
Submitted:
22 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
High Resolution Melting Analysis (HRM) has been pointed as a suitable alternative method to detect and identify Leishmania species. Herein, we aimed to evaluate the sensitivity, specificity, accuracy, and limitations of a HSP70-HRM protocol both as a diagnostic scheme applied in clinical samples and as a species typing tool for laboratory research and reference services. Our data reveal the pronounced species-typing potential of the HSP70-HRM in DNA from cultured parasites. For clinical samples, however, we advise caution due to the parasite load-dependent accuracy. In the light of these findings and considering the importance of parasite load determination for clinical and research purposes we recommend the integration of the presented typing scheme and the previously published Leishmania quantifying approach as combined tools for clinicians, surveillance, and research.
Keywords:
HRM
; Leishmania
; Americas
; leishmaniasis
; qPCR
; parasite typing
; HSP70
; diagnosis.
1. Introduction
Leishmaniasis is a worldwide distributed and neglected disease. caused by protozoan parasites of the genus Leishmania [1]. The complex of diseases encompasses various clinical forms; from visceral to disfiguring cutaneous lesions [2]. Genetic diversity in the Leishmania genus plays an important role within the complex factors that lead to the wide range of clinical outcomes. In the Americas. the visceral form is caused by Leishmania (Leishmania) infantum and the tegumentary leishmaniasis (TL) is associated with infection of 10 different species from the two subgenera (Leishmania and Viannia). In Brazil, the most common species causing TL are L. (V.) braziliensis, Leishmania (V.) guyanensis and L. (L.) amazonensis [3]. In the Amazon region of the country TL can also be caused by other species: L. (V.) naiffi. L. (V.) lainsoni. L. (V.) shawi and L. (V.) lindenbergi. The species L. (V.) utingensis, first detected in sandfly [4] is also present in the region and has been recently detected infecting human [5].
The diagnosis of leishmaniasis comprises the association of clinical, epidemiological and laboratory data. Clinical signs and symptoms, alone or in combination, are not always sufficient as they can be confused with other diseases. It is therefore essential to carry out the laboratory diagnosis, which can be done by parasitological diagnostic (microscopy), immunological, serological, and molecular tests. It is therefore essential to carry out the laboratory diagnosis, which can be done by direct (parasitological, culture, histopathological analysis, and PCR) or indirect methods based on the detection of anti-Leishmania antibodies - less used for TL diagnosis [6]. As stated, the species diversity within the Leishmania genus has a major role in the treatment, clinical outcome, and transmission cycles. Therefore, the identification of the infecting species directly in clinical samples (preferably) or by isolating the parasite, is still a relevant topic in the studies of leishmaniasis, especially in sympatric areas such as among countries and regions in the Americas.
Biological collections gather valuable material to explore microorganism diversity and its impact on the development of diagnostic and species typing tools. The Leishmania collection of Fiocruz (CLIOC; clioc.fiocruz) is a robust biobank, mainly for American Leishmania isolates, and is also a reference center for Leishmania species identification. The method applied for species typing is the Multilocus Enzyme Electrophosis (MLEE) - still the gold standard technique, but only applicable in successfully isolated and cultured parasites. MLEE has contributed hugely for typing [7] and for the epidemiology of leishmaniasis [8,9], but its drawbacks push researchers forward to the development of a cheaper and faster substitute method. Molecular-based approaches are currently the main target, and PCR-sequencing [10], PCR-RFLP are already applied in DNA from cultured parasites and from clinical material [11]. Both methods, however, involve a lot of steps, including gel electrophoresis. In this regard, protocols based on quantitative real-time PCR (qPCR) are good candidates because they eliminate the need for the detection of amplified product in gel, allow many samples to be assayed simultaneously and have the potential to be automatized. Moreover, it is especially relevant for clinical samples in which the parasite load affects sensitivity and accuracy, and the technique may potentially overcome this fact by detecting and quantifying minimal amounts of nucleic acids in a wide range of samples from different sources [12,13]. In this context, High Resolution Melting Analysis (HRM) has been pointed as a good and practical alternative to detect and identify Leishmania species, both as a diagnostic tool and as typing tool for laboratory work [14,15,16]. HRM is an adequate technique to detect mutations, single nucleotide polymorphisms (SNPs), and epigenetic differences in DNA samples. The analysis is based on the variance between the shape of melting curves and the difference between the samples melting temperatures (Tm). The Tm value of a sample can be obtained from the dissociation curve, defined as the temperature at which 50% of each DNA molecule is denatured [17].
A study developed by Zampieri et al [18] presented a HRM assay using as targets two regions of the HSP70 Leishmania gene [19] both within a 234 bp region already used to identify species, even from clinical samples [11]. The first pair of primers targets a conserved region of 144 bp that amplifies all Leishmania species; a second pair targets a smaller and more variable region of 104 bp, that identifies only Leishmania (Viannia) species [19]. Based on HRM analysis, these assays were able to differentiate eight Leishmania species found in the Americas: L. (L.) infantum, L. (L.) amazonenses, L. (L.) mexicana, L. (V.) lainsoni, L. (V.) braziliensis, L. (V.) guyanensis, L. (V.) naiffi and L. (V.) shawi; and 3 species found in Europe, Asia and Africa: L. (L.) tropica. L. (L.) donovani and L. (L.) major. The assays were validated in 16 experimentally infected golden hamster tissue samples, parasite strains isolated from humans, fresh tissue human samples or embedded in paraffin, experimentally infected BALB/c mice tissues and samples from naturally infected phlebotomines. The HRM analysis results were compared to previous genotyping by sequencing, and/or by MLEE, and presented a strong correlation between the methodologies. Finally, an algorithm was proposed to characterize by HRM the main Leishmania species circulating in Brazil [19].
Despite the potential offered by the published protocol [19], our group raised the concern on the following specific points: first, considering the well-known intraspecific genomic variation among L. (Viannia) strains, polymorphisms among distinct strains may affect the melting curve, melting temperature and ultimately the HRM typing efficiency. Additionally, some species were not assayed previously, and neither the intraspecific variation explored. Therefore, the effect of such diversity on the protocol must be investigated. By evaluating such aspects, we aim to appraise the potential of HRM to be applied as a substitute for MLEE. Secondly, an open question is how the accuracy, sensitivity and specificity vary when the method is applied on a panel of human clinical material displaying a broad range of parasite load. To address these subjects, we assayed a robust, distinctive and representative sampling of Leishmania DNA available at CLIOC and, equally importantly, human clinical samples with associated previous data of parasite load obtained by a HSP70 qPCR analytical study [20]. The samples were collected from patients with cutaneous leishmaniasis from a region known to present high intra and interspecific Leishmania diversity [20]. For the present study, we combine these described data and material aiming to evaluate the accuracy of the HRM protocol [18], both as a diagnostic method and as a possible substitute to the MLEE as a species typing tool for the reference service. The approach allowed unique and comprehensive inferences to be made on the HRM potential as a species typing tool and a diagnostic method for Leishmania.
2. Results
2.1. HRM HSP70 efficiently identifies the main Leishmania species with high reproducibility.
Sixteen Leishmania reference strains representing 14 distinct species (Supp. Table S1) were used for the adjustment of the HRM protocol. A sub-set of 13 samples was selected and the reproducibility test was attained by two independent experiments performed by the same operator in different days. An absolute reproducibility (100%) of the variable profiles generated by the software was observed (Table 1).
The typing outcome confirmed the separation of the main species circulating in Brazil, adding to the published information that L. (V.) lindenbergi and L. (V.) utingensis are also distinguishable by the algorithm. Clinically relevant co-circulating species in the Americas could not be distinguished using the strains evaluated in the present study: L. (V.) braziliensis vs L. (V.) peruviana, L. (V.) panamensis vs L. (V.) guyanensis. Based on the obtained data, some representative samples were selected as references for further assays. The schematic representation of the variant profiles obtained and melting curves are summarized in Figure 1 and Figure 2. respectively.
We further explored the possible effect of intraspecific diversity among Leishmania samples on HRM typing accuracy by assaying a larger and representative panel of strains hosted by CLIOC. Supplemental Table S2 depicts all strains used, including the information on whether samples were applied as a reference, as validation or during the reproducibility assays. All samples were previously characterized to determine the species by MLEE and/or DNA sequencing for the HSP70 target [10]. On total, 110 strains from distinct geographic areas of South America, mainly from the Amazon basin, were assayed, including three samples with hybrid profile by MLEE and by molecular typing methods (Table 2). To evaluate concordance between typing data, we excluded the references and the strains that could not be amplified by Primer 2. Results revealed concordance between MLEE/DNA sequencing and HRM typing data for 76/79 samples (96.2%). The HRM typing result of hybrid samples also converged with the other molecular typing approaches (Table 2).
Kappa index indicated values compatible to “Almost perfect agreement” (k=0.91) between MLEE/DNA sequencing and HRM HSP70 typing results. All DNA samples presenting divergent outcome (3/79) were sequenced a second time for the HSP70 region and BLAST hits (ncbi.gov) confirmed the MLEE-based described species (supplementary material). These samples, thus, represent appropriate examples of the intrinsic limits of the HRM species typing protocol.
2.2. HRM discordant results cannot be directly and exclusively associated with polymorphisms and/or Tms.
The Tms distribution of both Primers 1 and 2 were color-coded and plotted according to the groups of variants (Figure 3), which were determined by the software based on the reference set of strains. The distribution of Tms portrays the fact that even samples with convergent results may present slightly distinct Tms from the references. This suggests that Tm is not the only parameter to define the melting curve profile, and, ultimately, the species variants. The same conclusion applies for the occurrence of polymorphisms. Figure 4 and Figure 5 depict a set of nucleotide sequences of representative samples, including the references and strains with concordant and discordant HRM typing outcomes. The Tms and polymorphism within the amplified region are detailed for each sample.
Noteworthy, different combinations of various polimorphisms are observed among samples typed as the same variant, sharing similar Tm values and melting curve profile (e.g., Variant 1A). Conversely, the occurrence of very few polimorphims was associated with distinct Tms, melting curve profiles and divergent typing results, as observed for Primers 1 and 2 divergent IOCL 3310 (L. naiffi) strain (Figure 4 and Figure 5). For this sample, only one SNP T/A in Primer 1 region lead to Tm 85.7°C, representing 1.1°C variation from the reference strain L. naiffi (IOCL 1365; Tm 86.8°C). Distinct melting curve profiles were also observed for this sample (Figure 6). Primer 2 also revealed a divergent typing result for IOCL 3310 (L. naiffi) (Figure 5), despite the absence of polymorphism within the amplified region. The other two samples, both L. braziliensis strains, diverged in typing outcome, but only for the second set of primers (Figure 5 and Supp. Table S2). The polymorphisms observed for these samples (Figure 5) are the most likely cause for both the Tms and the melting curve profiles to diverge from the reference (Figure 6). These observations suggest that HRM divergent results cannot be directly and exclusively associated with polymorphisms and/or Tms, which although combined, potentially affect typing accuracy. Other undetectable factors may possibly be involved and would need further analysis.
2.3. HRM HSP70 presents high positivity and similar sensitivity and specificity to the previously proposed HSP70 qPCR protocol for parasite load determination.
To evaluate the accuracy of HRM HSP70 protocol as a diagnostic tool we assayed DNA obtained from clinical samples (n=60) collected from patients presenting different clinical forms of tegumentar leishmaniasis. Samples had been previously submitted to microscopy, HSP70 qPCR-based parasite load determination [20], conventional PCR (cPCR) and typing by RFLP to determine the infecting Leishmania species (Supp. Table S3). These previously published data allowed the current analytical assessment of HRM.
The HRM positivity percentage obtained for these samples was 76.6% (46/60), the same obtained for microscopy (46/60) and slightly lower than that of the HSP70 qPCR (47/60; 78.3%). Conventional PCR (HSP70 cPCR) presented the highest positivity rate (81.6 % - 49/60) for the same material. To determine the sensitivity and specificity, we plotted the data from the first set of HRM primers (P1) vs microscopy and vs cPCR as a gold standard (Table 3). Further, we compared these parameters with those from HSP70 qPCR. The HRM assay presented higher sensitivity (82.6%) and specificity (42.9%) than HSP70 qPCR (80.4% and 28.6%, respectively) when microscopy was chosen as the gold standard. By employing cPCR as the gold standard, HRM sensitivity and specificity increased to 85.7% and 63.6%, respectively. These results suggest that HRM represents a good diagnostic tool for Leishmania detection, independently of the species typing effectiveness.
2.4. HRM HSP70 as a species typing tool for clinical samples and the effect of parasite load on its accuracy.
We selected the cPCR positive samples successfully typed by RFLP and/or by DNA sequencing (45/60) to quantify the convergence with HRM typing results to evaluate its efficiency as a diagnostic-species tool. Among the samples, 5/45 revealed an inconclusive profile, by the generation of singular melt curves distinctive from all reference strains; 6/45 were negatives, i.e., not amplified by Primers 1 or 2. Therefore, HRM species typing was achieved for 34/45 samples, among which 26/34 (76.5%) converged with the RFLP species typing result and 8/34 (23.5%) diverged (Table 4). Kappa index >0 (K = 0.08) further confirmed the Moderate Agreement between RFLP data and HRM. This result on clinical samples is less accurate (76.5%) when compared to the successful outcome attained with DNA from cultured parasites (96.2 % concordance), an expected observation due to the nature of clinical samples of naturally infected individuals.
Considering the potential effect parasite load has on molecular typing outcomes, we plotted the distribution of normalized parasite load of the samples accordingly to the HRM-based species typing outcomes (Inconclusive, Divergent and Convergent). The result is represented in Figure 7. Most of the samples presenting inconclusive results by HRM (3/5) were likewise not quantifiable due to low parasite load, and therefore not represented in Figure 7. For the two remaining inconclusive samples, of which parasite load determination was possible, the median value of par.eq./μg human DNA was considerably higher (1.9 x 104 par. Eq./μg human DNA) than for the Divergent and Convergent groups, respectively (2.8 x 101 par. Eq./μg human DNA and 2.4 x 102 par. Eq./μg human DNA). Such association between typing success and Leishmania burden was also observed when Divergent and Convergent groups of samples were compared (Figure 7). We determine the quartiles and interquartile ranges to search for the parasite load interval that would provide better accuracy for HRM typing outcome. The interquartile range (25–75%) of parasite load for all assayed samples was determined as 5.9 x 101 - 6.0 x 102 (par. Eq./μg human DNA) (Figure 7, doted lines). The Convergent group interquartile range was narrowed within 1.1 x 102 – 5.7 x 102(par. Eq./μg human DNA), while for the Divergent group the range was widely dispersed outside this range (4.7 x 10-1 – 3.3 x 103 par. Eq./μg human DNA). Within the interquartile range (Figure 7, Quartile 2), 15/17 samples (88.2%) revealed an HRM convergent typing result, while for the first (Q1) and third (Q3) quartiles the accuracy was 5/8 (62.5 %) and 6/8 (75%) samples, respectively. The data reveal the parasite load interval with the best accuracy for the HRM typing scheme.
3. Discussion
Our group has been interested in developing and validating methodologies to detect, quantify and type Leishmania species, both as a laboratory/biobank protocol for the reference service and as a diagnostic tool to be applied in clinical material. The importance of detecting and typing Leishmania species is widely recognized, as well as the determination of parasite load for clinical and research purposes [20,21]. Therefore, combined molecular approaches merging these possibilities would be valuable tools for clinicians, surveillance, and research.
CLIOC, a Leishmania biobank, has been currently evaluating techniques with the potential to replace the MLEE, aiming to overcome the drawbacks of the technique such as its elevated cost, limited set of assayed samples per experiment, difficulty of inter-laboratory comparison, and time-consuming characteristics. High Resolution Melting analysis is simple and rapid, and its use in clinical or research samples offers many advantages, such as a lower total cost for species identification compared to MLEE. Moreover, as a real time PCR-based approach, there is no need for sequencing or gel electrophoresis to analyze the product, reducing the process and avoiding laboratory contamination with PCR products. It also reduces the need for trained personnel to analyze an electrophoretic gel or sequencing data to provide a result, and, additionally, presents the possibility of quantifying infecting parasites within samples since it can be applied as a quantitative PCR-based technique. The whole process can be automated as the analyzer software produces the result by comparing the tested sample and the reference sample profiles, which must always be included in the reactions [22]. Based on such, we consulted the promising results from Zampieri et. al., [18] and proposed to further evaluate the accuracy of the protocol as a typing and/or diagnostic tool.
First, we tested the reproducibility of the assay proposed by Zampieri and colleagues in our lab conditions, which include different operators and equipment, among other variables. The results of a reference set of samples fully reproduced the published data, by differentiating the main species with the first set of primers (P1), except for L. (L.) infantum vs L. (L.) donovani, and L. (L.) amazonensis vs L. (L.) mexicana. We included species not yet evaluated bonafide by the method, such as L. (V.) peruviana, L. (V.) panamensis, L. (V.) lindenbergi, L. (V.) utingensis, L. (P.) hertigi and the Paraleishmania colombiensis and P. equatorensis. The results precisely allowed the distinction of the main L. (Viannia) species, additionally revealing the profiles of the above-mentioned newly assayed species (Figure 1). By applying the second set of primers (variant 2) L. (V.) guyanensis and L. (V.) braziliensis were effectively differentiated, but L. (V.) peruviana vs L. (V.) braziliensis and L. (V.) panamensis vs L. (V.) guyanensis remained within the same variant and were, thus, not possible to distinguish. The separation of these species are specially relevant in sympatric regions such as Andean region in Peru [23,24], Colombia [25,26], Venezuela [27] and Bolivia [28]. Further steps with an additional primer design must be developed to specifically attend the purpose of typing these species.
We performed a reproducible assay with the same operator, executing experiments in distinct days and obtaining 100% reproducibility. The next necessary step was to test the efficacy of the protocol considering the intraspecific genetic variability - known to occur among Leishmania strains. Such assay allows to evaluate the effect of unanticipated DNA polymorphism on factors that affect the HRM typing outcome, such as mismatches at the primer alignment site, the melting curve and ultimately the melt temperature [22]. Therefore, a wider panel of samples was prepared, composed by DNA of 90 strains from widely different geographic origins representing 19 Leishmania species. Results converged with the MLEE and HSP70 sequencing data for 96.2% of samples, revealing a highly accurate outcome. Kappa index indicated Almost perfect agreement between HRM and MLEE. Results revealed the potential for the proposed method and protocol to be applied as a typing tool on DNA from isolated and cultured Leishmania, eventually substituting the current MLEE assay.
Raw data from the assays of the three divergent strains are a valuable opportunity to explore the intrinsic limiting features of HRM approach. The melting temperature is one of the parameters for the software to cluster samples within the window set defined by the reference strains profiles, though it is not the only metric. To express this subject, we plotted the Tm distribution with color-coded samples’ variants for both P1 and P2primers. The result revealed Tm as not being the only parameter to define the melting curve profile which ultimately defines the variants. Figure 3 summarizes the limited role Tm has on the clustering of samples as divergent and convergent. It has been reported that DNA methylation, concentration of the initial template for reaction – reflected by the Ct obtained, GC content within the amplified region and stoic metrics may influence the melt curve profile. Moreover, type and quality of the DNA source material (purity and integrity), isolation method and pipetting inconsistencies, might also influence outcome [22]. We managed to carefully avoid a few of these variables to deliberately focus on the effect of polymorphisms on melt profiles. Thus, DNAs were prepared by the same isolation method within at most six months interval; all assays were performed by the same experienced operator in the same equipment. Concentration of initial template for the reactions was not normalized, although parasites’ pellets for DNA isolation were prepared by the same laboratory protocol. Therefore, the Ct values varied slightly, mostly within the described 20-30 range presented as of optimal efficiency [22]. Indeed, the different quantities of initial DNA template did not compromise the accuracy, whereas sequencing of the amplified region of the divergent strains revealed polymorphisms as an important, but not unique, source of divergence between typing data. These samples presented Tm close to the upper-range of the correspondent references (Figure 3), but the G-C content and the polymorphisms position [22] were possibly a main-factor that produced distinct melt curve profiles, ultimately leading to the divergent result (Figure 4 and Figure 5). Shape of melting curves may also be affected by methylation and efficiency in primer alignment, independently of polymorphisms [22]. However, for the current data the polymorphisms and their location within the amplified region are the more parsimonious explanation for typing divergence. The intra and interspecific genetic diversity is a well-known trait of Leishmania and indeed a challenge for species typing and for diagnostic tools development. There are geographic regions of Brazil where hosts and circulating parasites present greater chances of variability. Indeed, this was the case for the divergent samples from the validation step. These strains were isolated from dogs and from a human host, from the northern region that harbors a highly diverse parasite population [29,30].
Our group recently proposed an HSP70 qPCR-based approach to determine parasite load in material collected from patients with confirmed clinical diagnostic for cutaneous leishmaniasis. The patients studied were from an endemic area in the Amazon region, in which parasite population is quite diverse [20]. Besides the HSP70 qPCR, the samples were also subjected to microscopy and conventional PCR; infecting species was identified by PCR-RFLP and/or by DNA sequencing. Herein, we applied the HSP70 HRM protocol on these samples to test the method’s accuracy as a diagnostic typing tool. Data revealed that HRM positivity percentage was the same obtained for microscopy and slightly lower than that of the HSP70 qPCR. Conventional PCR (cPCR) presented the highest positivity. To define sensitivity and specificity, microscopy and cPCR were used as gold standards. The HRM sensitivity and specificity were higher than that obtained by the HSP70 qPCR applied in these samples. By employing cPCR as the gold standard, HRM sensitivity and specificity increased significantly. These results suggest HRM represents a good diagnostic tool for Leishmania detection, independently of the species typing effectiveness.
In contrast to the Kappa index compatible to “Almost perfect agreement” and 96.2% concordance for the validation step, in which DNA from cultured parasites was used, for the clinical samples a “Moderate Agreement” was achieved, and convergent typing results were attained for 76.5% of samples. The effective outcome obtained by HRM with DNA from cultured parasites is expected due to the specific nature and purity of the samples. Even though predictable, we further explored the features underlining the reduction on HRM typing success in clinical material. Coinfections, for instance, could lead to inconclusive typing result, and such possibility cannot be excluded, given the geographic region where the patients were located (sympatric area for most Leishmania species). These conditions, however, are difficult to detect and characterize. The most likely - and testable – factor involved is the parasite load. As described in [20], the clinical samples assayed presented a wide range of parasites equivalents per human DNA, thus comprising a suitable panel to be tested by HRM (Figure 7). Therefore, to evaluate the effect of parasite load on typing accuracy, we plotted parasites equivalents per human DNA accordingly to HRM-based species typing outcome and defined the interquartile range of all samples. The result exposed the variable accuracy in each quartile, with the best outcome of 88.2% attained in a narrow range of parasite load. Therefore, HRM typing outcome was highly affected by parasite load, despite its high positivity for Leishmania DNA. Based on this conclusion, we would suggest a clinical-sample-typing workflow that first employs the HSP70 qPCR protocol [20] to evaluate whether the sample fits the most accurate parasite load range for HRM typing. The decision to further apply the HSP70-based HRM protocol as a typing tool must be taken considering this information.
There are recent studies in the literature addressing the use of HRM to identify Leishmania species [16,31,32]. None, however, have tested the protocol accuracy in a significantly bonafide strains panel representing the inter and intraspecific diversity of Leishmania. Important steps like this are vital to propose the technique as a typing tool. Moreover, the published studies avoid important validation steps by directly testing HRM-based protocols in biological material, not exploring the many factors that may influence the outcome. Herein, we reveal the great potential of the HSP70-HRM for species typing in DNA from cultured parasites. For clinical samples, however, the data raise the need for caution when using the HSP70-HRM protocol as a species typing tool. The variable accuracy dependent on parasite load imposes the need to first either determine the parasite load or, at least, establish a Ct-based cutoff value, before deciding for the typing approach.
The results presented herein open a venue of possibilities for evaluation of the HRM approach targeting other genes which might achieve better results related to Leishmania species identification, especially those targets with higher copy number than the HSP70 gene. Among different genes employed for Leishmania species typing, at least for neotropical regions, MPI [30] and Cytb [33] would be good alternatives to be tested.
4. Materials and Methods
Strains and DNA samples
All Leishmania strains were obtained from the Leishmania Collection of Oswaldo Cruz Foundation (CLIOC - http://clioc.fiocruz.br). Parasite promastigotes were grown at a biphasic medium, constituted in the liquid phase by Schneider medium (Sigma, Chemical Co., St. Louis, USA) supplemented with 20% inactivated Fetal Bovine Serum - FBS (Vitrocell, Campinas, SP, Brazil), and in the solid phase by 15% of rabbit blood and BHI-Agar (Sigma) [Novy-Nicolle-Mc Neal medium - NNN]. Strains were kept at BOD incubator at 25ºC. The cell viability was evaluated by microscopy. Sixteen Leishmania reference strains representing 14 distinct species (Supp. Table S1), were used for the standardization of HRM reactions. Additional strains representing the inter and intraspecific genetic diversity of Leishmania circulating in the Americas were selected, including hybrids detected by MLEE (Supp. Material Table S2) and the following species not previously examined: L. (V.) panamensis, L. (V.) utingensis, L. (V.) peruviana, L. (V.) lindenbergi, L. (Porcisia) hertigi, Paraleishmania colombiensis and P. equatorensis. In total, 110 strains representing 18 species were assayed. The DNA was extracted from cultures using the Wizard Genomic DNA purification kit (Promega, Madison, Wisconsin, EUA), according to the manufacturer's recommendations. The DNAs were quantified in NanoDrop® at 260 nm (Thermo Fisher Scientific, Waltham, Massachusetts, USA) and diluted in water to a concentration of 50ng/µL. The DNA purity was estimated by the ratio 260/280 nm.
Species typing
All samples from CLIOC are typed by Multilocus Enzyme Electrophoresis (MLEE) following the internal Standard Operational Procedures (SOPs). Additionally, for the present work, the HSP70 PCR products of the selected strains were obtained and sequenced, as previously reported, [9] as an additional method to identify the species and reveal the polymorphisms within the region analyzed by HRM. The DNA sequencing of HSP70 genomic region was performed by Fiocruz facilities. Briefly, PCR products were purified with the Wizard SV Clean-up System (Promega). The final DNA concentration was estimated by comparison with a DNA Ladder Marker (Promega) in 1,5% agarose gel. Sequencing was performed with the same primers used for amplification, using BigDyeTerminator v3.1 Cycle Sequencing Kit, and the products were analyzed in an automated DNA sequencer (ABI PRISM –3730 - Thermo Fisher Scientific). Consensus sequences were generated from forward and reverse strands using the Phred-Phrap-Consed package [20]. Only sequences with Phred values above 20 were used for contig construction.
HRM reactions and reproducibility assay
HRM reactions were designed to reproduce the protocol described by Zampieri et. al. 2016, [17] due to the reported effectiveness of the method. A few technical variations, however, were included to attend the laboratory routine. Briefly, the equipment used was the ViiA 7 Real-Time PCR System (Thermo Fisher Scientific) featured in 384-wells plataform; MeltDoctor™ HRM Master Mix (Thermo Fisher Scientific) was used in a 1x concentration per reaction, in 10 µL total reaction volume. Primer’s concentration was established as 200nM after standardization steps. All experiments were performed twice, in technical duplicates. Dissociation analysis is done by capturing fluorescence signals in 0.2°C intervals with the High-Resolution Melting Software. Protocols (primer 1 and primer 2, Table 1) were replicated for a larger sample panel of 110 strains, depicted above. The DNA of 16 reference strains (Supp. Material Table S1) was assayed to first confirm the findings from the published study. To keep a technical or experimental replicate within the analysis, the Silhouette Score was considered. For each melt curve, a modified silhouette score is presented, ranging between 0 to 100. A score closer to 100 indicates that a melt curve is more similar to curves assigned in the same variant call than to curves called differently. For the reproducibility assay, 13 representative samples from the main panel were selected and submitted twice to the protocol in experimental duplicates, by the same operator in two different days (depicted at Supp. Material Table S2).
Clinical samples
A panel of 60 DNAs from clinical samples was selected for the assays (Supp. Table S3). The ethical recommendations of the Brazilian National Council of Health were adhered to, wherein a research protocol was duly registered and approved under the Certificate of Presentation for Ethics Appreciation code CAAE No. 0020.0.046.000–11 by the Ethical and Research Committee of the Center of Research in Tropical Medicine. The clinical material was collected by two sterile cervical brushes, used to scrap the TL patients’ lesions, and included positive and negative samples by microscopy and/or conventional PCR (cPCR) (Supp. Table S3). Samples were stored in RNALater solution (Ambion®, Carlsbad, CA/USA) and kept at the Laboratory of Genetics Epidemiology from Fiocruz Rondônia (Fiocruz/RO), Brazil. The DNA was extracted using the PureLink DNA MiniKit (Invitrogen®, Carlsbad, CA/USA), according to manufacturer’s instructions, excluding the incubation at 55ºC step. Identification of the species involved in the infections was performed by RFLP for the HSP70c target [18], using HaeIII, BstuI and MboI enzimes [34]. As expected, most of them was identified as L. (V.) braziliensis, once this is the main species causing TL in that region. All clinical samples assays were performed blindly. The HRM protocol applied on these clinical DNAs was the same as previously described.
Analysis
Data of parasite load and species typing of clinical samples were obtained from our previous study [19]. Interquartile ranges of parasite load, Kappa index, sensitivity and specificity were determined by GraphPad Prism v.9 (GraphPad Software, Inc., San Diego, CA). Figures were designed in GraphPad Prism v.9 (GraphPad Software, Inc., San Diego, CA) and Biorender (https://www.biorender.com).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Reference strains assayed to reproduce the HSP70 HRM protocol.; Table S2: Strains selected to represent intra and intersepecific variation within Leishmania genus. The samples were donated by CLIOC and represent DNA from cultured parasites.; Table S3: Clinical samples assayed for the HRM validation step. Confirmation of Publication and Licensing Rights from Biorender for submitting to publication Figure 1. BLAST results obtained in August 2023 with the HSP70 nucleotide sequence data for the three divergent samples from the validation step.
Author Contributions
Conceptualization. M.C.B., C.B. and E.C.; methodology. O.C.M., M.C.B., D.P.P. and C.P.B.F.; validation. C.P.B.F., G.E.M.F, S.R., E.C., O.C.M. and L.M.C.; formal analysis. M.C.B.; investigation. C.P.B.F., S.R. and L.M.C.; resources. E.C. and C.B.; data curation. C.P.B.F., E.C., L.M.C., G.E.M.F. and S.R.; writing—original draft preparation. C.P.B.F. and M.C.B.; writing—review and editing. D.P.P, E.C, L.M.C., G.E.M.F., S.R., C.B., O.C.M., C.P.B.F, M.C.B.; visualization. L.M.C and M.C.B.; supervision. D.P.P., O.C.M. and M.C.B.; project administration. M.C.B.; funding acquisition. E.C., C.B.
Funding
This research was funded by (C.B.) Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro/FAPERJ (grant E-26/201.213/2022) and Conselho Nacional de Desenvolvimento Científico e Tecnológico/CNPq (grant 304308/2019-3). (E.C.) Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Finance Code 001; Conselho Nacional de Desenvolvimento Científico e Tecnológico, Research Fellow, 302622/2017-9; Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro, CNE, E26-202.569/2019; ColBio2020, E26-210.285/2021; Fundação de Amparo ao Desenvolvimento das Ações Científicas e Tecnológicas e à Pesquisa-FAPERO, Edital 16/2014. (L.M.C.) Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro, Pós-Doutorado Nota 10 E-26/205.730/2022 and 205.731/2022. (O.C.M.) research fellow of CNPq1D and FAPERJ (CNE).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data supporting reported results on the Leishmania samples available at CLIOC can be found at https://clioc.fiocruz.br/.
Acknowledgments
We thank the Rede de Plataformas Tecnológicas Fiocruz (RPT) for performing all the sequencing (RPT01A) and HRM reactions (RPT09A); Coleção de Leishmania da Fiocruz (CLIOC) for providing the strains; Caroline Batista Marques do Souza and Barbara Santos for laboratorial support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ross, R. FURTHER NOTES ON LEISHMAN’S BODIES. Br Med J 1903, 2, 1401. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.S.; Lockwood, D.N.J. Cutaneous leishmaniasis. Clinics in Dermatology 2007, 25, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.; Schubach, A.O.; Madeira, M.F.; Leal, C.A.; Pires, M.Q.; Oliveira, F.S.; et al. Leishmania (Viannia) braziliensis genotypes identified in lesions of patients with atypical or typical manifestations of tegumentary leishmaniasis: Evaluation by two molecular markers. Experimental Parasitology 2009, 121, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R. The Neotropical Leishmania species: a brief historical review of their discovery, ecology and taxonomy. Revista Pan-Amazônica de Saúde 2010, 1. [Google Scholar] [CrossRef]
- de Almeida, J.V.; de Souza, C.F.; Fuzari, A.A.; Joya, C.A.; Valdivia, H.O.; Bartholomeu, D.C.; et al. Diagnosis and identification of Leishmania species in patients with cutaneous leishmaniasis in the state of Roraima, Brazil’s Amazon Region. Parasit Vectors 2021, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization. Manual of procedures for leishmaniases surveillance and control in the Americas. 2019. Available online: https://iris.paho.org/handle/10665.2/51838.
- Cupolillo, E.; Momen, H.; Grimaldi, G. Genetic diversity in natural populations of New World Leishmania. Mem Inst Oswaldo Cruz 1998, 93, 663–668. [Google Scholar] [CrossRef]
- Brito, M.E.F.; Andrade, M.S.; Mendonça, M.G.; Silva, C.J.; Almeida, E.L.; Lima, B.S.; et al. Species diversity of Leishmania (Viannia) parasites circulating in an endemic area for cutaneous leishmaniasis located in the Atlantic rainforest region of northeastern Brazil. Trop Med Int Health 2009, 14, 1278–1286. [Google Scholar] [CrossRef]
- Romero G a, S.; Ishikawa, E.; Cupolillo, E.; Toaldo, C.B.; Guerra, M.V.F.; Paes, M.G.; et al. The rarity of infection with Leishmania (Viannia) braziliensis among patients from the Manaus region of Amazonas state, Brazil, who have cutaneous leishmaniasis. Ann Trop Med Parasitol 2002, 96, 131–136. [Google Scholar] [CrossRef]
- da Silva, L.A.; de Sousa, C.D.S.; da Graça, G.C.; Porrozzi, R.; Cupolillo, E. Sequence analysis and PCR-RFLP profiling of the hsp70 gene as a valuable tool for identifying Leishmania species associated with human leishmaniasis in Brazil. Infect Genet Evol 2010, 10, 77–83. [Google Scholar] [CrossRef]
- da Graça, G.C.; Volpini, A.C.; Romero, G.A.S.; Oliveira Neto MP de Hueb, M.; Porrozzi, R.; et al. Development and validation of PCR-based assays for diagnosis of American cutaneous leishmaniasis and identificatio nof the parasite species. Mem Inst Oswaldo Cruz 2012, 107, 664–674. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clinical Chemistry 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; et al. The real-time polymerase chain reaction. Mol Aspects Med 2006, 27, 95–125. [Google Scholar] [CrossRef] [PubMed]
- Talmi-Frank, D.; Nasereddin, A.; Schnur, L.F.; Schönian, G.; Töz, S.Ö.; Jaffe, C.L.; et al. Detection and Identification of Old World Leishmania by High Resolution Melt Analysis. PLoS Negl Trop Dis 2010, 4, e581. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Galluzzi, L.; Migliazzo, A.; Magnani, M. Detection and Characterization of Leishmania (Leishmania) and Leishmania (Viannia) by SYBR Green-Based Real-Time PCR and High Resolution Melt Analysis Targeting Kinetoplast Minicircle DNA. PLOS ONE 2014, 9, e88845. [Google Scholar] [CrossRef] [PubMed]
- Hernández, C.; Alvarez, C.; González, C.; Ayala, M.S.; León, C.M.; Ramírez, J.D. Identification of Six New World Leishmania species through the implementation of a High-Resolution Melting (HRM) genotyping assay. Parasites & Vectors 2014, 7, 501. [Google Scholar]
- Reed, G.H.; Kent, J.O.; Wittwer, C.T. High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 2007, 8, 597–608. [Google Scholar] [CrossRef]
- Zampieri, R.A.; Laranjeira-Silva, M.F.; Muxel, S.M.; Stocco de Lima, A.C.; Shaw, J.J.; Floeter-Winter, L.M. High Resolution Melting Analysis Targeting hsp70 as a Fast and Efficient Method for the Discrimination of Leishmania Species. PLoS Negl Trop Dis 2016, 10, e0004485. [Google Scholar] [CrossRef]
- Zampieri, R.A.; Laranjeira-Silva, M.F.; Muxel, S.M.; Lima ACS de Shaw, J.J.; Floeter-Winter, L.M. High Resolution Melting Analysis Targeting hsp70 as a Fast and Efficient Method for the Discrimination of Leishmania Species. PLOS Neglected Tropical Diseases 2016, 10, e0004485. [Google Scholar] [CrossRef]
- Filgueira, C.P.B.; Moreira, O.C.; Cantanhêde, L.M.; Farias HMT de Porrozzi, R.; Britto, C.; et al. Comparison and clinical validation of qPCR assays targeting Leishmania 18S rDNA and HSP70 genes in patients with American Tegumentary Leishmaniasis. PLOS Neglected Tropical Diseases 2020, 14, e0008750. [Google Scholar] [CrossRef]
- Moreira, O.C.; Yadon, Z.E.; Cupolillo, E. The applicability of real-time PCR in the diagnostic of cutaneous leishmaniasis and parasite quantification for clinical management: Current status and perspectives. Acta Tropica 2018, 184, 29–37. [Google Scholar] [CrossRef]
- Słomka, M.; Sobalska-Kwapis, M.; Wachulec, M.; Bartosz, G.; Strapagiel, D. High Resolution Melting (HRM) for High-Throughput Genotyping—Limitations and Caveats in Practical Case Studies. International Journal of Molecular Sciences 2017, 18, 2316. [Google Scholar] [CrossRef] [PubMed]
- Cortes, S.; Esteves, C.; Maurício, I.; Maia, C.; Cristovão, J.M.; Miles, M.; et al. In vitro and in vivo behaviour of sympatric Leishmania (V.) braziliensis, L. (V.) peruviana and their hybrids. Parasitology 2012, 139, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Seki, C.; Kubo, M.; Gonzales-Cornejo, L.; Caceres, A.G. Natural infections of Pintomyia verrucarum and Pintomyia maranonensis by Leishmania (Viannia) peruviana in the Eastern Andes of northern Peru. PLoS Negl Trop Dis 2021, 15, e0009352. [Google Scholar] [CrossRef]
- León, C.M.; Muñoz, M.; Hernández, C.; Ayala, M.S.; Flórez, C.; Teherán, A.; et al. Analytical Performance of Four Polymerase Chain Reaction (PCR) and Real Time PCR (qPCR) Assays for the Detection of Six Leishmania Species DNA in Colombia. Front Microbiol 2017, 8, 1907. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Chilama, M.; YOviedo, M.; KQuintero, Y.; LFernández, O.; Gómez, M.A. Leishmania RNA Virus Is Not Detected in All Species of the Leishmania Viannia Subgenus: The Case of L. (V.) panamensis in Colombia. Am J Trop Med Hyg 2023, 108, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Noguera, L.A.; Hernández-Pereira, C.E.; Castillo-Castañeda, A.C.; Patiño, L.H.; Castañeda, S.; Herrera, G.; et al. Diversity and geographical distribution of Leishmania species and the emergence of Leishmania (Leishmania) infantum and L. (Viannia) panamensis in Central-Western Venezuela. Acta Trop 2023, 242, 106901. [Google Scholar] [CrossRef]
- Torrico, M.C.; Fernández-Arévalo, A.; Ballart, C.; Solano, M.; Rojas, E.; Ariza, E.; et al. Tegumentary leishmaniasis by Leishmania braziliensis complex in Cochabamba, Bolivia including the presence of L. braziliensis outlier: Tegumentary leishmaniasis in Cochabamba, Bolivia: Tegumentary leishmaniasis in Cochabamba, Bolivia. Transbound Emerg Dis 2022, 69, 2242–2255. [Google Scholar] [CrossRef]
- Kuhls, K.; Cupolillo, E.; Silva, S.O.; Schweynoch, C.; Boité, M.C.; Mello, M.N.; et al. Population structure and evidence for both clonality and recombination among Brazilian strains of the subgenus Leishmania (Viannia). PLoS Negl Trop Dis 2013, 7, e2490. [Google Scholar] [CrossRef]
- Boité, M.C.; Mauricio, I.L.; Miles, M.A.; Cupolillo, E. New insights on taxonomy, phylogeny and population genetics of Leishmania (Viannia) parasites based on multilocus sequence analysis. PLoS Negl Trop Dis 2012, 6, e1888. [Google Scholar] [CrossRef]
- Garay, A.F.G.; Fraenkel, S.; Diaz, J.J.A.R.; Recalde, O.D.S.; Gómez, M.C.V.; Riquelme, J.A.M.; et al. Sensitivity comparison for the Leishmania spp. detection in different canine tissues using PCR-HRM. Rev Soc Bras Med Trop 2022, 55, e0069-2022. [Google Scholar] [CrossRef]
- Alaeenovin, E.; Parvizi, P.; Ghafari, S.M. Two Leishmania species separation targeting the ITS-rDNA and Cyt b genes by developing and evaluating HRM- qPCR. Rev Soc Bras Med Trop 2022, 55, e0186-2022. [Google Scholar] [CrossRef] [PubMed]
- Davila, M.; Pineda, V.; Calzada, J.E.; Saldaña, A.; Samudio, F. Evaluation of cytochrome b sequence to identify Leishmania species and variants: the case of Panama. Mem Inst Oswaldo Cruz 2021, 116, e200572. [Google Scholar] [CrossRef] [PubMed]
- Espada, C.R.; Ortiz, P.A.; Shaw, J.J.; Barral, A.M.P.; Costa, J.M.L.; Uliana, S.R.B.; et al. Identification of Leishmania (Viannia) species and clinical isolates of Leishmania (Leishmania) amazonensis from Brazil using PCR-RFLP of the heat-shock protein 70 gene reveals some unexpected observations. Diagn Microbiol Infect Dis 2018, 91, 312–318. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the species groups coupled in each variant obtained by PCR reactions with primers 1 and 2; suggestion of the algorithm to differentiate the main species circulating in the Americas.
Figure 1.
Schematic representation of the species groups coupled in each variant obtained by PCR reactions with primers 1 and 2; suggestion of the algorithm to differentiate the main species circulating in the Americas.
Figure 2.
Color-coded dissociation curves of reference strains selected as standards for primer 1 (a) and 2 (b). (a) The identified variants for primer 1: blue - variant 1A; green - variant 1B; red - variant 1C; orange - variant 1D; yellow - variant 1E. (b) The identified variants for Primer 2: Curves in grey - variant 2A; Curves in brown - variant 2B. Figures were exported from the High-Resolution Melting Software available at the ViiA 7 Real-Time PCR System ( Thermo Fisher Scientific).
Figure 2.
Color-coded dissociation curves of reference strains selected as standards for primer 1 (a) and 2 (b). (a) The identified variants for primer 1: blue - variant 1A; green - variant 1B; red - variant 1C; orange - variant 1D; yellow - variant 1E. (b) The identified variants for Primer 2: Curves in grey - variant 2A; Curves in brown - variant 2B. Figures were exported from the High-Resolution Melting Software available at the ViiA 7 Real-Time PCR System ( Thermo Fisher Scientific).
Figure 3.
Color-coded Melting Temperatures (Tms) accordingly to Primers 1 and 2 variants. Tms obtained during the validation assay. (a) blue circle = 1A, brown square = 1B, orange inverted triangle = 1C, green = 1D, pink diamond = 1E. (b) black circle = 2A, blue diamond = 2B. The three divergent results are represented in red: (a) red circle: IOCL 3310, L. naiffi clustered as 1A (expected to be 1E); (b) red circles correspond to the two L. braziliensis and the one L. naiffi erroneously clustered as variant 2A (expected 2B).
Figure 3.
Color-coded Melting Temperatures (Tms) accordingly to Primers 1 and 2 variants. Tms obtained during the validation assay. (a) blue circle = 1A, brown square = 1B, orange inverted triangle = 1C, green = 1D, pink diamond = 1E. (b) black circle = 2A, blue diamond = 2B. The three divergent results are represented in red: (a) red circle: IOCL 3310, L. naiffi clustered as 1A (expected to be 1E); (b) red circles correspond to the two L. braziliensis and the one L. naiffi erroneously clustered as variant 2A (expected 2B).
Figure 4.
Examples of the variants obtained by Primer 1 for different strains and species, the polymorphisms observed within the HRM analyzed region and the melting temperatures (Tm). The divergent strain IOCL 3310 is depicted in bold, marked*.
Figure 4.
Examples of the variants obtained by Primer 1 for different strains and species, the polymorphisms observed within the HRM analyzed region and the melting temperatures (Tm). The divergent strain IOCL 3310 is depicted in bold, marked*.

Figure 5.
Examples of the variants obtained by Primer 2 for different strains and species, the polymorphisms observed within the HRM analyzed region and the melting temperatures (Tm). The divergent strains IOCL 161, 167 and 3310 are depicted in bold, marked *.
Figure 5.
Examples of the variants obtained by Primer 2 for different strains and species, the polymorphisms observed within the HRM analyzed region and the melting temperatures (Tm). The divergent strains IOCL 161, 167 and 3310 are depicted in bold, marked *.
Figure 6.
(a) Aligned melt curve profiles for the divergent strain IOCL 3310 (L. naiffi) (yellow), compared to the reference strain IOCL 1365 (L. naiffi) named by the software as a new variant (brown). (b) Aligned melt curve profiles and amplification plot for the L. braziliensis strains IOCL 161 and IOCL 167 obtained from Primer 2. Melt curve for both strains are represented in grey and were assigned as a different variant from the reference strain in red. (c) and (d) Amplification plot demonstrates similar amplification efficiencies for samples and the reference, for Primer 1 and 2. Figures exported from the High-Resolution Melting Software available at the ViiA 7 Real-Time PCR System (Thermo Fisher Scientific).
Figure 6.
(a) Aligned melt curve profiles for the divergent strain IOCL 3310 (L. naiffi) (yellow), compared to the reference strain IOCL 1365 (L. naiffi) named by the software as a new variant (brown). (b) Aligned melt curve profiles and amplification plot for the L. braziliensis strains IOCL 161 and IOCL 167 obtained from Primer 2. Melt curve for both strains are represented in grey and were assigned as a different variant from the reference strain in red. (c) and (d) Amplification plot demonstrates similar amplification efficiencies for samples and the reference, for Primer 1 and 2. Figures exported from the High-Resolution Melting Software available at the ViiA 7 Real-Time PCR System (Thermo Fisher Scientific).
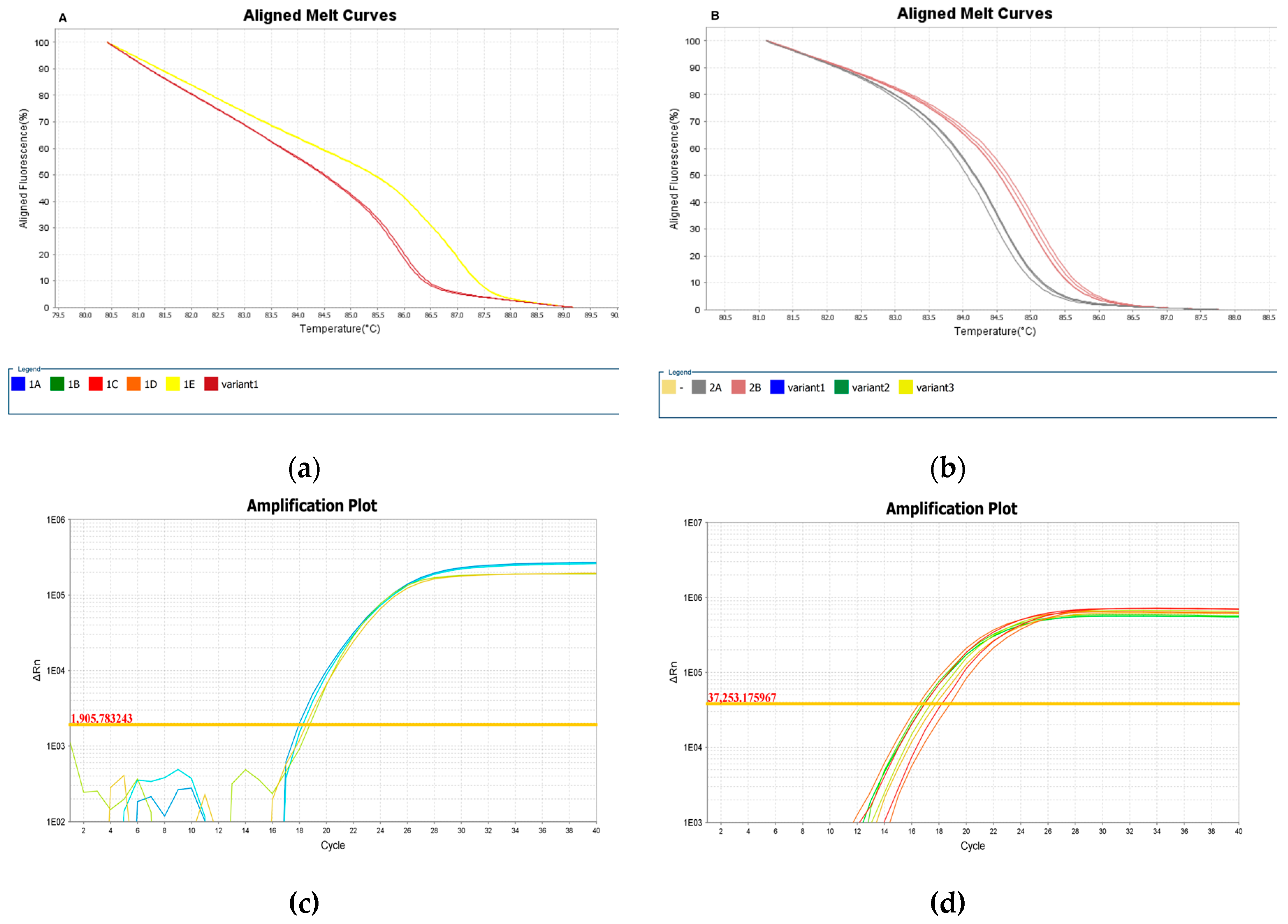
Figure 7.
Normalized parasite load distribution of the samples assayed plotted accordingly to species typing outcome. Convergent = HRM and RFLP convergent typing results. Divergent = HRM species typing was possible but did not match with previous RFLP characterization. Inconclusive = singular melting profile not correspondent to any of the reference strains. Doted lines mark the interquartile range of the total assayed samples separating quartiles Q1 (bottom) Q2 (in between) and Q3 (upper area).
Figure 7.
Normalized parasite load distribution of the samples assayed plotted accordingly to species typing outcome. Convergent = HRM and RFLP convergent typing results. Divergent = HRM species typing was possible but did not match with previous RFLP characterization. Inconclusive = singular melting profile not correspondent to any of the reference strains. Doted lines mark the interquartile range of the total assayed samples separating quartiles Q1 (bottom) Q2 (in between) and Q3 (upper area).
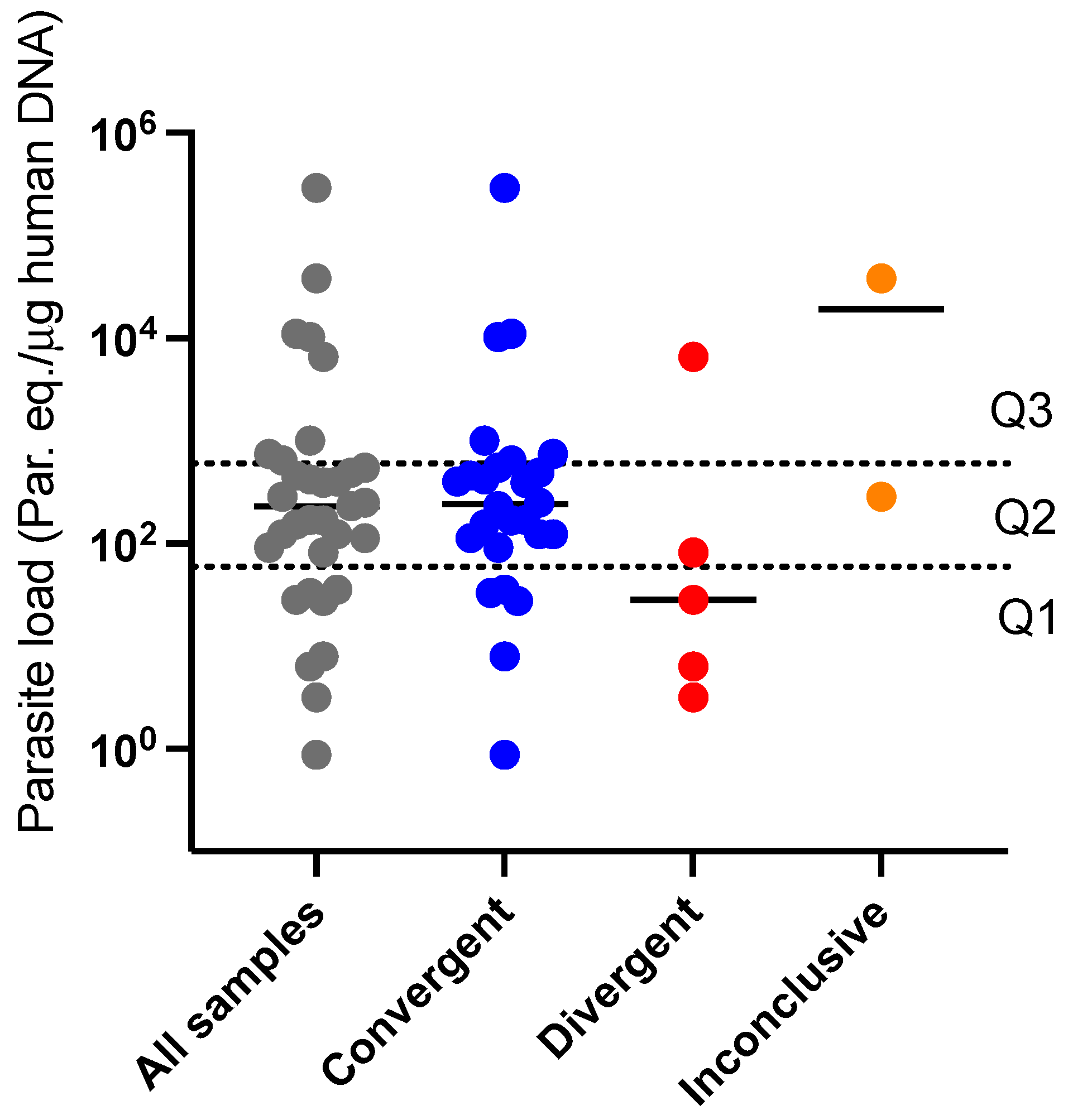

Table 1.
Reference strains, typing results during experiments 1 and 2 for Primer 1 and Primer 2; median of melting temperatures in °C (Tm) of replicates and standard variation (SD) in °C.
Table 1.
Reference strains, typing results during experiments 1 and 2 for Primer 1 and Primer 2; median of melting temperatures in °C (Tm) of replicates and standard variation (SD) in °C.
| Voucher | Species | Primer 1 | Primer 2 | Primer 1 | Primer 2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Exp 1 | Exp.2 | Exp 1 | Exp.2 | Tm | SD | Tm | SD | ||
| IOCL 565 | L. (V.) guyanensis | 1D | 1D | 2A | 2A | 86.1 | 0.2 | 84.1 | 0.0 |
| IOCL 566 | L. (V.) braziliensis | 1D | 1D | 2B | 2B | 85.8 | 0.1 | 84.7 | 0.1 |
| IOCL 575 | L. (L.) amazonensis | 1A | 1A | - | - | 85.2 | 0.1 | 83.9 | 0.2 |
| IOCL 579 | L. (L.) infantum | 1A | 1A | - | - | 85.0 | 0.1 | 85.1 | 0.0 |
| IOCL 1023 | L. (V.) lainsoni | 1A | 1A | 2A | 2A | 85.4 | 0.1 | 84.2 | 0.1 |
| IOCL 1365 | L. (V.) naiffi | 1E | 1E | 2B | 2B | 85.4 | 0.1 | 84.7 | 0.2 |
| IOCL 1545 | L. (V.) shawi | 1E | 1E | 2A | 2A | 86.7 | 0.1 | 84.0 | 0.1 |
| IOCL 2689 | L. (V.) utingensis | 1E | 1E | 2B | 2B | 87.0 | 0.1 | 84.8 | 0.2 |
| IOCL 2690 | L. (V.) lindenbergi | 1D | 1D | 2A | 2A | 85.9 | 0.1 | 84.1 | 0.1 |
| IOCL 3394 | L. (V.) braziliensis | 1D | 1D | 2B | 2B | 86.1 | 0.1 | 84.6 | 0.0 |
| IOCL 3398 | L. (V.) lainsoni | 1D | 1D | 2A | 2A | 85.7 | 0.2 | 84.1 | 0.0 |
| IOCL 3399 | L. (L.) amazonensis | 1A | 1A | - | - | 85.2 | 0.3 | 84.6 | 0.0 |
| IOCL 3538 | L. (V.) guyanensis | 1D | 1D | 2A | 2A | 86.2 | 0.1 | 84.2 | 0.0 |
1D: variant 1D; 1A: variant 1A; 1E: variant 1E; 2A: variant 2A and 2B: variant 2B.Exp. 1 and Exp. 2 refer to the experimental replicates 1 and 2 performed by the same operator in different days. .
Table 2.
Samples with hybrid profile by MLEE and respective variants and Tm obtained by HRM.
| V 1 | V 2 | Voucher (IOCL) | MLEE characterization | DNA sequencing | HRM | Tm V1 | Tm V2 | |
|---|---|---|---|---|---|---|---|---|
| 1D | 2A | 3358 | L. braziliensis/L. guyanensis (hybrid) | L guyanensis | L guyanensis | 86.5 | 84.1 | |
| 1A | 2A | 2490 | L. naiffi/L. lainsoni (hybrid) | L lainsoni | L lainsoni | 85.7 | 84.3 | |
| 1D | 2B | 390 | L. guyanensis/L. braziliensis (hybrid) | L. braziliensis | L. braziliensis | 86.5 | 84.8 |
V 1 variants observed after Primer 1 amplification. V 2 variants observed after Primer 2 amplification.
Table 3.
Sensitivity and specificity values for Leishmania detection by HRM (P1) and by HSP70 qPCR, compared to cPCR and microscopy as gold standards.
Table 3.
Sensitivity and specificity values for Leishmania detection by HRM (P1) and by HSP70 qPCR, compared to cPCR and microscopy as gold standards.
| Assays | Patient samples (n = 60) | ||
| HRM + | HRM - | Total | |
| Microscopy + | 38 (82.6%) | 8 (17.4%) | 46 (100%) |
| Microscopy – | 8 (57.1%) | 6 (42.9%) | 14 (100%) |
| cPCR + | 42 (85.7%) | 7 (14.3%) | 49 (100%) |
| cPCR - | 4 (36.4%) | 7 (63.6%) | 11 (100%) |
| Assays | HSP70 qPCR + | HSP70 qPCR - | Total |
| Microscopy + | 37 (80.4%) | 9 (19.6%) | 46 (100%) |
| Microscopy – | 10 (71.4%) | 4 (28.6%) | 14 (100%) |
| cPCR + | 40 (81.6%) | 9 (18.4%) | 49 (100%) |
| cPCR - | 4 (36.4%) | 7 (63.6%) | 11 (100%) |
Table 4.
Results from HRM HSP70 as a species typing tool in clinical samples.
| HRM outcome | Samples successfully typed by RFLP (n = 45/60) |
| Inconclusive | 5/45 (11.2%) |
| Negative | 6/45 (13.3%) |
| Species typing achieved | 34/45 (75.5%) |
| HRM outcome | Samples with HRM species typing data (n = 34) |
| Divergent typing result | 8/34 (23.5%) |
| Convergent typing result | 26/34 (76.5%) |
| Total | 45 (100%) |
Table 5.
Primers used for HRM reactions. Amplicon sequences and references.
| ID | Sequence | Fragment | Reference |
| P1 | HSP70 F2 5’-GGAGAACTACGCGTACTCGATGAAG-3’HSP70C R 5’- TCCTTCGACGCCTCCTGGTTG-3’ | 144 pb | Zampieri et al. 2016/ Graça et al. 2012 |
| P2 | HSP70 F1 5’-AGCGCATGGTGAACGATGCGTC-3’HSP70 R1 5’-CTTCATCGAGTACGCGTAGTTCTCC-3’ | 104 pb | Zampieri et al. 2016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



