
Submitted:
23 August 2023
Posted:
25 August 2023
You are already at the latest version
Abstract
Inflammatory Bowel Disease (IBD) is a chronic and progressive inflammatory disorder affecting the gastrointestinal tract (GT) caused by a wide range of genetic, microbial, and environmental factors. IBD is characterized by chronic inflammation, decreased gut microbial diversity and dysbiosis, with a lower amount of beneficial bacteria and a concomitant increase of pathogenic species. It is well known that dysbiosis is closely related to the induction of inflammation and oxidative stress, this last caused by an imbalance between reactive oxygen species (ROS) production and cellular antioxidant capacity that leads to cellular ROS accumulation. ROS are responsible for the intestinal epithelium oxidative damage and the increased intestinal permeability found in IBD patients and their reduction could represent a potential therapeutic strategy to limit the IBD progression and alleviate its symptom. Recent evidence highlighted that dietary polyphenols, natural antioxidants, are able to maintain redox equilibrium in the GT preventing gut dysbiosis, intestinal epithelium damage and radical inflammatory responses. Here, we suggest that the relatively new foodomics approaches, together with new technologies for promoting the antioxidative properties of dietary polyphenols, including novel delivery systems, chemical modifications and combination strategies, may provide critical insight to determine the clinical value of polyphenols for IBD therapy and a comprehensive perspective for implementing natural antioxidants as potential IBD candidate treatment.
Keywords:
inflammatory bowel diseases
; omics
; polyphenols
; inflammation
; antioxidative properties
; gut dysbiosis
1. Introduction
The first important interactions of nutrients with the human body occur at the level of the gastrointestinal tract (GT), whose primary function is the digestion and absorption of food, including solids and liquids. Notably, GT also displays an important role of barrier against many potentially harmful ingested substances. The epithelial barrier's functionality is ensured by tight junctions, adherens junctions, and desmosomes. Moreover, epithelial cells fully contribute to the defense of the intestine by secreting bactericidal substances, such as defensins, both naturally and in response to the pattern recognition receptors of bacteria (PRR) [1]. Thus, the health and defense of the human body depend on the integrity of the gut mucosa so that, for years now, many authors have identified a new condition: the ‘Leaky Gut Syndrome’ [2].
The integrity of GT is also correlated with the presence of the so-called intestinal microbiota. The number of microorganisms inhabiting the gastrointestinal tract is estimated to exceed 1014 (100 trillion) [3], with a rich bacterial diversity containing more than 1000 species [4] and the most represented phyla are: Firmicutes (49–76%), Bacteroidetes (16–23%), Proteobacteria, and Actinobacteria [5]. These bacteria are responsible for mineral absorption and carbohydrate degradation [6], as well as for aminoacids and vitamins synthesis [7]. In addition, gut microbiota enhances mucosal immunity tolerance through metabolizing dietary fiber into short-chain fatty acids (SCFAs) [8], such as butyrate, propionate, acetate and activating regulatory T cells (Tregs), a T cell subset specialized in suppressing immune response and maintaining self-tolerance and tissue homeostasis [9,10].
Furthermore, the production from intestinal commensal Clostridium sporogenes of indoles and its derivatives, such as tryptophan metabolites and indoleacrylic acid, is responsible for the activation of the aryl hydrocarbon receptor [11] and for the induction of mucin family gene expression and the activation of the nuclear factor erythroid 2-related factor 2 (Nrf2), a crucial regulator of the cellular defense mechanisms against xenobiotic and oxidative stress, thus enhancing the expression of antioxidant enzymes and molecules in the GT [12].
At the same time, Bacteroides species metabolize dietary fiber into succinate [13] which is responsible for the stabilization of hypoxia-inducible factor-1α (HIF-1α) and the activation of dendritic cells, thus promoting IL-1β production with consequent triggering of an inflammatory response [14]. In addition, inflammatory responses are also modulated by sphingolipids, especially ceramide, synthetized by Bacteroides via bacterial serine-palmitoil transferase, which turns off the inflammation in host cells [15,16]. Finally, the decrease in pH caused by the high concentration of SCFA prevents the growth of potentially pathogenic bacteria, such as Escherichia coli and other members of the Enterobacteriaceae [17]. Due to the close relationship between the intestinal barrier and microbiota, gastrointestinal system functions as efficiently as possible, making a significant contribution to homeostasis and human health. Everything that in any way affects or disrupts this equilibrium, such as molecules from foods, exposure to chemicals, or other types of stressors, has the potential to change how the intestinal barrier works and harm people's health [18].
2. Inflammatory Bowel Disease
Several studies highlighted how GT alterations at various levels can compromise its functions and also lead to the onset of chronic diseases as well as Inflammatory Bowel Disease (IBD) and autoimmune disorders [19,20].
Crohn's disease (CD) is the most common subtype of IBD both in adults and in children, comprising 59-73% of pediatric IBD, that affects the entire GT [21]. Weight loss, watery diarrhea, and abdominal pain are the traditional symptoms of this illness. Particularly, the clinical signs and symptoms resemble those of allergic and infectious gastroenteritis. The pain is often of the colicky type and is more frequently found in the right lower quadrant of the stomach due to the increased involvement of the terminal ileum [22]. Most often watery, bloody diarrhea is more common when the colon and rectum are involved [23].
While CD is defined by dispersed “jumpy” lesions in the entire intestinal tract, from the mouth to the anus, typically in different evolutionary stages, Ulcerative Colitis (UC) is a recurrent and remitting IBD which is represented only by mucosal inflammation starting distally, in the rectum, and potentially spreading to cover the entire intestine (continuous ascending colitis) [24,25]. Yet, compared to CD, UC has a higher frequency of bleeding (83–95%). The mucous layer of the intestinal wall typically suffers superficial damage as a result of UC inflammation, and bloody diarrhea is the most common symptom in those who are affected [26].
Although there are still many unclear points about the pathophysiology of IBD, it appears to entail a number of factors, such as genetic predisposition, epigenetic or gene activity regulating factors [27] altered immune responses, environmental factors and changed intestinal flora [26]. Recent studies underline the existence of an interesting connection between oxidative stress, gut microbiota, and immune response [28].
Reactive oxygen species (ROS), the main responsible for oxidative stress, exert dual functions in a dose-dependent way: low concentrations of ROS regulate cellular physiological processes, including redox signal transduction, gene expression, and receptor activation which are beneficial for tissue turnover and cell proliferation [29] while higher concentrations of ROS can damage cellular molecules such as DNA, proteins, and lipids, leading to cell senescence and even to death [30].
It is well known that the arising and progression of IBD are accompanied by persistent oxidative stress and inflammatory responses that has been correlated with dysbiosis of the gut microbiota [31]. External stimuli, including high-fat diet, smoking, circadian rhythms, and drug intervention, can induce oxidative stress which leads to gut dysbiosis [32]. Notably, IBD patients are characterized by a decreased gut microbial diversity and dysbiosis with a reduced proportion of Firmicutes and Bacteroidetes and an increased ratio of Proteobacteria [33]. GT of IBD patients contains lower levels of beneficial bacteria, such as Bacteroides, Lactobacillus, and Eubacterium, [34] and a concomitant increase of pathogenic species, including Enteroinvasive Escherichia coli (EIEC) [35]. The related dysbiosis has been correlated with the induction of oxidative stress and inflammation that are responsible for the intestinal epithelium damage and increased intestinal permeability [36]. This in turn results in the release in the blood of harmful bacterial metabolic toxins, such as lipopolysaccharide (LPS), and pro-inflammatory chemokines and cytokines, such as tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), and interferon-γ (IFN-γ) [32], causing a worsening of the inflammatory process that characterizes IBD patients [36].
Oxidative stress induced by dysbiosis can act as upstream stimuli to evoke aberrant activation of the intestinal immune system, causing destruction of tight junction and damage to the intestinal mucosal barrier with antimicrobial peptide secretion. The release of pro-inflammatory factors (e.g., TNF-α, IL-6, and IL-1β) and the activation of pro-inflammatory enzymes (e.g., iNOS, COX-2, and NOX) [37] via Nuclear Factor-κB (NF-κB), JAK/STAT, and Mitogen-Activated Protein Kinases (MAPKs) signaling pathways [38], leads to a further increase of the level of ROS in the GT [39] and a remodeling of the gut microbiota, in a vicious cycle in which dysbiosis and inflammation support each other. Restoring the homeostasis of gut microbiota of IBD patients in favor of beneficial versus pathogenic bacterial species could be helpful to relieve oxidative stress and inflammation. Thus, ROS are considered among the main culprits of IBD pathogenesis and a potential therapeutic target to limit its progression and alleviate its symptoms.
3. Potential Beneficial Effects of Dietary Polyphenols in IBD
Conventional IBD treatments are intent to turn-off the inflammation in the acute phase of disease and/or extend remission periods and these include aminosalicylates, corticosteroids, antibiotics, immunosuppressive agents, and biologics [40], such as therapeutic TNF-α antagonists. These last are able to restore the gut barrier [41], demonstrating a crucial role for TNF-α in the pathogenesis of IBD, due to its pro-inflammatory and pro-apoptotic effect.
These treatments could be effective for some patients, but many of them could lose the response over time or do not respond at all, not to mention their dangerous side effects, such as osteoporosis for corticosteroids [42] or increased risk of infection and gastrointestinal distress for immunosuppressants and aminosalicylates, respectively [40]. Finally, biologics, although very effective, are never first line medications because of their high cost [43] and, often, IBD patients require lifelong treatment.
Collectively, novel therapeutic strategies for IBD treatment able to reduce oxidative stress and inflammation are urgently required. In this scenario, polyphenols are among the most suitable candidates.
Polyphenols are characterized by a structure composed of multiple phenolic units and they are introduced with a diet made of vegetables, fruits, nuts, and beverages such as coffee, tea, wine, and beer [44,45]. They could have very low molecular weight, such as coumarins, or they could be polymerized compounds, as tannins, and conjugated or not with one or more sugar residues as glycosides. However, due to the presence of the hydroxyl group in their structure, polyphenols can scavenge ROS by hydrogen atom transfer or single-electron transfer reactions [46], having an important antioxidant and anti-inflammatory activity.
Several experimental evidences have linked polyphenol dietary intake to a reduced risk for cancer [47], obesity [48,49,50], type 2 diabetes [51] and metabolic syndrome [52]. In the recent years, multiple in vitro and in vivo studies supported the hypothesis of a protective role for polyphenols administration in IBD [53], by modulating the expression of transcription factor inflammatory targets, oxidative stress, intestinal barrier function, pathogen defense, composition of gut microbiota nd its metabolite production, crucial for maintaining intestinal homeostasis.
The ultimate outcome of these processes appears to be a reduction in the pro-inflammatory cytokine storm that characterizes IBD. Larussa T. et al analyzed biopsies obtained during colonoscopy from 14 patients with active UC, which were immediately placed in an organ culture chamber and treated with LPS from Escherichia coli, in the presence or absence of oleuropein (OLE), a glycosylated phenolic compound found in green olive skin, flesh, seeds, and leaves. The expression of IL-17 and COX-2 was significantly lower in the OLE-treated samples. Furthermore, OLE-treated colonic samples showed signs of reduced mucosal inflammation, with lower infiltration of B and T lymphocytes [54]. Similar effects were obtained in an ex vivo model of CD, in which exposure of intestinal biopsies to extracts from Olea europaea leaves reduced the production of pro-inflammatory mediators (IL-1β, IL-6, IL-8, and TNF-α) [55].
A recent metanalysis analyzed the effects of polyphenols from olives on murine models of UC. Authors proved that, although the use of these substances was not able to effectively prevent the development of the disease, in most UC models, olives constituents guaranteed a milder expression of the disease, better weight maintenance and a reduction in rectal bleeding. Of note, according to the original aim of this study, the metanalysis should include both animals and humans trials, but no human study met the eligibility criteria [56]. To date, only very few studies analyzed the effects of polyphenols intake in patients suffering from UC [57,58,59,60,61,62,63,64] and none on CD. Most of these considered the use of curcumin either as oral intake or as enema, other used sylmarine or resveratrol. The results obtained from these studies are controversial, regardless of the compound used, some of them demonstrating an improvement of both UC clinical [65], and inflammatory [62] parameters, others showing no effect [59]. These studies suffer of a great degree of heterogenicity, with different outcomes analyzed, different compounds used as well as variable concentration and way of administration, different UC activity status and, finally, low number of enrolled patients.
Other studies enquired the effectiveness of polyphenols on IBD; however, these used products in which coexists several polyphenols as well as other substances like extra virgin olive oil [66] and mango pulp [67], so that no specific conclusion could be drawn. Moreover, limiting the potential effectiveness of polyphenols anti-inflammatory effects, many polyphenols that we introduce with the diet are poorly absorbed, resulting in their poor bioavailability. Thus, an increase of the intestinal mucosal exposure, the target tissue of IBD treatment, is required. In addition, in the intestinal lumen polyphenols interact with intestinal microbiota that modify them, altering their pharmacokinetics and producing metabolites which are completely different from the polyphenols that generated them [68] and with a greater relevance and bioavailability than their precursors [31].
4. Molecular Mechanisms Underlying the Effects of Dietary Polyphenols in IBD
A great number of studies have shown that polyphenolic compounds showed inhibitory effects on NF-κB, an inducible transcription factor which regulates a large array of genes involved in different inflammatory processes, including flavones, isoflavones, flavonols, flavanones, chalcones, anthocyanins, lignans, stilbenes, and phenolic acids. Furthermore, many of them also positively regulate antioxidant signaling pathways, such as Nrf2 [72,73] to intensify the intestinal mucosal barrier, which is also beneficial for the homeostasis of the gut microbiota [74].
NF-κB, responsible for the transcription of several pro-inflammatory genes, including those for pro-inflammatory cytokines, (e.g., TNF-α and IFN-γ), has been found markedly overactivated in IBD, as evidenced by the increase of NF-κB p65 expression in biopsied tissue from IBD patients, particularly CD [75]. Thus, some authors have reported a direct and positive correlation between NF-κB activation in IBD patients and chronic mucosal inflammation [76]. The same pro-inflammatory cytokines, microbial endotoxins (e.g. LPS), and ROS [77] may be responsible for NF-κB activation, which is mediated by the phosphorylation of IkB, its inhibitory subunit, by a specific IkB kinase (IKK). Once phosphorylated, IkB is degraded and NF-κB is free to move to the nucleus of immune cells and enterocytes where induces the transcription of its target genes [77].
The NF-κB inhibitory effects of polyphenols were described largely in vitro [78] by comparing 36 flavonoids on NF-κB activation in murine J774 macrophages. Daidzein, genistein, isorhamnetin, kaempferol, quercetin, naringenin, and pelargonidin inhibited NF-κB activation, reducing nitric oxide (NO) production whereas stilbenes, such as resveratrol [79] and lignans, including arctigenin, and demethyltraxillagenin [80], inhibited NF-κB by similar mechanisms but with different degrees of potency. All these findings have suggested the idea that polyphenols may be more potent anti-inflammatory agents than some conventional IBD therapies, such as 5-aminosalicylic acid, a common IBD anti-inflammatory drug, since they were effective already at lower concentrations (micromolar) than the others (millimolar) [81].
Most of the polyphenols act on IKK complex, preventing IkB phosphorylation and NF-kB translocation to the nucleus. More specifically, some of them, such as apigenin and quercetin, mediate the inhibition of the IKK-γ regulatory subunit [82,83], whereas other flavonoids, such as morin [84], fisetin [85], and gossypin [86] inhibit the IKK-β regulatory subunit [87,88]. A further mechanism of control of NF-κB activity involves its phosphorylation on p50 subunit, via phosphatidylinositol 3-kinase (PI3K)/Akt pathway, influencing NF-κB DNA binding and its transcriptional activity. Several flavonoids have been shown to inhibit this pathway through the inhibition of Akt or PI3K [87,89].
Toll-like receptors (TLRs) are receptors through which immune cells, but also epithelial, endothelial cells and fibroblasts, recognize microbials molecules in order to protect us from pathogens or prevent an inflammatory response aberrantly triggered by commensal microbes [90].
Intestinal mucosa of IBD patients showed an elevated expression of LPS binding toll-like receptor 4 (TLR4), suggesting its involvement in the inflammation characterizing IBD patients. In fact, there is a close connection between TLRs and production of pro-inflammatory cytokines [91]. Once activated, TLRs signaling converge on NF-kB activation, that in turns leads to the transcription of pro-inflammatory genes [90], through two distinct signaling pathways, the MyD88-dependent and the TRIF-dependent pathway, that leads to the activation of the kinases TBK1 and RIPK1. Some polyphenols, such as Epigallocatechin-3-gallate (EGCG), act on both pathways suppressing the kinase activity of TBK1, a kinase whose activation leads to the production of Interferon type I inflammatory cytokines, as well as inhibiting IKK in murine RAW264.7 macrophages [92]. Other polyphenols, such as luteolin [93] and resveratrol [94], inhibit only the TRIF-dependent pathway through TBK1 inhibition.
Flavonoids have been reported to modulate MAPKs that influence NF-κB activity being involved in pro-inflammatory cytokines cellular production [95].
The mammalian MAPK family consists of three subfamilies: extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinases (JNK), and p38 MAPKs (p38). Some flavonoids, such as quercetin, perform its anti-inflammatory functions by inhibiting ERK and JNK, while JNK and p38 are inhibited from catechin in stimulated THP-1 cells [96], fisetin by attenuating phosphorylation of Akt and p38, but not of ERK and JNK, in dextran sulfate sodium (DSS)-induced colitis in mice [97] and proanthocyanidins acting on JNK, ERK, and PI3K/Akt phosphorylation in a rat hepatic stellate cell line [98]. Finally, quercetin [99], kaempferol [100], luteolin [101], apigenin [102], and EGCG [103] have been reported to function by modulating only one or all the three kinases together.
Nrf2 is the main transcriptional regulator of several detoxification and antioxidant enzymes belonging to the basic leucine zipper transcription factor family, which binds to the antioxidant response element (ARE) of DNA initiating transcription of many Phase II detoxifying and antioxidant genes. Given its function, Nrf2 regulates different processes such as metabolism, inflammation, immunity and autophagy [104]. It has two different localizations: in the cytosol it is bound to its repressor Kelch-like ECH-associated protein (Keap1), which tags Nrf2 for ubiquitination maintaining it at low levels [105]. When cellular oxidative stress occurs, Keap 1 activity is decreased and Nrf2 is free to move to the nucleus where it binds to ARE inducing the transcription of its target genes for detoxifying and antioxidant enzymes, such as superoxide dismutase (SOD), glutathione S-transferase (GST), UDP-glucuronyl transferase (UGT), and NAD(P)H: quinone oxidoreductase (NQO1).
Polyphenols regulate the Nrf2-Keap1-ARE pathway into a Keap1-dependent and independent way [106]. Polyphenols-dependent protection from oxidative stress consists in the induction of conformational changes in cysteine sulfhydryl residues of Keap1 that increase Nrf2 stability and accumulation, preventing its ubiquitination [107]. Resveratrol [108], quercetin [109], baicalein [110,111], xanthohumol [112,113], and others [106] have been shown to in vitro induce Nrf2. Alternatively, polyphenols act in a Keap1 independent way, modifying protein kinases that phosphorylate Nrf2 at different sites, such as Ser40 and Ser408, resulting in stability increase and activation [114,115,116].
In addition, MAPKs can also influence Nrf2 activation. Some polyphenols, such as quercetin [117], resveratrol [118], lycopene [119], luteolin [120], procyanidins [121], anthocyanins [122], hesperidin [123], EGCG [124], and epicatechin [125] seem to be responsible for the upregulation of the Nrf2-ARE pathway through ERK. Quercetin [117], lycopene [119], EGCG [126], and procyanidins [127] through p38 and kaempferol [128] and sappanchalcone [129] through JNK. Finally, resveratrol [118], EGCG [124], epicatechins [125], and procyanidins [127] activate Nrf2-ARE pathway via phosphatidylinositol 3-kinases (PI3K).
5. Foodomics Approach to Investigate the Relationship between Food and Health
In our days food is increasingly considered not only an important energy source but it also believed to play a crucial role in maintaining a body health condition by reducing risk disease. According to Hippocrates’s sentence “Let food be thy medicine and medicine be thy food”, today more than ever it is well known the importance of healthy eating and it can also be demonstrated by highlighting the mechanisms underlying these health effects [132].
Food science, which comprises food chemistry and food microbiology, has made several steps forward in the development of new food products with longer expiration dates, improved processes of production and packaging and better organoleptic characteristics, taking great advantages from the new analytical methods. These new analytical methods are those related to the holistic omics major types of technologies (genomics, transcriptomics, proteomics, and metabolomics) which gave life to a variety of different omics subdisciplines (epigenomics, lipidomics, interactomics, metallomics, diseasomics, etc.) each of them having its own set of tools, techniques, reagents and software and making it easier for the researchers to connect food nutrients, introduced with the diet, to the health and disease of the individual [133].
These technologies differ from each other not only for the laboratory techniques performed but also for the analysis of biological activities of food components, being very helpful to the development of novel biomarkers and to explore and understand how diet could affect gene transcription, protein expression and the entire human metabolism [134].
Foodomics is a scientific discipline that has as its purpose to improve the health of the individual by studying Food and Nutrition domains as a single domain through the omics technologies with nutrigenomics and nutrigenetics being part of it [135].
The genomics approach of Nutrition Science, that studies DNA structure and function, try to clarify how our genome could modify the different individual response to foods whereas the genomics approach of Food Science aims to develop genetically modified harvests and livestock with a higher growth performance, infection/disease resistance, and so a consequent improvement of food nutritional values [136]. So, we can refer to a new type of comprehensive approach called “foodomics approach” allowing to look the problem from the new perspective of using food science to improve human nutrition and its effects on human health. Our body can be influenced by the diet, as an external stimulus, in a positive or negative way, being a protective or a risk factor against certain types of human disease. Similarly, the diet can be influenced by external stimuli because we feed on meat, coming from living organisms, and vegetables which may be affected by breeding and agricultural techniques, respectively. Thus, the foodomics approach help us to relate all these factors to each other with the aim of improving human health.
6. Genomics Approach
Genomics tools include the sequencing, assembling and the analysis of genome structure and function within an organism. Genomics technologies are intended to lead to knowledge of as many genetic sequences as possible through different technologies such as high-density arrays of oligonucleotides or complementary DNAs (cDNAs), a collection of related DNA spots representing single genes attached to a solid surface by covalent or electrostatic binding with suitable chemical matrices. Furthermore, the next-generation sequencing (NGS) technology is able to process millions of sequencing reactions simultaneously without requiring a sequence library and so reducing the sequencing costs [137]. Each plant or farm animal gene composition can be well determined, leading to identify specific marker traits and beneficial alleles responsible for healthier food properties [138]. In fact, genomics technologies allow us to put on a “graphic genotype” which plant growers can use to find inheritable chromosome sections and to identify some beneficial marker traits and gene alleles leading to the selection of crops with greater nutritional and safety characteristics. Furthermore, in single-molecule sequencing, also called third-generation sequencing systems, samples are single DNA molecules that cannot be cloned or amplified during the preparation process [139], allowing high-density single molecules to expand asynchronously and with highly flexible chemical kinetics [140,141].
Genetics is the most common factor causing variability in the response to nutrition, as well as for eye and hair color [142], as in the case of PKU, or lactose intolerance [143], and metabolic syndrome [144], although if, differently from eye and hair color, nowadays is increasingly understood that genetics is unable to fully explain the response to food. Understanding how our genes interact with nutrients may lead to the prescription of tailor-made diets for each individual: for this purpose, nutrigenomics was introduced [145].
7. Transcriptomics Approach
Transcriptomics approach studies all the RNA transcripts, or transcriptome, inside a single cell or a group of cells through gene expression microarrays and large RNA sequencing (RNA-Seq) [145]. This leads to the knowledge of how the global gene expression is modulated by food and the role of single nutrients in inflammation, oxidative stress and cancer prevention [146], with the aim of implementing the strategies of microbial mitigation [147]. Gene expression microarray technology, introduced in the nineties and nowadays more and more widespread [148], it is used for producing new medicines and identifying the presence in the food of hazardous substances or contaminants, such as pesticides, estrogen-like chemicals and dioxins, thus improving its quality and safety [149].
This technology, also used to clarify the mechanisms underlying our response to them, can be performed on solid flat or microchip substrates or on cubic or particle substrates [146]. All the samples coming from different experimental conditions are tested and processed for the relative expression level of each transcript but to date it is still impossible to characterize unknown RNA sequences [150], fact which is overcome using RNA-Seq that allows us to analyze every type of RNA [151]. Finally, it also allows to evaluate how each food component affects the expression of host genes and to study metabolic transformations at molecular levels [150].
8. Proteomics Approach
Proteomics tools consist in the analysis of proteins in food, not only structurally or functionally but also elucidating their interactions and how their structure alterations could modify their role inside our cells [152]. Essentially proteomics techniques are divided into extraction of proteins from cells, enzymatic digestion, molecular separation by two-dimensional electrophoresis (2-DE) or multi-dimensional liquid chromatography (MDLC), and mass spectrometric (MS) analysis [153]. The 2-DE technique isolates and separates proteins thanks to their different isoelectric point (pI) [153] and molecular weight on polyacrylamide gels but the performance is poor and wasting time [154]. MS analysis is mainly divided into matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) and electrospray ion trap (ESI-IT) MS. Proteins are firstly ionized and then measured by MS [155]. Proteomics approach can be divided into that called bottom-up, where proteins, purified or mixed, are digested into peptides and analyzed by MS, or top-down, where unstable proteins are preserved and then analyzed by MS performed on intact proteins [156].
Proteomics technologies, that make use of high-performance separation techniques together with high-resolution MS, are widely exploited in Food Science for monitoring food quality and the presence of microbial contaminants [157] and animal health, allowing identification and characterization of proteins tissue or biological fluids [158].
9. Metabolomic Approach
Current metabolomic technologies are essential in both food and nutrition science [159]. Targeted metabolomics enables us to make precise chemical analyses of up to a thousand of metabolites, to investigate how they are affected by gene mutations causing proteins changes and to characterize metabolic processes that underlie several pathologies with the goal to find new functional food biomarkers, or therapeutic targets, of disease, with a leading role in our metabolism [160].
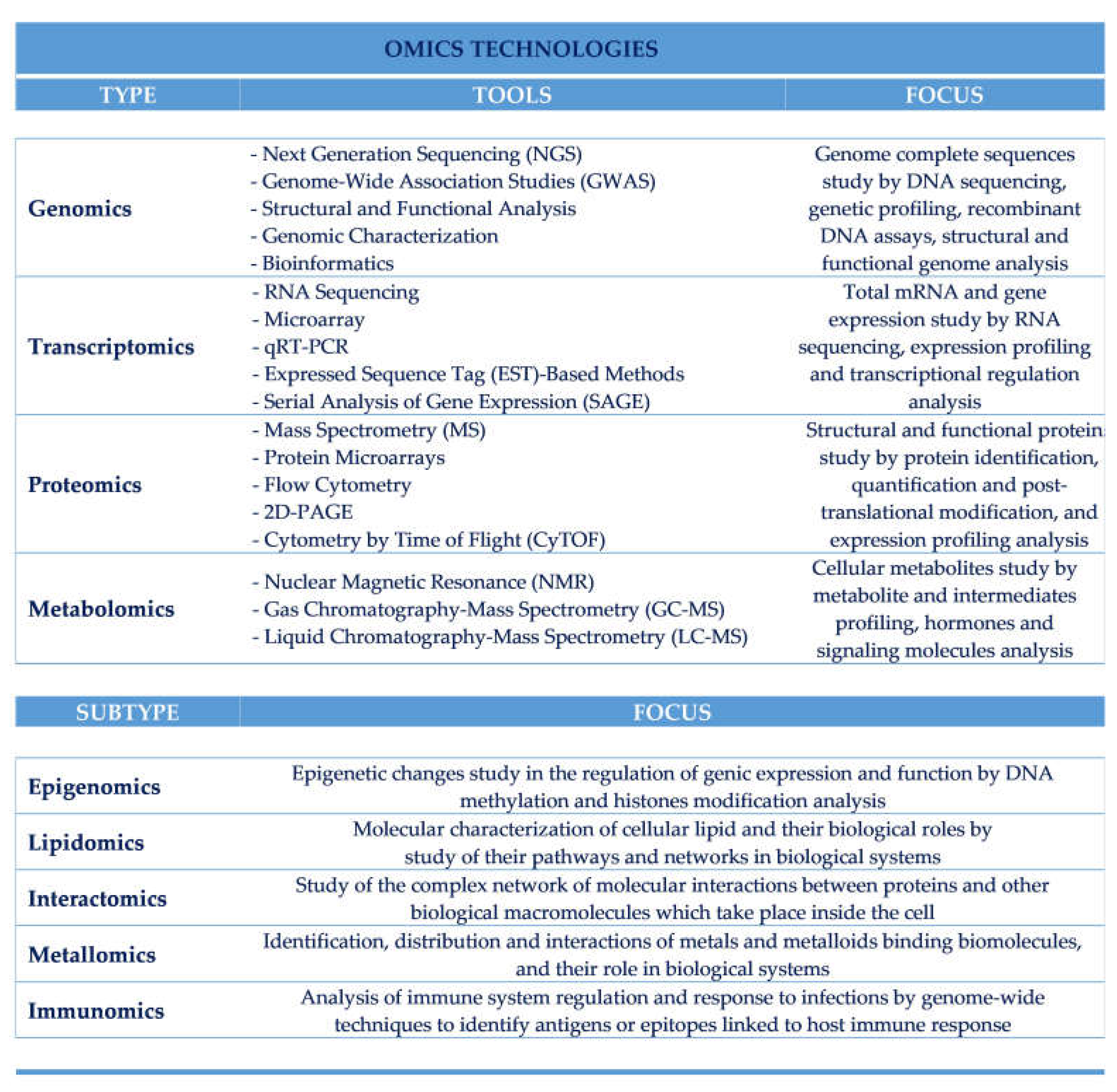
Metabolomics approach employs techniques such as sample preparation, metabolites extraction, sample testing, selection and use of appropriate analytical tools and collection of the data [161]. Metabolomics technologies include liquid and gas chromatography-MS (LC-MS and GC-MS), nuclear magnetic resonance (NMR) and capillary electrophoresis-MS (CE-MS) [162]. Despite having a very low sensitivity, NMR is mostly used for functional food studies aimed at the characterization of active ingredients as well as the effects of different biomarkers [163] through the quantization of metabolites and the analysis of their structure in detail. Requiring small sample size, and simple preparation procedures [164], MS technologies are especially used to characterize and quantify unknown metabolites, being very fast in separating them with high sensitivity and determining the composition and origin of foods during various processes of manufacturing [165]. Unlike CE-MS, which is used to analyze highly polar or charged compounds [160], GC-MS allows to analyze volatile, non-polar and thermally stable compounds very efficiently, with a high yield and reproducibility thanks to the introduction of capillary gas chromatography [166]. In addition, metabolomics technologies are also useful to detect individual metabolic changes and degraded food [167] and, for epidemiological studies in the field of Nutrition Science, to characterize novel biomarkers (Table 1).

Table 1.
Main omics types and subtypes technologies.
10. Trimethylamine-N-Oxide: A Novel Metabolomics-Based Biomarker of Inflammation and IBD Diagnosis
For a long time, trimethylamine-N-oxide (TMAO) was considered only a waste product of the metabolism of choline without any function but, surprisingly, nowadays there are more and more convincing evidence suggesting a positive correlation between TMAO plasma levels and inflammatory diseases, such as atherosclerosis and consequent cardiovascular disease (CVDs), and tumors [168]. TMAO is generated by a two-steps process: the conversion to TMA of the choline introduced with the foods, such as beef liver, eggs, beef, chicken, other types of meat and poultry, milk, beans, edamame, legumes and soy products, mushrooms, fish, cauliflower, broccoli, asparagus, quinoa and avocados, by choline TMA lyase, an enzyme characterizing bacteria from gut microenvironment. Once produced, TMA enters the portal circulation being rapidly oxidized in the liver by hepatic flavin monooxygenases (FMO) forming TMAO and then excreted mainly in the urine [169,170].
FMO3 is the most represented FMO enzyme in humans [171,172]. Genetic deficiency in FMO3 is reported to predispose to fish odor syndrome, a rare condition resulting from the failure to convert TMA to TMAO [173]. We ingest a small amount of TMA with food whereas the most is formed from dietary phosphatidylcholine and carnitine that are metabolized by anaerobes or facultative anaerobes bacteria species colonizing our GT [174], such as Clostridia, Proteus, Shigella, and Aerobacter [169].
Supporting this, Tang et al. recently demonstrated how GT microbiota is responsible for the production of TMAO from phosphatidylcholine introduced with food [175]. After they feed subjects a meal very rich in choline, a so called a “choline challenge”, TMAO plasma levels were measured over an 8-hour period. An increase of TMAO plasma levels was observed already within the first hour of meal intake. When antibiotics for gut microbiota suppression were given at the same subjects before a second “choline challenge” was administered, TMAO plasma levels disappeared, leading them to hypothesize that TMAO plasma concentration is dependent on gut microbiota and alterations of its composition, in terms of bacteria species presence, could lead to its variation. Some metabolites produced by gut microbiota during the degradation of nutrients introduced with the diet [176] are able to trigger an inflammatory response, thus activating the immune system improperly, often causing the onset of inflammation and diseases resulting therefrom [177,178].
Recently, a lot of evidences obtained from vascular inflammation models strongly support the hypothesis that TMAO can induce inflammation and immune regulation by directly inducing the expression of TNFα, NLRP3 inflammasome, mitochondrial ROS, and NF-κB which are critical pro-inflammatory mediators, and down-regulating anti-inflammatory cytokines, such as IL-10 [179,180]. By contrast, high plasma TMAO correlates with improved efficacy of immunotherapy in several types of tumors, boosting anti-tumor immune responses, thereby rendering pancreatic ductal adenocarcinoma (PDAC) [181] more responsive to checkpoint blockade immunotherapy (ICB) (anti-PD1 and/or anti-Tim3) as well as for triple-negative breast cancer (TNBC), probably in the first case potentiating the type-I interferon (IFN) pathway, while in the second activating the endoplasmic reticulum (ER) stress kinase PERK, which induced gasdermin E-mediated pyroptosis in tumor cells and enhanced CD8+ T cell-mediated antitumor immunity in TNBC in vivo. Thus, since the production of TMAO by gut microbiome could be regulated through an increased intake of choline with the food or using TMA lyase inhibitors, it turned out to be an excellent therapeutic target to be induced for the enhancement of the effectiveness of cancer treatment [182,183] and to be inhibited for the treatment of inflammatory diseases, being used also as diagnostic tool, as for IBD.
It is well known that an IBD patient is characterized by the loss of microbial diversity in its inflamed gut[184,185] although it is still unclear how this loss impact on IBD activity, phenotype, and severity. It’s noteworthy that many studies have focused on the alteration of microbiota in disease, but very few are intent to use such alterations as biomarker of diagnosis and progression of disease [186]. Today, IBD diagnosis involves screening for hematological features, such as inflammatory markers and elevated white blood cell count, but is confirmed only through endoscopic examination with multiple biopsies [187].
Based on their findings, Wilson et al. reported that plasma TMAO levels were significantly decreased in IBD patient when compared to non-IBD controls, suggesting that its plasma levels measurement might be clinically relevant not only for IBD treatment but also for its noninvasive diagnosis [188].
11. Gastrointestinal Protective Effects Mediated by Polyphenols: Foodomics-Based Approaches
Today more than ever, omics technologies are a valuable resource for polyphenols identification in foods and for the analysis of the changes that they induce inside our cells at molecular level when introduced with the diet. Thanks to foodomics [187]advanced technologies, today it is easier identifying bioactive compounds, like plant derived polyphenols, and analyze them in fruit, drinks, grain, oil and so on [189]. In the specific, gene-based genomics and transcriptomics can be used to study the interaction between polyphenols and the GT. Gene expression microarray technology is useful to investigate how genes interact with each other after introduction of bioactive compounds with food [190]. Some experimental evidences coming from gene expression microarray technology and reported by Valdès et al. showed that polyphenols from rosemary extract were not able to induce apoptosis in two colon adenocarcinoma cell lines [191]. Transcriptomics was used to evaluate the effects at molecular level of red wine polyphenols on colonic mucosa of F344 rats [192].
The authors, following the analysis of the expression of 5,707 genes, demonstrated that red wine polyphenols improve colonic mucosa functions and own an anti-cancer activity which is expressed through a reduction of oxidative stress, modulation of microbiota composition and down-regulation of genes involved in metabolism, transport and signaling transduction [192]. Transcriptomics studies were also performed by Wang et al. [193] using 16S ribosomal RNA (rRNA) amplicon sequencing (16S-Seq) and Shotgun metagenomic community sequencing (SMC-Seq) to investigate the interaction between green tea polyphenols and the gut microbiota of female Sprague-Dawley (SD) rats treated with green tea polyphenols for 6 months. Bacteroides and energy metabolism related genes in SD rats treated with green tea polyphenols resulted modified in a dose-dependent way, suggesting beneficial effects for consumers.
Other data reported by Yang et al. [194] have showed how a combined action of different polyphenols on mice colitis-related carcinogenesis (CRC) influences gut microbiota diversity and structure and how these changes in turn influence the success of treatment having a regulatory activity on 17 signaling pathways involved in related genes including several biomarkers associated with CRC, such as COX-2, EMR1, PCNA, and caspase-3 which were strongly changed by polyphenols treatment. Also, proteomics was used to clarify the interaction between probiotics and GT and the functional protein changes of probiotics that they induced, as well as to identify stress adaptation markers to better understand some probiotics features into food and also their ability to overcome the GT considering their immunomodulatory activity, colonization and host interaction [195].
A study on the effect of polyphenol-rich rosemary extract on HT-29 human colon cancer cells was reported by Valdes et al. [196], based on nanoliquid chromatography-tandem mass spectrometry (nano-LCMS/MS) in combination with stable isotope dimethyl labeling (DML) technology. The authors found an anti-proliferative effect exerted by this extract characterized by an increase of autophagy. Moreover, several altered proteins found are involved in the activation of Nrf2 and the unfolded protein response (UPR). Other studies used microarray and proteomics approach to evaluate the reduction of intestinal inflammation in a mouse model of IBD induced by green tea extract rich in polyphenols. This extract was able to reduce transcripts and proteins related with immune and inflammatory response pathways as well as increase those related with xenobiotic metabolism pathways. These effects seem to be regulated by Peroxisome Proliferator-Activated Receptor-α (PPAR-α) and signal transducer and activator of transcription 1 (STAT1) [197]. On the other hand, the anti-bacterial activity of catechin on Escherichia coli O157:H7 cell lines in vitro resulted in various changes in the proteins expression related with cell structure and processing of genetic information [198].
Metabolomics is an important part of foodomics, as it allows us to characterize biological functions and phenotypes [199], to target specific metabolic pathways and understand their molecular mechanisms. Therefore, how polyphenols interact with the GT was investigated through metabolomics approaches in terms of analysis of small molecule metabolites. In the specific, the phenolic extract from extra-virgin olive oil (EVOO-PE) was tested for its anti-proliferative effects on SW480 and HT29 human colon cancer cell lines by nano LC-ESITOF-MS technology. Several metabolites of phenolic compounds, especially quercetin and oleuropein aglycones (and their derivatives) were present in the cytoplasm of SW480 and HT29 cells where they induced apoptotic processes [200].
The protective effect of polyphenol-rich bee pollen (BP) extracts was also evaluated using UPLC-QTOF/MS metabolomics in the in vitro model of intestinal barrier Caco-2 cells during the early stages of dextran sulfate sodium (DSS)-induced colitis. Metabolites present in cells treated with BP, probably acting on the glycerophospholipid metabolic pathway, were totally different from those not treated, suggesting that BP suppress inflammation modulating cell metabolism. In particular, BP were able to up-regulate the mRNA expression levels of antioxidant factors such as NQO1, Txnrd1 and Nrf2. In contrast, the mRNA expression of inflammatory factors including TNF-α and IL-6 were down-regulated, in pair to MAPKs signaling inhibition [201]. Furthermore, transcriptomics and proteomics technologies together were used to study the anticarcinogenic effects of quercetin introduced with the diet on the colon mucosa F344 rats, an experimental model of colorectal cancer. Quercetin significantly down-regulated the oncogenic MAPK pathway, up-regulating the expression of cell cycle inhibitors, like MUTYH, and of tumor suppressor genes, such as PTEN, Tp53, and MSh2. Furthermore, dietary quercetin enhanced PPAR-α targets genes, and increased the expression of genes involved in mitochondrial fatty acids (FAs) degradation [202].
In their studies, Di Nunzio et al. showed the anti-inflammatory potential and the effects on the cell metabolome of the olive aqueous extract, in the Caco-2 cell line. In basal and inflamed condition, the olive extract supplementation decreased, not only the pro-inflammatory cytokine IL-8 secretion, but also induced modification in the metabolome leading to a change verse a glucose saving program with subsequent mitigation of appetite following the maintaining of anorexigenic hormone secretion [203]. A multi-omics approach proved to be the most suitable for the understanding of the possible effects of dietary polyphenols during GT disorders, which often involve both molecular and biochemical mechanisms for their metabolization. The omics together could be useful to characterize genes, proteins and metabolites involved in polyphenols metabolism carried out from intestinal microbiota and the anti-inflammatory or anti-proliferative effects that their metabolites have in IBD or GT cancer, respectively. In addition, multiple omics techniques could help to better understand the integrative effects of diet, host, and microbiota in order to develop new therapeutic strategies to use in a personalized nutrition [204].
As this regard transcriptomics, proteomics, and metabolomics technologies (microarray analysis, MALDI-TOF/TOF-MS and CE/LC-MS) were used together to evaluate the effectiveness of the anti-proliferative effects of polyphenols extracted from rosemary on human HT29 colon cancer cells. Based on the combination and comparation of different analytical platforms and in response to molecular changes induced by food ingredients, these studies demonstrated the anti-tumor effects of polyphenols increasing the interest to use an integrative strategy, as foodomics, in the reduction of cancer risk [205]. Finally, a study from Mayta-Apaza et al. demonstrated that polyphenol-rich tart cherries can increase the number of beneficial bacteria in our gut through their metabolites using bacterial fermentation assays performed on concentrate juices or pure polyphenols. 16S rRNA gene sequence and metabolomics experiments in combination showed that polyphenols, in vitro, were metabolized by gut microbiota into 4-hydroxyphenylpropionic acids, leading to an increase of Bacteroides, while in in vivo data to a decrease of Bacteroides and Bifidobacterium [206], suggesting that in vitro and in vivo data obtained using foodomics approach to evaluate the effect of polyphenolic extracts from natural foods or single polyphenolic compounds do not always match. This is because polyphenols digestive process and their metabolization from gut microbiota are very complex and in vitro experiments performed using integrating foodomics on colon cell lines or also in mice models can lead to a reduction of the in vivo physiological variability characterizing their behavior. Polyphenols present in different natural foods vary significantly and further research on their biological activities and studies at multiple molecular levels of genes, proteins, and metabolites are required to fully understand their effect on GT and to assess their effectiveness in the treatment of IBD.
12. Low Bioavailability and Bioaccessibility, Appropriate Doses and Side Effects of Polyphenols
Adequate bioavailability and bioaccessibility are key problems limiting the usage and effectiveness of polyphenols for the treatment of different diseases. The bioaccessibility of polyphenols is associated with the amount of compound accessible for absorption [207] that is different from that introduced by oral administration, depending on the site of the pathological process. In the colon, the distal part of digestive tract is affected during UC and low amounts of polyphenols are delivered, resulting in a low bioaccessibility. An increased bioaccessibility is found for diseases affecting the upper parts of digestive tract, as in CD, mainly small intestine. A low bioavailability also characterizes polyphenols, decreasing from phenolic acids to isoflavones, flavonols, catechins, flavanones, proanthocyanidins, and anthocyanins [208], probably caused by the same factors that reduce their bioaccesibility. An intrarectally administration could be carried out to overcome both problems affecting oral administration, which still remains the preferred and most widely used route of administration for its non-invasiveness, low cost and high patient compliance. Furthermore, polyphenols can also have adverse effects, as reported by some in vivo studies, including nephrotoxicity, hepatotoxicity and iron deficiency, especially at high doses [209], caused by antioxidant enzymes downregulation and a consequent increased production of malondialdehyde and 4-hydroxynonenal in liver and kidney [210].
Moreover, neuroprotective polyphenols used against amyloid β aggregation, own pro-oxidant activity, are able to damage DNA and other macromolecules of normal cells, including endothelial cells, leading them to apoptosis and to CVDs onset [211]. By contrast, low concentrations of polyphenols can prevent hepatic and renal damage [212].
Further studies are therefore necessary to highlight the side effects occurring after administration of polyphenols and determine the most appropriate doses with a pro-health activity and without toxicity. At this regard, to increase their bioavailability and reduce the dose at which they are effective and so their possible side effects, novel bioformulations using polyphenols are being developed for IBD treatment and encapsulation of polyphenolic compounds is strongly recommended, such as for resveratrol-encapsulated microsponges delivered by pectin-based matrix tablets that resulted more therapeutically effective than pure resveratrol in rats with colitis [213] as well as resveratrol delivered by silk fibroin nanoparticles in an experimental model of IBD in rats [214] and β-lactoglobulin nanospheres encapsulating resveratrol that alleviated inflammation in Winnie mice with spontaneous UC [215].
Furthermore, rosmarinic acid-derived nanoparticles conjugated with poly(ethylene glycol), rosmarinic acid-loaded nanovesicles, oleuropein loaded lipid nanocarriers, silica-installed redox nanoparticles with silymarin (compound being flavonolignan), or nanoparticles with curcumin were reported to be effective in lowering colonic inflammation in IBD in vivo [216,217,218] not only modulating the expression of the genes involved in the inflammatory response in a rat model of UC but also improving the mucosal lesions and preserving the distribution of telocytes, interstitial cells with a crucial role in colonic tissue homeostasis [219]. Taken together, these findings suggest that all the strong limitations for the use of polyphenols in the treatment of IBD can be overcome creating encapsulates but little is still known and research into synthesizing polyphenol encapsulates and assessing their utility in IBD are more and more developing.
13. Precision Nutrition and IBD Prevention and Treatment
The exact knowledge and understanding of how nutrients metabolism can affect the response of our body to the diet have been highlighted by many researchers [220]. In this respect, metabolomic helps us to evaluate the impact of specific nutrients introduced with the diet on individual’s health enabling us to characterize new food-derived biomarkers and to understand how the same foods are metabolized differently by different individuals in healthy or unhealthy conditions such as intolerances or allergies to some food components.
Precision nutrition (PN) is a relatively new discipline, and often the term precision nutrition is used instead of personalized nutrition [221] because the two terms overlap, while a distinction between the two is increasingly needed [222].
Biological variability between individuals in response to nutrition is the basis of PN [223]. So, knowing the factors causing this variability and its effect we could predict the outcome and translate it into nutrition advice. This area of nutrition focuses on the effects of the nutrients over the genome, proteome and metabolome [224] trying to clarify how genes expression may be affected by nutrients introduced with the diet, such as polyphenols and other bioactive compounds. Thus, the PN is closely related to a deep “metabolic phenotyping” achieved through omics technologies, including polymorphisms and genetic variants analysis (genomics), DNA methylation and histone modifications analysis (epigenomics), evaluation of microbiota composition (epigenomics), the complete set of RNA transcripts and protein analysis (transcriptomics and proteomics), and the study of chemical processes involving metabolites, intermediates, and products of cell metabolism (metabolomics) (Figure 2).
One of the earliest metabolomics methods centered on the investigation of single nucleotide polymorphisms that affect illnesses specifically linked to the metabolic state. Multiple polymorphisms can be now investigated in a single experiment thanks to modern sequencing technology. Studies being conducted right now evaluate the variations in responses to the same food patterns and concentrate on gene-diet interactions. This implies that many individuals respond differently to the same dietary ingredients (31-34). Analysis of the miRNA-410 gain-of-function mutant polymorphism (rs13702) in the Lipoprotein Lipase (LPL) 3'-untranslated region provides a stunning illustration. A Mediterranean-style diet decreased triglyceride concentrations and the risk of stroke, whereas this did not happen with the control diet, demonstrating a gene-diet interaction (31, 35). Another example concerns the intestinal microbiota, in particular how the Mediterranean diet influences it, modifying both the species present and at the metagenomic level (36). The Mediterranean diet has several systemic advantages, particularly since it contains a lot of antioxidants like vitamin E, β-carotene, vitamin C, and flavonoids, as well as minerals like selenium and natural folate. Dietary antioxidants have been demonstrated to have positive benefits on coronary heart disease in the case-control INTERHEART trial. In fact, a lack of these could change lipoprotein oxidation, which would favor the development of atherosclerotic events. A recent clinical trial also found that extra virgin olive oil supplementation together with a Mediterranean diet reduced levels of circulating oxidized low-density lipoprotein (LDL) and other inflammation-related indicators (31, 37-39). Therefore, nowadays it is increasingly assigned a crucial role to the gene-diet interaction both in maintaining a state of physical well-being and in developing and preventing a pathology. The antioxidants, undiscussed protagonists of the Mediterranean diet, protect us from several chronic, and especially inflammatory, diseases by modulating gene expression [225].
Data related to genome variations and expressed through single nucleotide polymorphisms (SNPs) and other inherited genetic variations are collected and analyzed by nutritional specialists and related to food, lifestyle, and environment which is generally but not primarily [226]. Therefore, IBD patients need PN plans which take into account individual biological, clinical and lifestyle characteristics responsible for different food effects on disease outcome providing “therapeutic diets” able to improve their quality of life.
14. Conclusions
Nowadays it is increasingly object of interest the evaluation of individual response to nutrients, a precision nutrition research initiative, and its association to the IBD management and clinical endpoints improvement. The long-term goal is the creation of predictive models, integrated with quantitative and qualitative measurement of the patient response, for a customized IBD management, with the awareness that diet could be as effective as medical therapy, such as steroids treatment, especially for controlling and limiting inflammation that often characterize IBD patients.
Notably, elimination diet, already used as a first-line therapy for pediatric CD in many countries, resulted effective, safe and inexpensive although it compromises the quality of life [227,228]. So, it might be useful to identify, and eventually eliminate, only those foods that trigger unwanted inflammatory responses in the patient, maximizing the intake of specific nutrients deficient in many patients with CD [229], personalizing it according to the patient’s needed. Clinical trials with polyphenols for IBD are promising, even if their experimental design is complex due to the several mechanisms involved and to the inherent difficulty in observing interactions between host metabolism, the gut microbiome, and the exposome in vivo. Metabolomics provides a comprehensive ‘snapshot’ of small-molecule metabolites and together with microbiome sequencing could help us to elucidate functional changes within our organism. Together, multi-omic integration methods may clarify the relationship between treatment and host, treatment and microbe, and the interactions between host and microbe playing a key role in IBD pathogenesis and response to treatment. These relatively new foodomics approaches may provide critical insight to determine the clinical value of polyphenols for IBD therapy.
Author Contributions
Conceptualization,; methodology,; investigation,; data curation,; software,; writing—original draft preparation,; writing—review and editing, all authors; supervision,. All authors have read and agreed to the published version of the manuscript.
Funding
This work was co-financed by the European Union - PON Research and Innovation 2014-2020 - DM1062/2021 and partially sustained by Finalized Research Funding (FFR 2023), FFR-D03-Di Liberto.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
Figures were created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Caruso, R.; Warner, N.; Inohara, N.; Núñez, G. NOD1 and NOD2: Signaling, Host Defense, and Inflammatory Disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar] [PubMed]
- Zhang, Y.; Zhang, J.; Duan, L. The role of microbiota-mitochondria crosstalk in pathogenesis and therapy of intestinal diseases. Pharmacol. Res. 2022, 186, 106530. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. , Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef]
- Hughes, R.L.; Holscher, H.D. Fueling Gut Microbes: A Review of the Interaction between Diet, Exercise, and the Gut Microbiota in Athletes. Adv. Nutr. Int. Rev. J. 2021, 12, 2190–2215. [Google Scholar] [CrossRef]
- Ding, J.; Ouyang, R.; Zheng, S.; Wang, Y.; Huang, Y.; Ma, X.; Zou, Y.; Chen, R.; Zhuo, Z.; Li, Z.; et al. Effect of Breastmilk Microbiota and Sialylated Oligosaccharides on the Colonization of Infant Gut Microbial Community and Fecal Metabolome. Metabolites 2022, 12, 1136. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Khalili, H.; Konijeti, G.G.; Higuchi, L.M.; de Silva, P.; Korzenik, J.R.; Fuchs, C.S.; Willett, W.C.; Richter, J.M.; Chan, A.T. , A prospective study of long-term intake of dietary fiber and risk of Crohn's disease and ulcerative colitis. Gastroenterology 2013, 145, 970–977. [Google Scholar] [CrossRef]
- Fernando, M.R.; Saxena, A.; Reyes, J.-L.; McKay, D.M. Butyrate enhances antibacterial effects while suppressing other features of alternative activation in IL-4-induced macrophages. Am. J. Physiol. Liver Physiol. 2016, 310, G822–G831. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.G.; Wu, G.D. Diet and the Intestinal Microbiome: Associations, Functions, and Implications for Health and Disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. , Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; D’hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37. [Google Scholar] [CrossRef]
- Fischbach, M.A.; Sonnenburg, J.L. Eating For Two: How Metabolism Establishes Interspecies Interactions in the Gut. Cell Host Microbe 2011, 10, 336–347. [Google Scholar] [CrossRef]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1beta through HIF-1alpha. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef]
- Hosomi, K.; Kiyono, H.; Kunisawa, J. Fatty acid metabolism in the host and commensal bacteria for the control of intestinal immune responses and diseases. Gut Microbes 2019, 11, 276–284. [Google Scholar] [CrossRef]
- Hartel, J.C.; Merz, N.; Grösch, S. How sphingolipids affect T cells in the resolution of inflammation. Front. Pharmacol. 2022, 13, 1002915. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Thomson, J.M.; Flint, H.J. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009, 11, 2112–2122. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, F.R. Intestinal Barrier in Human Health and Disease. Int. J. Environ. Res. Public Health 2021, 18, 12836. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, B.; La Manna, M.P.; La Barbera, L.; Mohammadnezhad, L.; Badami, G.D.; Azgomi, M.S.; Dieli, F.; Caccamo, N. Immunity and Nutrition: The Right Balance in Inflammatory Bowel Disease. Cells 2022, 11, 455. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Ferrante, A.; Guggino, G.; Triolo, G. The role of the gastrointestinal tract in the pathogenesis of rheumatic diseases. Best Pr. Res. Clin. Rheumatol. 2016, 30, 889–900. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Yu, Y.R.; Rodriguez, J.R. Clinical presentation of Crohn’s, ulcerative colitis, and indeterminate colitis: Symptoms, extraintestinal manifestations, and disease phenotypes. Semin. Pediatr. Surg. 2017, 26, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Gastroenterology, H.N.A.S.F.P.; America, C.F.O.; Bousvaros, A.; A Antonioli, D.; Colletti, R.B.; Dubinsky, M.C.; Glickman, J.N.; Gold, B.D.; Griffiths, A.M.; Jevon, G.P.; et al. Differentiating Ulcerative Colitis from Crohn Disease in Children and Young Adults: Report of a Working Group of the North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition and the Crohn's and Colitis Foundation of America. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 653–674. [Google Scholar] [CrossRef]
- Segal, J.P.; LeBlanc, J.-F.; Hart, A.L. Ulcerative colitis: An update. Clin. Med. 2021, 21, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Hibi, T.; Ogata, H. Novel pathophysiological concepts of inflammatory bowel disease. J. Gastroenterol. 2006, 41, 10–16. [Google Scholar] [CrossRef]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. , Ulcerative colitis. Nat. Rev. Dis. Primers 2020, 6, 74. [Google Scholar] [CrossRef]
- Petronis, A.; Petroniene, R. Epigenetics of inflammatory bowel disease. Gut 2000, 47, 302–306. [Google Scholar] [CrossRef]
- Ji, Y.; Yang, Y.; Sun, S.; Dai, Z.; Ren, F.; Wu, Z. Insights into diet-associated oxidative pathomechanisms in inflammatory bowel disease and protective effects of functional amino acids. Nutr. Rev. 2022, 81, 95–113. [Google Scholar] [CrossRef]
- Emanuele, S.; D’anneo, A.; Calvaruso, G.; Cernigliaro, C.; Giuliano, M.; Lauricella, M. The Double-Edged Sword Profile of Redox Signaling: Oxidative Events As Molecular Switches in the Balance between Cell Physiology and Cancer. Chem. Res. Toxicol. 2018, 31, 201–210. [Google Scholar] [CrossRef]
- Dodson, M.; Castro-Portuguez, R.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef]
- Li, H.; Christman, L.M.; Li, R.; Gu, L. Synergic interactions between polyphenols and gut microbiota in mitigating inflammatory bowel diseases. Food Funct. 2020, 11, 4878–4891. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef]
- Nagao-Kitamoto, H.; Kamada, N. Host-microbial Cross-talk in Inflammatory Bowel Disease. Immune Netw. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Khorsand, B.; Aghdaei, H.A.; Nazemalhosseini-Mojarad, E.; Nadalian, B.; Nadalian, B.; Houri, H. Overrepresentation of Enterobacteriaceae and Escherichia coli is the major gut microbiome signature in Crohn’s disease and ulcerative colitis; a comprehensive metagenomic analysis of IBDMDB datasets. Front. Cell. Infect. Microbiol. 2022, 12, 1015890. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, M.; Zirnbauer, R.; Schlager, S.; Mertens, D.; Gasche, N.; Sladek, B.; Herbold, C.; Bochkareva, O.; Emelianenko, V.; Vogelsang, H.; et al. coli are associated with disease activity in ulcerative colitis. Gut Microbes 2022, 14, 2143218. [Google Scholar] [CrossRef] [PubMed]
- Chervy, M.; Sivignon, A.; Dambrine, F.; Buisson, A.; Sauvanet, P.; Godfraind, C.; Allez, M.; Le Bourhis, L.; The Remind Group; Barnich, N. ; et al. Epigenetic master regulators HDAC1 and HDAC5 control pathobiont Enterobacteria colonization in ileal mucosa of Crohn’s disease patients. Gut Microbes 2022, 14, 2127444. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Gu, Y.; Jiang, T.; Che, H. Inhibition effect of PPAR-gamma signaling on mast cell-mediated allergic inflammation through down-regulation of PAK1/ NF-kappaB activation. Int. Immunopharmacol. 2022, 108, 108692. [Google Scholar] [CrossRef]
- Wang, L.; Hu, Y.; Song, B.; Xiong, Y.; Wang, J.; Chen, D. Targeting JAK/STAT signaling pathways in treatment of inflammatory bowel disease. Inflamm. Res. 2021, 70, 753–764. [Google Scholar] [CrossRef]
- Tsopmejio, I.S.N.; Yuan, J.; Diao, Z.; Fan, W.; Wei, J.; Zhao, C.; Li, Y.; Song, H. Auricularia polytricha and Flammulina velutipes reduce liver injury in DSS-induced Inflammatory Bowel Disease by improving inflammation, oxidative stress, and apoptosis through the regulation of TLR4/NF-κB signaling pathways. J. Nutr. Biochem. 2023, 111, 109190. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 2681. [Google Scholar] [CrossRef] [PubMed]
- Vulliemoz, M.; Brand, S.; Juillerat, P.; Mottet, C.; Ben-Horin, S.; Michetti, P. TNF-Alpha Blockers in Inflammatory Bowel Diseases: Practical Recommendations and a User’s Guide: An Update. Digestion 2020, 101, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Piodi, L.P.; Poloni, A.; Ulivieri, F.M. , Managing osteoporosis in ulcerative colitis: Something new? World J. Gastroenterol. 2014, 20, 14087–14098. [Google Scholar] [CrossRef] [PubMed]
- Park, K.T.; Bass, D. , Inflammatory bowel disease-attributable costs and cost-effective strategies in the United States: A review. Inflamm. Bowel. Dis. 2011, 17, 1603–1609. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Bin Dukhyil, A.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Metabolic Fate of Orally Ingested Proanthocyanidins through the Digestive Tract. Antioxidants 2022, 12, 17. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Lamuela-Raventos, R. ; Ray, S.; Micek, A.; Pajak, A.; Sciacca, S.; D'Orazio, N.; Del Rio, D.; Galvano, F. A comprehensive meta-analysis on dietary flavonoid and lignan intake and cancer risk: Level of evidence and limitations. Mol. Nutr. Food Res. 2016, 61. [Google Scholar] [CrossRef]
- De Blasio, A.; D’anneo, A.; Lauricella, M.; Emanuele, S.; Giuliano, M.; Pratelli, G.; Calvaruso, G.; Carlisi, D. The Beneficial Effects of Essential Oils in Anti-Obesity Treatment. Int. J. Mol. Sci. 2021, 22, 11832. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, G.; Di Liberto, D.; Carlisi, D.; Emanuele, S.; Giuliano, M.; Notaro, A.; De Blasio, A.; Calvaruso, G.; D’anneo, A.; Lauricella, M. Hypertrophy and ER Stress Induced by Palmitate Are Counteracted by Mango Peel and Seed Extracts in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2023, 24, 5419. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, G.; Carlisi, D.; D’anneo, A.; Maggio, A.; Emanuele, S.; Piccionello, A.P.; Giuliano, M.; De Blasio, A.; Calvaruso, G.; Lauricella, M. Bio-Waste Products of Mangifera indica L. Reduce Adipogenesis and Exert Antioxidant Effects on 3T3-L1 Cells. Antioxidants 2022, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Barbaresko, J.; Oluwagbemigun, K.; Schmid, M.; Nöthlings, U. Polyphenol exposure and risk of type 2 diabetes: Dose-response meta-analyses and systematic review of prospective cohort studies. Am. J. Clin. Nutr. 2018, 108, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Stepaniak, U.; Micek, A.; Stefler, D.; Bobak, M.; Pająk, A. Dietary polyphenols are inversely associated with metabolic syndrome in Polish adults of the HAPIEE study. Eur. J. Nutr. 2016, 56, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zamora-Ros, R.; Chan, S.; Cross, A.J.; Ward, H.; Jakszyn, P.; Luben, R.; Opstelten, J.L.; Oldenburg, B.; Hallmans, G.; et al. Dietary Polyphenols in the Aetiology of Crohn's Disease and Ulcerative Colitis-A Multicenter European Prospective Cohort Study (EPIC). Inflamm. Bowel. Dis. 2017, 23, 2072–2082. [Google Scholar] [CrossRef] [PubMed]
- Larussa, T.; Oliverio, M.; Suraci, E.; Greco, M.; Placida, R.; Gervasi, S.; Marasco, R.; Imeneo, M.; Paolino, D.; Tucci, L.; et al. Oleuropein Decreases Cyclooxygenase-2 and Interleukin-17 Expression and Attenuates Inflammatory Damage in Colonic Samples from Ulcerative Colitis Patients. Nutrients 2017, 9, 391. [Google Scholar] [CrossRef]
- Vezza, T.; Algieri, F.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Utrilla, M.P.; Talhaoui, N.; Gómez-Caravaca, A.M.; Segura-Carretero, A.; Rodríguez-Cabezas, M.E.; Monteleone, G.; et al. Immunomodulatory properties of Olea europaea leaf extract in intestinal inflammation. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Daniel, K.; Vitetta, L.; Singh, M.A.F. Effects of olives and their constituents on the expression of ulcerative colitis: A systematic review of randomised controlled trials. Br. J. Nutr. 2021, 127, 1153–1171. [Google Scholar] [CrossRef]
- Singla, V.; Pratap Mouli, V.; Garg, S.K.; Rai, T.; Choudhury, B.N.; Verma, P.; Deb, R.; Tiwari, V.; Rohatgi, S.; Dhingra, R.; et al. Induction with NCB-02 (curcumin) enema for mild-to-moderate distal ulcerative colitis - a randomized, placebo-controlled, pilot study. J. Crohns Colitis 2014, 8, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Salomon, N.; Wu, J.C.; Kopylov, U.; Lahat, A.; Har-Noy, O.; Ching, J.Y.; Cheong, P.K.; Avidan, B.; Gamus, D.; et al. Curcumin in Combination With Mesalamine Induces Remission in Patients With Mild-to-Moderate Ulcerative Colitis in a Randomized Controlled Trial. Clin. Gastroenterol. Hepatol. 2015, 13, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Kedia, S.; Bhatia, V.; Thareja, S.; Garg, S.; Mouli, V.P.; Bopanna, S.; Tiwari, V.; Makharia, G.; Ahuja, V. Low dose oral curcumin is not effective in induction of remission in mild to moderate ulcerative colitis: Results from a randomized double blind placebo controlled trial. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 147–154. [Google Scholar] [CrossRef]
- Masoodi, M.; Mahdiabadi, M.A.; Mokhtare, M.; Agah, S.; Kashani, A.H.F.; Rezadoost, A.M.; Sabzikarian, M.; Talebi, A.; Sahebkar, A. The efficacy of curcuminoids in improvement of ulcerative colitis symptoms and patients’ self-reported well-being: A randomized double-blind controlled trial. J. Cell. Biochem. 2018, 119, 9552–9559. [Google Scholar] [CrossRef]
- Sadeghi, N.; Mansoori, A.; Shayesteh, A.; Hashemi, S.J. The effect of curcumin supplementation on clinical outcomes and inflammatory markers in patients with ulcerative colitis. Phytotherapy Res. 2019, 34, 1123–1133. [Google Scholar] [CrossRef]
- Samsami-Kor, M.; Daryani, N.E.; Asl, P.R.; Hekmatdoost, A. Anti-Inflammatory Effects of Resveratrol in Patients with Ulcerative Colitis: A Randomized, Double-Blind, Placebo-controlled Pilot Study. Arch. Med Res. 2015, 46, 280–285. [Google Scholar] [CrossRef]
- Samsamikor, M.; Daryani, N.E.; Asl, P.R.; Hekmatdoost, A. Resveratrol Supplementation and Oxidative/Anti-Oxidative Status in Patients with Ulcerative Colitis: A Randomized, Double-Blind, Placebo-controlled Pilot Study. Arch. Med Res. 2016, 47, 304–309. [Google Scholar] [CrossRef]
- Rastegarpanah, M.; Malekzadeh, R.; Vahedi, H.; Mohammadi, M.; Elahi, E.; Chaharmahali, M.; Safarnavadeh, T.; Abdollahi, M. A randomized, double blinded, placebo-controlled clinical trial of silymarin in ulcerative colitis. Chin. J. Integr. Med. 2012, 21, 902–906. [Google Scholar] [CrossRef]
- Hanai, H.; Iida, T.; Takeuchi, K.; Watanabe, F.; Maruyama, Y.; Andoh, A.; Tsujikawa, T.; Fujiyama, Y.; Mitsuyama, K.; Sata, M.; et al. Curcumin Maintenance Therapy for Ulcerative Colitis: Randomized, Multicenter, Double-Blind, Placebo-Controlled Trial. Clin. Gastroenterol. Hepatol. 2006, 4, 1502–1506. [Google Scholar] [CrossRef]
- Morvaridi, M.; Jafarirad, S.; Seyedian, S.S.; Alavinejad, P.; Cheraghian, B. The effects of extra virgin olive oil and canola oil on inflammatory markers and gastrointestinal symptoms in patients with ulcerative colitis. Eur. J. Clin. Nutr. 2020, 74, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Venancio, V.P.; Fang, C.; Dupont, A.W.; Talcott, S.T.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr. Res. 2020, 75, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Peng, P.; Ding, N.; Jia, W.; Huang, C.; Tang, Y. Oxidative Stress, Inflammation, Gut Dysbiosis: What Can Polyphenols Do in Inflammatory Bowel Disease? Antioxidants 2023, 12, 967. [Google Scholar] [CrossRef] [PubMed]
- De Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.; Cassidy, A.; Kay, C. The pharmacokinetics of anthocyanins and their metabolites in humans. J. Cereb. Blood Flow Metab. 2014, 171, 3268–3282. [Google Scholar] [CrossRef]
- Crescenti, A.; Caimari, A.; Alcaide-Hidalgo, J.M.; Marine-Casado, R.; Valls, R.M.; Companys, J.; Salamanca, P.; Calderon-Perez, L.; Pla-Paga, L.; Pedret, A.; et al. Hesperidin Bioavailability Is Increased by the Presence of 2S-Diastereoisomer and Micronization-A Randomized, Crossover and Double-Blind Clinical Trial. Nutrients 2022, 14, 2481. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Liu, Y.; Kang, L.; Ye, H.; Zang, J.; Wang, J.; Han, D. Bifidobacterium animalis Promotes the Growth of Weaning Piglets by Improving Intestinal Development, Enhancing Antioxidant Capacity, and Modulating Gut Microbiota. Appl. Environ. Microbiol. 2022, 88, e0129622. [Google Scholar] [CrossRef]
- Fan, F.-Y.; Sang, L.-X.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, C. ; Abdullah; Tian, W. ; Qiu, Z.; Song, M.; Cao, Y.; Xiao, J., Hydroxytyrosol Alleviates Dextran Sulfate Sodium-Induced Colitis by Modulating Inflammatory Responses, Intestinal Barrier, and Microbiome. J. Agric. Food Chem. 2022, 70, 2241–2252. [Google Scholar] [PubMed]
- Astorga, J.; Gasaly, N.; Dubois-Camacho, K.; De la Fuente, M.; Landskron, G.; Faber, K.N.; Urra, F.A.; Hermoso, M.A. The role of cholesterol and mitochondrial bioenergetics in activation of the inflammasome in IBD. Front. Immunol. 2022, 13, 1028953. [Google Scholar] [CrossRef]
- Hegazy, S.K.; El-Bedewy, M.M. , Effect of probiotics on pro-inflammatory cytokines and NF-kappaB activation in ulcerative colitis. World J. Gastroenterol. 2010, 16, 4145–4451. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.D.; Goodlad, J.R.; Limb, G.A.; Powell, J.J.; Thompson, R.P.; Punchard, N.A. , Activation of nuclear factor kappa B in Crohn's disease. Inflamm. Res. 1998, 47, 440–445. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. , Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar]
- Cho, D.-I.; Koo, N.-Y.; Chung, W.J.; Kim, T.-S.; Ryu, S.Y.; Im, S.Y.; Kim, K.-M. Effects of resveratrol-related hydroxystilbenes on the nitric oxide production in macrophage cells: Structural requirements and mechanism of action. Life Sci. 2002, 71, 2071–2082. [Google Scholar] [CrossRef]
- Cho, M.K.; Park, J.W.; Jang, Y.P.; Kim, Y.C.; Kim, S.G. , Potent inhibition of lipopolysaccharide-inducible nitric oxide synthase expression by dibenzylbutyrolactone lignans through inhibition of I-kappaBalpha phosphorylation and of p65 nuclear translocation in macrophages. Int. Immunopharmacol. 2002, 2, 105–116. [Google Scholar] [CrossRef]
- Kennedy, M.; Wilson, L.; Szabo, C.; Salzman, A.L. 5-aminosalicylic acid inhibits iNOS transcription in human intestinal epithelial cells. Int. J. Mol. Med. 1999, 4, 437–480. [Google Scholar] [CrossRef]
- Comalada, M.; Ballester, I.; Bailón, E.; Sierra, S.; Xaus, J.; Gálvez, J.; de Medina, F.S.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mouse macrophages by naturally occurring flavonoids: Analysis of the structure–activity relationship. Biochem. Pharmacol. 2006, 72, 1010–1021. [Google Scholar] [CrossRef]
- Nicholas, C.; Batra, S.; Vargo, M.A.; Voss, O.H.; Gavrilin, M.A.; Wewers, M.D.; Guttridge, D.C.; Grotewold, E.; Doseff, A.I. , Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory cytokines expression by inactivating NF-kappaB through the suppression of p65 phosphorylation. J. Immunol. 2007, 179, 7121–7127. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Aggarwal, R.S.; Sethi, G.; Aggarwal, B.B.; Ramesh, G.T. , Morin (3,5,7,2',4'-Pentahydroxyflavone) abolishes nuclear factor-kappaB activation induced by various carcinogens and inflammatory stimuli, leading to suppression of nuclear factor-kappaB-regulated gene expression and up-regulation of apoptosis. Clin. Cancer Res. 2007, 13, 2290–2297. [Google Scholar] [CrossRef]
- Sung, B.; Pandey, M.K.; Aggarwal, B.B. Fisetin, an inhibitor of cyclin-dependent kinase 6, down-regulates nuclear factor-kappaB-regulated cell proliferation, antiapoptotic and metastatic gene products through the suppression of TAK-1 and receptor-interacting protein-regulated IkappaBalpha kinase activation. Mol. Pharmacol. 2007, 71, 1703–1714. [Google Scholar]
- Kunnumakkara, A.B.; Nair, A.S.; Ahn, K.S.; Pandey, M.K.; Yi, Z.; Liu, M.; Aggarwal, B.B. Gossypin, a pentahydroxy glucosyl flavone, inhibits the transforming growth factor beta-activated kinase-1-mediated NF-kappaB activation pathway, leading to potentiation of apoptosis, suppression of invasion, and abrogation of osteoclastogenesis. Blood 2007, 109, 5112–5121. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Peng, W.H.; Tsai, K.D.; Hsu, S.L. Luteolin suppresses inflammation-associated gene expression by blocking NF-kappaB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007, 81, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Zhang, J.F.; Guo, B.S.; He, Q.; Wang, B.Y.; He, B.; Wang, C.Q. , Resveratrol inhibits macrophage expression of EMMPRIN by activating PPARgamma. Vascul Pharmacol. 2007, 46, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, H.S.; Giang, P.M.; Jin, X.; Lee, S.; Son, P.T.; Lee, D.; Hong, Y.S.; Lee, K.; Lee, J.J. , Blockade of nuclear factor-kappaB signaling pathway and anti-inflammatory activity of cardamomin, a chalcone analog from Alpinia conchigera. J. Pharmacol. Exp. Ther. 2006, 316, 271–278. [Google Scholar] [CrossRef]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The Toll-Like Receptor 2 Pathway Establishes Colonization by a Commensal of the Human Microbiota. Science 2011, 332, 974–977. [Google Scholar] [CrossRef]
- Brown, M.; Hughes, K.R.; Moossavi, S.; Robins, A.; Mahida, Y.R. Toll-like receptor expression in crypt epithelial cells, putative stem cells and intestinal myofibroblasts isolated from controls and patients with inflammatory bowel disease. Clin. Exp. Immunol. 2014, 178, 28–39. [Google Scholar] [CrossRef]
- Youn, H.S.; Lee, J.Y.; Saitoh, S.I.; Miyake, K.; Kang, K.W.; Choi, Y.J.; Hwang, D.H. , Suppression of MyD88- and TRIF-dependent signaling pathways of Toll-like receptor by (-)-epigallocatechin-3-gallate, a polyphenol component of green tea. Biochem. Pharmacol. 2006, 72, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kim, S.Y.; Kim, Y.S.; Lee, W.-H.; Hwang, D.H.; Lee, J.Y. Suppression of the TRIF-dependent signaling pathway of Toll-like receptors by luteolin. Biochem. Pharmacol. 2009, 77, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.S.; Lee, J.Y.; Fitzgerald, K.A.; Young, H.A.; Akira, S.; Hwang, D.H. Specific Inhibition of MyD88-Independent Signaling Pathways of TLR3 and TLR4 by Resveratrol: Molecular Targets Are TBK1 and RIP1 in TRIF Complex. J. Immunol. 2005, 175, 3339–3346. [Google Scholar] [CrossRef]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. , Regulation of NF-kappa B activation by MAP kinase cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Huang, S.; Wu, C.; Yen, G. Effects of flavonoids on the expression of the pro-inflammatory response in human monocytes induced by ligation of the receptor for AGEs. Mol. Nutr. Food Res. 2006, 50, 1129–1139. [Google Scholar] [CrossRef]
- Sahu, B.D.; Kumar, J.M.; Sistla, R. , Fisetin, a dietary flavonoid, ameliorates experimental colitis in mice: Relevance of NF-kappaB signaling. J. Nutr. Biochem. 2016, 28, 171–82. [Google Scholar] [CrossRef]
- Jiang, M.; Wu, Y.L.; Li, X.; Zhang, Y.; Xia, K.L.; Cui, B.W.; Lian, L.H.; Nan, J.X. , Oligomeric proanthocyanidin derived from grape seeds inhibited NF-kappaB signaling in activated HSC: Involvement of JNK/ERK MAPK and PI3K/Akt pathways. Biomed. Pharmacother. 2017, 93, 674–680. [Google Scholar] [CrossRef]
- Cho, S.Y.; Park, S.J.; Kwon, M.J.; Jeong, T.S.; Bok, S.H.; Choi, W.Y.; Jeong, W.I.; Ryu, S.Y.; Do, S.H.; Lee, C.S.; et al. Quercetin suppresses proinflammatory cytokines production through MAP kinases andNF-kappaB pathway in lipopolysaccharide-stimulated macrophage. Mol. Cell Biochem. 2003, 243, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Jan, R.-L.; Kuo, C.-H.; Chu, Y.-T.; Wang, W.-L.; Lee, M.-S.; Chen, H.-N.; Hung, C.-H. Natural Flavone Kaempferol Suppresses Chemokines Expression in Human Monocyte THP-1 Cells through MAPK Pathways. J. Food Sci. 2010, 75, H254–H259. [Google Scholar] [CrossRef]
- Xagorari, A.; Roussos, C.; Papapetropoulos, A. Inhibition of LPS-stimulated pathways in macrophages by the flavonoid luteolin. Br. J. Pharmacol. 2002, 136, 1058–1064. [Google Scholar] [CrossRef]
- Masuelli, L.; Marzocchella, L.; Quaranta, A.; Palumbo, C.; Pompa, G.; Izzi, V.; Canini, A.; Modesti, A.; Galvano, F.; Bei, R. Apigenin induces apoptosis and impairs head and neck carcinomas EGFR/ErbB2 signaling. Front. Biosci. 2011, 16, 1060–8. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Choi, J.-S.; Choi, Y.-J.; Shin, S.-Y.; Kang, S.-W.; Han, S.J.; Kang, Y.-H. (−)Epigallocatechin gallate hampers collagen destruction and collagenase activation in ultraviolet-B-irradiated human dermal fibroblasts: Involvement of mitogen-activated protein kinase. Food Chem. Toxicol. 2008, 46, 1298–1307. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef]
- Qin, S.; Hou, D.X. , Multiple regulations of Keap1/Nrf2 system by dietary phytochemicals. Mol Nutr Food Res 2016, 60, 1731–1755. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Revel, J.S.; Maier, C.S.; Rada, P.; Pardo, V.; Mobasher, M.A.; Martínez, I.G.; Ruiz, L.; González-Rodríguez. ; Sanchez-Ramos, C.; et al. Mitochondria-Centric Review of Polyphenol Bioactivity in Cancer Models. Antioxidants Redox Signal. 2018, 29, 1589–1611. [Google Scholar] [CrossRef]
- Kode, A.; Rajendrasozhan, S.; Caito, S.; Yang, S.-R.; Megson, I.L.; Rahman, I. Resveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, L478–L488. [Google Scholar] [CrossRef]
- Tanigawa, S.; Fujii, M.; Hou, D.-X. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free. Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Chen, J.; Tanigawa, S.; Hou, D.-X. Gene expression profiling and pathway network analysis of hepatic metabolic enzymes targeted by baicalein. J. Ethnopharmacol. 2012, 140, 131–140. [Google Scholar] [CrossRef]
- Qin, S.; Deng, F.; Wu, W.; Jiang, L.; Yamashiro, T.; Yano, S.; Hou, D.X. , Baicalein modulates Nrf2/Keap1 system in both Keap1-dependent and Keap1-independent mechanisms. Arch. Biochem. Biophys. 2014, 559, 53–61. [Google Scholar] [CrossRef]
- Dietz, B.M.; Kang, Y.-H.; Liu, G.; Eggler, A.L.; Yao, P.; Chadwick, L.R.; Pauli, G.F.; Farnsworth, N.R.; Mesecar, A.D.; van Breemen, R.B.; et al. Xanthohumol Isolated from Humulus lupulus Inhibits Menadione-Induced DNA Damage through Induction of Quinone Reductase. Chem. Res. Toxicol. 2005, 18, 1296–1305. [Google Scholar] [CrossRef]
- Luo, Y.; Eggler, A.L.; Liu, D.; Liu, G.; Mesecar, A.D.; van Breemen, R.B. Sites of alkylation of human Keap1 by natural chemoprevention agents. J. Am. Soc. Mass Spectrom. 2007, 18, 2226–2232. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Nguyen, T.; Pickett, C.B. Phosphorylation of Nrf2 at Ser-40 by Protein Kinase C Regulates Antioxidant Response Element-mediated Transcription. J. Biol. Chem. 2002, 277, 42769–42774. [Google Scholar] [CrossRef]
- Nguyen, T.; Sherratt, P.J.; Huang, H.-C.; Yang, C.S.; Pickett, C.B. Increased Protein Stability as a Mechanism That Enhances Nrf2-mediated Transcriptional Activation of the Antioxidant Response Element. J. Biol. Chem. 2003, 278, 4536–4541. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Yang, C.S.; Pickett, C.B. The pathways and molecular mechanisms regulating Nrf2 activation in response to chemical stress. Free. Radic. Biol. Med. 2004. [Google Scholar] [CrossRef]
- Yao, P.; Nussler, A.; Liu, L.; Hao, L.; Song, F.; Schirmeier, A.; Nussler, N. Quercetin protects human hepatocytes from ethanol-derived oxidative stress by inducing heme oxygenase-1 via the MAPK/Nrf2 pathways. J. Hepatol. 2007, 47, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Jang, J.-H.; Li, M.-H.; Surh, Y.-J. Resveratrol upregulates heme oxygenase-1 expression via activation of NF-E2-related factor 2 in PC12 cells. Biochem. Biophys. Res. Commun. 2005, 331, 993–1000. [Google Scholar] [CrossRef]
- Yang, C.-M.; Huang, S.-M.; Liu, C.-L.; Hu, M.-L. Apo-8′-lycopenal Induces Expression of HO-1 and NQO-1 via the ERK/p38-Nrf2-ARE Pathway in Human HepG2 Cells. J. Agric. Food Chem. 2012, 60, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Wu, M.-J.; Liu, I.Y.-C.; Su, J.-D.; Yen, J.-H. Neurotrophic and Cytoprotective Action of Luteolin in PC12 Cells through ERK-Dependent Induction of Nrf2-Driven HO-1 Expression. J. Agric. Food Chem. 2010, 58, 4477–4486. [Google Scholar] [CrossRef]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M. . Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2011, 51, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.P.; Choi, J.H.; Choi, J.M.; Chung, Y.C.; Jeong, H.G. Protective mechanisms of anthocyanins from purple sweet potato against tert-butyl hydroperoxide-induced hepatotoxicity. Food Chem. Toxicol. 2011, 49, 2081–2089. [Google Scholar] [CrossRef]
- Chen, M.-C.; Ye, Y.-Y.; Ji, G.; Liu, J.-W. Hesperidin Upregulates Heme Oxygenase-1 To Attenuate Hydrogen Peroxide-Induced Cell Damage in Hepatic L02 Cells. J. Agric. Food Chem. 2010, 58, 3330–3335. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Hsu, M.; Hsieh, C.; Lin, J.; Lai, P.; Wung, B. Upregulation of heme oxygenase-1 by Epigallocatechin-3-gallate via the phosphatidylinositol 3-kinase/Akt and ERK pathways. Life Sci. 2006, 78, 2889–2897. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martín, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin induces NF-κB, activator protein-1 (AP-1) and nuclear transcription factor erythroid 2p45-related factor-2 (Nrf2) via phosphatidylinositol-3-kinase/protein kinase B (PI3K/AKT) and extracellular regulated kinase (ERK) signalling in HepG2 cells. Br. J. Nutr. 2009, 103, 168–179. [Google Scholar] [CrossRef]
- Pullikotil, P.; Chen, H.; Muniyappa, R.; Greenberg, C.C.; Yang, S.; Reiter, C.E.; Lee, J.-W.; Chung, J.H.; Quon, M.J. Epigallocatechin gallate induces expression of heme oxygenase-1 in endothelial cells via p38 MAPK and Nrf-2 that suppresses proinflammatory actions of TNF-α. J. Nutr. Biochem. 2011, 23, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.-J.; Jun, M.; Jeong, W.-S. Procyanidins from Wild Grape (Vitis amurensis) Seeds Regulate ARE-Mediated Enzyme Expression via Nrf2 Coupled with p38 and PI3K/Akt Pathway in HepG2 Cells. Int. J. Mol. Sci. 2012, 13, 801–818. [Google Scholar] [CrossRef]
- Gao, S.S.; Choi, B.-M.; Chen, X.Y.; Zhu, R.Z.; Kim, Y.; So, H.; Park, R.; Sung, M.; Kim, B.-R. Kaempferol Suppresses Cisplatin-Induced Apoptosis Via Inductions of Heme Oxygenase-1 and Glutamate-Cysteine Ligase Catalytic Subunit in HEI-OC1 cells. Pharm. Res. 2009, 27, 235–245. [Google Scholar] [CrossRef]
- Jeong, G.-S.; Lee, D.-S.; Li, B.; Lee, H.-J.; Kim, E.-C.; Kim, Y.-C. Effects of sappanchalcone on the cytoprotection and anti-inflammation via heme oxygenase-1 in human pulp and periodontal ligament cells. Eur. J. Pharmacol. 2010, 644, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Prasad, S.; Bhalerao, A.; Cucullo, L. NRF2 and NF-қB interplay in cerebrovascular and neurodegenerative disorders: Molecular mechanisms and possible therapeutic approaches. Redox Biol. 2019, 21, 101059. [Google Scholar] [CrossRef]
- Wu, S.; Liao, X.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Zhang, J.; Wu, Q.; Wang, J.; Ding, Y. , Antioxidant and anti-inflammation effects of dietary phytochemicals: The Nrf2/NF-kappaB signalling pathway and upstream factors of Nrf2. Phytochemistry 2022, 204, 113429. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, F.; Bordoni, A. Foodomics: A new comprehensive approach to food and nutrition. Genes Nutr. 2012, 8, 1–4. [Google Scholar] [CrossRef] [PubMed]
- LeVatte, M.; Keshteli, A.H.; Zarei, P.; Wishart, D.S. Applications of Metabolomics to Precision Nutrition. Lifestyle Genom. 2021, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Braconi, D.; Bernardini, G.; Millucci, L.; Santucci, A. Foodomics for human health: Current status and perspectives. Expert Rev. Proteom. 2017, 15, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Rodríguez, G.; Montero, L.; Herrero, M.; Cifuentes, A.; Castro-Puyana, M. Capillary electromigration methods for food analysis and Foodomics: Advances and applications in the period March 2021 to March 2023. ELECTROPHORESIS 2023. [Google Scholar] [CrossRef] [PubMed]
- German, J.B.; Zivkovic, A.M.; Dallas, D.C.; Smilowitz, J.T. , Nutrigenomics and personalized diets: What will they mean for food? Annu. Rev. Food Sci. Technol. 2011, 2, 97–123. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. , Next-generation sequencing platforms. Annu. Rev. Anal. Chem. 2013, 6, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.W.; Abell, G.C.; Kim, K.-H.; Nam, Y.-D.; Bae, J.-W. Comparing microarrays and next-generation sequencing technologies for microbial ecology research. Trends Biotechnol. 2010, 28, 291–299. [Google Scholar] [CrossRef]
- Berlin, K.; Koren, S.; Chin, C.-S.; Drake, J.P.; Landolin, J.M.; Phillippy, A.M. , Assembling large genomes with single-molecule sequencing and locality-sensitive hashing. Nat. Biotechnol. 2015, 33, 623–630. [Google Scholar] [CrossRef]
- Pushkarev, D.; Neff, N.F.; Quake, S.R. , Single-molecule sequencing of an individual human genome. Nat. Biotechnol. 2009, 27, 847–850. [Google Scholar] [CrossRef]
- Tuberosa, R.; Frascaroli, E.; Salvi, S. Leveraging plant genomics for better and healthier food. Curr. Opin. Food Sci. 2017, 16, 100–105. [Google Scholar] [CrossRef]
- Bashiardes, S.; Godneva, A.; Elinav, E.; Segal, E. Towards utilization of the human genome and microbiome for personalized nutrition. Curr. Opin. Biotechnol. 2018, 51, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Mattar, R.; de Campos Mazo, D.F.; Carrilho, F.J. , Lactose intolerance: Diagnosis, genetic, and clinical factors. Clin. Exp. Gastroenterol. 2012, 5, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Precision (Personalized) Nutrition: Understanding Metabolic Heterogeneity. Annu. Rev. Food Sci. Technol. 2020, 11, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, L.; Gabbianelli, R. Primers on nutrigenetics and nutri(epi)genomics: Origins and development of precision nutrition. Biochimie 2019, 160, 156–171. [Google Scholar] [CrossRef]
- Valdés, A.; Cifuentes, A.; León, C. Foodomics evaluation of bioactive compounds in foods. TrAC Trends Anal. Chem. 2017, 96, 2–13. [Google Scholar] [CrossRef]
- Soni, K.A.; Nannapaneni, R.; Tasara, T. , The contribution of transcriptomic and proteomic analysis in elucidating stress adaptation responses of Listeria monocytogenes. Foodborne Pathog. Dis. 2011, 8, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Malone, J.H.; Oliver, B. Microarrays, deep sequencing and the true measure of the transcriptome. BMC Biol. 2011, 9, 34–34. [Google Scholar] [CrossRef]
- Lancova, K.; Dip, R.; Antignac, J.-P.; Le Bizec, B.; Elliott, C.T.; Naegeli, H. , Detection of hazardous food contaminants by transcriptomics fingerprinting. TrAC Trends Anal. Chem. 2011, 30, 181–191. [Google Scholar] [CrossRef]
- Valdés, A.; Ibáñez, C.; Simó, C.; García-Cañas, V. Recent transcriptomics advances and emerging applications in food science. TrAC Trends Anal. Chem. 2013, 52, 142–154. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. , RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef]
- Wu, Q.; Yuan, H.; Zhang, L.; Zhang, Y. Recent advances on multidimensional liquid chromatography–mass spectrometry for proteomics: From qualitative to quantitative analysis—A review. Anal. Chim. Acta 2012, 731, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chandramouli, K.; Qian, P.-Y. Proteomics: Challenges, Techniques and Possibilities to Overcome Biological Sample Complexity. Hum. Genom. Proteom. 2009, 1, 239204. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, P. The future of analytical chemistry in foodomics. Curr. Opin. Food Sci. 2018, 22, 102–108. [Google Scholar] [CrossRef]
- Tholey, A.; Becker, A. , Top-down proteomics for the analysis of proteolytic events-Methods, applications and perspectives. Mol. Cell Res. 2017, 1864, 2191–2199. [Google Scholar] [CrossRef]
- D’alessandro, A.; Zolla, L. We Are What We Eat: Food Safety and Proteomics. J. Proteome Res. 2011, 11, 26–36. [Google Scholar] [CrossRef]
- Almeida, A.; Bassols, A.; Bendixen, E.; Bhide, M.; Ceciliani, F.; Cristobal, S.; Eckersall, P.D.; Hollung, K.; Lisacek, F.; Mazzucchelli, G.; et al. Animal board invited review: Advances in proteomics for animal and food sciences. Animal 2015, 9, 1–17. [Google Scholar] [CrossRef]
- Puiggròs, F.; Solà, R.; Bladé, C.; Salvadó, M.-J.; Arola, L. Nutritional biomarkers and foodomic methodologies for qualitative and quantitative analysis of bioactive ingredients in dietary intervention studies. J. Chromatogr. A 2011, 1218, 7399–7414. [Google Scholar] [CrossRef]
- Xu, Y.-J.; Wu, X. Foodomics in microbiological investigations. Curr. Opin. Food Sci. 2015, 4, 51–55. [Google Scholar] [CrossRef]
- Gómez, E.; Muñoz, M.; Simó, C.; Ibáñez, C.; Carrocera, S.; Martín-González, D.; Cifuentes, A. Corrigendum to "Non-invasive metabolomics for improved determination of embryonic sex markers in chemically defined culture medium" [J. Chromatogr. A 1474 (2016) 138-144]. J. Chromatogr. A 2017, 1526, 167. [Google Scholar] [CrossRef]
- Gowda, G.A.N.; Djukovic, D. Overview of Mass Spectrometry-Based Metabolomics: Opportunities and Challenges. In Mass Spectrometry in Metabolomics; 2014; Volume 1198, pp. 3–12. [CrossRef]
- Laghi, L.; Picone, G.; Capozzi, F. Nuclear magnetic resonance for foodomics beyond food analysis. TrAC Trends Anal. Chem. 2014, 59, 93–102. [Google Scholar] [CrossRef]
- Emwas, A.H. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar] [PubMed]
- Liu, Y.-R.; Xiao, B.-K.; Yang, J.-Y.; Guo, C.-H.; Shen, S.-J.; Tang, Z.-S.; Dong, J.-X.; Huang, R.-Q. , 1H-NMR and HPLC–MS/MS-based global/targeted metabolomic evaluation of Hypericum perforatum L. intervention for menopause. J. Funct. Foods 2015, 17, 722–741. [Google Scholar] [CrossRef]
- Kohler, I.; Giera, M. Recent advances in liquid-phase separations for clinical metabolomics. J. Sep. Sci. 2016, 40, 93–108. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Herrero, L.; González, M.; Gómara, B. Rapid and simultaneous determination of polychlorinated biphenyls and their main metabolites (hydroxylated and methyl sulfonyl) by gas chromatography coupled to mass spectrometry: Comparison of different ionisation modes. Anal. Chim. Acta 2013, 787, 148–154. [Google Scholar] [CrossRef]
- Jalandra, R.; Dalal, N.; Yadav, A.K.; Verma, D.; Sharma, M.; Singh, R.; Khosla, A.; Kumar, A.; Solanki, P.R. Emerging role of trimethylamine-N-oxide (TMAO) in colorectal cancer. Appl. Microbiol. Biotechnol. 2021, 105, 7651–7660. [Google Scholar] [CrossRef] [PubMed]
- Craciun, S.; Balskus, E.P. Microbial conversion of choline to trimethylamine requires a glycyl radical enzyme. Proc. Natl. Acad. Sci. USA 2012, 109, 21307–21312. [Google Scholar] [CrossRef]
- Campo, A.M.-D.; Bodea, S.; Hamer, H.A.; Marks, J.A.; Haiser, H.J.; Turnbaugh, P.J.; Balskus, E.P. Characterization and Detection of a Widely Distributed Gene Cluster That Predicts Anaerobic Choline Utilization by Human Gut Bacteria. Mbio 2015, 6, e00042–15. [Google Scholar] [CrossRef]
- Krueger, S.K.; Siddens, L.K.; Henderson, M.C.; Andreasen, E.A.; Tanguay, R.L.; Pereira, C.B.; Cabacungan, E.T.; Hines, R.N.; Ardlie, K.G.; Williams, D.E. Haplotype and functional analysis of four flavin-containing monooxygenase isoform 2 (FMO2) polymorphisms in Hispanics. Pharmacogenetics Genom. 2005, 15, 245–256. [Google Scholar] [CrossRef]
- Cashman, J.R.; Akerman, B.R.; Forrest, S.M.; Treacy, E.P. Population-specific polymorphisms of the human FMO3 gene: Significance for detoxication. Drug Metab. Dispos. 2000, 28, 169–173. [Google Scholar] [PubMed]
- Treacy, E.P.; Akerman, B.R.; Chow, L.M.L.; Youil, R.; Lin, C.B.; Bruce, A.G.; Knight, M.; Danks, D.M.; Cashman, J.R.; Forrest, S.M. Mutations of the flavin-containing monooxygenase gene (FMO3) cause trimethylaminuria, a defect in detoxication. Hum. Mol. Genet. 1998, 7, 839–845. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Suez, J.; Elinav, E. , You are what you eat: Diet, health and the gut microbiota. Nat Rev Gastroenterol Hepatol 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Wang, H.; Rong, X.; Zhao, G.; Zhou, Y.; Xiao, Y.; Ma, D.; Jin, X.; Wu, Y.; Yan, Y.; Yang, H.; et al. The microbial metabolite trimethylamine N-oxide promotes antitumor immunity in triple-negative breast cancer. Cell Metab. 2022, 34, 581–594. [Google Scholar] [CrossRef]
- Wu, K.; Yuan, Y.; Yu, H.; Dai, X.; Wang, S.; Sun, Z.; Wang, F.; Fei, H.; Lin, Q.; Jiang, H.; et al. The gut microbial metabolite trimethylamine N-oxide aggravates GVHD by inducing M1 macrophage polarization in mice. Blood 2020, 136, 501–515. [Google Scholar] [CrossRef]
- Chen, M.L.; Zhu, X.H.; Ran, L.; Lang, H.D.; Yi, L.; Mi, M.T. Trimethylamine-N-Oxide Induces Vascular Inflammation by Activating the NLRP3 Inflammasome Through the SIRT3-SOD2-mtROS Signaling Pathway. J Am Heart Assoc 2017, 6, e006347. [Google Scholar] [CrossRef]
- Mirji, G.; Worth, A.; Bhat, S.A.; El Sayed, M.; Kannan, T.; Goldman, A.R.; Tang, H.Y.; Liu, Q.; Auslander, N.; Dang, C.V.; et al. The microbiome-derived metabolite TMAO drives immune activation and boosts responses to immune checkpoint blockade in pancreatic cancer. Sci Immunol 2022, 7, eabn0704. [Google Scholar] [CrossRef]
- Roberts, A.B.; Gu, X.; Buffa, J.A.; Hurd, A.G.; Wang, Z.; Zhu, W.; Gupta, N.; Skye, S.M.; Cody, D.B.; Levison, B.S.; et al. Development of a gut microbe–targeted nonlethal therapeutic to inhibit thrombosis potential. Nat. Med. 2018, 24, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Denman, S.E.; Morrison, M.; Yu, Z.; Dore, J.; Leclerc, M.; McSweeney, C.S. , Dysbiosis of fecal microbiota in Crohn's disease patients as revealed by a custom phylogenetic microarray. Inflamm Bowel Dis 2010, 16, 2034–2042. [Google Scholar] [CrossRef]
- Manichanh, C.; Rigottier-Gois, L.; Bonnaud, E.; Gloux, K.; Pelletier, E.; Frangeul, L.; Nalin, R.; Jarrin, C.; Chardon, P.; Marteau, P.; et al. Reduced diversity of faecal microbiota in Crohn's disease revealed by a metagenomic approach. Gut 2006, 55, 205–211. [Google Scholar] [CrossRef]
- Patel, N.; Alkhouri, N.; Eng, K.; Cikach, F.; Mahajan, L.; Yan, C.; Grove, D.; Rome, E.S.; Lopez, R.; Dweik, R.A. Metabolomic analysis of breath volatile organic compounds reveals unique breathprints in children with inflammatory bowel disease: A pilot study. Aliment. Pharmacol. Ther. 2014, 40, 498–507. [Google Scholar] [CrossRef]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical Aspects and Pathophysiology of Inflammatory Bowel Disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef]
- Wilson, A.; Teft, W.A.; Morse, B.L.; Choi, Y.H.; Woolsey, S.; DeGorter, M.K.; Hegele, R.A.; Tirona, R.G.; Kim, R.B. , Trimethylamine-N-oxide: A Novel Biomarker for the Identification of Inflammatory Bowel Disease. Dig Dis Sci 2015, 60, 3620–3230. [Google Scholar] [CrossRef] [PubMed]
- Puiggròs, F.; Solà, R.; Bladé, C.; Salvadó, M.-J.; Arola, L. Nutritional biomarkers and foodomic methodologies for qualitative and quantitative analysis of bioactive ingredients in dietary intervention studies. J. Chromatogr. A 2011, 1218, 7399–7414. [Google Scholar] [CrossRef] [PubMed]
- García-Cañas, V.; Simó, C.; León, C.; Cifuentes, A. Advances in Nutrigenomics research: Novel and future analytical approaches to investigate the biological activity of natural compounds and food functions. J. Pharm. Biomed. Anal. 2010, 51, 290–304. [Google Scholar] [CrossRef]
- Valdés, A.; García-Cañas, V.; Rocamora-Reverte, L.; Gómez-Martínez. ; Ferragut, J.A.; Cifuentes, A. Effect of rosemary polyphenols on human colon cancer cells: Transcriptomic profiling and functional enrichment analysis. Genes Nutr. 2012, 8, 43–60. [Google Scholar] [CrossRef]
- Dolara, P.; Luceri, C.; De Filippo, C.; Femia, A.P.; Giovannelli, L.; Caderni, G.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Cresci, A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. Mol. Mech. Mutagen. 2005, 591, 237–246. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Zhou, H.; Zhou, J.; Glenn, T.C.; Shen, C.-L.; Wang, J.-S. Long-term treatment with green tea polyphenols modifies the gut microbiome of female sprague-dawley rats. J. Nutr. Biochem. 2018, 56, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Shan, S.; Zhang, C.; Shi, J.; Li, H.; Li, Z. Inhibitory Effects of Bound Polyphenol from Foxtail Millet Bran on Colitis-Associated Carcinogenesis by the Restoration of Gut Microbiota in a Mice Model. ACS Appl. Mater. Interfaces 2020, 68, 3506–3517. [Google Scholar] [CrossRef]
- Ruiz, L.; Hidalgo, C.; Blanco-Míguez, A.; Lourenço, A.; Sánchez, B.; Margolles, A. Tackling probiotic and gut microbiota functionality through proteomics. J. Proteom. 2016, 147, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Valdés, A.; Artemenko, K.A.; Bergquist, J.; García-Cañas, V.; Cifuentes, A. Comprehensive Proteomic Study of the Antiproliferative Activity of a Polyphenol-Enriched Rosemary Extract on Colon Cancer Cells Using Nanoliquid Chromatography–Orbitrap MS/MS. J. Proteome Res. 2016, 15, 1971–1985. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.P.; Cooney, J.M.; Dommels, Y.E.; Nones, K.; Brewster, D.T.; Park, Z.; Butts, C.A.; McNabb, W.C.; Laing, W.A.; Roy, N.C. , Modulation of colonic inflammation in Mdr1a(-/-) mice by green tea polyphenols and their effects on the colon transcriptome and proteome. J Nutr Biochem 2013, 24, 1678–1690. [Google Scholar] [CrossRef]
- Li, Z.; Xie, J.; Tian, X.; Li, K.; Hou, A.; Wang, Y. Proteomic changes in EHEC O157:H7 under catechin intervention. Microb. Pathog. 2018, 123, 9–17. [Google Scholar] [CrossRef]
- Pimentel, G.; Burton, K.J.; von Ah, U.; Bütikofer, U.; Pralong, F.P.; Vionnet, N.; Portmann, R.; Vergères, G. Metabolic Footprinting of Fermented Milk Consumption in Serum of Healthy Men. J. Nutr. 2018, 148, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Arroyo, S.; Gomez-Martinez, A.; Rocamora-Reverte, L.; Quirantes-Pine, R.; Segura-Carretero, A.; Fernandez-Gutierrez, A.; Ferragut, J.A. , Application of nanoLC-ESI-TOF-MS for the metabolomic analysis of phenolic compounds from extra-virgin olive oil in treated colon-cancer cells. J Pharm Biomed Anal 2012, 63, 128–34. [Google Scholar] [CrossRef]
- Li, Q.; Liang, X.; Guo, N.; Hu, L.; E. , M.P.; Wu, Y.; Xue, X.; Wu, L.; Wang, K. Protective effects of Bee pollen extract on the Caco-2 intestinal barrier dysfunctions induced by dextran sulfate sodium. BioMedicine 2019, 117, 109200. [Google Scholar] [CrossRef]
- Dihal, A.A.; van der Woude, H.; Hendriksen, P.J.M.; Charif, H.; Dekker, L.J.; Ijsselstijn, L.; de Boer, V.C.J.; Alink, G.M.; Burgers, P.C.; Rietjens, I.M.C.M.; et al. Transcriptome and proteome profiling of colon mucosa from quercetin fed F344 rats point to tumor preventive mechanisms, increased mitochondrial fatty acid degradation and decreased glycolysis. Proteomics 2008, 8, 45–61. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Picone, G.; Pasini, F.; Caboni, M.F.; Gianotti, A.; Bordoni, A.; Capozzi, F. Olive oil industry by-products. Effects of a polyphenol-rich extract on the metabolome and response to inflammation in cultured intestinal cell. Food Res. Int. 2018, 113, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Seidel, D.V.; Azcárate-Peril, M.A.; Chapkin, R.S.; Turner, N.D. Shaping functional gut microbiota using dietary bioactives to reduce colon cancer risk. Semin. Cancer Biol. 2017, 46, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, C.; Valdés, A.; Garcia-Cañas, V.; Simo, C.; Celebier, M.; Rocamora-Reverte, L.; Gómez-Martínez, Á.; Herrero, M.; Castro-Puyana, M.; Segura-Carretero, A.; et al. Global Foodomics strategy to investigate the health benefits of dietary constituents. J. Chromatogr. A 2012, 1248, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.-O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef]
- Gomez-Lopez, I.; Lobo-Rodrigo, G.; Portillo, M.P.; Cano, M.P. Characterization, Stability, and Bioaccessibility of Betalain and Phenolic Compounds from Opuntia stricta var. Dillenii Fruits and Products of Their Industrialization. Foods 2021, 10, 1593. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Samba-Mondonga, M.; Constante, M.; Fragoso, G.; Calvé, A.; Santos, M.M. Curcumin induces mild anemia in a DSS-induced colitis mouse model maintained on an iron-sufficient diet. PLoS ONE 2019, 14, e0208677. [Google Scholar] [CrossRef]
- Inoue, H.; Akiyama, S.; Maeda-Yamamoto, M.; Nesumi, A.; Tanaka, T.; Murakami, A. High-dose green tea polyphenols induce nephrotoxicity in dextran sulfate sodium-induced colitis mice by down-regulation of antioxidant enzymes and heat-shock protein expressions. Cell Stress Chaperon 2011, 16, 653–662. [Google Scholar] [CrossRef]
- Kobayashi, H.; Murata, M.; Kawanishi, S.; Oikawa, S. Polyphenols with Anti-Amyloid beta Aggregation Show Potential Risk of Toxicity Via Pro-Oxidant Properties. Int J Mol Sci 2020, 21, 3561. [Google Scholar] [CrossRef]
- Inoue, H.; Maeda-Yamamoto, M.; Nesumi, A.; Tanaka, T.; Murakami, A. Low and Medium but Not High Doses of Green Tea Polyphenols Ameliorated Dextran Sodium Sulfate-Induced Hepatotoxicity and Nephrotoxicity. Biosci. Biotechnol. Biochem. 2013, 77, 1223–1228. [Google Scholar] [CrossRef]
- Gandhi, H.; Rathore, C.; Dua, K.; Vihal, S.; Tambuwala, M.M.; Negi, P. Efficacy of resveratrol encapsulated microsponges delivered by pectin based matrix tablets in rats with acetic acid-induced ulcerative colitis. Drug Dev. Ind. Pharm. 2020, 46, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Pérez, A.A.; Rodriguez-Nogales, A.; Ortiz-Cullera, V.; Algieri, F.; Garrido-Mesa, J.; Zorrilla, P.; Rodriguez-Cabezas, M.E.; Garrido-Mesa, N.; Utrilla, M.P.; De Matteis, L.; et al. Silk fibroin nanoparticles constitute a vector for controlled release of resveratrol in an experimental model of inflammatory bowel disease in rats. Int. J. Nanomed. 2014, 9, 4507–4520. [Google Scholar] [CrossRef]
- Pujara, N.; Wong, K.Y.; Qu, Z.; Wang, R.; Moniruzzaman, M.; Rewatkar, P.; Kumeria, T.; Ross, B.P.; McGuckin, M.; Popat, A. , Oral Delivery of beta-Lactoglobulin-Nanosphere-Encapsulated Resveratrol Alleviates Inflammation in Winnie Mice with Spontaneous Ulcerative Colitis. Mol Pharm 2021, 18, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Huguet-Casquero, A.; Xu, Y.; Gainza, E.; Pedraz, J.L.; Beloqui, A. Oral delivery of oleuropein-loaded lipid nanocarriers alleviates inflammation and oxidative stress in acute colitis. Int. J. Pharm. 2020, 586, 119515. [Google Scholar] [CrossRef] [PubMed]
- Marinho, S.; Illanes, M.; Ávila-Román, J.; Motilva, V.; Talero, E. Anti-Inflammatory Effects of Rosmarinic Acid-Loaded Nanovesicles in Acute Colitis through Modulation of NLRP3 Inflammasome. Biomolecules 2021, 11, 162. [Google Scholar] [CrossRef]
- Nguyen, T.-H.T.; Trinh, N.-T.; Tran, H.N.; Tran, H.T.; Le, P.Q.; Ngo, D.-N.; Tran-Van, H.; Van Vo, T.; Vong, L.B.; Nagasaki, Y. Improving silymarin oral bioavailability using silica-installed redox nanoparticle to suppress inflammatory bowel disease. J. Control. Release 2020, 331, 515–524. [Google Scholar] [CrossRef]
- Arafat, E.A.; Marzouk, R.E.; Mostafa, S.A.; Hamed, W.H.E. Identification of the Molecular Basis of Nanocurcumin-Induced Telocyte Preservation within the Colon of Ulcerative Colitis Rat Model. Mediat. Inflamm. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Sébédio, J.-L. Metabolomics, Nutrition, and Potential Biomarkers of Food Quality, Intake, and Health Status. 2017, 82, 83–116. [CrossRef]
- Bush, C.L.; Blumberg, J.B.; El-Sohemy, A.; Minich, D.M.; Ordovás, J.M.; Reed, D.G.; Behm, V.A.Y. Toward the Definition of Personalized Nutrition: A Proposal by The American Nutrition Association. J. Am. Coll. Nutr. 2019, 39, 5–15. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361, bmj–k2173. [Google Scholar] [CrossRef]
- De Toro-Martín, J.; Arsenault, B.J.; Després, J.-P.; Vohl, M.-C. Precision Nutrition: A Review of Personalized Nutritional Approaches for the Prevention and Management of Metabolic Syndrome. Nutrients 2017, 9, 913. [Google Scholar] [CrossRef]
- Sales, N.M.R.; Pelegrini, P.B.; Goersch, M.C. Nutrigenomics: Definitions and Advances of This New Science. J. Nutr. Metab. 2014, 2014, 1–6. [Google Scholar] [CrossRef]
- Vázquez-Fresno, R.; Llorach, R.; Urpi-Sarda, M.; Lupianez-Barbero, A.; Estruch, R.; Corella, D.; Fitó, M.; Arós, F.; Ruiz-Canela, M.; Salas-Salvadó, J.; et al. Metabolomic Pattern Analysis after Mediterranean Diet Intervention in a Nondiabetic Population: A 1- and 3-Year Follow-up in the PREDIMED Study. J. Proteome Res. 2014, 14, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Rezzi, S.; Ramadan, Z.; Fay, L.B.; Kochhar, S. Nutritional Metabonomics: Applications and Perspectives. J. Proteome Res. 2007, 6, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Gerasimidis, K.; Nikolaou, C.K.; Edwards, C.A.; McGrogan, P. , Serial fecal calprotectin changes in children with Crohn's disease on treatment with exclusive enteral nutrition: Associations with disease activity, treatment response, and prediction of a clinical relapse. J Clin Gastroenterol 2011, 45, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Cameron, F.L.; Gerasimidis, K.; Papangelou, A.; Missiou, D.; Garrick, V.; Cardigan, T.; Buchanan, E.; Barclay, A.R.; McGrogan, P.; Russell, R.K. , Clinical progress in the two years following a course of exclusive enteral nutrition in 109 paediatric patients with Crohn's disease. Aliment Pharmacol Ther 2013, 37, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ney, M.; Eslamparast, T.; Vandermeer, B.; Ismond, K.P.; Kroeker, K.; Halloran, B.; Raman, M.; Tandon, P. Systematic review of nutrition screening and assessment in inflammatory bowel disease. World J. Gastroenterol. 2019, 25, 3823–3837. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular mechanisms activated or inhibited by dietary polyphenols in IBD.
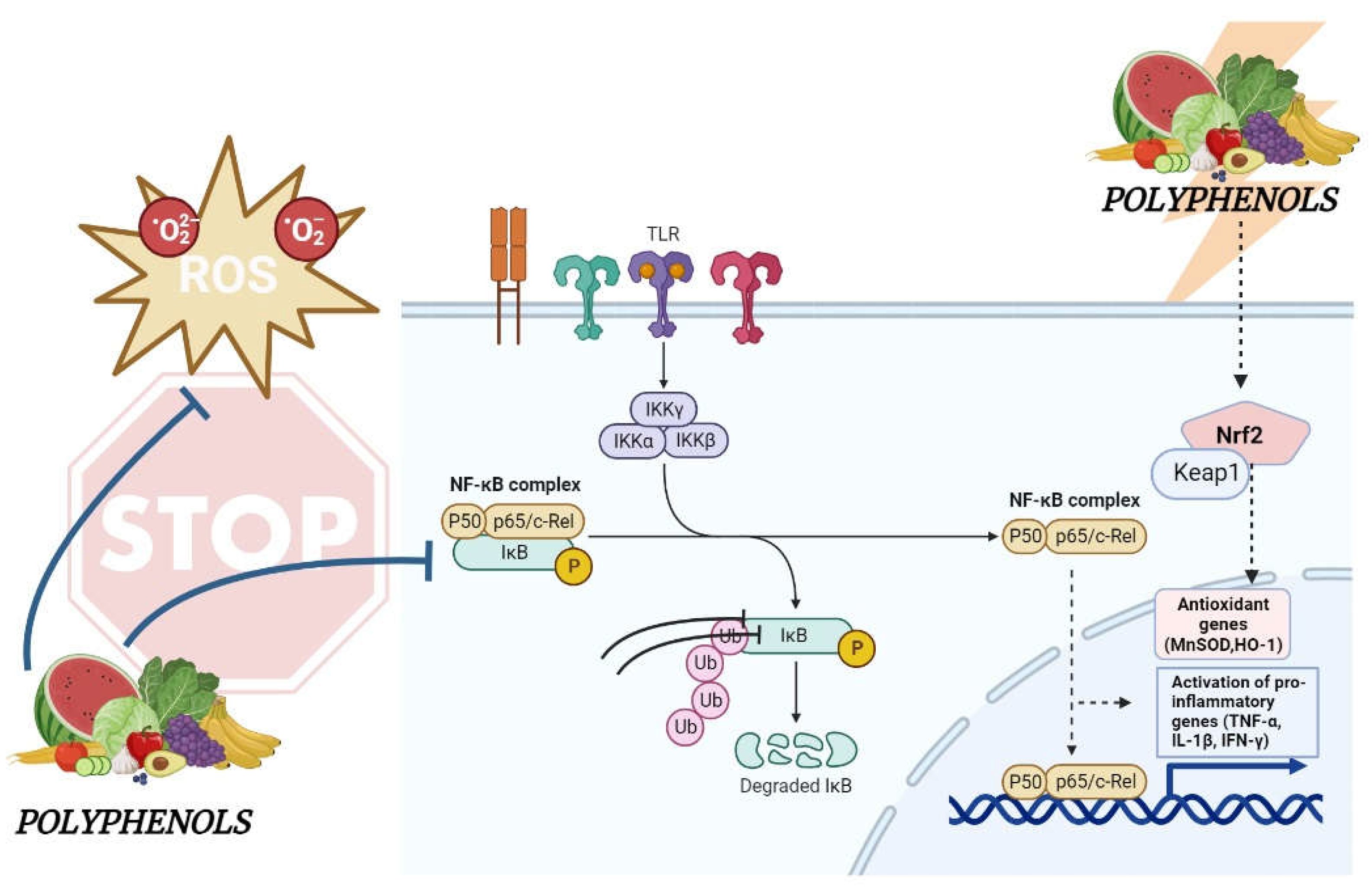
Figure 2.
Multiomics explain the close relationship between host, foods, microbiota, microenvironment and the biological variability between individuals in response to nutrition.
Figure 2.
Multiomics explain the close relationship between host, foods, microbiota, microenvironment and the biological variability between individuals in response to nutrition.
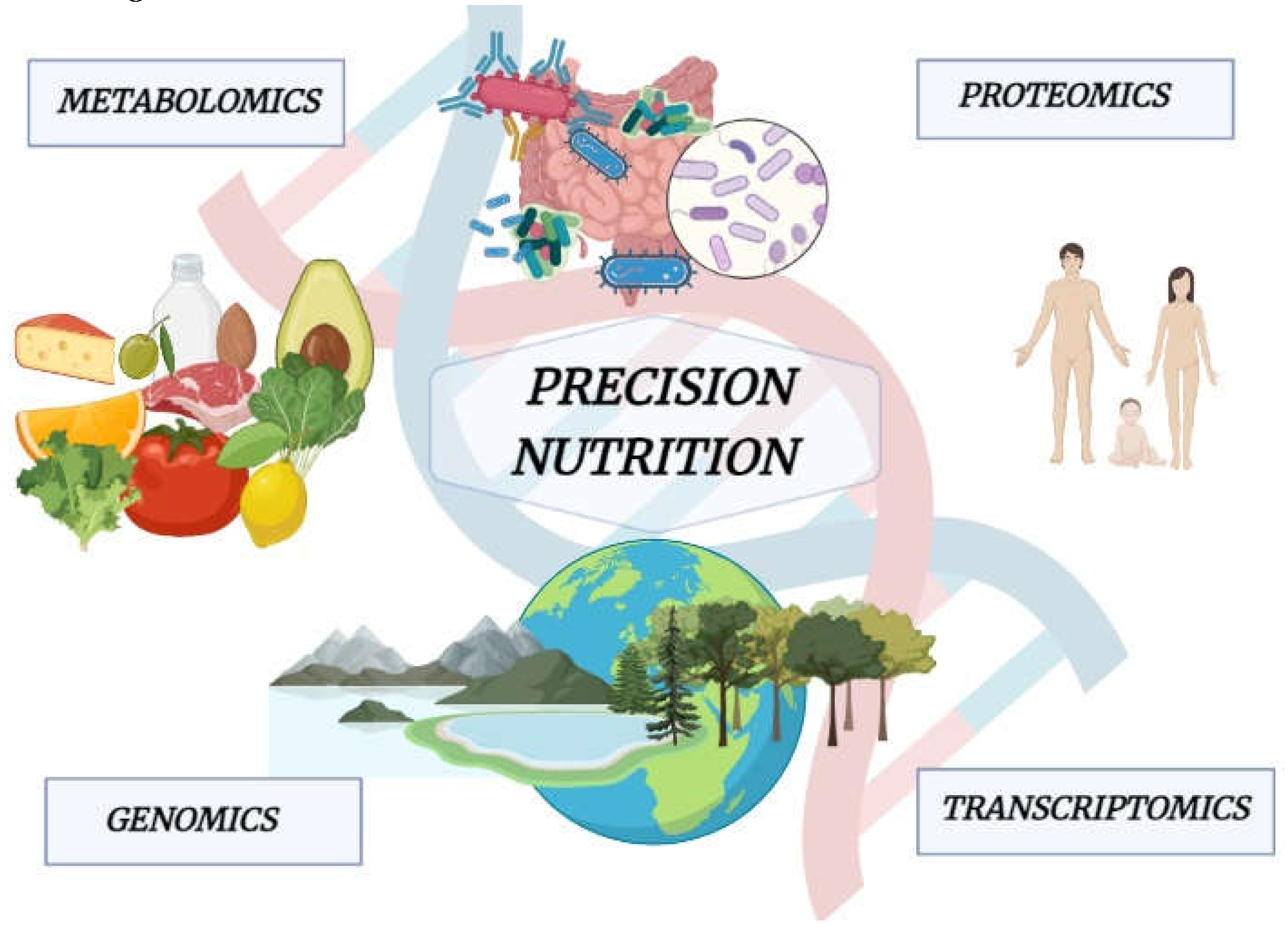
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



