
Submitted:
17 August 2023
Posted:
21 August 2023
You are already at the latest version
Abstract
Astragaloside IV (AIV) – triterpenoid saponin from the Astragalus mongholicus roots with the proved acetylcholinesterase (AChE)-inhibitory capability was used as the substance with the potential to attenuate memory impairment. Scopolamine (SCOP), antagonist of muscarinic cholinergic receptors and lipopolysaccharide (LPS), neuroinflammation inducer were used to impair memory processes in the passive avoidance (PA) test in mice. This memory impairment in the SCOP-treated mice was attenuated by the earlier intraperitoneal (ip) administration of AIV in the dose of 25 mg/kg. The reduction of memory impairment by LPS was not observed, therefore AIV does not reverse memory impairments through the anti-inflammatory mechanisms. All the studied doses of AIV did not affect basic locomotor activity of mice. As a result of the post-mortem analysis of mice body tissues, the largest content of AIV was determined in the kidneys, then in the spleen, liver and the smallest one in brain by mass spectrometry.
Keywords:
astragaloside IV
; passive avoidance
; memory impairment
; ADME-Tox
; blood-brain barrier
1. Introduction
Damage in neurons in the human brain may cause a number of neurodegenerative-nature disorders. There are approximately 10 million of new cases of dementia in the world recorded by the World Health Organization each year [1]. It is commonly known that aging is a crucial risk factor for dementia since it leads to the accumulation of various unrepaired cellular damages as well as impairment of the cellular repair, and compensatory mechanisms [2,3]. It is estimated that in 2050 16% of the world’s population will be seniors aged 65 or over. This is an increase of 7 percentage points compared to 2019. The trend of rapidly aging societies worldwide, called the Silver Tsunami, is important for the increase in the number of cases of neurodegenerative diseases associated with cognitive impairment [4]. Neuroprotection is closely related to the rescue, regeneration and preservation of the integrity of both neurons and the neurovascular unit for performing their physiological functions [5].
One of such disorders is the Alzheimer’s disease (AD), multi-factorial and heterogeneous one, characterized by a progressive loss of cognitive functions [6]. Accumulation of protein aggregates in the brain in the form of insoluble amyloid plaques as well as neurofibrillary tangles (NFTs) is the pathological hallmark of AD [7]. Fibrillar amyloid β (Aβ) peptides, which are senile plaques components, are produced by cleavage of the Aβ precursor protein whereas the constituents of NFTs are the paired helical filaments of abnormally phosphorylated microtubule-associated tau protein [6,8]. Currently the amyloid β secretion is believed to be the triggering occurrence in the pathogenesis of AD, while NFTs are considered to be a secondary event in the neurodegeneration [9]. The AD can be caused by mechanisms of neuroinflammation [10] as well as neurotoxicity [11], or induction of oxidative damage [12].
The cholinergic system is involved in many crucial physiological processes, i.e., learning and memory, attention, sleep, stress response, and sensory information [13]. Cholinergic neurons, innervating almost all areas of the brain [14,15] and therefore playing a very important role in the peripheral and central nervous systems, are particularly present in the spinal cord, hindbrain, medial habenula, mesopontine region, basal forebrain, striatum, olfactory tubercle and islands of Cajella complex [13,16,17].
In AD the cholinergic neurons in the central nervous system (CNS) are degenerated which leads to a functional loss [17,18,19]. Acetylcholine (ACh), a neurotransmitter secreted by all the cholinergic neurons, is that related to the memory process [20,21,22,23]. On one hand, the attention deficit can be caused by the injuries of the basal forebrain cholinergic neurons [24,25], whereas the attention improvement in humans can be obtained using the cholinesterase inhibitors facilitating the cholinergic neurotransmission [26,27]. In the treatment of senile dementia of the AD type, there are used the cholinergic receptor agonists (muscarinic and nicotinic) as well as the inhibitors of acetylcholine metabolizing enzyme, or synthesis promoters which enhance the endogenous level of ACh [28]. However, the most clinically successful approach is the use of acetylcholinesterase inhibitors [28]. The search for novel acetylcholinesterase inhibitors, also from the plant kingdom, which could be possible alternatives to the currently used therapeutics, is an important issue in the treatment of AD. One of the plants with the potential to improve cognitive functions, including memory and learning, is Astragalus mongholicus.
In our previous papers, the IC50 values using TLC-bioautography assay [29] as well as the pharmacokinetic parameters of the blood-brain barrier (BBB) permeation for AI-AIV using computational analysis [30] were obtained. Additionally, the logBB value (0.49 ± 0.03) of astragaloside IV was measured experimentally in the post-mortem studies on the brain tissues and plasma samples of mice. Based on the BBB-pharmacokinetic descriptors i.e., logBB (BBB-penetration descriptor), logPS (logarithmic permeability–surface area product), log(PSFubrain) (brain/plasma equilibration rate), Fu (fraction unbound in plasma), and Fb (fraction unbound in the brain), it was revealed that astragaloside IV can cross the BBB and can have a neuroactive potential.
Moreover, our previous paper [29] showed using the TLC-bioautography method that astragaloside IV is a potent AChE inhibitor. For further intensification of our previous findings, in the current paper the astragaloside IV was used as a substance with the potential to attenuate memory impairment taking advantage of passive avoidance test in mice. The astragaloside IV effect itself as well as that in mice with the experimentally induced memory-impairments were assessed. These studies were preceded by the analysis of the ADME-Tox parameters. For this purpose the passive human jejunum absorption, the Caco-2 absorption, the behaviour towards P-glycoprotein as well as the toxicity risk were determined in silico.
To characterize the passive absorption through the human intestinal epithelium, both the Caco-2 and jejunum models were used. The Caco-2 cell monolayer, being the human colon epithelial cancer cell line, is an excellent model of the passive transcellular pathway [31]. The tight junctions between the cells, which are formed during the Caco-2 cells differentiation, serve as a model of paracellular movement of compounds across the intestinal monolayer [32]. Therefore, it can mimic the human intestinal epithelium and can be compared to the extensively folded human jejunum [31]. Additionally, to assess the accumulation of AIV in the selected internal organs, there was performed the LC-MS analysis of kidneys, spleen, liver and brain mice tissues after the intraperitoneal administration of the substance.
2. Results
2.1. The ADME-Tox studies
In our previous paper [30] the promising results with regard to the astragalosides potential actions were presented. Therefore, we decided to carry on this research and extend it by including the in vivo tests on mice and the assessment of astragaloside IV accumulation on the other individual organs and tissues using both the computational and post-mortem analyses.
The ADME-Tox parameters calculated in silico are presented in Table 1 and Table S1 in the Supplementary Material section.
2.2. The influence of astragaloside IV on the acquisition and consolidation of long-term memory impairment induced by the SCOP administration in mice
Two-way ANOVA indicated the differences between the tested groups on animals both for acquisition [F(7,77) = 196.3, p < 0.001; Figure 1A) and consolidation [F(7,77) = 112.2, p < 0.001; Figure 1B] of long-term memory impaired by the SCOP administration. SCOP affected both memory acquisition and consolidation compared to the control mice (p < 0.001, Figure 1A,B). This memory impairment in the SCOP-treated mice was attenuated by the earlier administration of astragaloside IV but only in the largest dose i.e., 25 mg/kg, ip (acquisition: p < 0.001 and consolidation: p < 0.01; Figure 1A,B). The smaller doses were ineffective (p > 0.05). Interestingly, astragaloside IV itself in all tested doses improved acquisition (p < 0.001; Figure 1A) but not consolidation of long-term memory (p > 0.05; Figure 1B).
2.3. Influence of astragaloside IV on the acquisition and consolidation of long-term memory impairment induced by the LSP administration in mice
Two-way ANOVA indicated the differences between the tested groups on animals both for acquisition [F(4,55) = 17.65, p < 0.001; Figure 2A) and consolidation [F(4,55) = 17.69, p < 0.001; Figure 2B] of long-term memory impaired by the LSP administration. LSP affected both memory acquisition and consolidation compared to the control mice (p < 0.001, Figure 2A,B). In all tested doses astragaloside IV did not attenuate memory impaired by LPS in both the acquisition and consolidation phases (p > 0.05; Figure 2A,B) compared to the only LPS-treated mice.
2.4. Locomotor activity
One-way ANOVA indicated that astragaloside IV, in all tested doses, did not affect the locomotor activity of mice in the 30 min long measurement [F(3,44) = 7.57, p > 0.05] (Figure 3).
2.5. Post-mortem analysis of astragaloside IV distribution in soft organs
The analyses of the soft organs obtained from the experimental mice were performed using the HPLC-ESI-QTOF-MS/MS platform in the tailored chromatographic method. The results of these studies are presented in Figure 4 and Table 2. Moreover, the total ion chromatogram of liver extract is presented in Figure 5.
3. Discussion
Analyzing the values in Table 1, it can be stated that all the tested astragalosides showed poor or moderate passive intestinal absorption. The smallest passive jejunum as well as Caco-2 permeability [cm/s] is shown by both astragalosides III and IV. This can be due to a larger number of H-donor groups in comparison to astragalosides I and II and/or the relatively lowest lipophilicity of the analytes. However, there are no significant differences in the behaviour of compounds with respect to these two different models. This means that the same compounds, namely astragalosides III and IV, show the smallest permeability to both Caco-2 and jejunum models whereas astragalosides I and II show the largest absorption. This could suggest that both absorption models are appropriate for studying the human intestinal absorption of the tested saponins. This is in line with the hypothesis that only the villi tips which are a fraction of the anatomical surface of the intestine, participate in the absorption process [33]. It is generally assumed that the incompletely and slowly passively absorbed substances are transported across the intestinal epithelium via the paracellular pathway through the water-filled pores. In the drug research, it is assumed that the passage of molecules across the cellular barriers increases with the increase of the compound lipophilicity. However, there were observed some differences for the intestinal absorption when too high lipophilicity of a compound can result in the small intestinal absorption [34]. On the other hand, it is also possible that even very hydrophilic compounds can be transported through the intestinal barrier mainly by the transcellular route [35]. It is also assumed that the optimal transepithelial passage of drugs takes place when the n-octanol/water partition coefficient being the lipophilicity descriptor, is about 3000 [34]. In our case, the logPow values of astragalosides III and IV are in this optimal range, while those of astragalosides I and II are much higher. Furthermore, it should be remembered that before the compounds are absorbed, they will remain longer in the intestinal lumen. Therefore, in comparison with the compounds highly permeable through the intestinal endothelium which are rapidly and completely absorbed, they can diffuse further down the length of the villi [36].
The P-glycoprotein belonging to the ATP-binding cassette transport proteins plays a significant role as an efflux pump by translocating several endogenous and exogenous substances towards the extracellular compartment [37]. This protein having a mass 170 kD is produced by the multidrug resistance-1 gene. Moreover, it is a major component of the BBB which can be essential for the brain detoxification as well as protection against xenobiotics [6]. Expression of P-gp can lead to the reduced permeability of the BBB [38], which results in the brain protection against toxic substances. However, this effect can be also involved in the reduced effectiveness in a treatment of neurodegenerative diseases. The drugs which are P-gp substrates can reduce dramatically the permeability through the BBB [6], therefore the therapeutic effect cannot be achieved.
The in silico studies have estimated also the probability of astragalosides for being the P-gp substrates. In our case, astragalosides I and II are likely to be the P-gp substrates whereas III and IV are not. This may be due to the number of H-acceptors, which determines mainly the classification of the compound to the P-gp substrates [39]. However, as results from Table 1, the differences between these values are not significant. In addition, other physicochemical descriptors characterizing a molecule determine whether or not it is a P-gp substrate. Such parameters also include molecule size expressed as the molar weight or the volume as well as ionization given by the acid and base pKa values [39]. No significant differences were observed also in these parameters values. On the other hand, taking into account the lipophilic logPow parameter and the polarizability of the molecule, astragalosides I and II were found to be the most lipophilic and to possess higher topological polar surface area values compared to astragalosides III and IV. Therefore, the prediction of P-gp substrate specificity is a difficult task due to the various factors characterizing molecules. However, taking into account the risk assessment of adverse side effects obtained for astragalosides using the online OSIRIS Property Explorer (4/28/23), recommended by the U.S. Food and Drug Administration, no mutagenic, tumorigenic, irritating and reproductive adverse effects were found. This indicates that all the tested compounds can be safe for human health.
Herein, SCOP was used to impair memory processes in the PA test in mice. Scopolamine is a great affinity antagonist of muscarinic cholinergic receptors which can cross the BBB readily [40] and causes the cognitive impairment in humans, rodents, and non-human primates [41]. Blockade of muscarinic receptors by SCOP mimics the age-related degeneration of cholinergic neurons in the basal forebrain [42,43,44,45], therefore SCOP is extensively used in vivo as a model of cognitive impairment [46,47,48,49].
In our study, pretreatment with astragaloside IV in the largest dose reversed the SCOP-induced memory impairment in the PA task in mice. In our previous studies [29] it was shown that astragaloside IV is an inhibitor of AChE, the enzyme responsible for the degradation of ACh in the synaptic cleft [50]. Taking this into account, one can conclude that astragaloside IV, at least partially, reduce memory impairment through muscarinic receptors, increasing the affinity of ACh for muscarinic and probably also for the nicotinic cholinergic receptors. It can be presumed to work in the same way as the drugs used for Alzheimer’s disease, i.e., donepezil, rivastigmine, and galantamine – all being acetylcholinesterase inhibitors [51,52].
In this study, astragaloside IV pretreatment did not reverse the effect of the widely used neuroinflammation model [53] – lipopolysaccharide at any given dose. Therefore, it can be stated that it does not reverse memory impairments through the anti-inflamatory mechanisms.
Astragaloside IV was detected in the positive ionisation mode in all the biological samples as a signal with m/z of 817.4570 Da. At this measured m/z value the mass error was only 1.24 ppm. The compound was characterised by the double bond equivalent value of 8. The post-mortem analysis of the mice tissues is a part of ADME-Tox profiling. The concentration of astragaloside IV in the soft organs tissues was dose dependant (see Figure 4). Interestingly, the largest content of the saponin was determined in the kidneys, then in the spleen, liver and brain. At the dose of 25 mg/kg, the largest concentration of the substance was observed in the kidneys, while with the smaller doses, the largest concentration was found in the spleen.
The large concentration of AIV in the mice spleen tissues can indicate that this saponin can affect the functioning of the organ. The spleen being the peripheral immune organ is involved in the production of immune cells that act as immune effectors, including macrophages, monocytes, T cells, B cells, and neutrophils [54]. Numerous scientific studies prove that astragaloside IV can both control the immune cells such as macrophages, natural killer cells or lymphocytes and affect the production of diverse cytokines and chemokines [55]. In addition, AIV has also been shown to be able to alleviate polymicrobial sepsis and pathological spleen damage probably due to the inhibition of inflammatory response and apoptosis of lymphocytes [56].
Analyzing the astragaloside IV concentration in the mice kidneys one can conclude that the elimination of astragaloside IV can take place along with urine. However, the study by Zhang et al. [57] shows that AIV can be also slowly eliminated by the hepatic clearance. The smallest quantity of astragaloside IV was found in brain. The applied method did not detect the saponin in the mice treated with 6 mg/kg as the measured value was too low to provide meaningful data. In our previous paper [30] the concentration of AIV in the brain tissues was measured to calculate the logBB value expressed as the logarithmic ratio between the concentration of a compound in brain and blood. Calculation of this value is important mostly in the early estimation of BBB permeability in the drug design process. Taking into account the experimental value (0.49 ± 0.03) it can be stated that astragalaside can cross the blood-brain barrier, however, this passage can be impeded and slowed down. This can be due to the molecular weight of the compound of 785 g/mol and/or the topological polar surface area (TPSA) of 228.22 Ų (data calculated using the ACD/Percepta software). Besides lipophilicity, the TPSA is extremely important for the drugs ability to cross the BBB [58,59]. It is reported that for the orally bioavailable drugs the TPSA should be less than 120 Ų [60]. It is also recognized that the CNS drugs characterized by the good brain penetration have PSA<100Å2 or even smaller, <60–70 Å2 [60]. Nevertheless, the ability of astragaloside IV to cross the blood-brain barrier in the post-mortem studies, which we have proven [30], does not contradict the values of the above-mentioned physicochemical parameters. There is also the other theory that relates the BBB permeability to the molecular flexibility [61,62] of a compound which can be expressed e.g., by the number of rotatable bonds [63]. According to this theory, the more flexible molecules of a comparable molecular weight seem to fit to the membrane better than the rigid ones [62].
4. Materials and Methods
4.1. The ADME-Toxicity profiling
The ADME-Tox studies were carried out on the triterpenoid saponins from the Astragalus mongholicus roots (AI-AIV) with the highest proven neuroprotective potential [30]. To evaluate ADME-Tox properties, some physicochemical parameters of AI-AIV were calculated in silico using the ACD/Percepta software. There were calculated the following values: the logarithm of n-octanol/water partition coefficient (logPow), the human jejunum absorption, the Caco-2 absorption, the number of H-donors and H-acceptors, as well as the behaviour towards P-glycoprotein. Moreover, the toxicity risk i.e., mutagenic, tumorigenic, irritant, reproductive effectiveness was estimated using Osiris Property Explorer online.
4.2. Animal studies
4.2.1. Mice
Male adult Swiss mice (weight 25-35 g) were housed in the Experimental Medicine Centre, Medical University of Lublin, Poland. Mice were kept in groups of 5 per cage under the standard environmental conditions (12:12 h light:dark, room temperature 21 ± 1 °C). Animals had ad libitum access to tap water and mice chow (Agropol, Motycz, Poland). They were accustomed to the laboratory conditions for at least one week before the experiments. Each experimental group consisted of 12 animals. Throughout the experiments, all attempts were made to minimize animal suffering and to reduce a number of animals. The behavioural experiments were conducted regularly between 8:00 and 15:00, according to the National Institute of Health Guidelines for the Care and Use of Laboratory, and also based on the European Community Council Directive for the Care and Use of laboratory animals of 22 September 2010 (2010/63/EU). The tests were approved by the Ethics Committee at the University of Life Sciences in Lublin, Poland (No. 45/2018). Each mouse was used only once in the experiment.
4.2.2. Drugs
Astragaloside IV (6, 12.5 or 25 mg/kg), isolated from the extract of Astragalus mongholicus roots [30], was transferred to a mortar and dissolved in the physiological saline (0.9% NaCl) with the addition of 0.2% dimethylsulfoxide (DMSO). Scopolamine (Sigma Aldrich, St. Louis, USA; 1 mg/kg) and lipopolysaccharide (Sigma Aldrich, St. Louis, USA; 2.5 mg/kg) were dissolved in 0.9% NaCl (saline). The control groups were given injections of saline with the addition of 0.2% DMSO. All compounds were administered intraperitoneally at a volume of 10 mL/kg. Fresh drug solutions were prepared each day of the experimentation.
4.2.3. The passive avoidance test
The passive avoidance test in mice is used to assess the fear-related learning and memory [64,65]. The apparatus is made of acrylic box with two chambers: white (10 × 13 × 15 cm) and black (25 × 20 × 15 cm). During the experiment, the white chamber is illuminated with a bright light (8 W). The black chamber, equipped with an electric grid floor, is connected to the lit chamber.
Pre-test (training) was carried out prior to the experiments. To do so, each mouse was individually placed into the lit up chamber of the apparatus and allowed to explore it within 30 s. After that, the guillotine door which separated both chambers, was opened. The mouse was then allowed to enter the black chamber where they were immediately treated with the electric shock (0.2 mA) for 2 s. The latency for entering the black chamber was registered (TL1). During the retention trial, conducted 24 h after the pre-test, the same mouse was placed again in the lit chamber of apparatus. The time necessary for entering the black chamber was registered (TL2). Contrary to the pre-test, no foot-shock was provided this time. If the mouse did not enter the black chamber within 300 s, TL2 was registered as 300 s. The index of latency (IL) was calculated as follows:
IL=(TL2-TL1)/TL1
- Acquisition of long-term memory
Astragaloside IV (6, 12.5 or 25 mg/kg, ip) was injected 15 min before the administration of SCOP or LPS. After the 30 min delay, the pre-test was carried out.
- 2.
- Consolidation of long-term memory
Astragaloside IV (6, 12.5 or 25 mg/kg, ip) was injected immediately after the pre-test while SCOP or LPS 30 min later.
4.2.4. The locomotor activity test
The locomotor activity of mice was measured with the photo resistor actimeters (circular cages, 25 cm diameter, two light beams) kept in the sound-attenuated experimental room. Before the test, the mice were acclimatized to the new environment for at least one week. The animals were placed individually in the actimeters for 30 min. Two photocell beams, located across the axis, measured beam breaks automatically. During the experiment, animals were allocated into the following drug groups: the control group and the astragaloside IV (6, 12.5 or 25 mg/kg, ip) one. Immediately after the injection, animals were placed in the actimeters.
4.2.5. Statistical analysis
The behavioural data were analyzed by the one-way analysis of variance (ANOVA), with the Tukey’s post hoc test. The GraphPad Prism 9.3.1 (San Diego, USA) software was used for statistical purposes. The p-value < 0.05 was set as statistically significant. The data are depicted as individual measurements, mean ± standard error of the mean (SEM).
4.3. HPLC-ESI-QTOF-MS/MS determination of astragaloside IV in the animal tissues
The mice were decapitated 0.5 h after the locomotor activity test was finished (i.e., 1 h after astragaloside IV injection) and the brains, livers, kidneys, and spleens were collected, deep frozen, and kept for the quantitative analysis. Then, they were homogenized in the plastic Eppendorff vial kept in the ice. The homogenates of the organs were further vortexed for 10 min with 200 µL of 50% ethanol. The Eppendorf vials were further centrifuged at 3500 RPM for 5 min and the supernatants were filtered through a nylon syringe filter with the diameter of 0.22 µm to the autosampler vials with the glass inserts. Astragaloside IV was dissolved in 50% ethanol and transformed into 10 different concentrations in the range of 0.002–0.2 mg/mL to obtain the calibration curve. Due to the small sizes and masses of brains, AIV was quantified in these organs after the addition of the reference compound (Sigma Aldrich, St. Louis, MO, USA) to the samples. For this purpose, 100 µL of 0.5 mg/mL AIV was introduced to each brain sample. The peak areas of astragaloside IV in the brains were compared to the blank solution of the standard compound. The contents of astragaloside IV in the remaining organs were measured without the presence of the reference compound.
The detection of astragaloside IV in the mice brains, spleens, livers, and kidneys was performed using the liquid chromatograph (1200 Series) with the mass detector (6500 Series), namely the HPLC-ESI-QTOF-MS/MS instrument, that was produced by Agilent Technologies (Santa Clara, CA, USA). The liquid chromatograph was composed of a degasser, binary pump, column oven, autosampler, and the high resolution mass detector QTOF-MS/MS with an electrospray ionization source. The Zorbax Eclipse Plus chromatographic column (Agilent Technologies, CA, USA) (150 mm x 2.1. mm, 3.2 um, RP-18) was used to separate the metabolites of the injected matrix in the following gradient of acetonitrile with 0.1 % formic acid (solvent B) in the 0.1 % aqueous solution of formic acid (solvent A): 0 min—2 % B, 2 min—10 % B, 6 min—40 % B, 35-37 min—95 % B, 37.5 min—2 % B. The analysis time was set at 43 min, the post run—at 5 min, the flow rate at 0.2 mL/ min and the injection volume at 10 μL. The detailed settings of the mass spectrometer were as follows: the capillary voltage of 3000 V, the skimmer voltage of 65 V, the mass range of 100-1200 m/z, the collision energy of 10 and 20 V, the nebuliser pressure of 30 psig, the gas flow rate of 12 L/min, the gas and sheath gas temperatures were 250 and 325 oC. The Mass Hunter Workstation (version B.10.00) by Agilent Technologies was used to record and handle the obtained data. For the quantitative analysis the calibration curve equation of astragaloside IV was prepared (8 points) from the triple injections in the range between 0.0001 and 0.1 mg/mL (n = 3). For each of the concentrations, the average values of the content of the tested substance in the given organ were calculated. The standard deviation (SD) values were calculated in each case.
5. Conclusions
Memory impairment caused by the intraperitoneal administration of SCOP was attenuated by the AIV dose of 25 mg/kg. The AIV doses of 6 and 12.5 mg/kg did not affect acquisition and consolidation of long-term memory impaired by the SCOP in mice. None of astragaloside IV doses attenuated memory impairments induced by the LPS administration. Therefore, astragaloside IV did not reverse LPS-induced memory impairments through the anti-inflammatory mechanisms. The results of the in silico ADME-Tox studies on astragaloside IV show a probable safety of its use. The largest content of the saponin was determined in the kidneys, then in the spleen, liver and the smallest one in brain. Astragaloside IV seems to be an efficient drug candidate for the treatment of CNS diseases with memory impairment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Risk assessment of adverse side effects by OSIRIS Property Explorer of the tested astragalosides.
Author Contributions
Conceptualisation, K.S.; methodology, K.S., W.K.-K., A.B.-C. and K.G.; validation, K.S., W.K.-K., A.B.-C. and K.G.; formal analysis, K.S. and W.K.-K.; investigation, K.S., W.K.-K., A.B.-C. and K.G.; resources, K.S. and W.K.-K.; data curation, K.S.; writing—original draft preparation, K.S.; supervision, K.S.; project administration, K.S.; funding acquisition, K.S. and W.K.-K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by DS448, and DS24, Medical University of Lublin and Maria Curie-Skłodowska University in Lublin.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
AIV—astragaloside IV; ACh—acetylcholine; AChE—acetylcholinesterase; AD—Alzheimer’s disease; ADME-Tox—absorption-distribution-metabolism-excretion-toxicity; Aβ—amyloid β; BBB—blood-brain barrier; CNS—central nervous system; DMSO—dimethylsulfoxide; HPLC-MS—High Performance Liquid Chromatography coupled with mass spectrometry; IL—index of latency; ip—intraperitoneally; LPS—lipopolysaccharide; NFTs—neurofibrillary tangles; PA—passive avoidance; P-gp—P-glycoprotein; SCOP—scopolamine; SD—standard deviation; SEM—standard error of the mean; TPSA—topological polar surface area.
References
- World Health Organization, Global action plan on the public health response to dementia 2017–2025. Available online: https://www.who.int/publications/i/item/global-action-plan-on-the-public-health-response-to-dementia-2017---2025 (accessed on 10 April 2023).
- Hullinger, R.; Puglielli, L. Molecular and cellular aspects of age-related cognitive decline and Alzheimer’s disease. Behav. Brain Res. 2017, 322, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Kluever, V.; Fornasiero, E.F. Principles of brain aging: status and challenges of modeling human molecular changes in mice. Ageing Res. Rev. 2021, 72, 101465. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Phan, Ch.W.; Lim, S.J.; Babji, A.S. Insights on the molecular mechanism of neuroprotection exerted by edible bird’s nest and its bioactive constituents. Food Sci. Hum. Wellness 2023, 12, 1008–1019. [Google Scholar] [CrossRef]
- Farooqui, A.A. Contribution of neuroinflammation, resolution, and neuroprotection in neurotraumatic diseases. Farooqui, A.A., Ed.; In Neuroinflammation, Resolution, and Neuroprotection in the Brain; Elsevier: Amsterdam, 2022; pp. 83–119. [Google Scholar]
- Rapposelli, S.; Digiacomo, M.; Balsamo, A. P-gp Transporter and its Role in Neurodegenerative Diseases. Curr. Top. Med. Chem. 2009, 9, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Elufioye, T.O.; Berida, T.I.; Habtemariam, S. Plants-Derived Neuroprotective Agents: Cutting the Cycle of Cell Death through Multiple Mechanisms. Evid. Based Complement. Alternat. Med. 2017, 3574012. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s Disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, P.V.; Marzloff, K.; Hyman, B.T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer’s disease. Neurology 1992, 42, 1681–1688. [Google Scholar] [CrossRef]
- Mrak, R.E.; Griffin, W.S.T. Interleukin-1, neuroinflammation, and Alzheimer’s disease. Neurobiol. Aging. 2001, 22, 903–908. [Google Scholar] [CrossRef]
- Cole, G.; Teter, B.; Frautschy, S. Neuroprotective effects of curcumin. Adv. Exp. Med. Biol. 2007, 595, 595–197. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Dodwell, S.A.; Borrelli, L.A.; Raju, S.; Bacskai, B. In vivo reduction of plaque size in APPswe/PS1D9 mice treated with curcumin. Alzheimer’s & Dementia 2006, 2, S617. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; Khachaturian, Z.S. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain. 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Houser, C.R.; Crawford, G.D.; Barber, R.P.; Salvaterra, P.M.; Vaughn, J.E. Organization and morphological characteristics of cholinergic neurons: an immunocytochemical study with a monoclonal antibody to choline acetyltransferase. Brain Res. 1983, 266, 97–119. [Google Scholar] [CrossRef] [PubMed]
- Woolf, N.J.; Butcher, L.L. Cholinergic systems mediate action from movement to higher consciousness. Behav. Brain Res. 2011, 221, 488–498. [Google Scholar] [CrossRef]
- Woolf, N.J. Cholinergic systems in mammalian brain and spinal cord. Prog. Neurobiol. 1991, 37, 475–524. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef]
- Pepeu, G.; Giovannini, M.G. The fate of the brain cholinergic neurons in neurodegenerative diseases. Brain Res. 2017, 1670, 173–184. [Google Scholar] [CrossRef]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef]
- Dunnett, S.B.; Everitt, B.J.; Robbins, T.W. The basal forebraincortical cholinergic system: interpreting the functional consequences of excitotoxic lesions. Trends Neurosci. 1991, 14, 494–501. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Anderson, B.P.; Bower, J.M. Cholinergic modulation of cortical associative memory function. J. Neurophysiol. 1992, 67, 1230–1246. [Google Scholar] [CrossRef]
- Lucas-Meunier, E.; Fossier, P.; Baux, G.; Amar, M. Cholinergic modulation of the cortical neuronal network. Pflugers Arch. 2003, 446, 17–29. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P. Cognitive functions of cortical acetylcholine: toward a unifying hypothesis. Brain Res. Rev. 1997, 23, 28–46. [Google Scholar] [CrossRef]
- Bucci, D.J.; Holland, P.C.; Gallagher, M. Removal of cholinergic input to rat posterior parietal cortex disrupts incremental processing of conditioned stimuli. J. Neurosci. 1998, 18, 8038–8046. [Google Scholar] [CrossRef]
- Voytko, M.L.; Olton, D.S.; Richardson, R.T.; Gorman, L.K.; Tobin, J.R.; Price, D.L. Basal forebrain lesions in monkeys disrupt attention but not learning and memory. J. Neurosci. 1994, 14, 167–186. [Google Scholar] [CrossRef]
- Pepeu, G.; Giovannini, M.G.; Bracco, L. Effect of cholinesterase inhibitors on attention. Chem. Biol. Interact. 2013, 203, 361–364. [Google Scholar] [CrossRef]
- Bracco, L.; Bessi, V.; Padiglioni, S.; Marini, S.; Pepeu, G. Do cholinesterase inhibitors act primarily on attention deficit? A naturalistic study in Alzheimer’s disease patients. J. Alzheimers Dis. 2014, 40, 737–742. [Google Scholar] [CrossRef]
- Kim, D.H.; Jeon, S.J.; Jung, J.W.; Lee, S.; Yoon, B.H.; Shin, B.Y.; Son, K.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; Ko, K.H.; Ryu, J.H. Tanshinone congeners improve memory impairments induced by scopolamine on passive avoidance tasks in mice. Eur. J. Pharmacol. 2007, 574, 140–147. [Google Scholar] [CrossRef]
- Stępnik, K.; Kukula-Koch, W.; Plazinski, W.; Gawel, K.; Gaweł-Bęben, K.; Khurelbat, D.; Boguszewska-Czubara, A. Significance of Astragaloside IV from the Roots of Astragalus mongholicus as an Acetylcholinesterase Inhibitor-From the Computational and Biomimetic Analyses to the In Vitro and In Vivo Studies of Safety. Int J Mol Sci. 2023, 24, 9152. [Google Scholar] [CrossRef]
- Stępnik, K.; Kukula-Koch, W. In Silico Studies on Triterpenoid Saponins Permeation through the Blood-Brain Barrier Combined with Postmortem Research on the Brain Tissues of Mice Affected by Astragaloside IV Administration. Int. J. Mol. Sci. 2020, 21, 2534. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 1, 175–185. [Google Scholar] [CrossRef] [PubMed]
- DiMarco, R.L.; Hunt, D.R.; Dewi, R.E.; Heilshorn, S.C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials. 2017, 129, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Wilson, F.A.; Dietschy, J.M. The intestinal unstirred layer: Its surface area and effect on active transport kinetics. Biochim. Biophys. Acta. 1974, 363, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Wils, P.; Warnery, A.; Phung-Ba, V.; Legrain, S.; Scherman, D. High lipophilicity decreases drug transport across intestinal epithelial cells. J. Pharmacol. Exp. Ther. 1994, 269, 654–658. [Google Scholar] [PubMed]
- Artursson, P.; Palm, K.; Luthman, K. Caco-2 monolayers in experimental and theoretical predictions of drug transport. Adv. Drug Deliv. Rev. 1996, 22, 67–84. [Google Scholar] [CrossRef]
- New, R. Oral Delivery of Biologics via the Intestine. Pharmaceutics. 2020, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Weickert, C.S.; Garner, B. Role of ATP-binding cassette transporters in brain lipid transport and neurological disease. J. Neurochem. 2008, 104, 1145–1166. [Google Scholar] [CrossRef]
- Tatsuta, T.; Naito, M.; Oh-hara, T.; Sugawara, I.; Tsuruo, T. Functional involvement of P-glycoprotein in blood-brain barrier. J. Biol. Chem. 1992, 267, 20383–20391. [Google Scholar] [CrossRef]
- Didziapetris, R.; Japertas, P.; Avdeef, A.; Petrauskas, A. Classification Analysis of P-Glycoprotein Substrate Specificity. J. Drug Target. 2003, 11, 391–406. [Google Scholar] [CrossRef]
- Lochner, M.; Thompson, A.J. The muscarinic antagonists scopolamine and atropine are competitive antagonists at 5-HT3 receptors. Neuropharmacol. 2016, 108, 220–228. [Google Scholar] [CrossRef]
- Misane, I.; Ögren, S. Selective 5-HT1A Antagonists WAY 100635 and NAD-299 Attenuate the Impairment of Passive Avoidance Caused by Scopolamine in the Rat. Neuropsychopharmacol. 2003, 28, 253–264. [Google Scholar] [CrossRef]
- Araujo, D.M.; Lapchak, P.A.; Robitaille, Y.; Gauthier, S.; Quirion, R. Differential alteration of various cholinergic markers in cortical and subcortical regions of human brain in Alzheimer’s disease. J. Neurochem. 1988, 50, 1914–1923. [Google Scholar] [CrossRef]
- DeKosky, S.T.; Scheff, S.W.; Styren, S.D. Structural correlates of cognition in dementia: quantification and assessment of synapse change. Neurodegener. 1996, 5, 417–421. [Google Scholar] [CrossRef]
- Kuhl, D.E.; Koeppe, R.A.; Minoshima, S.; Snyder, S.E.; Ficaro, E.P.; Foster, N.L. In vivo mapping of cerebral acetylcholinesterase activity in aging and Alzheimer’s disease. Neurology 1999, 52, 691–699. [Google Scholar] [CrossRef]
- Shinotoh, H.; Namba, H.; Fukushi, K.; Nagatsuka, S.; Tanaka, N.; Aotsuka, A. Progressive loss of cortical acetylcholinesterase activity in association with cognitive decline in Alzheimer’s disease: a positron emission tomography study. Ann. Neurol. 2000, 48, 194–200. [Google Scholar] [CrossRef]
- Bartus, R.T. On neurodegenerative diseases, models, and treatment strategies: lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp. Neurol. 2000, 163, 495–529. [Google Scholar] [CrossRef] [PubMed]
- Bartolomeo, A.C.; Morris, H.; Buccafusco, J.J.; Kille, N.; Rosenzweig-Lipson, S.; Husbands, M.G.; Sabb, A.L.; Abou-Gharbia, M.; Moyer, J.A.; Boast, C.A. The preclinical pharmacological profile of WAY-132983, a potent M1 preferring agonist. J. Pharmacol. Exp. Ther. 2000, 292, 584–596. [Google Scholar] [PubMed]
- Elrod, K.; Buccafusco, J.J. An evaluation of the mechanism of scopolamine-induced impairment in two passive avoidance protocols. Pharmacol. Biochem. Behav. 1988, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Blin, O.; Audebert, C.; Pitel, S.; Kaladjian, A.; Casse-Perrot, C.; Zaim, M.; Micallef, J.; Tisne-Versailles, J.; Sokoloff, P.; Chopin, P.; Marien, M. Effects of dimethylaminoethanol pyroglutamate (DMAE p-Glu) against memory deficits induced by scopolamine: evidence from preclinical and clinical studies. Psychopharmacol. Berl. 2009, 207, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Blotnick-Rubin, E.; Anglister, L. Fine Localization of Acetylcholinesterase in the Synaptic Cleft of the Vertebrate Neuromuscular Junction. Front. Mol. Neurosci. 2018, 11, 123. [Google Scholar] [CrossRef]
- Gawel, K.; Labuz, K.; Gibula-Bruzda, E.; Jenda, M.; Marszalek-Grabska, M.; Silberring, J.; Kotlinska, J.H. Acquisition and reinstatement of ethanol-induced conditioned place preference in rats: Effects of the cholinesterase inhibitors donepezil and rivastigmine. J Psychopharmacol. 2016, 30, 676–687. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula-Bruzda, E.; Dziedzic, M.; Jenda-Wojtanowska, M.; Marszalek-Grabska, M.; Silberring, J.; Kotlinska, J.H. Cholinergic activation affects the acute and chronic antinociceptive effects of morphine. Physiol. Behav. 2017, 169, 22–32. [Google Scholar] [CrossRef]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Yao, J.; Liu, J.; He, Y.; Liu, L.; Xu, Z.; Lin, X.; Liu, N.; Kai, G. Systems pharmacology reveals the mechanism of Astragaloside IV in improving immune activity on cyclophosphamide-induced immunosuppressed mice. J. Ethnopharmacol. 2023, 313, 116533. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Jiang, H.; Tian, Y.; Zhao, W.; Wu, X. Astragaloside IV protects against polymicrobial sepsis through inhibiting inflammatory response and apoptosis of lymphocytes. J. Surg. Res. 2016, 200, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W-D; Zhang, Ch.; Liu, R-H.; Li, H-L.; Zhang, J-T.; Mao, Ch.; Moran, S.; Chen, Ch-L. Preclinical pharmacokinetics and tissue distribution of a natural cardioprotective agent astragaloside IV in rats and dogs. Life Sci. 2006, 79, 808–815. [CrossRef] [PubMed]
- Feher, M.; Sourial, E.; Schmidt, J.M. A simple model for the prediction of blood-brain partitioning. Int. J. Pharm. 2000, 201, 239–247. [Google Scholar] [CrossRef]
- Liu, R.; Sun, H.; So, S–S. Development of quantitative structure − property relationship models for early ADME evaluation in drug discovery. 2. Blood-brain barrier penetration. J Chem. Inf. Comput. Sci. 2001, 41, 1623–1632. [Google Scholar] [CrossRef]
- Van de Waterbeemd, H. In Silico Models to Predict Oral Absorption. Taylor, J.B., Triggle, D.J. Eds. In Comprehensive Medicinal Chemistry II.; Elsevier: Amsterdam, 2007; pp. 669–697. [Google Scholar]
- Fischer, H.; Gottschlich, R.; Seelig, A. Blood-brain barrier permeation: molecular parameters governing passive diffusion. J. Membr. Biol. 1998, 165, 201–211. [Google Scholar] [CrossRef]
- Muehlbacher, M.; Spitzer, G.M.; Liedl, K.R.; Kornhuber, J. Qualitative prediction of blood-brain barrier permeability on a large and refined dataset. J. Comput. Aided Mol. Des. 2011, 25, 1095–1106. [Google Scholar] [CrossRef]
- Hutter, M.C. Prediction of blood-brain barrier permeation using quantum chemically derived information. J. Comput. Aided Mol. Des 2003, 17, 415–433. [Google Scholar] [CrossRef]
- Kotlinska, J.H.; Lopatynska-Mazurek, M.; Gawel, K.; Gabka, P.; Jenda-Wojtanowska, M.; Kruk-Slomka, M.; Marszalek-Grabska, M.; Danilczuk, Z.; Kedzierska, E.; Talarek, S.; Listos, J.; Gibula-Tarlowska, E. Impact of the metabotropic glutamate receptor7 (mGlu7) allosteric agonist, AMN082, on fear learning and memory and anxiety-like behavior. Eur. J. Pharmacol. 2019, 858, 172512. [Google Scholar] [CrossRef] [PubMed]
- Marszalek-Grabska, M.; Zakrocka, I.; Budzynska, B.; Marciniak, S.; Kaszubska, K.; Lemieszek, M.K.; Winiarczyk, S.; Kotlinska, J.H.; Rzeski, W.; Turski, W.A. Binge-like mephedrone treatment induces memory impairment concomitant with brain kynurenic acid reduction in mice. Toxicol. Appl. Pharmacol. 2022, 454, 116216. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the acquisition (A) and consolidation (B) of long-term memory impairment induced by SCOP (1 mg/kg, ip) administration in mice.The data are shown as individual measurements, mean ± SEM (n = 12 per group). ***p<0.001 vs control, ^^^p<0.001, ^^p<0.01 vs SCOP. AIV- astragaloside IV, SCOP-scopolamine.
Figure 1.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the acquisition (A) and consolidation (B) of long-term memory impairment induced by SCOP (1 mg/kg, ip) administration in mice.The data are shown as individual measurements, mean ± SEM (n = 12 per group). ***p<0.001 vs control, ^^^p<0.001, ^^p<0.01 vs SCOP. AIV- astragaloside IV, SCOP-scopolamine.
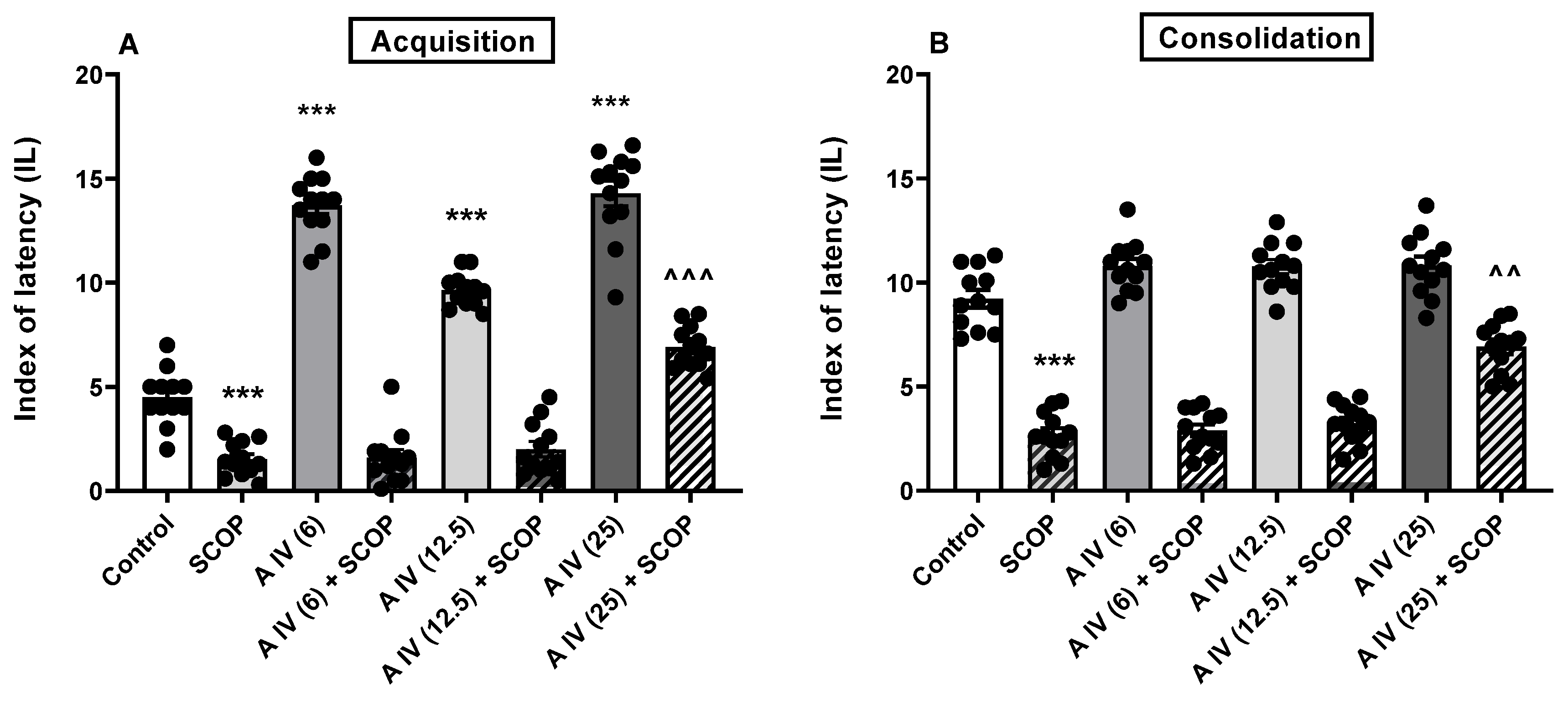
Figure 2.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the acquisition (A) and consolidation (B) of the long-term memory impairment induced by LPS (2.5 mg/kg, ip) administration in mice. The data are shown as individual measurements, mean ± SEM (n = 12 per group). ***p<0.001 vs control. AIV- astragaloside IV, LPS-lipopolysaccharide.
Figure 2.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the acquisition (A) and consolidation (B) of the long-term memory impairment induced by LPS (2.5 mg/kg, ip) administration in mice. The data are shown as individual measurements, mean ± SEM (n = 12 per group). ***p<0.001 vs control. AIV- astragaloside IV, LPS-lipopolysaccharide.
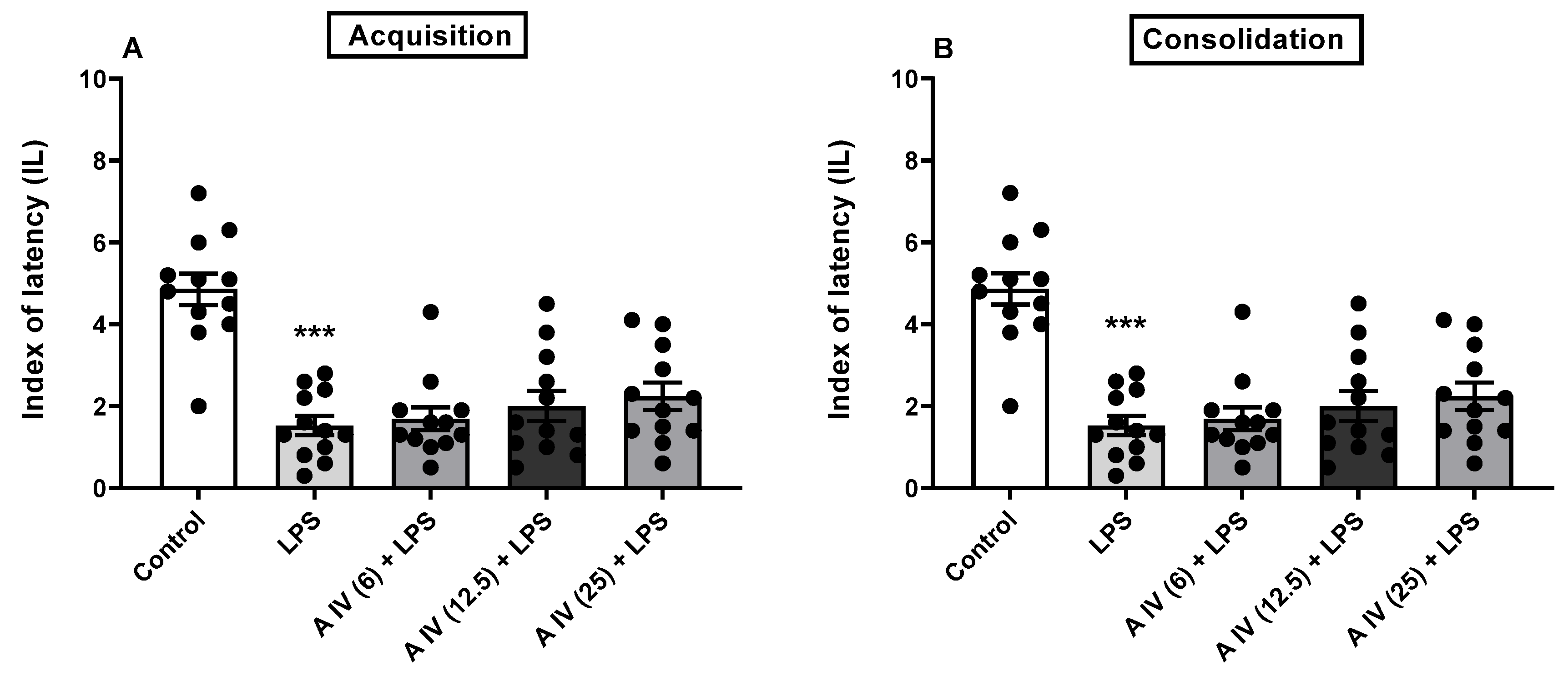
Figure 3.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the locomotor activity. The data are shown as the individual measurements, mean ± SEM. The locomotor activity was measured as the photocell beam breaks for 30 min. AIV- astragaloside IV.
Figure 3.
Influence of astragaloside IV (6, 12.5 or 25 mg/kg, ip) on the locomotor activity. The data are shown as the individual measurements, mean ± SEM. The locomotor activity was measured as the photocell beam breaks for 30 min. AIV- astragaloside IV.
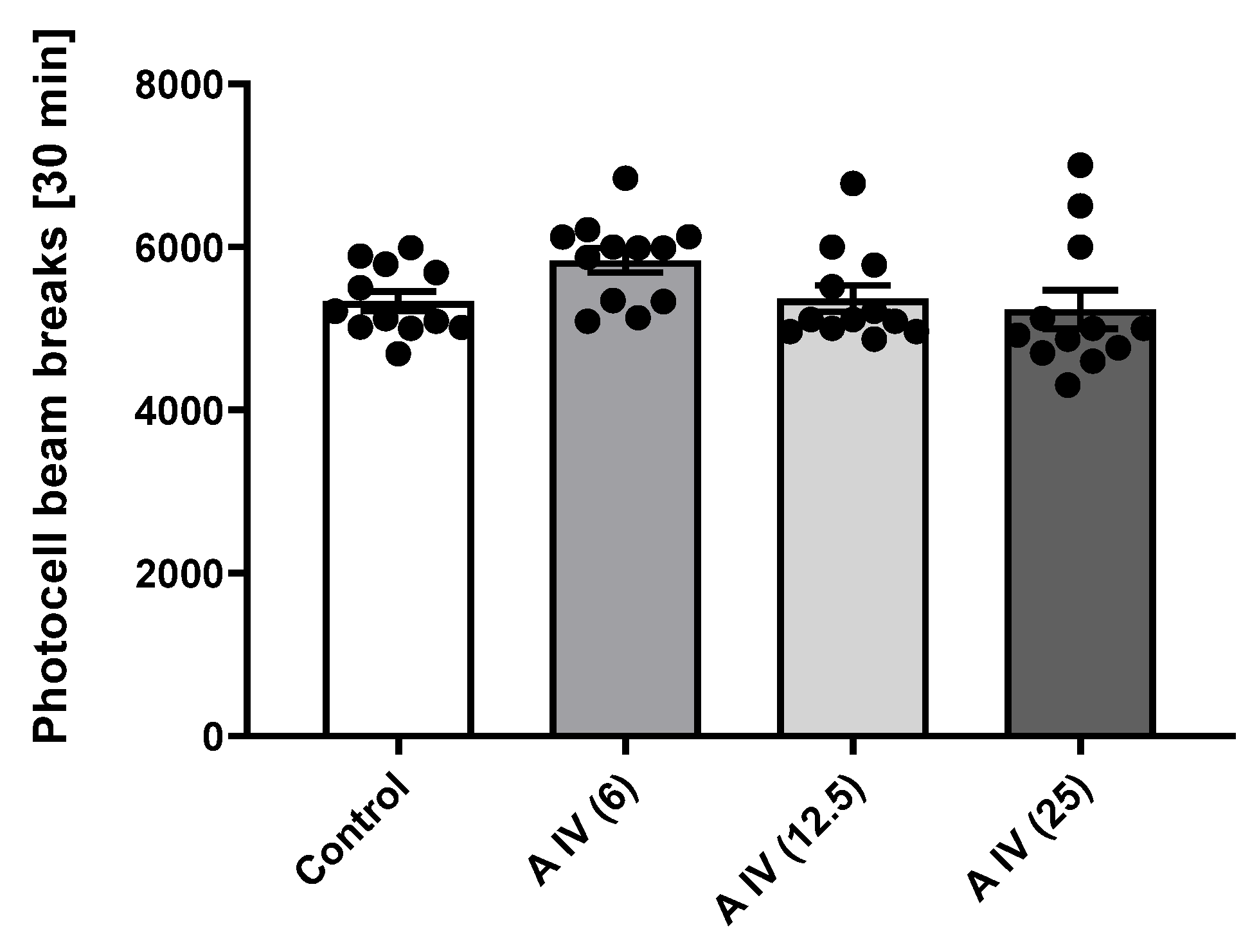
Figure 4.
Concentration of astragaloside IV (%/organ) after intraperitoneal injection of various concentrations of the compound.
Figure 4.
Concentration of astragaloside IV (%/organ) after intraperitoneal injection of various concentrations of the compound.
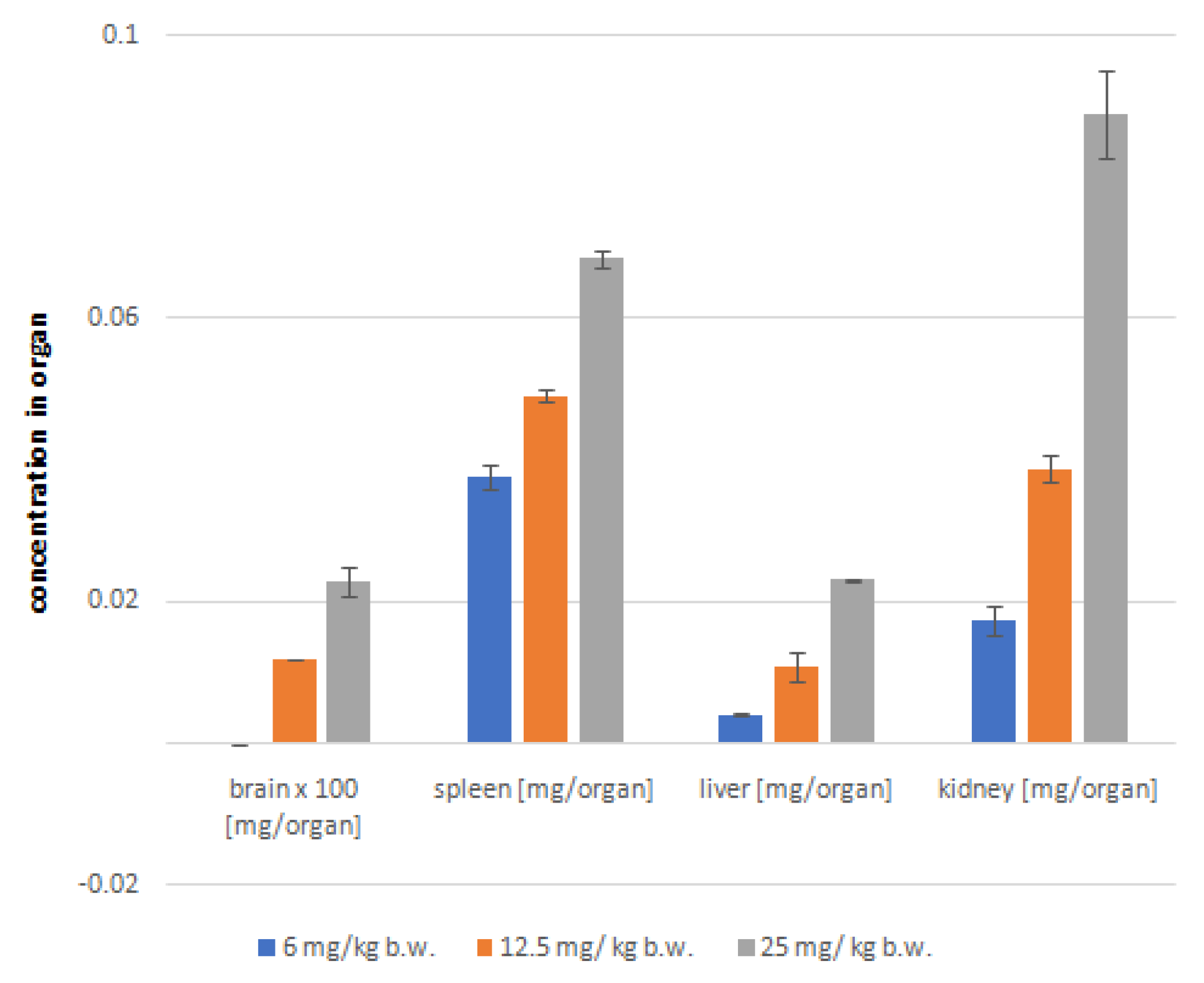
Figure 5.
The sample total ion chromatogram of liver extract recorded in the positive ionisation mode (A) together with the extracted ion chromatogram of astragaloside IV (11.6 min) (B) and the MS/MS chromatogram of the compound (C).
Figure 5.
The sample total ion chromatogram of liver extract recorded in the positive ionisation mode (A) together with the extracted ion chromatogram of astragaloside IV (11.6 min) (B) and the MS/MS chromatogram of the compound (C).
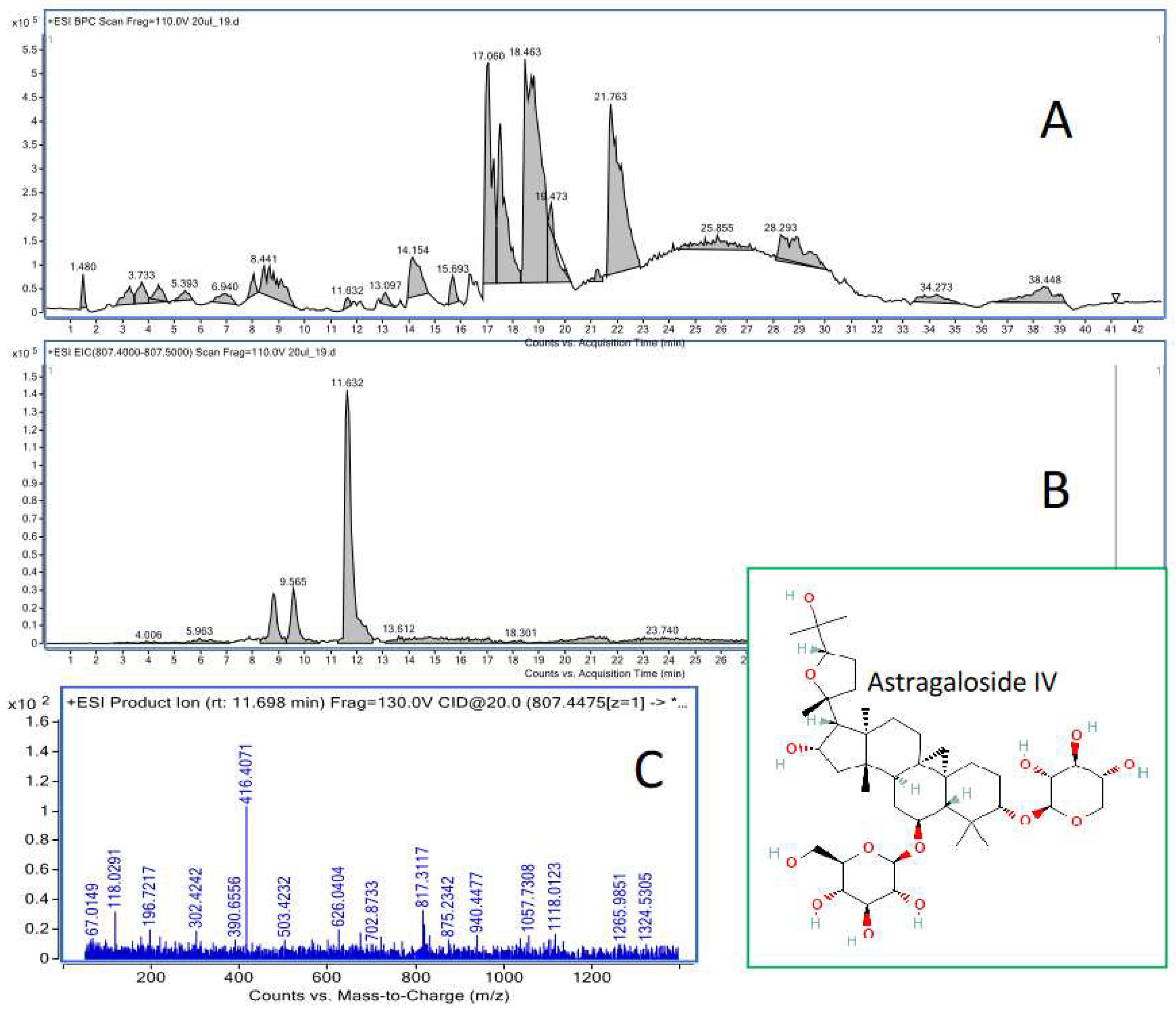

Table 1.
The logarithm of n-octanol/water partition coefficient (logPow), the logarithm of the brain/blood partition coefficient (logBB), the passive human jejunum absorption (Pe,jejunum), the Caco-2 absorption (Pe,Caco2), the number of H-donors, the number of H-acceptors, the P-glycoprotein substrate: (+) high probability (-) low probability of being P-gp substrate, calculated in silico using the ACD/Percepta software.
Table 1.
The logarithm of n-octanol/water partition coefficient (logPow), the logarithm of the brain/blood partition coefficient (logBB), the passive human jejunum absorption (Pe,jejunum), the Caco-2 absorption (Pe,Caco2), the number of H-donors, the number of H-acceptors, the P-glycoprotein substrate: (+) high probability (-) low probability of being P-gp substrate, calculated in silico using the ACD/Percepta software.
| Name | logPow | logBB | Pe,jejunum [10−4cm/s] | Pe,Caco-2 [10−6 cm/s] | Number of H-donors | Number of H-acceptors | P-gp substrate |
|---|---|---|---|---|---|---|---|
| AI | 5.020 | 0.46 | 1.05 | 2.0 | 7 | 16 | + |
| AII | 4.459 | 0.11 | 0.25 | 0.5 | 8 | 15 | + |
| AIII | 3.767 | 0.15 | 0.04 | 0.1 | 9 | 14 | - |
| AIV | 3.757 | 0.49* | 0.06 | 0.1 | 9 | 14 | - |
Table 2.
Content of astragaloside IV in the tested organs of mice regarding the administration dose (ip). (Av—the average value, SD—the standard deviation, n = 3).
Table 2.
Content of astragaloside IV in the tested organs of mice regarding the administration dose (ip). (Av—the average value, SD—the standard deviation, n = 3).
| Brain [mg/organ] |
Spleen [mg/organ] |
Liver [mg/organ] |
Kidney [mg/organ] |
|||||
|---|---|---|---|---|---|---|---|---|
| Animal group | Av | SD | Av | SD | Av | SD | Av | SD |
| 6 mg/kg | ND | ND | 0.03766 | 0.00186 | 0.00411 | 0.00011 | 0.01740 | 0.00209 |
| 12.5 mg/kg | 0.00012 | 0.00001 | 0.04919 | 0.00093 | 0.01084 | 0.00023 | 0.03882 | 0.00092 |
| 25 mg/kg | 0.00023 | 0.00002 | 0.06844 | 0.00133 | 0.02316 | 0.00016 | 0.08892 | 0.00613 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



