
Submitted:
20 August 2023
Posted:
22 August 2023
You are already at the latest version
Abstract
Polycystic ovary syndrome (PCOS) is a common endocrinopathy of reproductive-aged women characterized by hyperandrogenism, oligo-anovulation and insulin resistance closely linked with preferential abdominal fat accumulation. As an ancestral primate trait, PCOS was likely further selected in humans when scarcity of food in hunter-gatherers of the late Pleistocene additionally programmed for enhanced fat storage to meet the metabolic demands of reproduction in later life. As an evolutional model for PCOS, healthy normal-weight women with hyperandrogenic PCOS have subcutaneous (SC) abdominal adipose stem cells that favor fat storage through exaggerated lipid accumulation during development to adipocytes in vitro. In turn, fat storage is counterbalanced by reduced insulin sensitivity and preferential accumulation of highly-lipolytic intra-abdominal fat in vivo. This metabolic adaptation in PCOS balances energy storage with glucose availability and fatty acid oxidation for optimal energy use during reproduction; its accompanying oligo-anovulation allowed PCOS women from antiquity sufficient time and strength for childrearing of fewer offspring with a greater likelihood of childhood survival. Heritable PCOS characteristics are now affected by today’s contemporary environment through epigenetic events that predispose to lipotoxicity with excess weight gain and pregnancy complications, calling for an emphasis on preventive healthcare to optimize the long-term, endocrine-metabolic health of PCOS women in today’s obesogenic environment.
Keywords:
polycystic ovary syndrome
; hyperandrogenism
; insulin resistance
; adipocyte
; adipose stem cells
; evolution
; body fat distribution
; metabolic adaptation
1. Introduction
As the most common endocrinopathy of reproductive-aged women, polycystic ovary syndrome (PCOS) is characterized by hyperandrogenism, oligo-anovulation and insulin resistance closely linked with preferential abdominal fat accumulation [1]. Its clinical manifestations of hirsutism, menstrual irregularity, glucose intolerance and dyslipidemia worsen with obesity to increase the risks of developing subfertility, diabetes, metabolic syndrome and/or cardiovascular disease [2]. Almost one-half of women with PCOS in the United States have metabolic syndrome (i.e., increased abdominal [android] obesity, hyperglycemia, dyslipidemia and/or hypertension), with a prevalence higher than that of age-matched normal women in this country [1,3] and of PCOS women in other countries where obesity is less common [4,5].
Through an evolutionary perspective, the high worldwide prevalence of PCOS in today’s environment should have disappeared over millennia unless a beneficial effect favored both survival and reproduction [6]. Perhaps not surprisingly, therefore, ancestral traits resembling PCOS have been reported throughout antiquity [7] and in a nonhuman primate (i.e., female rhesus macaques) [8,9,10], that shares a common ancestor with humans [11]. One explanation is that an ancient female primate trait resembling PCOS may have been favored originally in the cooling, increasingly arid and less forested African environments of the Oligocene before ancestors of humans diverged from those of macaques [12,13], as the isolated continent of Africa contacted Euroasia [14], enabling inter-continental migration [15] (Figure 1). Such an ancestral trait may have been additionally favored in human hunter-gatherers of the late Pleistocene, or in more ancient human populations, when scarcity of food further selected for programming of enhanced fat storage to meet the metabolic demands of reproduction in later life (i.e., metabolic thrift) [7,16,17,18]. Parallel evolution in macaques, particularly rhesus macaques living in semi-desert and high-altitude environments [13], may have emulated selection in humans for programming of enhanced fat storage (Figure 1). Such evolutionary metabolic adaptations in female primates, including women, would complement the ancient sympathoadrenal response to stress, whereby altered glucocorticoid and catecholamine activities mobilize hepatic glucose and FFAs from visceral fat to act in concert with insulin resistance and ensure sufficient energy during a “fight or flight” response for survival [19,20,21].
Through this evolutionary perspective, the present review examines PCOS as an ancient metabolic adaptation that underwent additional selection pressure for survival of humans during ancient times of food deprivation, but now predisposes to metabolic-endocrine-reproductive dysfunction in today’s obesogenic environment [17,18,22]. A parallel obesogenic environmental change experienced by female rhesus macaques in their natural habitat [13], as well as by macaques removed from their natural habitat decades ago and housed in United States National Primate Research Centers (NPRC) [23,24], may emulate the current obesogenic environmental challenge confronting humans (Figure 1). Consistent with this notion, approximately 15% of adult female rhesus macaques at the Wisconsin NPRC are naturally hyperandrogenic and exhibit PCOS-like traits [9,10]. Polycystic ovarian syndrome and PCOS-like phenotypes may thus form a continuum of ancient primate traits. Understanding trait-related molecular mechanisms, including genetic, epigenetic, protein and lipid interactions leading to optimal energy utilization, along with the perspective of providing benefits for survival and reproductive in both humans and rhesus macaques, offers novel insight into more effective clinical management for women with PCOS.
2. Genetics and epigenetics of PCOS
Heritability of PCOS has been established by family and twin studies [25,26,27,28]; the prevalence of PCOS in female first-degree relatives of affected women is 20-40% [25,27,29] with monozygotic versus dizygotic twin studies showing heritability of PCOS as high as 70% [26]. Large genome-wide association studies (GWAS) in cohorts of PCOS women and controls have identified several PCOS susceptible loci in candidate genes involving gonadotropin secretion/action, androgen biosynthesis/gonadal function, insulin action/metabolism and follicle development [1,30,31,32,33,34,35,36,37,38]. Several PCOS candidate genes are shared among women with differing PCOS phenotypes (i.e., Rotterdam, National Institutes of Health [NIH] criteria, or self-reported) [36]. Some, such as thyroid adenoma associated (THADA) and insulin receptor (INSR), are associated with metabolic disorders in PCOS and type 2 diabetes mellitus (T2DM) [39], others with high bioavailable (unbound) circulating T levels [40]. Genetic correlations between PCOS status and components of metabolic syndrome, including childhood obesity, T2DM, and fasting insulin, high-density lipoprotein-cholesterol (HDL-C) as well as triglyceride (TG) levels, further suggest shared genetic and biological origins between these parameters and PCOS [36,38]. That similar PCOS risk genes are expressed in women with PCOS from Chinese and European populations points to ancient human origins of PCOS [37,38], potentially dating back before the migration of humans out of sub-Saharan Africa 300,000-50,000 years ago or earlier [41,42].
Importantly, women with NIH-defined PCOS have two distinct PCOS subtypes with different genetic heterogeneity: one defined as a “reproductive” group (23% of cases), characterized by higher luteinizing hormone (LH) and sex hormone binding globulin (SHBG) levels with relatively low body mass index (BMI) and insulin levels; the other defined as a “metabolic” group (37% of cases), characterized by higher BMI, glucose and insulin levels, with lower SHBG and LH levels [38,43]. These PCOS subtypes may differ in their developmental origins [43], with their heritability variably interacting with risk-increasing environmental factors to fully explain its prevalence.
Alternatively, rare variants in DENND1A; a gene encoding a 1009 amino acid protein with a clathrin-binding domain regulating endosome-mediated endocytosis, receptor cycling and calcium-dependent signaling cascades [44,45]; also have been associated with endocrine-metabolic traits in families of daughters with PCOS [46]. A posttranscription form of DENND1A, namely DENND1A.v2, is over-expressed in some PCOS women [47,48], with DENND1A.v2 over-expression in human theca cells increasing androgen biosynthesis/release, potentially by PCOS-candidate gene ZNF217 diminishing theca cell expression of microRNA mIR-130b-3p, a noncoding microRNA transcriptional repressor [49].
Genetic variants of anti-mullerian hormone (AMH) and its type 2 receptor (AMHR2) also have been identified in about 7% of women with PCOS by NIH criteria, with 37 such variants having reduced in vitro bioactivity and diminished AMH inhibition of CYP17A1 as a risk factor for PCOS [50,51]. Both AMH and AMHR2 gene variants regulate intra-ovarian follicle development and hypothalamic GnRH function, and possibly ovarian androgen production [52], and may underlie elevated circulating AMH levels and ovarian hyperandrogenism in PCOS women [51].
Considered together, the current understanding of the genetics of PCOS suggests multiple contributing risk genes within which different variants can contribute to a PCOS phenotype. Given the heterogeneity of PCOS phenotypic expression, the high prevalence of PCOS, and its complex gene associations that account for some PCOS women, PCOS may have multiple molecular underpinnings that arise from common or varied developmental origins.
Epigenetic changes coexist with many of these PCOS candidate genes [53,54]. In SC abdominal adipose, over-expression of the LHCG receptor and under-expression of the insulin receptor in nonobese and obese PCOS women, respectively, accompany reciprocal DNA methylation patterns [55], while reciprocal changes of gene expression and DNA methylation also coexist in adipogenic pathways of overweight PCOS women [56]. In PCOS theca cells, moreover, decreased expression of miR-130b-3b (i.e., a noncoding microRNA transcriptional represser) correlates with increased DENND1A.V2 and CYP17A1 expression as well as with androgen synthesis [49,57], while three PCOS-specific gene variants of AMHR2 occur in regions of higher methylation and acetylation activity [51]. PCOS susceptible loci alone, however, do not fully explain the majority of PCOS phenotypic expression [58], so that heritability of PCOS likely involves one or more PCOS candidate genes that have interacted with environmental factors throughout antiquity to modify target tissue phenotype through epigenetic events [5].
3. PCOS phenotypic expression
Most women with PCOS have systemic insulin resistance from perturbed insulin receptor/post receptor signaling, altered adipokine secretion and/or abnormal steroid metabolism [2] in combination with preferential abdominal fat accumulation worsened by obesity [1,59,60,61]. Most women with PCOS also have increased adiposity [62,63,64] that interacts with hyperandrogenism to worsen PCOS phenotypic expression [1,2,3,65,66,67] and insulin resistance [2,68,69]. Different PCOS phenotypes by Rotterdam criteria also vary in endocrine-metabolic dysfunction [70], with NIH-defined PCOS women (i.e., hyperandrogenism with oligo-anovulation) having the greatest risk of developing menstrual irregularity, anovulatory infertility, T2DM and metabolic syndrome [1]. Furthermore, women with PCOS from a referral population have a more severe phenotype than those from the general population [71,72].
To understand the origins of PCOS, the above variables underlying endocrine-metabolic dysfunction in PCOS need to be eliminated when comparing the clinical characteristics of healthy, normal-weight PCOS women by NIH criteria with age/BMI-balanced controls [68,69,71,73,74]. In doing so, healthy normal-weight PCOS women by NIH criteria show low-normal insulin sensitivity (Si) by frequently sampled intravenous glucose tolerance testing (FSIVGTT) in combination with preferential abdominal fat accumulation (i.e., android fat) by total body dual-energy x-ray absorptiometry (DXA) [59,75,76]. Compared to age- and BMI-matched controls, normal-weight PCOS women by NIH criteria also exhibit adipose insulin resistance (adipose-IR; defined by the product of fasting circulating free fatty acid [FFA] and insulin levels) [73,76,77].
4. Total abdominal (android) fat mass
Abdominal fat mass comprises two major adipose depots: subcutaneous (SC) and intra-abdominal adipose. In humans, SC abdominal adipose normally stores lipid as protection against insulin resistance, while intra-abdominal adipose has the opposite effect [78]. Total body dual-energy x-ray absorptiometry studies confirm that android fat mass and the percent android fat relative to total body fat are greater in normal-weight PCOS women than age- and BMI-matched controls [59,75]. In all women combined, android fat mass positively correlates with circulating levels of total testosterone (T), free T, androstenedione (A4), and fasting insulin, as does the percent android fat mass relative to total body fat with circulating levels of total T, free T, A4, and fasting insulin [59,76]. Android fat mass in these individuals also negatively correlates with circulating cortisol levels, demonstrating an opposing system interplay of testosterone with cortisol in the control of android fat mass in women with PCOS [76].
Adjusting for fasting insulin levels, android fat mass remains positively correlated with circulating total T levels, as does the percent android fat mass relative to total body fat with circulating levels of total T, free T, and A4 [59]. In these normal-weight PCOS women, moreover, androgen receptor blockade by low-dose flutamide simultaneously decreases percent android fat and increases fasting glucose levels, supporting the role of androgen excess in the metabolic adaptation of PCOS through body fat distribution [16,79].
4.1. Intra-abdominal adipose
Intra-abdominal (visceral) adipose in humans is normally highly lipolytic and resists androgen inhibition of catecholamine-induced lipolysis (lipid breakdown) despite expressing androgen receptors [80]. Intra-abdominal fat mass in normal-weight NIH-defined PCOS women is increased in proportion to circulating androgen concentrations, and fasting levels of insulin, TG and non-high-density lipoprotein (non-HDL) cholesterol [59]; it also exhibits exaggerated catecholamine-induced lipolysis in nonobese PCOS women [81,82]. These intra-abdominal fat characteristics favor enhanced FFA availability for hepatic lipid storage and utilization [83]. They also promote insulin resistance with obesity, however, when increased FFA availability exceeds the capacity of target tissues to oxidize fat or convert diacylglycerols to triacylglycerols [81,82,84].
4.2. Subcutaneous abdominal adipose
Subcutaneous abdominal adipose normally protects against insulin resistance by balancing lipogenesis (lipid formation) with lipolysis (lipid breakdown) in mature adipocytes in combination with adipogenesis (whereby adipose stem cells [ASCs] initially commit to preadipocytes and then differentiate into newly-formed adipocytes) (Figure 2) [85,86,87]. In this manner, SC adipose can increase its fat storage through enlargement of mature adipocytes (i.e., hypertrophy) and development of new adipocytes (i.e., hyperplasia) to buffer fatty acid influx as energy intake exceeds its expenditure [88,89].
Within SC adipose, androgen normally diminishes insulin-stimulated glucose uptake and impairs catecholamine-stimulated lipolysis through reduced β2-adrenergic receptor and hormone-sensitive lipase (HSL) protein expression [80,81,90]. Women with PCOS have similar SC abdominal adipose characteristics of diminished insulin-mediated glucose uptake, reduced glucose transporter type 4 (GLUT-4) expression [91] and catecholamine lipolytic resistance [92,93]. Importantly, catecholamine lipolytic resistance in normal-weight PCOS women [92,93] can be counterbalanced by impaired insulin suppression of lipolysis in overweight PCOS women [94].
Within SC adipose, an aldo-ketoreductase enzyme, namely aldo-ketoreductase type 1C3 (AKR1C3) generates local T from A4 [95,96]. AKR1C3 gene expression and activity are greater in SC gluteal than omental fat, with SC gluteal fat favoring androgen activation (i.e., AKR1C3), and omental cells favoring androgen inactivation (i.e., aldo-ketoreductase type 1C2 [AKR1C2]) [96]. In PCOS women, increased AKR1C3-mediated androgen activation enhances lipid storage through increased lipogenesis and decreased lipolysis [97,98], promoting fat accretion [75,98,99] despite diminished insulin-stimulated glucose uptake [90].
Figure 2.
Schematic representation of adipogenesis in subcutaneous abdominal adipose stem cells (ASCs) from normal-weight women with polycystic ovary syndrome. Adipogenesis involves ASC commitment to preadipocytes, followed by and early/late stage preadipocyte differentiation to immature/mature adipocytes [85,86,87]. Dynamic changes in chromatin accessibility of SC abdominal ASCs during adipogenesis activates different transcriptional factors/genes (zinc-finger protein 423 [Zfp423], activator protein-1 [AP-1], peroxisome proliferator-activated receptor γ [PPARγ], CCAAT enhancer binding protein a [CEBPα] and aldo-ketoreductase type 1C3 [AKR1C3]), leading to increased free fatty acid (FFA) incorporation/biosynthesis, thus forming triglycerides (TG) in newly-formed mature adipocytes [79,97,98,99,100].
Figure 2.
Schematic representation of adipogenesis in subcutaneous abdominal adipose stem cells (ASCs) from normal-weight women with polycystic ovary syndrome. Adipogenesis involves ASC commitment to preadipocytes, followed by and early/late stage preadipocyte differentiation to immature/mature adipocytes [85,86,87]. Dynamic changes in chromatin accessibility of SC abdominal ASCs during adipogenesis activates different transcriptional factors/genes (zinc-finger protein 423 [Zfp423], activator protein-1 [AP-1], peroxisome proliferator-activated receptor γ [PPARγ], CCAAT enhancer binding protein a [CEBPα] and aldo-ketoreductase type 1C3 [AKR1C3]), leading to increased free fatty acid (FFA) incorporation/biosynthesis, thus forming triglycerides (TG) in newly-formed mature adipocytes [79,97,98,99,100].
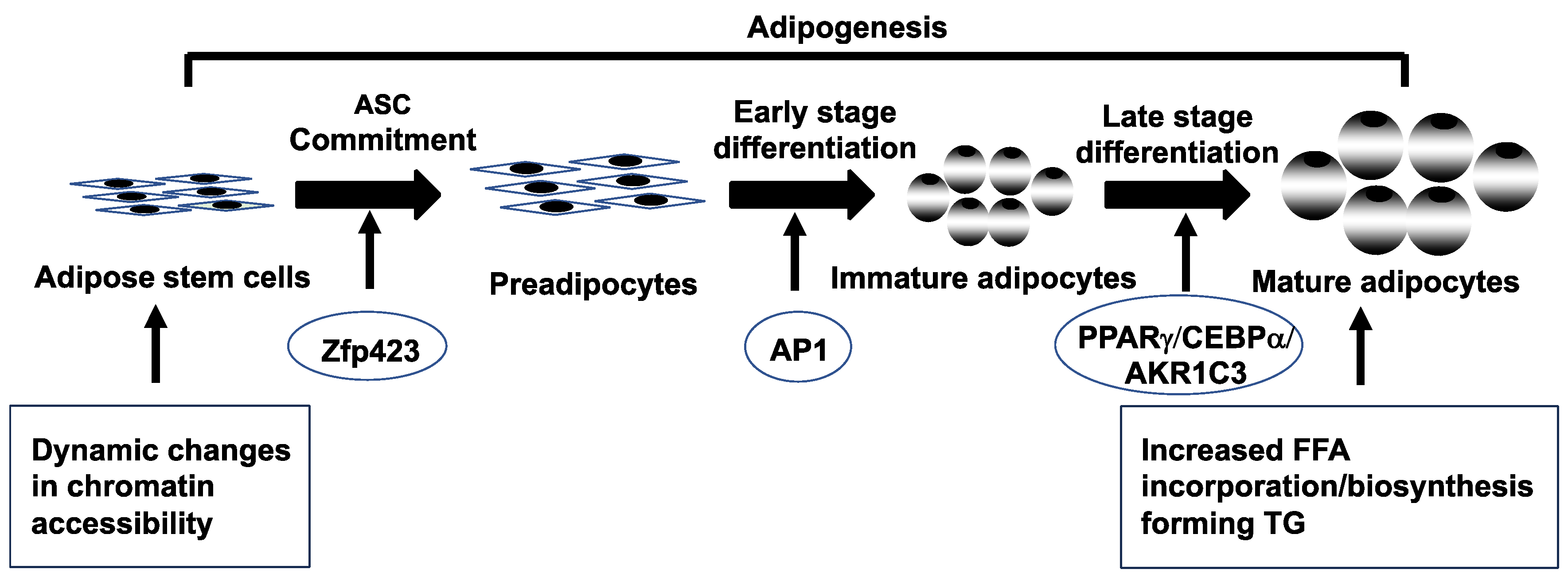
4.3. Subcutaneous abdominal stem cells and cellular reprogramming
Subcutaneous abdominal ASCs from normal-weight PCOS women exhibit altered dynamic chromatin accessibility during adipogenesis compared to control ASCs, and are characterized by limited chromatin accessibility in undifferentiated ASCs (quiescent stage) followed by exaggerated availability (active stage) in newly-formed adipocytes [100]. These chromatin remodeling patterns of PCOS stem cells accompany enrichment of binding motifs for transcription factors (TFs) of the activator protein-1 (AP-1) subfamily during early cell differentiation, with altered gene expression of adipogenic/angiogenic functions involving androgen-insulin interactions through transforming growth factor (TGF)-ß1 signaling [77].
In these SC abdominal ASCs of normal-weight PCOS women, an exaggerated commitment to preadipocytes via zinc-finger protein 423 (ZFP423) overexpression negatively correlates with fasting circulating glucose levels [99], and accompanies a greater proportion of small SC abdominal adipocytes [59,77], presumably to buffer against fatty acid influx [89,101]. Similar small SC abdominal adipocytes occur in other individuals [101,102,103], in whom they protect against insulin resistance through stem cell ZFP423 upregulation from epigenetic modifications [104].
Following exaggerated commitment to preadipocytes, these same abdominal ASCs from normal-weight PCOS exhibit accelerated lipid accumulation in newly-formed adipocytes in vitro that predicts reduced serum FFA levels and improved systemic insulin sensitivity in vivo [75,99]. These differentiating PCOS stem cells can overexpress the genes, peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT enhancer binding protein a (CEBPa), in combination with increased AKR1C3 gene expression during adipocyte maturation in vitro (Figure 2) [79,98,100].
From a causal perspective, administration of flutamide (an androgen receptor blocker) to healthy normal-weight PCOS women attenuates accelerated lipid accumulation within these newly-formed adipocytes in vitro and increases fasting circulating glucose levels (but within the normal range) [79]. In addition to intrinsic changes in PCOS stem cell characteristics, therefore, local androgen excess in PCOS appears to enhance lipid storage in SC abdominal adipocytes [79,98,99] and favor insulin sensitivity [75,105,106].
5. Lipotoxicity
Lipotoxicity refers to the ectopic lipid accumulation in non-adipose tissue, where it induces oxidative/endoplasmic reticulum stress linked with insulin resistance and inflammation [107]. Overweight/obese PCOS women; with greater preferential abdominal fat accumulation, hyperandrogenism, and insulin resistance [2]; are at particular risk of developing lipotoxicity due to excess FFA uptake into non-adipose cells, in part from increased highly-lipolytic intra-abdominal fat with impaired insulin suppression of lipolysis [81,82,94,108,109,110]. In these individuals, excess fatty acid influx in skeletal muscle and liver promotes diacylglycerol-induced insulin resistance, impairs insulin signaling via increased insulin receptor serine phosphorylation, and disrupts mitochondrial oxidative phosphorylation [84,111]. Enlarged SC abdominal mature adipocytes in overweight compared to normal-weight PCOS women also fosters a pro-inflammatory lipid depot environment [59,94].
Within today’s contemporary lifestyle, NIH-defined PCOS women have a 2- to 3-fold higher prevalence of metabolic syndrome (33-47%) than that of age-matched normal women [3,112,113,114], which is reduced by diminished abdominal fat accumulation [114]. Beginning in adolescence, an increased risk for developing metabolic syndrome [115] is evident in hyperandrogenic women [116] who preferentially increase abdominal adiposity with weight gain [61].
Increased abdominal fat in PCOS women also increases the risk of developing nonalcoholic fatty liver disease (NAFLD) [117,118,119], with non-alcoholic hepatic steatosis varying in inflammation and fibrosis [120]. Obesity in PCOS women is an important risk factor for hepatic steatosis [117], as is androgen excess per se, since the probability of hepatic steatosis (37%) and elevated serum aminotransferase levels are greater in hyperandrogenic women with PCOS than age- and weight-matched controls [121,122]. Magnetic resonance spectroscopy further confirms greater liver fat content in women with hyperandrogenic PCOS than non-hyperandrogenic PCOS [123].
6. Parallel evolution of PCOS-like traits in naturally hyperandrogenic female rhesus macaques
Ancestors of macaques migrated out of Africa before humans (Figure 1), about 5-6 million years ago [12,15]. Second only to humans, contemporary rhesus macaques occupy the largest habitat range of any primate, somewhat emulating humans in their diversity of habitats, including obesogenic urban environments [13]. Such close evolutionary history to humans bestows considerable similarities in genomic, developmental, physiological, anatomical, neurological, behavioral and aging characteristics, as well as comparable breadth of natural disease susceptibility [10], including female hyperandrogenism, PCOS [8,9] and obesity [124]. Obesity in rhesus macaques is heritable [125], emulates that in humans [126,127,128], and may associate with human obesity risk genes [125], increased risk of T2DM [127,129], dyslipidemia [12,126,128,130] and cardiometabolic disease [131,132]. In female rhesus macaques, as in women, hyperandrogenism enhances obesity outcomes [128,130,133]. Examining the etiology for female rhesus macaque hyperandrogenism and accompanying PCOS-like traits, including metabolic dysfunction, may thus provide supportive evidence for parallel evolution of these traits to humans and for a shared vulnerability to PCOS (Figure 1). In addition, female rhesus macaques and humans share menstrual cycle traits, including a relatively lengthy follicular or preovulatory phase, exposing selection of a single preovulatory follicle to hyperandrogenic anovulatory consequences of prolonged LH hypersecretion, FSH hyposecretion [134], as well as hyperinsulinemia [10].
Hyperandrogenic female rhesus monkeys with increased adiposity also emulate metabolic dysfunction as seen in women with PCOS. They exhibit increased abdominal subcutaneous and visceral adiposity [128,133,135], adipose insulin resistance and impaired insulin secretion [136], along with an increased incidence of T2DM [137]. Their SC abdominal adipocytes demonstrate an altered ability to store fat relative to BMI [128,130,135,138], with impaired preadipocyte differentiation into adipocytes accompanying decrease in C/EBPα mRNA. An associated enhancement of SC abdominal ASC commitment to preadipocytes through increased ZFP423 mRNA expression may indicate a compensatory mechanism for impaired preadipocyte differentiation [138]. Those with the highest testosterone values demonstrate increased BMI, central adiposity and insulin resistance [8,128].
Hyperandrogenism in female rhesus monkeys may have developmental origins, emulating PCOS in women. A positive correlation of adult anogenital distance with circulating testosterone levels in hyperandrogenic adult female rhesus monkeys suggests mid-gestational hyperandrogenic origins [9]. Increased anogenital distance has also been reported for girls born to women with PCOS [139] and in women with PCOS [140]. Elevated maternal circulating levels of anti-mullerian hormone from polycystic ovaries may enhance maternal hyperandrogenism and amplify epigenetic transgenerational transmission of hyperandrogenic phenotype in female offspring [141]. Consistent with these findings, gestational hyperandrogenism in rhesus monkeys induces maternal hyperinsulinemia and hyperglycemia, and reliably generates 75% of female offspring with heterogenous PCOS-like phenotypes [142], with gestational hyperinsulinemia inducing ectopic pericardial and perirenal fetal lipid accumulation [143]. Given these findings implicating hyperandrogenic developmental origins in the etiology of preferential fat storage, female rhesus monkeys may provide unique insight into proximate mechanisms amplifying outcomes from inheritance of PCOS risk genes, calling for gene editing studies of monkey embryos/cells to express female phenotypes generated by specific PCOS risk genes in individuals of known genetic backgrounds [10,144].
7. Conclusions
Polycystic ovary syndrome has persisted from antiquity to become the most common endocrine-metabolic disorder of reproductive-aged women. While its ancestral traits once favored abdominal fat deposition and increased energy availability through hyperandrogenism and insulin resistance for reproduction within hostile environments of food deprivation, these same traits now underlie different PCOS phenotypes with various risks for endocrine-metabolic dysfunction that are worsened by obesity. Healthy normal-weight women with NIH-defined PCOS have SC abdominal adipose characteristics that favor lipid storage in combination with low-normal insulin sensitivity accompanied by increased highly-lipolytic intra-abdominal fat deposition. As an ancestral trait programmed by genetic inheritance and epigenetic amplification during gestation, such an evolutionary metabolic adaptation in normal-weight PCOS women balances enhanced SC adipose storage with increased circulating glucose availability and free fatty acid oxidation as energy substrate for brain, muscle and other crucial target tissues. This metabolic adaptation in hyperandrogenic PCOS women also favors oligo-ovulation, which allowed women from antiquity sufficient time for childrearing of fewer offspring who in turn had a greater likelihood of childhood survival [6].
8. Future Directions
These heritable PCOS characteristics are now adversely affected by today’s contemporary environment through epigenetic events that predispose to lipotoxicity with excess weight gain and pregnancy complications. Understanding the evolutionary origins of PCOS emphasizes the need for a greater focus on preventive healthcare with early and appropriate lifestyle as well as therapeutic choices to optimize the long-term, endocrine-metabolic health of PCOS women in today’s obesogenic environment.
Author Contributions
Conceptualization, D.A.D., G.D.C. and D.H.A; methodology, D.A.D., G.D.C. and D.H.A.; formal analysis, D.A.D., G.D.C. and D.H.A.; writing—original draft preparation, D.A.D., G.D.C. and D.H.A; writing—review and editing, D.A.D., G.D.C. and D.H.A.; funding acquisition, D.A.D., G.D.C. and D.H.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Eunice Kennedy Shriver National Institute of Child Health & Human Development (NICHD), National Institutes of Health (NIH) under awards P50 HD071836 and P51 ODO11092 for the Endocrine Technologies Support Core (ETSC) through the Oregon National Primate Research Center; statistical analyses by the NIH National Center for Advancing Translational Science (NCATS) UCLA CTSI Grant Number UL1TR001881; and the Santa Monica Bay Woman’s Club. Nonhuman primate research was supported by awards R01 DK121559 (National Institute of Diabetes and Digestive and Kidney Diseases), R21 HD102172 (NICHD) and P51 OD011106 for the Office of Research Infrastructure Programs (ORIP) through the Wisconsin National Primate Research Center. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Institutional Review Board Statement
All studies were performed according to the Declaration of Helsinki after approval by the UCLA Institutional Review Board and signed informed consent by each subject (IRB number 12-001780; approval date January 18, 2013).
Informed Consent statement
Informed consent was obtained from all subjects involved in the study. Written informed consent was obtained from the patient(s) to publish this paper.
Data Availability Statement
Not applicable.
Acknowledgements
We thank Karla Largaespada at UCLA for subject recruitment strategies and administrative responsibilities that were crucial for the successful studies of the PCOS and control subjects; the veterinary, pathology, animal care and assays staff at the Wisconsin National Primate Research Center (WNPRC); and Jon Levine (WNPRC) for his highly valued contributions to refining and enabling our research into the developmental origins of PCOS.
Conflicts of Interests
D.A.D has consulted for Spruce Biosciences, Inc; Precede Biosciences, Inc; Ferring Research Institute; and Organon LLC. The funders had no role in the design of the study; in the collection, analysis or interpretation of data; in the writing of the manuscript; or in the decision to publish the results. G.D.C. and D.H.A have nothing to disclose.
References
- Chang RJ, Dumesic DA. Polycystic Ovary Syndrome and Hyperandrogenic States. In Yen and Jaffe’s Reproductive Endocrinology: Physiology, Pathophysiology and Clinical Management, 9th ed.; Strauss JF III, Barbieri RL, Dokras A, Williams CJ, Williams SZ, Eds.; Elsevier Saunders: Philadelphia, USA, 2024; pp. 517-547.
- Dumesic DA, Oberfield SE, Stener-Victorin E, Marshall JC, Laven JS, Legro RS. Scientific Statement on the Diagnostic Criteria, Epidemiology, Pathophysiology, and Molecular Genetics of Polycystic Ovary Syndrome. Endocr Rev. 2015,36(5),487-525. [CrossRef]
- Moran LJ, Misso ML, Wild RA, Norman RJ. Impaired glucose tolerance, type 2 diabetes and metabolic syndrome in polycystic ovary syndrome: a systematic review and meta-analysis. Hum Repro Update. 2010,16(4),347-363. [CrossRef]
- Carmina E, Napoli N, Longo RA, Rini GB, Lobo RA. Metabolic syndrome in polycystic ovary syndrome (PCOS): lower prevalence in southern Italy than in the USA and the influence of criteria for the diagnosis of PCOS. Eur J Endocrinol. 2006,154(1),141-145. [CrossRef]
- Dumesic DA, Hoyos LR, Chazenbalk GD, Naik R, Padmanabhan V, Abbott DH. Mechanisms of Intergenerational Transmission of Polycystic Ovary Syndrome. Reproduction, 2020,159(1),R1-R13. [CrossRef]
- Corbett S, Morin-Papunen L. Polycystic ovary syndrome and recent human evolution. Mol Cell Endocrinol. 2013,373(1-2),39-50. [CrossRef]
- Azziz R, Dumesic DA, Goodarzi M. Polycystic Ovary Syndrome: an ancient disorder? Fertil Steril 2011,95,1544-1548. [CrossRef]
- Arifin E, Shively CA, Register TC, Cline JM. Polycystic ovary syndrome with endometrial hyperplasia in a cynomolgus monkey (Macaca fascicularis). Vet Pathol. 2008,45(4),512-515. [CrossRef]
- Abbott DH, Rayome BH, Dumesic DA, Lewis KC, Edwards AK, Wallen K, Wilson ME, Appt SE, Levine JE. Clustering of PCOS-like traits in naturally hyperandrogenic female rhesus monkeys. Hum Reprod. 2017,32(4),923-936. [CrossRef]
- Abbott DH, Rogers J, Dumesic DA, Levine JE. Naturally Occurring and Experimentally Induced Rhesus Macaque Models for Polycystic Ovary Syndrome: Translational Gateways to Clinical Application. Med Sci (Basel). 2019,7(12),107. [CrossRef]
- Perelman P, Johnson WE, Roos C, Seuánez HN, Horvath JE, Moreira MA, Kessing B, Pontius J, Roelke M, Rumpler Y, Schneider MP, Silva A, O'Brien SJ, Pecon-Slattery J. A molecular phylogeny of living primates. PLoS Genet. 2011,7(3):e1001342. 1001. [CrossRef]
- Raaum RL, Sterner KN, Noviello CM, Stewart CB, Disotell TR. Catarrhine primate divergence dates estimated from complete mitochondrial genomes: concordance with fossil and nuclear DNA evidence. J Hum Evol. 2005,48(3):237-257. [CrossRef]
- Cooper EB, Brent LJN, Snyder-Mackler N, Singh M, Sengupta A, Khatiwada S, Malaivijitnond S, Qi Hai Z, Higham JP. The rhesus macaque as a success story of the Anthropocene. Elife. 2022,11:e78169. [CrossRef]
- Leakey M, Grossman A, Gutiérrez M, Fleagle JG. Faunal change in the Turkana Basin during the late Oligocene and Miocene. Evol Anthropol. 2011,20(6):238-253. [CrossRef]
- Stewart CB, Disotell TR. Primate evolution - in and out of Africa. Curr Biol. 1998,13;8(16):R582-R588. [CrossRef]
- Dumesic DA, Padmanabhan V, Levine J, Chazenbalk GD, Abbott DH. Polycystic Ovary Syndrome as a Plausible Evolutionary Metabolic Adaptation. Repro Biol Endocrinol 2022,20(1),12. [CrossRef]
- Parker J, O'Brien C, Hawrelak J, Gersh FL. Polycystic Ovary Syndrome: An Evolutionary Adaptation to Lifestyle and the Environment. Int J Environ Res Public Health. 2022,19(3),1336. [CrossRef]
- Parker J. Pathophysiological Effects of Contemporary Lifestyle on Evolutionary-Conserved Survival Mechanisms in Polycystic Ovary Syndrome. Life (Basel). 2023,13(4),1056. 4. [CrossRef]
- Tsatsoulis A, Mantzaris MD, Bellou S, Andrikoula M. Insulin resistance: an adaptive mechanism becomes maladaptive in the current environment - an evolutionary perspective. Metabolism. 2013,62(5),622-633. [CrossRef]
- Björntorp P. Metabolic implications of body fat distribution. Diabet Care. 1991,14(12):1132-1143. [CrossRef]
- Björntorp P. Visceral obesity: a "civilization syndrome". Obes Res. 1993,1(3):206-222. [CrossRef]
- López-Otín C, Kroemer G. Hallmarks of Health. Cell. 2021,184(1),33-63. [CrossRef]
- Klimentidis YC, Beasley TM, Lin HY, Murati G, Glass GE, Guyton M, Newton W, Jorgensen M, Heymsfield SB, Kemnitz J, Fairbanks L, Allison DB. Canaries in the coal mine: a cross-species analysis of the plurality of obesity epidemics. Proc Biol Sci. 2011,278(1712),1626-1632. [CrossRef]
- Terasawa E, Kurian JR, Keen KL, Shiel NA, Colman RJ, Capuano SV. Body weight impact on puberty: effects of high-calorie diet on puberty onset in female rhesus monkeys. Endocrinology. 2012,153(4),1696-1705. [CrossRef]
- Legro RS, Driscoll D, Strauss JF, 3rd, Fox J, Dunaif A. Evidence for a genetic basis for hyperandrogenemia in polycystic ovary syndrome. Proc Natl Acad Sci U S A 95,1998,14956-14960. [CrossRef]
- Vink JM, Sadrzadeh S, Lambalk CB, Boomsma DI. Heritability of polycystic ovary syndrome in a Dutch twin-family study. J Clin Endocrinol Metab. 2006,91,2100-2104. [CrossRef]
- Risal S, Pei Y, Lu H, Manti M, Fornes R, Pui HP, Zhao Z, Massart J, Ohlsson C, Lindgren E, Crisosto N, Maliqueo M, Echiburú B, Ladrón de Guevara A, Sir-Petermann T, Larsson H, Rosenqvist MA, Cesta CE, Benrick A, Deng Q, Stener-Victorin E. Prenatal androgen exposure and transgenerational susceptibility to polycystic ovary syndrome. Nat Med. 2019,25(12),1894-1904. [CrossRef]
- Shan D, Han J, Cai Y, Zou L, Xu L, Shen Y. Reproductive Health in First-degree Relatives of Patients With Polycystic Ovary Syndrome: A Review and Meta-analysis. J Clin Endocrinol Metab. 2022,107(1),273-295. [CrossRef]
- Kahsar-Miller MD, Nixon C, Boots LR, Go RC, Azziz R. Prevalence of polycystic ovary syndrome (PCOS) in first-degree relatives of patients with PCOS. Fertil Steril 2001,75(1),53-58. [CrossRef]
- Chen ZJ, Zhao H, He L, Shi Y, Qin Y, Shi Y, Li Z, You L, Zhao J, Liu J, Liang X, Zhao X, Zhao J, Sun Y, Zhang B, Jiang H, Zhao D, Bian Y, Gao X, Geng L, Li Y, Zhu D, Sun X, Xu JE, Hao C, Ren CE, Zhang Y, Chen S, Zhang W, Yang A, Yan J, Li Y, Ma J, Zhao Y. Genome-wide association study identifies susceptibility loci for polycystic ovary syndrome on chromosome 2p16.3, 2p21 and 9q33.3. Nat Genet. 2011,43(1),55-59. [CrossRef]
- Shi Y, Zhao H, Shi Y, Cao Y, Yang D, Li Z, Zhang B, Liang X, Li T, Chen J, Shen J, Zhao J, You L, Gao X, Zhu D, Zhao X, Yan Y, Qin Y, Li W, Yan J, Wang Q, Zhao J, Geng L, Ma J, Zhao Y, He G, Zhang A, Zou S, Yang A, Liu J, Li W, Li B, Wan C, Qin Y, Shi J, Yang J, Jiang H, Xu JE, Qi X, Sun Y, Zhang Y, Hao C, Ju X, Zhao D, Ren CE, Li X, Zhang W, Zhang Y, Zhang J, Wu D, Zhang C, He L, Chen ZJ. Genome-wide association study identifies eight new risk loci for polycystic ovary syndrome. Nat Genet. 2012,44(9),1020-1025. Nat Genet. 1025. [CrossRef]
- Goodarzi MO, Jones MR, Li X, Chua AK, Garcia OA, Chen YD, Krauss RM, Rotter JI, Ankener W, Legro RS, Azziz R, Strauss JF 3rd, Dunaif A, Urbanek M. Replication of association of DENND1A and THADA variants with polycystic ovary syndrome in European cohorts. J Med Genet. 2012,49(2),90-95. [CrossRef]
- Mutharasan P, Galdones E, Penalver Bernabe B, Garcia OA, Jafari N, Shea LD, Woodruff TK, Legro RS, Dunaif A, Urbanek M. Evidence for chromosome 2p16.3 polycystic ovary syndrome susceptibility locus in affected women of European ancestry. J Clin Endocrinol Metab. 2013,98(1),E185-E190. [CrossRef]
- Hayes MG, Urbanek M, Ehrmann DA, Armstrong LL, Lee JY, Sisk R, Karaderi T, Barber TM, McCarthy MI, Franks S, Lindgren CM, Welt CK, Diamanti-Kandarakis E, Panidis D, Goodarzi MO, Azziz R, Zhang Y, James RG, Olivier M, Kissebah AH; Reproductive Medicine Network; Stener-Victorin E, Legro RS, Dunaif A. Genome-wide association of polycystic ovary syndrome implicates alterations in gonadotropin secretion in European ancestry populations. Nat Commun. 2015,6,7502. [CrossRef]
- Day FR, Hinds DA, Tung JY, Stolk L, Styrkarsdottir U, Saxena R, Bjonnes A, Broer L, Dunger DB, Halldorsson BV, Lawlor DA, Laval G, Mathieson I, McCardle WL, Louwers Y, Meun C, Ring S, Scott RA, Sulem P, Uitterlinden AG, Wareham NJ, Thorsteinsdottir U, Welt C, Stefansson K, Laven JSE, Ong KK, Perry JRB. Causal mechanisms and balancing selection inferred from genetic associations with polycystic ovary syndrome. Nat Commun. 2015,6,8464. [CrossRef]
- Day F, Karaderi T, Jones MR, Meun C, He C, Drong A, Kraft P, Lin N, Huang H, Broer L, Magi R, Saxena R, Laisk T, Urbanek M, Hayes MG, Thorleifsson G, Fernandez-Tajes J, Mahajan A, Mullin BH, Stuckey BGA, Spector TD, Wilson SG, Goodarzi MO, Davis L, Obermayer-Pietsch B, Uitterlinden AG, Anttila V, Neale BM, Jarvelin MR, Fauser B, Kowalska I, Visser JA, Andersen M, Ong K, Stener-Victorin E, Ehrmann D, Legro RS, Salumets A, McCarthy MI, Morin-Papunen L, Thorsteinsdottir U, Stefansson K; 23andMe Research Team, Styrkarsdottir U, Perry JRB, Dunaif A, Laven J, Franks S, Lindgren CM, Welt CK. Large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for different diagnosis criteria. PLoS Genet. 2018,14(12),e1007813. 1007. [CrossRef]
- Dapas M, Dunaif A. The contribution of rare genetic variants to the pathogenesis of polycystic ovary syndrome. Curr Opin Endocr Metab Res 2020,12,26-32. [CrossRef]
- Dapas M, Dunaif A. Deconstructing a Syndrome: Genomic Insights Into PCOS Causal Mechanisms and Classification. Endocr Rev. 2022,43(6),927-965. [CrossRef]
- Tian Y, Li J, Su S, Cao Y, Wang Z, Zhao S, Zhao H. PCOS-GWAS Susceptibility Variants in THADA, INSR, TOX3, and DENND1A Are Associated With Metabolic Syndrome or Insulin Resistance in Women With PCOS. Front Endocrinol (Lausanne). 2020,11,274. Front Endocrinol (Lausanne). [CrossRef]
- Ruth KS, Day FR, Tyrrell J, Thompson DJ, Wood AR, Mahajan A, Beaumont RN, Wittemans L, Martin S, Busch AS, Erzurumluoglu AM, Hollis B, O'Mara TA; Endometrial Cancer Association Consortium, McCarthy MI, Langenberg C, Easton DF, Wareham NJ, Burgess S, Murray A, Ong KK, Frayling TM, Perry JRB. Using human genetics to understand the disease impacts of testosterone in men and women. Nat Med. 2020,26(2),252-258. 2. [CrossRef]
- Shriner D, Tekola-Ayele F, Adeyemo A, Rotimi CN. Ancient Human Migration after Out-of-Africa. Sci Rep. 2016,6,26565. [CrossRef]
- Nielsen R, Akey JM, Jakobsson M, Pritchard JK, Tishkoff S, Willerslev E. Tracing the peopling of the world through genomics. Nature. 2017,541(7637),302-310. [CrossRef]
- Dapas M, Lin FTJ, Nadkarni GN, Sisk R, Legro RS, Urbanek M, Hayes MG, Dunaif A. Distinct subtypes of polycystic ovary syndrome with novel genetic associations: An unsupervised, phenotypic clustering analysis. PLoS Med. 2020,17(6),e1003132. [CrossRef]
- Strauss JF, 3rd, McAllister JM, Urbanek M. Persistence pays off for PCOS gene prospectors. J Clin Endocrinol Metab. 2012,97(7),2286-2288. [CrossRef]
- Tee MK, Speek M, Legeza B, Modi B, Teves ME, McAllister JM, Strauss JF 3rd, Miller WL. Alternative splicing of DENND1A, a PCOS candidate gene, generates variant 2. Mol Cell Endocrinol. 2016,434,25-35. [CrossRef]
- Dapas M, Sisk R, Legro RS, Urbanek M, Dunaif A, Hayes MG: Family-based quantitative trait meta-analysis implicates rare noncoding variants in DENND1A in polycystic ovary syndrome. J Clin Endocrinol Metab 2019,104(9),3835-3850. 9. [CrossRef]
- McAllister JM, Modi B, Miller BA, Biegler J, Bruggeman R, Legro RS, Strauss JF 3rd. Overexpression of a DENND1A isoform produces a polycystic ovary syndrome theca phenotype. Proc Natl Acad Sci U S A. 2014,111(15),E1519-E1527. [CrossRef]
- McAllister JM, Legro RS, Modi BP, Strauss JF 3rd. Functional genomics of PCOS: from GWAS to molecular mechanisms. Trends Endocrinol Metab. 2015,26(3),118-124. [CrossRef]
- Waterbury JS, Teves ME, Gaynor A, Han AX, Mavodza G, Newell J, Strauss JF 3rd, McAllister JM. The PCOS GWAS Candidate Gene ZNF217 Influences Theca Cell Expression of DENND1A.V2, CYP17A1, and Androgen Production. J Endocr Soc. 2022,6(7),bvac078. [CrossRef]
- Gorsic LK, Kosova G, Werstein B, Sisk R, Legro RS, Hayes MG, Teixeira JM, Dunaif A, Urbanek M. Pathogenic Anti-Mullerian Hormone Variants in Polycystic Ovary Syndrome. J Clin Endocrinol Metab. 2017,102,2862-2872. [CrossRef]
- Gorsic LK, Dapas M, Legro RS, Hayes MG, Urbanek M. Functional Genetic Variation in the Anti-Mullerian Hormone Pathway in Women With Polycystic Ovary Syndrome. J Clin Endocrinol Metab 2019,104(7): 2855-2874. [CrossRef]
- Barbotin AL, Peigné M, Malone SA, Giacobini P. Emerging Roles of Anti-Müllerian Hormone in Hypothalamic-Pituitary Function. Neuroendocrinology. 2019,109(3):218-229. [CrossRef]
- Nilsson E, Benrick A, Kokosar M, Krook A, Lindgren E, Källman T, Martis MM, Højlund K, Ling C, Stener-Victorin E. Transcriptional and epigenetic changes influencing skeletal muscle metabolism in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2018,103(12),4465-4477. [CrossRef]
- Vázquez-Martínez ER, Gómez-Viais YI, García-Gómez E, Reyes-Mayoral C, Reyes-Muñoz E, Camacho-Arroyo I, Cerbón M. DNA methylation in the pathogenesis of polycystic ovary syndrome. Reproduction 2019,158(1),R27-R40. 1. [CrossRef]
- Jones MR, Brower MA, Xu N, Cui J, Mengesha E, Chen YD, Taylor KD, Azziz R, Goodarzi MO. Systems Genetics Reveals the Functional Context of PCOS Loci and Identifies Genetic and Molecular Mechanisms of Disease Heterogeneity. PLoS Genet. 2015,11(8),e1005455. 8. [CrossRef]
- Kokosar M, Benrick A, Perfilyev A, Fornes R, Nilsson E, Maliqueo M, Behre CJ, Sazonova A, Ohlsson C, Ling C, Stener-Victorin E. Epigenetic and Transcriptional Alterations in Human Adipose Tissue of Polycystic Ovary Syndrome. Sci Rep. 2016,6,22883. [CrossRef]
- McAllister JM, Han AX, Modi BP, Teves ME, Mavodza GR, Anderson ZL, Shen T, Christenson LK, Archer KJ, Strauss JF. miRNA Profiling Reveals miRNA-130b-3p Mediates DENND1A Variant 2 Expression and Androgen Biosynthesis. Endocrinology. 2019,160(8),1964-1981. [CrossRef]
- Abbott DH, Dumesic DA, Levine JE. Hyperandrogenic Origins of Polycystic Ovary Syndrome – Implications for Pathophysiology and Therapy. Expert Rev Endocrinol Metab. 2019,14(2),131-143. [CrossRef]
- Dumesic DA, Akopians AL, Madrigal VK, Ramirez E, Margolis DJ, Sarma MK, Thomas AM, Grogan TR, Haykal R, Schooler TA, Okeya BL, Abbott DH, Chazenbalk GD. Hyperandrogenism Accompanies Increased Intra-Abdominal Fat Storage in Normal Weight Polycystic Ovary Syndrome Women. J Clin Endocrinol Metab. 2016,101(11),4178-4188. [CrossRef]
- Tosi F, Di Sarra D, Kaufman JM, Bonin C, Moretta R, Bonoro E, Zanolin E, Mogetti P. Total body fat and central fat mass independently predict insulin resistance but not hyperandrogenemia in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2015,100(2),661-669. [CrossRef]
- Holte J, Bergh T, Berne C, Berglund L, Lithell H. Enhanced early insulin response to glucose in relation to insulin resistance in women with polycystic ovary syndrome and normal glucose tolerance. J Clin Endocrinol Metab. 1994,78(5),1052-1058. [CrossRef]
- Rosenzweig JL, Ferrannini E, Grundy SM, Haffner SM, Heine RJ, Horton ES, Kawamori R. Primary prevention of cardiovascular disease and type 2 diabetes in patients at metabolic risk: an endocrine society clinical practice guideline. J Clin Endocrinol Metab. 2008,93(10),3671-3689. [CrossRef]
- Wyatt HR. Update on treatment strategies for obesity. J Clin Endocrinol Metab. 2013,98(4),1299-1306. [CrossRef]
- Pasquali R, Pelusi C, Genghini S, Cacciari M, Gambineri A. Obesity and reproductive disorders in women. Hum Reprod Update. 2003,9(4),359-372. [CrossRef]
- Diamanti-Kandarakis E, Dunaif A. Insulin resistance and the polycystic ovary syndrome revisited: an update on mechanisms and implications. Endo Rev. 2012,33(6),981-1030. [CrossRef]
- Lim SS, Norman RJ, Davies MJ, Moran LJ. The effect of obesity on polycystic ovary syndrome: a systematic review and meta-analysis. Obes Rev. 2013,14(2):95-109. [CrossRef]
- Yildiz BO, Knochenhauer ES, Azziz R. Impact of obesity on the risk for polycystic ovary syndrome. J Clin Endocrinol Metab. 2008,93(1),162-168. 1. [CrossRef]
- Kakoly NS, Khomami MB, Joham AE, Corray SD, Misso ML, Norman RJ, Harrison CL, Ranasinha S, Teede HJ, Moran LJ. Ethnicity, obesity and the prevalence of impaired glucose tolerance and type 2 diabetes in PCOS: a systematic review and meta-regression. Hum Reprod Update. 2018,24(4),455-467. [CrossRef]
- Palaniappan LP, Carnethon MR, Fortmann SP. Heterogeneity in the relationship between ethnicity, BMI, and fasting insulin. Diabetes Care. 2002,25(8),1351-1357. [CrossRef]
- Teede HJ, Tay CT, Laven J, Dokras A, Moran LJ, Piltonen TT, Costello MF, Boivin J, M Redman L, A Boyle J, Norman RJ, Mousa A, Joham AE; International PCOS Network. Recommendations from the 2023 International Evidence-based Guideline for the Assessment and Management of Polycystic Ovary Syndrome. Fertil Steril. 2023 Aug 14:S0015-0282(23)00719-7. Epub ahead of print. [CrossRef]
- Ezeh U, Yildiz BO, Azziz R. Referral bias in defining the phenotype and prevalence of obesity in polycystic ovary syndrome. J Clin Endocrinol Metab. 2013,98(6),E1088-1096. 6. [CrossRef]
- Lizneva D, Kirubakaran R, Mykhalchenko K, Suturina L, Chernukha G, Diamond MP, Azziz R. Phenotypes and body mass in women with polycystic ovary syndrome identified in referral versus unselected populations: systematic review and meta-analysis. Fertil Steril. 2016,106(6),1510-1520.e2. [CrossRef]
- Søndergaard E, Espinosa De Ycaza AE, Morgan-Bathke M, Jensen MD. How to measure adipose tissue insulin sensitivity. J Clin Endocrinol Metab. 2017,102(4),1193-1199. 4. [CrossRef]
- Hershkop K, Besor O, Santoro N, Pierpont B, Caprio S, Weiss R. Adipose insulin resistance in obese adolescents across the spectrum of glucose tolerance. J Clin Endocrinol Metab. 2016,101(6):2423-2431. [CrossRef]
- Dumesic DA, Tulberg A, Leung KL, Fisch SC, Grogan TR, Abbott DH, Naik R, Chazenbalk GD. Accelerated subcutaneous abdominal stem cell adipogenesis predicts insulin sensitivity in normal-weight women with polycystic ovary syndrome. Fertil Steril. 2021,116(1),232-242. [CrossRef]
- Dumesic DA, Turcu AF, Liu H, Grogan TR, Abbott DH, Lu G, Dharanipragada D, Chazenbalk GD. Interplay of Cortisol, Testosterone, and Abdominal Fat Mass in Normal-weight Women With Polycystic Ovary Syndrome. J Endocr Soc. 2023,7(8):bvad079. [CrossRef]
- Dumesic DA, Phan JD, Leung KL, Grogan TR, Ding X, Li X, Hoyos LR, Abbott DH, Chazenbalk GD. Adipose Insulin Resistance in Normal-Weight Polycystic Ovary Syndrome Women. J Clin Endocrinol Metab. 2019,104(6),2171-2183. [CrossRef]
- McLaughlin T, Lamendola C, Liu A, Abbasi F. Preferential fat deposition in subcutaneous versus visceral depots is associated with insulin sensitivity. J Clin Endocrinol Metab. 2011,96911):E1756-E1760. [CrossRef]
- Dumesic DA, Winnett C, Lu, G, Grogan TR, Abbott DH, Naik R, Chazenbalk GD. Randomized Clinical Trial: Effect of Low-Dose Flutamide on Abdominal Adipogenic Function in Normal-Weight Polycystic Ovary Syndrome Women. Fertil Steril. 2023,119(1),116-126. [CrossRef]
- Dicker A, Ryden M, Naslund E, Muehlen IE, Wiren M, Lafontan M, Arner P. Effect of testosterone on lipolysis in human pre-adipocytes from different fat depots. Diabetologia. 2004,47(3),420-428. [CrossRef]
- Arner P. Effects of testosterone on fat cell lipolysis. Species differences and possible role in polycystic ovarian syndrome. Biochimie. 2005,87(1),39-43. 1. [CrossRef]
- Ek I, Arner P, Rydén M, Holm C, Thörne A, Hoffstedt J, Wahrenberg H. A unique defect in the regulation of visceral fat cell lipolysis in the polycystic ovary syndrome as an early link to insulin resistance. Diabetes. 2002,51(2),484-492. [CrossRef]
- Zhou MS, Wang A, Yu H. Link between insulin resistance and hypertension: What is the evidence from evolutionary biology? Diabetol Metab Syndr. 2014,6(1),12. 1. [CrossRef]
- Samuel VT, Petersen KF, Shulman GI. Lipid-induced insulin resistance: unraveling the mechanism. Lancet. 2010,375(9733),2267-2277. [CrossRef]
- Chazenbalk GD, Singh P, Irge D, Shah A, Abbott DH, Dumesic DA. Androgens inhibit adipogenesis during human adipose stem cell commitment to predipocyte formation. Steroids. 2013,78(9),920-926. 9. [CrossRef]
- Cristancho AG, Lazar MA. Forming functional fat: a growing understanding of adipocyte differentiation. Nat Rev Mol Cell Biol. 2011,12(11),722-734. [CrossRef]
- Tang QQ, Lane MD. Adipogenesis: from stem cell to adipocyte. Annual Rev Biochem. 2012,81,715-736. 2012; 81, 715–736. [CrossRef]
- Saponaro C, Gaggini M, Carli F, Gastaldelli A. The subtle balance between lipolysis and lipogenesis: a critical point in metabolic homeostasis. Nutrients. 2015,7(11),9453-9474. [CrossRef]
- Romacho T, Elsen M, Rohrborn D, Eckel J. Adipose tissue and its role in organ crosstalk. Acta Physiol (Oxf). 2014,210(4),733-753. [CrossRef]
- Corbould A. Chronic testosterone treatment induces selective insulin resistance in subcutaneous adipocytes of women. J Endocrinol. 2007;192(3),585-594. [CrossRef]
- Rosenbaum D, Harber RS, Dunaif A. Insulin resistance in polycystic ovary syndrome: decreased expression of GLUT-4 glucose transporters in adipocytes. Am J Physiol. 1993,264(2 Pt 1),E197-E202. [CrossRef]
- Faulds G, Rydén M, Ek I, Wahrenberg H, Arner P. Mechanisms behind lipolytic catecholamine resistance of subcutaneous fat cells in the polycystic ovarian syndrome. J Clin Endocrinol Metab. 2003,88(5),2269-2273. 5. [CrossRef]
- Ek I, Arner P, Bergqvist A, Carlstrom K Wahrenberg H. Impaired adipocyte lipolysis in nonobese women with the polycystic ovary syndrome: a possible link to insulin resistance? J Clin Endocrinol Metab. 1997,82(4),1147-1153. [CrossRef]
- Mannerås-Holm L, Leonhardt H, Kullberg J, Jennische E, Odén A, Holm G, Hellström M, Lönn L, Olivecrona G, Stener-Victorin E, Lönn M. Adipose tissue has aberrant morphology and function in PCOS: Enlarged adipocytes and low serum adiponectin, but not circulating sex steroids, are strongly associated with insulin resistance. J Clin Endocrinol Metab. 2011,96(2),E304-E311. [CrossRef]
- Blouin K, Veilleux A, Luu-The V, Tchernof A. Androgen metabolism in adipose tissue: recent advances. Mol Cell Endocrinol. 2009,301(1-2),97-103. [CrossRef]
- Quinkler M, Sinha B, Tomlinson JW, Bujalska IJ, Stewart PM, Arlt W. Androgen generation in adipose tissue in women with simple obesity--a site-specific role for 17beta-hydroxysteroid dehydrogenase type 5. J Endocrinol. 2004,183(2),331-342. [CrossRef]
- O’Reilly MW, Kempegowda P, Walsh M, Taylor AE, Manolopoulos KN, Allwood JW, Semple RK, Hebenstreit D, Dunn WB, Tomlinson JW, Arlt W. AKR1C3-Mediated Adipose Androgen Generation Drives Lipotoxicity in Women with Polycystic Ovary Syndrome. J Clin Endocrinol Metab. 2017,102(9),3327-3339. [CrossRef]
- Dumesic DA, Tulberg A, McNamara M, Grogan TR, Abbott DH, Naik R, Lu G, Chazenbalk GD. Serum Testosterone to Androstenedione Ratio Predicts Metabolic Health in Normal-Weight Polycystic Ovary Syndrome Women. J Endocr Soc. 2021,5(11),bvab158. [CrossRef]
- Fisch SC, Nikou AF, Wright EA, Phan JD, Leung KL, Grogan TR, Abbott DH, Chazenbalk GD, Dumesic DA. Precocious Subcutaneous Abdominal Stem Cell Development to Adipocytes in Normal-Weight Polycystic Ovary Syndrome Women. Fertil Steril. 2018,110(7),1367-1376. [CrossRef]
- Leung KL, Sanchita S, Pham CT, Davis BA, Okhovat M, Ding X, Dumesic P, Grogan TR, Williams KJ, Morselli M, Ma F, Carbone L, Li X, Pellegrini M, Dumesic DA, Chazenbalk GD. Dynamic changes in chromatin accessibility, altered adipogenic gene expression, and total versus de novo fatty acid synthesis in subcutaneous adipose stem cells of normal-weight polycystic ovary syndrome (PCOS) women during adipogenesis: evidence of cellular programming. Clin Epigenetics. 2020,23,12(1),181. [CrossRef]
- Spalding KL, Arner E, Westermark, PO, Bernard S, Buchholz BA, Bergmann O, Blomqvist L, Hoffsted J, Naslund, E, Britton, T, Concha H, Hassan M, Rydén M, Frisén J, Arner P. Dynamics of fat cell turnover in humans. Nature. 2008,453(7196),783-787. [CrossRef]
- Tandon P, Wafer R, Minchin JE. Adipose morphology and metabolic disease. J Expt Biol. 2018,221(Pt Suppl 1),jeb164970. [CrossRef]
- Arner E, Westermark PO, Spalding, KL, Britton T, Ryden M, Frisen J, Bernard S, Arner P. Adipocyte turnover: relevance to human adipose tissue morphology. Diabetes. 2010,59(1),105-109. [CrossRef]
- Longo M, Raciti GA, Zatterale F, Parrillo L, Desiderio A, Spinelli R, Hammarstedt A, Hedjazifar S, Hoffmann JM, Nigro C, Mirra P, Fiory F, Formisano P, Miele C, Smith U, Beguinot F. Epigenetic modifications of the Zfp/ZNF423 gene control murine adipogenic commitment and are dysregulated in human hypertrophic obesity. Diabetologia. 2018,61(2),369-380. [CrossRef]
- Nouws J, Fitch M, Mata M, Santoro N, Galuppo B, Kursawe R, Narayan D, Vash-Margita A, Pierpont B, Shulman GI, Hellerstein M, Caprio S. Altered In Vivo Lipid Fluxes and Cell Dynamics in Subcutaneous Adipose Tissues Are Associated with the Unfavorable Pattern of Fat Distribution in Obese Adolescent Girls. Diabetes. 2019,68(6),1168-77. [CrossRef]
- Umano GR, Shabanova V, Pierpont B, Mata M, Nouws J, Tricò D, Galderisi A, Santoro N, Caprio S. A low visceral fat proportion, independent of total body fat mass, protects obese adolescent girls against fatty liver and glucose dysregulation: a longitudinal study. Int J Obes (Lond). 2019,43(4),673-682. [CrossRef]
- Brennan KM, Kroener LL, Chazenbalk GD, Dumesic DA. Polycystic Ovary Syndrome: Impact of Lipotoxicity on Metabolic and Reproductive Health. Obstet Gynecol Surv. 2019,74(4),223-231. [CrossRef]
- Virtue S, Vidal-Puig A. It's not how fat you are, it's what you do with it that counts. PLoS Biol. 2008, 6(9),e237. [CrossRef]
- Unger RH, Clark GO, Scherer PE, Orci L. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim Biophys Acta. 2010,1801(3),209-214. [CrossRef]
- de Zegher F, Lopez-Bermejo A, Ibáñez L. Adipose tissue expandability and the early origins of PCOS. Trends Endocrinol Metab. 2009,20(9),418-423. [CrossRef]
- Shulman GI. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N Eng J Med. 2014;371(12),1131-1141. [CrossRef]
- Apridonidze T, Essah PA, Iuorno MJ, Nestler JE. Prevalence and characteristics of the metabolic syndrome in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2005,90(4),1929-1935. [CrossRef]
- Dokras A, Bochner M, Hollinrake E, Markham S, Vanvoorhis B, Jagasia DH. Screening women with polycystic ovary syndrome for metabolic syndrome. Obstet Gynecol. 2005,106(1),131-137. [CrossRef]
- Ehrmann DA, Liljenquist DR, Kasza K, Azziz R, legro RS, Ghazzi MN; PCOS/Troglitazone Study Group. Prevalence and predictors of the metabolic syndrome in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2006,91(1),48:-53. [CrossRef]
- Fazleen NE, Whittaker M, Mamun A. Risk of metabolic syndrome in adolescents with polycystic ovarian syndrome: A systematic review and meta-analysis. Diabetes Metab Syndr. 2018,12(6),1083-1090. 1090. [CrossRef]
- Yang R, Yang S, Li R, Liu P, Qiao J, Zhang Y. Effects of hyperandrogenism on metabolic abnormalities in patients with polycystic ovary syndrome: a meta-analysis. Reprod Biol Endocrinol. 2016,14(1),67. [CrossRef]
- Gambarin-Gelwan M, Kinkhabwala SV, Schiano TD, Bodian C, Yeh HC, Futterweit W. Prevalence of nonalcoholic fatty liver disease in women with polycystic ovary syndrome. Clin Gastroenterol Hepatol. 2007,5(4),496-501. [CrossRef]
- Macut D, Tziomalos K, Božić-Antić I, Bjekić-Macut J, Katsikis I, Papadakis E, Andrić Z, Panidis D . Non-alcoholic fatty liver disease is associated with insulin resistance and lipid accumulation product in women with polycystic ovary syndrome. Hum Reprod. 2016,31(6),1347-1353. [CrossRef]
- Vassilatou E, Vassiliadi DA, Salambasis K, Lazaridou H, Koutsomitopoulos N, Kelekis N, Kassanos D, Hadjidakis D, Dimitriadis G. Increased prevalence of polycystic ovary syndrome in premenopausal women with nonalcoholic fatty liver disease. Eur J Endocrinol. 2015,173(6),739-747. [CrossRef]
- Browning JD, Horton JD. Molecular mediators of hepatic steatosis and liver injury. J Clin Invest. 2004,114(2),147-152. 2. [CrossRef]
- Vassilatou E, Lafoyianni S, Vryonidou A, Ioannidis D, Kosma L, Katsoulis K, Papavassiliou E, Tzavara I . Increased androgen bioavailability is associated with non-alcoholic fatty liver disease in women with polycystic ovary syndrome. Hum Reprod. 2010,25(1),212-220. [CrossRef]
- Petta S, Ciresi A, Bianco J, Geraci V, Boemi R, Galvano L, Magliozzo F, Merlino G, Craxì A, Giordano C. Insulin resistance and hyperandrogenism drive steatosis and fibrosis risk in young females with PCOS. PLoS One. 2017,12(11),e0186136. [CrossRef]
- Jones H, Sprung VS, Pugh CJ, Daousi C, Irwin A, Aziz N, Adams VL, Thomas EL, Bell JD, Kemp GJ, Cuthbertson DJ. Polycystic ovary syndrome with hyperandrogenism is characterized by an increased risk of hepatic steatosis compared to nonhyperandrogenic PCOS phenotypes and healthy controls, independent of obesity and insulin resistance. J Clin Endocrinol Metab. 2012,97(10),3709-3716. 3709. [CrossRef]
- Schwartz SM, Kemnitz JW, Howard CF Jr. Obesity in free-ranging rhesus macaques. Int J Obes Relat Metab Disord. 1993,17(1),1-9. [PubMed]
- Raboin MJ, Letaw J, Mitchell AD, Toffey D, McKelvey J, Roberts CT Jr, Curran JE, Vinson A. Genetic Architecture of Human Obesity Traits in the Rhesus Macaque. Obesity (Silver Spring). 2019,27(3),479-488. [CrossRef]
- Kemnitz JW, Goy RW, Flitsch TJ, Lohmiller JJ, Robinson JA. Obesity in male and female rhesus monkeys: fat distribution, glucoregulation, and serum androgen levels. J Clin Endocrinol Metab. 1989,69(2),287-93. [CrossRef] [PubMed]
- Pound LD, Kievit P, Grove KL. The nonhuman primate as a model for type 2 diabetes. Curr Opin Endocrinol Diabetes Obes. 2014;21(2),89-94. [CrossRef]
- True C, Abbott DH, Roberts CT Jr, Varlamov O. Sex Differences in Androgen Regulation of Metabolism in Nonhuman Primates. Adv Exp Med Biol. 2017,1043,559-574. [CrossRef]
- Bodkin NL, Alexander TM, Ortmeyer HK, Johnson E, Hansen BC. Mortality and morbidity in laboratory-maintained Rhesus monkeys and effects of long-term dietary restriction. J Gerontol A Biol Sci Med Sci. 2003,58(3),212-219. [CrossRef]
- ishop CV, Takahashi D, Mishler E, Slayden OD, Roberts CT, Hennebold J, True C. Individual and combined effects of 5-year exposure to hyperandrogenemia and Western-style diet on metabolism and reproduction in female rhesus macaques. Hum Reprod. 2021,36(2),444-454. [CrossRef]
- Brown E, Ozawa K, Moccetti F, Vinson A, Hodovan J, Nguyen TA, Bader L, López JA, Kievit P, Shaw GD, Chung DW, Osborn W, Fu X, Chen J, Lindner JR. Arterial Platelet Adhesion in Atherosclerosis-Prone Arteries of Obese, Insulin-Resistant Nonhuman Primates. J Am Heart Assoc. 2021,10(9),e019413. [CrossRef]
- Newman LE, Testard C, DeCasien AR, Chiou KL, Watowich MM, Janiak MC, Pavez-Fox MA, Sanchez Rosado MR, Cooper EB, Costa CE, Petersen RM, Montague MJ, Platt ML, Brent LJN, Snyder-Mackler N, Higham JP. The biology of aging in a social world: insights from free-ranging rhesus macaques. bioRxiv [Preprint]. 2023 Jan 29:2023.01.28.525893. [CrossRef]
- Eisner JR, Dumesic DA, Kemnitz JW, Colman RJ, Abbott DH. Increased adiposity in female rhesus monkeys exposed to androgen excess during early gestation. Obes Res. 2003,11(2):279-286. [CrossRef]
- Barnett DK, Abbott DH. Reproductive adaptations to a large-brained fetus open a vulnerability to anovulation similar to polycystic ovary syndrome. Am J Hum Biol. 2003,15(3),296-319. [CrossRef]
- Bruns CM, Baum ST, Colman RJ, Dumesic DA, Eisner JR, Jensen MD, Whigham LD, Abbott DH. Prenatal androgen excess negatively impacts body fat distribution in a nonhuman primate model of polycystic ovary syndrome. Int J Obes (Lond). 2007,31(10),1579-1585. 10. [CrossRef]
- Zhou R, Bruns CM, Bird IM, Kemnitz JW, Goodfriend TL, Dumesic DA, Abbott DH. Pioglitazone improves insulin action and normalizes menstrual cycles in a majority of prenatally androgenized female rhesus monkeys. Reprod Toxicol. 2007,23(3),438-448. [CrossRef]
- Abbott DH, Barnett DK, Bruns CM, Dumesic DA. Androgen excess fetal programming of female reproduction: a developmental aetiology for polycystic ovary syndrome? Hum Reprod Update. 2005,11(4),357-374. [CrossRef]
- Keller E, Chazenbalk GD, Aguilera P, Madrigal V, Grogan T, Elashoff D, Dumesic DA, Abbott DH. Impaired preadipocyte differentiation into adipocytes in subcutaneous abdominal adipose of PCOS-like female rhesus monkeys. Endocrinology. 2014,155(7),2696-2703. [CrossRef]
- Barrett ES, Hoeger KM, Sathyanarayana S, Abbott DH, Redmon JB, Nguyen RHN, Swan SH. Anogenital distance in newborn daughters of women with polycystic ovary syndrome indicates fetal testosterone exposure. J Dev Orig Health Dis. 2018,9(3),307-314. 2018; 9, 3, 307–314. [CrossRef]
- Sánchez-Ferrer ML, Mendiola J, Hernández-Peñalver AI, Corbalán-Biyang S, Carmona-Barnosi A, Prieto-Sánchez MT, Nieto A, Torres-Cantero AM. Presence of polycystic ovary syndrome is associated with longer anogenital distance in adult Mediterranean women. Hum Reprod. 2017,32(11),2315-2323. 11. [CrossRef]
- ata B, Mimouni NEH, Barbotin AL, Malone SA, Loyens A, Pigny P, Dewailly D, Catteau-Jonard S, Sundström-Poromaa I, Piltonen TT, Dal Bello F, Medana C, Prevot V, Clasadonte J, Giacobini P. Elevated prenatal anti-Müllerian hormone reprograms the fetus and induces polycystic ovary syndrome in adulthood. Nat Med. 2018,24(6):834-846. [CrossRef]
- Abbott DH, Tarantal AF, Dumesic DA. Fetal, infant, adolescent and adult phenotypes of polycystic ovary syndrome in prenatally androgenized female rhesus monkeys. Am J Primatol. 2009,71(9):776-784. [CrossRef]
- Susa JB, Neave C, Sehgal P, Singer DB, Zeller WP, Schwartz R. Chronic hyperinsulinemia in the fetal rhesus monkey. Effects of physiologic hyperinsulinemia on fetal growth and composition. Diabetes. 1984,33(7):656-660. [CrossRef]
- Warren WC, Harris RA, Haukness M, Fiddes IT, Murali SC, Fernandes J, Dishuck PC, Storer JM, Raveendran M, Hillier LW, Porubsky D, Mao Y, Gordon D, Vollger MR, Lewis AP, Munson KM, DeVogelaere E, Armstrong J, Diekhans M, Walker JA, Tomlinson C, Graves-Lindsay TA, Kremitzki M, Salama SR, Audano PA, Escalona M, Maurer NW, Antonacci F, Mercuri L, Maggiolini FAM, Catacchio CR, Underwood JG, O'Connor DH, Sanders AD, Korbel JO, Ferguson B, Kubisch HM, Picker L, Kalin NH, Rosene D, Levine J, Abbott DH, Gray SB, Sanchez MM, Kovacs-Balint ZA, Kemnitz JW, Thomasy SM, Roberts JA, Kinnally EL, Capitanio JP, Skene JHP, Platt M, Cole SA, Green RE, Ventura M, Wiseman RW, Paten B, Batzer MA, Rogers J, Eichler EE. Sequence diversity analyses of an improved rhesus macaque genome enhance its biomedical utility. Science. 2020,370(6523),eabc6617. [CrossRef]
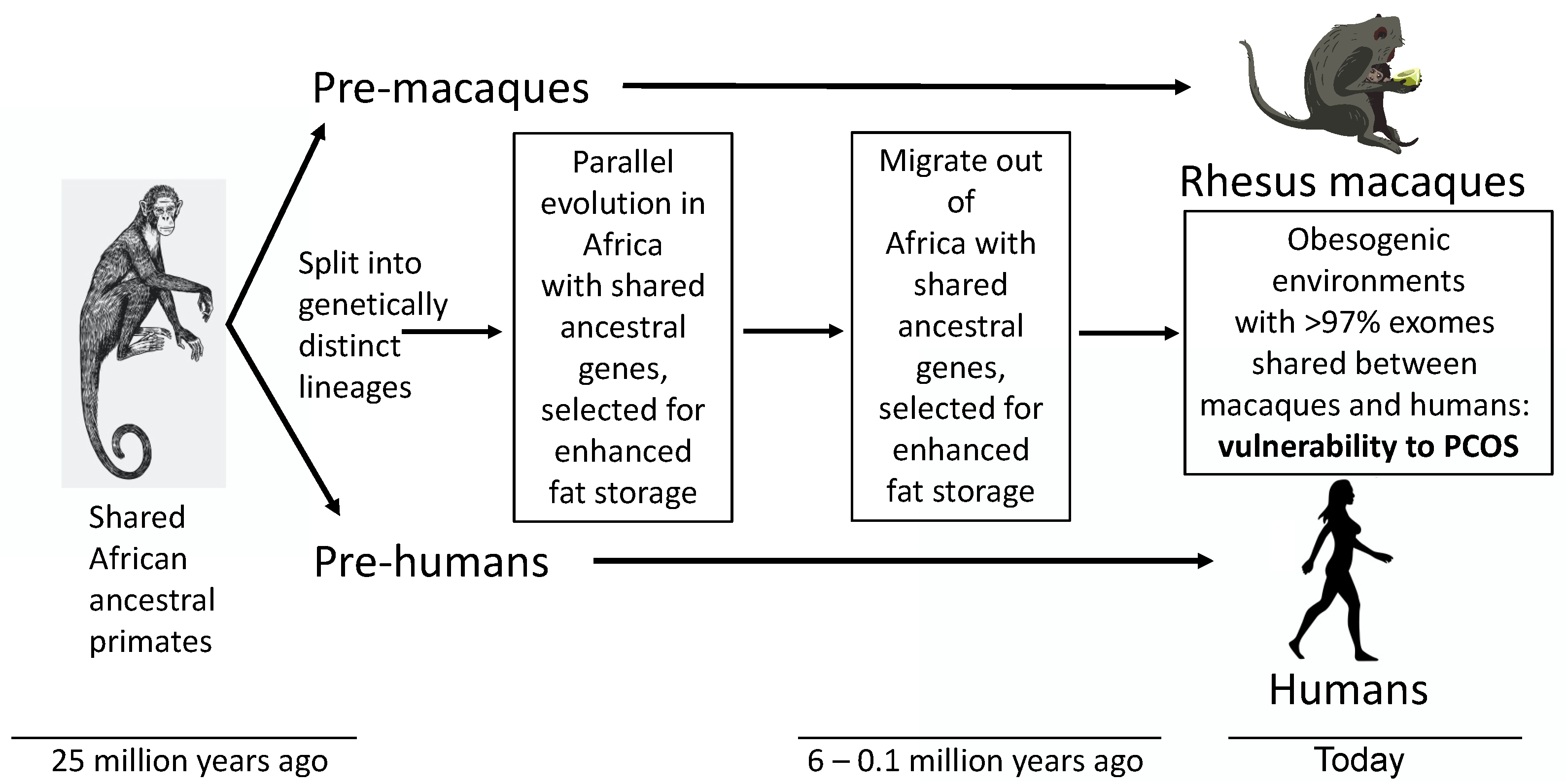
Figure 1.
Polycystic ovary syndrome as an ancient metabolic-reproductive adaptation that originally enhanced fat storage for survival of humans during ancient times of food deprivation, and also favored fewer offspring with a greater likelihood of childhood survival, but now predisposes to endocrine-reproductive dysfunction in today’s obesogenic environment (16). Ancestral traits resembling PCOS also exist in female rhesus macaques [8,9,10] that share a common ancestor with humans through parallel evolution.
Figure 1.
Polycystic ovary syndrome as an ancient metabolic-reproductive adaptation that originally enhanced fat storage for survival of humans during ancient times of food deprivation, and also favored fewer offspring with a greater likelihood of childhood survival, but now predisposes to endocrine-reproductive dysfunction in today’s obesogenic environment (16). Ancestral traits resembling PCOS also exist in female rhesus macaques [8,9,10] that share a common ancestor with humans through parallel evolution.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



