permeability by physical disruption, instead, they affect LPS production and its structure by reducing the hydroxylation of C14 and promote lipid A palmitoylation at Cl6 according to GC analysis, further examination of the LPS composition by gel electrophoresis reveals a structural change in the LPS treated by 5a [48].
Structure-activity relationship (SAR) analysis on the structural alterations produced by the compounds 5a and 5b to improve the activity of macrolide antibiotics was also conducted to establish the target for these molecules. Further modifications to the core phenyl ring of the compounds resulted in the augmentation of its potentiation activity of clarithromycin by 64- and 32-fold against the A. baumannii isolate AB5075. The minimum concentration of the lead compounds for its activity was only 10 and 7.5μM, respectively, while the required concentration of the original lead compounds was 30μM. Further modification of the amide linker of the compounds resulted in the development of two new adjuvants consisting of urea that inhibited resistance to clarithromycin at 7.5μM concentration in AB5075. The MIC of clarithromycin was reduced 64- and 128-fold [49]. The general mechanism of macrolide potentiators is diagrammatically represented in Figure 4.
4.2. Antimicrobial peptides as macrolide potentiators
Antimicrobial peptides (AMP) are very specific, fast-acting proteinaceous short peptide molecules, having 12-50 amino acid residues, which are known to have antimicrobial, anti-fungal, anti-parasitic, and anti-viral properties. These molecules act by disrupting both the outer and inner membrane of the bacterial cell and also display reduced cytotoxicity against the mammalian cell membrane. The emergence of resistant microbes, and the growing public concern about antibiotic use, prompted researchers to investigate the ability of AMPs to potentiate antibiotics and thereby re-sensitize MDR pathogens [50]. Peptides and peptidomimetics have displayed their action as potentiators of antibiotics targeting Gram-negative organisms. While most of them act by disrupting the OM, few of them reducing the efflux pump activity [29]. A hand full of antimicrobial peptides has been identified to potentiate macrolide antibiotics and are in different phases of clinical trials which need further research to understand the efficacy and mechanism of action [29,51]. She and her team recently explored the strong synergistic antimicrobial activity of the SPR741 molecule used in a triple combination with clarithromycin and erythromycin. The triple medication combination demonstrated significant efficacy against extremely drug-resistant K. pneumoniae and also showed low toxicity in vivo in a neutropenic mouse thigh infection model. The combination also successfully destroyed extremely resistant bacterial biofilms and persister cells in vitro [52].
Earlier, cathelicidin LL-37, a human AMP showed improved in vitro antimicrobial efficacy in vitro with daptomycin, a cationic antibiotic. It eradicated β-lactam and Vancomycin-resistant Enterococcus (VRE) and MRSA in patients [53]. A similar study was performed to validate the efficacy of cathelicidin LL-37 in potentiating azithromycin antibiotics against MDR Gram-negative rods [54]. In combination with cathelicidin LL-37, azithromycin had a strong bactericidal effect against MDR carbapenem-resistant isolates of Klebsiella pneumoniae, P. aeruginosa, and A. baumannii. This activity was attributed to the increased penetration of azithromycin when used with peptide LL-37. This implies that azithromycin, which is currently underutilized as the therapy option, can help patients with MDR bacterial infections, mainly when used in conjunction with other molecules. Although earlier studies have shown synergistic antibiotic-peptide interactions, the therapeutic implications of such results have rarely been investigated in depth.
In yet another study, the screening of thirty-four distinct peptides and four different antibiotics, including erythromycin, against E. coli ATCC 25922, four peptides that potentiated erythromycin was discovered. In MDR K. pneumoniae ST258 and E. coli ST131 the discovered peptides synergized with azithromycin and potentiated clindamycin. The low cytotoxicity of these two peptides such as (KLWKKWKKWLK-NH2 and GKWKKILGKLIR-NH2) toward eukaryotic cells (IC50 >50µM) led to the development and testing of all its D-analogues (D1 and D2). The growth of clinically important K. pneumoniae, E. coli, and A. baumannii strains used in the study were suppressed by lower concentrations of analogues D1 and D2 in conjunction with antibiotic azithromycin. The findings show that combinatorial screening at reduced peptide doses is an effective method for identifying therapeutically important peptide–antibiotic combinations. In vivo pharmacodynamic/pharmacokinetic as well as toxicity investigations are needed to verify the use of the peptides found in these studies with azithromycin antibiotics [55].
Another method is peptidomimetics, which is still in its early phases of development. Peptidomimetics are compounds whose pharmacophores mimic a natural peptide or protein while still interacting with the biological target and delivering the same biological effect. Peptidomimetics were created to solve peptides' shortcomings, as they are designed to have metabolic stability, high bioavailability, and high receptor affinity and selectivity [56]. A research showed that a peptidomimetic H-[NLys-tBuAla]6-NH2 (Figure 3: 6) (CEP-136; NLys = N-(4-aminobutyl) glycine; tBuAla = tert-butylalanine) when tested alone has low antimicrobial activity against a range of clinically important MDR strains including ESBL-producing isolates. The peptidomimetic reduced the needed effective bactericidal concentration of different macrolides (azithromycin or clarithromycin) to less than 1μg/mL in combination and this mechanistic insight concludes that the permeabilization of the outer membrane of the Gram-negative bacteria is a strong mechanism to potentiate existing antibiotics including macrolides. Another peptidomimetic, CEP-136 (Figure 3: 6) displayed low hemolytic activity with no substantial toxicity against mammalian HepG2 cells and also did not cause detrimental membrane disruption. In vivo confirmation of CEP-136's potentiation impact on the treatment of azithromycin in a mouse peritonitis model was possible due to less acute toxicity in healthy mice [29]. In another study, the pairwise screening of 42 synthetic peptidomimetics against MDR strain of E. coli ST131 and K. pneumoniae ST258 with the antibiotic azithromycin revealed two α-peptide/β-peptoid hybrids subclasses with fractional inhibitory concentration (FIC) indexes ranging from 0.03 to 0.38. Also, peptidomimetics that augmented erythromycin activity against E. coli and clindamycin activity against K. pneumoniae were discovered in the same screening with additional antibiotics. Six peptidomimetics were tested against P. aeruginosa, and five of these demonstrated antibiotic synergy. H-(Lys-NPhe)8-NH2, (Figure 3: 7) another promising peptidomimetic molecule, had only a minimal effect on mammalian cell viability and hence displayed the maximum selectivity. At sub-micromolar concentrations of 0.25–0.5μM, this chemical synergizes with azithromycin, generating sensitivity to the antibiotics at clinically suitable doses in MDR isolates. The identified peptidomimetic molecule and its analogues are attractive prospects for potentiating azithromycin against Gram-negative pathogens [57].
In another study, peptidomimetic-macrolide synergy has been analyzed by checkerboard assay in macrolide-resistant clinical strains of E. coli. The three peptidomimetics analyzed exhibited synergistic interactions with erythromycin, azithromycin, and tilmicosin against E. coli. With exposure to modest concentrations of peptidomimetic (0.5 to 8 mg/mL), the MICs of the three macrolides against these pathogens dropped by 4 to 32-fold. PEP-187 (Figure 3: 8a) showed faintly larger macrolide potentiation effects than PEP-387 (Figure 3: 8b) and CEP-136 among the three peptidomimetics examined.

Table 3.
Natural, synthetic and peptide-based compounds potentiating macrolide antibiotics.
| Sl. No. | Compound | Source | Antibiotic in combination | Organism tested | Current status | References |
|---|---|---|---|---|---|---|
| 1 |
1a: Methyl-1a-acetoxy-7a-14a-dihydroxy-8,15-isopimaradien-18-oate 1b: Methyl-1a,14a-diacetoxy-7a-hydroxy-8,15-isopimaradien-18-oate |
Natural- L. europaeus |
Erythromycin | S. aureus isolates expressing msr(A) multidrug efflux pump |
Lab study- in vitro |
Gibbons et al., 2003 |
| 2 |
2: Carnosic acid 3: Carnosol |
Natural - Rosmarinus officinalis L | Erythromycin | msr(A) & NorA expressing S. aureus strain |
Lab study- in vitro |
Oluwatuyi et al., 2004 |
| 3 | Allyl sulphide | Natural - Allium sativum | Erythromycin | EmrD-3 expressing V. cholerae |
Lab study- in vitro |
Bruns et al., 2017 |
| 4 | Conessine | Natural - Holarrhena antidysenterica | Erythromycin | P. aeruginosa PAO1 strain K767, MexAB-OprM overexpressed strain K1455, & MexB deleted strain K1523 |
Lab study- in vitro |
Siriyong et al., 2017 |
| 5 |
4a: Genistein, 4b: Biochanin A, 4c: Tectorigenin |
Cytisus striatus | Erythromycin | MRSA strains |
Lab study- in vitro |
Abreu et al., 2017 |
| 6 | Compound 5a & 5b | Nitrogen dense marine alkaloid scaffolds | Azithromycin, Erythromycin, Clarithromycin | A. baumannii AB5075 | Lab study - in vivo using a AB5075 infection model of Galleria mellonella | Martin et al., 2019 |
| 7 |
6: CEP-136 H-[NLys-tBuAla] 6-NH2 |
Peptide-based | Azithromycin, Clarithromycin | MDR strains including ESBL-producing isolates | Lab study - in vivo in mouse peritonitis model | Mood et al., 2021 |
| 8 | KLWKKWKKWLK-NH2 & GKWKKILGKLIR-NH2 | Peptide-based | Azithromycin, Erythromycin, Clarithromycin | K. pneumoniae, E. coli, & A. baumannii strains |
Lab study- in vitro |
Baker et al., 2019b |
| 9 | 7: H-(Lys-NPhe)8-NH2 | Peptide-based | Azithromycin, Erythromycin, Clindamycin | MDR strain of E. coli ST131 & K. pneumoniae ST258 |
Lab study- in vitro |
Baker et al., 2019a |
| 10 | 8: 4-isopentyloxy-2-naphthamide | Synthetic-2-naphthamide core | Erythromycin | AcrAB-TolC efflux pump expressing strains |
Lab study- in vitro |
Wang et al., 2017 |
| 11 |
9a: SLUPP-225 9b: SLUPP-417 |
Synthetic | Erythromycin | E. coli |
Lab study- in vitro |
Haynes et al., 2017 |
| 12 | 10: Bis-2-aminoimidazoles (bis-2-AIs) | Synthetic- nitrogen-dense heterocycles | Azithromycin, Clarithromycin | P. aeruginosa |
Lab study- in vivo |
Hubble et al., 2019 |
Abbreviations:MRSA: Methicillin-resistant Staphylococcus aureus, ST: Sequence type, MDR: Multi-drug resistant, ESBL: Extended-spectrum β-lactamase.
4.3. Synthetic potentiators
The ability of synthetic compounds to potentiate macrolide antibiotics has also been assessed and are discussed in this section. In E. coli, the AcrAB-TolC tripartite pump is the primary efflux pump that contributes to resistance. The pump comprises three different proteins, the first one is from the RND superfamily of proteins, the AcrB transporter which contributes to efflux substrate binding. The second protein is Porin TolC, the channel which bridges the outer membrane and permits substrates to move from the cell into the extracellular space, the third is AcrA, a membrane fusion protein (MFP) that provides a link among them to create a complete channel that precludes the periplasm. AcrB, a substrate-binding protein, has been studied extensively as a potential target for the development of new EPIs [58]. In the quest for AcrB EPI, a molecule viz. 2-naphthamide was synthesized. Conventional checkerboard assays revealed that the compound could reverse resistance to antibiotics imparted by the AcrAB-TolC efflux pump. The compound 4-isopentyloxy-2-naphthamide (Figure 3: 9) was identified to be the most efficient in lowering the erythromycin MIC in sensitive bacterial strains lacking efflux pumps [59]. Another study used in-silico virtual screening and experimental screening to find AcrA protein inhibitors. The study resulted in the discovery of two chemicals, SLUPP-225 (Figure 3: 10a) and SLUPP-417 (Figure 3: 10b), (Fig. 3) which showed promising EPI in E. coli cells. The compounds identified could traverse the outer membrane, improve efflux suppression, and augment erythromycin activity [58]. P. aeruginosa infection is a leading cause of death in cystic fibrosis (CF) patients, and antibiotic therapy remains the mainstay for the treatment of CF caused by P. aeruginosa. However, the growing AMR has limited the treatment efficacies and has rendered most antibiotics ineffective. A team of researchers examined a nitrogen-dense heterocycle compound library in search of potentiator compounds that would re-sensitize MDR P. aeruginosa. The study resulted in the identification of non-microbicidal bis-2-amino imidazoles (bis-2-AIs) 11a and 11b (Figure 3) which could augment the action of azithromycin against P. aeruginosa PAO1, a highly inherently resistant strain and other pseudomonal strains from CF patients [60].
Bis-2-AI 11b identified to be the most effective adjuvant in SAR experiments, increasing the activity of azithromycin by 1024 factor against PAO1 while presenting no evidence of hemolytic activity far above the effective dosage. Bis-2-AI 11b made the PAO1 strain sensitive to a variety of antimicrobials, particularly macrolide antibiotic (clarithromycin), doripenem, and rifampin, though it did not potentiate the membrane-active antibiotic colistin. The impact on membrane stability could be the mechanism of action as determined by BacLight assay; however, further mechanistic experiments for compound 11b are recently underway to determine if the observed effect is because of the direct interplay with cell membrane or an indirect impact because of the impaired cell membrane synthesis. Also, iron (Fe) and manganese (Mn) ions were identified to effect the efficacy of the compound, though their exact mode of action was not identified. Additionally, bis-2-AI 11b was effective to augment the activity of azithromycin in worm model without displaying any toxic or microbicidal activity. the survival rate of worms infected with PAO1 was identified to be 43%, similar to that seen with the positive control penicillin and clavulanic acid.
Apart from this, the recent study by Cui et. al have highlighted the potential of phentolamine as a potentiator of macrolide antibiotic. Phentolamine combined with the macrolide antibiotics erythromycin, clarithromycin, and azithromycin indicated a synergistic action against many Gram-negative isolates such as E. coli, S. typhimurium, K. pneumoniae, and A. baumannii . The fractional concentration inhibitory indices (FICI) of 0.375 and 0.5 indicated a synergic effect that was consistent with kinetic time–kill assays and the strong synergism has also been observed in the Galleria mellonella model [61].
5. Hindrances in taking macrolide potentiators from bench to bedside
5.1. Lack of comprehensive research and toxicity studies
The adjuvant strategy has the potential to overcome macrolide resistance by re-sensitizing MDR Gram-negative and Gram-positive bacteria, including species from the health-threatening ESKAPE category. However, number of studies on macrolide potentiators, is very limited. One major reason for this could be the toxicity of macrolide potentiator molecules identified [62,63]. Like the macrolide antibiotic itself, macrolide potentiators, particularly those targeting the efflux pumps, pose significant toxicity concerns which is a major roadblock for taking such molecules to clinical use. Additionally, because of the lack of definite MIC breakpoints for macrolide antibiotics against different pathogens, many macrolide potentiator compounds are used at high concentrations along with the high dosage of the antibiotic itself to completely inhibit bacterial growth. Another major issue of the macrolide-potentiating molecules identified so far is its acid instability [64]. This makes them suboptimal for oral administration, which further prompts the use of high concentrations of the compound.
5.2. Sensitivity and Specificity
Though there is no high homology between the macrolide resistance enzymes of different bacterial pathogens and human kinases, molecules that act via the resistance enzymes may follow a pathway similar to human serine/threonine (Ser/Thr) or tyrosine (Tyr) kinases and thus show non-specific inhibition of human targets. Thus, the cross-reactivity of macrolide phosphotransferases and macrolide esterases inhibitors with human enzymes represent an obstacle to their use as macrolide adjuvants to rescue macrolide resistance. However, no potential macrolide phosphotransferases and macrolide esterases inhibitors has been identified so far and no in-depth study regarding the structural homology between the bacterial macrolide resistance enzymes and human kinases is available, unlike the aminoglycoside resistance enzymes. Another aspect is the lack of complete understanding of macrolide resistance enzymes, as opposed to beta-lactamase enzymes.
5.3. Spectrum of activity
The outer membrane of Gram-negative bacteria, which serves as a permeability barrier, presents a challenge to the application of macrolide antibiotics itself. Hence, only those macrolide potentiators which can increase the penetration of erythromycin are effective against Gram-negative bacteria.
5.4. Macrolide potentiator resistance
Another major challenge in this strategy of antibiotic potentiation is the ability of bacterial pathogens to also evolve resistance against the potentiator molecules. Bacterial pathogens are known to accumulate resistance in their macrolide resistance machineries which prevents macrolide potentiators that act via modification of these machineries ineffective.
6. Future perspective
The emergence of multi- and pan-drug resistant organisms, colloquially known as "superbugs," causing illnesses has become an extremely concerning situation worldwide. Furthermore, increased antibiotic use, particularly macrolides like erythromycin or azithromycin, to treat secondary bacterial pneumonia in COVID-19 patients has aggravated macrolide antimicrobial resistance in both Gram-negative and Gram-positive bacteria [65]. Several strategies such as anti-virulence strategy, quorum inhibitors, phage cocktails, and CRISPR-Cas technology are being deployed in combination with antibiotics to combat the macrolide resistance threat. However, no such strategy has been identified to be the ultimate word to combat the growing AMR. In the past years, there have been a surge in screening and identification of molecules and compounds that in combination with the obsolete drugs could revive its lost activity. These studies could identify some very potential antibiotic adjuvants that could potentiate the antibiotic and re-sensitize MDR pathogens. Such studies were also extended for reviving macrolide antibiotics which identified several potential compounds. Despite the above-mentioned obstacles with potentiators, the scientific community is still working hard to discover more potential macrolide adjuvants. Macrolides are the primary option of medication for M. avium, H. pylori, Chancroid, and Diphtheria. Treatment of atypical and intracellular pathogenic infections like Chlamydia and Legionella spp. in parenchymal lower respiratory tract infections or nongonococcal urethritis are among the principal uses for macrolides. The safety of macrolides in beta-lactam-allergic individuals, oral bioavailability, and acceptability when administered during pregnancy, and efficacy in paediatric settings as well as in adult and geriatric patients are additional important benefits of macrolides compared to other drugs. Hence it is very important to maintain the efficacy of this drug class.
Till now, no macrolide potentiator has reached clinical trials though several natural and synthetic compounds have shown significant macrolide potentiating activity in vitro. However, the efficacy of very few compounds has been even performed in vivo. The efficacy of CEP-136's with azithromycin and nitrogen dense marine alkaloid compound 5a with clarithromycin combination were checked in-vivo. Comprehensive preclinical studies and optimization via a medicinal chemistry approach is yet to be done for these molecules to be further taken for clinical trials. Several peptide molecules has also shown significant macrolide potentiating activity. Despite this, their clinical usefulness is limited because of undesirable pharmacological characteristics. It is critical to understand the protein homology between bacterial-resistant enzymes and eukaryotic enzymes for better designing of macrolide adjuvants without adverse pharmacological properties. Structure-guided optimization of resistance enzyme inhibitors would be an effective technique to improve the efficacy and reduce off-target effects of compounds and small molecules, paving way for identification of macrolide resistance enzyme inhibitors and their human use. Peptidomimetics, allows the creation of molecules with finely tuned physicochemical features, resulting in increased antibacterial activity and a better pharmacological profile [66]. More studies into peptide-based potentiators could be beneficial. Multi-omics-assisted identification of resistance determinants and screening of small molecules that bind to these resistant determinants is a straightforward method for macrolide potentiator identification. Also, high throughput screening techniques such as artificial intelligence, machine learning could be deployed for the fast and less laborious identification of macrolide potentiators.
7. Concluding remarks
The rise in antimicrobial resistance is not only a medical crisis but also a scientific, economic, and interdisciplinary challenge, and it is very crucial to act before it’s too late. As AMR is almost becoming an insurmountable challenge, and antibiotics alone are not sufficient to fend off the implacably increasing tide of resistance, it is essential to weaponize the antibiotics and use them in medicine against pathogenic strains. The combinatorial activity of an antibiotic and its potentiator provides new insight into the therapeutic option for MDR clinical pathogens. Hence, there is a pressing need to carry out extensive research in the development of effective potentiators of macrolide antibiotics as the resistance against these antibiotics is growing dramatically among Gram-positive and Gram-negative pathogens. In the present review, the authors have recapitulated the different macrolide potentiators, their source of origin, the current clinical status of these molecules, challenges in macrolide potentiator development, and future prospects that can be undertaken for better screening and identification of macrolide potentiators. The search for macrolide inhibitors is a fascinating area of research which is currently expanding. It is not possible to develop a single compound capable of potentiating all classes of macrolides antibiotics due to their significant structural and mechanistic differences. The compounds discussed in the present review are futuristic and capable of potentiating different classes of macrolides antibiotics. The major fallback in the macrolide potentiator discovery research is the lack of in-depth knowledge of resistance mechanisms and systematic and extensive studies on the identified macrolide potentiators. Comprehensive mechanistic understanding of the mode of action of the macrolide potentiators discovered so far can shed light on its opportunities and shortcomings. Further research should be focused on these molecules and experiments to overcome their current limitations are warranted. More rigorous in-vitro and in-vivo studies to understand the safety and efficacy of the newly identified macrolide potentiating compounds have to be performed. Although the development of combinatorial therapy is complicated, considering the proven success of beta-lactam potentiators in clinics today, it can be speculated that the use of macrolide potentiators in clinical practice is also a possibility.
Author Contributions
All authors listed have made a substantial, and intellectual contribution to the work and approved it for publication.
Funding
This study received financial support from MK Bhan Fellowship Program 2020, under the Department of Biotechnology, Govt. of India.
Acknowledgments
Dr Deepjyoti Paul and Dr Lekshmi Narendrakumar MK Bhan Fellows, THSTI acknowledges the support of the Department of Biotechnology, Govt. of India and the MK Bhan fellowship program. Miss Meenal Chawla & Miss Taruna Ahrodia are thankful to CSIR & DBT, Govt. of India for the support. All the authors are thankful to the Executive Director, THSTI for the support and valuable suggestions.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Lenz KD, Klosterman KE, Mukundan H, et al. Macrolides: From Toxins to Therapeutics. Toxins 2021; 13. Available at:. [CrossRef]
- Manesh A, Varghese GM. Rising antimicrobial resistance: an evolving epidemic in a pandemic. The Lancet Microbe 2021; 2: e419–20. Available at:. [CrossRef]
- Knight GM, Glover RE, McQuaid CF, et al. Antimicrobial resistance and COVID-19: Intersections and implications. Elife 2021; 10. Available at:. [CrossRef]
- Sultana J, Cutroneo PM, Crisafulli S, Puglisi G, Caramori G, Trifiro G. Azithromycin in COVID-19 Patients: Pharmacological Mechanism, Clinical Evidence and Prescribing Guidelines. Drug Saf 2020; 43: 691–8. [CrossRef]
- Doan T, Worden L, Hinterwirth A, et al. Macrolide and Non-macrolide Resistance with Mass Azithromycin Distribution. N Engl J Med 2020; 383: 1941–50. [CrossRef]
- World Health Organization: WHO (2017). WHO publishes list of bacteria for which new antibiotics are urgently needed. World Health Organization: WHO. Available at: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed. /.
- Yu B, Roy Choudhury M, Yang X, et al. Restoring and Enhancing the Potency of Existing Antibiotics against Drug-Resistant Gram-Negative Bacteria through the Development of Potent Small-Molecule Adjuvants. ACS Infect Dis 2022; 8: 1491–508. [CrossRef]
- Narendrakumar L, Chakraborty M, Kumari S et al. β-Lactam potentiators to re-sensitize resistant pathogens: Discovery, development, clinical use and the way forward. Front Microbiol. 2023 Mar 10;13:1092556. [CrossRef]
- Chawla M, Verma J, Gupta R, et al. Antibiotic Potentiators Against Multidrug-Resistant Bacteria: Discovery, Development, and Clinical Relevance. Front Microbiol. 2022 Jul 1;13:887251. [CrossRef]
- Koya SF, Ganesh S, Selvaraj S, et al. Consumption of systemic antibiotics in India in 2019. The Lancet Regional Health - Southeast Asia; 2022; 4: 100025. [CrossRef]
- Hicks LA, Bartoces MG, Roberts RM, et al. US outpatient antibiotic prescribing variation according to geography, patient population, and provider speciality in 2011. Clin. Infect. Dis. 2015; 601308–1316. [CrossRef]
- Kumar M, Rao M, Mathur T, et al. Azithromycin Exhibits Activity Against Pseudomonas aeruginosa in Chronic Rat Lung Infection Model. Front. Microbiol. 2021; 12:603151. [CrossRef]
- Saini H, Chhibber S, Harjai K. Azithromycin and cipro- floxacin: a possible synergistic combination against Pseudomonas aeruginosa biofilm-associated urinary tract infections. Int J Antimicrob Agents 2015; 45:359–7. [CrossRef]
- Putnam SD, Castanheira M, Moet GJ, et al. CEM-101, a novel fluoroketolide: antimicrobial activity against a diverse collection of Gram-positive and Gram-negative bacteria. Diagn Microbiol Infect Dis 2010; 66:393–401.
- Gomes C, Martínez-Puchol S, Palma N, et al. Macrolide resistance mechanisms in Enterobacteriaceae: Focus on azithromycin. Crit Rev Microbiol. 2017; 43:1-30. [CrossRef]
- O’Brien KS, Emerson P, Hooper PJ et al. Antimicrobial resistance following mass azithromycin distribution for trachoma: a systematic review. Lancet Infect Dis 2019; 19: e14–25.
- Dinos, GP. The macrolide antibiotic renaissance. Br J Pharmacol 2017; 174: 2967-83. [CrossRef]
- Golkar T, Zielinski M, Berghuis AM. Look and Outlook on Enzyme-Mediated Macrolide Resistance. Front Microbiol 2018; 9: 1942.
- Gomes C, Martinez-Puchol S, Durand D, et al. Which mechanisms of azithromycin resistance are selected when efflux pumps are inhibited? Int J Antimicrob Agents 2013a; 4:307–11. [CrossRef]
- Gutierrez-Castrellon P, Mayorga-Buitron JL, Bosch-Canto V, et al. Efficacy and safety of clarithromycin in pediatric patients with upper respiratory infections: a systematic review with meta-analysis. Rev Invest Clin 2012; 64: 126–35.
- Bohnert JA, Schuster S, Feahnrich E, et al. Altered spectrum of multidrug resistance associated with a single point mutation in the Escherichia coli RND-type MDR efflux pump YhiV (MdtF). J Antimicrob Chemother 2007; 59:1216–22. [CrossRef]
- Pawlowski AC, Stogios PJ, Koteva K, et al. The evolution of substrate discrimination in macrolide antibiotic resistance enzymes. Nat Commun 2018; 9: 112. [CrossRef]
- Andersen JL, He GX, Kakarla P, et al. Multidrug efflux pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus bacterial food pathogens. Int J Environ Res Public Health. 2015 Jan 28;12(2):1487-547. [CrossRef]
- Xiang Y, Wu F, Chai Y, et al. A new plasmid carrying mphA causes prevalence of azithromycin resistance in enterotoxigenic Escherichia coli serogroup O6. BMC Microbiol. 2020 Aug 11;20(1):247. [CrossRef]
- Micoli F, Bagnoli F, Rappuoli R et al. The role of vaccines in combatting antimicrobial resistance. Nat Rev Microbiol. 2021;19:287-302.
- Eleraky NE, Allam A, Hassan SB, et al. Nanomedicine Fight against Antibacterial Resistance: An Overview of the Recent Pharmaceutical Innovations. Pharmaceutics 2020; 12. Available at:. [CrossRef]
- Morakul B, Suksiriworapong J, Chomnawang MT, et al. Dissolution enhancement and in vitro performance of clarithromycin nanocrystals produced by precipitation-lyophilization-homogenization method. Eur J Pharm Biopharm. 2014; 88:886-96. [CrossRef]
- Azhdarzadeh M, Lotfipour F, Zakeri-Milani P, et al. Anti-bacterial performance of azithromycin nanoparticles as colloidal drug delivery system against different Gram-negative and Gram-positive bacteria. Adv Pharm Bull 2012; 2: 17–24.
- Mood EH, Goltermann L, Brolin C, et al. Antibiotic Potentiation in Multidrug-Resistant Gram-Negative Pathogenic Bacteria by a Synthetic Peptidomimetic. ACS Infect Dis 2021; 7: 2152–63. [CrossRef]
- Hyun S, Choi Y, Jo D, et al. Proline Hinged Amphipathic α-Helical Peptide Sensitizes Gram-Negative Bacteria to Various Gram-Positive Antibiotics. J Med Chem 2020; 63: 14937–50. [CrossRef]
- MacNair CR, Brown ED. Outer Membrane Disruption Overcomes Intrinsic, Acquired, and Spontaneous Antibiotic Resistance. MBio 2020; 11. Available at:. [CrossRef]
- Tyers M, Wright GD. Drug combinations: a strategy to extend the life of antibiotics in the 21st century. Nat Rev Microbiol. 2019;17:141-155. [CrossRef]
- Du D, Wang-Kan X, Neuberger A, et al. Author Correction: Multidrug efflux pumps: structure, function and regulation. Nat Rev Microbiol 2018; 16: 577.
- Jeon B, Zhang Q. Sensitization of Campylobacter jejuni to fluoroquinolone and macrolide antibiotics by antisense inhibition of the CmeABC multidrug efflux transporter. J Antimicrob Chemother 2009; 63: 946–8.
- Gill EE, Franco OL, Hancock RE. Antibiotic adjuvants: diverse strategies for controlling drug-resistant pathogens. Chem Biol Drug Des. 2015;85:56-78. [CrossRef]
- 36. Mojica MF, Rossi MA, Vila AJ, Bonomo RA. The urgent need for metallo-β-lactamase inhibitors: an unattended global threat. Lancet Infect Dis 2022; 22: e28–34. [CrossRef]
- 37. Laws M, Shaaban A, Rahman KM. Antibiotic resistance breakers: current approaches and future directions. FEMS Microbiol Rev 2019; 43: 490–516. [CrossRef]
- Klobucar K, Brown ED. New potentiators of ineffective antibiotics: Targeting the Gram-negative outer membrane to overcome intrinsic resistance. Curr Opin Chem Biol 2022; 66: 102099. [CrossRef]
- Pandey P, Sahoo R, Singh K, et al. Drug Resistance Reversal Potential of Nanoparticles/Nanocomposites via Antibiotic’s Potentiation in Multi Drug Resistant P. aeruginosa. Nanomaterials 2021; 12: 117. Available at:. [CrossRef]
- . Pruul H, McDonald PJ. Potentiation of antibacterial activity of azithromycin and other macrolides by normal human serum. Antimicrob Agents Chemother. 1992; 36(1):10-6.
- Liu Y, Li R, Xiao X, Wang Z. Antibiotic adjuvants: an alternative approach to overcome multi-drug resistant Gram-negative bacteria. Crit Rev Microbiol 2019; 45: 301–14.
- Khameneh B, Iranshahy M, Soheili V, et al. Review on plant antimicrobials: a mechanistic viewpoint. Antimicrob Resist Infect Control 2019; 8: 118. [CrossRef]
- Gibbons S, Oluwatuyi M, Veitch NC, Gray AI. Bacterial resistance modifying agents from Lycopus europaeus. Phytochemistry 2003; 62: 83–7. [CrossRef]
- Oluwatuyi M, Kaatz GW, Gibbons S. Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 2004; 65: 3249–54. [CrossRef]
- Bruns MM, Kakarla P, Floyd JT, et al. Modulation of the multidrug efflux pump EmrD-3 from Vibrio cholerae by Allium sativum extract and the bioactive agent allyl sulfide plus synergistic enhancement of antimicrobial susceptibility by A. sativum extract. Arch Microbiol 2017; 199: 1103–12.
- Siriyong T, Srimanote P, Chusri S, et al. Conessine as a novel inhibitor of multidrug efflux pump systems in Pseudomonas aeruginosa. BMC Complement Altern Med 2017; 17: 405. [CrossRef]
- Abreu AC, Coqueiro A, Sultan AR, et al. Looking to nature for a new concept in antimicrobial treatments: isoflavonoids from Cytisus striatus as antibiotic adjuvants against MRSA. Sci Rep 2017; 7: 3777.
- Martin SE, Melander RJ, Brackett CM, et al. Small Molecule Potentiation of Gram-Positive Selective Antibiotics against. ACS Infect Dis 2019; 5: 1223–30. [CrossRef]
- Hubble VB, Bartholomew KR, Weig AW, et al. Augmenting the Activity of Macrolide Adjuvants against. ACS Med Chem Lett 2020; 11: 1723–31.
- Huan Y, Kong Q, Mou H, Yi H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front Microbiol 2020; 11: 582779. [CrossRef]
- Costa F, Teixeira C, Gomes P, et al. Clinical Application of AMPs. Adv Exp Med Biol 2019; 1117: 281–98.
- She P, Liu Y, Xu L, et al. SPR741, Double- or Triple-Combined With Erythromycin and Clarithromycin, Combats Drug-Resistant, Its Biofilms, and Persister Cells. Front Cell Infect Microbiol 2022; 12: 858606.
- Sakoulas G, Okumura CY, Thienphrapa W, et al. Nafcillin enhances innate immune-mediated killing of methicillin-resistant Staphylococcus aureus. J Mol Med 2014; 92: 139–49. [CrossRef]
- Lin L, Nonejuie P, Munguia J, et al. Azithromycin Synergizes with Cationic Antimicrobial Peptides to Exert Bactericidal and Therapeutic Activity Against Highly Multidrug-Resistant Gram-Negative Bacterial Pathogens. EBioMedicine. 2015 Jun 10;2(7):690-8. [CrossRef]
- Baker KR, Jana B, Hansen AM, et al. Repurposing azithromycin and rifampicin against Gram-negative pathogens by combination with peptide potentiators. Int J Antimicrob Agents 2019; 53: 868–72. [CrossRef]
- Lenci E, Trabocchi A. Peptidomimetic toolbox for drug discovery. Chem Soc Rev 2020; 49: 3262–77.
- Baker KR, Jana B, Hansen AM, et al. Repurposing Azithromycin and Rifampicin Against Gram-Negative Pathogens by Combination With Peptidomimetics. Front Cell Infect Microbiol 2019; 9: 236. [CrossRef]
- Haynes KM, Abdali N, Jhawar V, et al. Identification and Structure-Activity Relationships of Novel Compounds that Potentiate the Activities of Antibiotics in Escherichia coli. J Med Chem 2017; 60: 6205–19. [CrossRef]
- Wang Y, Mowla R, Guo L, et al. Evaluation of a series of 2-napthamide derivatives as inhibitors of the drug efflux pump AcrB for the reversal of antimicrobial resistance. Bioorg Med Chem Lett 2017; 27: 733–9.
- Hubble VB, Hubbard BA, Minrovic BM, et al. Using Small-Molecule Adjuvants to Repurpose Azithromycin for Use against Pseudomonas aeruginosa. ACS Infect Dis 2019; 5: 141–51.
- Cui ZH, He HL, Zheng ZJ et al. Phentolamine Significantly Enhances Macrolide Antibiotic Antibacterial Activity against MDR Gram-Negative Bacteria. Antibiotics 2023, 12, 760. [Google Scholar] [CrossRef]
- Ma TKW, Chow KM, Choy ASM, et al. Clinical manifestation of macrolide antibiotic toxicity in CKD and dialysis patients. Clin Kidney J 2014; 7: 507–12.
- Woodhead JL, Yang K, Oldach D, et al. Analyzing the Mechanisms Behind Macrolide Antibiotic-Induced Liver Injury Using Quantitative Systems Toxicology Modeling. Pharm Res 2019; 36: 48. [CrossRef]
- Fohner AE, Sparreboom A, Altman RB, et al. Pharm GKB summary: Macrolide antibiotic pathway, pharmacokinetics/pharmacodynamics. Pharmaco genet Genomics. 2017; 27:164-167.
- Pani A, Lauriola M, Romandini A, et al. Macrolides and viral infections: focus on azithromycin in COVID-19 pathology. Int J Antimicrob Agents 2020; 56: 106053.
- Mendez-Samperio, P. Peptidomimetics as a new generation of antimicrobial agents: current progress. Infect Drug Resist 2014; 7: 229–37.
Figure 1.
Structure of 14, 15, & 16-membered macrolide antibiotics. 14-membered macrolide antibiotics: Erythromycin, Clarithromycin, Roxithromycin, Flurithromycin, Dirithromycin, Cethromycin, Telithromycin; 15-membered macrolide antibiotic: Azithromycin, 16-membered macrolide antibiotic: Tylosin, Josamycin.
Figure 1.
Structure of 14, 15, & 16-membered macrolide antibiotics. 14-membered macrolide antibiotics: Erythromycin, Clarithromycin, Roxithromycin, Flurithromycin, Dirithromycin, Cethromycin, Telithromycin; 15-membered macrolide antibiotic: Azithromycin, 16-membered macrolide antibiotic: Tylosin, Josamycin.
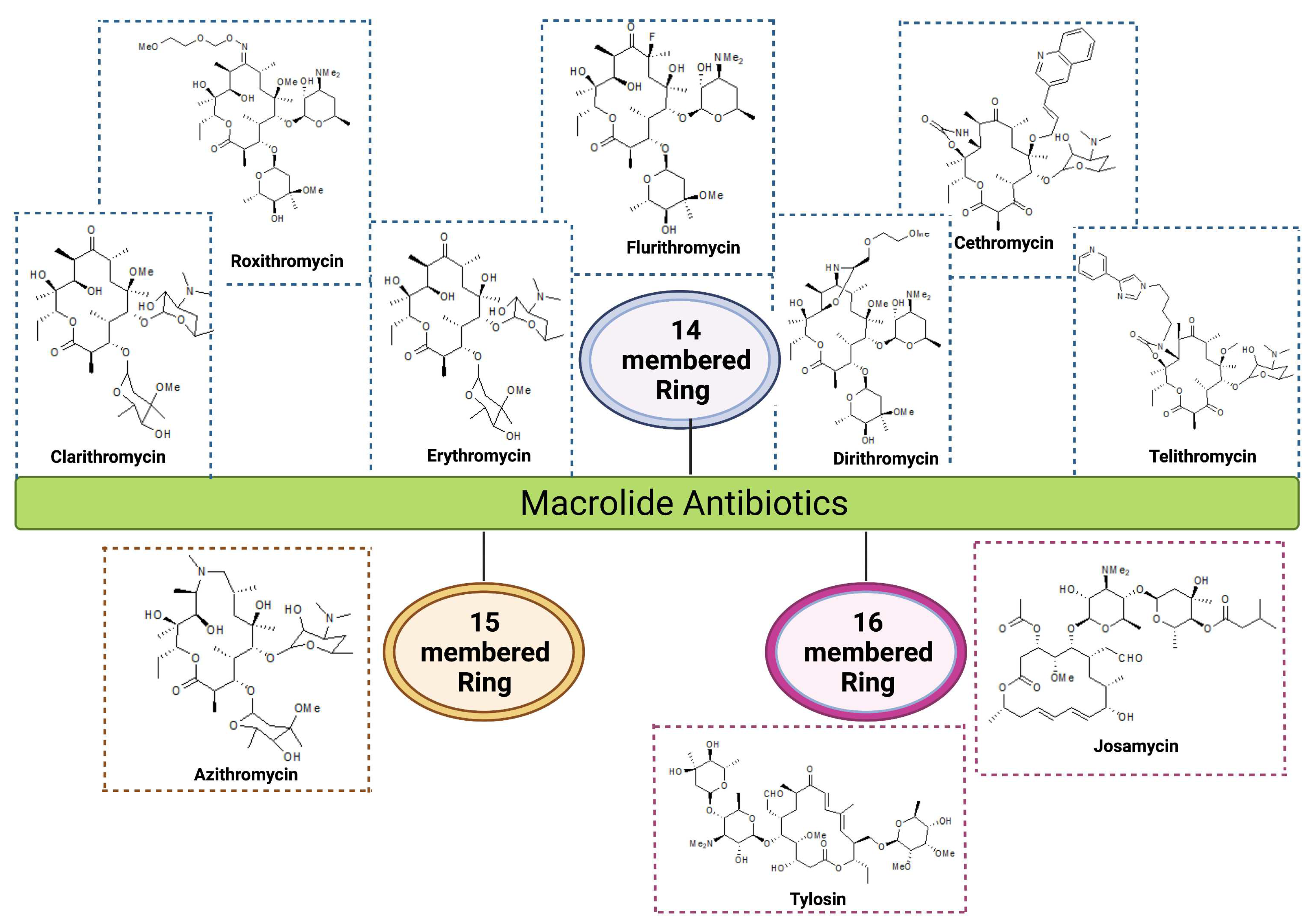
Figure 2.
Multiple strategies in fighting multi-drug resistant bacteria which might help in combating the worsening scenario of AMR.
Figure 2.
Multiple strategies in fighting multi-drug resistant bacteria which might help in combating the worsening scenario of AMR.
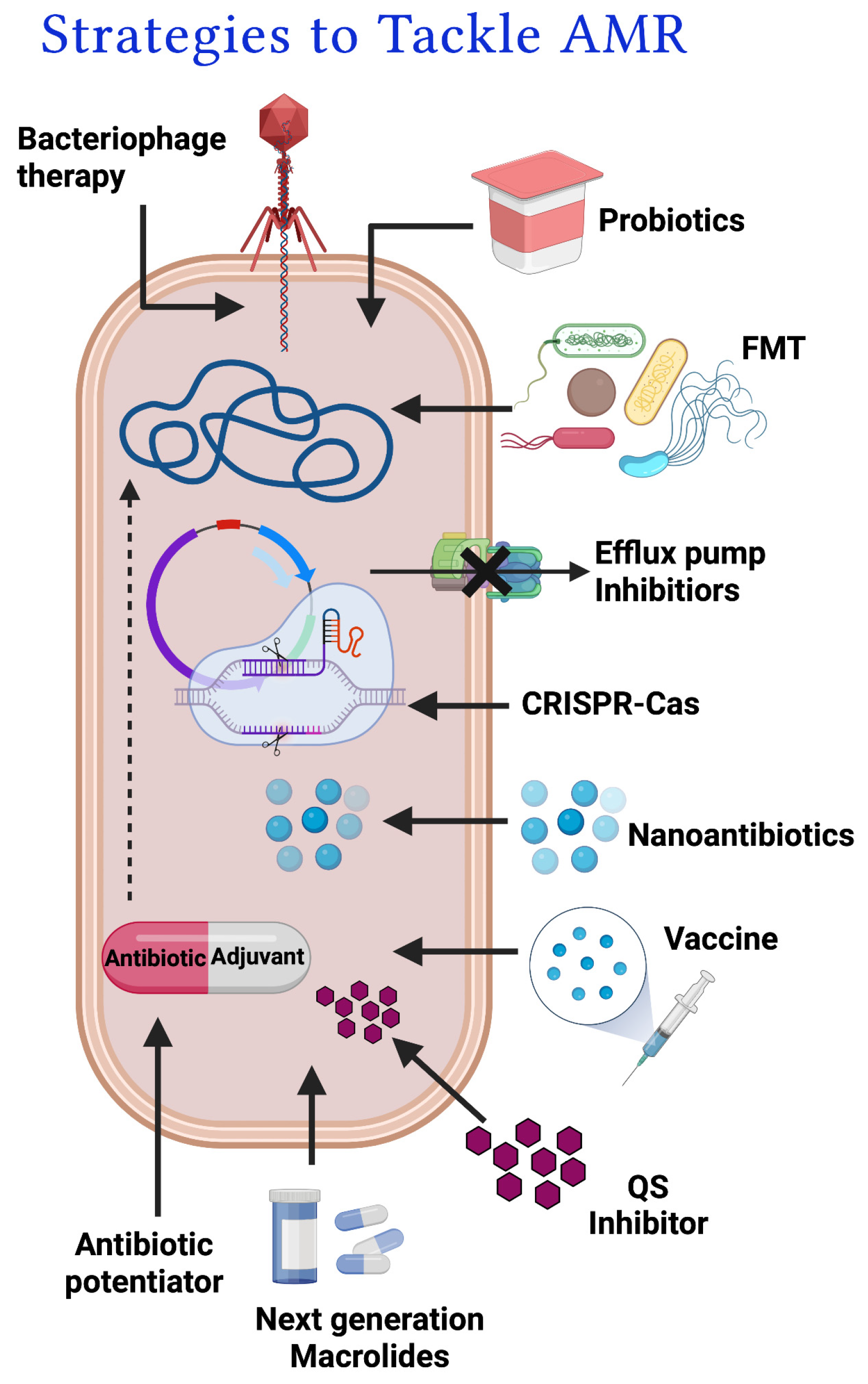
Figure 3.
Chemical structures of different macrolide potentiators. Macrolide potentiators are differentiated based on their source such as natural, peptide-based and synthetic. Natural potentiator includes 1) isopimarane diterpenes extracts, 2) Carnosic acid, 3) Carnosol, 4) Isoflavonoids, a) Genistein, b) Biochanin A, c) Tectorigenin, 5) Marine alkaloid scaffolds; antimicrobial peptides such as 6) CEP-136, 7) H-(Lys-NPhe)8-NH2, and synthetic potentiator comprises of 8) 4-isopentyloxy-2-naphthamide, 9a) SLUPP-225, 9b) SLUPP-417 10) bis-2- aminoimidazoles (bis-2-AIs).
Figure 3.
Chemical structures of different macrolide potentiators. Macrolide potentiators are differentiated based on their source such as natural, peptide-based and synthetic. Natural potentiator includes 1) isopimarane diterpenes extracts, 2) Carnosic acid, 3) Carnosol, 4) Isoflavonoids, a) Genistein, b) Biochanin A, c) Tectorigenin, 5) Marine alkaloid scaffolds; antimicrobial peptides such as 6) CEP-136, 7) H-(Lys-NPhe)8-NH2, and synthetic potentiator comprises of 8) 4-isopentyloxy-2-naphthamide, 9a) SLUPP-225, 9b) SLUPP-417 10) bis-2- aminoimidazoles (bis-2-AIs).
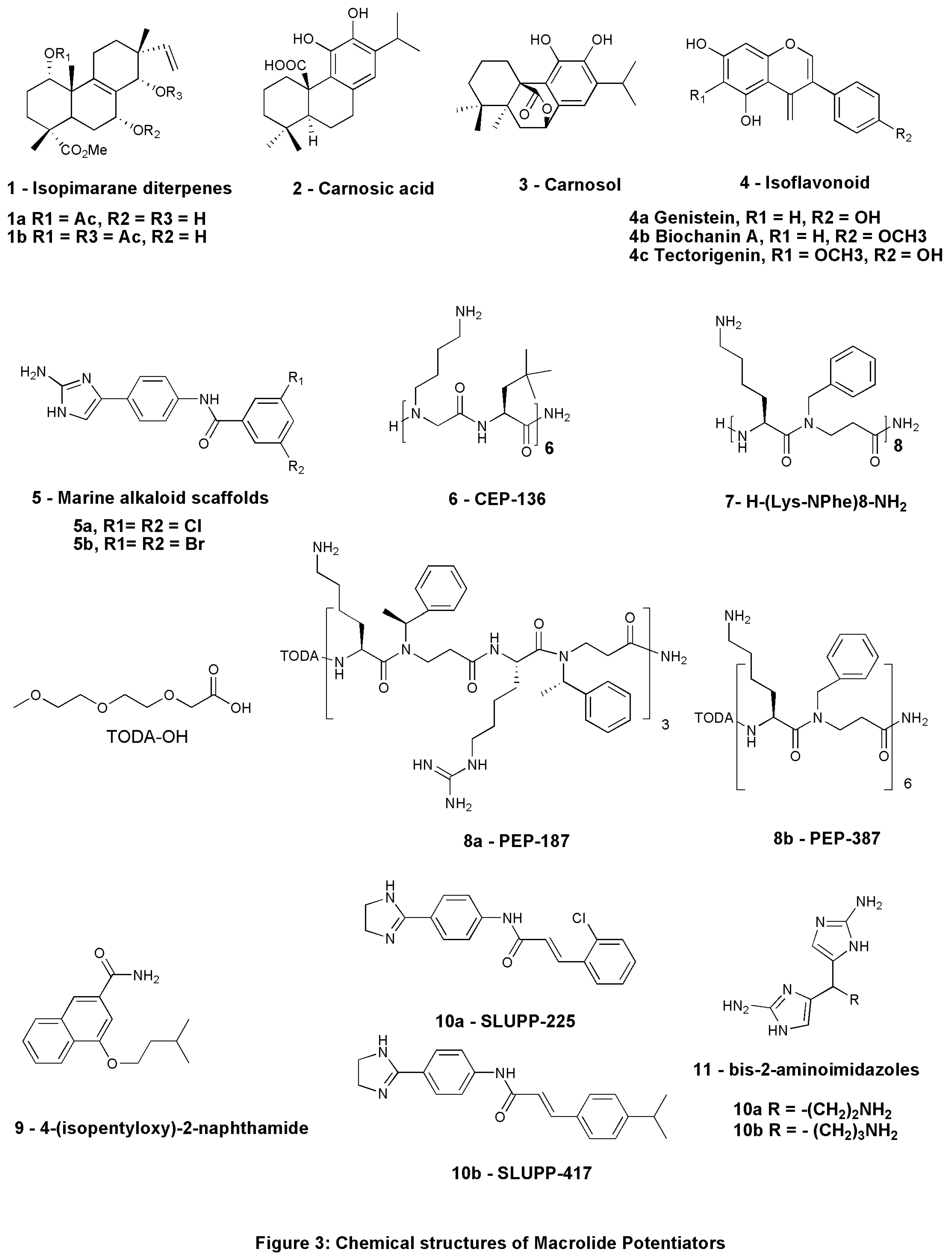
Figure 4.
Diagrammatic representation of the potentiation mechanism of macrolide antibiotics. This figure illustrates that macrolide antibiotic alone is unable to prevent the resistant bacteria, however, macrolide along with a potentiator molecule has the ability to enter into the bacterial cell and kill the pathogens. This figure also displays the different sources of macrolide potentiator molecules.
Figure 4.
Diagrammatic representation of the potentiation mechanism of macrolide antibiotics. This figure illustrates that macrolide antibiotic alone is unable to prevent the resistant bacteria, however, macrolide along with a potentiator molecule has the ability to enter into the bacterial cell and kill the pathogens. This figure also displays the different sources of macrolide potentiator molecules.
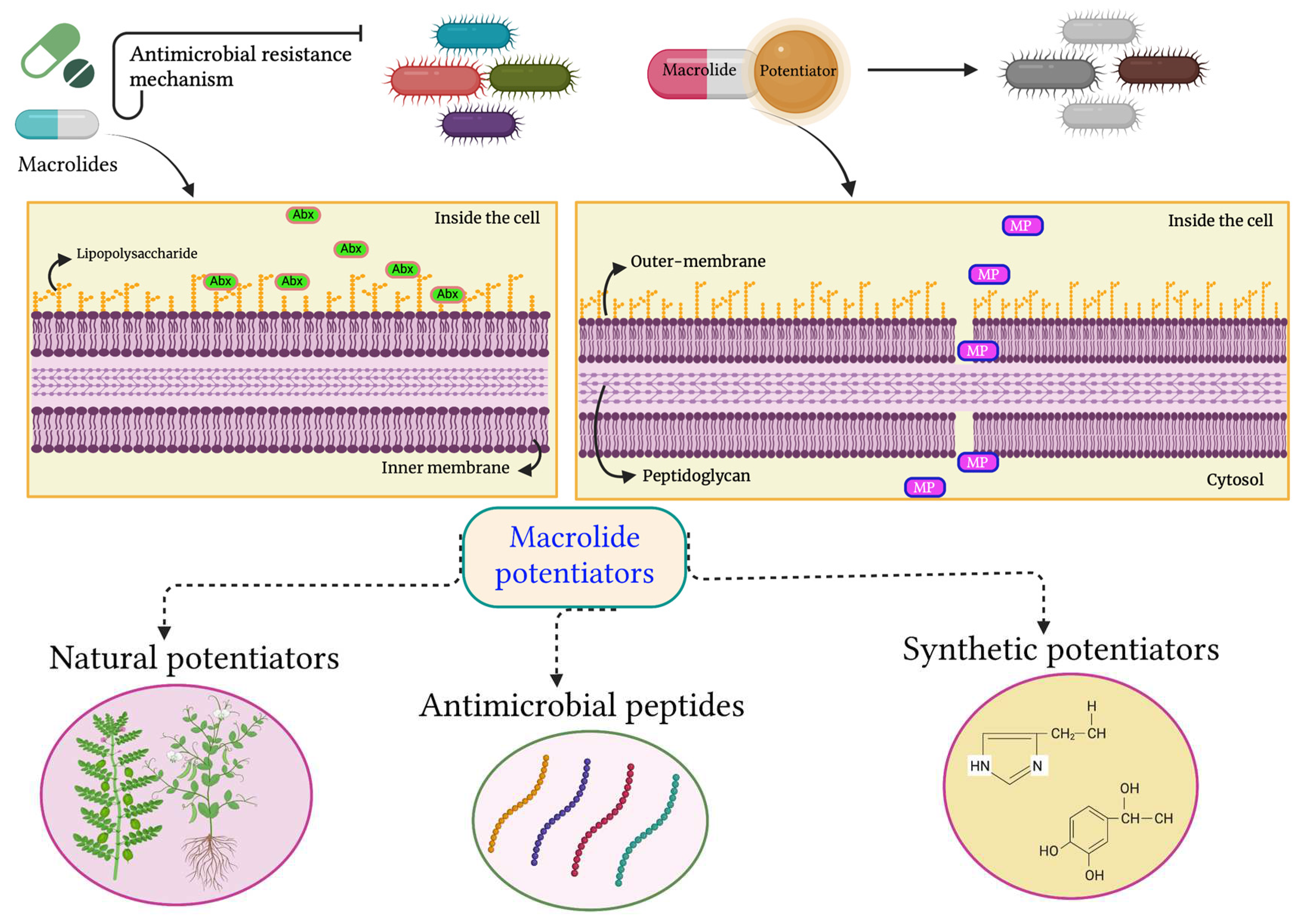
Table 1.
Spectrum of activity of different macrolide antibiotics.
| Group | Ring Structure | Molecule | Origin | Target pathogens | Treatment | Reference |
|---|---|---|---|---|---|---|
| First generation | 14-membered | Erythromycin | Streptomyces erythreus | Gram-positive bacteria: Staphylococcus aureus, Streptococcus pneumoniae, & S. pyogenes Gram-negative bacteria: Neisseria meningitis, N. gonorrhoeae, & Bordetella pertussis |
RTI, Skin, soft tissues, Urogenital tract and Middle ear infections |
Kaneko et al., 2007; Farzam et al., 2021 |
| Second generation | 14-membered | Clarithromycin |
Semi-synthetic conversion of erythromycin | Gram-positive bacteria: S. aureus, S. pneumoniae, & S. pyogenes Gram-negative bacteria : Mycoplasma pneumoniae, Legionella pneumophila, & Chlamydia pneumoniae, Helicobacter pylori, Pseudomonas aeroginosa |
RTI, Chronic inflammation of stomach ulcers, MAC infections in HIV patients |
Yanagihara et al., 2002; Lenz et al., 2021 |
| Roxithromycin | Semi-synthetic derivative of erythromycin | Gram-positive bacteria: S. aureus, S. pyogenes, S. pneumoniae, Listeria monocytogens Gram-negative bacteria: N. meningitidis, B. pertussis, Haemophilus influenzae |
RTI, skin and soft tissue infection and gastrointestinal infections |
Dinos et al., 2017 | ||
| Flurithromycin | Fluorinated derivative of erythromycin A |
H. pylori, Bacteroides forsythus |
Chronic gastritis, Periodontal disease | Fera et al., 2001; Blandino et al., 2013 |
||
| Dirithromycin | Semi-synthetic derivative of erythromycin | Gram-positive bacteria: S. aureus, S. pneumoniae, Gram-negative bacteria: H. influenzae, L. pneumophila, Moraxella catarrhalis, and M. pneumoniae |
Bronchitis, Pneumoniae, tonsillitis and skin infections | Dinos et al., 2017 | ||
| 15-membered | Azithromycin | Derivative of erythromycin | Gram-positive bacteria: S. aureus, S. pneumoniae Gram-negative bacteria: H. influenzae, M. catarrhalis, C. trachomatis, & H. pylori |
RTI, otitis media, skin and soft tissue infections, gastric and duodenal infections, trachoma eye infections & sexually transmitted diseases | Pawlowski et al., 2018 Leroy et al., 2021 |
|
| Third generation | 14-membered ketolides |
Telithromycin |
Semi-synthetic derivative of erythromycin | Gram-positive bacteria: S. pneumoniae Gram-negative bacteria: M. pneumoniae, C. pneumoniae, H. influenzae & L. pneumophilia |
Community-acquired respiratory tract infections |
Kaneko et al., 2007; Wolter et al 2008 |
| Cethromycin | Derivative of erythromycin | Gram-positive bacteria: Macrolide-resistant S. pneumoniae, S. pyogenes Gram-negative bacteria: H. influenzae |
Community acquired pneumonia | Mansour et al., 2016, Rafie et al., 2010 |
||
| 16-membered | Josamycin |
S. narbonensis var. josamyceticus | Gram-positive bacteria: S. aureus, S. pneumoniae, and S. pyogenes Gram-negative bacteria: H. influenzae, M. catarrhalis , M. genitalium, N. gonorrhea, N. meningitidis |
RTI, Urethritis |
Gupta et al., 2020 |
|
| Tylosin |
S. fradiae, H. influenzae |
H. influenzae, Gram-positive pathogens and mycoplasma | Respiratory diseases, mastitis, and dysentery in cattle and other farm animals |
Arsic et al., 2012 |
Abbreviations: MAC: Mycobacterium Avium Complex, HIV: Human Immunodeficiency Virus, RTI: Respiratory Tract Infection.
Table 2.
Mechanism of macrolide resistance and their association with mobile genetic elements.
| Mechanism of resistance | Target/ Enzymes |
Genes | Organisms | Location/Associated MGEs/ Co-resistance determinants | Accession number | References |
|---|---|---|---|---|---|---|
| Efflux pumps | Mef proteins | mef(A) | Streptococcus pyogenes, S. pneumoniae, Staphylococcus aureus, Enterococcus spp. | Plasmid, Tn1207.1, Tn1207.2 |
U70055, AF227520, AF227521 |
Miklasinska-Majdanik 2021; Dinos 2017; Roberts et al., 1999 |
| mef(B) | Escherichia coli | Plasmid, Class 1 integron, IS440 | FJ196385 | |||
| mef(C) | Photobacterium damselae | Plasmid, tet(M), flo(R), Tet(C), Tet(D) | AB571865 | |||
| mef(D) | S. aureus | Chromosome | LR130509 | |||
| mef(E) | S. pneumoniae | Plasmid, Chromosome | U83667, AF274302 | |||
| mef(F) | Macrococcus canis | Plasmid | CP046364 | |||
| mef(G) | S. mitis | Chromosome | HG423652 | |||
| mef(H) | Clostridioides difficile | Chromosome | MW269960.1 | |||
| mef(I) | S. pneumoniae | Chromosome, Tn916, tet(M) | AJ971089 | |||
| mef(J) | S. pyogenes | Chromosome | CP065927.1 | |||
| mef(O) | S. pyogenes | Chromosome | DQ016305 | |||
| Msr proteins | msr(A) | S. epidermidis, S. aureus | Plasmids | X52085 | Fyfe et al., 2016; Roberts et al., 1999; Schwendener et al., 2020 |
|
| msr(B) | S. xylosus | Plasmid | M81802.1 | |||
| msr(C) | E. faecium | Chromosome | AF313494 | |||
| msr(D) | S. pyogenes | Tn1207.2 | AF227521 | |||
| msr(E) | Pasteurella multocida, S. pneumoniae, Klebsiella pneumoniae | Chromosome, Plasmid, Tn1207.1, Sul1, blaKPC-2, blaDHA-1, qnrB4, & armA |
FR751518, FJ628167.2, AF227520 | |||
| msr(F) | Macrococcus canis | Chromosome | MN728681 | |||
| msr(G) | M. canis | Plasmid | CP046364 | |||
| msr(H) | M. canis | Chromosome | BK011995 | |||
| msr(I) | S. pyogenes | Chromosome, tet(M) | CP065927.1 | |||
| Enzymatic degradation | Phosphotransferases | mph(A) | E. coli, | Plasmid, IS26 | D16251 |
Pawlowski et al., 2018; Golkar et al., 2018 |
| mph(B) | E. coli, | Plasmid | D85892 | |||
| mph(C) | S. aureus | Plasmid, msrA | AF167161 | |||
| mph(D) | Pseudomonas aeruginosa | Chromosome | AB048591 | |||
| mph(E) |
P. multocida, K. pneumoniae, Serratia marcescens, E. coli,Acinetobacter baumannii,Citrobacter freundii |
Chromosome, plasmid, IS26, ISCR1, ISEc28, ISEc29, IS26, ISAba125, Tn5393, IS18, ISAba3, Class 1 integron, ISEcp1, msr(E) blaKPC-2, blaDHA-1, qnrB4, armA, blaOXA-58, ant3/, linF, sul1, blaCTX-M, aac3, dhfr, aadA2, qac |
FR751518, FJ628167.2 FJ917355.1 FJ187822.1 EU294228.1 AY522431.4 AF550415.2 |
|||
| mph(F) | Uncultured bacterium | Plasmid | AM260957 | |||
| mph(G) |
Photobacterium damselae |
Plasmid, floR, tet (B), tet(C) tet (D), tet (M) | AB571865 | |||
| mph(H) |
Brachybacterium faecium |
Chromosome | NC_013172.1 | |||
| mph(I) | Paenibacillus sp | Chromosome | KX531056.1 | |||
| mph(J) |
Brevibacillus brevis |
Chromosome | KY753883.1 | |||
| mph(K) | Bacillus subtilis | Chromosome | NC_000964.3 | |||
| mph(L) |
B. cereus |
Plasmid, FosB, TetV, | ACMJ01000036.1 | |||
| mph(M) | B. cereus | Chromosome | AHFH01000066.1 | |||
| Macrolide esterases | ere(A) |
E. coli,Providencia stuartii, Enterobacter aerogenes |
Plasmids, Integrons, Tn7, sat, aadA1, dfr16, aac(6')-Ib |
M11277, AY183453 DQ157752. 1 AF512546.1 |
Zielinski et al., 2021 |
|
| ere(B) |
E. coli |
Plasmids | A15097, X03988 | |||
| ere(C) |
Riemerella anatipestifer |
Chromosome | CP004020 | |||
| ere(D) |
R. anatipestifer |
Chromosome | KP265721 | |||
| Ribosomal Modification | Mutation in 23S rRNA genes | erm(A) |
S. aureus |
Tn554 | KT803896.1 |
Gupta et al., 2013 |
| erm(B) |
S. pneumoniae |
Tn551, IS1216v | LC125351.1 |
|||
| erm(C) | ˇ S. aureus, S. cohnii | Tn917 | JQ219851.1 |
|||
| Mutation in ribosomal proteins | Mutation in ribosomal protein L4 & L22 |
S. pneumoniae, H. influenzae, and E. coli |
Chromosome | AF126059 |
Zaman et al,, 2007; Schroeder et al., 2016 |
|
| Mutation in ribosomal RNA | Mutation in A2058 & A2059 | S. pneumoniae, Helicobacter pyroli, E.coli | Chromosome, Plasmid | CP000936.1 |
Jiang et al., 2015; Dinos et al., 2017 |
Abbreviations: Mef: Macrolide efflux pump, Tn: Transposon, IS: Insertion sequence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



