Submitted:
17 August 2023
Posted:
21 August 2023
You are already at the latest version
Abstract
Signaling modules that integrate the diverse extra- and intracellular inputs to control cell proliferation are essential during both development and adult stages to guarantee organism homeostasis. Mobs are small adaptor proteins that participate in several of these signalling pathways. Here we review recent advances unraveling Mob4 cellular functions, a highly conserved non-catalytic protein, that plays a diversity of roles in cell proliferation, sperm cell differentiation and simultaneously is involved in synapse formation and neural development. In addition, the gene is often overexpressed in a wide range of tumors and is linked to poor clinical outcomes. Nevertheless, Mob4 molecular functions remain poorly defined, although it integrates the core structure of STRIPAK, a kinase/phosphatase protein complex, that can act upstream of the Hippo pathway. In this review we focus on the recent findings of Mob4 functions, that have begun to clarify its critical role on cell proliferation and development of tissues and individuals.
Keywords:
Mob4
; Cell proliferation
; neurogenesis
; spermatogenesis
; Hippo pathway
; STRIPAK
; MOB-like protein
1. Introduction
Tissue homeostasis requires a fine balance between cellular proliferation, diferentiation and cell death. Throughout evolution, several mechanisms have evolved to ensure the fine-tuning of cell proliferation in multicellular organisms. The Mps-One Binder (MOB) family of genes encodes for a highly conserved group of proteins with a central role in regulating some of these mechanisms [1,2]. Mob1p, initially identified in S.cerevisiae, was the first MOB gene to be identified and described to be a part of the mitotic exit network (MEN) through its interaction with Dbf2-like kinases [3,4]. In the last twenty five years, the sequencing of several model organisms has led to the identification of several MOB homologues. In eukaryotes, the MOB family is divided into four classes – Mob1, Mob2, Mob3 and Mob4 – with non-overlapping functions. Seven MOB genes are encoded in the human genome, all with a high degree of sequence similarity [5,6]. Molecularly, MOB protein act as adaptor proteins without catalitical activity, that can bind kinases and modulate their activity.
Mob1, the first MOB identified in metazoans, functions as a core component of the Hippo signaling pathway [7], which regulates cell proliferation and organ size [8,9]. Interestingly, the other members of the MOB family can also modulate the activation of the Hippo signaling pathway. For example, Mob2 displays an antagonist function of Mob1 by negatively restricting Hippo signaling [10,11]. On the other hand, in response to apoptotic stimuli and cell-cell contact, Mob3 protects against the induction of apoptosis, thereby sustaining cell proliferation and tumor growth [12]. Finally, Mob4, the focus of this review, can compete with MOB1, thereby restricting Hippo signaling.
The Hippo signaling pathway is a major regulator of cell proliferation, organ size, cellular homeostasis and regeneration. The pathway is evolutionarily conserved and is modulated by a variety of signals such as cell-cell contact, ligands of G-protein coupled receptors, cell polarity, mechanical cues, and cellular energy status [13]. The core of Hippo pathway is composed of a kinase cascade wherein MST1/2 kinases phosphorylates and activates the complex formed by LATS and Mob1, that in turn phosphorylates and inactivates the oncoprotein YAP/TAZ that promotes the expression of cell proliferative and antiapoptotic genes [13].
The Hippo pathway is negatively regulated by a large protein complex named STRIPAK (Striatin-Interacting Phosphatase and Kinase complex). Core members of STRIPAK include the catalytical (PP2A-C) and scaffolding (PP2A-A) subunits of serine/threonine protein phosphatase 2A (PP2A); MST3/4 kinases and the adaptor proteins CCM3, Mob4 and STRIP1/2 [14,15,16]. The STRIPAK complex can also associate with a number of proteins in different signaling pathways, such as dynein, sarcolemal membrane–associated protein (SLMAP), and cortactin-binding proteins, presumably targeting them for phosphorylation and/or dephosphorylation [17,18]. The diversity of the STRIPAK-associated proteins suggests a key roles for STRIPAK in a range of biological systems [19,20] Importantly, MOB4 as a core component of the STRIPAK complex, can restrict Hippo signaling through this second mechanism.
2. MOB4: from the gene to the function
Initially named as Phocein [21] it was soon renamed Mob4 because of the high homology it displays with other MOB proteins. The Mob4 gene has been referred in the literature as Mob1, 2C4D, CGI-95, Class II mMOB1, Mob1 homolog 3, Mob3, Mps one binder kinase activator-like 3 (MOBKL3), and Preimplantation protein 3 (PREI3). In this work, we follow the HUGO Gene Nomenclature Committee (HGNC) and use MOB4 as the product of the gene ID 17261 (NCBI Entrez Gene: 25843).
Structurally, Mob4 adopts the canonical MOB fold that consists of a four-helix bundle at its core with three short α-helices at the N-terminal extension (aa 1−61) [22]. Of note, Mob4 share a high structural homology with Mob1, even at the N-terminal, a region where the various MOB proteins diverge.
The human Mob4 gene is predicted to generate alternatively spliced transcriptional variants producing three predicted protein isoforms: a canonical isoform with 26 KDa (isoform 1) and two smaller variants. One variant results from an alternative exon, containing a different start codon and thus generating a smaller protein with a different N-terminal with 22.3 KDa (isoform 2); and a second variant lacking an in-frame exon generating an isoform with 23.5 KDa (isoform 3) but sharing the same N- and C-termini as the canonical isoform (Figure 1). Like all MOB proteins, Mob4 has a high degree of conservation across evolution; for example there is 80% aminoacid (aa) identity and 88% similarity between Drosophila melanogaster Mob4 (dMob4) and its human ortholog (hMob4).
3. Neuronal functions of Mob4
Mob4 was initially identified on screenings in neural cell libraries as a potential interactor of striatins, by the groups of Dr. Ariane Monneron and of Dr. David Pallas, independently [21,23]. These initial biochemical studies led to the characterization of Mob4 as a core component of the STRIPAK complex (Figure 2) and indicated that Mob4 had a putative role in neuronal function. In agreement with this idea, it was found that mammalian Mob4 is highly expressed in melanized dopamine neurons as well as in the central and peripheral human nervous system [21,24]. In addition, it is highly enriched in dendritic spines, the actin-rich protrusions emerging from dendrites [24,25,26].
Immunogold labeling detected Mob4 in close vicinity to endocytic-like membranes in the neuronal dendritic spines suggesting vesicular trafficking functions [24,25]. Immunocytochemical studies also confirmed that Mob4 is strictly somato-dendritic extending down to the neuronal spines [21]. Noticeably, Mob4 is detected in cell bodies and dendrites but not in axons [21] , similarly to what has been described for striatin. In addition, immunoreactivity for Mob4 was only detected in neurons, not in glial cells. Recents studies in zebrafish confirms that Mob4 is highly expressed in the central nervous system: Mob4 is detected in the full extension of the neuroectoderm and in the brain in the initial stages of zebrafish development. Prior to hatching, mob4 is almost exclusively expressed in the brain and its expression remains enriched in the brain after hatching [27]. A neural role for Mob4 was first demonstratedby genetic studies in Drosophila, where it was shown to be a key regulator of neuronal structure and development [26]. Importantly, these studies showed that Mob4 is an essential gene, as Drosophila individuals lacking Mob4 do not survive past the larval stages. Selective somatic downregulation of Drosophila Mob4 resulted in a disruption of neuronal morphology. Neurons that developed without Mob4 showed abnormal branching patterns, hyperbranching and established incorrect connections to target cells. In addition, lack of Mob4 resulted in severe defects in microtubule organization, synaptic development and axonal transport. Finally, mutant Drosophila individuals display a defective neuronal transport of cargo from the cell body to the synapses [26]. DMob4 mutants have neuromere clusters smaller in size but increased complexity of the neurite branching with shorter neurites and an increased thickness of neurite bundles.
Synapses are the principal sites of neuronal cell-cell communication in the brain. These cell contacts are highly adaptable structures that are assembled, pruned, and modified throughout our lives. At a cellular level, most excitatory synapses in the central nervous system occur onto protrusions from neuronal dendrites called dendritic spines. The association of mammalian Mob4 with endocytic-like membranes and neuronal spinessuggests a function on endocytosis and vesicular trafficking [24,25]. Intracellular traffic depends on cytoskeleton functioning as rails for the cargo. Mutant Mob4 individuals have a disorganized microtubule cytoskeleton [26] indicating a possible association of Mob4 function with cytoskeleton organization and stability.
Recently, Florindo et al. have looked for a role of Mob4 in vertebrate neurodevelopment by addressing the consequences of the downregulation of Mob4 in the initial stages of zebrafish embryogenesis. They found that knockdown of zfMob4 using translation-blocking morpholinos in young embryos lead to severe neurologic defects. Zebrafish embryos at 24 hours post-fertilization lacked the midbrain-hindbrain boundary and showed reduced eye size. A quantitative analysis on the morphological consequences of mob4 downregulation revealed that Mob4 morphants are significantly smaller than their control morpholino siblings. Notably, these differences are not found in the axial trunk but only in the head [27]. This evidence strongly supports that Mob4 action is related to neurodevelopment of the brain but not the spinal cord. In addition, the authors have observed that, in these morphant embryos, the number of cell divisions in the hindbrain and eye are severely diminished. The hindbrain region is also significantly smaller in mob4 morphants arguing that the reduced hindbrain size is a consequence of impaired cell divisions. The developing eye had also a much smaller size than controls, as well as a severe decrease in the number of mitotic cells [27]. Altogether, these results strongly suggests that Mob4 is involved in cell proliferation during neurodevelopment in vertebrates. In agreement with these observations for Mob4 and supporting such functions in neurons, it has been shown that the STRIPAK complex promotes the organization, development and maturation of striatal neurons. In fact, dysfunction of STRIPAK has been linked to a range of clinical neurological conditions [28]. For example, knockdown of Striatin, that binds directly to Mob4 (FIGURE) , blocks dendrite formation [29], an observation that suggests that knockdown of Mob4 affects neuronal morphology at least in part by affecting STRIPAK function.
Taken these observations altogether, Mob4 seems to be required for the regulation of neuronal functions at different levels. Firstly, by being required for the assembly of a normal microtubule cytoskeleton [26]. Second, biochemical studies indicated Mob4 to be involved in endocytosis and vesicular trafficking [24,25,26]. It seems therefore that Mob4 contributes both to postnatal synaptogenesis and to the dendritic activity dependent plasticity in the adult. It is noteworthy to recall that Mob4 is highly conserved throughout the animal kingdom. For such, it is tempting to speculate that a deficient MOB4 function may be related to mechanisms of neurological diseases in humans.
4. Mob4 and cytoskeleton
Recent reports indicate that Mob4 has important roles in cytoskeletal regulation. Studies in Drosophila cells, reported mob4 to be involved in mitotic spindle microtubule focusing [Vale 2009], while studies in the fly nervous system showed Mob4 to be important for the organization of microtubule networks within postmitotic neurons [26]. A recent report shows that in Zebrafish, Mob4 function is required for the incorporation of skeletal muscle α-actin into organised sarcomeres [31]. In this work, the authors found an interaction between Mob4 and the actin-folding chaperonin TriC, suggesting that Mob4 impacts on TRiC to control actin biogenesis and thus myofibril growth. This report supports a previously established molecular interaction between Mob4 and TriC, in human cells [14].
The proper folding of proteins is essential to cellular function and, consequently, defective protein folding is linked to multiple pathological disorders. In eukaryotes, folding of misfolded or unfolded proteins is mediated by molecular chaperones, one of which is the protein complex TRiC (TCP-1 ring complex, also called chaperonin containing TCP-1). Cytoplasmic chaperonins are key elements for the proper folding of proteins. TRiC chaperonin display limited substrate specificity and is required for folding of essential proteins, including the cytoskeletal proteins myosin, actin and α- and β-tubulin [32,33,34]. The importance of TRiC complex in protein folding was first shown in Caenorhabditis elegans where individuals with reduced TRiC function display defective microtubule cytoskeletons [35]. Co-immunoprecipitations assays had shown that Mob4 and TriC are part of a multiprotein complex and that a direct interaction between Mob4 protein and TriC was demonstrated to occur in nematods [14,36]. Recently, Berger et al. showed that Zebrafish mob4 mutants also display defective microtubule networks, which is in-line with tubulin being a main folding substrate of TRiC. Therefore, it is possible that such defective microtubule networks are the cause of the observed compromised neuronal connectivity in zebrafish mob4 mutantes. In line with these observations, neuronal neurite formation is severely reduced in TRiC-deficient zebrafish [32,33].
Additionally, Berger et al. also found that within wildtype zebrafish, Mob4 protein localises to the sarcomere’s Z-discs, co-localising with the marker α-actinin. Importantly, whereas loss of Mob4 function lead to a reduction in the amount of myofibril, increased expression of Mob4 induced an increase in the amount of organised myofibrils. These findings indicate that Mob4 function might be required for the regulation of the amount of organised myofibril. Lack of Mob4 function results in reduced numbers of myofribrils and impaired movement due to skeletal muscle defects.
As Mob4 functions within the protein complex STRIPAK, Berger and collegues looked for similar defects in strn3-deficient mutants. In fact, the authors found that strn3-deficient mutants featured both neuronal as well as muscle defects. These observations confirm not just Mob4 as a core component of STRIPAK in Zebrafish but also imply a role for the STRIPAK complex in sarcomerogenesis. Surprisingly, zebrafish TRiC mutants develop into relatively normal larvae although with highly specific skeletal muscle defects, resulting from impaired folding of α-actin at Z-disks in the skeletal muscle, with the result of a reduced sarcomere assembly.
The work of Berger et al. highlight the fact that Mob4 function involves, at least two different protein complexes, the STRIPAK and the TRiC. The first with a large diversity of cellular functions and the later required for actin and tubulin biogenesis. Whereas actin is the main component of the muscle’s thin filaments, tubulin constitutes the microtubule network essential for neuronal axons. It is therefore no surprise that mutants in mob4 and strn3 featured neuronal as well as muscle defects, leading the authors to conclude that the two protein complexes STRIPAK and TRiC may interact through Mob4 to coordinate growth of the myofibril and of microtubule network in neural development.
5. Cell proliferation function of Mob4
From a physiological point of view, Mob4 plays important roles in the control of cell proliferation. As previously mentioned, downregulation of zfMob4 results in a severe decrease in the number of mitotic cells in the brain and in the developing eye [27]. Control of stem cell number also seems to be one function of Mob4, as studies in planarian indicated that mob4 function limits body size through limiting stem cell numbers [37]. Planarians are well known for their ability to regenerate body parts after injury using neoblast adult pluripotent stem cells and keeping body proportionality [38]. The inhibition of mob4 in planaria dramatically increased posterior length after injury, afecting the polarity along the apical posterior axis [37].
This an important step forward in deciphering the molecular mechanisms that allow animals to reconstruct body parts while simultaneously integrating newly regenerated tissues into pre-existing old ones after injury. The process of regeneration in planarians is complex and involves the regulation of multiple cellular signalling pathways essential for normal physiological functions, such as asymmetric division, cell cycle, apoptosis, tissue/organ size and polarity determinations [39]. One such signalling pathway is the evolutionarily conserved Wnt pathway, which is involved in a myriad of processes namely cell proliferation, differentiation, and apoptosis, as well as in stem cell maintenance [40]. In planaria, the Wnt signalling pathway is the key regulator of the head-tail polarity, and it is at the core of the decision-making process to regenerate a head or a tail [41,42; reviewed in 39]. Constitutive localised expression of Wnt ligands (posteriorly) and of Wnt inhibitors (anteriorly) control regionalization of the planarian Antero-Posterior axis. The results obtained by REF et al, support a model in which wnt pole cells determine tail proportionality along the Antero-Posterior axis, with mob4 controlling wnt pole cell numbers through limiting stem-cell-dependent production [37]. These results therefore indicate that mob4 is involved in the scaling of tail size with respect to body size via regulation of wnt1. Considering that Mob4 is constitutively expressed, even when no injury has occurred, the recovery of normal proportions through regeneration might then involve a process that achieves a stable equilibrium of local signaling processes. The authors results indicate that Mob4 (and STRIPAK) suppression of Wnt signaling is a critical pathway regulating whole-body proportional scaling in planarians. Interestingly, planarian Hippo pathway components do not seem to function within antero-posterior tissue proportionality suggesting that Mob4 and STRIPAK exert their action independently of the Hippo signaling pathway. On the other hand, a crosstalk between the Hippo pathway and the Wnt pathway has been previously described [43], and Hippo plays an important role in the regulation of the cell cycle, equally crucial for planarian regeneration.
Importantly, MOB proteins have been demonstrated to be involved in defining cell polarity both in humans and in Tetrahymena [44,45]. The importance of cell polarity in a number of physiological processes, including cell diferentiation, cell migration, asymmetric cell division, cancer progression and immune response, has been extensively described [reviewed in [46]]. In Tetrahymena, an unicellular organism, the single Mob protein encoded in the genome, is required for correct division plane placement by establishing the anterior-posterior axis. Downregulation of Mob in Tetrahymena induces a misplacement of the division plane with consequent abcission failure; daughter cells fail to separate and form trails of interconnected abnormal cells [44]. Interestingly, the authors found that the Mob protein accumulates at the future site of cell division prior to constriction start, this way defining the anterior and posterior end of the future new daughter cells. This finding highlights the importance of Mob in cell polarity inception through a cell-intrinsic mechanism and how polarity is coupled to growth in Tetrahymena. Likewise, in human HeLa cultured cells, downregulation of Mob1 also results in abscission failure and, importantly, cell polarity is affected in such a way that allows cells to become motile [45].
Finally, a role for MOB4 in cell proliferation and tissue formation has also been described in the filamentous fungi Sodaria macrospora, where downregulation of SmMob3 (the MOB4 homologue in Sodaria) results in impaired vegetative growth accompanied by a sexually sterile strain unable to undergo self-fusion and fusion [47]. On the other hand, in Caenorhabditis elegans, mob-4 deficient mutant individuals do not show an obvious abnormal phenotype under the normal growth conditions. Nevertheless, life span and thermotolerance are compromised in these individuals [48]. These observations, together with a previous report showing that C. elegans YAP-1 overexpression shortens life span (whereas yap-1 deficiency promotes health life span) [49] suggests that, in contrast to humans and Drosophila where Mob4 activates YAP1 [50], in worms mob-4 does not act upstream of Yap-1. Taken together, these observations led to suggestion that in C. elegans the Hippo pathway is not conserved [49].
MOB4 may be involved in cell proliferation through it’s function within the STRIPAK complex or indenpendently of it. STRIPAK negatively regulates the Hippo signaling pathway thereby participating in the control of cell proliferation [28,51,52]. Deletion of the N-terminal residues of Mob4 abolishes the formation of STRIPAK [22] and disruption of the interacting sites between Mob4 and STRN3 causes aberrant regulation of Hippo signaling. Thus, Mob4 can affect cell proliferation by participating in STRIPAK assembly and Hippo pathway activity.
Given the role of Mob4 in regulating cell proliferation it is not surprising that Mob4 may be involved in cancer initiation/progression. Cancer development, due to excessive cell proliferation, is highly associated to the activation of oncogenic pathways [53,54,55] or the deregulation of genes with tumour suppression functions [56,57,58,59]. There is a moderate/high protein expression of Mob4 in most human cancers and a higher expression of Mob4 is associated with a poor prognosis in the cases of renal and liver cancers (https://www.proteinatlas.org). The Hippo pathway, per se, is an important regulator of cell proliferation and tissue growth, and mice mutant in Hippo components (Sav, Mst1/2, Lats and Mob1) are prone to develop malignant growths [60,61,62,63]. In addition, is has been described that the different MOB family members behave either as tumor suppressors or oncogenes: the complete loss of Mob1, a component of the Hippo pathway, in mice promotes tumorigenesis and embryonic death [60]; Mob2 has been recently reported as a tumor suppressor in glioblastoma [64]; in contrast, Mob3 has been found upregulated in glioblastoma multiforme and it is proposed as oncoprotein by suppressing MST1 activity [12].
Interestingly, Mob4 can form a complex with the protein kinase MST4 forming a complex that antagonizes the complex formed by Mob1 and the kinases MST1/2 [50] (Figure 3). But while MST1-Mob1 acts as a tumor suppressor, MST4-Mob4 is oncogenic by activating YAP signaling and promoting cell proliferation. In fact, Mob4 can alternatively pair with either MST4 or MST1 due to the high structural similarities of both MOBs and of both MST kinases. Mob4 can therefore sequester MST1, consequently inhibiting the Hippo pathway and promoting cell proliferation [50], this way acting as an oncogene. It is worth remembering that simultaneously, on an alternative mechanism and as a component of STRIPAK complex, Mob4 is a negative regulator of the Hippo signaling pathway and therefore actsas a tumour suppressor gene.
6. Mob4 and spermatogenesis
Spermatogenesis is the process by which germinal stem cells give rise to haploid spermatozoa. In Drosophila, the morphogenesis of spermatids generally takes place within a syncytium, in which all spermatid nuclei remain interconnected via an extensive network of cytoplasmic bridges. As spermiogenesis progresses, the physical resolution of the syncytium (or cyst) into individual cells is necessary, a process referred to as sperm individualization. Although with some differences in the control of hormonal regulation and in testicular structure, the different stages of spermatogenesis are highly conserved from fly to human. Importantly, many of the genes involved in Drosophila spermatogenesis were shown to be conserved in humans.
Santos et al. recently described a requirement for Mob4 during spermatogenesis in Drosophila: males without Mob4 function in the gonads are sterile while females are fully fertile. In the absence of mob4 function cyst elongation occurs, meaning spermatids capable of elongating an axoneme, but spermatid individualization fails. Consequently, migration of sperm into the seminal vesicle does not occur and males aresterile. To determine defects in spermiogenesis underlying the failure in spermatid individualization the authors have examined the ultrastructure of the developing axoneme and found that the formation of the mitochondrial derivatives is defective. Importantly, defects in the axonemal structure were also found. These defects include loss of microtubule doublets and, most interestingly, preservation of stereotypical 9+2 microtubule-doublets of the axoneme but with large radial expansion [65].
The use of a GFP:Mob4 transgene reveals that Mob4 has a dynamic sub-cellular localization in different cell types throughout spermatogenesis. In meiotic cells, Mob4 showsa reticular localization accumulating in membranous fibers surrounding the meiotic spindle structures. Later, durind the early canoe stage of spermatid differentiation, Mob4 strongly accumulates in individual punctae at the basal side of nuclei, in close vicinity to the basal body. This punctate localization is transientand can only be observed in this particular stage of spermatid differentiation. Such a dynamic behaviour of Mob4 at different stages of spermatogenesis is suggestive of multiple functions in the parafusorial membranes and associated microtubules in meiosis, at the basal body or transition zone in the initiation of axoneme elongation, and in the cystic bulge during individualization per se.
As referred, spermatogenesis is a highly conserved process, and many of the genes involved in spermatogenesis have their function conserved across species. Mob4 seem to follow the trend as insertion of the human Mob4 paralog gene into Drosophila was capable of rescuing all the meiotic defects in mob4 mutant, including full fertility [65], suggesting that human Mob4 and Drosophila Mob4 are functionally equivalent orthologs.
Considering that Mob4 is a component of STRIPAK, one may wonder if the defective spermatogenesis results from lack of STRIPAK function, or is an isolated function of Mob4. To answer this question Santos et al. looked for a function of Strip and Cka (two other components of STRIPAK) in Drosophila testes. They found that, like for Mob4, either Strip or Cka downregulation in testes results equally in male sterility; on the other hand, Strip or Cka downregulation did not seem to affect female ovaries and are not required for female oogenesis,. In addition, the investigators also showed that downregulation of either Strip or Cka results in similar failures in sperm individualization to Mob4, suggesting that STRIPAK complex activity is required for spermatogenesis and that Mob4 is probably acting through STRIPAK.
- Other Mob4 functions
A different set of studies indicated that Mob4 is also required for mitosis progression. In Drosophila S2 cultured cells, Mob4 was found to associate with centrioles and kinetochores and its downregulation resulted in the formation of monopolar spindles and defective mitosis [30]. Moreover, in human cells Frost and collaborators showed that the downregulation of Mob4 causeds DNA content increase, abnormal spindle formation and mitosis failure triggering cell death [66]. In addition the authors found that Mob4, together with the STRIPAK complex (Figure 3), bridges the cis-Golgi, the centrosome, and the outer nuclear membrane [66]. Disruption of these bridges may be the cause of the defective mitotic progression.
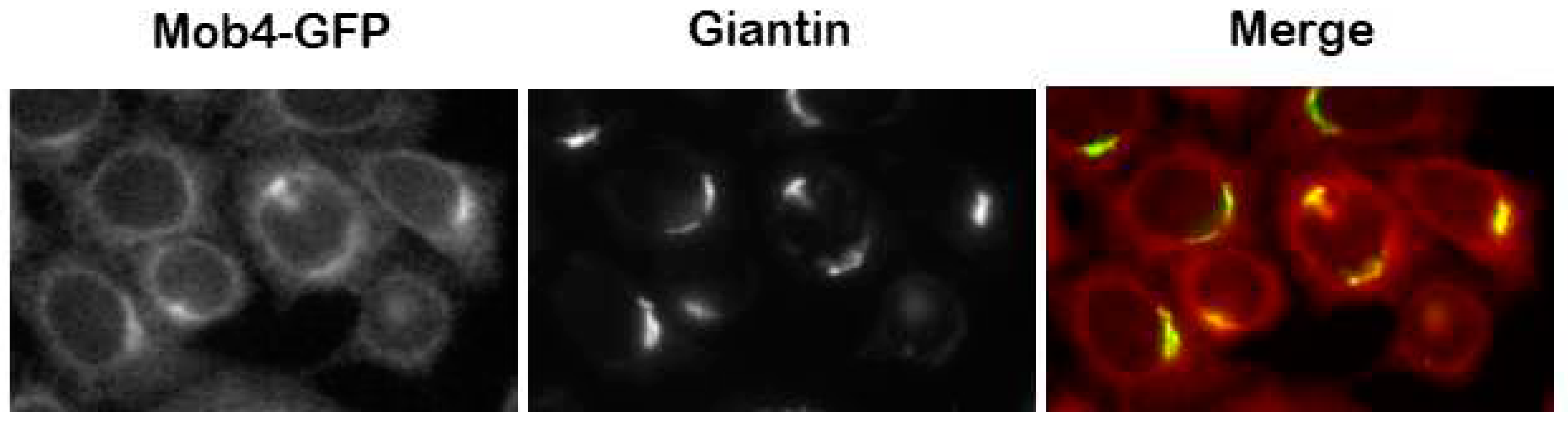
Figure 4.
Mob4 accumulates on the Golgi complex in human cells (Baillat 2001 [13]). In the figure, HeLa cells stably expressing Mob4-GFP were treated for immunofluorescence with anti-giantin antibody for identification of the Golgi complex. (photos by Andreia Colaço and Álvaro Tavares).
Figure 4.
Mob4 accumulates on the Golgi complex in human cells (Baillat 2001 [13]). In the figure, HeLa cells stably expressing Mob4-GFP were treated for immunofluorescence with anti-giantin antibody for identification of the Golgi complex. (photos by Andreia Colaço and Álvaro Tavares).
Human Mob4 may also have a role in DNA damage signaling since its downregulation results in γH2AX phosphorylation in cells [67], which is an early marker for DNA damage [68]. Moreover, Ser147 of human Mob4 is a possible target of ATM kinase suggesting a possible role for Mob4 within the DNA damage response [69]. Other MOB family members are also involved in the DNA damage signaling. For example, Mob2 interacts with RAD50 promoting the assembly of the MRE11-RAD50-NBS1 (MRN) DNA damage sensor complex and activates ATM kinase, a well-known protein orchestrating DNA damage response. Mob2 can also compete with Mob1 for NDR1/2 binding, and NDR-mediated phosphorylation plays a role in the G2/M DNA damage checkpoint by promoting the degradation of the CDC25A phosphatase [70].
Apoptosis is a process important for the maintenance of cell numbers, and it’s regulation has been previously associated with Mob4 and Mob1 due to their association with MST1 kinase. MST1 modulates oxidative stress-induced neuronal death [71] and several studies have reported how MST1 phosphorylation of FOXO proteins enhances their nuclear translocation promoting the transcription of apoptosis-related genes [[72], Valis 2011]. A neuronal-specific isoform of YAP (the end target of the Hippo pathway), YAPdeltaC, acts as neuronal apoptosis protector that decreases with progression for amyotrophic lateral sclerosis (ALS) whereas the full-length YAP remains constant during the late and severe stage of the disease [74].
Specific connections between MOB proteins and neurodegeneration can be established through their interaction with NDR/LATS kinases. All four Drosophila MOB genes genetically interact with tricornered (Trc) (Drosophila homologue of NDR kinases) [75]. Trc is required for morphological changes such as outgrowth of epidermal hair and dendritic tiling in sensory neurons and Wts (Drosophila homologue of LATS) play a role in dendritic maintenance in sensory neurons [76,77,78]. Mob2 has been directly associated to neuronal functions. In neuronal cell lines, Mob2 is required for sustaining neurite formation [79]. Mob2 expression is required to regulate the growth of larval neuromuscular junction in Drosophila [80], and for normal neuronal distribution in mice developmental cortex [81]. In addition, human Mob2 has been identified as a specific protein of cerebral amyloid angiopathy (CAA), a condition where amyloid plaques deposits on the walls of cortical and leptomeningeal blood vessels of the brain [82]. On the other hand, increasing evidence shows a potential role for Mob3 in neurodegenerative diseases. Human Mob3A gene was identified as a target for the nuclear respiratory factor 1 (NRF1) [83], a transcription factor that activates the expression of a wide range of nuclear genes essential for mitochondrial biogenesis and function, whose function is severely compromised in the brains of aging-related neurodegenerative diseases [84]. On a second study, human Mob3 proteins were found to bind to the oligomeric Aβ42, a hallmark protein complex of Alzeimer’s disease, suggesting a potential involvement of Mob3 proteins with this condition [85]. Therefore Mob4 is not the only MOB with a role in the control of cell proliferation and apoptosis, relevant for the clearence of excessive neural cells during normal development [86].
In summary, proteins of the MOB family have important functions in the control of cell proliferation. Mob4 is fast becoming an important member of the family with a range of diverse essential functions for normal neuronal development, spermatogenesis and for the control of cell proliferation both in vertebrates and in Drosophila (Figure 5). These functions are most probably exerted through different mechanisms. Some of Mob4 action is exerted as a scaffold on the assembly of the STRIPAK complex (Figure 2 and Figure 3) and this way, like other STRIPAK components, participates in dendritic development, axonal transport, and synapse assembly. But the action of Mob4 seems to extend beyond STRIPAK functions, like interacting with the TriC complex, or direct interactions with MST1. Most human cancers show a moderate/high protein expression of Mob4, and renal and liver cancer patients with a high expression of Mob4 showed a reduced survival probability compared to those with low expression suggesting that Mob4 is an unfavorable prognostic marker for renal and liver cancers. Therefore, elucidating the underlying mechanisms of Mob4 action and regulation may be of help to identify novel therapeutic targets and diagnostic markers for cancer and neurodegeneration diseases those diseases.
Author Contributions
Original draft preparation, writing, review and editing: J.G-M., I.B.S., B.I.O., C.G. e A.A.T. Funding acquisition: A.A.T. All authors have read and agreed to the published version of the manuscript.
Funding
Work funded by Algarve 2020 Program, grant number ALG-01-0145-FEDER-030014, and co-financed by FEDER Funds through the Operational Program for Competitiveness Factors—COMPETE 2020 and by National Funds through FCT—Foundation for Science and Technology under the Project PTDC/BIA-CEL/30014/2017. I.B.Santos was funded by FCT fellowship SFRH/BD/141734/2018.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Delgado, I.; Carmona, B.; Nolasco, S.; Santos, D.; Leitão, A.; Soares, H. MOB: Pivotal Conserved Proteins in Cytokinesis, Cell Architecture and Tissue Homeostasis. Biology 2020, 9, 413. [Google Scholar] [CrossRef] [PubMed]
- Duhart, J.C.; Raftery, L.A. Mob Family Proteins: Regulatory Partners in Hippo and Hippo-Like Intracellular Signaling Pathways. Front. Cell Dev. Biol. 2020, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Luca, F.C.; Winey, M. Regulation of Mob1p, an essential budding yeast protein required for completion of mitosis and spindle pole body duplication. Mol. Biol. Cell 1998, 9, 12A. [Google Scholar] [CrossRef] [PubMed]
- Luca, F.C.; Winey, M. MOB1, an essential yeast gene required for completion of mitosis and maintenance of ploidy. Mol. Biol. Cell 1998, 9, 29–46. [Google Scholar] [CrossRef]
- Ye, X.; Nikolaidis, N.; Nei, M.; Lai, Z. Evolution of the mob Gene Family. The Open Cell Signaling Journal 2009, 1, 1–11. [Google Scholar] [CrossRef]
- Vitulo, N.; Vezzi, A.; Galla, G.; Citterio, S.; Marino, G.; Ruperti, B.; Zermiani, M.; Albertini, E.; Valle, G.; Barcaccia, G. Characterization and evolution of the cell cycle-associated mob domain-containing proteins in eukaryotes. Evol Bioinform Online 2007, 3, 121–158. [Google Scholar] [CrossRef]
- Lai, Z.C.; Wei, X.; Shimizu, T.; Ramos, E.; Rohrbaugh, M.; Nikolaidis, N.; Ho, L.L.; Li, Y. Control of cell proliferation and apoptosis by mob as tumor suppressor, mats. Cell 2005, 120, 675–685. [Google Scholar] [CrossRef]
- Pan, D. Hippo signaling in organ size control. Genes and Development. 2007, 21, 886–897. [Google Scholar] [CrossRef]
- Halder, G.; Johnson, R.L. Hippo signaling: Growth control and beyond. Development. 2011, 138, 9–22. [Google Scholar] [CrossRef]
- Weiss, E.L.; Kurischko, C.; Zhang, C.; Shokat, K.; Drubin, D.G.; Luca, F.C. The Saccharomyces cerevisiae Mob2p-Cbk1p kinase complex promotes polarized growth and acts with the mitotic exit network to facilitate daughter cell-specific localization of Ace2p transcription factor. J Cell Biol. 2002, 158, 885–900. [Google Scholar] [CrossRef]
- Kohler, R.S.; Schmitz, D.; Cornils, H.; Hemmings, B.A.; Hergovich, A. Differential NDR/LATS interactions with the human MOB family reveal a negative role for human MOB2 in the regulation of human NDR kinases. Mol Cell Biol. 2010, 30, 4507–4520. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Zhang, L.; Xue, G.; Hynx, D.; Wang, Y.; Cron, P.D.; Hundsrucker, C.; Hergovich, A.; Frank, S.; Hemmings, B.A.; Schmitz-Rohmer, D. hMOB3 modulates MST1 apoptotic signaling and supports tumor growth in glioblastoma multiforme. Cancer Res. 2014, 74, 3779–3789. [Google Scholar] [CrossRef]
- Ma, S.; Meng, Z.; Chen, R.; Guan, K.L. The Hippo Pathway: Biology and Pathophysiology. Annu Rev Biochem. 2019, 88, 577–604. [Google Scholar] [CrossRef] [PubMed]
- Goudreault, M.; D'Ambrosio, L.M.; Kean, M.J.; Mullin, M.J.; Larsen, B.G.; Sanchez, A.; Chaudhry, S.; Chen, G.I.; Sicheri, F.; Nesvizhskii, A.I.; Aebersold, R.; Raught, B.; Gingras, A.C. A PP2A phosphatase high density interaction network identifies a novel striatin-interacting phosphatase and kinase complex linked to the cerebral cavernous malformation 3 (CCM3) protein. Mol Cell Proteomics. 2009, 8, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Glatter, T.; Wepf, A.; Aebersold, R.; Gstaiger, M. An integrated workflow for charting the human interaction proteome: Insights into the PP2A system. Mol Syst Biol. 2009, 5, 237. [Google Scholar] [CrossRef]
- Ribeiro, P.S.; Josué, F.; Wepf, A.; Wehr, M.C.; Rinner, O.; Kelly, G.; Tapon, N.; Gstaiger, M. Combined functional genomic and proteomic approaches identify a PP2A complex as a negative regulator of Hippo signaling. Mol Cell. 2010, 39, 521–534. [Google Scholar] [CrossRef]
- Gordon, J.; Hwang, J.; Carrier, K.J.; Jones, C.A.; Kern, Q.L.; Moreno, C.S.; Karas, R.H.; Pallas, D.C. Protein phosphatase 2a (PP2A) binds within the oligomerization domain of striatin and regulates the phosphorylation and activation of the mammalian Ste20-Like kinase Mst3. BMC Biochem. 2011, 12, 54. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014, mutations, PTMs and recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef]
- Gil-Ranedo, J.; Gonzaga, E.; Jaworek, K.J.; Berger, C.; Bossing, T.; Barros, C.S. STRIPAK Members Orchestrate Hippo and Insulin Receptor Signaling to Promote Neural Stem Cell Reactivation. Cell Rep. 2019, 27, 2921–2933. [Google Scholar] [CrossRef]
- Shi, Z.; Jiao, S.; Zhou, Z. STRIPAK complexes in cell signaling and cancer. Oncogene. 2016, 35, 4549–4557. [Google Scholar] [CrossRef]
- Baillat, G.; Moqrich, A.; Castets, F.; Baude, A.; Bailly, Y.; Benmerah, A.; Monneron, A. Molecular cloning and characterization of phocein, a protein found from the Golgi complex to dendritic spines. Mol. Biol. Cell 2001, 12, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.C.; Bae, S.J.; Ni, L.; Zhang, X.; Bai, X.C.; Luo, X. Cryo-EM structure of the Hippo signaling integrator human STRIPAK. Nat Struct Mol Biol. 2021, 28, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.S.; Lane, W.S.; Pallas, D.C. A mammalian homolog of yeast MOB1 is both a member and a putative substrate of striatin family-protein phosphatase 2A complexes. J Biol Chem. 2001, 276, 24253–24260. [Google Scholar] [CrossRef] [PubMed]
- Haeberlé, A.M.; Castets, F.; Bombarde, G.; Baillat, G.; Bailly, Y. Immunogold localization of MOB4 in dendritic spines. J. Comp. Neurol. 2006, 495, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Bailly, Y.J.; Castets, F. Phocein: A potential actor in vesicular trafficking at Purkinje cell dendritic spines. Cerebellum. 2007, 6, 344–352. [Google Scholar] [CrossRef]
- Schulte, J.; Sepp, K.J.; Jorquera, R.A.; Wu, C.; Song, Y.; Hong, P.; Littleton, J.T. DMob4/Phocein regulates synapse formation, axonal transport, and microtubule organization. J Neurosci. 2010, 30, 5189–5203. [Google Scholar] [CrossRef]
- Florindo, C.; Mimoso, J.M.; Palma, S.L.; Gonçalves, C.; Silvestre, D.; Campinho, M.; Tavares, Á.A. Mob4 is required for neurodevelopment in zebrafish. MicroPubl Biol 2023, micropub.biology.000762. [Google Scholar] [CrossRef]
- Hwang, J.; Pallas, D.C. STRIPAK complexes: Structure, biological function, and involvement in human diseases. Int J Biochem Cell Biol. 2014, 47, 118–148. [Google Scholar] [CrossRef]
- Li, D.; Musante, V.; Zhou, W.; Picciotto, M.R.; Nairn, A.C. Striatin-1 is a B subunit of protein phosphatase PP2A that regulates dendritic arborization and spine development in striatal neurons. J Biol Chem. 2018, 293, 11179–11194. [Google Scholar] [CrossRef]
- Trammell, M.A.; Mahoney, N.M.; Agard, D.A.; Vale, R.D. Mob4 plays a role in spindle focusing in Drosophila S2 cells. J Cell Sci. 2008, 121, 1284–1292. [Google Scholar] [CrossRef]
- Berger, J.; Berger, S.; Currie, P.D. Mob4-dependent STRIPAK involves the chaperonin TRiC to coordinate myofibril and microtubule network growth. PLoS Genet. 2022, 18, e1010287. [Google Scholar] [CrossRef]
- Yaffe, M.B.; Farr, G.W.; Miklos, D.; Horwich, A.L.; Sternlicht, M.L.; Sternlicht, H. TCP1 complex is a molecular chaperone in tubulin biogenesis. Nature. 1992, 358, 245–248. [Google Scholar] [CrossRef]
- Berger, J.; Berger, S.; Li, M.; Jacoby, A.S.; Arner, A.; Bavi, N.; et al. In Vivo Function of the Chaperonin TRiC in alpha-Actin Folding during Sarcomere Assembly. Cell Rep. 2018, 22, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Ghozlan, H.; Cox, A.; Nierenberg, D.; King, S.; Khaled, A.R. The TRiCky Business of Protein Folding in Health and Disease. Front Cell Dev Biol. 2022, 10, 906530. [Google Scholar] [CrossRef] [PubMed]
- Saegusa, K.; Sato, M.; Sato, K.; Nakajima-Shimada, J.; Harada, A.; Sato, K. Caenorhabditis elegans chaperonin CCT/TRiC is required for actin and tubulin biogenesis and microvillus formation in intestinal epithelial cells. Mol Biol Cell. 2014, 25, 3095–3104. [Google Scholar] [CrossRef] [PubMed]
- Khabirova, E.; Moloney, A.; Marciniak, S.J.; Williams, J.; Lomas, D.A.; Oliver, S.G.; et al. The TRiC/CCT chaperone is implicated in Alzheimer’s disease based on patient GWAS and an RNAi screen in Abeta-expressing Caenorhabditis elegans. PLoS ONE. 2014, 9, e102985. [Google Scholar] [CrossRef]
- Schad, E.G.; Petersen, C.P. STRIPAK Limits Stem Cell Differentiation of a WNT Signaling Center to Control Planarian Axis Scaling. Curr. Biol. 2020, 30, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.E.; Wang, I.E.; Reddien, P.W. Clonogenic neoblasts are pluripotent adult stem cells that underlie planarian regeneration. Science. 2011, 332, 811–816. [Google Scholar] [CrossRef]
- Reddien, P.W. The Cellular and Molecular Basis for Planarian Regeneration. Cell 2018, 175, 327–345. [Google Scholar] [CrossRef]
- Clevers, H.; Loh, K.M.; Nusse, R. Stem cell signaling. An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Science. 2014, 346, 1248012. [Google Scholar] [CrossRef]
- Gurley, K.A.; Rink, J.C.; Sanchez Alvarado, A. Beta-catenin defines head versus tail identity during planarian regeneration and homeostasis. Science 2008, 319, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.P.; Reddien, P.W. Smed-betacatenin-1 is required for anteroposterior blastema polarity in planarian regeneration. Science. 2008, 319, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Sileo, P.; Simonin, C.; Melnyk, P.; Chartier-Harlin, M.C.; Cotelle, P. Crosstalk between the Hippo Pathway and the Wnt Pathway in Huntington's Disease and Other Neurodegenerative Disorders. Cells. 2022, 11, 3631. [Google Scholar] [CrossRef]
- Tavares, A.; Gonçalves, J.; Florindo, C.; Tavares, A.A.; Soares, H. Mob1, defining cell polarity for proper cell division. J Cell Sci. 2012, 125, 516–527. [Google Scholar] [CrossRef]
- Florindo, C.; Perdigão, J.; Fesquet, D.; Schiebel, E.; Pines, J.; Tavares, A.A. Human Mob1 proteins are required for cytokinesis by controlling microtubule stability. J Cell Sci. 2012, 125, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Piroli, M.E.; Blanchette, J.O.; Jabbarzadeh, E. Polarity as a physiological modulator of cell function. Front Biosci (Landmark Ed). 2019, 24, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Bernhards, Y.; Pöggeler, S. The phocein homologue SmMOB3 is essential for vegetative cell fusion and sexual development in the filamentous ascomycete Sordaria macrospora. Curr Genet. 2011, 57, 133–149. [Google Scholar] [CrossRef]
- Jahan, M.; Iwasa, H.; Kuroyanagi, H.; Hata, Y. Loss of Caenorhabditis elegans homologue of human MOB4 compromises life span, health life span and thermotolerance. Genes Cells. 2021, 26, 798–806. [Google Scholar] [CrossRef]
- Iwasa, H.; Maimaiti, S.; Kuroyanagi, H.; Kawano, S.; Inami, K.; Timalsina, S.; Ikeda, M.; Nakagawa, K.; Hata, Y. Yes-associated protein homolog, YAP-1, is involved in the thermotolerance and aging in the nematode Caenorhabditis elegans. Exp Cell Res. 2013, 319, 931–945. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, H.; Shi, Z.; Li, Y.; Zhang, X.; Gao, Z.; Zhou, L.; Ma, J.; Xu, Q.; Guan, J.; Cheng, Y.; Jiao, S.; Zhou, Z. The MST4-MOB4 complex disrupts the MST1-MOB1 complex in the Hippo-YAP pathway and plays a pro-oncogenic role in pancreatic cancer. J Biol Chem. 2018, 293, 14455–14469. [Google Scholar] [CrossRef]
- Pan, D. The hippo signaling pathway in development and cancer. Dev Cell. 2010, 19, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Kück, U.; Radchenko, D.; Teichert, I. STRIPAK, a highly conserved signaling complex, controls multiple eukaryotic cellular and developmental processes and is linked with human diseases. Biol Chem. 2019, 400, 1005–1022. [Google Scholar] [CrossRef] [PubMed]
- Chiu, I.M. Growth factor genes as oncogenes. Mol Chem Neuropathol. 1989, 10, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.S.; Miller, L.D. RAS Mutations and Oncogenesis: Not all RAS Mutations are Created Equally. Front Genet. 2012, 2, 100. [Google Scholar] [CrossRef] [PubMed]
- Morrish, F.; Neretti, N.; Sedivy, J.M.; Hockenbery, D.M. The oncogene c-Myc coordinates regulation of metabolic networks to enable rapid cell cycle entry. Cell Cycle. 2008, 7, 1054–1066. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, D.W. The retinoblastoma tumor-suppressor gene, the exception that proves the rule. Oncogene. 2006, 25, 5233–5243. [Google Scholar] [CrossRef]
- Smith, A.L.; Robin, T.P.; Ford, H.L. Molecular pathways: Targeting the TGF-β pathway for cancer therapy. Clin Cancer Res. 2012, 18, 4514–4521. [Google Scholar] [CrossRef]
- Savage, K.I.; Harkin, D.P. BRCA1, a ‘complex’ protein involved in the maintenance of genomic stability. FEBS J. 2015, 282, 630–646. [Google Scholar] [CrossRef]
- Mantovani, F.; Collavin, L.; Del Sal, G. Mutant p53 as a guardian of the cancer cell. Cell Death Differ. 2019, 26, 199–212. [Google Scholar] [CrossRef]
- Nishio, M.; Hamada, K.; Kawahara, K.; Sasaki, M.; Noguchi, F.; Chiba, S.; Mizuno, K.; Suzuki, S.O.; Dong, Y.; Tokuda, M.; Morikawa, T.; Hikasa, H.; Eggenschwiler, J.; Yabuta, N.; Nojima, H.; Nakagawa, K.; Hata, Y.; Nishina, H.; Mimori, K.; Mori, M.; Sasaki, T.; Mak, T.W.; Nakano, T.; Itami, S.; Suzuki, A. Cancer susceptibility and embryonic lethality in Mob1a/1b double-mutant mice. J Clin Invest. 2012, 122, 4505–4518. [Google Scholar] [CrossRef]
- St John, M.A.; Tao, W.; Fei, X.; Fukumoto, R.; Carcangiu, M.L.; Brownstein, D.G.; Parlow, A.F.; McGrath, J.; Xu, T. Mice deficient of Lats1 develop soft-tissue sarcomas, ovarian tumours and pituitary dysfunction. Nat Genet. 1999, 21, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Conrad, C.; Xia, F.; Park, J.S.; Payer, B.; Yin, Y.; Lauwers, G.Y.; Thasler, W.; Lee, J.T.; Avruch, J.; Bardeesy, N. Mst1 and Mst2 maintain hepatocyte quiescence and suppress hepatocellular carcinoma development through inactivation of the Yap1 oncogene. Cancer Cell. 2009, 16, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Lee, J.H.; Kim, T.S.; Kim, T.H.; Park, H.D.; Byun, J.S.; Kim, M.C.; Jeong, W.I.; Calvisi, D.F.; Kim, J.M.; Lim, D.S. The Hippo-Salvador pathway restrains hepatic oval cell proliferation, liver size, and liver tumorigenesis. Proc Natl Acad Sci U S A. 2010, 107, 8248–8253. [Google Scholar] [CrossRef]
- Jiang, K.; Yao, G.; Hu, L.; Yan, Y.; Liu, J.; Shi, J.; Chang, Y.; Zhang, Y.; Liang, D.; Shen, D.; Zhang, G.; Meng, S.; Piao, H. MOB2 suppresses GBM cell migration and invasion via regulation of FAK/Akt and cAMP/PKA signaling. Cell Death Dis. 2020, 11, 230. [Google Scholar] [CrossRef]
- Santos, I.B.; Wainman, A.; Garrido-Maraver, J.; Pires, V.; Riparbelli, M.G.; Kovács, L.; Callaini, G.; Glover, D.M.; Tavares, Á.A. Mob4 is essential for spermatogenesis in Drosophila melanogaster. Genetics. 2023. [Google Scholar] [CrossRef]
- Frost, A.; Elgort, M.G.; Brandman, O.; Ives, C.; Collins, S.R.; Miller-Vedam, L.; Weibezahn, J.; Hein, M.Y.; Poser, I.; Mann, M.; Hyman, A.A.; Weissman, J.S. Functional repurposing revealed by comparing S. pombe and S. cerevisiae genetic interactions. Cell. 2012, 149, 1339–1352. [Google Scholar] [CrossRef]
- Paulsen, R.D.; Soni, D.V.; Wollman, R.; Hahn, A.T.; Yee, M.C.; Guan, A.; Hesley, J.A.; Miller, S.C.; Cromwell, E.F.; Solow-Cordero, D.E.; Meyer, T.; Cimprich, K.A. A genome-wide siRNA screen reveals diverse cellular processes and pathways that mediate genome stability. Mol Cell. 2009, 35, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia. 2010, 24, 679–686. [Google Scholar] [CrossRef]
- Wong, Y.H.; Lee, T.Y.; Liang, H.K.; Huang, C.M.; Wang, T.Y.; Yang, Y.H.; Chu, C.H.; Huang, H.D.; Ko, M.T.; Hwang, J.K. KinasePhos 2.0, a web server for identifying protein kinase-specific phosphorylation sites based on sequences and coupling patterns. Nucleic Acids Res. 2007, 35, W588–W594. [Google Scholar] [CrossRef]
- Fukasawa, T.; Enomoto, A.; Miyagawa, K. Serine-Threonine Kinase 38 regulates CDC25A stability and the DNA damage-induced G2/M checkpoint. Cell Signal. 2015, 27, 1569–1575. [Google Scholar] [CrossRef]
- Lehtinen, M.K.; Yuan, Z.; Boag, P.R.; Yang, Y.; Villén, J.; Becker, E.B.; DiBacco, S.; de la Iglesia, N.; Gygi, S.; Blackwell, T.K.; Bonni, A. A conserved MST-FOXO signaling pathway mediates oxidative-stress responses and extends life span. Cell. 2006, 125, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Sanphui, P.; Biswas, S.C. FoxO3a is activated and executes neuron death via Bim in response to β-amyloid. Cell Death Dis. 2013, 4, e625. [Google Scholar] [CrossRef] [PubMed]
- Valis, K.; Prochazka, L.; Boura, E.; Chladova, J.; Obsil, T.; Rohlena, J.; Truksa, J.; Dong, L.F.; Ralph, S.J.; Neuzil, J. Hippo/Mst1 stimulates transcription of the proapoptotic mediator NOXA in a FoxO1-dependent manner. Cancer Res. 2011, 71, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, N.; Nagai, M.; Miyazaki, K.; Kurata, T.; Takehisa, Y.; Ikeda, Y.; Kamiya, T.; Okazawa, H.; Abe, K. Progressive decrease in the level of YAPdeltaCs, prosurvival isoforms of YAP, in the spinal cord of transgenic mouse carrying a mutant SOD1 gene. J Neurosci Res. 2009, 87, 928–936. [Google Scholar] [CrossRef]
- He, Y.; Emoto, K.; Fang, X.; Ren, N.; Tian, X.; Jan, Y.N.; Adler, P.N. Drosophila Mob family proteins interact with the related tricornered (Trc) and warts (Wts) kinases. Mol Biol Cell. 2005, 16, 4139–4152. [Google Scholar] [CrossRef]
- Emoto, K.; He, Y.; Ye, B.; Grueber, W.B.; Adler, P.N.; Jan, L.Y.; Jan, Y.N. Control of dendritic branching and tiling by the Tricornered-kinase/Furry signaling pathway in Drosophila sensory neurons. Cell. 2004, 119, 245–256. [Google Scholar] [CrossRef]
- Norkett, R.; Del Castillo, U.; Lu, W.; Gelfand, V.I. Ser/Thr kinase Trc controls neurite outgrowth in Drosophila by modulating microtubule-microtubule sliding. Elife. 2020, 9, e52009. [Google Scholar] [CrossRef]
- Wang, L.H.; Baker, N.E. Salvador-Warts-Hippo pathway regulates sensory organ development via caspase-dependent nonapoptotic signaling. Cell Death Dis. 2019, 10, 669. [Google Scholar] [CrossRef]
- Lin, C.H.; Hsieh, M.; Fan, S.S. The promotion of neurite formation in Neuro2A cells by mouse Mob2 protein. FEBS Lett. 2011, 585, 523–530. [Google Scholar] [CrossRef]
- Campbell, M.; Ganetzky, B. Identification of Mob2, a novel regulator of larval neuromuscular junction morphology, in natural populations of Drosophila melanogaster. Genetics. 2013, 195, 915–926. [Google Scholar] [CrossRef]
- O'Neill, A.C.; Kyrousi, C.; Einsiedler, M.; Burtscher, I.; Drukker, M.; Markie, D.M.; Kirk, E.P.; Götz, M.; Robertson, S.P.; Cappello, S. Mob2 Insufficiency Disrupts Neuronal Migration in the Developing Cortex. Front Cell Neurosci. 2018, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Hondius, D.C.; Eigenhuis, K.N.; Morrema, T.H.J.; van der Schors, R.C.; van Nierop, P.; Bugiani, M.; Li, K.W.; Hoozemans, J.J.M.; Smit, A.B.; Rozemuller, A.J.M. Proteomics analysis identifies new markers associated with capillary cerebral amyloid angiopathy in Alzheimer's disease. Acta Neuropathol Commun. 2018, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Satoh, J.; Kawana, N.; Yamamoto, Y. Pathway Analysis of ChIP-Seq-Based NRF1 Target Genes Suggests a Logical Hypothesis of their Involvement in the Pathogenesis of Neurodegenerative Diseases. Gene Regul Syst Bio. 2013, 7, 139–152. [Google Scholar] [CrossRef] [PubMed]
- McInnes, J. Insights on altered mitochondrial function and dynamics in the pathogenesis of neurodegeneration. Transl Neurodegener. 2013, 2, 12. [Google Scholar] [CrossRef]
- Oláh, J.; Vincze, O.; Virók, D.; Simon, D.; Bozsó, Z.; Tõkési, N.; Horváth, I.; Hlavanda, E.; Kovács, J.; Magyar, A.; Szũcs, M.; Orosz, F.; Penke, B.; Ovádi, J. Interactions of pathological hallmark proteins: Tubulin polymerization promoting protein/p25, beta-amyloid, and alpha-synuclein. J Biol Chem. 2011, 286, 34088–34100. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; ffrench-Constant, C. The control of cell number during central nervous system development in flies and mice. Mech Dev. 2003, 120, 1311–1325. [Google Scholar] [CrossRef]
Figure 1.
hMob4 gene structure and predicted encoded proteins. A) hMob4 genetic locus at 2q33.1. hMob4 gene is predicted to generate three different isoforms: the canonical isoform 1 contains 225 aa; Isoform 2 loses the first 32 aa in respect to the canonical isoform 1; Isoform 3 lacks an alternate in-frame exon (exon 2, light blue) generating a smaller isoform (aa 20-40 missing). B) Comparative alignment of the three Mob4 protein isoforms. Dark and light blue regions correspond to exon 1 and 2, respectively.
Figure 1.
hMob4 gene structure and predicted encoded proteins. A) hMob4 genetic locus at 2q33.1. hMob4 gene is predicted to generate three different isoforms: the canonical isoform 1 contains 225 aa; Isoform 2 loses the first 32 aa in respect to the canonical isoform 1; Isoform 3 lacks an alternate in-frame exon (exon 2, light blue) generating a smaller isoform (aa 20-40 missing). B) Comparative alignment of the three Mob4 protein isoforms. Dark and light blue regions correspond to exon 1 and 2, respectively.
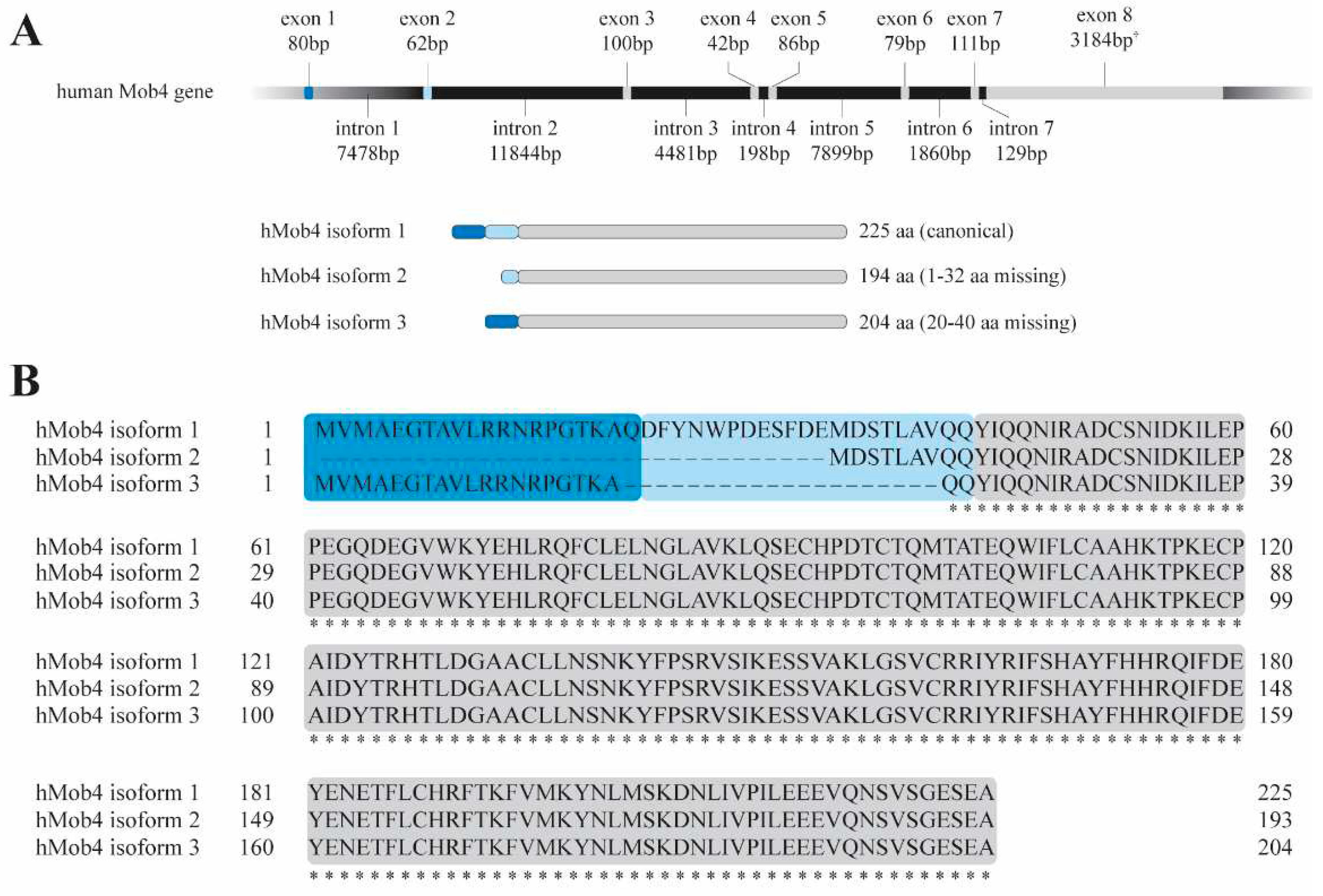
Figure 2.
Representative scheme of core mammalian STRIPAK complex. The STRIPAK complex is assembled on a tretramere of striatin (Striatin 3 in this figure), the phosphatase PP2A, a kinase (Mst3 or Mst4), and adaptor proteins interacting at different regions of the complex, like MOB4. Mob4 connects STRIP1 to STRN3 (adaptated and modified from [22]).
Figure 2.
Representative scheme of core mammalian STRIPAK complex. The STRIPAK complex is assembled on a tretramere of striatin (Striatin 3 in this figure), the phosphatase PP2A, a kinase (Mst3 or Mst4), and adaptor proteins interacting at different regions of the complex, like MOB4. Mob4 connects STRIP1 to STRN3 (adaptated and modified from [22]).
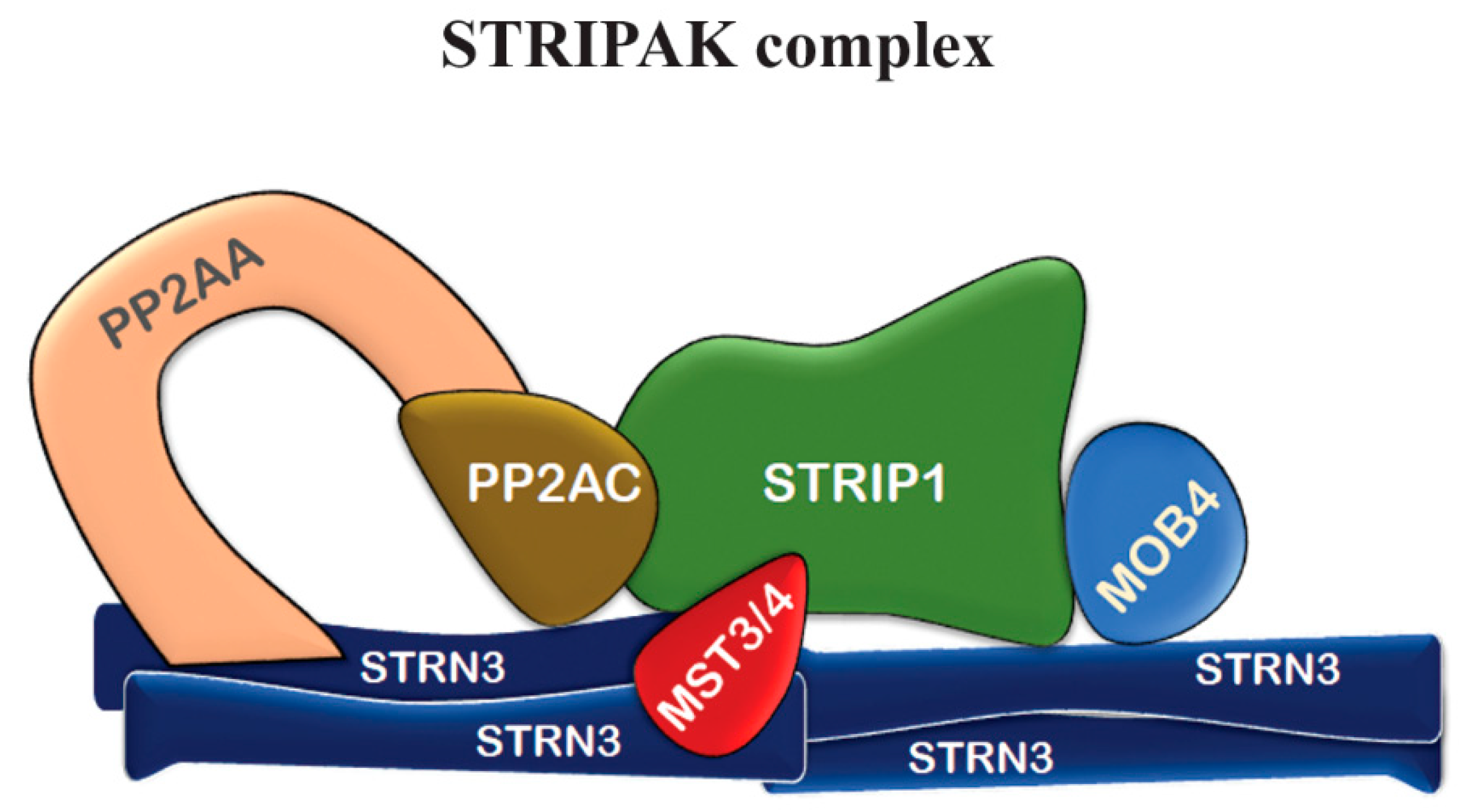
Figure 3.
Hippo signaling pathway regulation by MOB proteins. When active, the Hippo pathway blocks cell proliferation. The core of Hippo pathway is composed of a kinase cascade wherein MST1/2 together with SAV1, phosphorylates and activates the complex formed by LATS and MOB1 that, in turn, phosphorylates and inactivates the oncoprotein YAP/TAZ. Active YAP/TAZ (non-phosphorilated)migrates to the nucleus andpromotes the expression of cell proliferative and antiapoptotic genes. MOB family proteins interact with Hippo pathway at different levels regulating its activity. Mob2 negatively regulates Hippo pathway by competing with Mob1 for LATS1/2 binding. Mob3 appears to be a MST1 suppressor. Mob1, beside interacting with LATS1/2, can also form a complex with MST1 with tumor supressor functions. Mob4 forms a complex with MST4 that antagonises Mob1-MST1 functions. Mob4 also takes part of STRIPAK complex, a complex that acts upstream of Hippo signaling and therefore modulates MST1 activation. The core Hippo pathway is indicated by the traced square. See text for references.
Figure 3.
Hippo signaling pathway regulation by MOB proteins. When active, the Hippo pathway blocks cell proliferation. The core of Hippo pathway is composed of a kinase cascade wherein MST1/2 together with SAV1, phosphorylates and activates the complex formed by LATS and MOB1 that, in turn, phosphorylates and inactivates the oncoprotein YAP/TAZ. Active YAP/TAZ (non-phosphorilated)migrates to the nucleus andpromotes the expression of cell proliferative and antiapoptotic genes. MOB family proteins interact with Hippo pathway at different levels regulating its activity. Mob2 negatively regulates Hippo pathway by competing with Mob1 for LATS1/2 binding. Mob3 appears to be a MST1 suppressor. Mob1, beside interacting with LATS1/2, can also form a complex with MST1 with tumor supressor functions. Mob4 forms a complex with MST4 that antagonises Mob1-MST1 functions. Mob4 also takes part of STRIPAK complex, a complex that acts upstream of Hippo signaling and therefore modulates MST1 activation. The core Hippo pathway is indicated by the traced square. See text for references.
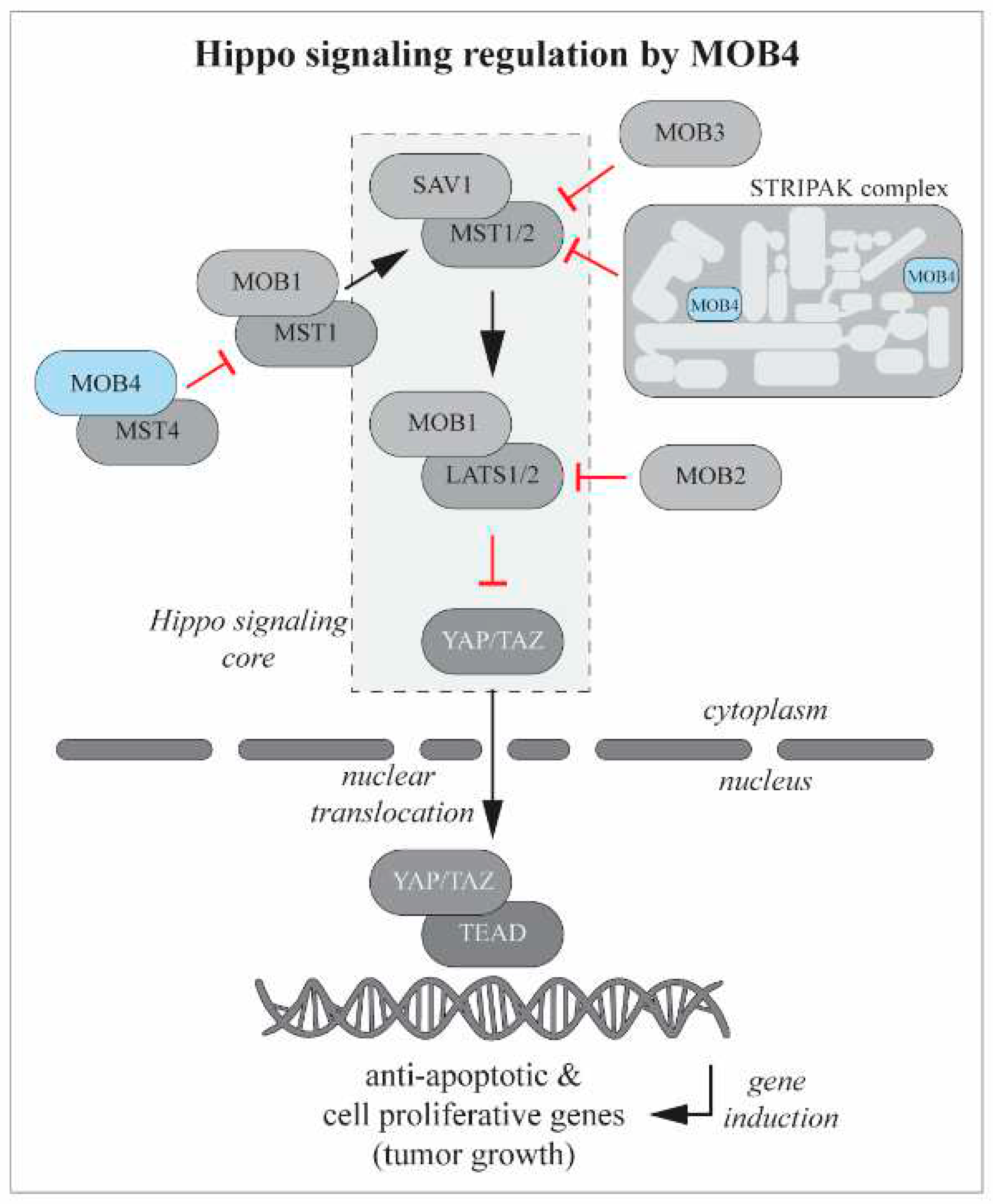
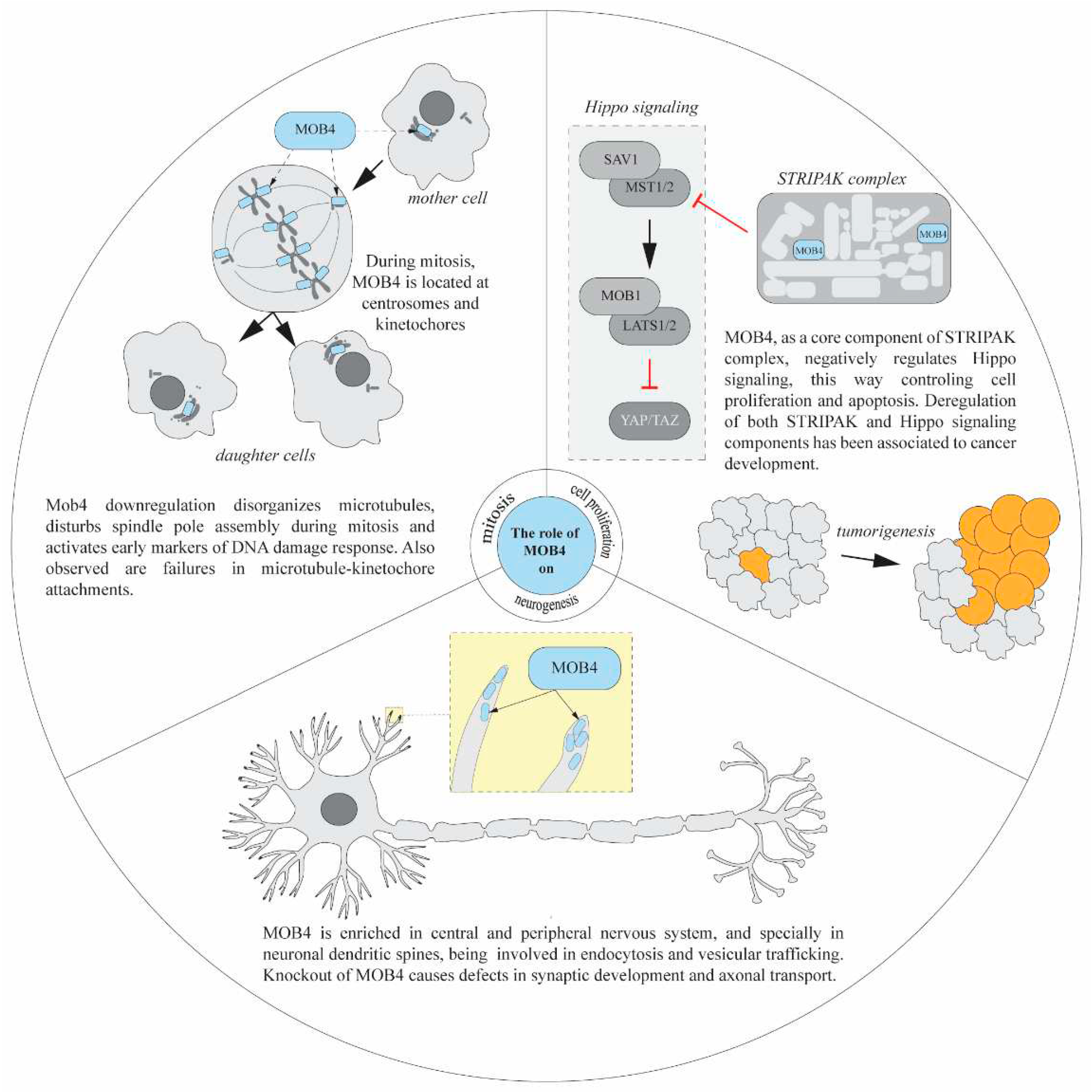
Figure 5.
An integrated view of some MOB4 functions (mitosis, cell proliferation and neurogenesis). MOB4 has been reported to participated in mitotic spindle assembly and cell division; in the control of cell proliferation regulating the Hippo signalling pathway as a member of STRIPAK complex; and in neural development regulating axonal transport, dendrite branching and controlling apoptosis in neurons. See main text for details.
Figure 5.
An integrated view of some MOB4 functions (mitosis, cell proliferation and neurogenesis). MOB4 has been reported to participated in mitotic spindle assembly and cell division; in the control of cell proliferation regulating the Hippo signalling pathway as a member of STRIPAK complex; and in neural development regulating axonal transport, dendrite branching and controlling apoptosis in neurons. See main text for details.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



