
Submitted:
18 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
The need for multi-gene analysis in evolutionary and taxonomic studies is generally accepted. However, sequencing of multiple genes is not always possible. For various reasons, short mito-chondrial DNA barcodes are the only source of molecular information for some species in many genera, although multilocus data are available for other species of the same genera. In particular, such a situation exists in the species-rich butterfly subgenus Polyommatus (Agrodiaetus). Here, we analysed partitioning of this subgenus into species groups by using three data sets. The first data set was represented by short mitochondrial DNA barcodes for all analysed samples. The second and third data sets were represented by combination of short mitochondrial DNA barcodes for a part of the taxa with longer mitochondrial sequences COI+tRNA-Leu+COII (data set 2) and with longer mitochondrial COI+tRNA-Leu+COII and nuclear 5.8S rDNA+ITS2+28S rDNA sequences (data set 3) for the remaining species. We show that DNA barcoding approach (data set 1) failed to reveal the taxonomic structure resulting in numerous polytomies in the phylogenetic tree ob-tained. Combined analysis of the mitochondrial and nuclear sequences (data sets 2 and 3) re-vealed the species groups and position within these species groups even for the taxa for which only short DNA barcodes were available.
Keywords:
biodiversity
; COI
; DNA barcoding
; insects
; Lepidoptera
; Polyommatus
; taxonomy
1. Introduction
Ideally, analysis of evolutionary history and taxonomic structure of living organisms requires comparative analysis of data obtained from multiple sources of evidence (morphological, multilocus molecular, ecological, karyological, etc.) [1,2,3,4,5]. In practice, such a comprehensive analysis is not always possible. Many species are extremely rare and represented in collections by limited number of specimens. Usually, such museum material is hardly suitable for comprehensive multilocus molecular analysis due its old age resulting in DNA degradation, and consideration that unique samples (especially type-specimens) should be rather preserved as important standard vouchers than destroyed in the course of molecular studies.
In this situation, massive single-locus sequencing studies, such as the DNA-barcoding research [6,7], became the only real way to obtain regular molecular information that is available for multispecies comparisons and, thus, can be incorporate into taxonomic research and taxonomic revisions. The situation, when for some species of a genus there are only mitochondrial DNA barcodes, and for other species of the same genus there are multi gene-data, is ordinary [4]. Recently, a novel approach has been suggested for phylogenetic analysis of such genera [8]. This approach is based on the combined analysis of short mitochondrial DNA barcodes for all species of a genus with multilocus data for several representative taxa of the same genus.
In our study, we applied this approach [8] to analysis of taxonomic structure in the species-rich butterfly subgenus Polyommatus (Agrodiaetus) Hübner, 1822 (Lepidoptera, Lycaenidae). This subgenus represents a distinct monophyletic lineage within the diverse genus Polyommatus Latreille, 1804 [4]. The subgenus Agrodiaetus was estimated to have originated only about 3 million years ago [9] and radiated rapidly in the Western Palaearctic [10]. The last published review of the subgenus includes 120 valid species [11]. Although this group has attracted the attention of numerous researchers [4,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23], a large number of unresolved taxonomic problems persist in Agrodiaetus. One of these problems is the taxonomic structure of the subgenus as a whole, namely the division of the subgenus into natural monophyletic lineages (14].
This subgenus has been studied relatively well with respect to molecular markers, and for many species multilocus molecular data are available including such genes as mitochondrial COI, tRNA-leu, COII, cytochrome b and NADH dehydrogenase sequences and nuclear 5.8S rDNA, ITS2, 28S rDNA and EF1-α sequences [9,10,21,23,24,25,26,27,28]. At the same time, for many taxa, especially for rare species from Turkey, Iran, Pakistan and Afghanistan, only mitochondrial DNA barcodes are available [29,30] or molecular data are absent.
In this work we
1) obtain and analyse standard mitochondrial DNA barcodes for five deviated and most enigmatic taxa of the subgenus Agrodiaetus: P. muellerae Eckweiler, 1997 from Pakistan, P. afghanicus (Forster, 1973) and P. frauvartianae Balint, 1997 from Afghanistan, P. bogra Evans, 1932 from Afghanistan and Iran, and P. anticarmon (Koçak, 1983) (= charmeuxi Pages, 1984) from SE Turkey,
2) analyse partitioning of the subgenus Agrodiaetus into species groups by using three data sets. The first data set is represented by short mitochondrial DNA barcodes for all analysed samples. The second and third data sets are represented by combination of short mitochondrial DNA barcodes for a part of the taxa with longer mitochondrial sequences COI+tRNA-Leu+COII (data set 2) and with longer mitochondrial COI+tRNA-Leu+COII and nuclear 5.8S rDNA + ITS2 +28S rDNA sequences (data set 3) for the remaining species,
3) show that DNA barcoding approach (data set 1) failed to reveal the taxonomic structure of the subgenus whereas combined analysis of the mitochondrial and nuclear sequences (data sets 2 and 3) revealed the species groups and position within these species groups even for taxa for which only mitochondrial sequences were available,
4) provide list of the species groups of the subgenus Agrodiaetus, and
5) discuss the status and taxonomic position of P. muellerae, P. afghanicus, P. frauvartianae, P. bogra and P. anticarmon.
2. Materials and Methods
Standard mitochondrial DNA barcodes (658 bp fragments of the cytochrome c oxidase subunit I gene) were obtained for five samples of P. afghanicus, one sample of P. anticarmon (=charmeuxi), six samples of P. bogra, seven samples of P. frauvartianae and one sample of P. muellerae (Table 1). The specimens (except the samples BPAL2125-BPAL2128) were processed at the Department of Karyosystematics of the Zoological Institute of the Russian Academy of Sciences. DNA extraction from a single leg removed from each specimen was done using the QIAamp DNA Investigator Kit (Qiagen, Venlo, The Netherlands) according to the manufacturer’s protocol. Standard lepidopteran barcode primers [7] failed to amplify a sufficient product. Thus, two self-designed forward primers (Nz_COI_b - TAC AAT TTA TCG CTT ATA AACTCA; DRD4F - TAGAAAATGGAGCAGGAA) and two reverse primers, MH-MR1 [31] and Nancy [32] were used for DNA amplification and resulted in a 671 bp fragment of the mitochondrial cytochrome oxidase I gene (COI). The PCR amplifications were performed in a 50 µl reaction volume containing ca. 10-20 ng genomic DNA and 0.5 mM each of forward and reverse primer, 1 mM dNTPs, 10x PCR Buffer (0.01 mM Tris-HCl, 0.05MKCl, 0.1% Triton X-100: pH 9.0), 1 unit Taq DNA Polymerase (Thermo Fisher Scientifics, Waltham, MA, USA), 5m MMgCl2. The temperature profile was as follows: initial denaturation at 94°C for 1 min, followed by 30 cycles of denaturation at 94°C for 45 s, annealing at 50°C for 45 s, and extension at 72°C for 1 min with a final extension at 72°C for 10 min. Amplified fragments were purified using GeneJET Gel Extraction Kit (Thermo Fisher Scientifics, Waltham, MA, USA). Purification was carried out according to the manufacturer’s protocol. The success of PCR amplification and purification was evaluated by electrophoresis of the products in 1% agarose gel. Purified PCR product was used for direct sequencing. Sequencing of the double stranded product was carried out at the Research Resource Center for Molecular and Cell Technologies (St. Petersburg State University).
The samples BPAL2125-BPAL2128 were processed at the Canadian Centre for DNA Barcoding (CCDB, Biodiversity Institute of Ontario, University of Guelph) using their standard high-throughput protocol described by deWaard et al. [33].
Comparison of the obtained COI barcodes revealed 11 unique haplotypes within the five studied species (Table 1).
For the other 130 species and well-differentiated subspecies of the subgenus Agrodiaetus, all available sequences of the mitochondrial (COI, leu-tRNA complete and COII partial) and nuclear (5.8S rDNA partial gene, ITS2 complete and 28S rDNA partial) loci were extracted from GenBank (see Supplementary Material for the GenBank numbers). Sequences of each locus (gene) were aligned separately by using the clustalW algorithm, and then the alignments were checked and corrected manually using BioEdit [34]. Since within Agrodiaetus, the previous phylogenetic analyses of the nuclear sequences 5.8S rDNA + ITS2 +28S rDNA recovered clades which are mostly congruent to those obtained from analyses of the mitochondrial genes COI + COII [25], the nuclear and mitochondrial sequences were concatenated for subsequent phylogenetic study. This concatenation was then combined with the 11 unique haplotypes revealed within the five studied species (Table 1) as well as with the sequences of P. icarus (Rottemburg, 1775). The latter species was used as an outgroup, as previously established by Kandul et al. [10]. This resulted in the final matrix consisting of 142 taxa. For 87 of these 142 taxa, the matrix contained data for both mitochondrial and nuclear genes. For 55 out of these 142 taxa, only mitochondrial gene(s) were available in the matrix.
Since ITS2 sequence has multiple indels which are highly specific on the species-level, it provides additional information for phylogenetic analysis, so we treated all ITS2 indels as binary characters (insertion - 1, deletion - 0). The final concatenated alignment had length of 2948 nucleotides (COI – 1-1539 bp, leu-tRNA – 1540-1604 bp, COII – 1605-2281 bp, 5.8S rDNA + ITS2 + 28S rDNA – 2282-2948 bp) and 23 binary characters.
Three data sets were prepared from the final concatenated alignment. In the data set 1 for all 142 studied samples, only short COI barcodes were presented. In the data set 2, for 13 samples (shown by red asterisk (*) in Figure 1, Figure 2 and Figure 3) only short COI barcodes were available and for the remaining 129 species the longer mitochondrial sequence COI+tRNA-Leu+COII was presented. In the data set 3, the mitochondrial matrix (data set 2) was supplemented by 5.8S rDNA + ITS2 + 28S rDNA nuclear sequences for 87 samples.
Substitution models were inferred using jModeltest, version 2 [35] separately for each gene (locus). Bayesian analysis was conducted using MrBayes 3.2 [36] on four molecular (COI, COII, leu-tRNA and 5.8S rDNA + ITS2 + 28S rDNA genes) and one “standard” (binary) partitions using 20 000 000 generations.
We used the following command block for the first data set:
begin mrbayes;
set autoclose=yes nowarn=yes;
lset nst=6 rates=invgamma;
unlink statefreq=(all) revmat=(all) shape=(all) pinvar=(all);
prset applyto=(all) ratepr=variable;
mcmcp ngen= 20000000 relburnin=yes burninfrac=0.05 printfreq=20000 samplefreq=20000 nchains=4 savebrlens=yes;
mcmc;
sumt;
end;
We used the following command block for the second data set:
begin mrbayes;
charset COI = 1-1539;
charset tRNA = 1540-1604;
charset COII = 1605-2281;
partition Names = 3: COI, tRNA, COII;
end;
begin mrbayes;
set partition=Names;
lset applyto=(1) nst=6 rates=invgamma;
lset applyto=(2) nst=2 rates=equal;
lset applyto=(3) nst=6 rates=invgamma;
unlink shape=(all) pinvar=(all) statefreq=(all) revmat=(all);
prset applyto=(all) ratepr=variable;
mcmcp ngen= 20000000 relburnin=yes burninfrac=0.05 printfreq=20000 samplefreq=20000 nchains=4 savebrlens=yes;
mcmc;
sumt;
end;
We used the following command block for the third data set:
begin mrbayes;
charset COI = 1-1539;
charset tRNA = 1540-1604;
charset COII = 1605-2281;
charset ITS2 = 2282-2948;
charset gaps=2949-2971;
partition Names = 5: COI, tRNA, COII, ITS2, gaps;
end;
begin mrbayes;
set partition=Names;
lset applyto=(1) nst=6 rates=invgamma;
lset applyto=(2) nst=2 rates=equal;
lset applyto=(3) nst=6 rates=invgamma;
lset applyto=(4) nst=6 rates=equal;
unlink shape=(all) pinvar=(all) statefreq=(all) revmat=(all);
prset applyto=(all) ratepr=variable;
mcmcp ngen= 20000000 relburnin=yes burninfrac=0.05 printfreq=20000 samplefreq=20000 nchains=4 savebrlens=yes;
mcmc;
sumt;
end;
3. Results
Analysis of DNA barcodes (Figure 1) did not reveal the structure of the subgenus Agrodiaetus. Only 33 supported clades (posterior probability 0.9 or higher) were recovered, and the position of the majority of species, particularly of our target taxa P. muellerae, P. afghanicus, P. frauvartianae, P. bogra and P. anticarmon was unresolved. This was manifested in the facts that (1) relationships with sister species were not identified (P. muellerae), (2) sister relationships had low support (P. afghanicus, P. frauvartianae, P. bogra), and (3) it was not clear which species groups the target species belonged to (P. muellerae and P. afghanicus).
Combined analysis of the mitochondrial (Figure 2) and mitochondrial + nuclear sequences (Figure 3) resulted in significant increase of the resolution of the phylogenetic tree, with 53 highly supported clades (posterior probability 0.9 or higher) for the dataset 2 and with 65 highly supported clades (posterior probability 0.9 or higher) for the dataset 3. Thus, increasing the alignment length by adding additional mitochondrial and nuclear loci resulted in a dramatic increase in the number of highly supported clades, which is not unexpected. More interesting is that this approach resulted in increased support and tree position detection for those branches for which additional mitochondrial and nuclear data were not available (shown by red asterisks on the trees). It was established that the taxon P. frauvartianae was included in the same clade together with the species P. faramarzii Skala 2001, P. shahrami Skala, 2002 and P. achaemenes Skala, 2002 (Figure 3), while this relationship had extremely low support (0.54) on the DNA barcode tree (Figure 1). It was shown that P. anticarmon was not only a sister species to P. turcicus (Koçak, 1977) (Figure 1), but that both of these taxa were members of the same clade together with P. iphigenia (Herrich-Schäffer, 1847) (Figure 3). The position of the taxon P. australorossicus Lukhtanov & Dantchenko, 2017 on the DNA barcode tree (Figure 1) was unclear due to low support. On the combined tree (Figure 2 and Figure 3), this species is placed with high support in the same clade along with P. hamadanensis (de Lesse, 1959).
Our analysis revealed 11 major lineages, showed by different colours (Figure 3). Two lineages were represented by single species. Seven lineages were highly supported (posterior probability value from 0.91 to 1). The target species P. afghanicus (Figure 4(4-7)) appeared as a lineage distantly related to the lineage P. antidolus – P. iphidamon (species group 8); however, the sister relationship between them was weakly supported. Polyommatus muellerae (Figure 4(8-11)) appeared as a distinct lineage (species group 1). The target species P. frauvartianae (Figure 4(12,13)), P. bogra, and P. anticarmon (Figure 4(14,15)) appeared as members of the species groups 5 and 4.
4. Discussion
The methodology proposed by Talavera et al [8] allowed us to identify positions on the phylogenetic tree for the rare south Central Asian taxa for which molecular data were available only in the form of short DNA barcodes. On the one hand, this is an empirical test of the previously proposed methodology [8]. On the other hand, the data obtained provide the opportunity to discuss in more detail the taxonomy of the studied species.
Polyommatus frauvartianae was described from Afghanistan as a distinct species by Balint [37], but then, on the basis of external morphological similarity, was interpreted as a subspecies of the Iranian-Turkmen species P. glaucias (Lederer, 1871) [11]. Our data unequivocally show that this is an independent species, phylogenetically distant from P. glaucias, but having an undoubted relationship with P. faramarzii, P. shahrami and P. achaemenes, endemics of the Zagros Mts in Iran. From the last three species, P. frauvartianae is well distinguished by the brown (not blue) colouration of the upper side of the wings in males. Thus, these data shed light on the origin of the enigmatic Iranian taxa P. faramarzii, P. shahrami and P. achaemenes, which, having dot-like distribution ranges in the Zagros, did not show close relationships with any other species of the subgenus Agrodiaetus. The new data show that the four species P. faramarzii, P. shahrami, P. achaemenes, and P. frauvartianae are members of the same phylogenetic sub-lineage, spread over a wide area from western Iran to central Afghanistan.
Our data show the conspecificity of two geographically distant population groups identified as P. bogra birjandensis Eckweiler, 2003 (E Iran) and P. bogra afghanistanus (Forster, 1972) (Afghanistan) [11]. Thus, the polytypic concept of the P. bogra species as proposed by Eckweiler and Bozano [11] is confirmed.
The taxon P. anticarmon is also the subject of hot taxonomic debates [11]. Butterflies of this taxon inhabiting SE Turkey are similar in appearance to P. turcicus from NE Turkey and Armenia, differing in larger size. In addition, there is a difference in ecological preferences between P. turcicus and P. anticarmon: while P. turcicus is an alpine species, P. anticarmon is found at relatively low altitudes. Our data show that P. anticarmon is indeed closely related to P. turcicus. According to Eckweiler and Bozano [11], the taxon P. charmeuxi described from SE Turkey is a synonym of P. anticarmon.
In addition, the conducted phylogenetic analysis allows us to discuss another very controversial issue of Agrodiaetus taxonomy, namely, the division of the subgenus into groups of species. In most cases, species delimitation and recognizing monophyletic species groups within Agrodiaetus is difficult because of low number of differentiated morphological characters. The morphology of male genitalia is uniform throughout most of the species, with a few exceptions [11,38,39]. Some Agrodiaetus species show considerable variability in male wing colour in both in visible and ultraviolet wavelength ranges [24,40]. Despite this variation, this character is difficult to use for phylogenetic purposes since in the great majority of species is represented by plesiomorphic state (blue colour), and derived states are found mostly as unique apomorphies characterizing single species but not species groups[24].
The same can be said about chromosomal characters. Although chromosome numbers in the Polyommatus (Agrodiaetus) possess a strong phylogenetic signal [10,28], generally, in the genus Polyommatus, as well as in the closely related genus Lysandra, karyotypes are extremely variable on interspecific level and found mostly as unique apomorphies characterizing single species but not species groups [24,28,41].
de Lesse [12] divided Agrodiaetus into three species complexes based on male colouration and the presence/absence of well-developed tufts formed by androconial scales. In the classification of Hesselbarth et al. [42], Agrodiaetus was divided into eight species groups (actis, admetus, carmon, damon, damone, dolus, poseidon, and transcaspicus) named after their oldest members. Eckweiler and Häuser [14] recognized the admetus and dolus groups but argued that available evidence was too weak to support the remaining groups. Instead, they erected a more inclusive damon group that combined the membership of Hesselbarth et al.’s [42] actis, carmon, damon, damone and transcaspicus groups with some species from the poseidon group. The remainder of the poseidon group was renamed the dama group, and three additional species groups, the dagmara, erschoffii (=Paragrodiaetus Rose & Schurian, 1977) and iphigenides groups, were erected to accommodate species from eastern Iran and Central Asia that had not been considered by Hesselbarth et al. [42].
Similarly, Balint [37] proposed a more fractional division and separated Agrodiaetus into the following groups: actinides, actis, admetus, carmon, dama, damon, damone, dolus, nadira, poseidon, poseidonides and transcaspicus. He also considered Paragrodiaetus as a complex separated from Agrodiaetus, and divided Paragrodiaetus into two groups: erschoffii and glaucias.
Molecular studies [9,10,21,24,25] revealed that the previously recognized species groups [12,14,37,42] were mostly non-monophyletic assemblages. This resulted in creation of a new division of the subgenus Agrodiaetus including of 10 clades: the P. damocles (Herrich-Schäffer, 1844) group, the P. actis (Herrich-Schäffer, 1851) group, the P. erschoffii (Lederer, 1869) group, the P. carmon (Herrich-Schäffer, 1851) group, the P. admetus (Esper, 1783) group, the P. dolus (Hübner, 1823) group, the P. damone (Eversmann, 1841) group, the P. magnificus (Grum-Grshimailo, 1885) group, the P. iphigenia (Herrich-Schäffer, 1847) group and the P. damon (Denis & Schiffermüller, 1775) group [10].
Here, using analysis of additional taxa and additional molecular markers, we demonstrate that the subgenus Agrodiaetus consists of 11 lineages. In particular, we also show that the enigmatic taxon P.(A.) muellerae from Pakistan represents a distinct evolutionary lineage and cannot be included in the previously recognized species groups.
Koçak and Kemal (2001) [43] divided Agrodiaetus into 13 subsections and proposed the following names for these subsections: Actisia, Admetusia, Antidolus, Dagmara, Damaia, Juldus, Musa, Paragrodiaetus, Peileia, Phyllisia, Transcaspius, Xerxesia, and Agrodiaetus s.str. Here we demonstrate that these subsections do not reflect correctly the evolutionary and taxonomic structure of the subgenus (see the list below). Three species groups discovered in our study (erschoffii, damocles, and carmon) are represented by two (erschoffii, damocles groups) and even by four (carmon group) names from the list proposed by Koçak and Kemal [43], whereas five species groups has no names and only five names are unambiguously associated with species groups (one name corresponds to one species group). According to the Code of Zoological Nomenclature (ICZN 10.4) [44] “a uninominal name proposed for a genus-group division of a genus, even if proposed for a secondary (or further) subdivision, is deemed to be a subgeneric name even if the division is denoted by a term such as "section" or "division"”. Thus, the names proposed by Koçak and Kemal (2001) [43] should be considered subgeneric names and therefore subjective junior synonyms of Agrodiaetus since the subgeneric rank of Agrodiaetus is well-founded [4].
5. Conclusions
1. Here, we show that DNA barcoding approach failed to reveal the taxonomic structure of the subgenus Agrodiaetus whereas combined analysis of the mitochondrial and nuclear sequences revealed the species groups and position within these species groups even for taxa for which only short DNA barcodes were available.
2. The Afghani taxon Polyommatus frauvartianae is a distinct species, most closely related to West Iranian endemics P. faramarzii, P. shahrami and P. achaemenes.
3. P. bogra birjandensis (E Iran) and P. bogra afghanistanus (Afghanistan) are confirmed as members of the polytypic species P. bogra.
4. Polyommatus anticarmon (= charmeuxi) is identified as a taxon which is a sister to P. turcicus.
5. The enigmatic Pakistani taxon P. muellerae represents a distinct evolutionary lineage and cannot be included in previously recognized species groups.
6. The subgenus Agrodiaetus consists of 11 lineages and the following species.
Checklist of the species groups and species of Agrodiaetus
(1) muellerae Eckweiler, 1997 species group
muellerae Eckweiler, 1997
(2) admetus (Esper, 1783) species group
admetus (Esper, 1783) Type species of Admetusia Koçak & Seven, 1998
nephohiptamenos (Brown & Coutsis, 1978)
ripartii (Freyer, 1830)
khorasanensis (Carbonell, 2001)
pseudorjabovi Lukhtanov, Dantchenko, Vishnevskaya & Saifitdinova, 2015
yeranyani (Dantchenko & Lukhtanov, 2005)
demavendi (Pfeiffer, 1938)
(3) damone (Eversmann, 1841) species group
damone (Eversmann, 1841)
juldusus (Staudinger, 1886) Type species of Juldus Koçak & Kemal, 2001
mediator Dantchenko & Churkin, 2003
karatavicus Lukhtanov, 1990
iphigenides (Staudinger, 1886)
phyllides (Staudinger, 1886)
(4) iphigenia (Herrich-Schäffer, 1847) species group
baytopi (de Lesse, 1959)
rovshani Dantchenko & Lukhtanov, 1994
iphicarmon Eckweiler & Rose, 1993
tankeri (de Lesse, 1960)
turcicus (Koçak, 1977)
anticarmon (Koçak, 1983) (= charmeuxi Pages, 1984)
iphigenia (Herrich-Schäffer, 1847)
(5) erschoffii (Lederer, 1869) species group
achaemenes Skala, 2002
shahrami Skala, 2001
faramarzii Skala, 2001
frauvartianae Balint, 1997
ardschira (Brandt, 1938)
eckweileri ten Hagen, 1998
pfeifferi (Brandt, 1938)
luna Eckweiler, 2002
birunii Eckweiler & ten Hagen, 1998
darius Eckweiler & ten Hagen, 1998
posthumus (Christoph, 1877)
masulensis ten Hagen & Schurian, 2000
bogra Evans, 1932
caeruleus (Staudinger, 1871)
dagestanicus (Forster, 1960)
phyllis (Christoph, 1877) Type species of Phyllisia Koçak & Kemal, 2001
vanensis (de Lesse, 1957)
erschoffii (Lederer, 1869)
glaucias (Lederer, 1871) Type species of Paragrodiaetus Rose & Schurian, 1977
klausschuriani ten Hagen, 1999
tenhageni Schurian & Eckweiler, 1999
(6) actis (Herrich-Schäffer, 1851) species group
actis (Herrich-Schäffer, 1851) Type species of Actisia Koçak & Kemal, 2001
artvinensis (Carbonell, 1997)
bilgini (Dantchenko & Lukhtanov, 2002)
haigi (Dantchenko & Lukhtanov, 2002)
firdussii (Forster, 1956)
pseudactis (Forster, 1960)
sigberti (Olivier et al. 2000) (? athis Freyer, 1851)
sertavulensis (Koçak, 1979)
hopfferi (Herrich-Schäffer, 1851)
lycius (Carbonell, 1996)
poseidon (Herrich-Schäffer, 1851)
putnami (Dantchenko & Lukhtanov, 2002)
baltazardi (de Lesse, 1962)
(7) damocles (Herrich-Schäffer, 1844) species group
altivagans (Forster, 1956)
damocles (Herrich-Schäffer, 1844)
ciscaucasicus (Forster, 1956)
cyaneus (Staudinger, 1899)
cyaneus musa Koçak & Hosseinpour, 1996 Type species of Musa Koçak & Kemal, 2001
cyaneus xerxes (Staudinger, 1899) Type species of Xerxesia Koçak & Kemal, 2001
sennanensis (de Lesse, 1959)
urartua (Carbonell, 2003)
ectabanensis (de Lesse, 1964)
gorbunovi (Dantchenko & Lukhtanov, 1994)
kendevani (Forster, 1956)
maraschi (Forster, 1956)
wagneri (Forster, 1956)
mofidii (de Lesse, 1963)
sorkhensis Eckweiler, 2003
shamil (Dantchenko, 2000)
shirkuhensis ten Hagen & Eckweiler, 2001
merhaba (de Prins, van der Poorten, Borie, van Oorschot, Riemis & Coenen 1991)
mithridates (Staudinger, 1878)
vaspurakani (Lukhtanov & Dantchenko, 2003)
shahkuhensis (Lukhtanov, Shapoval & Dantchenko, 2008)
barmifiruze (Carbonell, 2000) (no molecular data)
cilicius (Carbonell, 1998) (no molecular data)
sephidarensis Karbalaye, 2008 (no molecular data)
esfahensis (Carbonell, 2000) (no molecular data)
(8) carmon (Herrich-Schäffer, 1851) species group
australorossicus Lukhtanov & Dantchenko, 2017
huberti (Carbonell, 1993)
zapvadi (Carbonell, 1993)
ninae (Forster, 1956)
turcicolus (Koçak, 1977)
antidolus (Rebel, 1901) Type species of Antidolus Koçak & Kemal, 2001
kurdistanicus (Forster, 1961)
morgani (Le Cerf, 1909)
femininoides (Eckweiler, 1987)
karindus (Riley, 1921)
peilei Bethune-Baker, 1921 Type species of Peileia Koçak & Kemal, 2001
dama (Staudinger, 1892) Type species of Damaia Koçak & Kemal, 2001
carmon (Herrich-Schäffer, 1851)
schuriani (Rose, 1978)
surakovi Dantchenko & Lukhtanov, 1994
arasbarani (Carbonell & Naderi, 2000)
damonides (Staudinger, 1899) (=elbursicus Forster, 1956)
lukhtanovi (Dantchenko, 2004)
zarathustra Eckweiler, 1997
pierceae (Lukhtanov & Dantchenko, 2002)
guezelmavi Olivier, Puplesiene, van der Poorten, de Prins & Wiemers, 1999
theresiae Schurian, van Oorschot & van den Brink, 1992
hamadanensis (de Lesse, 1959)
transcaspicus (Heyne, 1895) Type species of Transcaspius Koçak & Kemal, 2001
dizinensis (Schurian, 1982)
iphidamon (Staudinger, 1899)
afghanicus (Forster, 1973)
larseni (Carbonell, 1994) (no molecular data)
zardensis Schurian & ten Hagen, 2001 (no molecular data)
alibali (Carbonell, 2015) (no molecular data)
kashani Eckweiler, 2013 (no molecular data)
lori Eckweiler, 2013 (no molecular data)
(9) dolus (Hübner, 1823) species group
alcestis (Zerny, 1932)
dantchenkoi Lukhtanov & Wiemers, 2003
karacetinae (Lukhtanov & Dantchenko, 2002)
interjectus (de Lesse, 1960)
eriwanensis (Forster, 1960)
menalcas (Freyer, 1837)
aroaniensis (Brown, 1976)
humedasae (Toso & Balletto, 1976)
lurae Parmentier, Vila & Lukhtanov, 2022
orphicus Kolev, 2005
timfristos Lukhtanov, Vishnevskaya & Shapoval, 2016
dolus (Hübner, 1823)
fabressei (Oberthür, 1910)
fulgens (de Sagarra,1925)
violetae (Gómez-Bustillo, Expósito Hermosa & Martínez Borrego, 1979)
rjabovianus (Koçak, 1980)
valiabadi (Rose & Schurian, 1977)
(10) damon ([Denis & Schiffermüller], 1775) species group
damon ([Denis & Schiffermüller], 1775) Type species of Agrodiaetus, type species of Hirtusina Tutt, [1909]
(11) magnificus (Grum-Grshimailo, 1885) species group
actinides (Staudinger, 1886)
dagmara (Grum-Grshimailo, 1888) Type species of Dagmara Koçak & Kemal, 2001
florenciae (Tytler, 1926)
poseidonides (Staudinger, 1886)
pulchellus (Bernardi, 1951)
magnificus (Grum-Grshimailo, 1885)
7. We confirm the previous conclusion [11] that the following subgeneric names are subjective junior synonyms of Agrodiaetus:
Actisia Koçak & Kemal, 2001;
Admetusia Koçak & Seven, 1998;
Antidolus Koçak & Kemal, 2001;
Dagmara Koçak & Kemal, 2001;
Damaia Koçak & Kemal, 2001;
Hirsutina Tutt, [1909]);
Juldus Koçak & Kemal, 2001;
Musa Koçak & Kemal, 2001;
Paragrodiaetus Rose & Schurian, 1977;
Peileia Koçak & Kemal, 2001;
Phyllisia Koçak & Kemal, 2001;
Transcaspius Koçak & Kemal, 2001;
Xerxesia Koçak & Kemal, 2001.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The final alignment of the analysed samples with their GenBank numbers (file in fasta-format).
Author Contributions
Project design, V.A.L.; conceptualization, V.A.L.; PCR amplification and sequencing, N.A.S.; molecular analysis, N.A.S., V.A.L.; writing—original draft preparation, V.A.L.; writing—review and editing, V.A.L., N.A.S., A.V.D., W.E.; collecting material, W.E., A.V.D, V.A.L. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Russian Science Foundation Grant No 19-14-00202 (Continuation) to the Zoological Institute of the Russian Academy of Sciences (St. Petersburg, Russian Federation) (molecular analysis of P. muellerae, P. afghanicus, P. frauvartianae, and P. bogra) and by the state research project 122031100272-3 (molecular analysis of P. anticarmon).
Data Availability Statement
All the analyzed DNA sequences are available via the GenBank links provided
Acknowledgments
We thank Vasiliy Tuzov, Jury Skrylnik and Igor Pljuschtch who provided us the samples from Afghanistan and Pakistan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rubinoff, D.; Holland, B.S. Between two extremes: Mitochondrial DNA is neither the panacea nor the nemesis of phylogenetic and taxonomic inference. Syst. Biol. 2005, 54, 952–961. https://doi.org/10.1080/10635150500234674. [CrossRef]
- Will, K.W.; Mishler, B.D.; Wheeler, Q.D. The perils of DNA barcoding and the need for integrative taxonomy. Syst. Biol. 2005, 54, 844–851. https://doi.org/10.1080/10635150500354878. [CrossRef]
- Knowles, L. L.; Carstens, B. C. Delimiting species without monophyletic gene trees. Syst. Biol. 2007, 56, 887–895. https://doi.org/10.1080/10635150701701091. [CrossRef]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.E.; Vila, R. Establishing criteria for higher-level classification using molecular data: the systematics of Polyommatus blue butterflies (Lepidoptera, Lycaenidae). Cladistics 2013, 29, 166–192. https://doi.org/10.1111/j.1096-0031.2012.00421.x. [CrossRef]
- Hernández-Roldán, J.L.; Dapporto, L.; Dincă, V.; Vicente, J.C.; Hornett, E.A.; Šíchová, J.; Lukhtanov, V.A.; Talavera, G.; Vila, R. Integrative analyses unveil speciation linked to host plant shift in Spialia butterflies. Mol. Ecol. 2016, 25, 4267–4284. https://doi.org/10.1111/mec.13756. [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Royal Soc. B: Biol. Sci. 2003, 270, 313–321. https://doi.org/10.1098/rspb.2002.2218. [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Nat. Acad. Sci. USA 2004, 101, 14812–14817. https://doi.org/10.1073/pnas.0406166101. [CrossRef]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.; Vila, R. DNA barcodes combined with multilocus data of representative taxa can generate reliable higher-level phylogenies. Syst. Biol. 2022, 71, 382–395. https://doi.org/10.1093/sysbio/syab038. [CrossRef]
- Kandul, N.P.; Lukhtanov, V.A.; Dantchenko, A.V.; Coleman, J.W.S.; Sekercioglu, C.H.; Haig, D.; Pierce, N.E. Phylogeny of Agrodiaetus Hübner 1822 (Lepidoptera: Lycaenidae) inferred from mtDNA sequences of COI and COII and nuclear sequences of EF1-α: Karyotype diversification and species radiation. Syst. Biol. 2004, 53, 278–298. https://doi.org/10.1080/10635150490423692. [CrossRef]
- Kandul, N.P.; Lukhtanov, V.A.; Pierce, N.E. Karyotypic diversity and speciation in Agrodiaetus butterflies. Evolution 2007, 61, 546-559. https://doi.org/10.1111/j.1558-5646.2007.00046.x. [CrossRef]
- Eckweiler, W.; Bozano, G.C. Guide to the butterflies of the Palearctic region. Lycaenidae part IV. Omnes artes, Milano, Italy, 2016.
- de Lesse, H. Les nombres de chromosomes dans la classification du groupe d’Agrodiaetus ripartii Freyer (Lepidoptera, Lycaenidae). Rev. Fran. Entomol. 1960, 27, 240–264.
- de Lesse, H. Spéciation et variation chromosomique chez les Lépidoptères Rhopaloceres. Ann. Sci. Nat. Zool. Biol. Anim, 12e série. 1960, 2: 1–223.
- Eckweiler, W.; Häuser, C.L. An illustrated checklist of Agrodiaetus Hübner, 1822, a subgenus of Polyommatus Latreille, 1804 (Lepidoptera: Lycaenidae). Nachr. Entomol. Ver. Apollo 1997, Suppl. 16, 113–168.
- Olivier, A.; Puplesiene, J.; van der Pooten, D.; De Prins, W.; Wiemers, M. (1999) Revision of some taxa of Polyommatus (Agrodiaetus) transcaspicus group with description of a new species from Central Anatolia (Lepidoptera, Lycaenidae). Phegea 1999, 27, 1–24.
- Carbonell, F. Contribution a la conaissance du genre Agrodiaetus Hübner (1822), A. barmifiruze n. sp. et A. musa esfahensis n. ssp, en Iran méridional. Linneana Belgica 2000, 17, 211–217.
- Carbonell, F. (2001) Contribution a la connaissance du genre Agrodiaetus Hübner (1822), A. ahmadi et A. khorasanensis nouvelles espèces dans le Nord de l’Iran (Lepidoptera, Lycaenidae). Linneana Belgica 2001, 18, 105–110.
- Dantchenko, A.V. Genus Agrodiaetus. In: Tuzov, V.K. (ed.) Guide to the butterflies of Russia and adjacent territories, Vol. 2. Pensoft, Sofia & Moscow, 2000, 196–214.
- ten Hagen, W.; Eckweiler, W. Eine neue Art von Polyommatus (Agrodiaetus) aus Zentraliran (Lepidoptera, Lycaenidae). Nachr. Entomol. Ver. Apollo 2001, 22, 53–56.
- Skala, P. New taxa of the subgenus Agrodiaetus Hübner, 1822 from Iran: Polyommatus (Agrodiaetus) faramarzii sp. n., P. (A.) shahrami sp. n. and P. (A.) pfeifferi astyages ssp. n. (Lepidoptera, Lycaenidae). Nachr. Entomol. Ver. Apollo 2001, 22, 101–108.
- Wiemers, M. Chromosome differentiation and the radiation of the butterfly subgenus Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus) a molecular phylogenetic approach. Ph.D. Dissertation, University of Bonn, Bonn, Germany, 2003. 203 pp. http://nbn-resolving.de/urn:nbn:de:hbz:5n-02787.
- Schurian, K.G., ten Hagen W. Polyommatus (Agrodiaetus) urmiaensis sp. n. aus Nordwestiran (Lepidoptera: Lycaenidae). Nachr. Entomol. Ver. Apollo 2003, 24, 1–5.
- Vila, R.; Lukhtanov, V.A.; Talavera, G.; Gil-T.F.; & Pierce, N. E. (2010). How common are dot-like distribution ranges? Taxonomical oversplitting in Western European Agrodiaetus (Lepidoptera, Lycaenidae) revealed by chromosomal and molecular markers. Biol. J. Linn. Soc. 2010, 101, 130–154. https://doi.org/10.1111/j.1095-8312.2010.01481.x. [CrossRef]
- Lukhtanov, V.A.; Kandul, N.P.; Plotkin, J.B.; Dantchenko, A.V.; Haig, D.; Pierce, N.E. Reinforcement of pre-zygotic isolation and karyotype evolution in Agrodiaetus butterflies. Nature 2005, 436, 385–389. https://doi.org/10.1038/nature/03704. [CrossRef]
- Wiemers, M.; Keller, A.; Wolf, M. ITS2 secondary structure improves phylogeny estimation in a radiation of blue butterflies of the subgenus Agrodiaetus (Lepidoptera: Lycaenidae: Polyommatus). BMC Evol. Biol. 2009, 9, 300. https://doi.org/10.1186/1471-2148-9-300. [CrossRef]
- Przybyłowicz, Ł.; Lukhtanov, V.A.; Lachowska-Cierlik, D. Towards the understanding of the origin of the Polish remote population of Polyommatus (Agrodiaetus) ripartii (Lepidoptera: Lycaenidae) based on karyology and molecular phylogeny. J. Zool. Syst. Evol. Res. 2014, 52, 44–51. https://doi.org/10.1111/jzs.12040. [CrossRef]
- Vishnevskaya, M.S.; Saifitdinova, A.F.; Lukhtanov, V.A. Karyosystematics and molecular taxonomy of the anomalous blue butterflies (Lepidoptera, Lycaenidae) from the Balkan Peninsula. Comp. Cytogen. 2016, 10, 1–85. https://doi.org/10.3897/CompCytogen.v10i5.10944. [CrossRef]
- Vershinina, A.O.; Lukhtanov, V.A. Evolutionary mechanisms of runaway chromosome number change in Agrodiaetus butterflies. Sci. Rep. 2017, 7, 8199. https://doi.org/10.1038/s41598-017-08525-6. [CrossRef]
- Lukhtanov, V.A.; Shapoval, N.A.; Dantchenko, A.V. Taxonomic position of several enigmatic Polyommatus (Agrodiaetus) species (Lepidoptera, Lycaenidae) from Central and Eastern Iran: insights from molecular and chromosomal data. Comp.Cytogen. 2014, 8, 313–322. https://doi.org/10.3897/CompCytogen.v8i4.8939. [CrossRef]
- Lukhtanov, V.A.; Dantchenko, A.V. 2017. A new butterfly species from south Russia revealed through chromosomal and molecular analysis of the Polyommatus (Agrodiaetus) damonides complex (Lepidoptera, Lycaenidae). Comp. Cytogen. 2017, 11, 769–795, doi:10.3897/CompCytogen.v11i4.20072. [CrossRef]
- Lukhtanov, V.A.; Sourakov, A.; Zakharov, E.V.; Hebert, P.D.N. DNA barcoding Central Asian butterflies: increasing geographical dimension does not significantly reduce the success of species identification. Mol. Ecol. Resour. 2009, 9, 1302–1310. https://doi.org/10.1111/j.1755-0998.2009.02577.x. [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299.
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: analytical protocols. In Environmental Genomics, Methods in Molecular Biology; Martin, C.C., Ed.; Humana Press: Totowa, N.J., United States, 2008; Volume 410, pp. 275–283, ISBN-13: 978-1588297778.
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61.
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. https://doi.org/10.1038/nmeth.2109. [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. https://doi.org/10.1093/sysbio/sys029. [CrossRef]
- Balint, Z. Reformation of the Polyommatus section with a taxonomic and biogeographic overview (Lepidoptera, Lycaenidae, Polyommatini). Neue Entomol. Nachricht. 1997, 40, 1–67.
- Coutsis, J.G. Notes concerning the taxonomic status of Agrodiaetus tankeri de Lesse (Lepidoptera: Lycaenidae). Nota Lep. 1985, 8, 8-14.
- Coutsis, J.G. The blue butterflies of the genus Agrodiaetus Hübner (Lep., Lycaenidae): symptoms of taxonomic confusion. Nota Lep. 1986, 9, 159-169.
- Parmentier, L.; Vila, R.; Lukhtanov, V.A. Integrative analysis reveals cryptic speciation linked to habitat differentiation within Albanian populations of the anomalous blues (Lepidoptera, Lycaenidae, Polyommatus Latreille, 1804). Comp. Cytogen. 2022, 16, 211–242. https://doi.org/10.3897/compcytogen.v16.i4.90558. [CrossRef]
- Pazhenkova, E.A.; Lukhtanov, V.A. Сhromosomal conservatism vs chromosomal megaevolution: enigma of karyotypic evolution in Lepidoptera. Chromosome Research 2023, 31, 16. https://doi.org/10.1007/s10577-023-09725-9. [CrossRef]
- Hesselbarth, G.; Van Oorschot, H.; Wagener, S. Die Tagfalter der Türkei; Bocholt: Selbstverlag Sigbert Wagener, 1995; vol. 1, 754 pp.
- Koçak, A.O.; Kemal, M. 2001. A study on the biodiversity, zoogeography and taxonomy of the section Agrodiaetus Hbn. in the genus Polyommatus Latr. (Lycaenidae, Lepidoptera). Centre for Entomological Studies Miscellaneous Papers 2001, 78-79, 1-11.
- International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature. The International Trust for Zoological Nomenclature, 1999. https://www.iczn.org/the-code/the-code-online/.
Figure 1.
The Bayesian tree of Agrodiaetus species based on analysis of the short mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (higher than 0.5). Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate samples for which longer mitochondrial and/or nuclear sequences were unavailable.
Figure 1.
The Bayesian tree of Agrodiaetus species based on analysis of the short mitochondrial COI barcodes. Numbers at nodes indicate Bayesian posterior probabilities (higher than 0.5). Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate samples for which longer mitochondrial and/or nuclear sequences were unavailable.

Figure 2.
The Bayesian tree of Agrodiaetus species based on combined analysis of the mitochondrial COI+tRNA-Leu+COII sequences. Numbers at nodes indicate Bayesian posterior probability higher than 0.5. Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate species for which only short DNA barcodes were available.
Figure 2.
The Bayesian tree of Agrodiaetus species based on combined analysis of the mitochondrial COI+tRNA-Leu+COII sequences. Numbers at nodes indicate Bayesian posterior probability higher than 0.5. Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate species for which only short DNA barcodes were available.
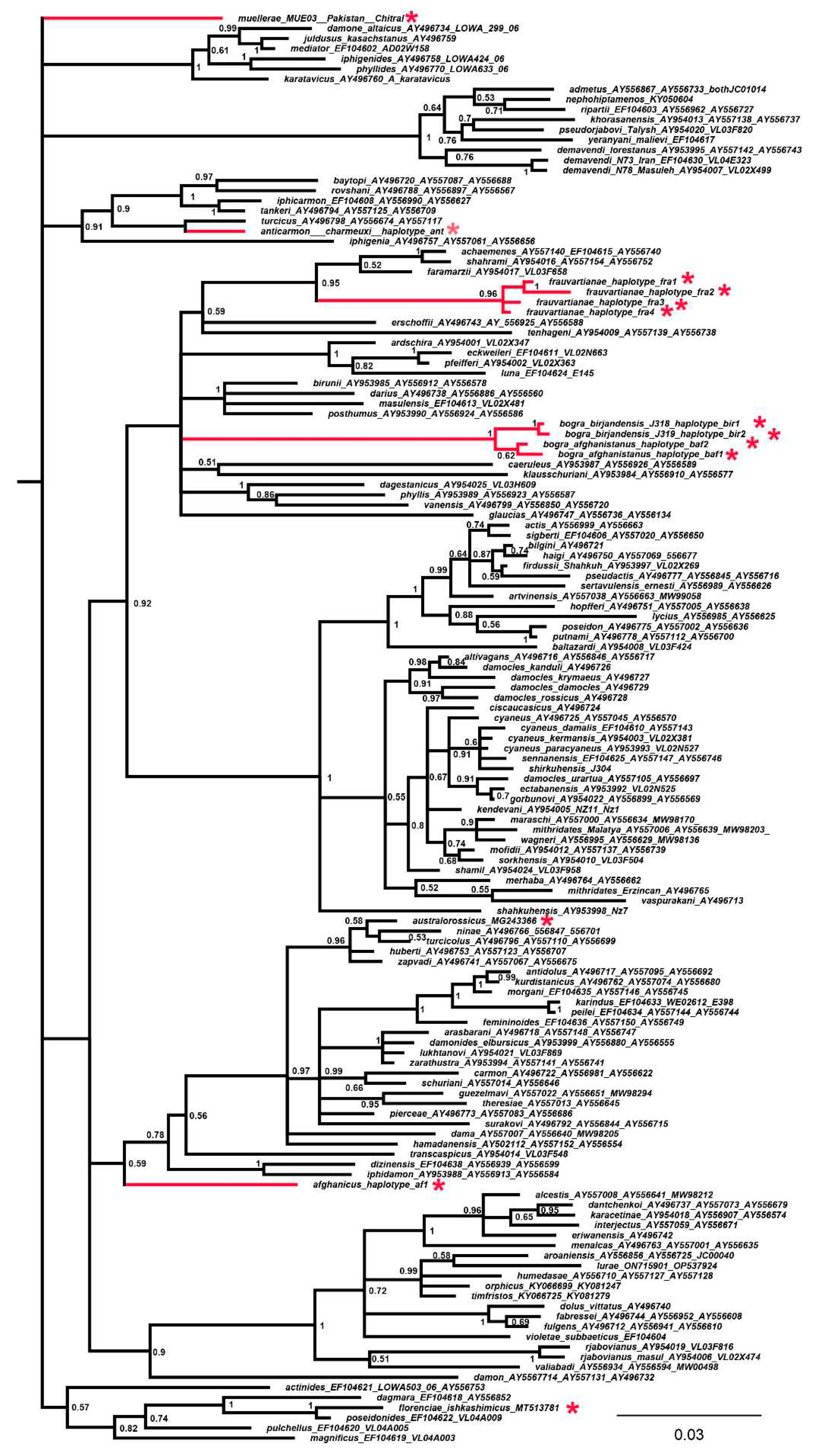
Figure 3.
The Bayesian tree of Agrodiaetus species based on combined analysis of the mitochondrial COI+tRNA-Leu+COII and nuclear 5.8S rDNA + ITS2 +28S rDNA sequences. Numbers at nodes indicate Bayesian posterior probabilities higher than 0.5. Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate species for which only short DNA barcodes were available. 1 – 11 are species groups.
Figure 3.
The Bayesian tree of Agrodiaetus species based on combined analysis of the mitochondrial COI+tRNA-Leu+COII and nuclear 5.8S rDNA + ITS2 +28S rDNA sequences. Numbers at nodes indicate Bayesian posterior probabilities higher than 0.5. Polyommatus icarus (not shown) was used to root the tree. Red asterisks indicate species for which only short DNA barcodes were available. 1 – 11 are species groups.
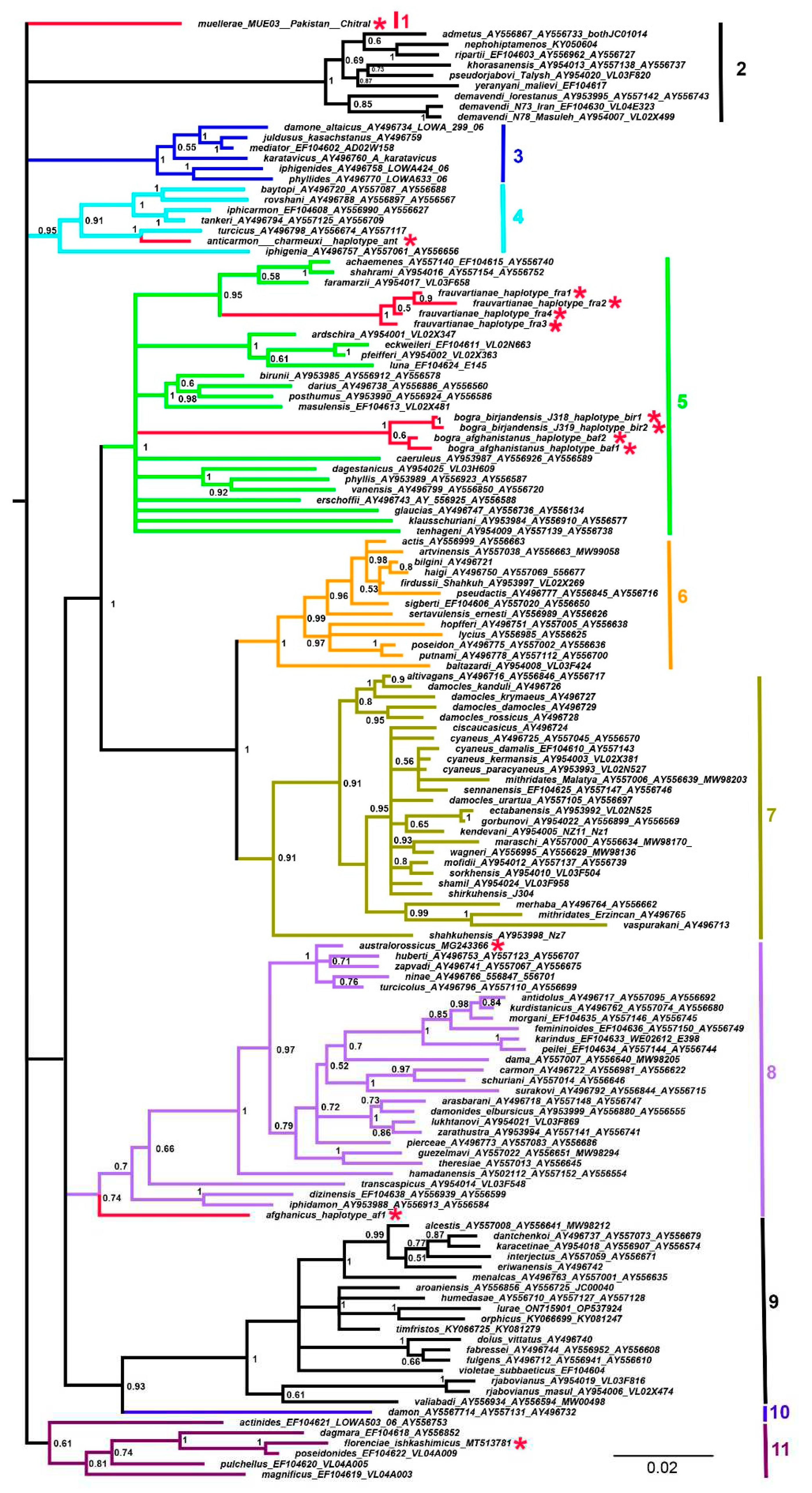
Figure 4.
Polyommatus (Agrodiaetus) afghanicus, P. (A.) muellerae, P. (A.) frauvartianae and P. (A.) anticarmon. 4-7. P. (A.) afghanicus, Afghanistan, Koh-i-Baba, Band-i-Amir, 34.8294° N, 67.1805° E, 2900-3000m., 2.07.2009, Yu. Skrylnik leg. (4, 5 male; 6, 7 female). 8-11. P. (A.) muellerae, Pakistan, Chitral, Birmogh Lasht, 35.8981°N, 71.7712°E, 2600-3000m, 1.07.2001, leg. V. Tuzov (8, 9 male, mue02; 10, 11 female, mue03). 12-13. P. (A.) frauvartianae, male, Afghanistan, Kotale Altimur, 2900m, 10.07.1973, leg. Dr. Resholt. 14-15. P. (A.) anticarmon (=charmeuxi), male CHAR01, Turkey, Hakkari Prov., vic. Üzümcū 1300 m, 28.06-04.07.1976.
Figure 4.
Polyommatus (Agrodiaetus) afghanicus, P. (A.) muellerae, P. (A.) frauvartianae and P. (A.) anticarmon. 4-7. P. (A.) afghanicus, Afghanistan, Koh-i-Baba, Band-i-Amir, 34.8294° N, 67.1805° E, 2900-3000m., 2.07.2009, Yu. Skrylnik leg. (4, 5 male; 6, 7 female). 8-11. P. (A.) muellerae, Pakistan, Chitral, Birmogh Lasht, 35.8981°N, 71.7712°E, 2600-3000m, 1.07.2001, leg. V. Tuzov (8, 9 male, mue02; 10, 11 female, mue03). 12-13. P. (A.) frauvartianae, male, Afghanistan, Kotale Altimur, 2900m, 10.07.1973, leg. Dr. Resholt. 14-15. P. (A.) anticarmon (=charmeuxi), male CHAR01, Turkey, Hakkari Prov., vic. Üzümcū 1300 m, 28.06-04.07.1976.


Table 1.
List of the studied samples and obtained COI sequences.
| Species | Laboratory ID | GeneBank | Haplotype | Country | Locality |
|---|---|---|---|---|---|
| P. (A.) afghanicus | AF05 | OR413713 | af1 | Afghanistan | Koh-i-Baba, Band-i-Amir, 34.8294° N, 67.1805° E, 2900-3000m., 2.07.2009, Yu. Skrylnik leg. |
| BPAL2125 | OR413714 | af1 | Afghanistan | near Kabul, July 2010, I. Pljushch leg. | |
| BPAL2126 | OR413715 | af1 | Afghanistan | the same locality | |
| BPAL2127 | OR413716 | af1 | Afghanistan | the same locality | |
| BPAL2128 | OR413717 | af1 | Afghanistan | the same locality | |
| P. (A.) anticarmon (=charmeuxi) | CHAR01 | OR424389 | ant | Turkey | Hakkari Prov., vic. Üzümcū 1300m, 28.06-04.07.1976 |
| P. (A.) bogra afghanistanus | XX21 | OR424390 | baf1 | Afghanistan | Bamyan prov., 8 km S Bamyan, 2700m, 31.05.2010, O. Pak leg. |
| AAF02 | OR424391 | baf2 | Afghanistan | the same locality | |
| AAF10 | OR424392 | baf1 | Afghanistan | Bamiyan prov., 34.2155° N, 66.8994° E, 2545m, 23.06.2016. O. Pak leg. |
|
| AAF11 | OR424393 | baf1 | Afghanistan | the same locality | |
| P. (A.) bogra birjandensis | J318 | OR413718 | bir1 | Iran | South Khorasan Prov., 26 km N of Birjand, 1900- 2000m, 14.07.2005, V. Lukhtanov leg. |
| J319 | OR413719 | bir2 | Iran | the same locality | |
| P. (A.) frauvartianae | AAF01 | OR424394 | fra1 | Afghanistan | Bamiyan prov., Yakawlang District, Bandi-Amir env., 3300m, 03.08.2011, O. Pak leg. |
| AAF03 | OR424395 | fra2 | Afghanistan | the same locality | |
| AAF04 | OR424396 | fra1 | Afghanistan | the same locality | |
| AAF05 | OR424397 | fra1 | Afghanistan | the same locality | |
| AAF07 | OR424398 | fra3 | Afghanistan | the same locality | |
| AAF08 | OR424399 | fra4 | Afghanistan | Bamiyan prov., Panjab District, Kohi-Baba Mts., Rashak Mts., Shatu Pass, 3490m, 06.08.2011, O. Pak leg. | |
| AAF09 | OR424400 | fra1 | Afghanistan | the same locality | |
| P. (A.) muellerae | MUE03 | OR413720 | mu1 | Pakistan | Chitral, Birmogh Lasht, 35.8981°N, 71.7712°E, 2600-3000m, 01.07.2001, leg. V. Tuzov |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



