Submitted:
16 August 2023
Posted:
21 August 2023
Read the latest preprint version here
Abstract
Single-particle cryo-electron microscopy (cryo-EM SPA) has recently emerged as an exceptionally well-suited technique for determining the structure of membrane proteins (MPs). Indeed, in the last years, it was observed a huge increase in the number of MPs solved by cryo-EM SPA at a resolution better than 3.0 Å in the Protein Data Bank (PDB). However, sample preparation remains a significant challenge in the field. Here, we evaluated in the MPs solved by cryo-EM SPA deposited in the PDB in the last two years at a resolution below 3.0 Å the most critical parameters for sample preparation: i) the surfactant used for protein extraction from the membrane, ii) the surfactant, amphiphiles, nanodiscs or other molecules present in the vitrification step, iii) the vitrification method employed, and iv) the type of grids used. The aim is not to provide a definitive answer on the optimal sample conditions for cryo-EM SPA of MPs, but rather assess the current trends in the MP structural biology community towards obtaining high-resolution cryo-EM structures.
Keywords:
membrane proteins
; cryo-electron microscopy
; detergents
; nanodiscs
; amphipols
; structural biology
1. Introduction
Membrane proteins (MPs) play a crucial role in various essential functions in the cells, including regulation of molecule transport across the membrane, cell-cell recognition, signal transduction, cell adhesion, and control of membrane lipid composition [1]. MPs, when dysfunctional, are also involved in several diseases, including cancers, viral infections, channelopathies and neurodegenerative disorders, being the target of more than 50% of currently marketed drugs [2,3]. The obtention of high-resolution structures of MPs are essential to a comprehensive understanding of their mechanisms of actions and for successful design of new drugs [4].
Over the past three decades, X-ray crystallography has been the prevailing technique for determining high-resolution structures of MPs, as well as other macromolecules and complexes. However, this technique faces a significant challenge in obtaining well-ordered 3D crystals, which require large amounts of sample. This challenge is particularly pronounced for MPs due to their low yield per liter of cell culture in heterologous expression systems [1]. Furthermore, the surfactants employed to extract MPs from the host cell membrane have a detrimental effect on protein stability, impairing the crystallization process [5].
In this context, single-particle cryo-electron microscopy (cryo-EM SPA) has emerged as an exceptionally suitable technique for determining the structure of MPs. Unlike X-ray crystallography, cryo-EM SPA eliminates the need for crystal formation, requires much less sample quantities, and allows the use of a wide variety of hydrophobic environments [6,7]. The impact of cryo-EM SPA in the field of structural biology of MP can be illustrated by some numbers since the first atomic structures of a MP were determined by this technique in 2013 [8,9]. While X-ray crystallography has yielded to a total of 5,828 deposits of MP structures in the Protein Data Bank (PDB) in all history, cryo-EM SPA has led to 3,556 deposits of MP structures in just the last ten years (2013-2022). Notably, this huge progress has been concentrated in the last four years (2019-2022; Figure 1A), which account for 90% of the total number of the cryo-EM SPA deposits of MP structures. Furthermore, there has been a remarkable increase in MP structures solved by cryo-EM SPA at better than 3.0 Å resolution in the last three years (a 174% increase in 2022 compared to 2020; Figure 1B), with 11 structures reaching better than 2.0 Å resolution appearing in the last two years.
Cryo-EM SPA is a technique that involves the application of purified macromolecules onto cryo-EM grids, which are then rapidly frozen to embed the molecules in a thin layer of vitreous ice (vitrification step). The ice layer thickness must be thin enough to support all orientations of the vitrified molecule as thicker layers can lead to a significant increase in noise in the high-resolution information [10]. Then, the cryo-EM grid is transferred to a transmission electron microscope (TEM), where images are acquired under cryogenic conditions [6,11]. The image processing pipeline to obtain the final 3D cryo-EM map of a target molecule encompasses several steps [12]. Briefly, each captured image consists of a movie composed of dose-fractionated frames, which requires motion estimation and correction. The corrected images, known as micrographs, are then used to estimate the contrast transfer function (CTF) parameters, which describes how aberrations in the TEM affect the image of the samples. Additionally, the micrographs are utilized to locate and select single particles. The particles are extracted from the micrographs and subjected to iterative classifications to sort out suitable ones for constructing the 3D cryo-EM map. Finally, the atomic structure of the molecule is built within the 3D cryo-EM map, resulting in the final structure of the target molecule [12].
The high increase in popularity of cryo-EM SPA among structural biologists in the last ten years is a result of several technical advancements, often referred to as the ‘resolution revolution’ [13]. One of the most significant technical advancements was the introduction of complementary metal-oxide semiconductor (CMOS) direct detector device (DDD) cameras, which improved the detective quantum efficiency (DQE) compared to charged-coupled device (CCD) and other detectors [14]. Moreover, the DDD cameras have a high frame rate that allows the data to be captured as dose-fractionated image stacks rather than a single micrograph. This approach enables the correction of motion induced by stage or beam movements, leading to a substantial improvement in image quality by recovering the high-resolution signal that may have been compromised by such motion [15,16]. Moreover, the data processing has become more robust and user friendly with automated pipelines in different software [17,18,19]. Other technical advances continue to be continuously made in recent years, which has allowed obtaining high resolution data to occur more and more routinely. Notably, the new generation of field emission fun (cold FEG) and the introduction of post-column imaging filters, such as Selectris and Selectris X significantly enhanced the signal-to-noise ratio (SNR) and contrast of TEM images [20]. Additionally, the latest generation of DDD cameras (Falcon 4) allows for the acquisition of images with super-resolution information at a significantly faster rate, up to ten times faster, while also producing smaller file sizes [20,21].
In view of these great advancements, sample preparation arises as the most import challenge for cryo-EM SPA, especially for MPs, which require their extraction and purification from the membrane, and their stabilization in an aqueous environment. This is typically achieved by solubilizing the MPs in surfactants that can be later replaced by more suitable surfactants or amphiphiles molecules prior to the vitrification step. A recent quantitative analysis examined the use of surfactants and other types of molecules employed for protein extraction from the membrane and during the vitrification step in all unique cryo-EM structures of MPs solved up to 2020 [22]. However, at that time, only 192 MP structures were available at near-atomic resolution (better than 3.0 Å resolution). However, in the last two years (2021-2022), new 623 MP structures have been released at this resolution level (more than three times the number of structures released up to the year 2020 with ≤ 3.0 resolution) (Figure 1B). Thus, in light of the significant increase in the number of MP structures solved by cryo-EM SPA at very high resolution in the last two years, we present an updated and expanded analysis about surfactants and other molecules used for protein extraction and at vitrification step, focused solely on MP structures solved at near-atomic resolution. Additionally, we assessed the vitrification method and the type of the grids employed for obtaining these structures. Our objective is not to provide a definitive answer regarding the perfect sample conditions for cryo-EM SPA for MPs, but rather evaluate the current trends in the MP structural biology community towards obtaining cryo-EM structures at resolution better than 3.0 Å.
To conduct these analysis, we searched on the Protein Data Bank (PDB) for deposited structures of MPs (annotated as PDBTM, MemPROTMD, OPM or mpstruc in PDB) solved by cryo-EM SPA at a resolution better than 3.0 Å. We then examined in the published manuscripts referring to these structures the following sample preparation parameters: i) the surfactant used for MP extraction from the membrane, ii) the surfactant, amphiphiles, nanodiscs or other molecules present in the vitrification step, iii) the vitrification method employed, and iv) the type of grids used. It is worth noting that some manuscripts may include multiple structures obtained under different states (e.g., open and closed conformations, presence of different ligands, and others). To avoid bias of more extensively studied proteins and provide greater representativeness of all MPs structural molecular biological groups, a single structure from each manuscript was analyzed unless when one of the parameters analyzed above was different (unique reports). As a result, we analyzed a total 313 different manuscripts, resulting in 313 unique reports analyzed for the surfactant used for MP extraction, 316 unique reports analyzed for the surfactant present in the vitrification step, 313 unique reports analyzed for the vitrification method employed, and 320 unique reports analyzed for the type of grids employed.
2. Sample Preparation
2.1. Amphiphiles Used for Membrane Protein Extraction
MPs are characterized by their dual hydrophobic-hydrophilic nature, which arises from the presence of a highly hydrophobic transmembrane domain, along with an hydrophilic extracellular and cytoplasmic domains within their structures. Consequently, maintaining MPs in a soluble and stable state outside the lipid biological bilayer poses a considerable challenge, as they have a heightened tendency to aggregate or denature in an aqueous medium [23]. Therefore, upon expression of MPs in heterologous systems, it becomes necessary to extract them from the lipid bilayer and replace it with another amphiphilic system for subsequent structural and functional studies. Various surfactants have been developed to provide a suitable environment for MPs extraction from the membrane, with detergents being the historically first ones used for this purpose [24]. Detergents are amphipathic molecules consisting of a hydrophilic head group (typically polar, sometimes charged) and a lipophilic or hydrophobic (apolar) tail. This architecture allows detergents to insert their lipophilic tails into the lipidic membrane, thereby disrupting it and extracting the MP from the membrane as the detergent concentration increases. As a result of the hydrophilic-lipophilic balance of detergent molecules, they form spontaneously micelles, which are pseudo-spherical assemblies. Micelle formation occurs once the critical micelle concentration (CMC) is reached and when the sample is above critical micellar temperature. Through micelle formation, a MP becomes part of the detergent-protein complex, sometimes leading to the complete loss of surrounding lipids [25].
Among the MP structures solved by cryo-EM SPA at near-atomic resolution (better than 3.0 Å resolution), almost all molecules employed for membrane protein extraction are detergents (309 of 313 reports). The only exceptions where the four cases where styrene-maleic acid copolymer (SMA) was used (Figure 2A). This is a free detergent system where SMA copolymers solubilize MPs directly from the membrane keeping the native lipid environment forming polymer-bounded nanodiscs, often called as native nanodiscs [26]. The four most common detergents used for membrane protein extraction are, in order of frequency of use (Figure 2A):
- 1)
- Detergents with a single maltose-based polar moiety and single alkyl chain (DDM, UDM, DM): 131 of 313 reports (41%). In this group, the most widely used detergent is DDM, which is present in 120 of 131 reports of this detergent class. DM is present in 10 of 131 reports and UDM in 1 of 131 reports;
- 2)
- Detergents belonging to the maltoside-neopentyl glycol (MNG) family (MNG, LMNG and DMNG): 107 of 313 reports (34%). In this group, the most widely used detergent is LMNG, which is present in 104 of 107 reports of this detergent class. MNG is present in 2 of 107 reports and DMNG is present in 1 of 107 reports;
- 3)
- Glyco-diosgenin (GDN): 18 of 313 reports (6%);
- 4)
- Digitonin: 16 of 313 reports (5%).
DDM (n-Dodecyl-β-D-maltopyranoside), as well as their relatives DM (n-Decyl-β-maltoside and UDM (n-Undecyl-β-maltoside), is a detergent that belongs to alkyl maltoside class. DDM was developed in 1980 and has been widely used in X-ray crystallography [25]. As it is demonstrated here, it is still the preferred detergent for extracting membrane proteins for high-resolution cryo-EM studies. The polar head of DDM is provided by a disaccharide (maltose), while its hydrophobic portion consists of 12-carbon alkyl chain (Figure 3A). DDM proves to be highly effective for membrane protein extraction, and being non-ionic, it is able to maintain the stable native state of numerous proteins. Additionally, DDM exhibits a low CMC value (0.17 mM), which allows a reduced detergent usage. It forms well-defined micelles, albeit relatively large in size and surrounds protein with a significant and mobile belt-like structure [24].
LMNG (lauryl maltose neopentyl glycol) is a detergent that belongs to the maltoside-neopentyl glycol (MNG) family. This detergent class is characterized by have a maltoside head group and two alkyl chains and two carbohydrate head groups connected with each other via a neopentyl glycol unit in the central region, that result in a dimeric architecture compared with detergents from alkyl maltoside class. LMNG, as example, is a dimer of DDM, thus, presenting two maltoside units in their hydrophilic domain and two alkyl chains (Figure 3B). This molecular rearrangement of LMNG results in a CMC value even lower than DDM (0.01 mM of rather than 0.17 mM), although it has been reported to form larger micelles [24,27,28].
Although DDM is still the most widely used detergent for membrane protein extraction even for cryo-EM studies, its once clear dominance seems to have slightly decreased in recent years. In comparison to a previous report [22], our analysis focused on only high-resolution cryo-EM structures indicates a slight decrease in DDM usage from membrane protein extraction (~41% compared to ~43% from the previous report) and a significant rise in the utilization of detergents from the MNG family, particularly LMNG (~34% compared to ~22% from the previous report). Originally employed with remarkable success for the solubilization and stabilization of GPCRs (G protein-coupled receptors) in comparison to DDM [27], the utilization of LMNG has extended to numerous other classes of membrane proteins, as showed by our data and the existing literature [24]. A relevant practice worth highlighting is the addition of cholesteryl hemisuccinate (CHS) to detergent-solubilized MPs. CHS is a water-soluble cholesterol analog able to provide a more native and stabilizing environment for many purified MPs [29,30]. This addition of CHS has been reported mostly in combination with detergents from the MNG family, especially LMNG [75 of the 107 reports using MNG detergents (70%) contain CHS; Figure 2B].
Two other detergents that rank among the top four most commonly used surfactants for membrane protein extraction here identified are the steroidal detergents GDN and digitonin (6% and 5% of the reports, respectively). Digitonin is a natural steroidal saponin produced by the purple foxglove plant Digitalis purpurea. Its commercial form is in fact a mixture of about five different glycosides, with digitonin and digalonin, another saponin of similar structure, as the main components [31]. GDN is a recently developed synthetic variant of digitonin with some modifications. In digitonin, the head group comprises two galactoses, two glucoses, and a xylose, whereas GDN features a branched maltoside head group (Figure 3C and 3D, respectively). Digitonin and GDN have CMC values of 0.25–0.50 mM and 0.018 mM, respectively. GDN, in particular, demonstrates higher homogeneity, enhanced water solubility, and lower toxicity compared to digitonin [24]. The presence of steroid hydrophobic groups in these detergents allows their usage in challenging cases involving membrane proteins that are unstable when solubilized with other classes of detergents [24]. In comparison to a previous report [22], we observed a decrease in the digitonin usage (5% compared to 10% from the previous report) and an increase in the utilization of GDN (6% compared to 3% from the previous report).
2.2. Amphiphiles and Other Molecules Used in the Vitrification Step
The six most classes of amphiphiles (or other types of molecules) present at vitrification step of MP structures solved by cryo-EM SPA at near-atomic resolution (better than 3.0 Å resolution) are, in order of frequency of use (Figure 4A):
- 1)
- Mixed detergents: 64 of 316 reports (20%). In this group, the most widely used association of detergents was the combination of LMNG and GDN, which is present in 48 of 64 reports.
- 2)
- Nanodiscs: 61 of 316 reports (19%). In this group, the most widely used type is the membrane scaffold proteins (MSP)-based nanodiscs, which is present in 50 of 61 reports.
- 3)
- Detergents with a single maltose-based polar moiety and single alkyl chain (DDM, UDM, DM): 52 of 316 reports (16%). In this group, the most widely used detergent is DDM, which is present in 43 of the 53 reports of this detergent class. DM is responsible for the 9 remaining reports;
- 4)
- Glyco-diosgenin (GDN): 52 of 316 reports (16%).
- 5)
- Detergents belonging to the maltoside-neopentyl glycol (MNG) family (MNG and LMNG): 42 of 316 reports (13%). In this group, the most widely used detergent is LMNG, which is present in 40 of the 43 reports of this detergent class. MNG is responsible for the 3 remaining reports.
- 6)
- Digitonin: 29 of 316 reports (9%).
These data indicate that a step of surfactant exchange or reconstitution into nanodiscs is often performed after extraction from the membrane and prior to vitrification of the membrane protein. While there is a clear predominance of DDM and LMNG usage during the protein membrane extraction step, no particular surfactant or other type of molecule exhibits distinct dominance during the vitrification step. There is similar frequency of use between the mixed detergents, nanodiscs, DDM and GDN (20%, 19%, 16% and 16% of the reports respectively). There is a significant increase in the usage of association between detergents in the vitrification step (20% of the reports) compared to membrane protein extraction step (4.8% reports, Figure 4A and Figure 2A, respectively). Notably, among the detergent associations used present in the vitrification step, the combination of LMNG and GDN stands out as the most prevalent, accounting for 48 out of the 64 reports (75%).
The combination of different detergents can be useful to explore their stabilizing properties of detergent-protein and detergent-detergent interactions and increase the effectiveness of the surfactant power obtained from this association without denaturing the membrane proteins. Strong detergent-protein interactions enhances membrane protein stabilization and strong detergent-detergent interactions prevent protein aggregation and may minimize protein denaturation [24]. However, it is important to note that excessively strong detergent-protein interactions can lead to protein denaturation [32]. In the case of association between LMNG and GDN, LMNG interacts more strongly with the hydrophobic surface of MPs than DDM, GDN and digitonin due to its two alkyl chains, providing a strong detergent-protein interaction, but without causing protein denaturation [24]. On the other hand, GDN has similar effects on protein-detergent interactions than DDM; however the rigid hydrophobic group, resulted by the presence of a steroidal unit, promotes an enhanced detergent-detergent interactions than LMNG [24]. It is important to note that the addition of CHS to detergent-solubilized MPs was mostly reported when the mixture of LMNG and GDN was employed at the vitrification step (45 out of the 48 reports, 93%) (Figure 4B). In comparison to the protein membrane extraction step, addition of CHS is also extensively adopted in combination with detergents from the MNG family (24 out of 42 reports, 57%) (Figure 4B).
Nanodiscs is a detergent free-environment that represent another prominent type of preparation present in the vitrification step (19% of the reports) (Figure 4A), where the protein is reconstituted in a lipid environment [33]. While 4 reports of SMA copolymer nanodiscs (native nanodiscs) were identified at the protein membrane extraction step (Figure 2A), the vitrification step featured the same 4 reports of native nanodiscs in addition to 49 reports of membrane scaffold proteins (MSP)-based nanodiscs, 4 of saposin-lipid-protein complexes (Salipro) and 3 of circularized nanodiscs (Figure 4A and Figure 5A). MSP-based nanodiscs (50 of the 61 reports of nanodiscs, 82%) are small (7-50 nm in diameter) disc-shaped structure formed by self-assembled bilayer of lipid bilayer (which mimic the native lipid bilayer) stabilized by two encircling amphipathic helical proteins derived from apolipoprotein A1 (ApoA1), called Membrane Scaffold Proteins (MSP) [33]. In contrast to other surfactants and molecules that are supplied by commercial sources, MSPs can be expressed in-house using E.coli. Besides, MSPs have been engineered to form nanodiscs with different transmembrane domain sizes, such as the MSPs here identified MSP1E3D1 (18 of the 49 reports of MSP-nanodiscs, 37%), MSP2N2 (18 of the 49 reports of MSP-nanodiscs, 37%) and MSP1D1 (14 of the 49 reports of MSP-nanodiscs, 28%) (Figure 5A). The first MSP, MSP1, was engineered with its sequence based on ApoA1, but without the globular N-terminal of its protein [34]. The MSP1D1 variant of MSP1 does not contain the first 11 residues of the helix 1 and generates nanodiscs ~9.7 nm in diameter [35]. The MSP1E3D1 is a variant of MSP1 that contains additional 3 helix sequences and generate nanodiscs of ~12.9 nm. MSP2N2 is a fusion of MSP2D1 and another MSP variant, MSP1D2, which features a complete deletion of the helix 1. MSP2N2 generates larger nanodiscs of 15.0-16.5 nm in diameter [35]. Regarding the lipid composition of the MSP-based nanodiscs of the MP structures solved cryo-EM SPA below 3.0 Å resolution, total lipid extracts of E. coli and soybean polar lipid extracts are the most widely used (13 and 11 of the 49 reports of MSP-nanodiscs, respectively) (Figure 5B).
Circularized nanodiscs (3 of the 61 reports of nanodiscs) have been recently designed based on engineered scaffold proteins based on Apo1 scaffold protein where the N and C-terminus are covalently linked [36,37]. As a result of the covalent circularization, larger (up to 80 nm diameter) and more homogeneous and stable nanodiscs were produced compared to the standard ones [37]. Two MP structures identified by this study were obtained at high-resolution using circularizable scaffold protein NW9 (~8.5 nm of diameter) and one using NW11 (~11 nm of diameter). In terms of lipid composition, a report that used NW9 and the report that used NW11 employed soybean polar lipid extract. The NW9 remaining report utilized E. coli total extract lipid. Finally, 4 out of 61 reports of nanodiscs have been identified using a system related to MSP-nanodiscs, known as Salipro, wherein another protein, Saposin A, replaces MSPs. Regarding the lipid composition of MP structures that used Salipro at the vitrification step, two reports employed soybean polar lipid extract (one of them mixed with 25% cholesterol), while the other two reports used brain polar lipids.
Although nanodiscs have emerged as the predominant detergent-free environment of high-resolution MP structures, it is worth noting that two other detergent-free systems, namely amphipols and peptidiscs, have also been employed, albeit to a lesser extent. Similar to nanodiscs, amphipols and peptidiscs replace the detergent used for membrane protein extraction prior to the sample vitrification. Among the 316 unique reports analyzed, amphipols were employed in the vitrification step in four cases (Figure 4A). Amphipols represent a class of short and flexible amphipathic polymers that directly bind to the hydrophobic surface of MPs and effectively replace detergents [38]. Unlike detergents that require 50-200 to associate to with a single MP, only a small number of amphipols molecules are needed to cover the entire hydrophobic surface of a MP [24,39]. The four identified cases presenting these molecules used amphipols A8-35, although other types of amphipols can also be used for MP structural studies [38]. Even when examining the vitrification step of MP structures resolved at lower resolution ranges (3-5 Å) in the past two years, the usage of amphipols remained limited, with a low number of reports (3) may be due to their cost. In addition, a single report was found regarding the use of peptidiscs (classified within “others” in Figure 4A) among the MP structures resolved at below 3.0 Å resolution. Peptidiscs constitute a detergent-free surfactant system very recently developed (2018) that uses multiple copies of a short sequence of a unique helical peptides (NSPr) redesigned to have optimal hydrophobic and hydrophilic properties. These helical peptides wrap around the hydrophobic transmembrane domains of the MPs, and shield them from the watery solution [40]. Although these two systems offer easier manipulation compared to nanodiscs and have shown promising results, they are not yet widely adopted by the structural biology community for cryo-EM studies of MPs.
As mentioned above, a surfactant exchange or nanodiscs reconstitution step is frequently realized after membrane extraction and prior to vitrification of the membrane protein samples. There is a strong decrease in the use of DDM and LMNG (when used alone, without mixing with another detergent) in the vitrification step. Analysis of the substitutions among the four most common surfactants (DDM, LMNG, GDN and digitonin) used for membrane protein extraction revealed that DDM had the highest rate of exchange prior to the vitrification steps. Out of the 120 unique reports that used DDM for membrane protein extraction, only 42 retained it for the vitrification step (Figure 6A), representing a 65% replacement of DDM. The most frequently used substitute for DDM in the vitrification step was nanodiscs (30 unique reports), followed by GDN (20 unique reports), association between detergents (13 unique reports), detergents from the MNG family (9 unique reports), and digitonin (6 unique reports) (Figure 6A). Interestingly, in all cases but one, where DDM was present in the vitrification step, it was also employed for protein extraction from the membrane. Regarding LMNG, of the 104 unique reports that used this detergent for membrane protein extraction, 33 retained it as the sole surfactant in the vitrification step (Figure 6B). Although the use of LMNG alone has significantly decreased in the vitrification step, it has not been completely replaced. Instead, in 33 unique reports, LMNG was used in combination with other detergents, with 31 reports including a combination of LMNG and GDN and 2 combining LMNG with both GDN and digitonin. Thus, LMNG was completely exchanged before sample vitrification in 37 out the 104 unique reports (35% of total replacement). The most common substitute for LMNG in the vitrification step was nanodiscs (13 unique reports), followed by GDN (12 unique reports), digitonin (11 unique reports) and DDM (1 unique report) (Figure 6B). GDN was the detergent less exchanged between membrane protein extraction and vitrification steps. Out of the 18 unique reports that utilized this detergent for membrane protein extraction, it was exchanged before sample vitrification in only 4 cases, all of which were nanodiscs (22% replacement) (Figure 6C). Regarding digitonin, out of the 16 unique reports that used this detergent for membrane protein extraction, 7 retained it as the sole surfactant in the vitrification step, and 1 used it in combination with GDN. In 3 cases, digitonin was replaced by Amphipols A8-35, and in 5 cases by nanodiscs before sample vitrification (50% of total replacement) (Figure 6D).
The usage profile of detergent, nanodiscs and other molecules in the vitrification step presented in this study focused on analysis of high-resolution cryo-EM structures obtained between 2021 and 2022 is quite different from a previous report conducted two years ago [22], which analyzed cryo-EM structures solved at all resolution ranges. In this previous report, digitonin was the most preferred choice at the vitrification step, accounting for 18% of the reports [22]. However, in the current study, digitonin is only the sixth most frequent choice at the vitrification step (Figure 4A) and accounts for 9% of the reports. On the other hand, we observed a substantial increase in the usage of mixed detergents, particularly those containing LMNG, in comparison to the previous report. We identified in the present study that mixed detergents was the preferred choice for the sample vitrification, comprising 20% of the reports (Figure 4A), whereas in the previous study, mixed detergents accounted for only 5% of the reports [22]. Furthermore, we noted a slight increase in the usage of GDN in the current report in comparison to the previous one, with this detergent representing 16% of the reports, as opposed to the 11% in the previous study. The usage of single-maltose detergents (DDM, UDM, DM), detergents from MNG family and MSP-based nanodiscs were similar in both studies (~17%, ~13% and ~16% of the reports, respectively). Regarding the exchange of the amphiphiles between membrane protein extraction and vitrification steps, both studies observed a similar replacement rate of DDM by other detergents or nanodiscs (65-68%) and that digitonin was found to be the least exchanged detergent. However, a notable difference was observed between the two studies concerning LMNG. Whereas in the previous study observed a high level of LMNG substitution for other detergents or amphiphiles before samples vitrification (~59% of the reports) [22], here we observed that LMNG was maintained in the vitrification step, alone or in combination with other detergents, particularly GDN (Figure 6B).
A frequently employed strategy to improve the quality of the samples for cryo-EM SPA is the addition of some specific detergents before the vitrification step to overcome common problems observed in the vitrified samples, such as protein aggregation, high particle adsorption at the air-water interface, and, preferential particle orientation [7,11,41]. Through the cryo-EM sample preparation, after the sample is deposited onto the grids prior to freezing, a significant number of protein molecules diffuse to the air-water interface within milliseconds. This phenomenon can lead to certain protein regions preferentially bind to the air-water interface, resulting in protein particles molecules aligning in a preferred orientation [42,43]. This orientation bias hinders the achievement of high-resolution structures and adversely affects the reliability and interpretability of the obtained structures [43,44]. Also, protein molecules adsorbed onto the air-water interface may suffer partial damage or even form a layer of denatured protein [42,43]. Here, we were able to identify 5 reports where fluorinated detergents (3 using fluorinated fos-choline-8 at 1.5-3 mM, and 2 using fluorinated octyl maltoside, FOM, at 60 μM and 5 mM, respectively) and 1 where a high CMC detergent (0.2% CHAPS) were added before vitrification step to mitigate these issues. Finally, we also identified 1 report that used for the first time megabodies to prevent preferential particle orientation [45].
3. Sample Vitrification
The vitrification step consists of applying approximately 3 μl of protein solution onto a grid, followed by blotting with filter paper to eliminate excess solution and create a very thin layer of protein suspension on the grid. The grid is then is rapidly frozen by plunging it into liquid ethane cooled by liquid nitrogen [6]. The plunge-freezing could be done manually or using some commercial devices, such as VitrobotTM (Thermo Fischer ScientificTM), EM GP2 (Leica Microsystems GmnH) and CryoplungeTM 3 (Gatan, Inc.). In our analysis, we identified 294 reports that used VitrobotTM (95.2%), 16 that used EM GP2 (5.2%), 1 report that used CryoplungeTM 3 (0.3%), and 1 report that performed the blotting and plunge freezing manually (0.3%).
However, optimal ice thickness is difficult to reproduce from one grid to another due to the uneven surface properties of the blotting papers [46]. Moreover, the dwell time of the samples on the grids before freezing at level of seconds is sufficient to favor the particle adsorption at the air-water interface [47]. In order to solve these limitations, several efforts have been made recently to develop alternative blotting-free methods that guarantee a more reliable and reproducible grid preparation and that diminish the deleterious effects of the air-water interface. For these blotting-free methods, several automated devices have been developed employing inkjet printing (Spotiton) [48], gas pressure spray (microfluidic devices) [49,50], ultrasonic spray [51,52,53], electrostatic spray [54], and sample scribing [55,56] (for a detailed description of these methods, see [11]). Here, we did not identify the use the any of these new blotting-free methods for obtention of MP cryo-EM structures at high-resolution, likely due to the limited commercial availability of most of these new devices. However, very recently, the commercialized form of Spotiton, known as Chamaleon (SPT Labtech Ltd)[57], has been acquired by several cryo-EM facilities worldwide. This system employs an inkjet mechanism to spray picoliter-sized droplets of the sample solution are sprayed onto self-wicking nanowire grids. The entire sample volume can be accurately dispensed through the piezoelectric inkjet head by applying voltage pulses to control ice thickness [48,58,59]. This method potentially provides more reproducible grid preparations than blotting-dependent methods and reduces the dwell time on the grids before freezing for seconds to hundreds of milliseconds [60]. This minimizes the number of particles adsorbed at the air-water interface and increase the number of particles adsorbed in the vitreous ice. Consequently, particle distribution is enhanced in the grids and preferred particle orientation problem is alleviated [58,60]. The Chamaleon system has been tested in several cryo-EM facilities. For example, vitrification experiments using Chamaleon conducted with soluble proteins demonstrated improved particle distribution onto the grids, and enhanced stability and reduced dissociation level of a protein/DNA complex compared to gold grids [61]. In the future, it will be possible to evaluate its overall impact in the cryo-EM field in terms of quality gains in sample vitrification and in the collected micrographs from several types of samples, including MPs.
4. Type of the Grids
Grids serve as the sample carriers for TEM and cryo-EM. They have a standard diameter of 3 nm and typically consist of two main components: a mesh base and a foil [11]. The mesh base is made by a metal that is capable of offering mechanical stability, electron beam conduction and heat dissipation. The foil coats the grid by being placed on top of this mesh and contains micrometer-sized holes that support the vitreous ice with the embedded protein particles and allow the electrons passage during microscope imaging [62]. The foil can contain a regular repeating array of circular holes (Quantifoil® or C-flatTM grids) or an irregular geometry of holes (Lacey grids) [6,11]. These varying designs of mesh base and foils meet specific experimental needs and preferences in TEM and cryo-EM applications.
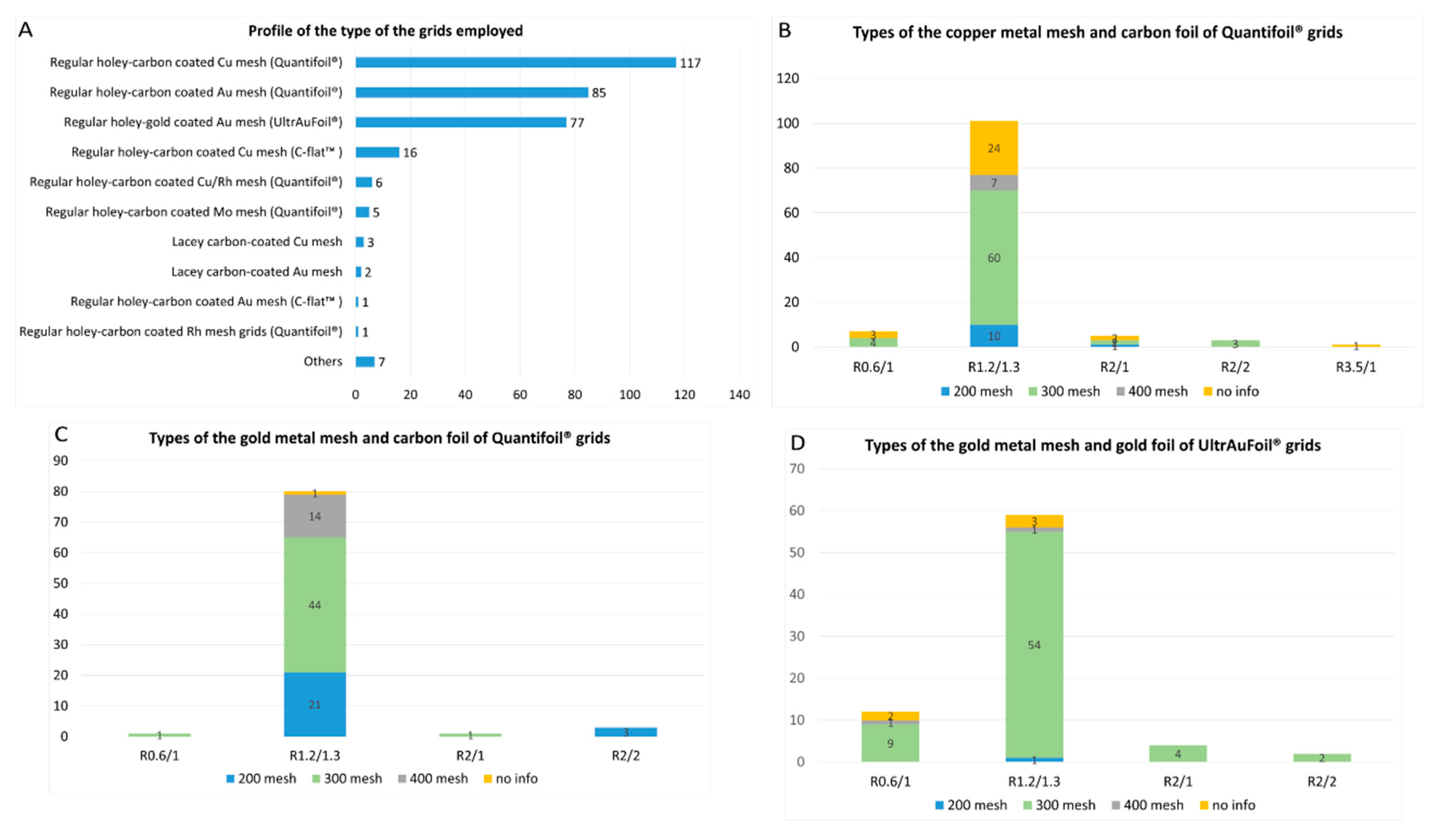
The three major types of the grids used for cryo-EM data collection of MP solved by cryo-EM SPA at near-atomic resolution (below 3.0 Å resolution) in the last two years (2021-2022) are, in order of frequency of use (Figure 7A):
- 1)
- Regular holey-carbon coated copper mesh grids (Quantifoil®): 117 of 320 reports (36.5%);
- 2)
- Regular holey-carbon coated gold mesh grids (Quantifoil®): 85 of 320 reports (26.5%);
- 3)
- Regular holey-gold coated gold mesh grids (UltrAuFoil®): 77 of 320 reports (24%).
Copper mesh grids are the most popular choice identified in this study, as would be expected as they are the historically most popular grids used in cryo-EM [62]. Copper mesh grids are inexpensive and display a good electron conductivity, which is an important feature in preventing detrimental sample charging of the samples, that would lead to a significant loss of the micrographs quality [6,7,11]. Gold mesh grids are even more conductive [15], although more expensive. Concerning the foil, amorphous carbon-film coated grids are the also the most popular choice for cryo-EM SPA of MP here identified, as well as for cryo-EM in general, because they are also inexpensive and easily manufactured into foils [6,11,62]. However, they have challenging limitations [63,64]. The difference in thermal contraction between the carbon foil and the metal mesh (copper, gold and other metals) on cooled temperatures during vitrification may shrink the carbon foil and induce deformations in the ice layer. This results in a significant beam-induced motion during microscope imaging, resulting in a loss of resolution in the micrographs [63,64]. The substitution of carbon films to gold films on gold mesh grids (UltrAuFoil® grids) results in an entire grid structure composed of the same material that eliminates the retraction of the foil caused by the differential thermal contraction on cooled temperatures. As a result, a drastic reduction in the beam-induced motion during microscope imaging is obtained, significantly improving the micrographs quality [63,64]. Besides the 70 UltrAuFoil grid reports here identified, we noticed 2 reports that used homemade nanofabricated holey-gold grids with holes of 1.5 and 2 μm respectively and 2 reports that employed of CryoMatrix® gold grids. Recently, it was demonstrated that amorphous nickel-titanium alloy film has similar effects in reducing beam-induced motion and improving image quality than gold films. Here, we identified two reports that employed this film (CryoMatrix® nickel titanium alloy film).
C-flatTM grids have demonstrated good performance in the cryo-EM field as they are potentially able to provide, thanks to their ultra-flat surface architecture, a even and lower ice thickness and better particle dispersion in the grids compared to Quantifoil® grids [65,66]. Despite these advantages, they did not represent a popular choice for cryo-EM of MP at near-atomic resolution, as evidenced by their usage in only 17 of 320 reports (5.3%) identified here (16 reports for C-flatTM holey-carbon coated copper mesh grids and 1 report for homemade C-flatTM holey-gold coated gold mesh grids) (Figure 7A). Grids with irregular geometry of the holes (Lacey grids) were used only in 6 reports (1.8%), being 5 for carbon coated grids and 1 report for gold coated grids (Figure 7A), although these grids are quite popular for the cryo-EM analysis in general.
Foil films can be categorized based on the diameter of the holes and the width between them, while metal meshes are classified by the number of squares per inch. For instance, the notation R1.2/1.3 denotes a foil film with a hole size of 1.2 μm hole size and a distance of 1.3 μm between the holes. The advantage of using foils with larger holes is the possibility of collecting more images per hole resulting in faster automated data collection. However, larger hole sizes increase the chances of obtaining deformation and inhomogeneity in the ice layer, resulting in beam-induced motion of the specimen and producing blurred images [15]. The use of smaller distances between the holes reduces the amount of foil on the metal support of the grids, which enhances the chances of the protein particles entering in the holes. However, it results in more fragile grids that require more careful sample preparation manipulation. Among the three major types of the grids used for cryo-EM SPA of MP proteins identified in this study, the most typical foil films chosen were R1.2/1.3 (77-94% of the reports). All other reports used R2/1, R2/2 and R0.6/1 foils, except for a single case where R3.5/1 foils were used (Figure 7B–D). The choice of using 0.6-2 μm hole size foils can be attributed to the nature of the samples deposited on the grids, which did not require larger foil holes. Regarding the metal mesh, a grid designed as 400 mesh indicates that it contains 400 squares per inch. Smaller mesh grids consequently have larger squares with a greater number of foil holes, which makes the setting of the automated data collection faster and easier. For example, a 100 mesh grid contains squares of 204 μm and 6,659 foil holes per grid square, while a 400 mesh grid contains squares of 38 μm and 231 foil holes per grid square [67]. On the other hand, the smaller the mesh, the more fragile the grid, which requires very careful and delicate handling of the grid during sample preparation. In this study, we noticed that the cryo-EM community of MPs has a preference for Quantifoil® and UltrAuFoil® grids of 300 mesh (Figure 7B–D).
A common challenge in cryo-EM sample preparation is the propensity of certain samples to adsorb onto the carbon film rather than entering the foil holes. This behavior results in a collection of an insufficient number of particles to achieve high-resolution structures [6,7,11]. To address this issue, a common used strategy involves creating conditions that promote the entry of protein particles into the holes by adding a continuous film on top of the foil. This provides an extra physical support for the protein particles to adsorb onto [6,7,11]. Moreover, these additional films can also improve particle distribution on the grid and significantly decrease the preferential particle orientation issue. Here, we identified 8 reports where a thin layer of continuous carbon film (2-4 nm) was added on top of the grids, being 5 on carbon-coated copper mesh grids (Quantifoil®), 2 on carbon-coated gold mesh grids (Quantifoil®) and 1 on UltrAuFoil® grids. We also identified 11 reports where the grids were covered with a layer of graphene oxide, being 5 on carbon-coated copper mesh grids (Quantifoil®), 5 on carbon-coated gold mesh grids (Quantifoil®) and 1 on Lacey carbon supported copper grids. The thin carbon layer reduces the signal-to-noise ratio of micrographs, leading to loss of high-resolution information, particularly for small proteins. On the other hand, this problem is not observed with the introduction of the graphene oxide layer [68,69]. Finally, we identified 1 report that a lysine-coated graphene oxide layer was applied on carbon-coated copper mesh grids (Quantifoil®). This extra-layer of lysine is very effective to attract negatively charged particles onto grid surface [70].
5. Conclusions and Perspectives
The present study provided an overview of the prevailing choices for the most critical parameters in sample preparation of membrane protein structures solved by cryo-EM SPA at near-atomic resolution over the past two years. We observed that DDM continues to be the most commonly used surfactant extracting MPs from the membrane. However, its prominent dominance appears to have decreased in recent years as we noticed a significant increase in the use of LMNG for membrane protein extraction, often combined with the cholesterol analog CHS. We also observed that there is a high rate of substitutions between the types of surfactants used for membrane protein extraction and those present in the sample vitrification step. These detergents are being replaced in the vitrification step mainly by the detergent-free environment MSP-based nanodiscs and by a mixture of detergents containing LMNG, GDN and the cholesterol analog CHS. However, other detergent-free environments, such as amphipols and circularized nanodiscs, have been providing excellent results for high-resolution membrane protein structure determination by cryo-EM SPA, and their usage could increase in the future. Moreover, the potential development of new detergents that focus on enhancing detergent-detergent interactions by promoting hydrogen-bonding interaction between detergent molecules [24] can provide new surfactants with higher membrane extraction efficiency and enhanced protein-stabilizing effect. Additionally, the recent discovery of two small diglucoside amphiphiles able of forming discoidal assemblies containing lipids, similar to the nanodiscs [71], could impact the field in the future. This discovery may open possibilities for obtaining new surfactants, where any surfactant exchange would be unnecessary between membrane protein extraction and vitrification steps.
All reports analyzed here employed the blotting technique for sample vitrification, with a clear predominance for utilizing the VitrobotTM automated device. Moreover, analysis of the reports showed a dominant trend in using cooper or gold 300 mesh grids coated with regular holey-carbon (Quantifoil®) or holey-gold (UltrAuFoil®) R1.3/1.2 foils. Despite of the good performance of the C-flatTM grids in obtaining high-resolution cryo-EM structures, their utilization is not yet widespread in the MP field. The popularization of the recent new blotting-free methods for sample vitrification, such as Chamaleon and others, holds the potential to make obtaining high-resolution structures more routine in the future, once these methods confirm their initial promising results in obtaining reduced particle adsorption in the air-water interface in a wide range of samples. Other strategies to decrease the particle adsorption in the air-water interface and mitigate the preferential orientation problem include the addition of some specific detergents to the samples before vitrification. Here, we identified a few cases where the addition of fluorinated detergents, high CMC detergents, and megabodies proved effective in addressing these challenges. The use of this strategy could be further explored by the MPs cryo-EM community. Strategies to prevent the particle sticking in the foil films and which can also mitigate the preferential orientational problem were also identified in this study. We identified some cases where a thin layer of continuous carbon film or graphene oxide was applied to avoid preferred orientation. However, other successful strategies aimed at the same purpose in the cryo-EM studies of different proteins, such as cryo-EM grids with Ni-NTA functionalized films to bind histidine-tagged proteins [72,73], 2D streptavidin crystal films [74] or PEGylation treatment of the grids [75] or of the protein samples [76], were not identified in this study. These strategies may have limited awareness within the MP cryo-EM community and could be further explored in the future.
Despite the remarkable advances in cryo-EM SPA in recent years, it remains an evolving technique. The continuous development of novel sample preparation approaches has the potential to usher in groundbreaking progress in the structural knowledge at near-atomic level of an increasing number of MPs. Moreover, it can offer novel accessible experimental conditions for obtaining high-resolution structures of challenging samples and large membrane protein complexes.
Author Contributions
Conceptualization: CAHF and CVB; data curation: CAHF; formal analysis: CAHF and CVB; writing – original draft preparation: CAHF; writing – review and editing: CVB; supervision: CVB; funding acquisition: CAHF and CVB. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by AFM-Téléthon #23207 for CVB and CAHF and CAHF has received funding from the European Union’s Horizon Europe Research and Innovation Program under grant agreement no. 101026386.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ApoA1 | Apolipoprotein A1 |
| C12E9 | Nonaethylene glycol monododecyl ether |
| C12E8 | Octaethylene glycol monododecyl ether |
| CCD | Charged-coupled device |
| CHAPS | 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate |
| CHS | Cholesteryl hemisuccinate |
| CMC | Critical micelle concentration |
| CMOS | Complementary metal-oxide semiconductor |
| Cryo-EM | Cryo-electron microscopy |
| Cymal-4 | 4-cyclohexyl-1-butyl-β-D-maltoside |
| Cymal-6 | 6-cyclohexyl-1-hexyl-β-D-maltoside |
| DDD | Direct detector device |
| DDM | n-Dodecyl-β-D-maltopyranoside |
| DM | Decyl-b-D-Maltopyranoside |
| DQE | Detective quantum efficiency |
| FOM | Fluorinated octyl maltoside |
| GDN | Glyco-diosgenin |
| LDAO | N,N-Dimethyl-n-dodecylamine N-oxide |
| LMNG | Lauryl Maltose Neopentyl Glycol |
| MNG | Maltose-Neopentyl Glycol |
| MP | Membrane Protein |
| MSP | Membrane Scaffold Protein |
| OGNG | Octyl glucose neopentyl glycol |
| t-PCCαM | 4-trans-(4-trans-Propylcyclohexyl)-cyclohexyl α-maltoside |
| PDB | Protein Data Bank |
| POPC | 1-palmitoyl-2-oleoyl-glycero-3-phosphocholine (16:0-18:1PC) |
| POPE | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine (16:0-18:1PE) |
| POPG | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoglycerol (16:0-18:1PG) |
| Salipro | Saposin-lipid-protein complexes |
| SMA | Styrene maleic acid copolymer |
| SPA | Single-particle analysis |
| SPME | Soybean polar lipid extract |
| SNR | Signal-to-noise ratio |
| TEM | Transmission electron microscopy |
| TLE | Total lipid extract |
| UDM: | n-Undecyl-β-D-maltopyranoside |
References
- Pandey, A.; Shin, K.; Patterson, R.E.; Liu, X.-Q.; Rainey, J.K. Current Strategies for Protein Production and Purification Enabling Membrane Protein Structural Biology. Biochem. Cell Biol. 2016, 94, 507–527. [Google Scholar] [CrossRef]
- Gong, J.; Chen, Y.; Pu, F.; Sun, P.; He, F.; Zhang, L.; Li, Y.; Ma, Z.; Wang, H. Understanding Membrane Protein Drug Targets in Computational Perspective. CDT 2019, 20, 551–564. [Google Scholar] [CrossRef]
- Arinaminpathy, Y.; Khurana, E.; Engelman, D.M.; Gerstein, M.B. Computational Analysis of Membrane Proteins: The Largest Class of Drug Targets. Drug Discovery Today 2009, 14, 1130–1135. [Google Scholar] [CrossRef]
- Vénien-Bryan, C.; Li, Z.; Vuillard, L.; Boutin, J.A. Cryo-Electron Microscopy and X-Ray Crystallography: Complementary Approaches to Structural Biology and Drug Discovery. Acta Crystallogr F Struct Biol Commun 2017, 73, 174–183. [Google Scholar] [CrossRef]
- Carpenter, E.P.; Beis, K.; Cameron, A.D.; Iwata, S. Overcoming the Challenges of Membrane Protein Crystallography. Current Opinion in Structural Biology 2008, 18, 581–586. [Google Scholar] [CrossRef]
- Sgro, G.G.; Costa, T.R.D. Cryo-EM Grid Preparation of Membrane Protein Samples for Single Particle Analysis. Front. Mol. Biosci. 2018, 5, 74. [Google Scholar] [CrossRef]
- Xu, Y.; Dang, S. Recent Technical Advances in Sample Preparation for Single-Particle Cryo-EM. Front. Mol. Biosci. 2022, 9, 892459. [Google Scholar] [CrossRef]
- Liao, M.; Cao, E.; Julius, D.; Cheng, Y. Structure of the TRPV1 Ion Channel Determined by Electron Cryo-Microscopy. Nature 2013, 504, 107–112. [Google Scholar] [CrossRef]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 Structures in Distinct Conformations Reveal Activation Mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef]
- Wu, S.; Armache, J.-P.; Cheng, Y. Single-Particle Cryo-EM Data Acquisition by Using Direct Electron Detection Camera. Microscopy (Tokyo) 2016, 65, 35–41. [Google Scholar] [CrossRef]
- Weissenberger, G.; Henderikx, R.J.M.; Peters, P.J. Understanding the Invisible Hands of Sample Preparation for Cryo-EM. Nat Methods 2021, 18, 463–471. [Google Scholar] [CrossRef]
- Cheng, Y.; Grigorieff, N.; Penczek, P.A.; Walz, T. A Primer to Single-Particle Cryo-Electron Microscopy. Cell 2015, 161, 438–449. [Google Scholar] [CrossRef]
- Kühlbrandt, W. The Resolution Revolution. Science 2014, 343, 1443–1444. [Google Scholar] [CrossRef]
- McMullan, G.; Faruqi, A.R.; Henderson, R. Direct Electron Detectors. In Methods in Enzymology; Elsevier, 2016; Volume 579, pp. 1–17. ISBN 978-0-12-805382-9. [Google Scholar]
- Brilot, A.F.; Chen, J.Z.; Cheng, A.; Pan, J.; Harrison, S.C.; Potter, C.S.; Carragher, B.; Henderson, R.; Grigorieff, N. Beam-Induced Motion of Vitrified Specimen on Holey Carbon Film. Journal of Structural Biology 2012, 177, 630–637. [Google Scholar] [CrossRef]
- Li, X.; Mooney, P.; Zheng, S.; Booth, C.R.; Braunfeld, M.B.; Gubbens, S.; Agard, D.A.; Cheng, Y. Electron Counting and Beam-Induced Motion Correction Enable near-Atomic-Resolution Single-Particle Cryo-EM. Nat Methods 2013, 10, 584–590. [Google Scholar] [CrossRef]
- Scheres, S.H.W. RELION: Implementation of a Bayesian Approach to Cryo-EM Structure Determination. Journal of Structural Biology 2012, 180, 519–530. [Google Scholar] [CrossRef]
- Punjani, A.; Rubinstein, J.L.; Fleet, D.J.; Brubaker, M.A. CryoSPARC: Algorithms for Rapid Unsupervised Cryo-EM Structure Determination. Nat Methods 2017, 14, 290–296. [Google Scholar] [CrossRef]
- Gómez-Blanco, J.; De La Rosa-Trevín, J.M.; Marabini, R.; Del Cano, L.; Jiménez, A.; Martínez, M.; Melero, R.; Majtner, T.; Maluenda, D.; Mota, J.; et al. Using Scipion for Stream Image Processing at Cryo-EM Facilities. Journal of Structural Biology 2018, 204, 457–463. [Google Scholar] [CrossRef]
- Nakane, T.; Kotecha, A.; Sente, A.; McMullan, G.; Masiulis, S.; Brown, P.M.G.E.; Grigoras, I.T.; Malinauskaite, L.; Malinauskas, T.; Miehling, J.; et al. Single-Particle Cryo-EM at Atomic Resolution. Nature 2020, 587, 152–156. [Google Scholar] [CrossRef]
- Guo, H.; Franken, E.; Deng, Y.; Benlekbir, S.; Singla Lezcano, G.; Janssen, B.; Yu, L.; Ripstein, Z.A.; Tan, Y.Z.; Rubinstein, J.L. Electron-Event Representation Data Enable Efficient CryoEM File Storage with Full Preservation of Spatial and Temporal Resolution. IUCrJ 2020, 7, 860–869. [Google Scholar] [CrossRef]
- Le Bon, C.; Michon, B.; Popot, J.-L.; Zoonens, M. Amphipathic Environments for Determining the Structure of Membrane Proteins by Single-Particle Electron Cryo-Microscopy. Quart. Rev. Biophys. 2021, 54, e6. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, K.; Yan, N. The Recombinant Expression Systems for Structure Determination of Eukaryotic Membrane Proteins. Protein Cell 2014, 5, 658–672. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, H.S.; Youn, T.; Byrne, B.; Chae, P.S. Impact of Novel Detergents on Membrane Protein Studies. Chem 2022, 8, 980–1013. [Google Scholar] [CrossRef]
- Stetsenko, A.; Guskov, A. An Overview of the Top Ten Detergents Used for Membrane Protein Crystallization. Crystals 2017, 7, 197. [Google Scholar] [CrossRef]
- Dörr, J.M.; Scheidelaar, S.; Koorengevel, M.C.; Dominguez, J.J.; Schäfer, M.; Van Walree, C.A.; Killian, J.A. The Styrene–Maleic Acid Copolymer: A Versatile Tool in Membrane Research. Eur Biophys J 2016, 45, 3–21. [Google Scholar] [CrossRef]
- Lee, S.; Ghosh, S.; Jana, S.; Robertson, N.; Tate, C.G.; Vaidehi, N. How Do Branched Detergents Stabilize GPCRs in Micelles? Biochemistry 2020, 59, 2125–2134. [Google Scholar] [CrossRef]
- Breyton, C.; Javed, W.; Vermot, A.; Arnaud, C.-A.; Hajjar, C.; Dupuy, J.; Petit-Hartlein, I.; Le Roy, A.; Martel, A.; Thépaut, M.; et al. Assemblies of Lauryl Maltose Neopentyl Glycol (LMNG) and LMNG-Solubilized Membrane Proteins. Biochimica et Biophysica Acta (BBA) - Biomembranes 2019, 1861, 939–957. [Google Scholar] [CrossRef]
- Magnani, F.; Shibata, Y.; Serrano-Vega, M.J.; Tate, C.G. Co-Evolving Stability and Conformational Homogeneity of the Human Adenosine A 2a Receptor. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 10744–10749. [Google Scholar] [CrossRef]
- Thompson, A.A.; Liu, J.J.; Chun, E.; Wacker, D.; Wu, H.; Cherezov, V.; Stevens, R.C. GPCR Stabilization Using the Bicelle-like Architecture of Mixed Sterol-Detergent Micelles. Methods 2011, 55, 310–317. [Google Scholar] [CrossRef]
- Fukunaga, K.; Szabo, L.; Fales, H.M.; Pitha, J. 2-Hydroxypropyldigitonin: Synthesis and Properties of Preparations Differing in Degree of Substitution. Journal of Pharmaceutical Sciences 1988, 77, 640–642. [Google Scholar] [CrossRef]
- Otzen, D.E. Protein Unfolding in Detergents: Effect of Micelle Structure, Ionic Strength, PH, and Temperature. Biophysical Journal 2002, 83, 2219–2230. [Google Scholar] [CrossRef] [PubMed]
- Sligar, S.G.; Denisov, I.G. Nanodiscs: A Toolkit for Membrane Protein Science. Protein Science 2021, 30, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Bayburt, T.H.; Grinkova, Y.V.; Sligar, S.G. Self-Assembly of Discoidal Phospholipid Bilayer Nanoparticles with Membrane Scaffold Proteins. Nano Lett. 2002, 2, 853–856. [Google Scholar] [CrossRef]
- Denisov, I.G.; Grinkova, Y.V.; Lazarides, A.A.; Sligar, S.G. Directed Self-Assembly of Monodisperse Phospholipid Bilayer Nanodiscs with Controlled Size. J. Am. Chem. Soc. 2004, 126, 3477–3487. [Google Scholar] [CrossRef]
- Nasr, M.L.; Baptista, D.; Strauss, M.; Sun, Z.-Y.J.; Grigoriu, S.; Huser, S.; Plückthun, A.; Hagn, F.; Walz, T.; Hogle, J.M.; et al. Covalently Circularized Nanodiscs for Studying Membrane Proteins and Viral Entry. Nat Methods 2017, 14, 49–52. [Google Scholar] [CrossRef]
- Nasr, M.L.; Wagner, G. Covalently Circularized Nanodiscs; Challenges and Applications. Current Opinion in Structural Biology 2018, 51, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Zoonens, M.; Popot, J.-L. Amphipols for Each Season. J Membrane Biol 2014, 247, 759–796. [Google Scholar] [CrossRef]
- Le Maire, M.; Champeil, P.; Møller, J.V. Interaction of Membrane Proteins and Lipids with Solubilizing Detergents. Biochimica et Biophysica Acta (BBA) - Biomembranes 2000, 1508, 86–111. [Google Scholar] [CrossRef]
- Carlson, M.L.; Young, J.W.; Zhao, Z.; Fabre, L.; Jun, D.; Li, J.; Li, J.; Dhupar, H.S.; Wason, I.; Mills, A.T.; et al. The Peptidisc, a Simple Method for Stabilizing Membrane Proteins in Detergent-Free Solution. eLife 2018, 7, e34085. [Google Scholar] [CrossRef]
- Kampjut, D.; Steiner, J.; Sazanov, L.A. Cryo-EM Grid Optimization for Membrane Proteins. iScience 2021, 24, 102139. [Google Scholar] [CrossRef]
- Glaeser, R.M. Preparing Better Samples for Cryo–Electron Microscopy: Biochemical Challenges Do Not End with Isolation and Purification. Annu. Rev. Biochem. 2021, 90, 451–474. [Google Scholar] [CrossRef] [PubMed]
- Naydenova, K.; Russo, C.J. Measuring the Effects of Particle Orientation to Improve the Efficiency of Electron Cryomicroscopy. Nat Commun 2017, 8, 629. [Google Scholar] [CrossRef] [PubMed]
- Sorzano, C.O.S.; Semchonok, D.; Lin, S.-C.; Lo, Y.-C.; Vilas, J.L.; Jiménez-Moreno, A.; Gragera, M.; Vacca, S.; Maluenda, D.; Martínez, M.; et al. Algorithmic Robustness to Preferred Orientations in Single Particle Analysis by CryoEM. Journal of Structural Biology 2021, 213, 107695. [Google Scholar] [CrossRef]
- Uchański, T.; Masiulis, S.; Fischer, B.; Kalichuk, V.; López-Sánchez, U.; Zarkadas, E.; Weckener, M.; Sente, A.; Ward, P.; Wohlkönig, A.; et al. Megabodies Expand the Nanobody Toolkit for Protein Structure Determination by Single-Particle Cryo-EM. Nat Methods 2021, 18, 60–68. [Google Scholar] [CrossRef]
- Armstrong, M.; Han, B.-G.; Gomez, S.; Turner, J.; Fletcher, D.A.; Glaeser, R.M. Microscale Fluid Behavior during Cryo-EM Sample Blotting. Biophysical Journal 2020, 118, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Noble, A.J.; Wei, H.; Dandey, V.P.; Zhang, Z.; Tan, Y.Z.; Potter, C.S.; Carragher, B. Reducing Effects of Particle Adsorption to the Air–Water Interface in Cryo-EM. Nat Methods 2018, 15, 793–795. [Google Scholar] [CrossRef]
- Razinkov, I.; Dandey, V.P.; Wei, H.; Zhang, Z.; Melnekoff, D.; Rice, W.J.; Wigge, C.; Potter, C.S.; Carragher, B. A New Method for Vitrifying Samples for CryoEM. Journal of Structural Biology 2016, 195, 190–198. [Google Scholar] [CrossRef]
- Kaledhonkar, S.; Fu, Z.; White, H.; Frank, J. Time-Resolved Cryo-Electron Microscopy Using a Microfluidic Chip. In Protein Complex Assembly; Marsh, J.A., Ed.; Methods in Molecular Biology; Springer New York: New York, NY, 2018; ISBN 978-1-4939-7758-1. [Google Scholar]
- Kontziampasis, D.; Klebl, D.P.; Iadanza, M.G.; Scarff, C.A.; Kopf, F.; Sobott, F.; Monteiro, D.C.F.; Trebbin, M.; Muench, S.P.; White, H.D. A Cryo-EM Grid Preparation Device for Time-Resolved Structural Studies. IUCrJ 2019, 6, 1024–1031. [Google Scholar] [CrossRef]
- Rubinstein, J.L.; Guo, H.; Ripstein, Z.A.; Haydaroglu, A.; Au, A.; Yip, C.M.; Di Trani, J.M.; Benlekbir, S.; Kwok, T. Shake-It-off: A Simple Ultrasonic Cryo-EM Specimen-Preparation Device. Acta Crystallogr D Struct Biol 2019, 75, 1063–1070. [Google Scholar] [CrossRef]
- Tan, Y.Z.; Rubinstein, J.L. Through-Grid Wicking Enables High-Speed CryoEM Specimen Preparation. Acta Crystallogr D Struct Biol 2020, 76, 1092–1103. [Google Scholar] [CrossRef]
- Ashtiani, D.; De Marco, A.; Neild, A. Tailoring Surface Acoustic Wave Atomisation for Cryo-Electron Microscopy Sample Preparation. Lab Chip 2019, 19, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- White, H.D.; Thirumurugan, K.; Walker, M.L.; Trinick, J. A Second Generation Apparatus for Time-Resolved Electron Cryo-Microscopy Using Stepper Motors and Electrospray. Journal of Structural Biology 2003, 144, 246–252. [Google Scholar] [CrossRef]
- Ravelli, R.B.G.; Nijpels, F.J.T.; Henderikx, R.J.M.; Weissenberger, G.; Thewessem, S.; Gijsbers, A.; Beulen, B.W.A.M.M.; López-Iglesias, C.; Peters, P.J. Cryo-EM Structures from Sub-Nl Volumes Using Pin-Printing and Jet Vitrification. Nat Commun 2020, 11, 2563. [Google Scholar] [CrossRef]
- Rima, L.; Zimmermann, M.; Fränkl, A.; Clairfeuille, T.; Lauer, M.; Engel, A.; Engel, H.-A.; Braun, T. CryoWriter: A Blotting Free Cryo-EM Preparation System with a Climate Jet and Cover-Slip Injector. Faraday Discuss. 2022, 240, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Darrow, M.C.; Moore, J.P.; Walker, R.J.; Doering, K.; King, R.S. Chameleon: Next Generation Sample Preparation for CryoEM Based on Spotiton. Microsc Microanal 2019, 25, 994–995. [Google Scholar] [CrossRef]
- Dandey, V.P.; Wei, H.; Zhang, Z.; Tan, Y.Z.; Acharya, P.; Eng, E.T.; Rice, W.J.; Kahn, P.A.; Potter, C.S.; Carragher, B. Spotiton: New Features and Applications. Journal of Structural Biology 2018, 202, 161–169. [Google Scholar] [CrossRef]
- Wei, H.; Dandey, V.P.; Zhang, Z.; Raczkowski, A.; Rice, W.J.; Carragher, B.; Potter, C.S. Optimizing “Self-Wicking” Nanowire Grids. Journal of Structural Biology 2018, 202, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Levitz, T.S.; Weckener, M.; Fong, I.; Naismith, J.H.; Drennan, C.L.; Brignole, E.J.; Clare, D.K.; Darrow, M.C. Approaches to Using the Chameleon: Robust, Automated, Fast-Plunge CryoEM Specimen Preparation. Front. Mol. Biosci. 2022, 9, 903148. [Google Scholar] [CrossRef]
- Vilella, P. Étude Des Mécanismes Moléculaires Régulant l’activité Des Complexes Formés Par Le Récepteur Nucléaire ERRa; University of Strasbourg, 2023. [Google Scholar]
- Russo, C.J.; Passmore, L.A. Progress towards an Optimal Specimen Support for Electron Cryomicroscopy. Current Opinion in Structural Biology 2016, 37, 81–89. [Google Scholar] [CrossRef]
- Russo, C.J.; Passmore, L.A. Ultrastable Gold Substrates for Electron Cryomicroscopy. Science 2014, 346, 1377–1380. [Google Scholar] [CrossRef]
- Russo, C.J.; Passmore, L.A. Ultrastable Gold Substrates: Properties of a Support for High-Resolution Electron Cryomicroscopy of Biological Specimens. Journal of Structural Biology 2016, 193, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Quispe, J.; Damiano, J.; Mick, S.E.; Nackashi, D.P.; Fellmann, D.; Ajero, T.G.; Carragher, B.; Potter, C.S. An Improved Holey Carbon Film for Cryo-Electron Microscopy. Microsc Microanal 2007, 13, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-J.; Hyun, J.-K.; Kim, J.-G.; Jeong, H.S.; Park, H.N.; You, D.-J.; Jung, H.S. Measurement of Ice Thickness on Vitreous Ice Embedded Cryo-EM Grids: Investigation of Optimizing Condition for Visualizing Macromolecules. J Anal Sci Technol 2013, 4, 7. [Google Scholar] [CrossRef]
- Park, J.; Park, J.; Lee, S.; Roh, S.-H. Grid Selection Strategy for High-Resolution Cryo-EM. Korean Soc Struct Biol 2020, 8, 41–48. [Google Scholar] [CrossRef]
- Naydenova, K.; Jia, P.; Russo, C.J. Cryo-EM with Sub–1 Å Specimen Movement. Science 2020, 370, 223–226. [Google Scholar] [CrossRef]
- Naydenova, K.; Peet, M.J.; Russo, C.J. Multifunctional Graphene Supports for Electron Cryomicroscopy. Proc. Natl. Acad. Sci. U.S.A. 2019, 116, 11718–11724. [Google Scholar] [CrossRef]
- Guo, F.; Jiang, W. Single Particle Cryo-Electron Microscopy and 3-D Reconstruction of Viruses. In Electron Microscopy; Kuo, J., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, 2014; ISBN 978-1-62703-775-4. [Google Scholar]
- Mahler, F.; Meister, A.; Vargas, C.; Durand, G.; Keller, S. Self-Assembly of Protein-Containing Lipid-Bilayer Nanodiscs from Small-Molecule Amphiphiles. Small 2021, 17, 2103603. [Google Scholar] [CrossRef]
- Kelly, D.F.; Abeyrathne, P.D.; Dukovski, D.; Walz, T. The Affinity Grid: A Pre-Fabricated EM Grid for Monolayer Purification. Journal of Molecular Biology 2008, 382, 423–433. [Google Scholar] [CrossRef]
- Llaguno, M.C.; Xu, H.; Shi, L.; Huang, N.; Zhang, H.; Liu, Q.; Jiang, Q.-X. Chemically Functionalized Carbon Films for Single Molecule Imaging. Journal of Structural Biology 2014, 185, 405–417. [Google Scholar] [CrossRef]
- Han, B.-G.; Watson, Z.; Kang, H.; Pulk, A.; Downing, K.H.; Cate, J.; Glaeser, R.M. Long Shelf-Life Streptavidin Support-Films Suitable for Electron Microscopy of Biological Macromolecules. Journal of Structural Biology 2016, 195, 238–244. [Google Scholar] [CrossRef]
- Meyerson, J.R.; Rao, P.; Kumar, J.; Chittori, S.; Banerjee, S.; Pierson, J.; Mayer, M.L.; Subramaniam, S. Self-Assembled Monolayers Improve Protein Distribution on Holey Carbon Cryo-EM Supports. Sci Rep 2014, 4, 7084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Shigematsu, H.; Shimizu, T.; Ohto, U. Improving Particle Quality in Cryo-EM Analysis Using a PEGylation Method. Structure 2021, 29, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Number of structures of membrane proteins deposited at Protein Data Bank (PDB) solved by cryo-EM single particle analysis since the first atomic structure of a membrane protein was determined by this technique in 2013. (A) Number of membrane protein structure deposits per year; (B) Number of membrane protein structure deposits per year and per resolution range.
Figure 1.
Number of structures of membrane proteins deposited at Protein Data Bank (PDB) solved by cryo-EM single particle analysis since the first atomic structure of a membrane protein was determined by this technique in 2013. (A) Number of membrane protein structure deposits per year; (B) Number of membrane protein structure deposits per year and per resolution range.
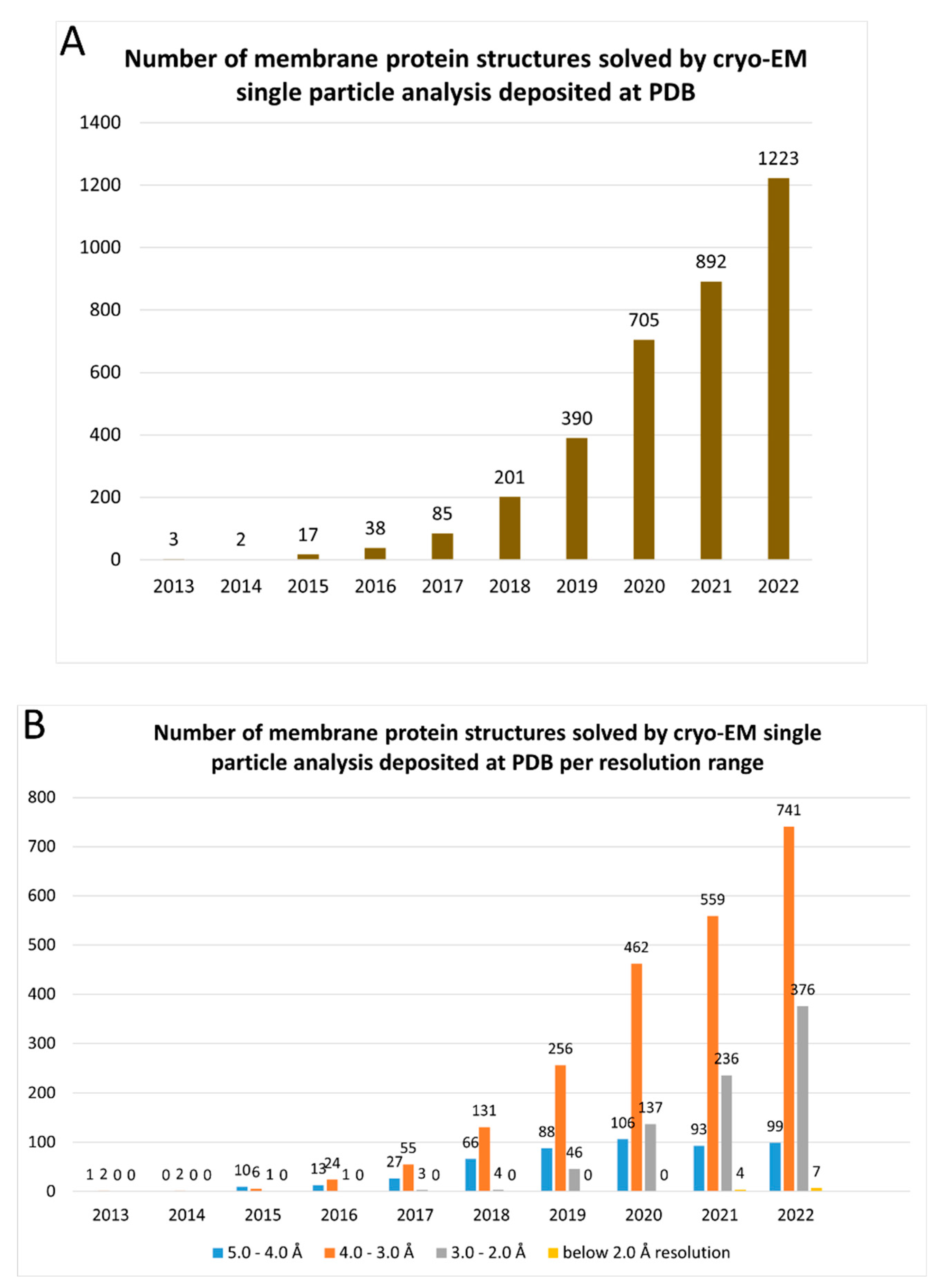
Figure 2.
Profile of detergents and other types of molecules usage during the membrane extraction step of the membrane proteins whose structure has been solved at a resolution better than 3 Å by cryo-EM SPA during the last two years (2021-2022) available at Protein Data Bank. (A) Number of reports found for each surfactant or other molecules used for the protein membrane extraction. Alkyl maltoside detergents include DDM, DM and UDM. MNG family detergents include LMNG, MNG and DMNG. Mixed detergents include association between different detergents. In this class, we noticed the reports of the association between DDM and sodium cholate (4 reports), DDM and LMNG (2 reports), DDM and DM-NPG (2 reports), DDM and GDN (1 report), LMNG and digitonin (2 reports), LMNG and GDN (1 report), LMNG and sodium cholate (1 report), digitonin and LDAO (1 report), DDM and Triton X-100 (1 report). Other detergents include DDM analogue cyclohexyl-α maltoside (t-PCCαM; 3 reports), Cymal-4 (1 report), Cymal-6 (1 report), LDAO (2 reports), Sodium cholate (1 report), OGNG (1 report), C12E9 (1 report), C12E8 (1 report), and glycerol (1 report). (B) Analysis of the addition of cholesteryl hemisuccinate (CHS) on surfactant-solubilized MPs at the five most used detergents used for protein membrane extraction step.
Figure 2.
Profile of detergents and other types of molecules usage during the membrane extraction step of the membrane proteins whose structure has been solved at a resolution better than 3 Å by cryo-EM SPA during the last two years (2021-2022) available at Protein Data Bank. (A) Number of reports found for each surfactant or other molecules used for the protein membrane extraction. Alkyl maltoside detergents include DDM, DM and UDM. MNG family detergents include LMNG, MNG and DMNG. Mixed detergents include association between different detergents. In this class, we noticed the reports of the association between DDM and sodium cholate (4 reports), DDM and LMNG (2 reports), DDM and DM-NPG (2 reports), DDM and GDN (1 report), LMNG and digitonin (2 reports), LMNG and GDN (1 report), LMNG and sodium cholate (1 report), digitonin and LDAO (1 report), DDM and Triton X-100 (1 report). Other detergents include DDM analogue cyclohexyl-α maltoside (t-PCCαM; 3 reports), Cymal-4 (1 report), Cymal-6 (1 report), LDAO (2 reports), Sodium cholate (1 report), OGNG (1 report), C12E9 (1 report), C12E8 (1 report), and glycerol (1 report). (B) Analysis of the addition of cholesteryl hemisuccinate (CHS) on surfactant-solubilized MPs at the five most used detergents used for protein membrane extraction step.
Figure 3.
Chemical structures of the four most common detergents used for membrane protein extraction step of the membrane protein structures solved by cryo-EM single particle analysis in the last two years (2021-2022) available at Protein Data Bank. (A) n-Dodecyl-β-D-maltopyranoside (DDM); (B) Lauryl maltose neopentyl glycol (LMNG); (C) Digitonin; (D) Glyco-diosgenin (GDN).
Figure 3.
Chemical structures of the four most common detergents used for membrane protein extraction step of the membrane protein structures solved by cryo-EM single particle analysis in the last two years (2021-2022) available at Protein Data Bank. (A) n-Dodecyl-β-D-maltopyranoside (DDM); (B) Lauryl maltose neopentyl glycol (LMNG); (C) Digitonin; (D) Glyco-diosgenin (GDN).
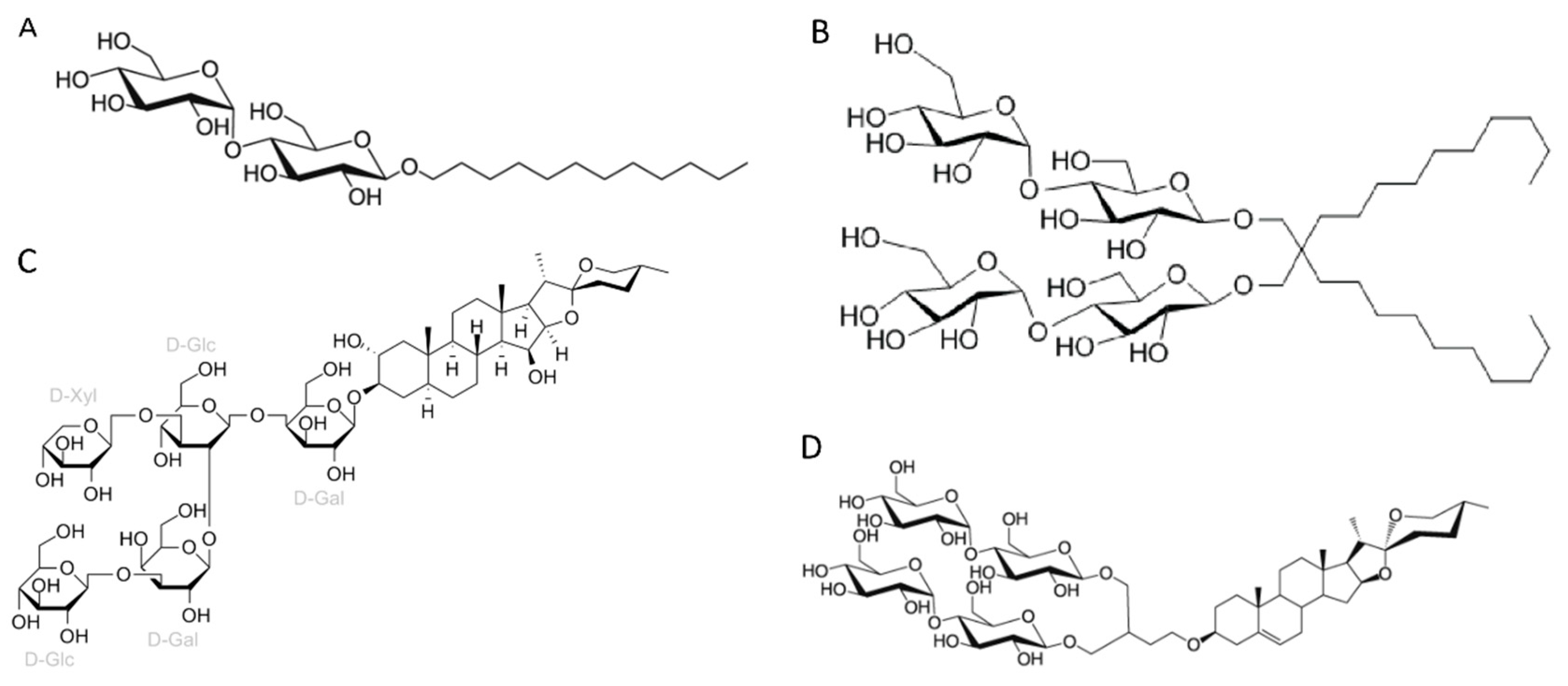
Figure 4.
Profile of the surfactants and other types of molecules usage in the vitrification step of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. A) Number of reports for each surfactant used in the vitrification step. Alkyl maltoside detergents include DDM and DM. MNG family detergents include LMNG and MNG. Mixed detergents include association between different detergents. In this class, we notice the reports of the association between LMNG and GDN (48 reports), LMNG, GDN and digitonin (3 reports), MNG and GDN (2 reports), GDN and digitonin (2 reports), DM-NPG and digitonin (2 reports), DDM and LMNG (1 report), DDM and Cymal-5 (1 report), DDM and Triton X-100 (1 report), DDM and CHAPS (1 report), LMNG and CHAPS (1 report), digitonin and sodium cholate (1 report), digitonin and LDAO (1 report). Nanodiscs include native nanodiscs (SMA copolymer nanodiscs, 4 reports), MSP-nanodiscs (50 reports), circularized nanodiscs (3 reports) and saposin-lipid-protein complexes (Salipro, 4 reports). Other surfactants include: LDAO (2 reports), Cymal-5 (1 report), C12E9 (1 report), glycerol (1 report), and peptidisc (1 report). B) Analysis of the addition of cholesteryl hemisuccinate (CHS) on detergent-solubilized MPs at the vitrification step.
Figure 4.
Profile of the surfactants and other types of molecules usage in the vitrification step of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. A) Number of reports for each surfactant used in the vitrification step. Alkyl maltoside detergents include DDM and DM. MNG family detergents include LMNG and MNG. Mixed detergents include association between different detergents. In this class, we notice the reports of the association between LMNG and GDN (48 reports), LMNG, GDN and digitonin (3 reports), MNG and GDN (2 reports), GDN and digitonin (2 reports), DM-NPG and digitonin (2 reports), DDM and LMNG (1 report), DDM and Cymal-5 (1 report), DDM and Triton X-100 (1 report), DDM and CHAPS (1 report), LMNG and CHAPS (1 report), digitonin and sodium cholate (1 report), digitonin and LDAO (1 report). Nanodiscs include native nanodiscs (SMA copolymer nanodiscs, 4 reports), MSP-nanodiscs (50 reports), circularized nanodiscs (3 reports) and saposin-lipid-protein complexes (Salipro, 4 reports). Other surfactants include: LDAO (2 reports), Cymal-5 (1 report), C12E9 (1 report), glycerol (1 report), and peptidisc (1 report). B) Analysis of the addition of cholesteryl hemisuccinate (CHS) on detergent-solubilized MPs at the vitrification step.
Figure 5.
Profile of the type and composition of nanodiscs present on the vitrification step of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. (A) Profile of the type of nanodiscs used in the vitrification step. Membrane scaffold proteins (MSP)-based nanodiscs: MSP1E3D1, MSP2N2, and MSP1D1. Salipro: saposin-lipid-protein complexes. SMA: Styrene-maleic acid copolymer (native nanodiscs). (B) Profile of the lipid component of MSP-based nanodiscs. E. coli TLE: Total lipid extract of E.coli. SPLE: soybean polar lipid extract. POPC associations include: POPC+POPE+POPG (3 reports), POPC+POPE+cholesterol (1 report), and POPC+POPG (1 report).
Figure 5.
Profile of the type and composition of nanodiscs present on the vitrification step of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. (A) Profile of the type of nanodiscs used in the vitrification step. Membrane scaffold proteins (MSP)-based nanodiscs: MSP1E3D1, MSP2N2, and MSP1D1. Salipro: saposin-lipid-protein complexes. SMA: Styrene-maleic acid copolymer (native nanodiscs). (B) Profile of the lipid component of MSP-based nanodiscs. E. coli TLE: Total lipid extract of E.coli. SPLE: soybean polar lipid extract. POPC associations include: POPC+POPE+POPG (3 reports), POPC+POPE+cholesterol (1 report), and POPC+POPG (1 report).
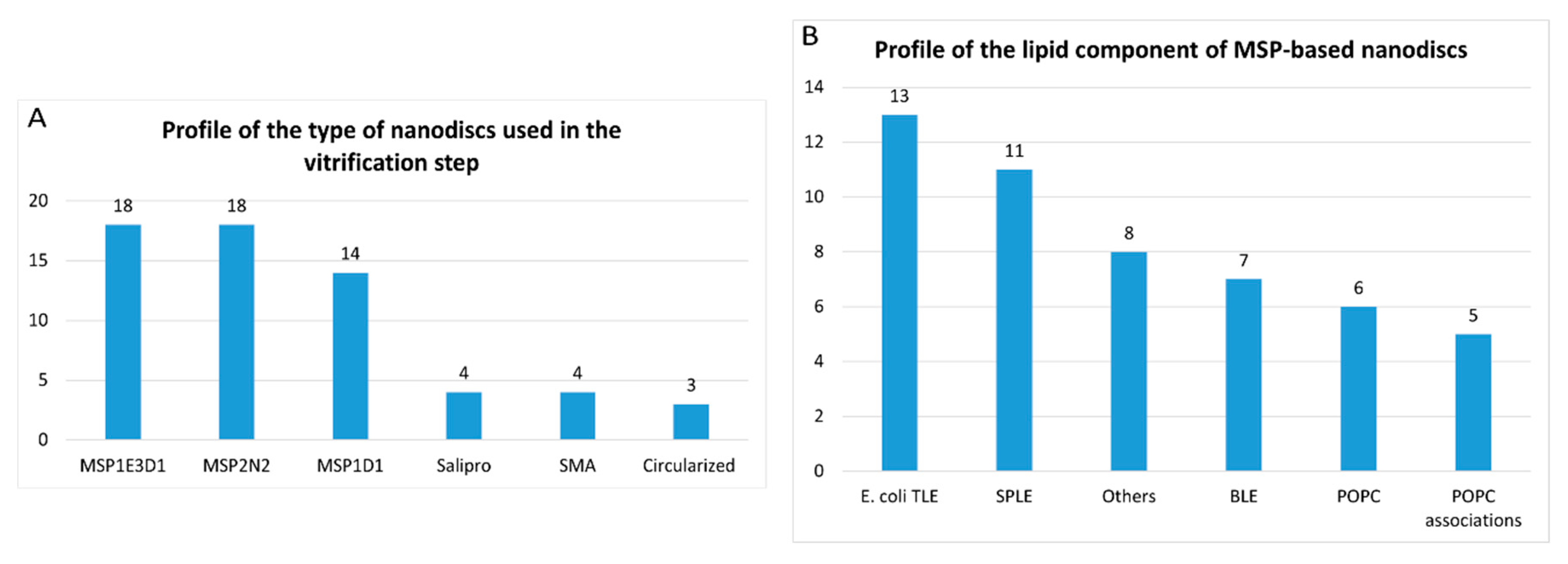
Figure 6.
Profile of detergent substitution between protein membrane extraction and vitrification steps of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. Numbers of DDM (A), LMNG (B), GDN (C) and digitonin (D) substitution between protein membrane extraction and vitrification steps. Mixed detergent reports in panel A include: LMNG and GDN (11 reports), MNG and GDN (1 report), DDM and Cymal-5 (1 report). Mixed detergent reports in panel B include: LMNG and GDN (30 reports) and LMNG, GDN and digitonin (2 reports). Mixed detergent reports in panel D include 1 report of GDN and digitonin.
Figure 6.
Profile of detergent substitution between protein membrane extraction and vitrification steps of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. Numbers of DDM (A), LMNG (B), GDN (C) and digitonin (D) substitution between protein membrane extraction and vitrification steps. Mixed detergent reports in panel A include: LMNG and GDN (11 reports), MNG and GDN (1 report), DDM and Cymal-5 (1 report). Mixed detergent reports in panel B include: LMNG and GDN (30 reports) and LMNG, GDN and digitonin (2 reports). Mixed detergent reports in panel D include 1 report of GDN and digitonin.
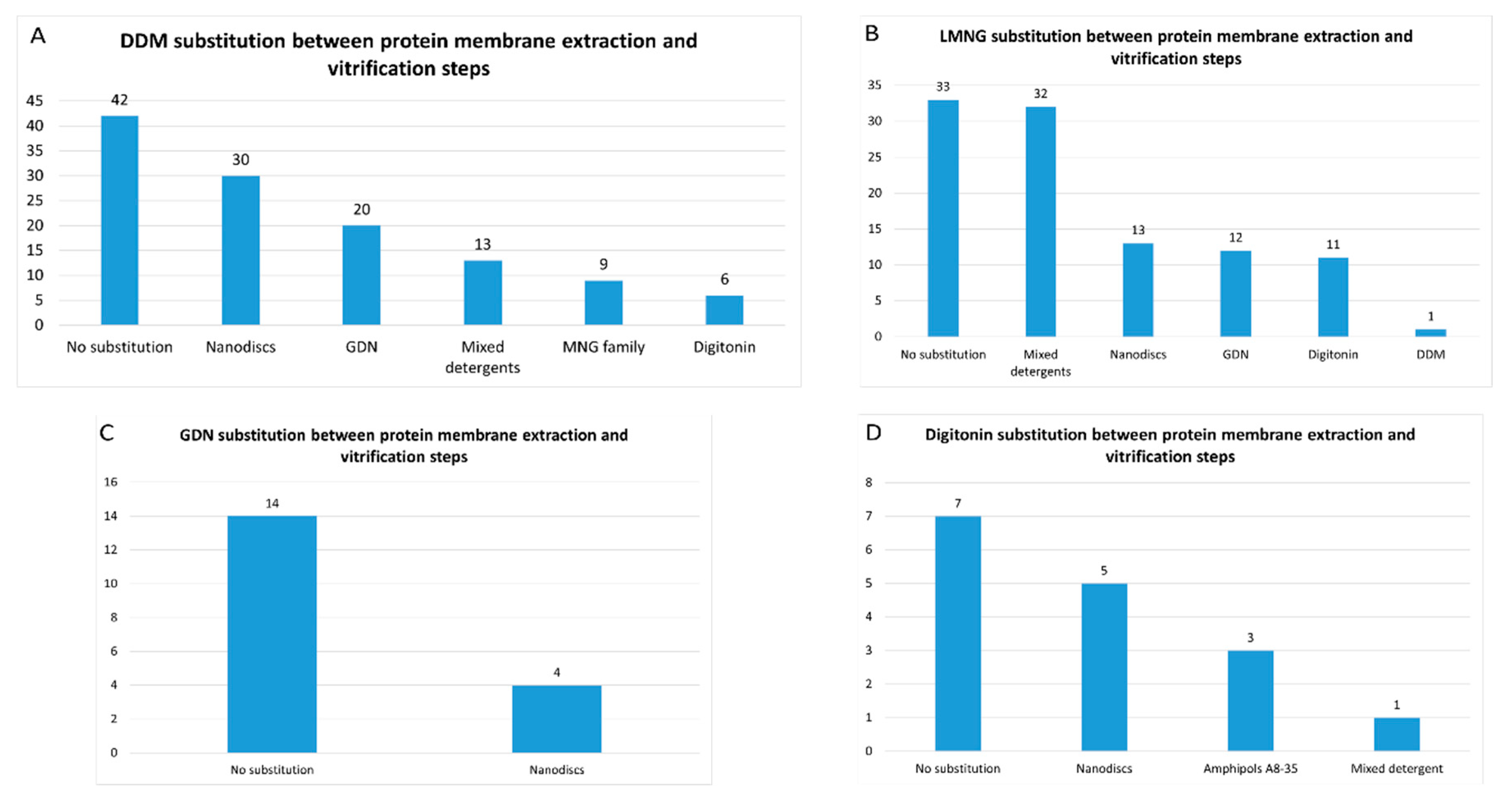
Figure 7.
Profile of the grids used for determination of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. (A) Type of the grids employed; (B) Types of the cooper metal mesh and carbon foil of Quantifoil® grids; (C) Types of the gold metal mesh and carbon foil of Quantifoil® grids; (D) Types of the gold metal mesh and gold foil of UltrAuFoil® grids.
Figure 7.
Profile of the grids used for determination of membrane protein structures solved by cryo-EM single particle analysis below 3.0 Å resolution in the last two years (2021-2022) available at Protein Data Bank. (A) Type of the grids employed; (B) Types of the cooper metal mesh and carbon foil of Quantifoil® grids; (C) Types of the gold metal mesh and carbon foil of Quantifoil® grids; (D) Types of the gold metal mesh and gold foil of UltrAuFoil® grids.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



