Submitted:
23 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
The application of melatonin (MT) has been shown to improve the quality during the storage of fruits and vegetables. The primary objective of this study is to investigate the effects of MT on the quality of fresh-cut Gastrodia elata during low temperature (4°C) storage. The results indicate that MT treatment could suppress the respiratory rate and malondialdehyde content of fresh-cut G. elata, while also slowing down the decline in total acidity and total soluble solids. The treatment with MT reduced the superoxide anions and hydrogen peroxide production, inhibited the activity and expression of peroxidase and polyphenol oxidase. Additionally, it led to increased activity and expression of antioxidant-related enzymes, including superoxide dismutase, catalase, ascorbate peroxidase, glutathione reductase, monodehydroascorbate reductase, and dehydroascorbate reductase, while also resulted in elevated levels of ascorbic acid and glutathione. Furthermore, the treatment with MT induced an increase in total phenolic and flavonoid content of fresh-cut G. elata and enhanced the activity and expression of key enzymes involved in phenylpropanoid pathway (phenylalanine ammonia-lyase, cinnamate-4-hydroxylase, 4-coumarate: CoA ligase). In summary, MT enhances the antioxidant capacity by activating both ROS metabolism and phenylpropanoid pathway, thus maintaining the quality of fresh cut G. elata.
Keywords:
fresh-cut Gastrodia elata
; melatonin
; ROS metabolism
; phenylpropanoid pathway
1. Introduction
Gastrodia elata, belonging to the Orchidaceae family, is rich in functional components such as gastrodin, amino acids, and polysaccharides [1]. In 2019, G. elata was designated as a pilot variety for the homology of medicine and food by the Chinese health commission. With G. elata being recognized as a food with medicinal properties, its applications as a food with medicinal properties are rapidly expanding [2]. Storing fresh G. elata for extended periods is challenging, and it is prone to rot [3]. Currently, research on the storage and preservation of fresh G. elata is relatively limited.
Melatonin (MT), a derivative of the essential amino acid tryptophan, is recognized as a novel plant hormone due to its diverse functions [4]. Prior research has shown that MT can scavenge reactive oxygen species (ROS) and enhance the quality of fruits and vegetables. Postharvest MT treatment has been shown to maintain the quality and delay senescence in jujube [5], plum [6], apple [7], and eggplant [8] during storage. However, the impact of MT treatment on the quality of fresh cut G. elata remains unknown.
It has been confirmed that ROS can directly interact with pathogens or act as signaling molecules in plant defense responses. However, excessive ROS can damage cells, disrupt cell membrane integrity, and accelerate plant aging [9]. Plants regulate ROS metabolism-related enzymes, such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), and glutathione reductase (GR), along with antioxidant substances like ascorbic acid (AsA) and glutathione (GSH), to maintain the balance of ROS [10]. Previous studies have proven that postharvest inducer treatments could enhance the enzymatic activity of oxygen metabolism-related enzymes, thereby improving the antioxidant capacity of apples [11], ginger rhizomes [12], pears [13], and blueberries [14].
The phenylpropanoid pathway, an integral secondary metabolic route within plants, assumes a pivotal responsibility in bolstering plant antioxidant mechanisms and fortifying resilience against diseases. A plethora of indispensable antimicrobial agents present in plants, encompassing flavonoids and phenolic compounds, originate from the intricate progression of the phenylpropanoid pathway, either through direct or indirect biosynthetic routes [15]. Pre-treatment with hot air has been shown to enhance the biosynthesis of phenolic compounds and maintain high antioxidant activity in fresh-cut pitaya fruits [16]. Furthermore, previous studies have demonstrated that the accumulation of phenylpropanoid pathway can enhance the antioxidant capacity of fresh-cut Agaricus bisporus [17], fresh-cut pitaya [18,19], and fresh-cut potato strips [20]. However, the impact of MT on the antioxidant capacity and phenylpropanoid pathway in fresh-cut G. elata has not been investigated yet.
The primary objective of the present study was to examine the impacts of MT treatment on the following: (1) the quality of fresh-cut G. elata during low temperature (4°C) storage, (2) the metabolism of antioxidant and phenylpropanoid of fresh-cut G. elata, that is to assess the efficacy of MT in improving the overall quality of fresh-cut G. elata.
2. Results
2.1. Effect of MT on phenotypic observation, weight loss, respiratory rate, total soluble solid (TSS), titratable acid (TA) and malondialdehyde (MDA) in fresh-cut G. elata
As depicted in Figure 1, the control group of fresh-cut G. elata exhibited significant browning on the surface after 6 days of storage at 4°C. However, MT treatment effectively inhibited the degree of browning in fresh-cut G. elata during the same period (Figure 1). In both groups, the fresh-cut G. elata exhibited an increasing trend in weight loss throughout the storage duration. However, in the MT-treated group showed a notable increase in weight loss at 4-10 days (Figure 2A). After MT treatment, the respiratory rate of fresh-cut G. elata remained consistently lower than the control throughout the storage stage and substantially lower at 2 and 6-10 days compared to the control (Figure 2B). Both the MT treatment and control groups of fresh cut G. elata exhibited a decreasing trend in TSS and TA throughout the entire experimental stage. However, the MT treatment significantly delayed the decrease in TSS and TA of fresh-cut G. elata from days 4-10 and 4-6, 10 (Figure 2C, D). The MDA content showed an increasing trend in both the control and MT treatment groups. However, the MT treatment markedly reduced the MDA content throughout the entire storage period (Figure 2E).
2.2. Effect of MT on ROS metabolism
As shown in Figure 3 (A, B), both the Superoxide anion (O2•-) production rate and hydrogen peroxide (H2O2) content in both MT treatment and control exhibited a gradual increase trend. Specifically, the group treated with MT exhibited a notable reduction in the production rate of O2•- compared to the control group on days 2, 5-7 (Figure 3A). Additionally, throughout the entire experimental period, the MT-treated group consistently demonstrated significantly lower levels of H2O2 content compared to the control group (Figure 3B). The MT-treated group exhibited a rapid increase in SOD activity within the first 2 days, reaching its peak at day 2. Although there were fluctuations in SOD activity afterward, it consistently remained significantly higher than the control group (Figure 3C). During the storage period, both the MT-treated and control groups demonstrated a progressive increase in CAT activity. Notably, the MT-treated group exhibited significantly higher CAT activity than the control group on days 2, 6-10 (Figure 3D). The control group displayed an upward trend in peroxidase (POD) and polyphenol oxidase (PPO) activities. In contrast, the MT-treated group exhibited significantly suppressed POD and PPO activities compared to the control group (Figure 3E, F).
In the MT treatment, APX activity rapidly increased from 0 to 4 days, followed by a decrease from 4 to 8 days. Specifically, the MT-treated group showed considerably higher APX activity compared to the control on days 2-8. On the other hand, the control exhibited an increase in APX activity from 0 to 6 days, followed by fluctuations (Figure 4A). The GR activity in the MT treatment group elevated from 0 to 8 days, followed by a decrease. Throughout the entire stage, the MT-treated group consistently exhibited greater GR activity compared to the control, with remarkable differences observed on days 2 and 6-10 (Figure 4B). The monodehydroascorbate reductase (MDHAR) activity raised from 0 to 6 days in the MT-treated group, followed by fluctuations. Throughout the entire storage period, the MT treatment consistently exhibited significantly higher MDHAR activity (Figure 4C). The control group showed a progressive increase in dehydroascorbate reductase (DHAR) activity from the initial day until day 6, followed by a subsequent decline. In contrast, the MT-treated group demonstrated a noteworthy rise in DHAR activity, notably surpassing the control on days 6 to 10 (Figure 4D).
2.3. Determination of AsA and GSH content
Both the MT-treated and control groups showed a similar trend in the change of AsA content, reaching their respective peak values at 6 and 4 days, followed by a decrease. Throughout the entire storage duration, the MT treatment consistently showcased notably elevated levels of AsA content in comparison to the control group, as illustrated in Figure 4E. The GSH content consistently reduced in the control. In contrast, the MT-treated group exhibited a continuous increase in GSH content from 0 to 2 days, followed by a decrease. Additionally, during days 2-6 and 10, the MT-treated group demonstrated notably higher GSH content compared to the control (Figure 4F).
2.4. Effects of MT treatment on total phenolic and flavonoid contents
The total phenolic content in both the control and MT treatment groups increased from 0 to 4 days and then declined. Nonetheless, over the entirety of the storage interval, the MT treatment consistently demonstrated significantly augmented levels when juxtaposed with the control group, as depicted in Figure 5A. The flavonoid content in the control exhibited fluctuations from 0 to 6 days, followed by a decrease. In contrast, the MT-treated group showed an increase in flavonoid content from 0 to 4 days, reaching its maximum level. Additionally, the MT-treated group exhibited an essential increase on days 4 and 8-10 (Figure 5B).
2.5. Effect of MT on phenylpropanoid pathway
The phenylalanine ammonia-lyase (PAL) activity of MT treatment increased consistently throughout the entire storage period and was considerably greater than the control. In contrast, the PAL activity in the control increased from 0 to 6 days and then fluctuated thereafter (Figure 5C). After MT treatment, the cinnamate-4-hydroxylase (C4H) activity of fresh-cut G. elata was markedly higher than the control at 2-4 and 8-10 days (Figure 5E). The control showed no significant changes in 4-coumarate: CoA ligase (4CL) activity throughout the entire experimental stage. However, the MT treatment exhibited a rapid increase in 4CL activity from 0 to 2 days, reaching its peak, followed by a gradual decline, and it was substantially greater than the control group from 2 to 8 days (Figure 5G).
2.6. Gene expressions of ROS metabolism of fresh-cut G. elata
In the MT treatment, GeSOD and GeCAT expressions rapidly increased from 2 days and reached their peak at 6 days. In particular, the GeSOD expression in the MT treatment remained substantially higher than the control throughout the storage period (Figure 6A), while the GeCAT expression was markedly higher than the control from 2 to 6 days (Figure 6B). As depicted in Figure 6, After MT treatment, GePOD and GePPO expressions in fresh-cut G. elata were considerably lower compared to the control throughout the experimental stage (Figure 6C, D). According to Figure 6, it is evident that MT treatment considerably enhanced the expression levels of key enzymes involved in ascorbate-glutathione cycle (AsA-GSH) (APX, GR, MDHAR, DHAR) to varying degrees (Figure 6E-I). As shown in Figure 5, MT treatment markedly elevated the expression of key enzymes involved in phenylpropane metabolism (PAL, C4H, 4CL) throughout the entire storage stage (Figure 5D, F, H).
3. Discussion
After plant wounds, the respiration and transpiration rates accelerate, which is an important factor leading to the aging and quality deterioration of fresh-cut fruits [21]. Slowing down respiration and transpiration is an effective approach to preserve the quality of fresh-cut fruits and vegetables. The study found that MT treatment effectively reduces the respiration rate of fresh-cut G. elata, delays quality deterioration, and significantly decreases the MDA content during storage, thereby slowing down the process of cell membrane lipid peroxidation. Tan et al. found that MT treatment effectively reduces the respiration rate of Chinese flowering cabbage, delaying the increase in weight loss and thus ensuring its postharvest quality [22]. In addition, in a study on wax apple, it was found that postharvest MT treatment significantly delayed the increase in weight loss and effectively reduced MDA content, alleviating cell membrane damage in wax apple [23].
During postharvest respiration, both TA and TSS are utilized as substrates, and TA and TSS are important indicators of plant quality after harvest. However, excessive respiration rate accelerates the depletion of TSS and TA, leading to a decline in plant quality [24]. This study found that MT effectively reduces the depletion of TSS and TA in fresh-cut G. elata, which is consistent with the significant decrease in respiration rate observed after treatment (Figure 3C, D). Likewise, similar results were obtained in studies on Chinese flowering cabbage [22] and wax apple [23] with MT treatment after harvest.
When plants encounter abiotic stress, they produce a notable quantity of ROS. Moderate levels of ROS can function as signaling molecules that trigger the defense system in plant, whereas elevated concentrations of ROS can lead to oxidative damage in plants [25]. In response to this situation, plants possess a comprehensive antioxidant system, comprising antioxidant enzymes like SOD and CAT, along with the AsA-GSH cycle. These mechanisms play a vital role in neutralizing excessive ROS and preserving a harmonious ROS equilibrium [9]. This study found that MT treatment effectively enhances the activities and expressions of SOD and CAT in fresh-cut G. elata. Simultaneously, it significantly reduces the rate of O2•- generation and the content of H2O2 during storage. Prior studies have found that post-treatment with MT significantly reduces H2O2 levels and enhances the activities of SOD and CAT in 'Newhall' navel oranges. This effectively delays the postharvest senescence of the oranges and maintains their quality [4]. According to Chen et al., the application of MT in wax apples has been shown to effectively decrease ROS accumulation while simultaneously enhancing the activities of antioxidant enzymes like SOD and CAT. These findings suggest that the external application of MT mitigates postharvest decay in wax apples by maintaining a delicate equilibrium between ROS generation and the activity of antioxidant system [23]. Furthermore, postharvest treatment with MT has been demonstrated to delay the senescence of various fruits and vegetables and enhance their disease resistance by regulating ROS metabolism and antioxidant capacity. This includes fresh-cut broccoli [26], sweet cherries [27], and mango fruit [28]. Therefore, we hypothesize that MT reduces the excessive ROS levels in fresh-cut G. elata by activating SOD and CAT.
In plant, the AsA-GSH cycle serves as an essential ROS clearance system, where key enzymes such as APX, GR, and glutathione peroxidase play critical roles. These enzymes participate directly or indirectly in the AsA-GSH cycle, protecting cells from oxidative damage [29]. The function of this cycle is to neutralize ROS by converting AsA and GSH to their oxidized forms, thereby maintaining the redox balance within the cell. APX converts AsA to dehydroascorbate [30], while GR reduces dehydroascorbate back to AsA and converts oxidized glutathione to reduced glutathione [31]. The synergistic action of these enzymes helps maintain the redox balance within plant cells, protecting them from oxidative damage. This study discovered that the application of MT treatment significantly boosts the activity and expression of APX and GR in fresh-cut G. elata. According to the research conducted by Aghdam et al., the application of MT treatment has been found to effectively boost the antioxidant capacity of postharvest pomegranate. This positive effect is attributed to the significant increase in the activities of two important antioxidant enzymes, APX and GR, in the pomegranate fruit [32]. Furthermore, similar results have been observed in researches of sweet cherries [27], jujubes [5], and white mushroom [33] with MT treatment.
The AsA-GSH cycle is a crucial system for maintaining cellular redox homeostasis, with AsA and GSH serving as major non-enzymatic antioxidants. AsA exhibits potent antioxidant properties and can be recycled back to its reduced form (AsA) from its oxidized form (Dehydroascorbic Acid) through interactions with DHAR and GSH [34]. Additionally, MDHAR is also involved in the AsA-GSH cycle, facilitating the conversion of monodehydroascorbate to AsA and thereby contributing to the normal functioning of this cycle [26]. During this investigation, it was observed that MT treatment could enhance the activities and expressions of MDHAR and DHAR to varying degrees, effectively increasing the contents of AsA and GSH during storage. Previous studies have revealed that exogenous MT can enhance AsA-GSH cycle by increasing AsA content, thereby maintaining quality of guava fruit and enhancing its disease resistance [35]. Treatment with MT has been shown to increase the AsA content in papaya fruit and improve its quality [36]. Aghdam et al. demonstrated that the application of MT effectively improved the levels of AsA and GSH in pomegranate fruit [32]. This resulted in improved antioxidant capacity and the preservation of fruit quality. Furthermore, Wang et al. demonstrated that exogenous MT treatment enhanced the activities of AsA-GSH cycle-related enzymes, MDHAR and DHAR, during the storage period of sweet cherries. It also increased the contents of AsA and GSH, thereby maintaining the postharvest quality of sweet cherries [27]. The aforementioned results suggest that MT treatment could activate the activity and expression of AsA-GSH cycle-related enzymes, such as APX, GR, MDHAR, and DHAR, further increasing the levels of AsA and GSH. Consequently, this helps eliminate excessive ROS in fresh-cut G. elata and maintain ROS balance.
When fresh-cut fruits and vegetables are subjected to cutting or other forms of mechanical stress, they respond to this injury by synthesizing higher levels of phenolic compounds, thereby enhancing their antioxidant capacity [37]. These phenolic compounds include flavonoids, anthocyanins, and phenolic acids, which exhibit strong antioxidant properties and can neutralize ROS, thereby reducing cellular oxidative damage [16]. Through this mechanism, fresh-cut fruits and vegetables can rapidly respond to cutting, enhance their antioxidant defense, prolong shelf life, and maintain nutritional value and texture. Based on this research, the application of MT treatment has been observed to significantly boost the accumulation of total phenolics and flavonoids in fresh-cut G. elata during storage, thus improving its antioxidant capacity. A previous experiment has demonstrated that exogenous MT treatment could enhance the total phenolic content in guava fruit, thereby maintaining its antioxidant capacity and enhancing its disease resistance [35]. MT treatment has also been shown to enhance the accumulation of total phenolics in sweet cherry fruit, thereby enhancing fruit antioxidant capacity and maintaining storage quality [38]. Furthermore, in studies conducted on cherry tomato fruit [39] and pomegranate fruits [32], it has been found that MT treatment could increase the content of phenolic compounds.
The phenylpropanoid pathway in plants is an important secondary metabolic pathway involved in the synthesis of various phenolic compounds. Within this pathway, PAL, 4CL, and C4H are three key rate-limiting enzymes [15]. PAL functions as the primary regulatory enzyme in phenylpropanoid metabolism, converting L-phenylalanine into cinnamic acid. Subsequently, C4H and 4CL catalyze the hydroxylation of different types of hydroxycinnamic acids into their corresponding thioesters, which then enter the synthesis pathway of phenolic compounds, flavonoids, and anthocyanins [40]. In this research, it was observed that the application of MT treatment led to notable increases in both the activities and expressions of PAL, C4H, and 4CL. These enhancements indirectly resulted in elevated levels of phenolic compounds. Previous research has revealed that MT treatment of blueberries Fruit leads to increased PAL, C4H, and 4CL activities and expressions, ensuring enhanced antioxidant capacity and disease resistance [41]. Similar findings in litchi fruit demonstrate that MT treatment is associated with the maintenance of fruit quality and the activities of PAL, C4H, and 4CL [42]. Additionally, similar conclusions have been obtained in studies conducted on cherry tomato [39], tomato fruits [43], and pomegranate fruits [32]. However, at the same time, certain phenolic compounds such as caffeic acid and catechins are susceptible to oxidation by PPO and POD, resulting in the formation of quinone polymers and brown pigments, leading to rapid browning of fresh-cut fruits and vegetables [18]. This phenomenon is attributed to the oxidation of the catechol structure present in the phenolic compounds, which generates quinone structures that subsequently polymerize to form oligomers and brown pigments. Our research results indicate that MT treatment effectively inhibits the activities and expressions of POD and PPO, leading to a significant reduction in the browning extent of fresh-cut G. elata during storage, which is consistent with the results depicted in Figure 1. Li et al. conducted research on fresh-cut white pitaya fruit and reported similar results. They found that subjecting the fruit to hot air pre-treatment resulted in enhanced activities of PAL, C4H, and 4CL, while simultaneously inhibiting the activities of POD and PPO [16]. This treatment effectively prevented the reduction of total phenolic content and successfully controlled browning in fresh-cut white pitaya fruit. Therefore, we hypothesize that MT treatment could enhance the antioxidant capacity of fresh-cut G. elata by activating the rapid accumulation of total phenolics through the phenylpropanoid pathway. Simultaneously, it effectively inhibited the activities of POD and PPO, preventing premature browning in fresh-cut G. elata and thus maintaining its postharvest quality.
4. Materials and Methods
4.1. Fruit and treatment
The G. elata was harvested from Zhaotong (103◦ 79′ E, 27◦ 38′ N), Yunnan Province, China, and taken to the laboratory by car. G. elata that were free from diseases, pests and mechanical damage and of uniform size were selected, washed with water, then soaked in 50 μM MT solutions for 5 min, which was confirmed by pre-experiment based on the inhibition effect of browning, air-dried, and sliced into 1 cm-thick pieces in a sterilized environment. At 0, 2, 4, 6, 8 and 10 days of 4°C storage, phenotypic observations, physiological and biochemical indexes were performed and fresh-cut G. elata was snap-frozen with liquefied nitrogen and stored in an ultra-low-temperature refrigerator (-80°C) for subsequent experiments. Three biological replicates, each containing four sections, were used for all index determinations.
4.2. Determination of weight loss, respiratory rate, TSS, TA and MDA
The weight loss rate (%) is calculated as: [(initial fresh-cut G. elata weight- present fresh-cut G. elata weight)] / initial fresh-cut G. elata weight] × 100. The respiration rate was assessed using a dual-wavelength infrared carbon dioxide analyzer (HM-GX20, Shandong Hengmei Electronic Technology Co., Ltd., China). Changes in CO2 concentration were tracked by enclosing four slices of fresh-cut G. elata in a sealed 100 mL container linked to the analyzer, forming a closed gas circulation system. Measurements were recorded at 1-minute intervals over five consecutive instances, and the respiration rates were expressed as mg CO2 kg-1h-1 fresh weight. TSS was measured using an ATAGO-PAL-1 digital refractometer and TA was titrated using NaOH (pH 13.0, 0.1 mol L-1) and then expressed as a percentage relative to TSS and TA. MDA content was determined to the method of Gao et al., where absorbance was measured at 450, 532 and 600 nm and expressed as mmol kg-1 FW [44].
4.3. Assays for ROS production and the activity of antioxidant scavenging enzymes.
O2•- production rate and H2O2 were measured on a UV spectrophotometer using O2•- and H2O2 Quantification Kits (Beyotime Biotechnology Co. Ltd., Beijing, China), respectively, in accordance with the specific procedures offered by the producer. The rate of O2•- generation and H2O2 content were indicated in mol kg−1 min−1 and mmol kg−1 FW, respectively.
For assessment the activity of antioxidant scavenging enzyme, SOD, APX, CAT, and GR extraction procedures are detailed in Ding et al. [45].
DHAR and MDHAR and were extracted according to the instructions of the kit (Solarbio Life Sciences. Co. Ltd., Beijing, China).
SOD activity was established by assessing the capacity to suppress the reductive effect of nitro blue tetrazolium in the presence of light [45]. 1U represents the amount of enzyme required to depress 50% of the photo-reduction reaction of NBT.
CAT activity was defined by examining the change in absorbance caused by H2O2 content at 240 nm [14]. 1U of CAT activity was defined as the amount of enzyme per minute that caused the decomposition of 1 μmol of H2O2.
APX activity was evaluated by the changed absorbance (290 nm) caused by the oxygenation of AsA after the addition of H2O2 [14]. 1U of APX activity was the amount of enzyme per minute that caused the oxidation of 1 μmol of AsA.
GR activity was measured change in absorbance due to NADPH oxidation in the absence of oxidized glutathione [14]. GR activity of 1U was calculated as the enzyme amount per minute leading to 1 nmol of NADPH oxidation.
The activities of DHAR and MDHAR were identified according to the kit instructions. 1 U of the activity of DHAR was defined as the enzyme amount catalyzing the production of 1 μmol of AsA per second and MDHAR was determined as the enzyme amount per second catalyzing 1 μmol NADPH oxidation. All the above antioxidant scavenging enzyme activities were shown in U kg-1 FW.
4.4. Activities of POD and PPO
In this study, we conducted the extraction of frozen tissues weighing 2.0 g using a 50 mM pre-chilled phosphate buffer solution (PBS) with a pH of 7.0, supplemented with 1% polyvinylpyrrolidone (PVP). The resultant supernatants served as the enzymatic source for the assessment of POD and PPO activities. To measure POD (EC 1.11.1.7) activity, the method outlined by Li et al. was adopted [16]. The reaction system consisted of a 1 mL mixture containing 50 mM PBS (pH 7.0), 20 mM H2O2, 1% guaiacol, and 50 μL of the extracted supernatant. The monitoring of absorbance changes at 560 nm allowed us to determine the enzymatic activity, with one unit of POD defined as the catalysis of 1 μmol of guaiacol oxidation per minute. For the assessment of PPO (EC 1.10.3.1) activity, we prepared a reaction mixture of 1 mL comprising 50 mM catechol, 50 μL of the extracted supernatant, and 0.1 M PBS at pH 6.8. The observation of absorbance changes at 420 nm facilitated the determination of enzymatic activity, with one unit of PPO defined as the oxidation of 1 nmol of catechol per minute. All the above enzyme activities were shown in 106 U kg-1 protein.
4.5. Determination of AsA and GSH content
The contents of AsA and GSH content were determined by reference to Dong et al. and expressed as mmol kg-1 FW, g kg-1 FW, mg kg-1 FW, and mmol kg-1 FW, respectively [46].
4.6. Analysis of the Metabolite Content in the Phenylpropanoid Pathway
Total phenolic, flavonoid, and lignin contents were extracted and measured using the method described by Liu et al. Total phenolic and lignin concentrations were measured at OD280 and OD325, respectively and determined as mg·Kg-1 FW [40].
4.7. Activities of key enzymes in phenylpropanoid metabolism
To extract key enzymes associated with phenylpropanoid metabolism in fresh-cut G. elata, the following procedure was employed: Fresh-cut tissues (2 g) were finely ground and mixed with distinct extraction solutions. For the PAL, an extraction solution was prepared using a borate buffer (0.1 mol L-1) adjusted to pH 8.7. This buffer contained 2 mmol L-1 of ethylene diamine tetraacetic acid, 5 mmol L-1 of β-mercaptoethanol, and 40 g L-1 of polyvinylpyrrolidone. To isolate C4H, the extraction solution comprised Tris-HCl buffer (50 mmol L-1) at pH 8.9, supplemented with 15 mmol L-1 of β-mercaptoethanol, 5 mmol L-1 of AsA, 4 mmol L-1 of MgCl2, 1 mmol L-1 of phenylmethanesulfonyl fluoride, and 10 μmol L-1 of leupeptin. For 4CL, the extraction solution consisted of Tris-HCl buffer (50 mmol L-1) at pH 8.0, containing 0.1 mol L-1 of DTT and 25% (v/v) glycerol. The activities of PAL, C4H, and 4CL were assessed using the methodology outlined by Li et al. and were expressed as 106 units per kg of protein (106 U kg-1 protein) [16].
4.8. Determination of gene expression
Gene expression was determined by quantitative Real-Time PCR (qRT-PCR) analysis. An RNeasy Plant Mini Kit (Takara, Japan) was used to extract RNA from tissue (1.0 g) in strict adherence to the exact procedure. RNA was purified using the manufacturer's instructions of TRIzolTM Plus RNA Purification Kit (Invitrogen™, Carlsbad, USA) to remove DNA contamination. Synthesis of first-strand cDNA was performed using the Prime-ScriptTM RT-PCR kit (TaKaRa, Dalian, China). qRT-PCR was performed using the KAPA 1-step qRT-PCR Kit (Bio-rad, California, USA) by CFX384 Touch Real-Time PCR Detection System. The ACTIN gene was used for quantitative normalization. Primers are shown in Table S1.
4.9. Data analysis
All physiological data were examined in triplicate and gene expression data were replicated in quadruplicate, with results shown as mean ± SE. These data were subjected to one-way analysis of variance (ANOVA) using SPSS version 17.0, and differences of P<0.05 were considered significant.
5. Conclusions
In conclusion, this study demonstrates that MT treatment enhanced the activity of antioxidant enzymes and effectively inhibits the activities of PPO and POD, thereby maintaining the postharvest quality of fresh-cut G. elata. Furthermore, MT treatment activates the phenylpropanoid pathway, increasing the content of total phenolics and flavonoids, and enhancing the antioxidant capacity of fresh-cut G. elata. Figure 7 illustrates the potential mechanisms by which MT treatment maintains the quality of fresh-cut G. elata.
Author Contributions
Boyu Dong: Writing - original draft, Writing - review & editing, Conceptualization, Data curation, Formal analysis, Funding acquisition. Fangfang Da: Software, Formal Analysis. Yulong Chen: Investigation, Formal Analysis. Xiaochun Ding: Funding acquisition, Formal analysis, Writing - review & editing.
Funding and Acknowledgments
This study was supported by the National Natural Science Foundation of China (32260795, 32002103); Guizhou Provincial Science and Technology Projects [Qian G. Z. K. J. (2021) 172]; Guangdong Basic and Applied Basic Research Foundation (2020A1515110092).
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Li, Y.; Zhang, Y.; Zhang, Z.; Hu, Y.; Cui, X.; Xiong, Y. Quality evaluation of Gastrodia elata tubers based on HPLC fingerprint analyses and quantitative analysis of multi-components by single marker. Molecules 2019, 24, 1521. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Chen, Z.; Guo, L.; Cui, X.; Xu, T.; Wan, F.; Yang, Y. Evaluate how steaming and sulfur fumigation change the microstructure, physicochemical properties and in vitro digestibility of Gastrodia elata Bl. starch. Front. Nutr. 2023, 9, 1087453. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Tan, C.; Tang, X.; Chu, Z.; Min, J.; Luo, Y. Study on Quality Influence of Hot Air Drying and Heat pump drying of Xiaocaoba Gastrodia elata. J. Phy. Confere. Ser. 2020, 4, 1601. [Google Scholar] [CrossRef]
- Ma, Q.; Lin, X.; Wei, Q.; Yang, X.; Zhang, Y.N.; Chen, J. Melatonin treatment delays postharvest senescence and maintains the organoleptic quality of ‘Newhall’navel orange (Citrus sinensis (L.) Osbeck) by inhibiting respiration and enhancing antioxidant capacity. Sci. Hortic. 2021, 286, 110236. [Google Scholar] [CrossRef]
- Tang, Q.; Li, C.; Ge, Y.; Li, X.; Cheng, Y.; Hou, J.; Li, J. Exogenous application of melatonin maintains storage quality of jujubes by enhancing anti-oxidative ability and suppressing the activity of cell wall-degrading enzymes. LWT-Food Sci. Technol. 2020, 127, 109431. [Google Scholar] [CrossRef]
- Lin, X.; Huang, S.; Huber, D.J.; Zhang, Q.; Wan, X.; Peng, J.; Zhu, S. Melatonin treatment affects wax composition and maintains storage quality in ‘Kongxin’Plum (Prunus salicina L. cv) during postharvest. Foods 2022, 11, 3972. [Google Scholar] [CrossRef]
- Fan, Y.; Li, C.; Li, Y.; Huang, R.; Guo, M.; Liu, J.; Ge, Y. Postharvest melatonin dipping maintains quality of apples by mediating sucrose metabolism. Plant Physiol. Bioch. 2022, 174, 43–50. [Google Scholar] [CrossRef]
- Song, L.; Zhang, W.; Li, Q.; Jiang, Z.; Wang, Y.; Xuan, S.; Chen, X. Melatonin alleviates chilling injury and maintains postharvest quality by enhancing antioxidant capacity and inhibiting cell wall degradation in cold-stored eggplant fruit. Postharvest Biol. Tec. 2022, 194, 112092. [Google Scholar] [CrossRef]
- Ge, Y.; Li, X.; Li, C.; Tang, Q.; Duan, B.; Cheng, Y.; Li, J. Effect of sodium nitroprusside on antioxidative enzymes and the phenylpropanoid pathway in blueberry fruit. Food Chem. 2019, 295, 607–612. [Google Scholar] [CrossRef]
- Chen, Y.; Hung, Y.C.; Chen, M.; Lin, M.; Lin, H. Enhanced storability of blueberries by acidic electrolyzed oxidizing water application may be mediated by regulating ROS metabolism. Food Chem. 2019, 270, 229–235. [Google Scholar] [CrossRef]
- Wei, M.; Ge, Y.; Li, C.; Han, X.; Qin, S.; Chen, Y.; Li, J. G6PDH regulated NADPH production and reactive oxygen species metabolism to enhance disease resistance against blue mold in apple fruit by acibenzolar-S-methyl. Postharvest Biol. Tec. 2019, 148, 228–235. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Wei, M.; Ge, Y.; Hou, J.; Cheng, Y.; Chen, J. Methyl jasmonate maintained antioxidative ability of ginger rhizomes by regulating antioxidant enzymes and energy metabolism. Sci. Hortic. 2019, 256, 108578. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Cheng, Y.; Hou, J.; Zhang, J.; Ge, Y. Postharvest application of acibenzolar-S-methyl delays the senescence of pear fruit by regulating reactive oxygen species and fatty acid metabolism. J. Agric. Food Chem. 2020, 68, 4991–4999. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Duan, B.; Li, C.; Tang, Q.; Li, X.; Wei, M.; Li, J. γ-Aminobutyric acid delays senescence of blueberry fruit by regulation of reactive oxygen species metabolism and phenylpropanoid pathway. Sci. Hortic. 2018, 240, 303–309. [Google Scholar] [CrossRef]
- Ge, Y.; Tang, Q.; Li, C.; Duan, B.; Li, X.; Wei, M.; Li, J. Acibenzolar-S-methyl treatment enhances antioxidant ability and phenylpropanoid pathway of blueberries during low temperature storage. LWT-Food Sci. Technol. 2019, 110, 48–53. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Li, M.; Fu, X.; Zhao, X.; Min, D.; Zhang, X. Hot air pretreatment alleviates browning of fresh-cut pitaya fruit by regulating phenylpropanoid pathway and ascorbate-glutathione cycle. Postharvest Biol. Tec. 2022, 190, 111954. [Google Scholar] [CrossRef]
- Zhang, L.; Li, S.; Wang, A.; Li, J.; Zong, W. Mild heat treatment inhibits the browning of fresh-cut Agaricus bisporus during cold storage. LWT-Food Sci. Technol. 2017, 82, 104–112. [Google Scholar] [CrossRef]
- Li, B.; Li, M.; Liu, J.; Sun, W.; Min, D.; Li, F.; Li, X. Methyl salicylate pretreatment maintains quality and antioxidant capacity of fresh-cut pitaya fruit by modulating phenylpropanoid metabolism and antioxidant system. Sci. Hortic. 2023, 309, 111705. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Ji, N.; Jin, P.; Zhang, J.; Zheng, Y.; Li, F. Cold plasma treatment induces phenolic accumulation and enhances antioxidant activity in fresh-cut pitaya (Hylocereus undatus) fruit. LWT-Food Sci. Technol. 2019, 115, 108447. [Google Scholar] [CrossRef]
- Zhou, F.; Xu, D.; Liu, C.; Chen, C.; Tian, M.; Jiang, A. Ascorbic acid treatment inhibits wound healing of fresh-cut potato strips by controlling phenylpropanoid metabolism. Postharvest Biol. Tec. 2021, 181, 111644. [Google Scholar] [CrossRef]
- Chen, Y.; Ge, Y.; Zhao, J.; Wei, M.; Li, C.; Hou, J.; Chen, J. Postharvest sodium nitroprusside treatment maintains storage quality of apple fruit by regulating sucrose metabolism. Postharvest Biol. Tec. 2019, 154, 115–120. [Google Scholar] [CrossRef]
- Tan, X.L.; Fan, Z.Q.; Zeng, Z.X.; Shan, W.; Kuang, J.F.; Lu, W.J.; Zhao, Y.T. Exogenous melatonin maintains leaf quality of postharvest Chinese flowering cabbage by modulating respiratory metabolism and energy status. Postharvest Biol. Tec. 2021, 177, 111524. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Nawaz, G.; Zhao, C.; Li, Y.; Dong, T.; Xu, T. Exogenous melatonin attenuates post-harvest decay by increasing antioxidant activity in wax apple (Syzygium samarangense). Front. Plant Sci. 2020, 11, 569779. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lin, H.; Shi, J.; Neethirajan, S.; Lin, Y.; Chen, Y.; Lin, Y. Effects of a novel chitosan formulation treatment on quality attributes and storage behavior of harvested litchi fruit. Food Chem. 2018, 252, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Du, C.; Xu, Y.; Wu, X. Exogenous nitric oxide improves chilling tolerance of Chinese cabbage seedlings by affecting antioxidant enzymes in leaves. Hortic. Environ. Biote. 2014, 55, 159–165. [Google Scholar] [CrossRef]
- Wei, L.; Liu, C.; Wang, J.; Younas, S.; Zheng, H.; Zheng, L. Melatonin immersion affects the quality of fresh-cut broccoli (Brassica oleracea L.) during cold storage: Focus on the antioxidant system. J. Food Process. Pres. 2020, 44, e14691. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.; Yang, Q.; Zhao, Q. Exogenous melatonin delays postharvest fruit senescence and maintains the quality of sweet cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Pareek, S.; Mani, S.; Domínguez-Avila, J.A.; González-Aguilar, G.A. A melatonin treatment delays postharvest senescence, maintains quality, reduces chilling injury, and regulates antioxidant metabolism in mango fruit. J. Food Quality, 2022, 2022, 1–18. [Google Scholar] [CrossRef]
- Ruíz-Torres, C.; Feriche-Linares, R.; Rodríguez-Ruíz, M.; Palma, J.M.; Corpas, F.J. Arsenic-induced stress activates sulfur metabolism in different organs of garlic (Allium sativum L.) plants accompanied by a general decline of the NADPH-generating systems in roots. J. Plant Physiol. 2017, 211, 27–35. [Google Scholar] [CrossRef]
- Yu, J.; Cang, J.; Lu, Q.; Fan, B.; Xu, Q.; Li, W.; Wang, X. ABA enhanced cold tolerance of wheat ‘dn1’via increasing ROS scavenging system. Plant Signal. Behav. 2020, 15, 1780403. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Moradi, M.; Razavi, F.; Rabiei, V. Exogenous phenylalanine application promotes chilling tolerance in tomato fruits during cold storage by ensuring supply of NADPH for activation of ROS scavenging systems. Sci. Hortic. 2019, 246, 818–825. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Luo, Z.; Li, L.; Jannatizadeh, A.; Fard, J.R.; Pirzad, F. Melatonin treatment maintains nutraceutical properties of pomegranate fruits during cold storage. Food Chem. 2020, 303, 125385. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kitazawa, H.; Zhang, X.; Zhang, L.; Sun, Y.; Wang, X.; Yu, S. Melatonin retards senescence via regulation of the electron leakage of postharvest white mushroom (Agaricus bisporus). Food Chem. 2021, 340, 127833. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Cheng, Y.; Li, C.; Guo, M.; Zhu, J.; Ge, Y.; Sun, T. Postharvest application of acibenzolar-S-methyl delays the senescence of pears by mediating the ascorbate-glutathione cycle. Sci. Hortic. 2022, 293, 110741. [Google Scholar] [CrossRef]
- Fan, S.; Xiong, T.; Lei, Q.; Tan, Q.; Cai, J.; Song, Z.; Zhu, X. Melatonin treatment improves postharvest preservation and resistance of guava fruit (Psidium guajava L.). Foods 2022, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Li, Q.; Feng, S.; Lei, Q.; Abbas, F.; Yao, Y.; Zhu, X. Melatonin maintains fruit quality and reduces anthracnose in postharvest papaya via enhancement of antioxidants and inhibition of pathogen development. Antioxidants 2022, 11, 804. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; Wang, J.; Wu, Z.; Xu, Y.; Wu, Z. Ozone treatment promotes physicochemical properties and antioxidant capacity of fresh-cut red pitaya based on phenolic metabolism. Front. Nutr. 2022, 9, 1016607. [Google Scholar] [CrossRef]
- Carrión-Antolí, A; Martínez-Romero, D.; Guillén, F.; Zapata, P.J.; Serrano, M.; Valero, D. Melatonin pre-harvest treatments leads to maintenance of sweet cherry quality during storage by increasing antioxidant systems. Front. Plant Sci. 2022, 13, 863467. [Google Scholar] [CrossRef]
- Li, S.; Xu, Y.; Bi, Y.; Zhang, B.; Shen, S.; Jiang, T.; Zheng, X. Melatonin treatment inhibits gray mold and induces disease resistance in cherry tomato fruit during postharvest. Postharvest Biol. Tec. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Liu, Y.; Ge, Y.; Bi, Y.; Li, C.; Deng, H.; Hu, L.; Dong, B. Effect of postharvest acibenzolar-S-methyl dipping on phenylpropanoid pathway metabolism in muskmelon (Cucumis melo L.) fruits. Sci. Hortic. 2014, 168, 113–119. [Google Scholar] [CrossRef]
- Qu, G.; Wu, W.; Ba, L.; Ma, C.; Ji, N.; Cao, S. Melatonin enhances the postharvest disease resistance of blueberries fruit by modulating the jasmonic acid signaling pathway and phenylpropanoid metabolites. Front. Chem. 2022, 10, 957581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, T.; Liu, G.; Hu, M.; Yun, Z.; Duan, X.; Jiang, G. Inhibition of downy blight and enhancement of resistance in litchi fruit by postharvest application of melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, Y.; Aghdam, M.S.; Luo, Z.; Jannatizadeh, A.; Razavi, F.; Fard, J.R.; Farmani, B. Melatonin treatment promotes endogenous melatonin accumulation and triggers GABA shunt pathway activity in tomato fruits during cold storage. Sci. Hortic. 2019, 254, 222–227. [Google Scholar] [CrossRef]
- Gao, H.; Lu, Z.; Yang, Y.; Wang, D.; Yang, T.; Cao, M.; Cao, W. Melatonin treatment reduces chilling injury in peach fruit through its regulation of membrane fatty acid contents and phenolic metabolism. Food Chem. 2018, 245, 659–666. [Google Scholar] [CrossRef]
- Ding, X.; Zhu, X.; Zheng, W.; Li, F.; Xiao, S.; Duan, X. BTH treatment delays the senescence of postharvest pitaya fruit in relation to enhancing antioxidant system and phenylpropanoid pathway. Foods 2021, 10, 846. [Google Scholar] [CrossRef]
- Dong, B.; Zhu, D.; Yao, Q.; Tang, H.; Ding, X. Hydrogen-rich water treatment maintains the quality of Rosa sterilis fruit by regulating antioxidant capacity and energy metabolism. LWT-Food Sci. Technol. 2022, 161, 113361. [Google Scholar] [CrossRef]
Figure 1.
Visual appearance of fresh-cut G. elata after 0 and 50 μmol/L melatonin treatment during storage at 4℃.
Figure 1.
Visual appearance of fresh-cut G. elata after 0 and 50 μmol/L melatonin treatment during storage at 4℃.
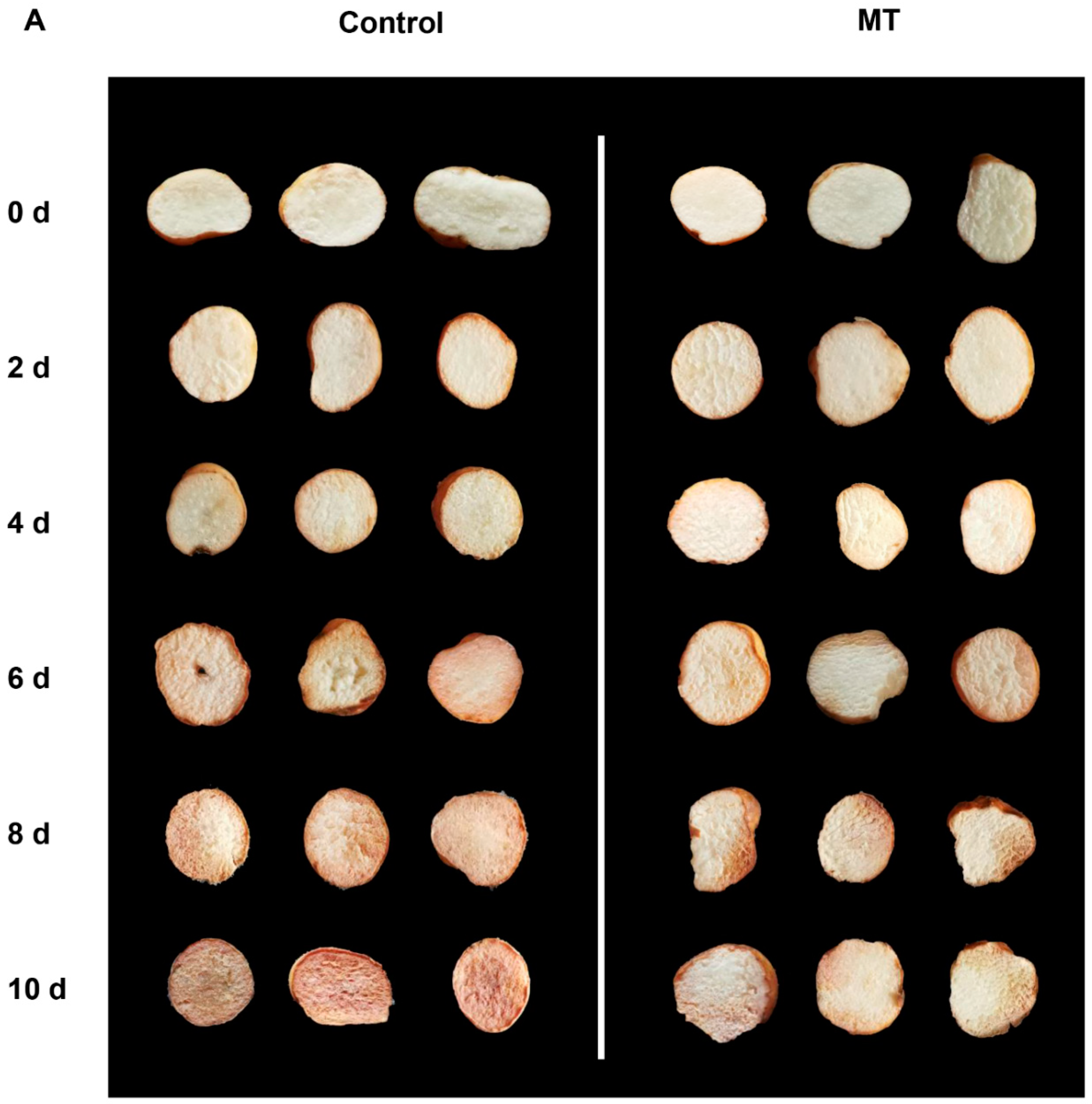
Figure 2.
Effect of melatonin treatment on weight loss (A), respiratory rate (B), TSS (C), TA (D), and MDA (E). * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
Figure 2.
Effect of melatonin treatment on weight loss (A), respiratory rate (B), TSS (C), TA (D), and MDA (E). * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
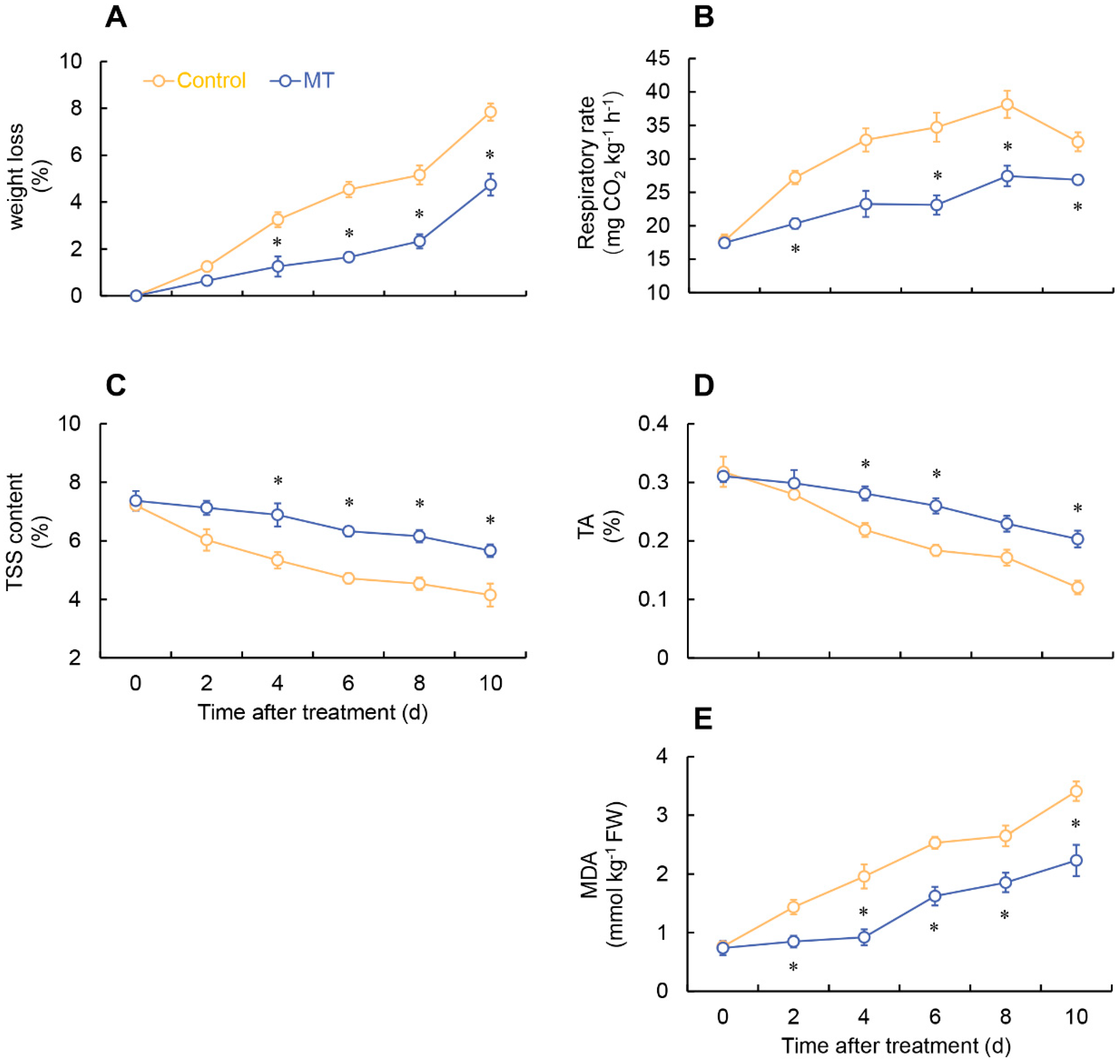
Figure 3.
Changes in O2•- production rate (A), H2O2 content (B), and the activity of SOD (C), CAT (D), POD (E) and PPO (E) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
Figure 3.
Changes in O2•- production rate (A), H2O2 content (B), and the activity of SOD (C), CAT (D), POD (E) and PPO (E) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
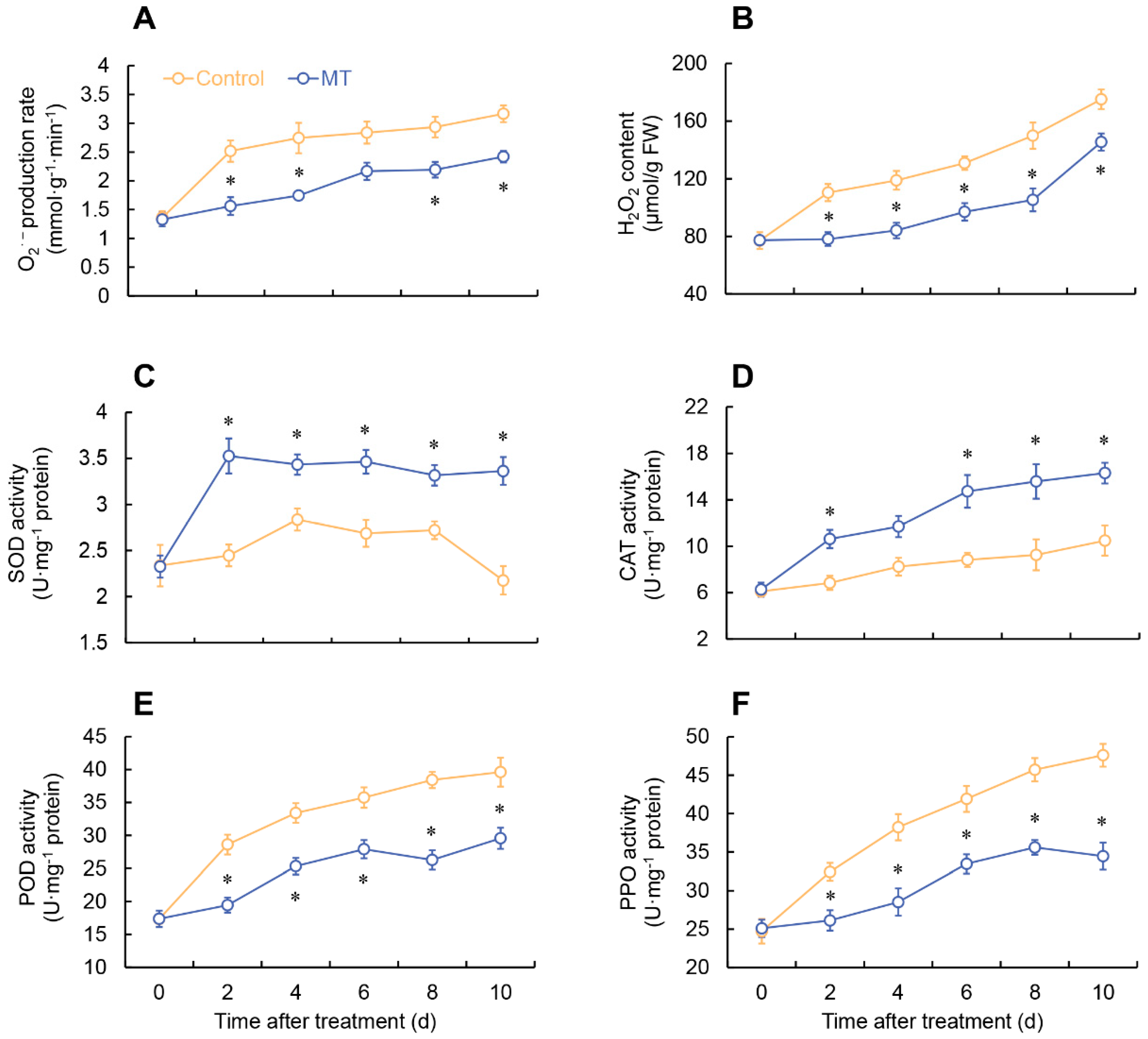
Figure 4.
Changes in the activity of APX (A), GR (B), MDHAR (C) and DHAR (D), and the content of AsA (E), and GSH (F) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
Figure 4.
Changes in the activity of APX (A), GR (B), MDHAR (C) and DHAR (D), and the content of AsA (E), and GSH (F) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
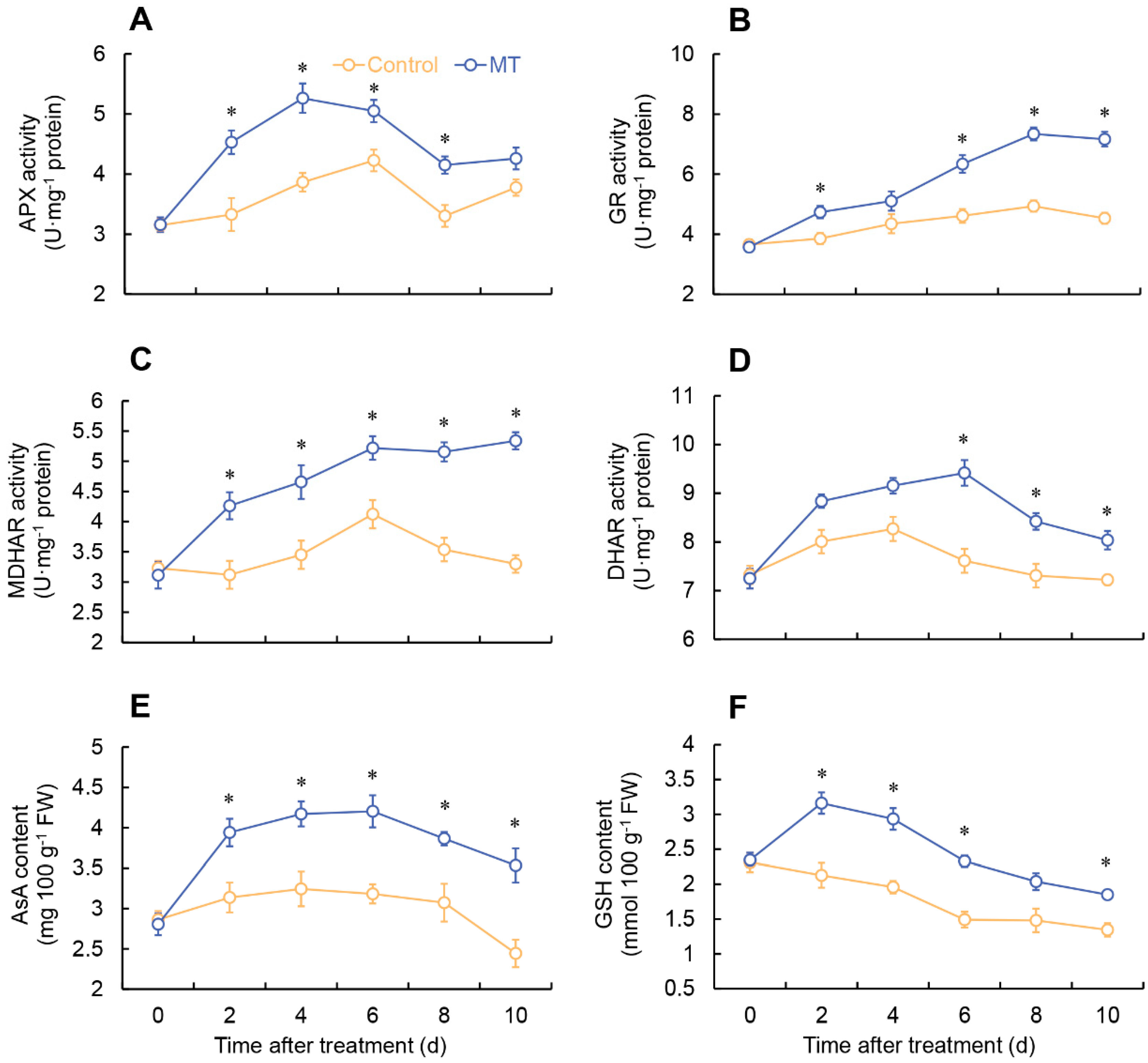
Figure 5.
Changes in the content of total phenols (A) and flavonoids (B), and the activity and expression of PAL (C, D), C4H (E, F) and 4CL (G, H) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
Figure 5.
Changes in the content of total phenols (A) and flavonoids (B), and the activity and expression of PAL (C, D), C4H (E, F) and 4CL (G, H) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
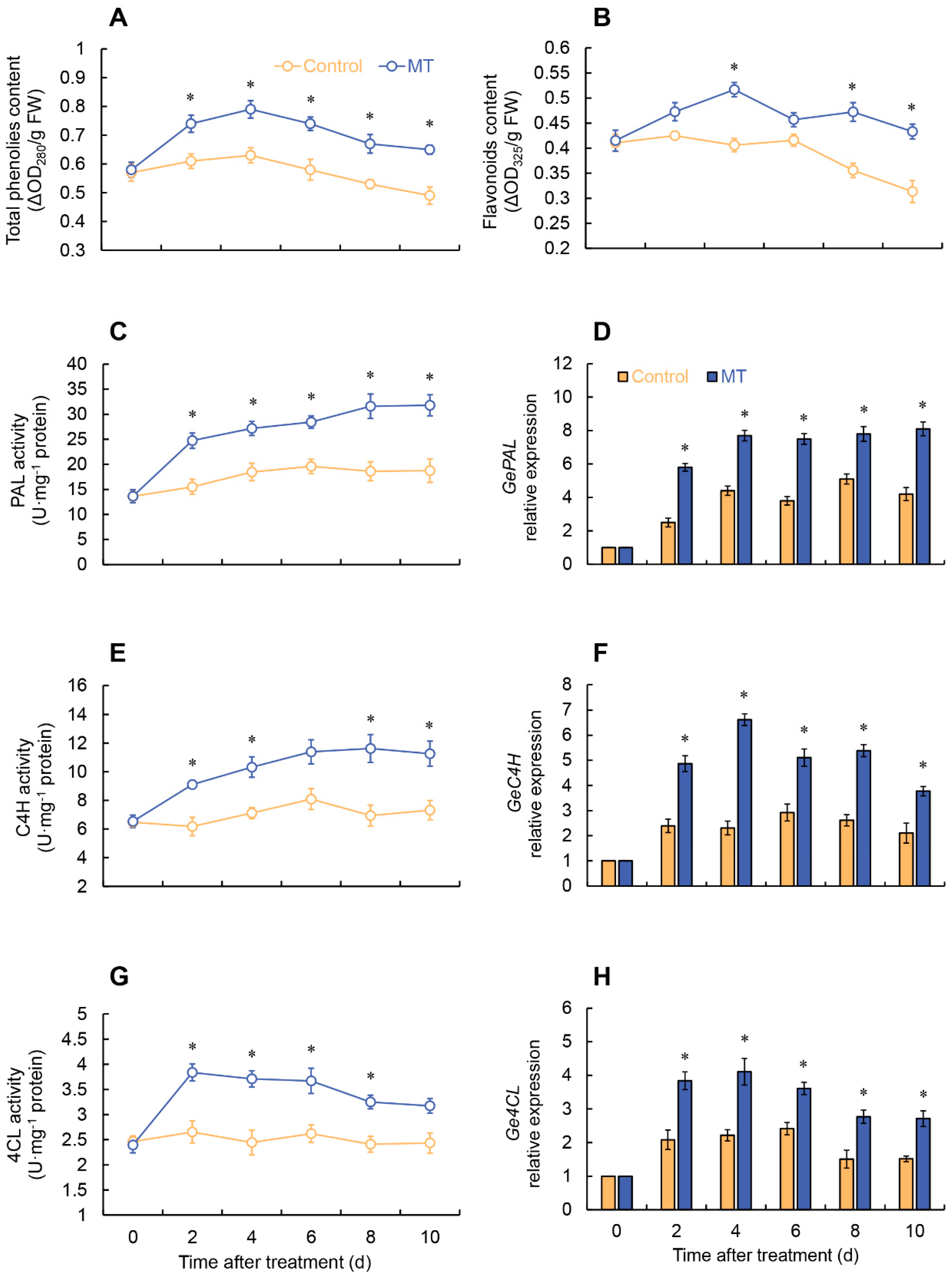
Figure 6.
Expression of GeSOD (A), GeCAT (B), GePOD (C), GePPO (D), GeAPX (E), GeGR (F), GeMDHAR (G) and GeDHAR (H) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
Figure 6.
Expression of GeSOD (A), GeCAT (B), GePOD (C), GePPO (D), GeAPX (E), GeGR (F), GeMDHAR (G) and GeDHAR (H) in fresh-cut G. elata after melatonin treatment during storage at 4℃. * denotes significant difference at the level of P < 0.05. Vertical bars represent the standard errors of the means (±SE).
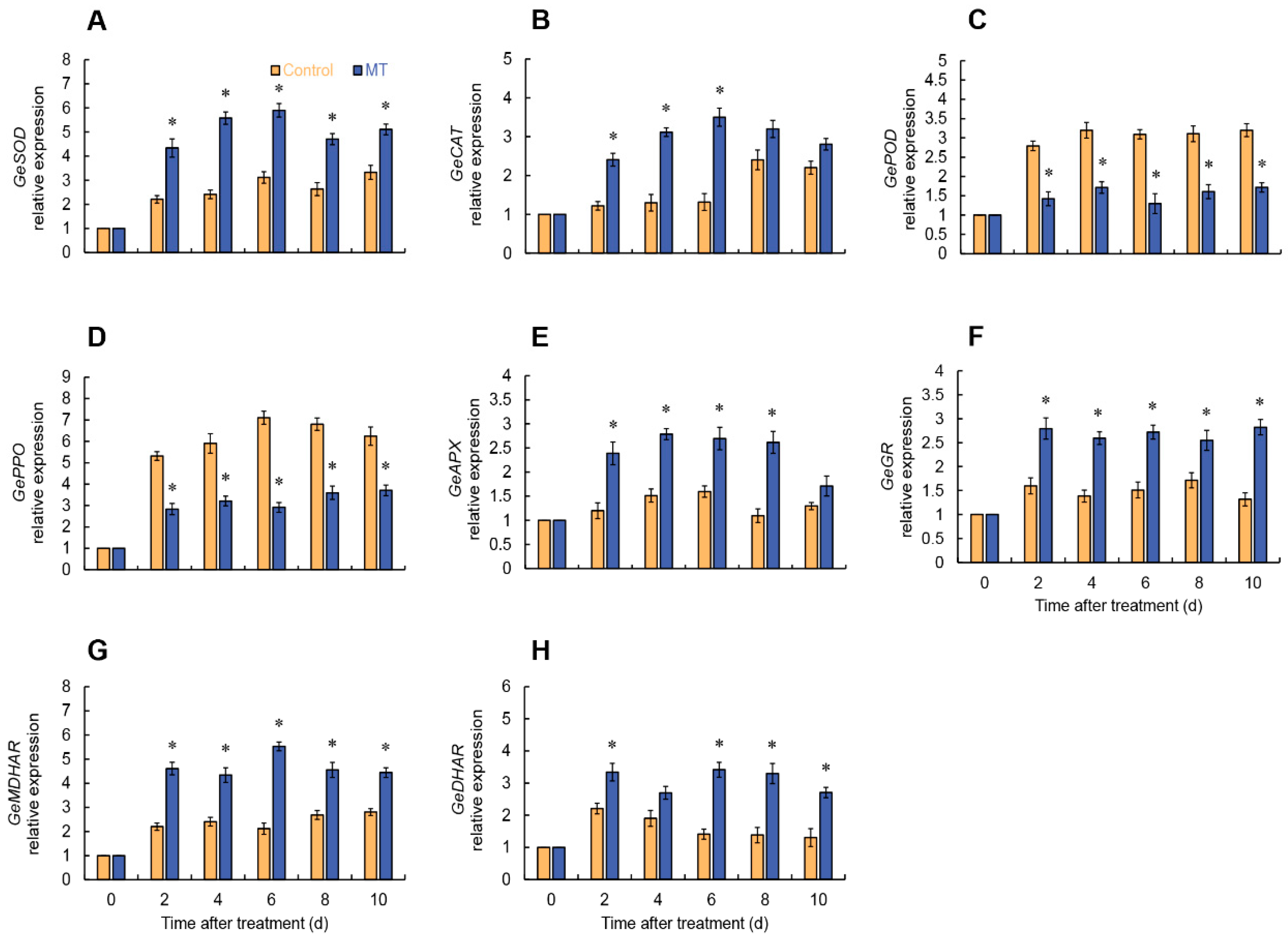
Figure 7.
A speculation mechanism to explain the role of melatonin in mainting quality in fresh-cut G. elata.
Figure 7.
A speculation mechanism to explain the role of melatonin in mainting quality in fresh-cut G. elata.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



