
Submitted:
21 August 2023
Posted:
22 August 2023
You are already at the latest version
Abstract
Since 2017, a fishing moratorium has been enforced in Lake Honghu as part of Chang-Jiang’s biodiversity conservation strategy. However, given that the lake is a semi-closed aquatic ecosystem, and no longer serves as a habitat for certain fishes of the mid-lower Chang-Jiang basin, the efficacy of this moratorium remains uncertain. To address the question from a trophic ecology perspective, a stomach content analysis was performed on captured anchovies C. brachygnathus, a current predominant species in Lake Honghu, from November 2020 to October 2021. Results showed that copepods, shrimps, and macrophytes were the main components of this anchovy's diet, and had size-related shifts in diets. The dominance of C. brachygnathus, a pelagic-littoral omnivore in this lake, implies a lacustrine environment shift triggered by continued human disturbances. The utilization of macrophytes as the main food items by large-sized anchovies of Lake Honghu partially contributed to the rapid degradation of submerged vegetation. This highlights the need to remove large-sized individuals of C. brachygnathus and Carassius auratus, another benthic-omnivorous dominant fish, in order to effectively restore the aquatic vegetation and ecosystem of Lake Honghu. The current implementation of fishing moratoriums in subtropical shallow floodplain lakes such as Lake Honghu should be reviewed critically.
Keywords:
Anchovy
; omnivore
; plant-origin food
; fisheries management
; fish ban
1. Introduction
Lake Honghu is the seventh-largest freshwater lake in China, located in the middle Yangtze River (or Chang-Jiang in Chinese) basin of southern Hubei Province [1]. As a water-carrying system, it performs a crucial function in flood regulation, providing water for industry and domestic use, agricultural irrigation, transportation, and fisheries. It also serves as an important breeding and wintering habitat for East Asian migrating birds, such as Mergus squamates and Ciconia boyciana [2,3]. Lake Honghu has been listed in the Ramsar wetlands of international importance, and it is a key biodiversity region and one of the priority conservation areas set up by the Chinese Government.
Nevertheless, Lake Honghu’s wetland has undergone severe human perturbations since the mid-1950s, disrupting its connectivity to the Chang-Jiang mainstem [2,3]. Prior to this, it served as nurseries, spawning, and feeding grounds to many potamodromous (e.g., silver, grass, bighead, and blunt carps) and diadromous (e.g., Japanese eel) fishes. However, due to the construction of sluice gates from 1955-1975, it has transformed into a river-isolated lake under artificial water level regulation, resulting in the decline of its aquatic environment and ecology. As a consequence, there has been a decrease in the species diversity of fish and fishery resources [1,4]. The number of fish species has drastically decreased from 73 in the 1950s-60s to 49 in the 1990s. Furthermore, fish assemblages have shown a trend toward early sexual maturation and smaller body sizes [5]. Fish farming on a large scale from the 1990s to 2010s has caused environmental and ecological problems, including reduced water quality and aquatic vegetation [6]. These issues have been worsened by exogenous wastewater discharge into the lake, causing a continuous degradation of water quality and aquatic habitat. The lake's altered environment has had significant impacts on its fish assemblages.
Under the current biodiversity conservation strategy in the Chang-Jiang basin, Lake Honghu is designated as a non-fishing zone at both the national and provincial levels. To restore its aquatic biodiversity and fish resources, strict regulations have been implemented by the national and local governments, including a fishing ban enforced since 2017. However, as a semi-closed or river-isolated aquatic ecosystem, Lake Honghu no longer serves as nursing, feeding, or breeding ground for many diadromous and potamodromous fish species. Therefore, the question of whether a fishing moratorium remains necessary in Lake Honghu remains debatable.
Coilia brachygnathus is a significant freshwater economic fish species in China, due to its substantial biomass, especially in affiliated lakes of the mid-lower Chang-Jiang [7,9]. The anchovy, previously regarded as the landlocked form of the anadromous species C. nasus, has been recently confirmed as a distinct species via molecular evidence, with its occurrence extending to Lakes Taihu, Poyang, and Dongting [10,11]. While the anchovy was reported to inhabit Lake Honghu, according to local fishermen, it was not the primary catch in the lake prior to 2015. Recent field surveys in Lake Honghu conducted between 2019 and 2021 revealed that C. brachygnathus, followed by benthic omnivorous Carassius auratus, were the most abundant fish species in the lake [12]. The anchovy's flourishing in Lake Honghu may be due to life history characteristics such as a short life span, early maturity, fast growth rate, availability of ample food, low predator pressure, and the fishing moratorium [13]. Fisheries management is therefore proposed for this fish. Understanding the trophic ecology of C. brachygnathus in Lake Honghu would aid in the rational management and utilization of fisheries resources.
Several studies have investigated the diet of C. brachygnathus in China. For instance, stomach content analyses of 52 individuals caught between March and June 1958 from the middle Chang-Jiang revealed that copepods and insect larvae, followed by small fish and shrimps, were the anchovy's primary food items [14]. Cladocerans and oligochaetes were only occasionally consumed. Similarly, stomach content analyses of 146 individuals collected from Lake Taihu between 1982 and 1984, 87 individuals fished from Meiliang Bay from September to October 2004, and 43 individuals landed from the entire lake from September to December 2004 demonstrated that the anchovy mainly fed on animal diets such as copepods, cladocerans, aquatic insects, shrimps, and small fish [15,16,17]. The same findings were observed for the fish in Lake Chaohu [18]. Nonetheless, these studies mainly used snapshot surveys, and the diet composition of the anchovy was shown to be influenced by habitat trophic status [19], with the fish displaying a temporal dietary shift linked to hydrological fluctuation levels [9].
Hence, the present study aims to (1) conduct a stomach content analysis of a large number of C. brachygnathus caught from Lake Honghu in different sampling seasons, (2) to provide insight into the optimal utilization and management of the lake’s fisheries resources, taking into account the dietary preferences of this omnivorous fish, and (3) address the issue on the necessity for fishing in this semi-closed aquatic environment.
2. Materials and Methods
2.1. Study Site
Lake Honghu, located in the southeast region of Hubei province (29o41’–29o58’N, 113o12’–113o28’E) and at an altitude of 22.75 m above sea level (Figure 1), is the largest shallow lake in the Jianghan Plain. It spans 23.4 km from east to west, 20.8 km from north to south, and has a mean water depth of 1.91m. The lake has a water surface area of 348.2 km2 and a watershed area of 8265 km2 [2,20]. The climate in Lake Honghu is subtropical monsoon, characterized by average annual precipitation ranging from 1000 to 1300 mm and an annual water temperature of 15.9 to 16.6oC [2].
2.2. Fish Sampling and Gut Content Analysis
From November 2020 to December 2021, samples of C. brachygnathus were collected from Lake Honghu seasonally. To collect these samples, eight different sites were used and 30- and 60-mm-stretched mesh-size monofilament gill nets and < 20 mm hoop nets were employed. Nets were set up between 14:00 and 16:00 h and were collected between 8:00 and 11:00 h the following morning. Individual fish were measured for total length to the nearest 0.1 mm, and the total weight was determined to the nearest 0.1 g using an electronic scale. Of the 800 fish sampled, their stomachs (guts) were removed, and the degree of gut fullness was recorded. Each fish was preserved individually in a buffered 4% formaldehyde solution and then transported for further diet analysis to the College of Fisheries laboratory at Huazhong Agricultural University.
At the laboratory, the entire stomach contents were emptied onto a Petri dish, and subsamples were taken to scrutinize under a microscope [21]. For large-sized food items, such as shrimp and macrophyte fragments, visual identification was performed. The inverted compound microscope (magnification: x400) was utilized to count and identify small food items up to the lowest possible taxonomic level [22,23]. Any prey items that were partially digested and consequently unidentifiable were reported as unidentified [24].
Diet composition was quantified as percentage frequency of occurrence (% O, the proportion of stomachs containing a particular prey item irrespective of amount) and percentage composition by number (%N; the proportion of the number of a particular prey item to the total number of all prey items in the entire stomach contents [25]. Given the difficult task to quantify macrophytes, the frequency of occurrence in each fish stomach was computed. The diet diversity was expressed by the Shannon-Wiener diversity index (H'). Furthermore, Pielou’s evenness index (J') was calculated to measure how evenly fish rely on food resources in different seasons.
Hierarchical cluster analysis based on the Euclidean distance and prey occurrence (%O) and abundance (%N) were used for petitioning size classes into different length groups. The matrix similarity was constructed using the Bray-Curtis distance after the abundance data were log (x+1) transformed. Canonical analysis of principal coordinates (CAP) [26] was used to distinguish variations in the diet compositions among sampling seasons and fish size classes. Besides, non-parametric permutational multivariate analysis of variance (PERMANOVA) was run to test whether the variations in diet composition among sampling periods and fish size classes were significant at 999 permutations [26]. All the statistical analyses were performed using OriginPro®2017 software (www.OriginLab.com), and PAST software.
3. Results
3.1. Food Items in Gut Contents
A total of 800 stomachs from anchovies measuring between 50 and 359 mm TL were dissected for diet analyses, which were seasonally caught from November 2020 to October 2021. Of the examined stomachs, 480 (60%) contained food, while the remaining 320 (40%) were empty. The study revealed 12 different food items, including macrophyte fragments (1 item), copepods (2 items), cladocerans (2 items), rotifers (5 items), shrimp (1 item), and the unrecognized (1 item). Table 1 presents the frequency of occurrence (%O) and abundance (%N) of these food items. Zooplankton groups were dominated by Copepods, followed by rotifers. dominated. The most frequently observed prey item in copepods remained consistent, with Calanus sp., making up 51.16% to 76.66% of observations. The most commonly observed prey item in cladocerans was Bosmina sp. The dominant prey item among rotifers was Brachionus calyciflorus, representing between 30.59% to 59.67% of observations throughout the sampling period. Macrophytes and shrimp were also among the most important food items throughout the entire duration of the study (Table 1). The study demonstrated a commonality of food items across all seasons, with Calanus sp., nauplii, B. calyciflorus, macrophytes, and shrimps appearing consistently as main food items.
3.2. Size-Related Dietary Variation
The cluster analysis conducted on collated anchovies, based on the occurrence of food items, showed size-related differences in the diet of the fish. As a result, three-sized fish classes were identified and outlined in Table 2 and Figure 3. The first group of individuals (<125 mm TL) subsisted exclusively on Calanus sp., Nauplii, Brachionus spp., Keratella sp., Tricocerca sp., Bosmina sp., and Alona sp. (Table 2). The second group (125-235 mm TL) fed mainly on Calanus sp., Nauplii, Keratella sp., Brachionus spp., Tricocerca sp., Bosmina sp., Alona sp., macrophytes, and shrimps. The third group (> 235 mm TL) foraged on larger food items such as big-sized Calanus sp., a few amounts of nauplii, macrophytes, and shrimps (Table 2).
In larger-sized anchovies, it was found that only a single food source, either macrophytes or shrimps, constituted the entirety of their diet (Table 3). Out of the more than 235 mm TL individuals of C. brachygnathus studied, only 20 were found to feed exclusively on macrophytes, another 3 individuals of more than 248 mm TL relied entirely on shrimps, and 14 individuals with sizes ranging from181-358 mm TL (with an average of 206 mm TL) consumed both macrophytes and shrimps. However, individuals with a smaller size (125 mm TL) were observed to have a higher rate of feeding on mixed diets consisting of macrophytes, zooplankton, and shrimps (Table 3).
The findings from this study demonstrate that there are significant differences in the food composition between the three length groups (P < 0.05), as confirmed by the PERMANOVA analysis (Table 4). It was also affirmed by the CAP ordination plot that size differences existed among the sampled individuals (Figure 4). As shown in Figure 5, the proportion of food groups varied with fish size. In the smaller fish class, copepods and rotifers were the predominant food groups. For the medium-sized fish class, copepods, rotifers, macrophytes, and shrimps were the top food groups. Finally, macrophytes and shrimp were the two most proportioned food groups for the large-sized fish classes.
4. Discussion
4.1. Dietary Variation of Anchovies in Lake Honghu
This study provides evidence supporting the omnivorous feeding habit and size-related dietary variation of C. brachygnathus in Lake Honghu. The size at which C. brachygnathus reached 50% maturity was 172 mm TL for females and 190 mm TL for males, which corresponded to one and 1.6 years, respectively [12]. Based on this, the stomach content analysis of our study, which examined a large number of anchovies (800) fished from Lake Honghu in four sampling seasons, proved that small-sized anchovies (less than 125 mm TL) were juveniles with a generalized feeding habit of zooplanktivory, while large-sized anchovies (more than 235 mm TL) were adults with a specialized feeding habit of omni-invertivory, with a greater reliance on aquatic invertebrates (shrimps) and food items of plant origin (macrophytes). Medium-sized anchovies adopted a mixed feeding strategy with varying degrees of specialization of different food items. The transition from zooplanktivory to omni-invertivory in their diet occurred in anchovies reaching 125 to 235 mm TL. This range is consistent with the previous report for C. brachygnathus of the middle Chang-Jiang mainstem [14]. The minimum transition length of 131 mm TL reported by Tang [15] for Lake Taihu’s anchovies was consistent with our study, but the maximum (170 mm TL) was much shorter than the 235 mm TL reported here. This variation could be explained by the smaller body size of the anchovies in Lake Taihu compared to Lake Honghu, where the anchovy has long been a dominant capture fish [15]. The trend toward body miniaturization in Lake Taihu’s anchovies is likely due to high fishing pressure [17].
The study also revealed that the size-related dietary shift of C. brachygnathus in Lake Honghu was accompanied by a shift in spatial habitat utilization from the pelagic to the littoral zone. While small-sized fish preferred feeding on zooplankton in the pelagic zone of the lake, large-sized fish showed a preference for shrimps and/or macrophytes, indicating a preference for the well-vegetated littoral zone (Tables 1-2). The density and distribution of shrimps in Lake Bao’an, located in the middle Chang-Jiang basin of Hubei Province, was known to depend on water depth and aquatic vegetation [28]. The shrimps were found to routinely use macrophytes as a refuge under pressure from predators such as Culter spp. and fed on attached algae. It is, therefore, likely that large-sized C. brachygnathus preferentially used the well-vegetated littoral zone of Lake Honghu. Unlike in previous studies [17,19,29], zoobenthos and demersal small fish were not observed in the stomachs of anchovies from Lake Honghu (Tables 1-2). This clearly indicates that anchovies of the lake were not benthic feeders and should be referred to as pelagic or littoral omnivores instead.
In this study, no seasonal dietary difference was observed for C. brachygnathus of Lake Honghu (Table 4). This is in contrast to the findings of Zhang et al.’s [9] from Lake Poyang, where stable isotope analysis indicated a seasonal variation in the diet of this anchovy. The difference can be attributed to the fact that, unlike Lake Poyang, Lake Honghu is a semi-closed aquatic ecosystem where the natural sources of food are not regulated by the season. Besides, being a shallow, eutrophic floodplain lake with high productivity, Lake Honghu provides abundant food sources throughout the year for C. brachygnathus.
4.2. Plant-Origin Food Items
Food items of plant origin, like aquatic macrophytes, have never been reported for C. brachygnathus of Lakes Chaohu [18], Lake Dongting [9], and Taihu [19], and the congeneric species C. nasus of the Qingcaosha reservoir located near the estuary of the Chang-Jiang [6]. However, there have been some reports of phytoplankton and algae being found in the stomachs of anchovies. Diatoms and spirogyra have been sporadically observed in the diet of C. brachygnathus from the middle Chang-Jiang basin [14], while algae and a significant amount of Microcystis spp. were found in the diet of C. ectenes taihuensis (currently rendered synonymic to C. brachygnathus) from Lake Taihu [15] [17]. Variations in feeding habits within the same species may be due to variations in prey availability as a response to the existence of differing hydrographic conditions among surveyed areas [30]. In Lake Honghu, the presence of macrophytes in the guts of medium- to large-sized individuals of C. brachygnathus may be attributed to a restricting of feeding habits in response to a modified environment and available resources [30]. This shift in feeding strategy may also be driven by energetic demands, as larger-sized individuals need more energy to survive [31].
It cannot be ruled out that C. brachygnathus accidentally ingests macrophytes when chasing after shrimps that are attached to them. However, the majority of anchovies found with macrophytes in their guts had mixed diets consisting of macrophytes, zooplankton, and shrimps. The number of anchovies feeding only on shrimps or a combination of macrophytes and shrimps was lower than the number of anchovies that solely fed on macrophytes (Table 3). This implies that C. brachygnathus intentionally prefers macrophytes as a part of its diet instead of incorporating them accidentally or at random.
4.3. No Need for Fishing Ban
Omnivores are more adaptable to varying conditions than specialists [32], they are expected to be prevalent in ecosystems affected by anthropogenic and natural factors [33]. Lake Honghu exemplifies this trend, as it has undergone significant perturbations since the 1950s, resulting in the disappearance of aquatic vegetation and the deterioration of water quality. The construction of some sluice gates in its outlets during the 1950s-1980s resulted in the hydrological disconnection from the middle Chang-Jiang mainstem, which had great influences on its fish assemblage. Following this disconnection, Carassius auratus, a benthic omnivore, has become one of the three dominant capture fish of Lake Honghu. So far, C. brachygnathus and Carassius auratus, two omnivorous fish species, have become the dominant capture fish of the lake [34]. The rise of C. brachygnathus reflects further anthropogenic disturbances [12], indicating intensified fish stocking, water pollution, and over-exploitation of aquatic vegetation during the past two decades or more [6]. As a result, Lake Honghu has already shifted from a macrophyte-dominate to an algae-dominated eutrophic lake.
Omnivorous fishes can accelerate the eutrophication process in shallow lakes and their biomass tends to increase with intensified natural and anthropogenic disturbances, such as fish stocking, water pollution, and global warming [35]. There was a positive relation between the capture of C. brachygnathus and the concentration of TN, TP, and DOC in Taihu Lake [17]. The removal of omnivorous fish from shallow lakes could be beneficial for improving water clarity and promoting benthic primary production [2]. In this context, the harvest of economically important omnivorous fish, such as C. brachygnathus and C. auratus, in Lake Honghu could be a viable restoration approach for this shallow eutrophic lake.
As shown here, C. brachygnathus of Lake Honghu can utilize macrophytes as its food items. Whether the anchovy assimilates this food item of plant origin or not, the persistence of medium- or large-sized individuals has adverse impacts on submerged macrophytes and affects the aquatic vegetation’s rehabilitation and habitat promotion of the lake. The current dominance of this anchovy in the lake is partially attributed to the fishing ban implemented in 2017, which has led to a decrease in competition among fish species. This ban may have unintentionally created a breeding ground for C. brachygnathus, resulting in the degradation of the aquatic ecosystem [12]. Therefore, it is crucial to reevaluate the fishing ban and implement a rational approach to managing the resources of this semi-closed aquatic ecosystem. A sustainable management plan for the dominant omnivorous fish is urgently needed to promote the rehabilitation and sustainable development of Lake Honghu.
Predation by planktivorous fish on zooplankton has an adverse impact on the abundance, size, and species composition of the zooplankton community [36,37]. This, in turn, indirectly affects the phytoplankton community and physico-chemical conditions, leading to increases in phytoplankton abundance [38]. Due to weak “top-down” control exerted by zooplankton on phytoplankton communities, this can result in the flourishing of phytoplankton and the deterioration of water quality. When zooplankton abundance is low, phytoplankton can thrive, and water quality can suffer [39,40]. Fishing of big-sized anchovy in Lake Honghu is predicted to increase high zooplankton abundance, thereby providing a selective constraint on phytoplankton, ultimately facilitating the improvement of water quality and the promotion of aquatic habitats.
Author Contributions
A.Y.M., E.Z., and L.C. designed the study, A.Y.M. and L.C. collected the survey data, E.Z.and J-Z.S. equipment, logistic support, and edited the manuscript, A.Y.M. analysed the data and elaborated the statistical analysis and figures, A.Y.M. and E.Z. wrote and modified the initial draft of the manuscript with the contribution of all the authors.
Institutional Review Board Statement
Fishes used for this study were collected in accordance with the Chinese Laboratory Animal Welfare Laws (GB/T 35892–2018).
Data Availability Statement
Data will be made available on request.
Acknowledgments
We thank the Lake Honghu wetland protection and restoration demonstration office for their cooperation during our sampling period. We would like to thank all the Huazhong Agricultural University, College of Fisheries laboratory students for their invaluable support during the field data collection and laboratory analysis. We are also grateful to the Institute of Hydrobiology (IHB) fish taxonomy and evolution research group for their immense support during data collection and laboratory analysis. This study was funded by the scientific investigation program of Lake Honghu.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Han M, Dsouza M, Zhou C, Li H, Zhang J, Chen C, et al. Agricultural Risk Factors Influence Microbial Ecology in Honghu Lake. Genomics Proteomics Bioinformatics, 2019; 17 (1): 76–90. [CrossRef]
- Zhang T, Ban X, Wang X, Cai X, Li E, Wang Z, et al. Analysis of nutrient transport and ecological response in Honghu Lake, China by using a mathematical model. Science of the Total Environment, 2017: 418–428. [CrossRef]
- Zhu M, Wang X, Ning L. Ecological impacts of water conservancy activity on fish resources in Honghu Lake. Journal of water resources and water engineering, 2008; 19 (1):2007–18.
- Zhao S, Fang J, Ji W, Tang Z. Lake restoration from impoldering: Impact of land conversion on the riparian landscape in Honghu Lake area, Central Yangtze. Agriculture, Ecosystem and Environment, 2003; 95:111–8. [CrossRef]
- Chen Y, Xu Y, Yang H, Ni X. Hydrobiology and Resources Exploitation in Lake Honghu. Science Press, Beijing,1995.
- Li L, Rong D, Qigen L, Shubo F, Liping C, Shihui S, et al. The ontogenetic dietary shift of Japanese grenadier anchovy (Coilia nasus) in the Qingcaosha reservoir near the Yangtze River estuary. Journal of Fishery Sciences of China, 2019; 26:765–73.
- Qin X, Wang T, Lin P, Wang X, Liu H. Age, growth, mortality and movement patterns of short jaw tapertail anchovy, Coilia brachygnathus, in the channel connecting Dongting Lake and the Yangtze River in central China. Journal of Aquatic living resources, 2018; 31 (3):2-9. [CrossRef]
- Xue D, Yang Q, Li Y, Zong S, Gao T. Comprehensive assessment of population genetic structure of the overexploited Japanese grenadier anchovy (Coilia nasus): Implications for fisheries management and conservation. Fisheries Research, 2019: 113–120. [CrossRef]
- Zhang H, Wu G, Xie P, Xu J, Zhou Q. Role of body size and temporal hydrology in the dietary shifts of short jaw tapertail anchovy. Hydrobiologia, 2013; 703:247–56.
- Cheng F, Wang Q, Maisano Delser P, Li C. Multiple freshwater invasions of the tapertail anchovy (Clupeiformes: Engraulidae) of the Yangtze River. Ecology and Evolution, 2019;9:1–4. [CrossRef]
- Xuan Z, Jiang T, Liu H, Chen X, Yang J. Mitochondrial DNA and microsatellite analyses reveal strong genetic differentiation between two types of estuarine tapertail anchovies (Coilia) in Yangtze River Basin, China. Hydrobiologia, 2021;848, 1409–1431. [CrossRef]
- Yimer M A, Cao L, Shen J, Zhang E. Age, growth, maturity and mortality of the tapetail anchovy Coilia brachygnathus (Engraulidae) in Lake Honghu, China. Journal of Fish Biology, 2023;2 (102):1–12.
- Zhang E. Fishes. in Li EH, Li, FY, Zhu JH (eds). Comprehensive scientific report on the national natural reserve of Lake Honghu, Hubei Province, 2023.
- Anonymous. Fishes of the Yangtze River. Science Press, Beijing, 1976.
- Tang Y. On the population dynamics of lake anchovy in Taihu Lake and its rational exploitation. Journal of fisheries of China, 1987;11:62–72.
- Ye J, Liu Z, Wang W. Comparative study on the feeding habits of Coilia ectenes and Hyporhamphus intermidius in the Meilliang Bay of Lake Taihu. Journal of Lake Sciences, 2007; 19:218–22.
- Liu E. A study on diet composition of dominant fishes in Lake Taihu. Journal of Fisheries of China, 2008; 32:396–400.
- Diao Z, Wu X. Dynamic of lake anchovy resource and suggestion of fishery utilization. Research Report on the Enhancement of Fisheries Resources in Lake Chaohu. 1982; 1: 62–73.
- Sha YC, Su GH, Zhang PY, Zhang H, Xu J. Diverse dietary strategy of lake anchovy Coilia ectenes taihuensis in lakes with different trophic status. Journal of Ichthyology, 2015; 55:866–73. [CrossRef]
- Yao S, Xue B, Xia W, Zhu Y, Li S. Lead pollution recorded in sediments of three lakes located at the middle and lower Yangtze River basin, China. Quaternary International journal, 2009; 208:145–50. [CrossRef]
- Costalago D, Palomera I, Tirelli V. Seasonal comparison of the diets of juvenile European anchovy Engraulis encrasicolus and sardine Sardina pilchardus in the Gulf of Lions. Journal of Sea Research, 2014; 89:64–72. [CrossRef]
- Botes L. Phytoplankton Identification Catalogue Saldanha Bay, South Africa. Marine and Coastal Management, Rogge Bay, 2001.
- MRC. Identification Handbook of Freshwater Zooplankton of the Mekong River and its Tributaries Identification Handbook of Freshwater Zooplankton of the Mekong River and its Tributaries. Technical Paper, 2015.
- Bacha M, Amara R. Spatial, temporal and ontogenetic variation in diet of anchovy (Engraulis encrasicolus) on the Algerian coast (SW Mediterranean). Estuary, Coast Shelf Science, 2009; 85:257–64. [CrossRef]
- Hyslop EJ. Stomach contents analysis—a review of methods and their application. Journal of Fish Biology, 1980; 17:411–29.
- Anderson MJ, Gorley RN, Clarke KR. PERMANOVA+ Primer V7: User Manual. Primer-E Ltd., Plymouth, UK, 2008; 93.
- Xu J, Zhang M, Xie P. Size-related shifts in reliance on benthic and pelagic food webs by lake anchovy. Ecoscience, 2007; 14:170–7. [CrossRef]
- Qin H M, Zhang T, Li Z, Hong Y J. Species composition, spatial distribution, and biomass of shrimp community in the Biandangtang Lake. Acta Hydrobiologica Sinica, 2005; 29: 379-384.
- Li Y, Bordinhon AM, Allen D, Zhang W, Zhu X. Protein: Energy ratio in practical diets for Nile tilapia Oreochromis niloticus. Aquaculture International, 2013; 21:1109–19. [CrossRef]
- Mustać B, Hure M. The diet of the anchovy Engraulis encrasicolus (Linnaeus, 1758) during the spawning season in the Eastern Adriatic Sea. Acta Adriatica, 2020; 61:57–66. [CrossRef]
- Zorica B, Čikeš KV, Vidjak O, Mladineo I, Ezgeta BD. Feeding habits and helminth parasites of sardine (S. pilchardus) and anchovy (E. encrasicolus) in the Adriatic Sea. Mediterranean Marnie Science, 2016; 17:216–29.
- Fagan WF. Omnivory as a stabilizing feature of natural communities. American Naturalist, 1997; 150:554–67. [CrossRef]
- Wootton KL. Omnivory and stability in freshwater habitats: Does theory match reality? Freshwater Biology, 2017; 62:821–32.
- He H, Hu E, Yu J, Luo X, Li K, Jeppesen E, et al. Does turbidity induce by Carassius carassius limit phytoplankton growth? A mesocosm study. Environmental Science and Pollution Research, 2017; 24:5012–8. [CrossRef]
- Mei X, Vladimir R, Lars GR, Erik J, Tang Y, Zhang X, et al. Effects of omnivorous fish on benthic-pelagic habitats coupling in shallow aquatic ecosystems: A minireview. Journal of Lake Sciences, 2021; 33:667–74.
- Brooks JL, Dodson SI. Predation, body size, and composition of zooplankton. Science, 1965; 150:28–35.
- Jeppesen E, Jensen JP, Sondergaard M, Lauridsen T, Landkildehus F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshwater Biology, 2000; 45:201–18. [CrossRef]
- Jeppesen E, Søndergaard M, Lauridsen TL, Davidson TA, Liu Z, Mazzeo N, et al. Biomanipulation as a Restoration Tool to Combat Eutrophication. Recent Advances and Future Challenges. Advances in Ecological Research, 2012;47:411–88.
- Liu Z, Hu J, Zhong P, Zhang X, Ning J, Larsen SE, et al. Successful restoration of a tropical shallow eutrophic lake: Strong bottom-up but weak top-down effects recorded. Water Research, 2018; 146:88–97. [CrossRef]
- Yu J, Xia M, Kong M, He H, Guan B, Liu Z, et al. A small omnivorous bitterling fish (Acheilognathus macropterus) facilitates dominance of cyanobacteria, rotifers, and Limnodrilus in an outdoor mesocosm experiment. Environmental Science and Pollution Research, 2020; 27:23862–70. [CrossRef]
Figure 1.
Location of Lake Honghu with the spatial distribution of the eight sampling sites.

Figure 2.
Major food items identified in the stomach of C. brachygnathus from Lake Honghu.
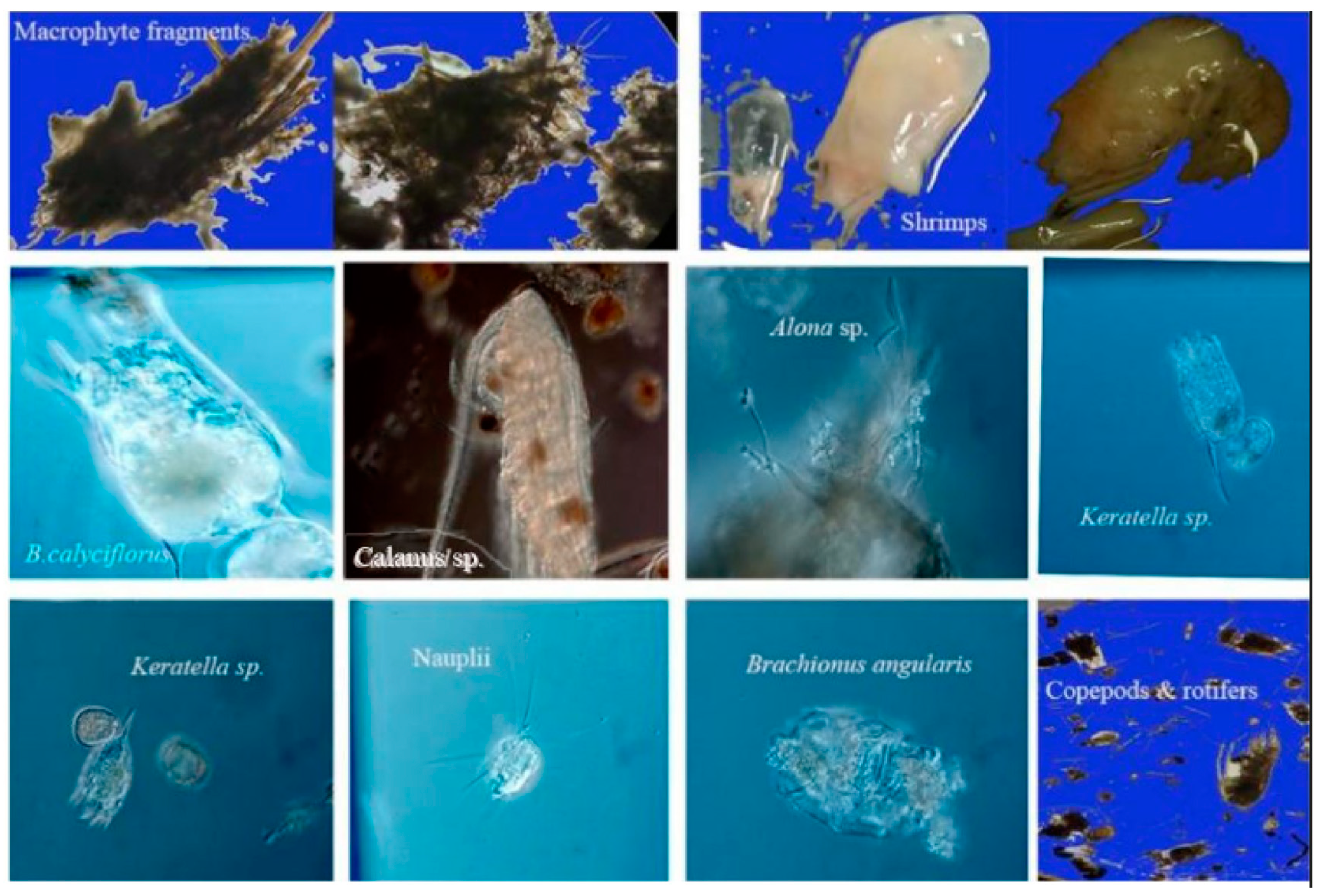
Figure 3.
Cluster analysis of fish size groups based on prey occurrence based on the Euclidean distance.
Figure 3.
Cluster analysis of fish size groups based on prey occurrence based on the Euclidean distance.
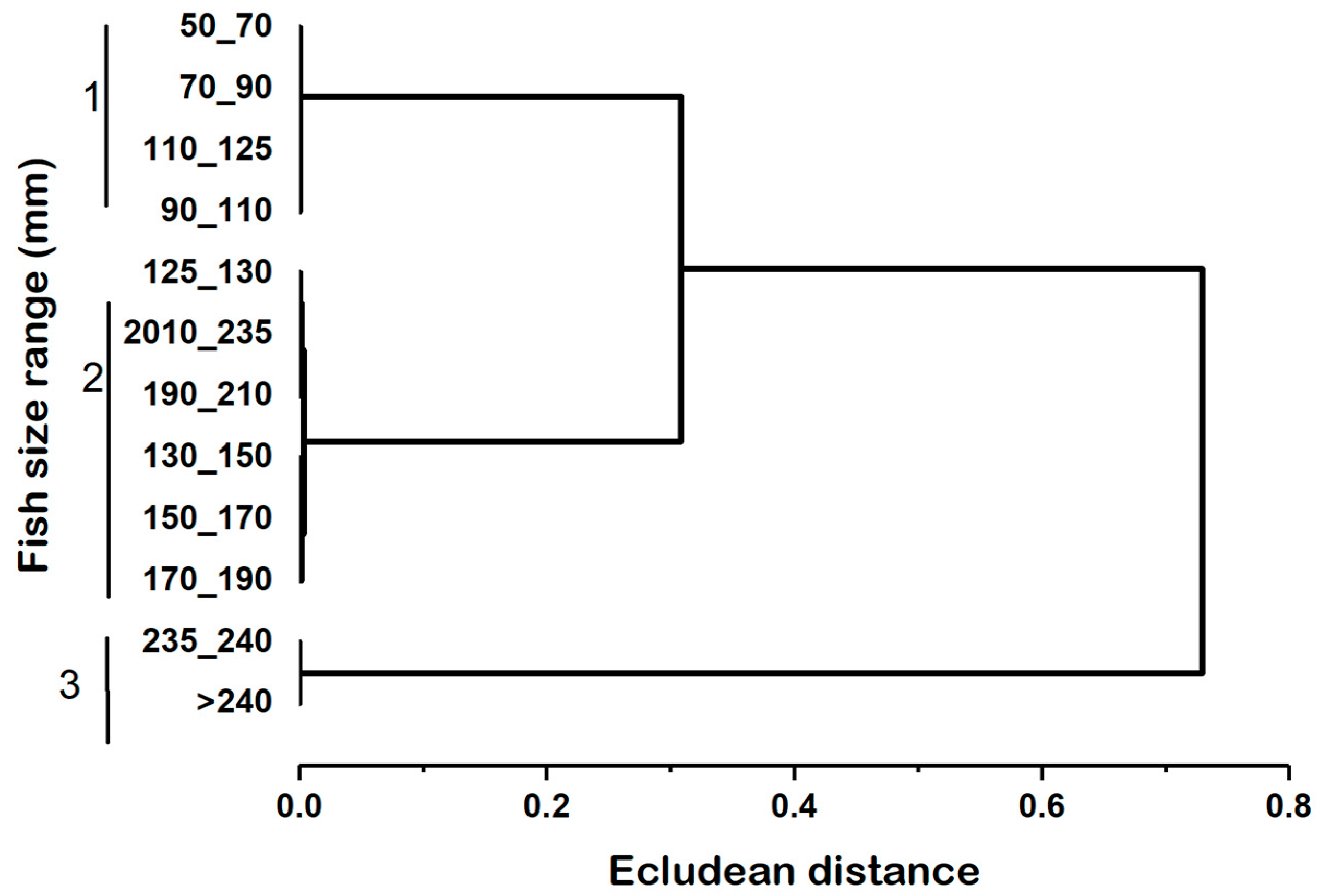
Figure 4.
Canonical analysis of principal coordinates (CAP) ordination plots based on the proportion of feed compositions of the three length classes of fish.
Figure 4.
Canonical analysis of principal coordinates (CAP) ordination plots based on the proportion of feed compositions of the three length classes of fish.

Figure 5.
The percentage contribution of different food items consumed by C. brachygnathus in different length classes in Lake Honghu.
Figure 5.
The percentage contribution of different food items consumed by C. brachygnathus in different length classes in Lake Honghu.
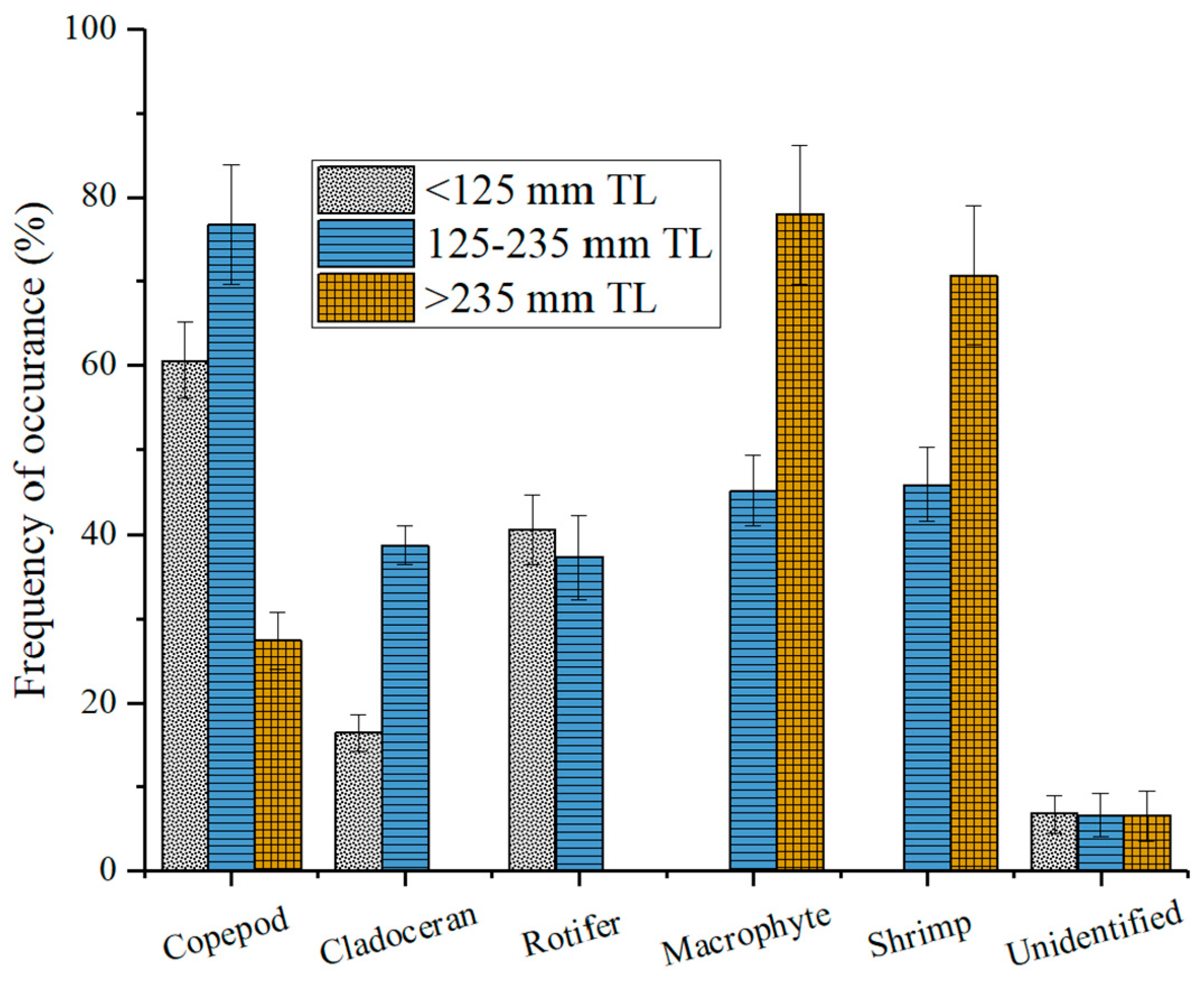

Table 1.
Frequency of occurrence (%O) and number (%N) of food items found in the stomach of C. brachygnathus between November 2020 and October 2021 in Lake Honghu.
Table 1.
Frequency of occurrence (%O) and number (%N) of food items found in the stomach of C. brachygnathus between November 2020 and October 2021 in Lake Honghu.
| November | January | June | October | |||||
|---|---|---|---|---|---|---|---|---|
| Food items | %O | %N | %O | %N | %O | %N | %O | %N |
| Calanus sp. | 75.20 | 23.33 | 54.17 | 23.02 | 51.16 | 20.55 | 76.66 | 23.61 |
| Nauplii | 66.43 | 19.22 | 37.75 | 19.16 | 46.99 | 21.84 | 67.23 | 23.22 |
| Bosmina sp. | 45.57 | 0.57 | - | - | - | - | - | - |
| Alona sp. | - | - | - | - | - | - | 35.42 | 1.64 |
| Brachionus calyciflorus | 50.98 | 15.83 | 30.59 | 21.66 | 59.67 | 28.64 | 43.98 | 10.07 |
| Brachionus angularis | 44.64 | 8.48 | 31.37 | 14.94 | - | - | 39.88 | 12.06 |
| Brachionus caudatus | 32.06 | 8.99 | - | - | - | - | 25.06 | 1.12 |
| Keratella sp. | 11.61 | 8.65 | - | - | - | - | - | - |
| Tricocerca sp. | 15.77 | 2.18 | - | - | - | - | 6.25 | 2.1 |
| Macrophytes | 60.76 | - | 70.34 | - | 58.46 | - | 65.28 | - |
| Shrimps | 55.21 | 12.39 | 51.84 | 21.11 | 45.96 | 24.13 | 68.94 | 25.98 |
| Unidentified | 10.23 | 0.36 | 9.85 | 0.11 | 8.96 | 4.84 | 8.53 | 0.2 |
| Number of preys | 11 | 7 | 6 | 10 | ||||
| Shannon-Wiener index | 2.25 | 1.85 | 1.7 | 2.09 | ||||
| Pielou’s evenness index | 0.94 | 0.95 | 0.95 | 0.91 | ||||
Table 2.
Frequency of occurrence (%O) and number (%N) of food items of the three length groups of C. brachygnathus in Lake Honghu.
Table 2.
Frequency of occurrence (%O) and number (%N) of food items of the three length groups of C. brachygnathus in Lake Honghu.
| Food items | Fish length groups | |||||
|---|---|---|---|---|---|---|
| < 125 mm TL | 125 – 235 mm TL | >235 mm TL | ||||
| O% | %N | O% | %N | %O | %N | |
| Calanus sp. | 57.35 | 27.02 | 76.10 | 30.55 | 58.33 | 35.69 |
| nauplii | 64.50 | 26.16 | 76.29 | 26.74 | 23.61 | 15.63 |
| Bosmina sp. | 23.11 | 0.13 | 47.06 | 0.04 | - | - |
| Alona sp. | 9.60 | 1.81 | 30.64 | 1.77 | - | - |
| Brachionus calyciflorus | 62.39 | 17.61 | 60.51 | 14.67 | - | - |
| Brachionus angularis | 52.66 | 12.8 | 63.24 | 10.45 | - | - |
| Brachionus caudatus | 22.38 | 6.1 | 36.03 | 5.06 | - | - |
| Keratella sp. | 9.52 | 5.35 | 6.07 | 2.03 | - | - |
| Tricocerca sp. | 9.52 | 2.11 | 20.34 | 1.9 | - | - |
| Macrophytes | - | - | 45.15 | - | 77.08 | - |
| Shrimps | - | - | 45.15 | 6.06 | 70.14 | 47.45 |
| Unidentified | 6.55 | 0.91 | 6.66 | 0.73 | 6.23 | 1.23 |
Table 3.
Food item contributions among different length groups of C. brachygnathus of Lake Honghu.
| Food items | Number | Percent (%) | Total length (mm) | ||
|---|---|---|---|---|---|
| Min. | Max. | Mean | |||
| Only shrimps | 3 | 0.38 | 248 | 308 | 283.2 |
| Only macrophytes | 20 | 2.50 | 235 | 306 | 278.75 |
| Macrophytes + shrimps | 14 | 1.75 | 181 | 359 | 247.71 |
| Macrophytes + zooplankton + shrimps | 121 | 15.13 | 125 | 309 | 206.92 |
| Macrophytes + zooplankton | 42 | 5.25 | 131 | 309 | 215 |
Table 4.
Results of permutational multivariate analysis of variance (PERMANOVA) applied to diet composition.
Table 4.
Results of permutational multivariate analysis of variance (PERMANOVA) applied to diet composition.
| Sources | Df | SS | MS | Pseudo-F | P(perm) |
|---|---|---|---|---|---|
| Fish size | 2 | 32593 | 16297 | 685.52 | 0.001 |
| Season | 3 | 1619.8 | 539.94 | 0.499 | 0.708 |
| Fish size * season | 6 | 1436.1 | 239.35 | 0.199 | 0.989 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



