
Submitted:
23 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
In order to enhance understanding of RNA-binding proteins in rice, a comprehensive investigation was conducted on the RRM1 gene family of rice, encompassing genome-wide identification and exploration of its role in rice blast resistance. Physical and chemical properties of the OsRRM1 gene family in rice was analyzed, including conserved domain, motif, location information, gene structure, phylogenetic tree, collinearity analysis, cis-acting elements, GO, and KEGG. Furthermore, the expression patterns of the OsRRM1 gene were examined at different time intervals following rice blast treatment. Furthermore, the alterations in expression patterns of selected OsRRM1 genes were assessed using quantitative real-time PCR(qRT-PCR). A total of 212 members of the OsRRM1 gene family were identified, which were dispersed across 12 chromosomes. Many of these genes exhibit multiple exons and introns, all of which encompass the conserved RRM1 domain and share analogous motifs. This observation suggests a high degree of conservation within the encoded sequence domain of these genes. Phylogenetic analysis revealed the existence of five subfamilies within the OsRRM1 gene family. Furthermore, the investigation of the promoter region identified homeopathic elements that are involved in nucleic acid binding and interaction with multiple transcription factors. By employing GO and KEGG analysis, four RRM1 genes were tentatively identified as crucial contributors to plant immunity, while the RRM1 gene family was also found to have a significant involvement in the complex of alternative splicing. Additionally, gene expression analysis indicated that the majority of OsRRM1 genes exhibited constitutive expression. The results of the qRT-PCR analysis revealed distinct temporal changes in the expression pattern of the OsRRM1 gene following rice blast treatment. These findings contribute to the existing knowledge of the OsRRM1 gene family, establish a foundation for further investigation into the role of the OsRRM1 gene in response to rice blast infection, and hold theoretical significance for future studies on the functionality of the OsRRM1 gene.
Keywords:
rice
; gene family
; rice blast
; bioinformatics.
1. Introduction
Gene expression must abide by strict laws, and each step needs to be strictly regulated, which is often regulated at the transcription level through DNA cis-acting elements and transcription factor binding[1,2]. Studies have shown that post-transcriptional regulation plays an important role in regulating gene expression of plant. Post-transcriptional regulation involves multiple processes, namely alternative splicing, RNA editing, RNA transport from the nucleus to the cytoplasm, RNA stabilization, and translation, which require the help of RNA binding proteins (RBPs) [1,3]. In order to achieve sequence-specific recognition of regulation in different levels and regulatory targets, there are several RNA binding domains with conserved characteristics in RBPs, such as RRM (RNA Recognition motif) domains[4,5].
The RNA recognition motif (RRM), also known as the RNA binding domain (RBD) or ribonucleoprotein domain (RNP), is one of the most abundant protein domains in eukaryotes and was first identified in the late 1980s[6,7,8,9]. The RNA recognition motif (RRM) domain is an important player in the regulation of development, signaling, gene expression, and cell differentiation[10,11,12,13]. RRM is a structurally conserved region consisting of about 80-90 amino acids, consisting of two short consensus sequences: RNP1 (hexapeptide) and RNP2 (octapeptide) [14]. It folds into a αβ sandwich with a typical β1α1β2β3α2β4 topology that forms a four-stranded antiparallel β-sheet packed against two α-helices[15]. The specificity of RNA binding is determined by multiple exposures to surrounding amino acids[14,16]. In some cases, a third helix is present during RNA binding[17]. The largest single-stranded RNA-binding proteome is the eukaryotic RNA recognition motif (RRM) family, which contains eight amino acid RRM1 consensus sequences[8,18]. RRM proteins have a variety of RNA-binding preferences and functions, including heteroribonucleo proteins (hnRNPs), proteins associated with alternative splicing regulation (SR, U2AF, Sxl), protein components of small ribonucleoproteins (U1 and U2 snRNPs), and proteins that regulate RNA stability and translation (PABP) [18,19,20]. The RRM in the heterodimer splicing factor U2 snRNP cofactor (U2AF) appears to have two RRM-like domains with special features for protein recognition[21]. This motif also appears in some single-stranded DNA-binding proteins[16].
Rice (Oryza sativa Japonica) is one of the main food crops in the world, which plays an irreplaceable role in China’s food security and is also an important model crop selected by biological research. However, there are few reports on OsRRM1 gene family. Previously unknown RRM1 transcription factors have been identified that interact directly with NLR to activate plant defense, establishing a direct link between transcriptional activation of immune responses and NRL-mediated pathogen perception[22]. Although the rice genome encodes a large number of OsRRM1 proteins, the exact number and function of these gene families in rice remains unclear.
Magnaporthe Oryza is one of the most widespread and harmful worldwide fungal diseases caused by rice blast fungus. It may infect rice at all stages of growth and development, seriously affecting the yield and quality of rice, and thus threatening the global food security. Although the traditional chemical control means can quickly and effectively control diseases and pests, long-term use of pesticide will not only bring severe environmental problems, but also increase economic costs, which is not conducive to the sustainable development of agriculture[23]. The resistance of germplasm resources has a wide range of genetic variation, and thus the host plant’s own resistance is the most effective, economical and environmentally friendly method to against Magnaporthe Oryza [24]. Many of studies have shown that the adaptability of rice blast fungus to the host changes frequently, and the resistance of rice varieties can only be maintained for 3 to 5 years[23,25,26]. Plant genomes express a large number of RRM-containing proteins, but only a few RRM proteins have been elucidated for their roles in plants, including immunity in plants, possibly through RNA processing[27,28,29,30]. Some researchers have identified possible members of the RRM transcription factor family, but have not predicted the role of all RRM genes in transcriptional activation in rice and other plants[31]. Therefore, it is necessary to further study the regulation of gene network during rice blast occurrence and to explore and identify new blast resistance genes, which has important theoretical and practical significance for the breeding of new varieties resistant to rice blast.
In this study, bioinformatics was used to identify and characterize the whole genome of RRM1 gene family in rice. The gene structure, physical and chemical properties, domain and phylogenetic characteristics of RRM1 gene family in rice were studied. In addition, RNA-seq was used to analyze the expression patterns of RRM1 gene family in different time periods after rice blast fungus treatment. At the same time, the expression changes of RRM1 family genes in response to stress resistance were analyzed by quantitative real-time PCR. This study increased the understanding of OsRRM1 gene family, and provided a basis for further investigation of the function of OsRRM1 gene under infection of rice blast fungus, and played a certain theoretical role for the subsequent study of the function of OsRRM1 gene family.
2. Materials and Methods
2.1. Identification and physicochemical properties of RRM1 gene family members in rice
Rice(Oryza sativa Japonica) genome sequence, annotation files, protein sequences, and gene structure file were downloaded from the Ensembl Plants database (http://plants.ensembl.org/index.html). Download the HMM (Hidden Marov Model) PF00076.24 (RRM1 domain) of the RRM1 gene family from the Pfam database (http://pfam.xfam.org/). Using HMM SEARCH tool sequence in HMMER3.2 software to search and analyze, RRM1 gene family in rice was predicted, and the E-value was less than 1×10-5. Domain analysis of identified RRM1 candidate sequences was performed using conserved RRM1 domain sequences in Pfam database (PF00076.24) and SMART online analysis software (http://smart.embl.de/). Using ExPASy (https://www.expasy.org/protparam/) online tools to predict protein isoelectric point, molecular size, length of protein sequences of amino acids. Use the Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/) online tools for protein subcellular localization prediction analysis.
2.2. Chromosome location of OsRRM1 gene family and construction of phylogenetic tree
The position of RRM1 gene on chromosome was analyzed through the rice gene sequence file downloaded from the Ensembl Plants database, and the chromosome location map was drawn by TBtools software. The NJ (neighbor-joining method) phylogenetic tree of RRM1 protein was constructed using MEGA11.0(Molecular Evolutionary genetics Analysis11.0) with the Bootstrap value of 1000 and use default for other parameters, and then the online software Itol(https://itol.embl.de/) was used to beautify the tree.
2.3. Analysis of conserved domain, gene structure and motif of OsRRM1 gene family
Conserved domains of the identified gene families were analyzed using the online tool Pfam (https://pfam.xfam.org/), and visualized by TBtools[32].
Through the plant genome database Ensembl plant rice gene structure annotation files downloaded (http://plants.ensembl.org/), the structure information of the members of the RRM1 gene family identified were analyzed using TBtools software for drawing genetic structure.
The conserved motif location of the identified RRM1 gene family was predicted using online MEME (https://meme-suite.org/meme/doc/meme). The parameter was set to 10 motifs and the other parameters were default. The prediction results were plotted using TBtools software.
2.4. Interspecies collinearity analysis of OsRRM1 gene family
The collinearity analysis and prediction of RRM1 gene in rice and Arabidopsis Thaliana were carried out, and the collinearity map was drawn by TBtools software.
2.5. GO and KEGG analysis of OsRRM1 gene family
GO and KEGG analysis of the OsRRM1 gene family was performed using PlantRegMap(http://plantregmap.gao-lab.org/) and Kobas(http://kobas.cbi.pku.edu.cn/kobas3/), respectively. All analysis results were calculated with q<0.05. Prism8.0 was used to plot the path name as the ordinate and -log10 (q-value) as the abscissa.
2.6. Analysis of presumptive cis-regulatory elements in the promoter region of OsRRM1 gene
Use TBtools software to predict cis acting elements in the 2000bp upstream gene promoter region of OsRRM1 in the PlantCARE Database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html).
2.7. Expression pattern analysis of RRM1 gene in rice treated with blast fungus
TBtools software was used to map the expression patterns of OsRRM1 gene family members identified under rice blast fungus treatment.
2.8. Plant materials and rice blast stress treatment
The experimental material is rice(O.Sativa L.spp.japonica,var nippobare). The mature seeds were placed in petri dishes, sterilized with 2%NaClO, soaked at 28 ° C for 48 hours, and then placed in perforated PCR plates. PCR plates were isolated by placing 24 seeds as three biological replicates. The treatment group and control group were repeated with two plates. All seedlings were placed in a growth chamber with a photoperiod of 14 h (day) /10 h (night) and a temperature cycle of 28/24 ℃.
For rice blast stress, rice seedlings were cultivated in artificial growth chamber until the three-leaf stage. Finally, suspensions of blast fungus with a concentration of 1*105 times were used as stress treatment. The rice blast strain was guy11. At 0, 12, 24, 36 and 48h after infecting Magnaporthe Oryza (guy11), the young rice leaves were immediately frozen in liquid nitrogen and stored at -80℃ for later use.
2.9. Analysis of OsRRM1 gene expression by qRT-PCR
RNA was extracted from treated plant leaves. Total RNA was extracted using the RNA Total RNA Extraction Kit (Takara). The first strand cDNA was synthesized using PrimeScript First strand cDNA Synthesis Kit (Takara) with a reaction volume of 20 μl, consisting of 1 μg total RNA, 4 μl 5xPrime Script RT premix and RNA-free enzyme ddH2O. The PCR procedure is as follows: 95℃ for 2 minutes, then 35 cycles, 95 ℃ for 5 seconds, 60℃ for 30 seconds.
Quantitative real-time PCR is performed on the ABI 7500 quantitative real-time fluorescent quantitative PCR system according to the manufacturer’s instructions. Primer Premier 5 was used to design specific primers targeting the OsRRM1 gene (Table 1). Actin is used as a reference gene. The qRT-PCR was performed in a final volume of 20 μl and consisted of 2 μl cDNA, 10 μl 2*SYBR green premix (Takara), and 1 μl forward and reverse primers. The amplification procedure is as follows: initial denaturation at 95 ℃ for 5 minutes; 35 cycles, denatured at 95 ℃ for 10 seconds, annealed at 60 ℃ for 20 seconds; And finally extended for 20 seconds at 72 ℃. Three biological replicates and three technical replicates were performed for each cDNA sample. Relative expression values are calculated by the 2 −ΔΔCT method.
3. Results
3.1. Screening and identification of RRM1 gene family members in rice
In this study, domains (Pfam: PF00076.24) predicted that 212 RRM1 genes (all with E values less than 1×10-5) were identified in the whole genome of rice (Oryza sativa Japonica), and their conserved domain was analyzed by Pfam (Figure 1). The results showed that all 212 OsRRM1 genes contained RRM1, but the location in the gene was different. These genes are named OsRRM1-1-OsRRM1-212 based on their physical location on the chromosome (Table 2). Use Expasy (https://web.expasy.org/protparam/) article analyzed 212 OsRRM1 gene molecular weight, length, isoelectric point, amino acids, et al. The results showed that the length of amino acids encoding the 212 rice RRM1 genes ranged from 53aa to 1160aa, the molecular weight ranged from 5837Da to 127816Da, and the theoretical isoelectric point distribution ranged from 3.97 to 12.37, which made the study of the OsRRM1 gene family more difficult. Subcellular localization prediction showed that OsRRM1 was mainly located in the nucleus, followed by the extracellular matrix, mitochondria, chloroplast, cell membrane, and intracytoplasmic matrix. This suggests that these proteins function were mainly in the nucleus. In addition, functional reports of several previously studied genes are listed.
3.2. Chromosome localization and phylogenetic tree analysis of OsRRM1 gene family
The positions of 212 OsRRM1 genes on chromosomes were mapped using TBtools software (Figure 2). There were 212 OsRRM1 genes distributed on all 12 chromosomes, among which 31 OsRRM1 genes were the most distributed on chromosome 3, and only 8 OsRRM1 genes were the least distributed on chromosome 10. Distinct gene clusters were formed on chromosomes 1, 2, and 3.
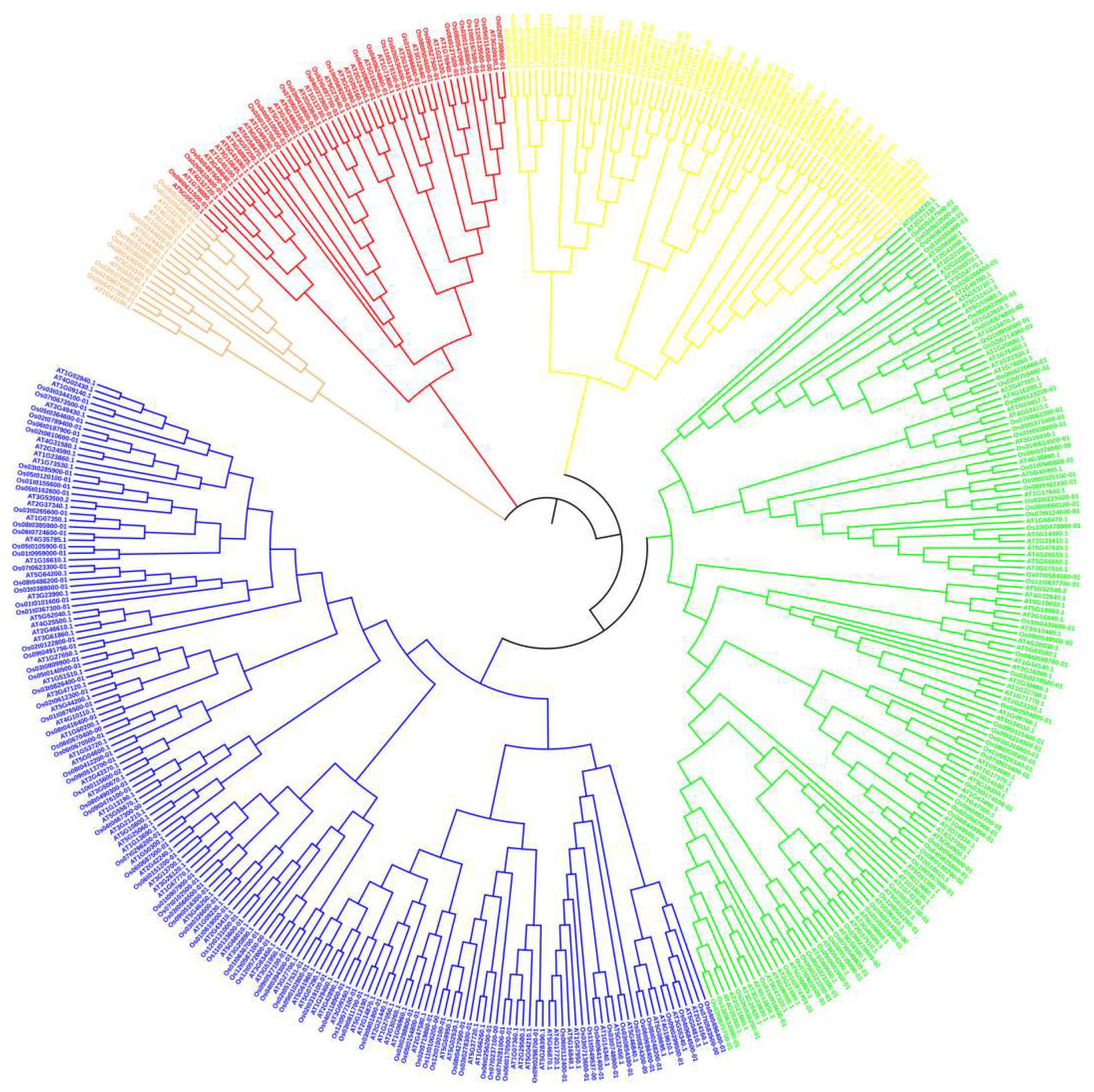
In order to study the phylogenetic relationship of OsRRM1 protein, a phylogenetic tree was constructed for 21 OsRRM1 protein sequences in rice (Figure 3). According to the topological structure of the evolutionary tree, 212 OsRRM1 proteins can be divided into 5 groups. The fifth group (Branch marks green) contained the highest amount of OsRRM1 protein and 61 proteins in total; The third group (Branch marks dark green) contained 58 RRM1 proteins, the second group (Branch marks red) and the fourth group (Branch marks blue) contained 33 RRM1 proteins and 53 RRM1 proteins respectively. The first group (Branch marks orange) had the lowest number of RRM1 proteins, seven in total.
3.3. Motif analysis and gene structure analysis of OsRRM1 gene family
The evolution of a family is mainly manifested by the diversity of gene structure and the change of conserved motifs. In order to better understand the structure of OsRRM1 gene, the exon intron structure of OsRRM1 gene was analyzed using annotated information from the rice reference genome (Figure 4). The results showed that 212 OsRRM1 genes had large differences in sequence length and exons and introns, but the same as the clustering results of evolutionary tree, genes in the same group usually had similar structure, but their intron lengths were different. It was also found that exon-intron patterns in the same phylogenetic taxa showed great similarity. This may be the result of replication of these sequences, which may also prove that the classification results are reliable.
Then, the online prediction tool MEME was used to identify the conserved motifs of OsRRM1 protein. Multiple motifs exist in 212 OsRRM1 protein sequences (Figure 4), and the types and numbers of motifs are highly overlapping. In addition, gene families within the same subfamily in the evolutionary tree are composed similarly on the motif.
3.4. Evolutionary analysis of OsRRM1 gene family and collinearity analysis of RRM1 gene family between rice and Arabidopsis Thaliana
Phylogenetic tree was constructed by comparing 212 OsRRM1 and 230 AtRRM1 sequences, with a total of 442 members (Figure A1). According to the topological structure of the evolutionary tree, RRM1 proteins of the two species can be divided into five groups. Most of the RRM1 protein members of rice and Arabidopsis do not cluster into their own clades. Each subfamily contains members of the RRM1 family of Arabidopsis and rice, and the members of each subfamily may have similar functions and domains. According to the phylogenetic relationship of protein sequences, the function of OsRRM1 protein can be predicted by the function of plant RRM1 protein with known function.
In order to further explore the evolutionary relationship of OsRRM1 gene family, collinearity analysis was conducted between rice and Arabidopsis Thaliana. The results showed (Figure 5) that 20 pairs of RRM1 genes in the two species were collinear, and no collinearity was found on chromosomes 8, 9, 10, 11 and 12 of rice and chromosome 4 of Arabidopsis thaliana.
3.5. GO and KEGG analysis of OsRRM1 gene family
GO annotation results showed (Figure 6) that the OsRRM1 gene family plays an important role in biological processes such as innate immune response, immune response, stimulus response, defense response of biological processes, regulatory transcription negative regulation, DNA template negative regulation of gene expression, epigenetic immune system processes, and alternative splicing. These results further confirm the reported functions of RRM1 gene in these aspects. KEGG analysis showed (Figure 6) that OsRRM1 gene family plays an important role in alternative splicing, messenger RNA surveillance pathway, RNA transport, and RNA degradation.
3.6. Characterization of presumptive cis-regulatory elements in the promoter region of OsRRM1 gene
The cis-regulatory elements in the promoter region play an important role in plant response to stress. Using the PlantCARE database, we identified five stress response cy-regulatory elements in the upstream 2000bp of these OsRRM1 genes, including TGACG motif (involved in JA response), CGTCA motif (involved in MeJA response), ABRE motif (involved in abscisic acid stress), TCA element (involved in salicylic acid reactivity), TGACG motif (involved in JA response), and CGTCA motif (involved in salicylic acid reactivity). WUN motif (wound response element). In the OsRRM1 gene family, the element associated with the largest number of stress response elements was ABRE (Figure 7), and ABA was synthesized mainly in response to blast stress. These results indicate that the OsRRM1 gene and stress-related response elements are relatively intact, but the types and amounts of stress-related elements contained in the promoter of each OsRRM1 gene are different, suggesting that members of the OsRRM1 gene family respond differently to rice blast stress.
3.7. Expression pattern of RRM1 gene family in rice after treatment with blast fungus
Using RNA-seq data, heat maps of 212 OsRRM1 genes represented by log2foldchange values were constructed at different time periods after infecting Magnaporthe Oryza(Figure 8). All OsRRM1 genes were expressed, and three major clusters of expression patterns were distinguished according to the expression specificity at different time periods after treatment. The RRM1 gene in two clusters showed an obvious up-regulation trend.
3.8. Expression analysis of OsRRM1 gene in response to biological stress
In order to further explore the expression changes of OsRRM1 gene in response to biological stress, qRT-PCR was performed on 4 OsRRM1 genes through the analysis of GO and KEGG results combined with the expression heat map to measure the transcription level of OsRRM1 gene. There were differences in the expression levels of four OsRRM1 genes under rice blast stress, all of which were up-regulated after treatment (Figure 9), indicating that four OsRRM1 genes played a certain role in the development of rice blast.
4. Discussion
Rice is one of the most important food crops and a monocotyledonous plant. In this study, bioinformatics was used to analyze the whole genome of rice OsRRM1 gene family. The length of amino acids encoded by 212 OsRRM1 genes ranges from 53aa to 1160aa, while Antoine Cler’s study indicated that the length of RRM is 90 amino acids, which is because there are a large number of introns in the RRM1 gene in rice, and these introns are largely discarded during transcription and translation. Therefore, the RRM1 protein encoded by the RRM1 gene actually has only about 90 amino acids (Figure 4)[54]. It is now clear that RRM1 is an important domain that needs to be further understood and that further biochemical and structural studies are needed to obtain a complete model of its role in cells[16]. The RRM1 gene family is distributed in many species, 230 of which have been identified in Arabidopsis and 212 in rice. One study investigated the complete Arabidopsis genome containing proteins containing RRM and KH RNA binding domains, and the Arabidopsis genome encodes 196 RRM proteins[55]. The phylogenetic tree analysis of RRM1 protein in rice and Arabidopsis showed that there were multiple pairs of RRM1 homologous genes in rice and Arabidopsis, suggesting that these genes have similar amino acid sequences in rice and Arabidopsis and may have similar functions. Since rice is a monocotyledonous plant and Arabidopsis is a dicotyledonous plant, it can be inferred that the time of RRM1 gene evolution may be earlier than the time of species differentiation. Subcellular localization prediction showed that OsRRM1 gene was mainly located in the nucleus, followed by the extracellular matrix, mitochondria, chloroplast, cell membrane, and intracytoplasmic matrix, indicating that the above proteins mainly function in the nucleus. According to subcellular localization prediction tools, 23 Arabidopsis RRM proteins were reported to be located in chloroplasts and 10 in mitochondria[56]. This result may be due to the fact that the main site of DNA replication is in the nucleus, with a small amount of DNA replication in mitochondria and chloroplasts. In chromosome localization, 212 RRM1 genes were found to be distributed on 12 chromosomes. In addition, there were multiple gene clusters on some chromosomes, which may be attributed to tandem duplication, resulting in gene amplification, which is of great significance in evolution. Among 212 OsRRM1 gene sequences, CDS and introns had different numbers and large spans. However, analysis of 10 amino acid conserved motifs of 212 OsRRM1 family proteins showed that the conserved sequences of OsRRM1 were mostly similar, especially in homologous sequences (Figure 4).
RRM1 gene was enriched by analysis of GO and KEGG, and this family gene was mainly enriched in biological processes related to stress resistance, such as rice blast immune pathway. As previous studies have shown, the RRM protein in plant organelles is involved in various RNA processes, regulating plant development (such as flowering) and plant stress response[57]. Moreover, this gene family is highly enriched in alternative splicing and mRNA assembly processes[58]. Studies have shown that both PSRP2 and ORRM5 have RNA-binding activity, and it is speculated that RRM proteins increase their RNA-binding energy as RNA chaperone under stress conditions[59,60,61]. They are also involved in plant development and stress responses, sometimes acting as proteins or RNA-binding proteins[62,63]. In addition, several RRM proteins have been reported to be involved in plant development and stress response[59,64,65,66]. It can be inferred that this gene family may be involved in immunity of rice by regulating downstream gene alternative splicing. The cis-regulatory elements in the promoter region play an important role in plant response to stress. We identified five stress response cis-regulatory elements (Figure 7) in the upstream 2000bp of these OsRRM1 genes, including TGACG motif (involved in JA response), CGTCA motif (involved in MeJA response), ABRE motif (involved in ababolic acid stress), TCA element (involved in salicylic acid reactivity), TGACG motif (involved in JA response), and TCA motif (involved in salicylic acid reactivity), WUN motif (wound response element). These results indicate that the stress-related response elements of OsRRM1 gene are relatively complete, suggesting that members of the OsRRM1 gene family regulate stress to a certain extent. In order to further explore the expression changes of OsRRM1 gene in response to biological stress, qRT-PCR was performed on four OsRRM1 gene candidates through the analysis of GO and KEGG results and combined with the expression heat map to measure the transcription level of OsRRM1 gene. There were differences in the expression levels of four OsRRM1 genes under rice blast stress, all of which were up-regulated after treatment (Figure 9), indicating that OsRRM1 played a certain role in the development of rice blast, which also verified the results of S. Wang and X. Shi[67].
This study offers an initial comprehension of the RRM1 gene family in rice, elucidating the potential roles of these genes in rice resistance, and establishing a basis for future investigations into the functions of individual members within this gene family. Subsequent steps will involve cloning, analysis of expression patterns, and functional verification of relevant genes in order to deepen our understanding and explore the significant contribution of RRM1 genes to the growth and development of rice.
Author Contributions
Conceptualization, Xinlei Jiang, Shaochun Liu and Junying Huang; methodology, Shangwei Yu; software, Shangwei Yu; validation, Xinlei Jiang and Yuhan Huang; formal analysis, Xinlei Jiang; investigation, Junru Fu; resources, Haihui Fu and Dewei Ynag; data curation, Haihui Fu; writing—original draft preparation, Xinlei Jiang; writing—review and editing, Haihui Fu; visualization, Xinlei Jiang; supervision, Haihui Fu; project administration, Haihui Fu; funding acquisition, Haihui Fu. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Double Thousand Plan of Jiangxi Province to Haihui Fu, grant number jxsq2019101057.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funder of this paper is Professor Fu, who supervised all the experiments in the paper and played a key role in review and editing.
Appendix A
Figure A1.
Phylogenetic tree of RRM1 gene families in Arabidopsis and rice.
References
- Latchman, D.S. Transcriptional Gene Regulation in Eukaryotes. In Els, 2011.
- Jeune, E.L.; Ladurner, A.G. Book Review. Protein Science 2004, 13, 1950–1952. [Google Scholar] [CrossRef]
- Jackson, D.A.; Pombo, A.; Iborra, F. The Balance Sheet for Transcription: An Analysis of Nuclear Rna Metabolism in Mammalian Cells. Faseb j 2000, 14, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Lorković, Z.J.; Barta, A. Genome Analysis: Rna Recognition Motif (Rrm) and K Homology (Kh) Domain Rna-Binding Proteins from the Flowering Plant Arabidopsis Thaliana. Nucleic Acids Res 2002, 30, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Burd, C.G.; Dreyfuss, G. Conserved Structures and Diversity of Functions of Rna-Binding Proteins. Science 1994, 265, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Dreyfuss, G.; Swanson, M.S.; Piñol-Roma, S. Heterogeneous Nuclear Ribonucleoprotein Particles and the Pathway of Mrna Formation. Trends in Biochemical Sciences 1988, 13, 86–91. [Google Scholar] [CrossRef]
- Adam, S.A.; Nakagawa, T.; Swanson, M.S.; Woodruff, T.K.; Dreyfuss, G. Mrna Polyadenylate-Binding Protein: Gene Isolation and Sequencing and Identification of a Ribonucleoprotein Consensus Sequence. Mol Cell Biol 1986, 6, 2932–2943. [Google Scholar] [PubMed]
- Bandziulis, R.J.; Swanson, M.S.; Dreyfuss, G. Rna-Binding Proteins as Developmental Regulators. Genes Dev 1989, 3, 431–437. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Kim, V.N.; Kataoka, N. Messenger-Rna-Binding Proteins and the Messages They Carry. Nat Rev Mol Cell Biol 2002, 3, 195–205. [Google Scholar] [CrossRef]
- Gomes, J.-E.; Encalada, S.E.; Swan, K.A.; Shelton, C.A.; Carter, J.C.; Bowerman, B. The Maternal Gene Spn-4 Encodes a Predicted Rrm Protein Required for Mitotic Spindle Orientation and Cell Fate Patterning in Early C. Elegans Embryos. Development 2021, 21, 4301–4314. [Google Scholar]
- Zhan, X.; Qian, B.; Cao, F.; Wu, W.; Yang, L.; Guan, Q.; Gu, X.; Wang, P.; Okusolubo, T.A.; Dunn, S.L.; Zhu, J.K.; Zhu, J. An Arabidopsis Pwi and Rrm Motif-Containing Protein Is Critical for Pre-Mrna Splicing and Aba Responses. Nat Commun 2015, 6, 8139. [Google Scholar] [CrossRef]
- Paukku, K.; Backlund, M.; De Boer, R.A.; Kalkkinen, N.; Kontula, K.K.; Lehtonen, J.Y. Regulation of At1r Expression through Hur by Insulin. Nucleic Acids Res 2012, 40, 5250–5261. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, M.K.; Clark, B.J.; McLaughlin, E.A.; D’Sylva, R.J.; O’Donnell, L.; Wilce, J.A.; Sutherland, J.; O’Connor, A.E.; Whittle, B.; Goodnow, C.C.; Ormandy, C.J.; Jamsai, D. Rbm5 Is a Male Germ Cell Splicing Factor and Is Required for Spermatid Differentiation and Male Fertility. PLoS Genet 2013, 9, e1003628. [Google Scholar] [CrossRef] [PubMed]
- Maris, C.; Dominguez, C.; Allain, F.H. The Rna Recognition Motif, a Plastic Rna-Binding Platform to Regulate Post-Transcriptional Gene Expression. Febs j 2005, 272, 2118–2131. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Oubridge, C.; Jessen, T.H.; Li, J.; Evans, P.R. Crystal Structure of the Rna-Binding Domain of the U1 Small Nuclear Ribonucleoprotein A. Nature 1990, 348, 515–520. [Google Scholar] [CrossRef]
- Cléry, A.; Blatter, M.; Allain, F.H. Rna Recognition Motifs: Boring? Not Quite. Curr Opin Struct Biol 2008, 18, 290–298. [Google Scholar] [CrossRef]
- Birney, E.; Kumar, S.; Krainer, A.R. Analysis of the Rna-Recognition Motif and Rs and Rgg Domains: Conservation in Metazoan Pre-Mrna Splicing Factors. Nucleic Acids Research 1993, 21, 5803–5816. [Google Scholar] [CrossRef]
- Query, C.C.; Bentley, R.C.; Keene, J.D. A Common Rna Recognition Motif Identified within a Defined U1 Rna Binding Domain of the 70k U1 Snrnp Protein. Cell 1989, 57, 89–101. [Google Scholar] [CrossRef]
- Chambers, J.C.; Kenan, D.; Martin, B.J.; Keene, J.D. Genomic Structure and Amino Acid Sequence Domains of the Human La Autoantigen. J Biol Chem 1988, 263, 18043–18051. [Google Scholar] [CrossRef]
- 20. Sachs, Davis and Kornberg. A Single Domain of Yeast Poly(a)-Binding Protein Is Necessary and Sufficient for Rna Binding and Cell Viability. Mol.cell.biol.
- Kielkopf, C.L.; Lücke, S.; Green, M.R. U2af Homology Motifs: Protein Recognition in the Rrm World. Genes Dev 2004, 18, 1513–1526. [Google Scholar] [CrossRef]
- Zhai, K.; Deng, Y.; Liang, D.; Tang, J.; Liu, J.; Yan, B.; Yin, X.; Lin, H.; Chen, F.; Yang, D. Rrm Transcription Factors Interact with Nlrs and Regulate Broad-Spectrum Blast Resistance in Rice. Mol Cell 2019. [CrossRef]
- Jeon, J.; Lee, G.W.; Kim, K.T.; Park, S.Y.; Kim, S.; Kwon, S.; Huh, A.; Chung, H.; Lee, D.Y.; Kim, C.Y.; Lee, Y.H. Transcriptome Profiling of the Rice Blast Fungus Magnaporthe Oryzae and Its Host Oryza Sativa During Infection. Mol Plant Microbe Interact 2020, 33, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, H.K.; Jørgensen, H.J.L.; Mathur, S.B.; Smedegaard-Petersen, V. Suppression of Rice Blast by Preinoculation with Avirulent Pyricularia Oryzae and the Nonrice Pathogen Bipolaris Sorokiniana. Phytopathology 1998, 7, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Nasir, F.; Tian, L.; Chang, C.; Li, X.; Gao, Y.; Tran, L.P.; Tian, C. Current Understanding of Pattern-Triggered Immunity and Hormone-Mediated Defense in Rice (Oryza Sativa) in Response to Magnaporthe Oryzae Infection. Semin Cell Dev Biol 2018, 83, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, H.; Urayama, S.I.; Higashiura, T.; Le, T.M.; Komatsu, K. Chrysoviruses in Magnaporthe Oryzae. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, D.S.; Hwang, B.K. The Pepper Rna-Binding Protein Carbp1 Functions in Hypersensitive Cell Death and Defense Signaling in the Cytoplasm. Plant J 2012, 72, 235–248. [Google Scholar] [CrossRef]
- Nina and F. J. C. O. i., P. Biology. Rna-Binding Proteins in Plants: The Tip of an Iceberg? 2002.
- Lorković, Z.J. Role of Plant Rna-Binding Proteins in Development, Stress Response and Genome Organization. 2009, 14, 229–236.
- Woloshen, V.; Huang, S.; Li, X. Review Article Rna-Binding Proteins in Plant Immunity. 2011.
- Zhai, K.; Deng, Y.; Liang, D.; Tang, J.; Liu, J.; Yan, B.; Yin, X.; Lin, H.; Chen, F.; Yang, D.; Xie, Z.; Liu, J.-Y.; Li, Q.; Zhang, L.; He, Z. Rrm Transcription Factors Interact with Nlrs and Regulate Broad-Spectrum Blast Resistance in Rice. Mol Cell 2019, 74, 996–1009. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. Tbtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Molecular Plant 2020, 13. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, N.; Wang, X.; Yi, Q.; Zhu, D.; Lai, Y.; Zhao, Y. Analysis of Rice Snf2 Family Proteins and Their Potential Roles in Epigenetic Regulation. Plant Physiol Biochem 2013, 70, 33–42. [Google Scholar] [CrossRef]
- Mimura, M.; Itoh, J. Genetic Interaction between Rice Plastochron Genes and the Gibberellin Pathway in Leaf Development. Rice (N Y) 2014, 7, 25. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Itoh, J.-I.; Miyoshi, K.; Kurata, N.; Alvarez, N.; Veit, B.; Nagato, Y. Plastochron2 Regulates Leaf Initiation and Maturation in Rice. The Plant Cell 2006, 18, 612–625. [Google Scholar] [CrossRef]
- Xiong, G.S.; Hu, X.M.; Jiao, Y.Q.; Yu, Y.C.; Chu, C.C.; Li, J.Y.; Qian, Q.; Wang, Y.H. Leafy Head2, Which Encodes a Putative Rna-Binding Protein, Regulates Shoot Development of Rice. Cell Res 2006, 16, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Wang, D.; Duan, P.; Liu, Y.; Huang, K.; Zeng, D.; Zhang, L.; Dong, G.; Li, Y.; Xu, R.; Zhang, B.; Huang, X.; Li, N.; Wang, Y.; Qian, Q.; Li, Y. . Control of Grain Size and Weight by the Gsk2-Large1/Oml4 Pathway in Rice. Plant Cell 2020, 32, 1905–1918. [Google Scholar] [CrossRef]
- Isshiki, M.; Matsuda, Y.; Takasaki, A.; Wong, H.L.; Satoh, H.; Shimamoto, K.J.P.B. Du3, a Mrna Cap-Binding Protein Gene, Regulates Amylose Content in Japonica Rice Seeds. Plant Biotechnology 2008. [CrossRef]
- Yano, M.; Okuno, K.; Satoh, H.; Omura, T. Chromosomal Location of Genes Conditioning Low Amylose Content of Endosperm Starches in Rice, Oryza Sativa L. Theor Appl Genet 1988, 76, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zhai, K.; Deng, Y.; Liang, D.; Tang, J.; Liu, J.; Yan, B.; Yin, X.; Lin, H.; Chen, F.; Yang, D.; Xie, Z.; Liu, J.Y.; Li, Q.; Zhang, L.; He, Z. Rrm Transcription Factors Interact with Nlrs and Regulate Broad-Spectrum Blast Resistance in Rice. Mol Cell 2019, 74, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, X.; Zhang, H.; Cheng, Z.; Liu, J.; Zhou, C.; Luo, S.; Luo, W.; Li, S.; Xing, X.; Chang, Y.; Shi, C.; Ren, Y.; Zhu, S.; Lei, C.; Guo, X.; Wang, J.; Zhao, Z.; Wang, H.; Zhai, H.; Lin, Q.; Wan, J. Dwarf and High Tillering1 Represses Rice Tillering through Mediating the Splicing of D14 Pre-Mrna. Plant Cell 2022, 34, 3301–3318. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lan, J.; Huang, Y.; Cao, P.; Zhou, C.; Ren, Y.; He, N.; Liu, S.; Tian, Y.; Nguyen, T.; Jiang, L.; Wan, J. Wsl5, a Pentatricopeptide Repeat Protein, Is Essential for Chloroplast Biogenesis in Rice under Cold Stress. J Exp Bot 2018, 69, 3949–3961. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Ding, S.; Reiland, S.; Rödiger, A.; Roschitzki, B.; Xue, P.; Gruissem, W.; Lu, C.; Baginsky, S. Identification and Characterization of Chloroplast Casein Kinase Ii from Oryza Sativa (Rice). J Exp Bot 2015, 66, 175–187. [Google Scholar] [CrossRef]
- Conservation and Divergence of Fca Function between Arabidopsis and Rice. Plant Molecular Biology 2005, 58, 823–838. [CrossRef]
- Chen, S.Y.; Wang, Z.Y.; Cai, X.L. Osrrm, a Spen-Like Rice Gene Expressed Specifically in the Endosperm. Cell Res 2007, 17, 713–721. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Wang, Z.-Y.; Cai, X.-L. Osrrm, a Spen-Like Rice Gene Expressed Specifically in the Endosperm. Cell Res 2007, 17, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Kwak, K.J.; Jung, H.J.; Lee, K.H.; Kim, Y.S.; Kim, W.Y.; Ahn, S.J.; Kang, H. The Minor Spliceosomal Protein U11/U12-31k Is an Rna Chaperone Crucial for U12 Intron Splicing and the Development of Dicot and Monocot Plants. PLoS One 2012, 7, e43707. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Vega-Sánchez, M.E.; Park, C.H.; Bellizzi, M.; Guo, Z.; Wang, G.L. Rbs1, an Rna Binding Protein, Interacts with Spin1 and Is Involved in Flowering Time Control in Rice. PLoS One 2014, 9, e87258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Ossro1a Interacts with Rna Binding Domain-Containing Protein (Osrbd1) and Functions in Abiotic Stress Tolerance in Yeast. Front Plant Sci 2016, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Ren, L.; Zhao, Y.; You, H.; Zhou, Y.; Tang, D.; Du, G.; Shen, Y.; Li, Y.; Cheng, Z. Reproductive Cells and Peripheral Parietal Cells Collaboratively Participate in Meiotic Fate Acquisition in Rice Anthers. Plant J 2021, 108, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.; Sato, Y.; Asano, T.; Nagamura, Y.; Nonomura, K. Rice Mel2, the Rna Recognition Motif (Rrm) Protein, Binds in Vitro to Meiosis-Expressed Genes Containing U-Rich Rna Consensus Sequences in the 3’-Utr. Plant Mol Biol 2015, 89, 293–307. [Google Scholar] [CrossRef]
- Nonomura, K.; Eiguchi, M.; Nakano, M.; Takashima, K.; Komeda, N.; Fukuchi, S.; Miyazaki, S.; Miyao, A.; Hirochika, H.; Kurata, N. A Novel Rna-Recognition-Motif Protein Is Required for Premeiotic G1/S-Phase Transition in Rice (Oryza Sativa L.). PLoS Genet 2011, 7, e1001265. [Google Scholar] [CrossRef]
- Sahi, C.; Agarwal, M.; Singh, A.; Grover, A. Molecular Characterization of a Novel Isoform of Rice (Oryza Sativa L.) Glycine Rich-Rna Binding Protein and Evidence for Its Involvement in High Temperature Stress Response. Plant Science 2007, 173, 144–155. [Google Scholar] [CrossRef]
- Cléry, A.; Blatter, M.; Allain, F.H.T. Rna Recognition Motifs: Boring? Not Quite. Curr Opin Struct Biol 2008, 18, 290–298. [Google Scholar] [CrossRef]
- Lorković, Z.J.; Barta, A. Genome Analysis: Rna Recognition Motif (Rrm) and K Homology (Kh) Domain Rna-Binding Proteins from the Flowering Plant Arabidopsis Thaliana. Nucleic Acids Research 2002, 30, 623–635. [Google Scholar] [CrossRef]
- Shi, X.; Hanson, M.R.; Bentolila, S. Functional Diversity of Arabidopsis Organelle-Localized Rna-Recognition Motif-Containing Proteins. Wiley Interdiscip Rev Rna 2017, e1420. [Google Scholar] [CrossRef]
- Shi, X.; Hanson, M.R.; Bentolila, S. Functional Diversity of Arabidopsis Organelle-Localized Rna-Recognition Motif-Containing Proteins. Wiley Interdiscip Rev Rna 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Qian, B.; Cao, F.; Wu, W.; Yang, L.; Guan, Q.; Gu, X.; Wang, P.; Okusolubo, T.A.; Dunn, S.L.; Zhu, J.-K.; Zhu, J. An Arabidopsis Pwi and Rrm Motif-Containing Protein Is Critical for Pre-Mrna Splicing and Aba Responses. Nat Commun 2015, 6, 8139. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.J.; Jang, B.; Jung, C.H.; Ahn, S.-J.; Goh, C.H.; Cho, K.; Han, O.; Kang, H. Functional Characterization of a Glycine-Rich Rna-Binding Protein 2 in Arabidopsis Thaliana under Abiotic Stress Conditions. The Plant journal 2007, 3, 439–451. [Google Scholar] [CrossRef]
- Xu, T.; Lee, K.; Gu, L.; Kim, J.-I.; Kang, H. Functional Characterization of a Plastid-Specific Ribosomal Protein Psrp2 in Arabidopsis Thaliana under Abiotic Stress Conditions. Plant Physiology and Biochemistry 2013, 73, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Cold Shock Domain Proteins and Glycine-Rich Rna-Binding Proteins from Arabidopsis Thaliana Can Promote the Cold Adaptation Process in Escherichia Coli. Nucleic Acids Research 2007.
- Wang, S.; Bai, G.; Wang, S.; Yang, L.; Yang, F.; Wang, Y.; Zhu, J.K.; Hua, J. Chloroplast Rna-Binding Protein Rbd1 Promotes Chilling Tolerance through 23s Rrna Processing in Arabidopsis. PLoS Genet 2016, 12, e1006027. [Google Scholar] [CrossRef]
- Shi, X.; Germain, A.; Hanson, M.R.; Bentolila, S. Rna Recognition Motif-Containing Protein Orrm4 Broadly Affects Mitochondrial Rna Editing and Impacts Plant Development and Flowering. Plant Physiol 2016, 170, 294–309. [Google Scholar] [CrossRef]
- Vermel, M.; Guermann, B.; Delage, L.; Grienenberger, J.M.; Maréchal-Drouard, L.; Gualberto, J.M. A Family of Rrm-Type Rna-Binding Proteins Specific to Plant Mitochondria. Proc Natl Acad Sci U S A 2002, 99, 5866–5871. [Google Scholar] [CrossRef]
- Kwak, K.J.; Kim, Y.O.; Kang, H. Characterization of Transgenic Arabidopsis Plants Overexpressing Gr-Rbp4 under High Salinity, Dehydration, or Cold Stress. J Exp Bot 2005, 56, 3007–3016. [Google Scholar] [CrossRef]
- Lorković, Z.J. Role of Plant Rna-Binding Proteins in Development, Stress Response and Genome Organization. Trends Plant Sci 2009, 14, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Shuai, *!!! REPLACE !!!*; Wan, *!!! REPLACE !!!*; Shu, *!!! REPLACE !!!*; Wan, *!!! REPLACE !!!*; Leiyun, *!!! REPLACE !!!*; Yang, *!!! REPLACE !!!*; Fen, *!!! REPLACE !!!*; Genetics, Y.J.P. Shuai; Wan; Shu; Wan; Leiyun; Yang; Fen; Genetics, Y.J.P. Chloroplast Rna-Binding Protein Rbd1 Promotes Chilling Tolerance through 23s Rrna Processing in Arabidopsis. 2016.
Figure 1.
The conserved domain of OsRRM1 gene family.
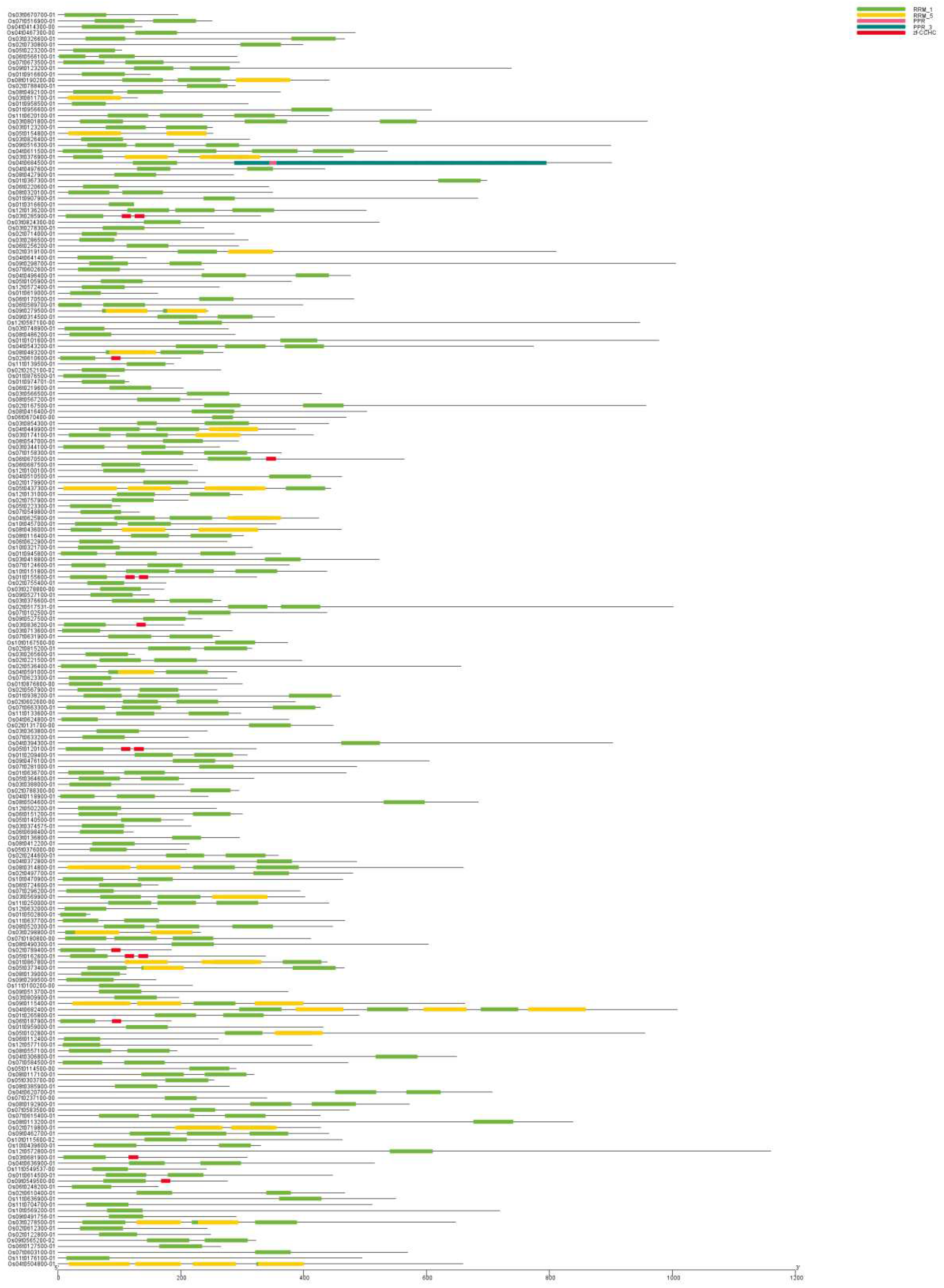
Figure 2.
Chromosome mapping of OsRRM1 gene family.
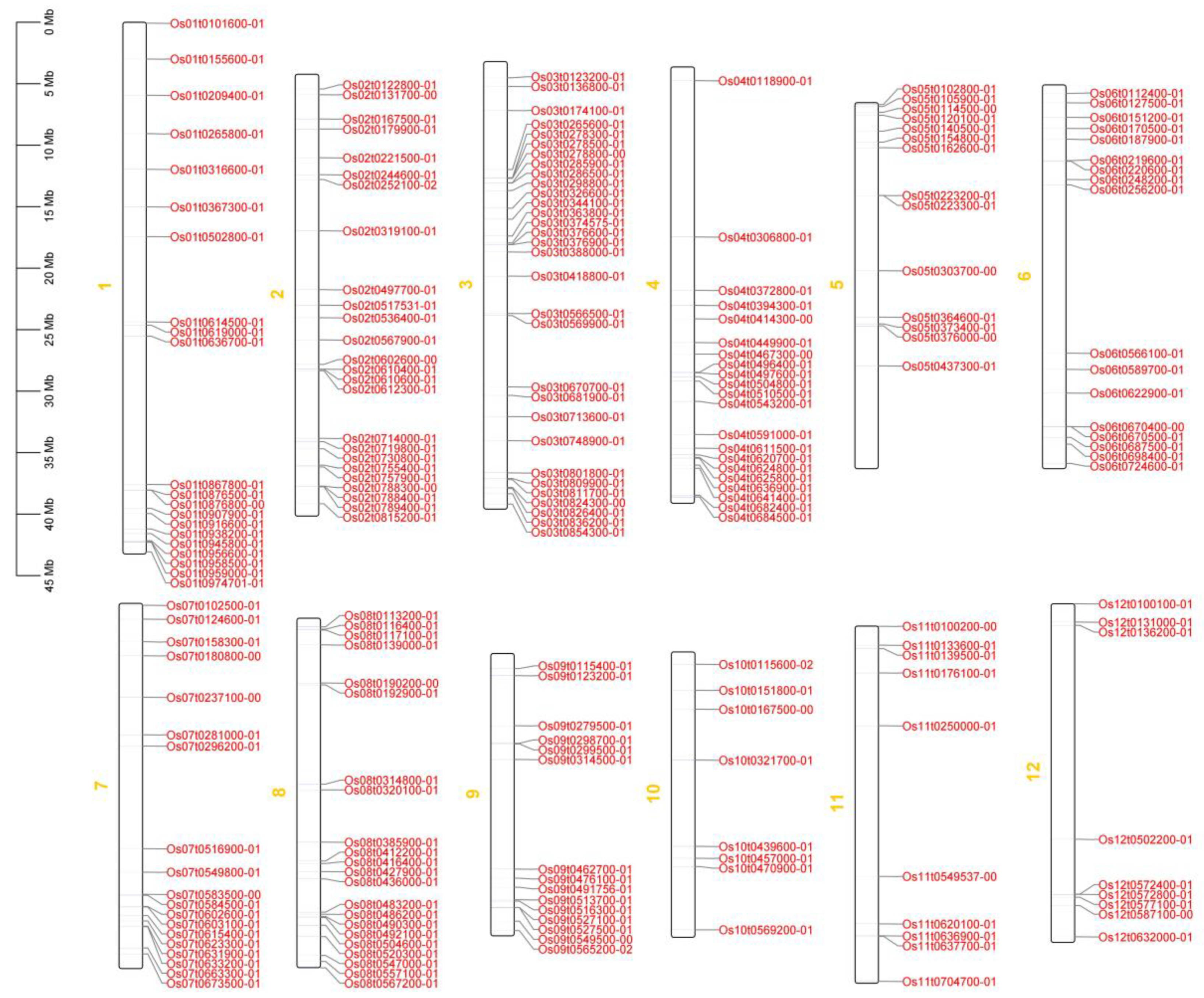
Figure 3.
Phylogenetic tree of OsRRM1 gene family.
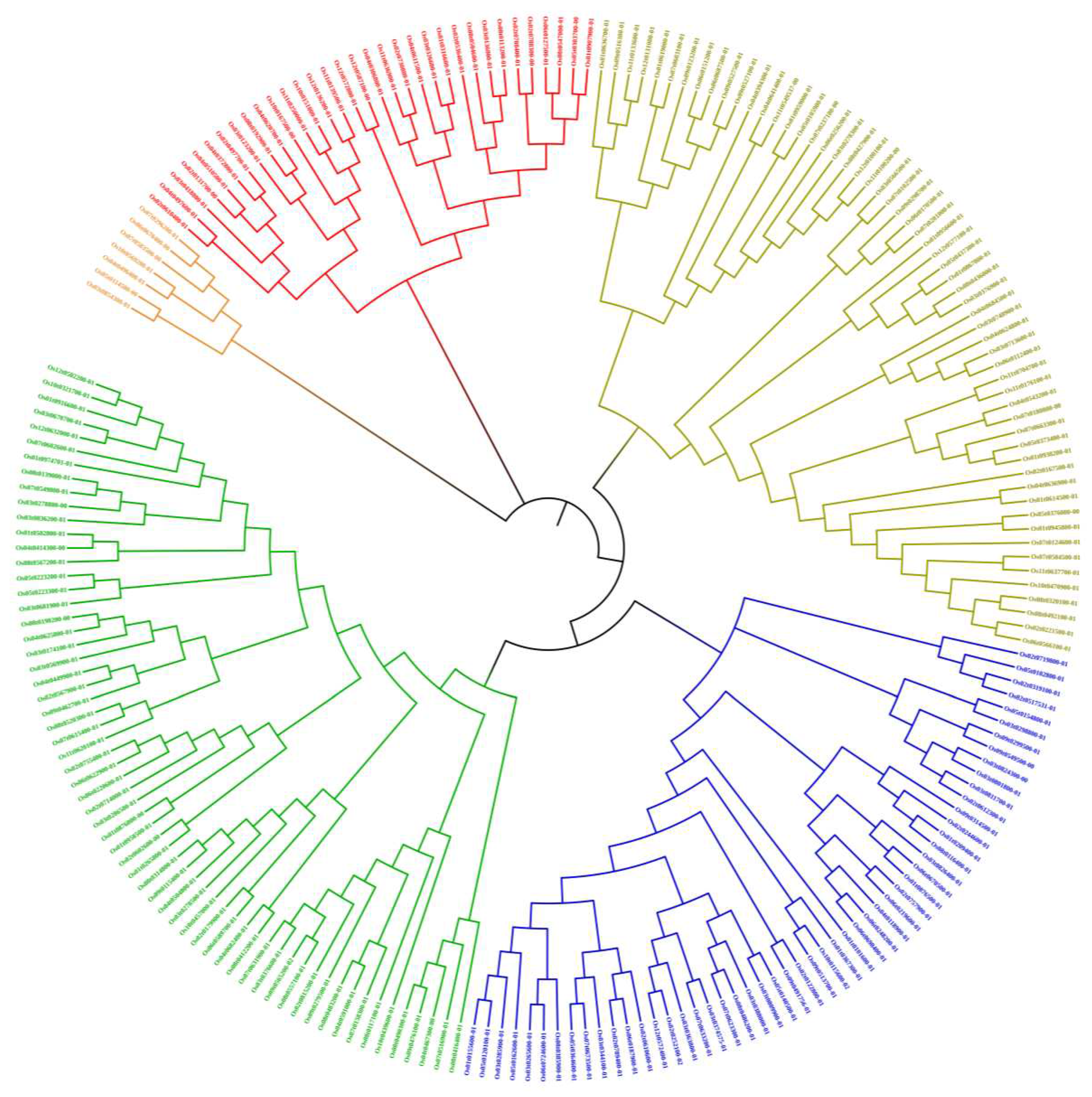
Figure 4.
Motif analysis and prediction of OsRRM1 gene family(left) and Schematic diagram of gene structure of OsRRM1 gene family(right).
Figure 4.
Motif analysis and prediction of OsRRM1 gene family(left) and Schematic diagram of gene structure of OsRRM1 gene family(right).

Figure 5.
The synteny analysis RRM1 gene family in Arabidopsis thaliana and rice.
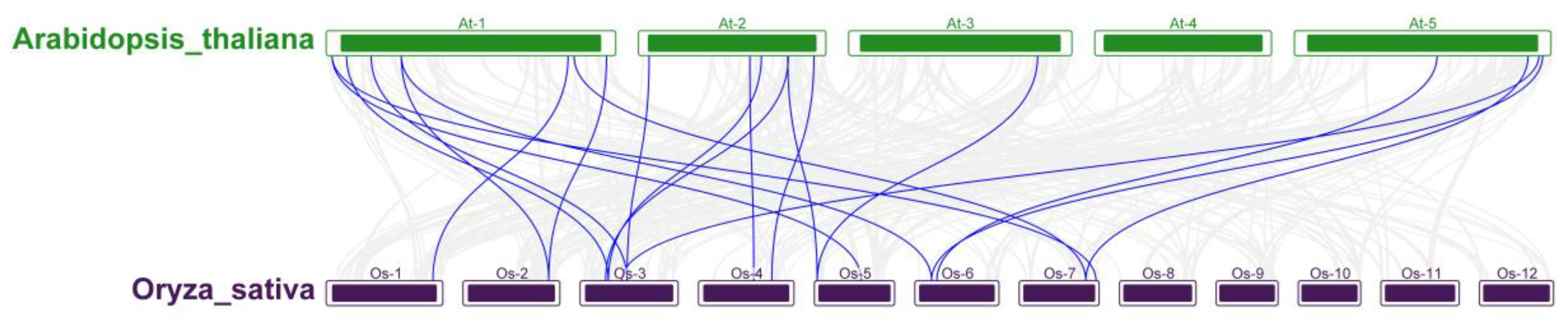
Figure 6.
Geng function enrichment analysis. (A) Analysis of GO of OsRRM1 gene family; (B) KEGG annotation of OsRRM1 gene family.
Figure 6.
Geng function enrichment analysis. (A) Analysis of GO of OsRRM1 gene family; (B) KEGG annotation of OsRRM1 gene family.
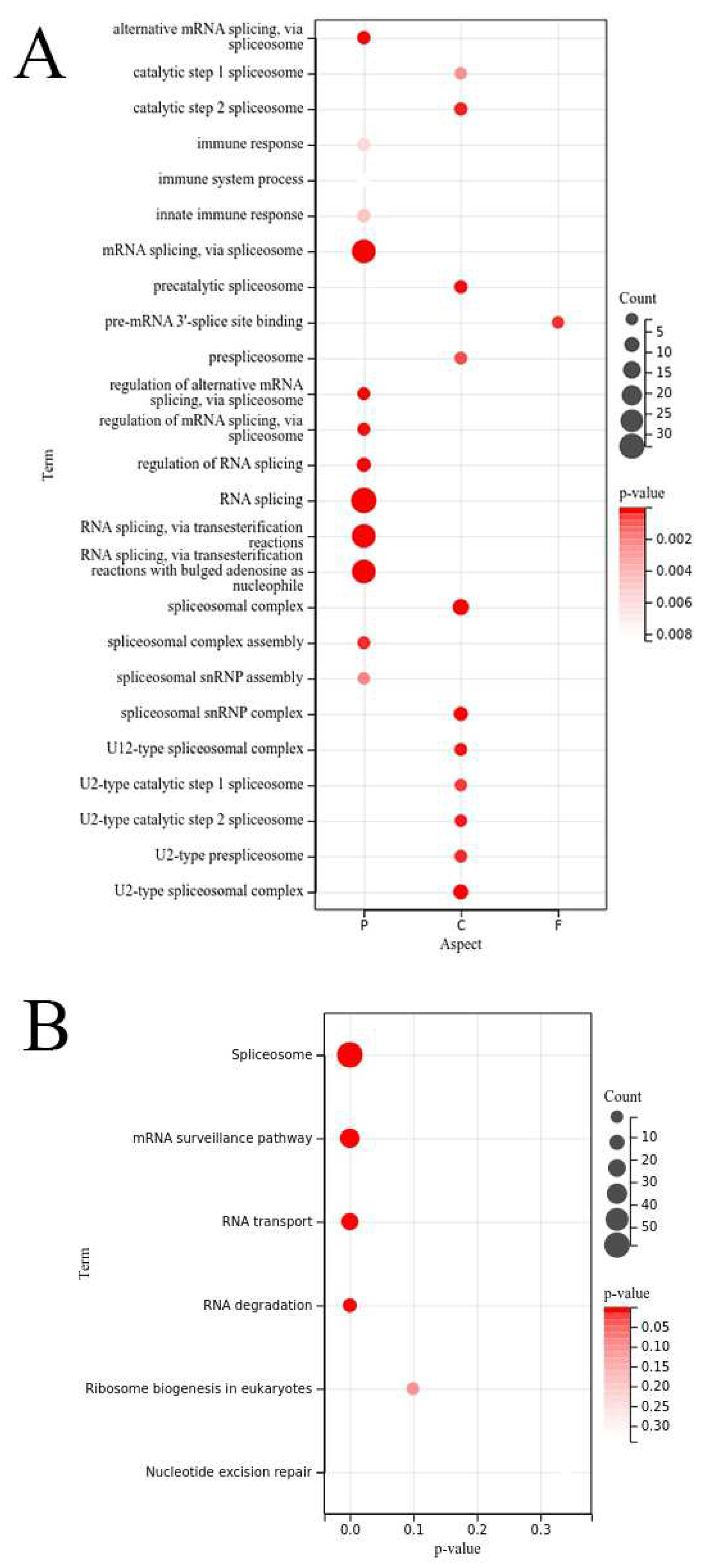
Figure 7.
Predictive cis-regulatory elements in the promoter of OsRRM1 gene family.
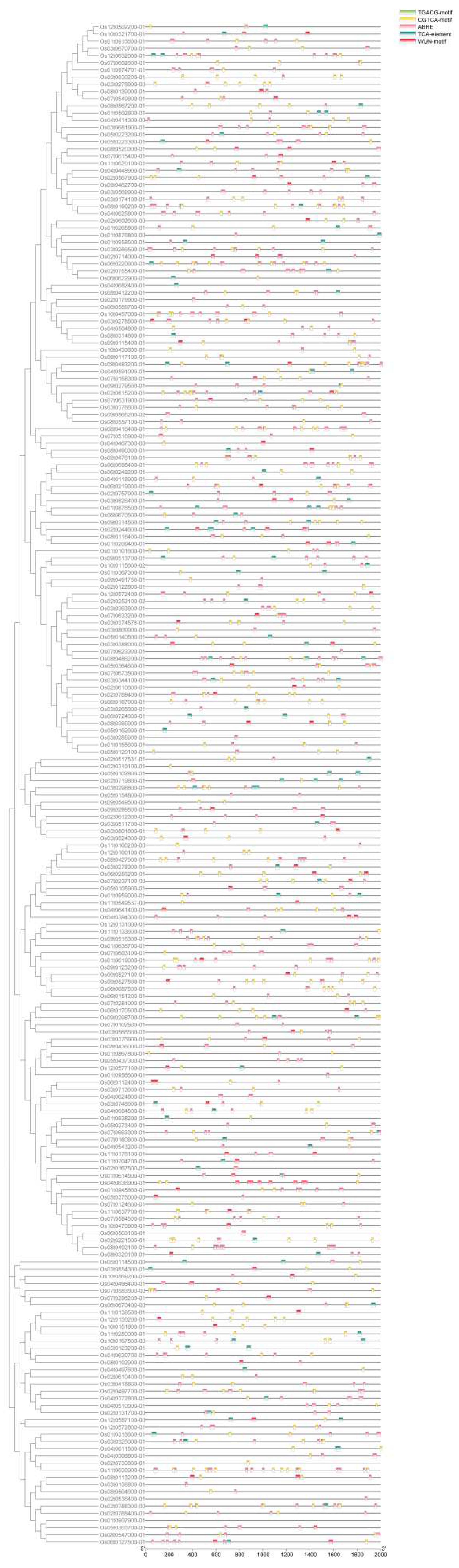
Figure 8.
Expression heat map of OsRRM1 gene family treated with Magnaporthe oryzae.
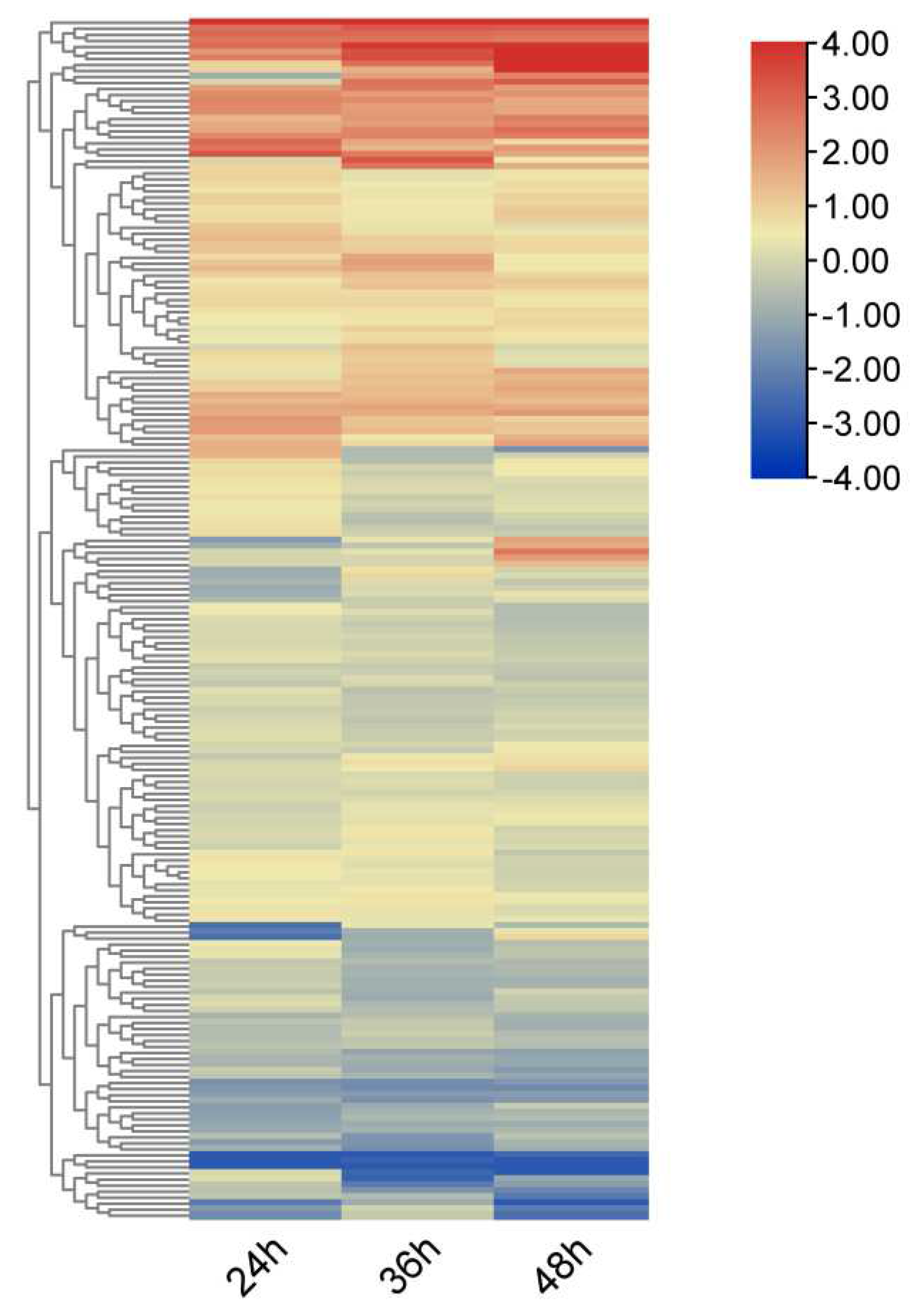
Figure 9.
Expression levels of OsRRM1 gene at different time periods under rice blast stress.
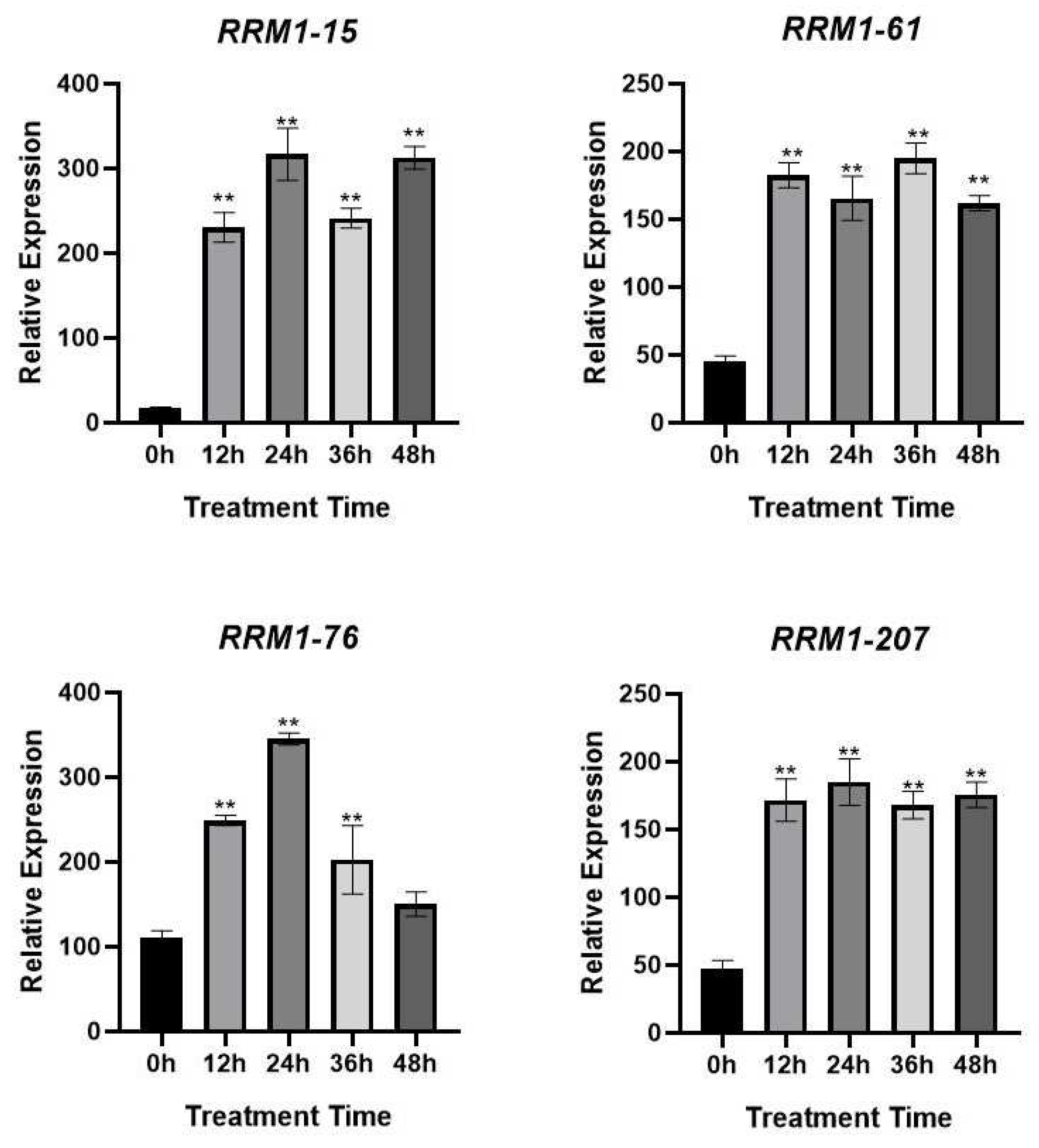

Table 1.
Specific primers for OsRRM1 gene.
| Gene | F-primer | R-primer |
|---|---|---|
| RRM1-15 | GGATGTGACTGAAGCTCGGGTGATC | GGATGTGACTGAAGCTCGGGTGATC |
| RRM1-61 | GGAGGTCTTGGAAGCCAAGGTCATC | CCATCCATGTCAGCGCCATCAAG |
| RRM1-76 | CACTGAAGCAAAGGTGGTTTTTGAC | GAGCTTTATCGACAGTGATCGCC |
| RRM1-207 | CTTGGATGGAAAGGATCTCGATGG | CATAGCCACCGCCTCCATAG |
| Actin | CCAATCGTGAGAAGATGACCCA | CCATCAGGAAGCTCGTAGCTCT |
Table 2.
Basic information of OsRRM1 gene identified in rice.
| Gene | RAP | Number of Amino Acid | Molecular Weight | pI | subcellular localization |
|---|---|---|---|---|---|
| RRM1-1 | Os01g0101600 | 978 | 106323.33 | 9.56 | nucleus |
| RRM1-2 | Os01g0155600 | 324 | 36893.16 | 11.27 | nucleus |
| RRM1-3 | Os01g0209400 | 308 | 33656.38 | 8.94 | nucleus |
| RRM1-4 | Os01g0265800 | 490 | 49279.96 | 5.09 | nucleus |
| RRM1-5 | Os01g0316600 | 124 | 13965.3 | 9.91 | chloroplast |
| RRM1-6 | Os01g0367300 | 698 | 79763.67 | 10.57 | nucleus |
| RRM1-7 | Os01g0502800 | 53 | 5837.75 | 10.27 | chloroplast |
| RRM1-8 | Os01g0614500 | 447 | 44269.69 | 8.43 | nucleus |
| RRM1-9 | Os01g0619000 | 163 | 18016.54 | 5.76 | extracellular space |
| RRM1-10[33] | Os01g0636700 | 469 | 52108.23 | 8.63 | nucleus |
| RRM1-11 | Os01g0867800 | 439 | 49316.8 | 6.46 | nucleus |
| RRM1-12 | Os01g0876500 | 100 | 11385 | 7.72 | chloroplast |
| RRM1-13 | Os01g0876800 | 300 | 31951.79 | 8.91 | extracellular space |
| RRM1-14[34,35,36] | Os01g0907900 | 683 | 71779.19 | 6.37 | nucleus |
| RRM1-15 | Os01g0916600 | 150 | 15546.9 | 8.01 | chloroplast thylakoid lumen |
| RRM1-16 | Os01g0938200 | 460 | 48888.22 | 8.72 | nucleus |
| RRM1-17 | Os01g0945800 | 363 | 40073.63 | 6.61 | nucleus |
| RRM1-18 | Os01g0956600 | 608 | 68184.84 | 7.8 | nucleus |
| RRM1-19 | Os01g0958500 | 310 | 31802.78 | 8.34 | nucleus |
| RRM1-20 | Os01g0959000 | 432 | 48111.49 | 12.37 | chloroplast thylakoid lumen |
| RRM1-21 | Os01g0974701 | 116 | 12459.19 | 9.74 | mitochondrion |
| RRM1-22 | Os02g0122800 | 249 | 28953.55 | 10 | nucleus |
| RRM1-23 | Os02g0131700 | 448 | 49261.45 | 5.02 | nucleus |
| RRM1-24 | Os02g0167500 | 957 | 105767.07 | 7.88 | extracellular space |
| RRM1-25 | Os02g0179900 | 240 | 28105.95 | 8.85 | nucleus |
| RRM1-26 | Os02g0221500 | 397 | 40265.08 | 5.63 | nucleus |
| RRM1-27 | Os02g0244600 | 359 | 38737.53 | 5.62 | nucleus |
| RRM1-28 | Os02g0252100 | 265 | 30466.57 | 11.09 | nucleus |
| RRM1-29 | Os02g0319100 | 811 | 90295.23 | 6.27 | nucleus |
| RRM1-30 | Os02g0497700 | 480 | 50879.51 | 5.01 | nucleus |
| RRM1-31[37] | Os02g0517531 | 1001 | 110368.83 | 6.39 | nucleus |
| RRM1-32 | Os02g0536400 | 656 | 74812.42 | 9.43 | nucleus |
| RRM1-33 | Os02g0567900 | 259 | 28284.5 | 9.18 | nucleus |
| RRM1-34 | Os02g0602600 | 386 | 41584.82 | 7.67 | nucleus |
| RRM1-35 | Os02g0610400 | 467 | 51689.83 | 5.56 | nucleus |
| RRM1-36 | Os02g0610600 | 200 | 22797.31 | 11.33 | nucleus |
| RRM1-37[38,39] | Os02g0612300 | 243 | 28573.22 | 5.44 | chloroplast |
| RRM1-38 | Os02g0714000 | 287 | 30609.94 | 9.32 | nucleus |
| RRM1-39 | Os02g0719800 | 428 | 47331.12 | 5.57 | nucleus |
| RRM1-40 | Os02g0730800 | 399 | 43547.8 | 6.15 | extracellular space |
| RRM1-41 | Os02g0755400 | 176 | 18512.61 | 9.99 | mitochondrion |
| RRM1-42 | Os02g0757900 | 212 | 24083.82 | 5.07 | nucleus |
| RRM1-43 | Os02g0788300 | 295 | 32235.18 | 7.72 | nucleus |
| RRM1-44 | Os02g0788400 | 289 | 32009.09 | 8.66 | nucleus |
| RRM1-45 | Os02g0789400 | 185 | 21023.33 | 11.24 | nucleus |
| RRM1-46 | Os02g0815200 | 316 | 34612.01 | 5.17 | chloroplast thylakoid lumen |
| RRM1-47 | Os03g0123200 | 252 | 28108.69 | 7.64 | nucleus |
| RRM1-48 | Os03g0136800 | 296 | 32305.94 | 9.02 | nucleus |
| RRM1-49 | Os03g0174100 | 416 | 46056.33 | 5.35 | nucleus |
| RRM1-50 | Os03g0265600 | 125 | 13993.55 | 7.86 | chloroplast |
| RRM1-51 | Os03g0278300 | 238 | 24720.42 | 9.83 | chloroplast |
| RRM1-52 | Os03g0278500 | 647 | 72627.76 | 8.43 | nucleus |
| RRM1-53 | Os03g0278800 | 173 | 18433.86 | 9.3 | chloroplast outer membrane |
| RRM1-54 | Os03g0285900 | 330 | 37042.2 | 11 | nucleus |
| RRM1-55 | Os03g0286500 | 310 | 32704.09 | 9 | extracellular space |
| RRM1-56 | Os03g0298800 | 232 | 26100.86 | 9.44 | chloroplast |
| RRM1-57 | Os03g0326600 | 467 | 51073.78 | 9.06 | nucleus |
| RRM1-58 | Os03g0344100 | 264 | 29782.1 | 10.08 | nucleus |
| RRM1-59 | Os03g0363800 | 243 | 27781.69 | 10.83 | nucleus |
| RRM1-60 | Os03g0374575 | 217 | 25589.48 | 11.17 | nucleus |
| RRM1-61 | Os03g0376600 | 265 | 28556.57 | 4.5 | chloroplast outer membrane |
| RRM1-62 | Os03g0376900 | 464 | 49564.37 | 6.39 | nucleus |
| RRM1-63 | Os03g0388000 | 205 | 24739.51 | 10.27 | nucleus |
| RRM1-64 | Os03g0418800 | 523 | 56761.18 | 8.75 | chloroplast |
| RRM1-65 | Os03g0566500 | 429 | 46194.37 | 9.62 | chloroplast |
| RRM1-66 | Os03g0569900 | 402 | 43945.82 | 5.34 | extracellular space |
| RRM1-67 | Os03g0670700 | 196 | 20375.4 | 6.73 | nucleus |
| RRM1-68 | Os03g0681900 | 308 | 34036.6 | 9.05 | nucleus |
| RRM1-69[40] | Os03g0713600 | 284 | 30904.71 | 5.06 | nucleus |
| RRM1-70 | Os03g0748900 | 278 | 29986.94 | 9.23 | nucleus |
| RRM1-71 | Os03g0801800 | 959 | 105396.52 | 9.48 | nucleus |
| RRM1-72 | Os03g0809900 | 197 | 21969.34 | 5.2 | nucleus |
| RRM1-73 | Os03g0811700 | 130 | 14710.82 | 9.49 | chloroplast |
| RRM1-74 | Os03g0824300 | 523 | 58186.08 | 7.22 | nucleus |
| RRM1-75 | Os03g0826400 | 312 | 36258.57 | 9.25 | nucleus |
| RRM1-76 | Os03g0836200 | 205 | 21823.38 | 8.29 | nucleus |
| RRM1-77 | Os03g0854300 | 441 | 48288.94 | 10.11 | nucleus |
| RRM1-78 | Os04g0118900 | 245 | 28783.89 | 9.94 | nucleus |
| RRM1-79 | Os04g0306800 | 649 | 72026.14 | 9.09 | nucleus |
| RRM1-80 | Os04g0372800 | 486 | 51446 | 5.1 | nucleus |
| RRM1-81 | Os04g0394300 | 903 | 97243.83 | 8.7 | nucleus |
| RRM1-82 | Os04g0414300 | 137 | 15074.25 | 9.93 | chloroplast |
| RRM1-83 | Os04g0449900 | 387 | 41807.64 | 8.68 | extracellular space |
| RRM1-84 | Os04g0467300 | 484 | 51314.72 | 7.33 | nucleus |
| RRM1-85 | Os04g0496400 | 476 | 53576.63 | 4.69 | nucleus |
| RRM1-86 | Os04g0497600 | 435 | 48295.81 | 5.49 | nucleus |
| RRM1-87 | Os04g0504800 | 659 | 71231.24 | 8.95 | extracellular space |
| RRM1-88 | Os04g0510500 | 462 | 51785.72 | 5.01 | nucleus |
| RRM1-89 | Os04g0543200 | 774 | 86649.43 | 5.64 | nucleus |
| RRM1-90 | Os04g0591000 | 291 | 31672.86 | 6.05 | mitochondrion |
| RRM1-91 | Os04g0611500 | 536 | 60240.64 | 9.16 | nucleus |
| RRM1-92 | Os04g0620700 | 707 | 75253.44 | 4.85 | nucleus |
| RRM1-93 | Os04g0624800 | 376 | 40858.93 | 5.59 | nucleus |
| RRM1-94 | Os04g0625800 | 425 | 46195.8 | 5.99 | extracellular space |
| RRM1-95[41] | Os04g0636900 | 515 | 52204.84 | 5.79 | nucleus |
| RRM1-96 | Os04g0641400 | 144 | 16026.58 | 4.61 | nucleus |
| RRM1-97 | Os04g0682400 | 1008 | 110200.99 | 6.17 | nucleus |
| RRM1-98[42] | Os04g0684500 | 901 | 101135.53 | 6.65 | chloroplast inner membrane |
| RRM1-99 | Os05g0102800 | 955 | 104522 | 6.01 | nucleus |
| RRM1-100 | Os05g0105900 | 380 | 42434.11 | 12.18 | nucleus |
| RRM1-101 | Os05g0114500 | 290 | 32890.31 | 6.85 | nucleus |
| RRM1-102 | Os05g0120100 | 323 | 36222.41 | 10.83 | nucleus |
| RRM1-103 | Os05g0140500 | 204 | 22104.33 | 5.18 | nucleus |
| RRM1-104 | Os05g0154800 | 253 | 28203.66 | 9.2 | cytoplasm |
| RRM1-105 | Os05g0162600 | 338 | 39019.1 | 9.83 | nucleus |
| RRM1-106 | Os05g0223200 | 104 | 11486.44 | 8.03 | nucleus |
| RRM1-107 | Os05g0223300 | 102 | 11702.99 | 5.06 | nucleus |
| RRM1-108 | Os05g0303700 | 254 | 29800.11 | 8.77 | nucleus |
| RRM1-109 | Os05g0364600 | 319 | 36105.16 | 11.2 | nucleus |
| RRM1-110 | Os05g0373400 | 466 | 50213.29 | 8.1 | nucleus |
| RRM1-111 | Os05g0376000 | 209 | 23394.61 | 9.14 | nucleus |
| RRM1-112 | Os05g0437300 | 444 | 49754.23 | 6.41 | nucleus |
| RRM1-113[40] | Os06g0112400 | 261 | 27763.35 | 6.23 | nucleus |
| RRM1-114 | Os06g0127500 | 265 | 28209.55 | 7.14 | nucleus |
| RRM1-115 | Os06g0151200 | 300 | 32650.85 | 5 | nucleus |
| RRM1-116 | Os06g0170500 | 482 | 54009.89 | 8.12 | nucleus |
| RRM1-117 | Os06g0187900 | 185 | 21183.36 | 11.29 | nucleus |
| RRM1-118 | Os06g0219600 | 204 | 23178.94 | 5.19 | nucleus |
| RRM1-119 | Os06g0220600 | 343 | 36170.91 | 9.63 | chloroplast outer membrane |
| RRM1-120 | Os06g0248200 | 164 | 17952.57 | 5.98 | nucleus |
| RRM1-121 | Os06g0256200 | 294 | 31817.7 | 10.97 | nucleus |
| RRM1-122 | Os06g0566100 | 292 | 29810.49 | 9.33 | nucleus |
| RRM1-123 | Os06g0589700 | 399 | 43823.12 | 9.17 | nucleus |
| RRM1-124 | Os06g0622900 | 275 | 29594.2 | 8.39 | nucleus |
| RRM1-125 | Os06g0670400 | 469 | 53864.27 | 5.38 | nucleus |
| RRM1-126 | Os06g0670500 | 564 | 64975.32 | 5.63 | nucleus |
| RRM1-127 | Os06g0687500 | 219 | 23922.07 | 9.52 | endomembrane system |
| RRM1-128 | Os06g0698400 | 123 | 13222.7 | 5 | nucleus |
| RRM1-129 | Os06g0724600 | 164 | 18503.88 | 10.31 | nucleus |
| RRM1-130 | Os07g0102500 | 438 | 47703.93 | 9.53 | nucleus |
| RRM1-131 | Os07g0124600 | 377 | 41006.31 | 6.68 | nucleus |
| RRM1-132 | Os07g0158300 | 364 | 39084.91 | 4.61 | mitochondrion |
| RRM1-133 | Os07g0180800 | 411 | 46253.74 | 9.65 | nucleus |
| RRM1-134 | Os07g0237100 | 340 | 36144.67 | 10.27 | chloroplast |
| RRM1-135 | Os07g0281000 | 486 | 54334.99 | 6.72 | nucleus |
| RRM1-136 | Os07g0296200 | 394 | 43291.14 | 8.3 | nucleus |
| RRM1-137 | Os07g0516900 | 251 | 27613.79 | 6.3 | extracellular space |
| RRM1-138 | Os07g0549800 | 133 | 14421.25 | 9.41 | chloroplast outer membrane |
| RRM1-139 | Os07g0583500 | 474 | 54197.46 | 6.55 | extracellular space |
| RRM1-140 | Os07g0584500 | 472 | 50477.44 | 5.94 | nucleus |
| RRM1-141 | Os07g0602600 | 238 | 23564.23 | 8.54 | mitochondrion |
| RRM1-142 | Os07g0603100 | 569 | 62175.74 | 6.15 | nucleus |
| RRM1-143 | Os07g0615400 | 427 | 46723.56 | 7.19 | nucleus |
| RRM1-144 | Os07g0623300 | 275 | 32242.91 | 11.35 | nucleus |
| RRM1-145[43] | Os07g0631900 | 264 | 28099.31 | 4.75 | chloroplast thylakoid lumen |
| RRM1-146 | Os07g0633200 | 213 | 24820.57 | 10.68 | nucleus |
| RRM1-147 | Os07g0663300 | 427 | 46493.89 | 9.17 | nucleus |
| RRM1-148 | Os07g0673500 | 296 | 33141.48 | 10.64 | nucleus |
| RRM1-149 | Os08g0113200 | 838 | 95016.62 | 5.47 | endomembrane system |
| RRM1-150 | Os08g0116400 | 302 | 32739.26 | 6.4 | nucleus |
| RRM1-151 | Os08g0117100 | 319 | 35941.88 | 6.02 | chloroplast outer membrane |
| RRM1-152 | Os08g0139000 | 111 | 11938.8 | 9.55 | chloroplast outer membrane |
| RRM1-153 | Os08g0190200 | 442 | 47809.63 | 5.86 | extracellular space |
| RRM1-154 | Os08g0192900 | 572 | 60393.68 | 4.98 | nucleus |
| RRM1-155 | Os08g0314800 | 660 | 71558.46 | 7.55 | nucleus |
| RRM1-156 | Os08g0320100 | 350 | 36738.65 | 9.22 | nucleus |
| RRM1-157 | Os08g0385900 | 279 | 32947.48 | 11.88 | nucleus |
| RRM1-158 | Os08g0412200 | 214 | 25104.07 | 10.05 | chloroplast |
| RRM1-159 | Os08g0416400 | 503 | 54742.2 | 7.66 | nucleus |
| RRM1-160 | Os08g0427900 | 286 | 30491.17 | 11.05 | nucleus |
| RRM1-161 | Os08g0436000 | 461 | 49888.14 | 6.46 | nucleus |
| RRM1-162 | Os08g0483200 | 269 | 29132.14 | 9.39 | mitochondrion |
| RRM1-163 | Os08g0486200 | 289 | 33541.09 | 11.8 | nucleus |
| RRM1-164 | Os08g0490300 | 603 | 64733.85 | 6.09 | nucleus |
| RRM1-165 | Os08g0492100 | 362 | 38125.83 | 9.22 | nucleus |
| RRM1-166 | Os08g0504600 | 684 | 75299.38 | 6.19 | nucleus |
| RRM1-167 | Os08g0520300 | 447 | 48765.28 | 6.86 | nucleus |
| RRM1-168 | Os08g0547000 | 294 | 31708.05 | 7.08 | nucleus |
| RRM1-169 | Os08g0557100 | 194 | 21388.83 | 4.95 | chloroplast |
| RRM1-170 | Os08g0567200 | 235 | 26254.56 | 9.77 | nucleus |
| RRM1-171 | Os09g0115400 | 662 | 71630.27 | 6.45 | mitochondrion |
| RRM1-172[44] | Os09g0123200 | 738 | 79658.29 | 9.09 | nucleus |
| RRM1-173 | Os09g0279500 | 245 | 26681.14 | 8.53 | chloroplast thylakoid lumen |
| RRM1-174[45] | Os09g0298700 | 1005 | 110844.59 | 6.79 | nucleus |
| RRM1-175 | Os09g0299500 | 160 | 17315.17 | 5.76 | extracellular space |
| RRM1-176 | Os09g0314500 | 353 | 38868.39 | 5.96 | nucleus |
| RRM1-177 | Os09g0462700 | 441 | 46949.78 | 8.52 | chloroplast |
| RRM1-178 | Os09g0476100 | 604 | 64263.07 | 6.3 | nucleus |
| RRM1-179 | Os09g0491756 | 290 | 34087.52 | 8.92 | nucleus |
| RRM1-180 | Os09g0513700 | 375 | 43193.42 | 9.74 | nucleus |
| RRM1-181[46] | Os09g0516300 | 900 | 97198.57 | 6.85 | nucleus |
| RRM1-182 | Os09g0527100 | 149 | 16616.66 | 8.8 | nucleus |
| RRM1-183 | Os09g0527500 | 235 | 25960.25 | 8.81 | nucleus |
| RRM1-184[47] | Os09g0549500 | 276 | 29500.33 | 9.18 | nucleus |
| RRM1-185 | Os09g0565200 | 322 | 35425.05 | 4.41 | mitochondrion |
| RRM1-186 | Os10g0115600 | 463 | 55113.96 | 9.1 | nucleus |
| RRM1-187 | Os10g0151800 | 438 | 47821.62 | 4.98 | nucleus |
| RRM1-188 | Os10g0167500 | 374 | 40267.56 | 3.97 | nucleus |
| RRM1-189 | Os10g0321700 | 317 | 32244.11 | 4.59 | chloroplast thylakoid lumen |
| RRM1-190 | Os10g0439600 | 330 | 34829.59 | 4.96 | nucleus |
| RRM1-191 | Os10g0457000 | 355 | 38849.39 | 8.55 | nucleus |
| RRM1-192 | Os10g0470900 | 464 | 45620.47 | 6.24 | nucleus |
| RRM1-193 | Os10g0569200 | 719 | 83181.8 | 4.98 | nucleus |
| RRM1-194 | Os11g0100200 | 219 | 24033.05 | 9.87 | nucleus |
| RRM1-195 | Os11g0133600 | 298 | 32998.39 | 7.65 | nucleus |
| RRM1-196 | Os11g0139500 | 189 | 21471.25 | 4.13 | extracellular space |
| RRM1-197 | Os11g0176100 | 495 | 52955.01 | 6.43 | extracellular space |
| RRM1-198[48] | Os11g0250000 | 441 | 48446.94 | 5.68 | nucleus |
| RRM1-199 | Os11g0549537 | 242 | 26479.77 | 6.08 | chloroplast |
| RRM1-200 | Os11g0620100 | 441 | 47561.26 | 6.86 | nucleus |
| RRM1-201 | Os11g0636900 | 550 | 61141.76 | 7.78 | nucleus |
| RRM1-202 | Os11g0637700 | 467 | 49048.64 | 8.44 | nucleus |
| RRM1-203 | Os11g0704700 | 511 | 57960.23 | 10.14 | chloroplast |
| RRM1-204[49] | Os12g0100100 | 228 | 24809.9 | 9.87 | nucleus |
| RRM1-205 | Os12g0131000 | 300 | 33277.87 | 8.81 | chloroplast |
| RRM1-206 | Os12g0136200 | 502 | 55072.87 | 5.03 | nucleus |
| RRM1-207 | Os12g0502200 | 258 | 25044.52 | 4.74 | mitochondrion |
| RRM1-208 | Os12g0572400 | 263 | 30186.19 | 10.9 | nucleus |
| RRM1-209[50,51,52] | Os12g0572800 | 1160 | 127816.97 | 8.61 | plasma membrane |
| RRM1-210 | Os12g0577100 | 414 | 47380.57 | 9.1 | nucleus |
| RRM1-211 | Os12g0587100 | 947 | 106893.09 | 9.14 | nucleus |
| RRM1-212[53] | Os12g0632000 | 162 | 16083.1 | 6.31 | nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



