
Submitted:
24 August 2023
Posted:
25 August 2023
You are already at the latest version
Abstract
Emerging evidence has indicated the involvement of extra-hypothalamic kisspeptin and GnRHR in reproductive function. In this study, we evaluate if maternal exposure to the pesticide chlorpyrifos (CPF) and/or a high-fat diet (HFD) have an impact on the expression of kisspeptin and GnRHR in reproductive organ of offspring’s rats. 16 Pregnant rats are divided into 4 groups, control group (n=4), CPF group (4 rats exposed daily to 1/mg/kg/day), HFD group (4 rats randomly fed a 5,25 kcal/g HFD), co-exposed group (4 rats exposed to CPF and HDF). At postnatal development PND60, males and female offspring were sacrificed. The reproductive organs (ovary and testis) were removed and histological, immunohistological analysis and in silico quantification (software Tissuefax, HistoQuest) was assessed to investigate the impact of different treatment on kisspeptin and GnRHR expression in reproductive organs. The main outcomes of the study showed a significant decrease in rat offspring’s body weight in CPF group from PND30 andPND60 (p<0.05 and p<0.01, respectively). Histological analysis shows a significant increase in atretic follicle and abnormal testis structure with germ cell desquamation at CPF exposed group. The immunodetection quantification of protein shows a significant decrease in GnRHR and kisspeptin in HFD and CPF exposed group, respectively in testis rat offspring. Perinatal exposure to CPF and HFD exposure affect the reproduction function of rat offspring.
Keywords:
chlorpyrifos
; high-fat diet
; kisspeptin
; GnRHR
; ovary
; testis
; rat
1. Introduction
The World Health organization defines the infertility as the inability to achieve a successful pregnancy within 12 months of unprotected intercourse [1,2]. It affects approximately 17 to 20% of couples worldwide [2]. According to a 2015 epidemiological study, the World Health Organization estimates that nearly around 186 million individuals worldwide affected with infertility globally [3]. Numerous factors can affect fertility including environmental and lifestyle factors such as smoking, excessive alcohol intake and obesity [4].
Since the last decade, the literature reported that the exposure to pesticides and the issue of obesity have emerged as pressing public health concerns. They are interconnected and have been associated with various adverse effects, including a decline in reproductive health and a detrimental impact on the effectiveness of assisted reproduction technology [5]. The mechanisms behind this relationship are not yet fully elucidated, but it is believed that pesticide exposure and high fat diet disrupt metabolic processes [6] and alter the reproductive health [7]. Our previous studies have shown that pesticides exposure alters some sperm parameters including motility and vitality and increase the sperm DNA fragmentation [8]. The Anses (L’Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail) and EPA (United States Environmental Protection Agency) reported that certain pesticides are considered as an endocrine disruptor including, the chlorpyrifos polychlorinated bisphenyls (PCB), bisphenol A (BPA), dibutyl phthalates (DBP) and alkyl phenols, Di-(2-ethylhexyl) phthalate (DEHP), Glyphosate, Phthalates, dichlorodiphenyltrichloroethane DDT and Methoxychlor and chlorpyrifos (CPF) [9,10].
Chlorpyrifos is an organophosphate insecticide widely used in agriculture to control pests on crops [11]. However, studies raised concerns about its potential effects on human health, including reproductive toxicity which includes both male and female reproductive organs as well as the development of embryos and fetuses [12,13]. CPF is an insecticide that disrupts the nervous system by inhibiting the activity of an enzyme called acetylcholinesterase (AChE) [14,15], which plays a critical role in transmitting nerve signals in target insects, and it can also affect non-target organisms, including humans [16]. The Developmental Origins of Health and Disease (DOHaD) hypothesis suggests that environmental exposures and experiences during early life, particularly during fetal development and early childhood, can have a profound impact on an individual’s lifelong health and disease risk and can lead to long-term health consequences [6,17,18]. The main aspects of DOHaD include Fetal Programming [19], Critical Windows of Development [20], Epigenetic Changes [21], Long-Term Health Consequences [22], Interactions with Genetics, Transgenerational Effects [23]. Guidelines and regulations set by authorities like EFSA (European Food Safety Authority) and EPA (United States Environmental Protection Agency) regarding the use of pesticides like chlorpyrifos and to take necessary precautions to ban exposure, especially for pregnant women and young children [11,24].
Emerging evidence indicates that endocrine disruptors alter the synthesis and functions of androgens and estrogens, which are essential sex hormones responsible for the development and maintaining of the male reproductive system. It can disturb the hypothalamic and pituitary (HPG) axis function by altering synthesis of hormones regulating the reproduction including GnRH and kisspeptin [25].
Kisspeptin, a neuropeptide with 145-aminoacid, encoded by the KISS1 gene. It considered as a central regulator of the hypothalamic-pituitary-gonadal (HPG) axis [26]. It plays a crucial role in the regulation of reproductive function. It act as a potent stimulator of the release of gonadotropin-releasing hormone (GnRH) in the brain [27]. GnRH serves as the key mediator, conveying these cues throughout the HPG axis [28]. Over the past twenty years, significant research has been dedicated studying kisspeptin (KISS1) neurotransmitter, and a growing body of evidence supports their involvement in regulating the secretion of GnRH and gonadotropin hormones throughout the ovulatory cycle [29].
It’s believed that kisspeptin neurotransmitters were only found in the hypothalamus. However, emerging evidence have revealed interesting findings indicates that kisspeptin-producing cells are also present in several extra-hypothalamic regions of the brain, as well as in peripheral tissues such as the placenta, ovary, uterus, testis [30,31], and gastrointestinal tract. It was detected for the first time in rat tissue [32,33,34,35]. Its implicated in the regulation of puberty onset, fertility, and pregnancy [36]. Extra hypothalamic Kisspeptin has been found to modulate the secretion of reproductive hormones, influence the development and function of reproductive organs, and participate in the establishment of reproductive behaviors [37,38]. Siminara et al. 2003 has reported that mutation in kisspeptin receptor knock out (kiss1r -/- or GPR54 -/-) in mice lead to an idiopathic hypogonadotropic hypogonadism (small testes in male mice and an absence of follicular maturation in female mice) [39]. Furthermore, Feng et al. 2021 have reported that the expression of the KISS-1/GPR54 system in the testes of rats is influenced by a high-fat diet, leading to a decrease in the expression level of the KISS-1 protein [30]. A case-control study published by Abdelkareem et al. 2023, included a total of 30 patients (10 patients with unexplained recurrent pregnancy loss, 10 patients with pregnancy loss due to aneuploidy and 10 patients for control group). The study has investigated the role of KISS1 and KISS1R in early placentation. The outcome revealed a significant decrease in KISS1R expression in the chorionic tissues of (euploid and aneuploid) recurrent pregnancy loss compared with control group [40].
Several studies reported that GnRHR was founded not only in the hypothalamus but also in various extra-hypothalamic tissues, including ovary, oviduct [41], testis [42], mammary gland [43], placenta and endometrium [44]. While the specific mechanisms and the exact function of kisspeptin and GnRHR of extra-hypothalamic origin in mammals remains unknown and still under investigation. Additional research is required to better understand the relationship between environmental factors, diet and the implication of Kisspeptin in reproductive function.
The primary aim of this research is to identify the presence of Kisspeptin and GnRHR expression within the reproductive organs of rat offspring using Immunohistochemistry. Additionally, the study aims to evaluate how exposure to chlorpyrifos, and a high-fat diet impacts the expression of these proteins through in silico quantification. Furthermore, the study investigates the morpho-pathological effects on the reproductive organs using histological analysis.
2. Materials and Methods
a) Chlorpyrifos preparation and High Fat Diet
Chlorpyrifos, O, O-diethyl-O-(3,5,6-trichloro-2-pyridinyl) phosphorothioate, was purchased from LGC Standards (Molsheim, France). A quantity of 250 mg of Chlorpyrifos was combined with 25 ml of rapeseed oil (MP Biomedicals, Illkirch, France) as a vehicle, resulting in a concentration of 1 mg/ml. This mixture was then administered to the rats (the CPF group and the co-exposed group) via gavage at a dosage of 1 mg/kg body weight per day [45]. The HFD group and the co-exposed group were fed a high fat diet (60% Kcal as fat) until PND21.
b) Experimental design and ethical approval
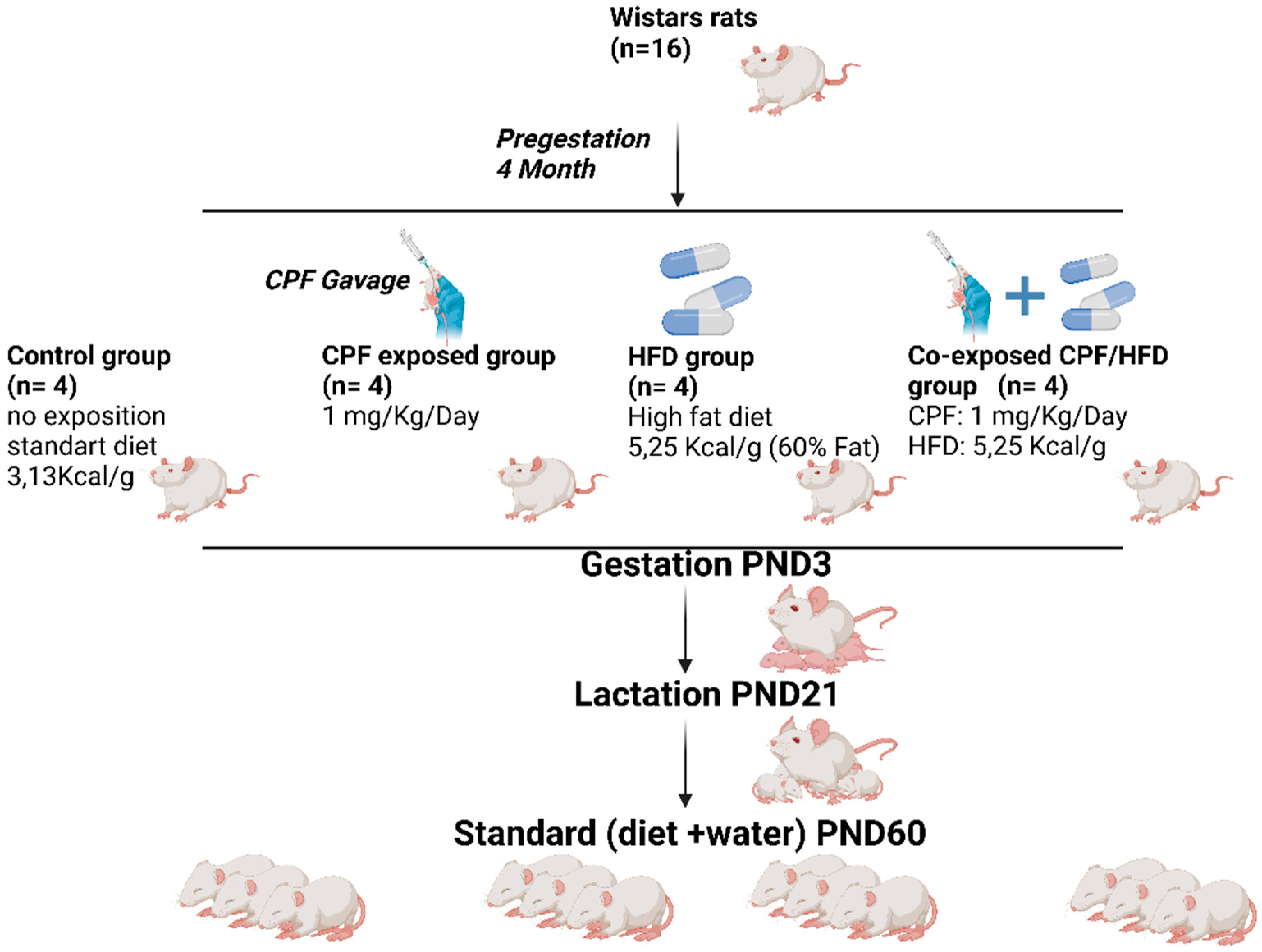
16 female Wistar rats (Janvier Labs, Le Genest Saint Isle, France,), upon arrival, aged 7 weeks, were placed in cages and kept under consistent conditions in a controlled environment. The room was maintained at a temperature of 23°C to ensure a stable and regulated environment for the rats. Then, rats were housed in NexGen Max cage system with 81 in2 /523 cm2 floor area mounted on EcoFlow rack system (Allentown Inc, Bussy Saint Georges, France). Following a week-long acclimation period, the female rats, with an average body weight of 225 ± 4.9 g, were randomly assigned to four groups. To facilitate social interaction, two rats were housed together in each cage. This housing arrangement allowed for appropriate socialization among the animals during the study (Figure 1).
Figure 1.
Study design and animal’s preparation (personal source).
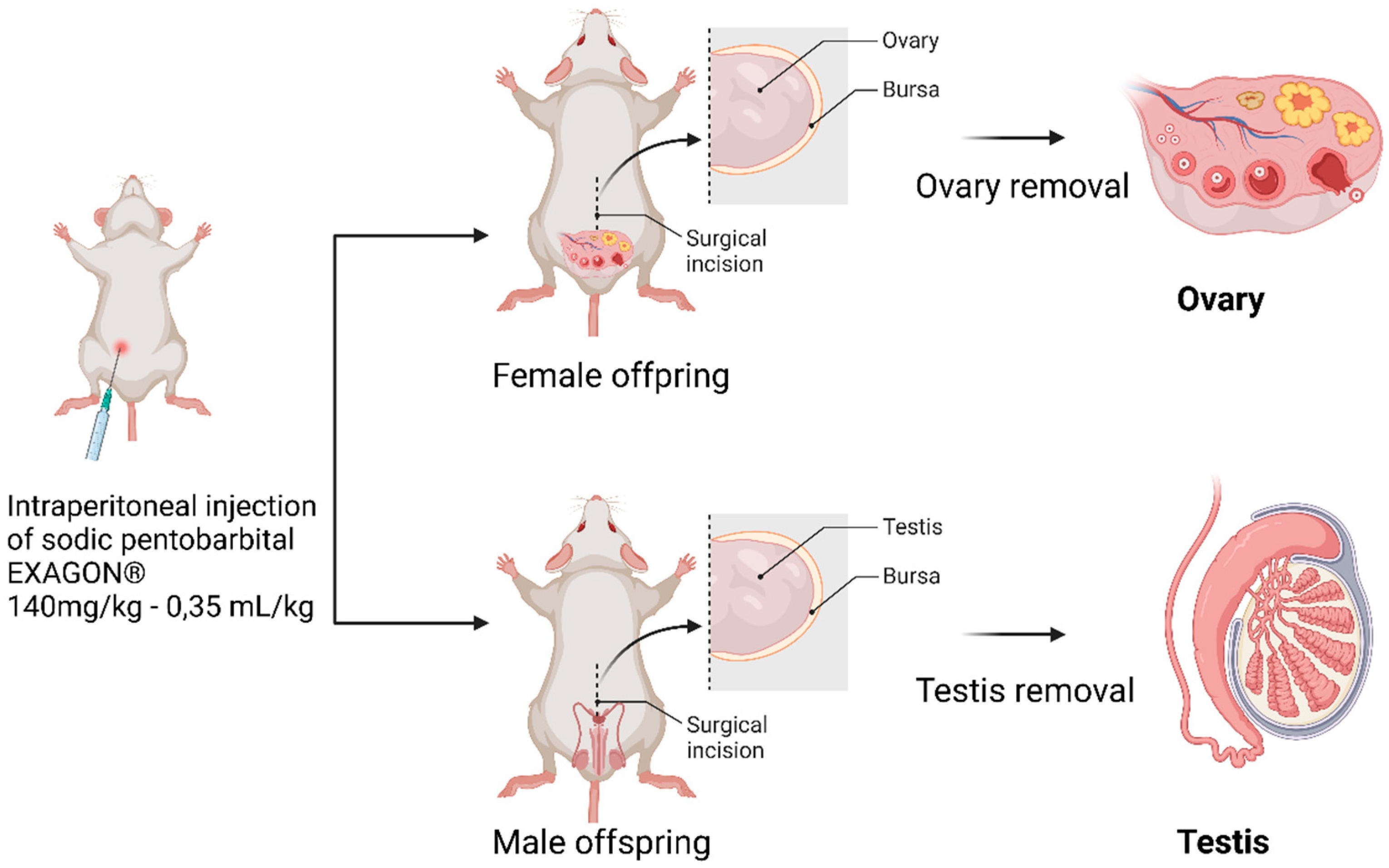
At PND60, the offspring rats were euthanized using an injection of (1 ml.kg-1; 200 mg.ml-1 solution) sodium pentobarbital from EXAGON (Axience, France). After euthanasia, the reproductive organs were dissected and placed directly into Krebs Henseleit solution from Sigma Aldrich (Saint Quentin Fallavier, France). This solution is commonly used to maintain the viability and functionality of isolated tissues during experimental procedures (Figure 2).
Figure 2.
offspring’s sacrifice and reproductive organ dissection (personal source).
The study protocol obtained approval from the Regional Directorate for Health, Animal and Environment Protection, an accredited organization in France. Additionally, it was also approved by the French Ministry of Research under the reference number APAFIS#8207-2016121322563594 v2. These regulatory approvals ensured that the study adhered to the necessary ethical and scientific standards regarding animal welfare and research conduct.
c) Immunohistochemistry (kisspeptin and GnRHR)
Tissue sections were deparaffinized in xylene (Sigma Aldrich, Germany) and rehydrated through decreasing concentrations of ethanol (TechniSolv, France). After deparaffinization the slides were incubated in Tris EDTA buffer (10mM tris-base (Cat # 161-0719, Biorad, US), 1mM EDTA solution (Cat # 161-0729, Biorad, US), 0.05% tween20® (Sigma-Aldrich, UE), pH=9) 20 min (2*10 min) in a microwave (850W) followed by washing with cold water for 10 min. To quench the endogenous peroxidase activity, the slides were incubated in water peroxide H202 (Gilbert) for 30 min then then washed twice (each 5 min delicately) in TBS (ThermoScientific, Belgium) 0,025 Triton and blocked with 10% normal Goat Serum (ab7481) with 1% BSA (ThermoScientific, USA) in TBS for 2 hours [C] between 0.5 and 10 ug/ml. thereafter, the sections were incubated overnight in a humidity room at 4 °C with primary antibody:
- -
- For kisspeptin detection: Primary rabbit polyclonal anti kisspeptin antibody (1:300 dilution; product no.251265; Abbiotec. Escondido, CA, USA)
- -
- For GnRHR detection: Primary rabbit polyclonal to GnRHR (1:200 dilution; abcam, ab202848, UK)
The sections were washed in TBS/0,025 Triton 2*(5min) under gentle agitation, then incubated with goat anti-rabbit IgG-HRP (1:1000 dilution; product no.252237; Abbiotec. Escondido, CA, USA) for 1 hour at room temperature. After washing step 3*(5min) in TBS, labeling was visualized by incubating with DAB (3,3’Diaminobenzidine) substrate kit (ab64238, abcam, UK) for 4 min then washed in 5 min in TBS. Sections were counterstained with Mayer’s hematoxylin (VWR Q-Path Chemicals, France) for 2 min and mounted with Aqueous Mounting Medium (ab64230, Abcam, UK).
The slides were observed under a light microscope, specifically the Nikon ECLIPSE Ci model from Nikon Europe B.V. This microscope offers magnifications of X4, X20, and X40, allowing for different levels of details observation. To capture the images, the NIS Elements version 1.10.00 imaging software was used. This software provides a platform for acquiring and processing images obtained from the microscope, enabling researchers to analyze and document their observations.
d) Histological analysis
Histopathological analysis of the testicular and ovary sections (4um-thick) was performed using a hematoxyline-eosine staining. 5 slides for each group and each slide contains 2~3 sections of an organ.
The paraformaldehyde-fixed paraffin-embedded sections were incubated in Xylene 2*(5 min) then rehydrated using different concentrations of ethanol (2 min ethanol 100%, 2 min ethanol 95%, 6min ethanol 70% and 2min osmosed water, respectively). The slides were stained with hematoxylin of Mayer 5% (2min) and after a washing step for2 min in osmosed water. The slides were soaked 5 times in acid/alcohol 37%. After the washing step, an eosin 1% staining was realized for 1~3 min. Finally, the slides were washed and dehydrated (2 min ethanol 95%, 2 min ethanol 100%, 2min in xylene, respectively). Morphological Analysis was performed under a light microscopy (Nikon ECLIPSE Ci, Nikon Europe B.V) with X4, X20 and X40 magnifications. The image was captured using an imaging software NIS Elements version 1.10.00.
The morphometric measurements assess the number of cells per seminiferous tubule (average of 10 selected seminiferous tubules per section); area of seminiferous tubules (average in 10 randomly selected seminiferous tubules per section) [46].
e) In silico quantification of expression
The C, CPF, HFD, CPF/HFD immunodetected sections were scanned and photographed usingTissue Faxs plus system (Tissue Gnostics Medical & Biotech Solutions, Vienna, Austria. At least 5 randomly selected slides per animal were used for immunostaining and scanned. Regions of interest (ROIs) were manually delineated using the zooming and mark-up tools available in the TissueFAXS viewer. This process allows us to focus on specific regions for further analysis or measurements. The ROIs were then digitized at 20× using HistoQuest® (TissueGnostics GmbH, Austria) and the expression of kisspeptin and GnRHR (the intensity of signals detected onto the sections) were quantified. The typical image was selected for presentation [47,48].
f) Statistical analysis
Statistical data was analyzed using StatView software version 5.0. The results were presented as mean values ± standard deviations (SD), number (n) and percentage (%). We perform a Two-way ANOVA to compare the difference between the groups. The null hypothesis of the Two-way ANOVA is that all four variables being compared have the same mean. The alternative hypothesis, on the other hand, suggests that at least one of the variables has a different average. The statistical Figure s were generated by GraphPad 9 software.
3. Results
1- Impacts of treatment on pup’s body weight
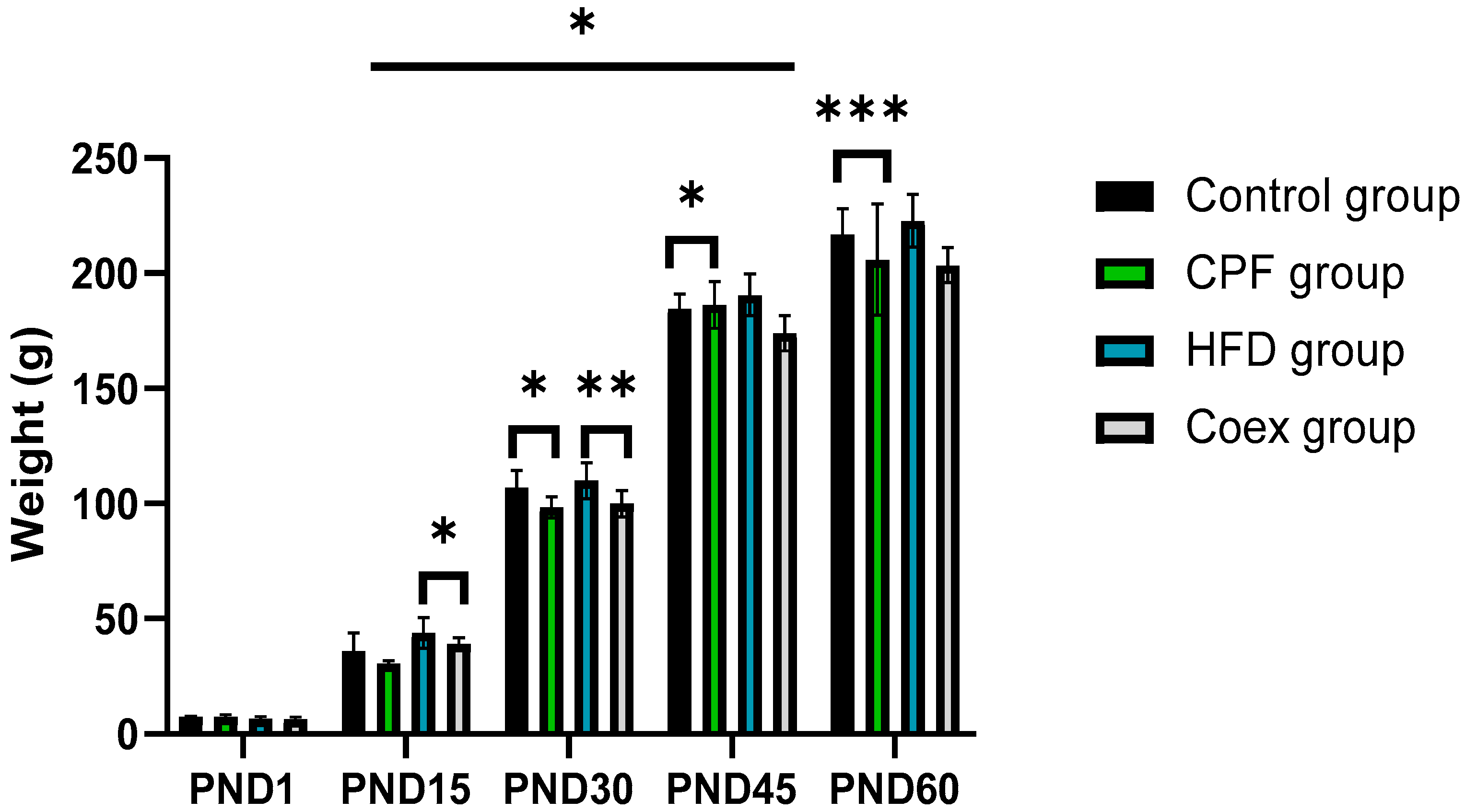
Statistical data analysis shows a significant increase in rat offspring’s body weight in postnatal period P=0.03. The high fat diet group presents the heaviest body weight groups in the different PND period.
The intergroup comparison between the control and the CPF exposed group shows a highly significant decrease in the body weight in the CPF exposed group at the period of (PND30 and PND 60) p<0.05 and p<0.001, respectively).
At PND 15 and PND 30, the body weight of rat offsprings shows a decrease in the co-exposed group to CPF/HFD compared to the HFD group (p<0.05 and p<0.01, respectively) Figure 3.
No significant differences were observed between the different groups at PND1, between control group and CPF group at PND15 and between HFD group and Coex group at PND 45 and PND 60.
Figure 3.
Evolution of the pup’s body weight (male + female) at the postnatal periods.
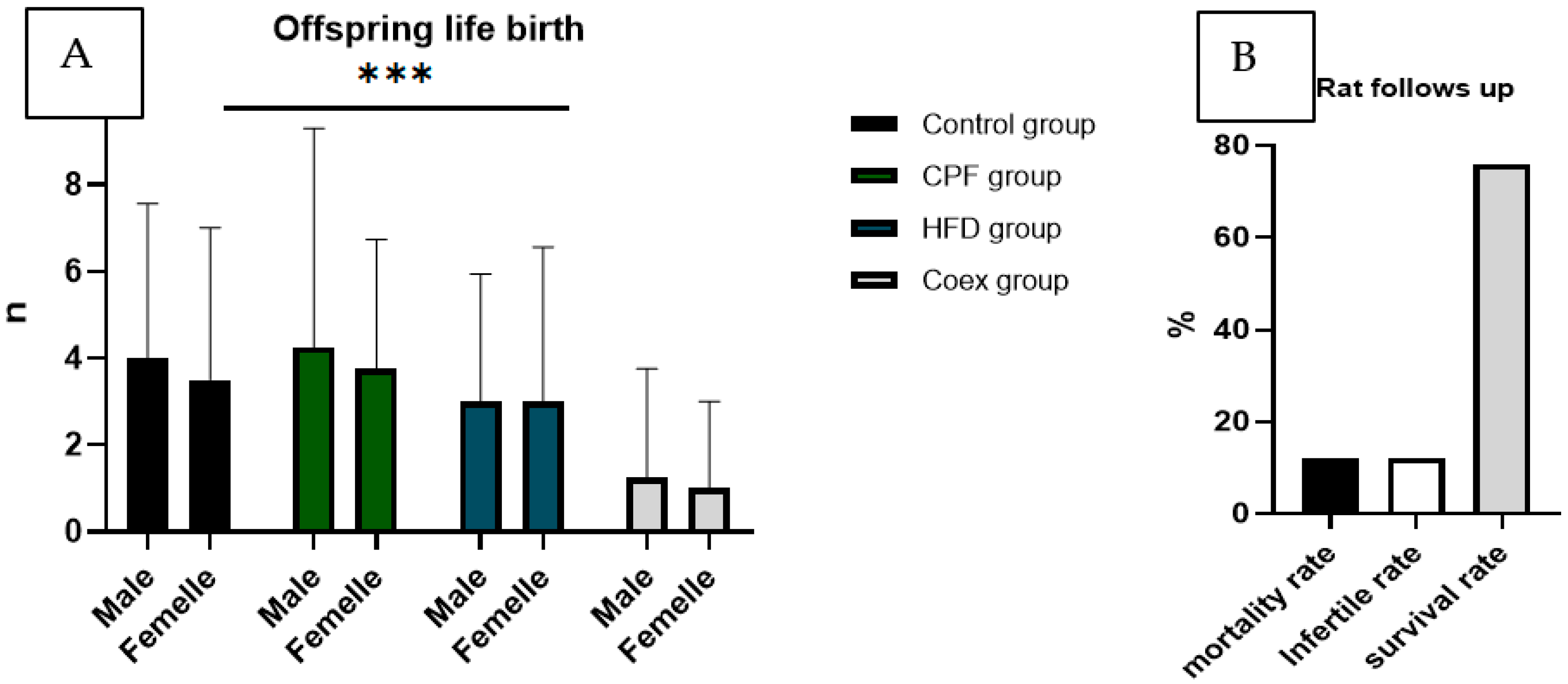
The outcomes of the rat study were showed in Figure 4, illustrating the count of male and female offspring within each group, alongside the mortality rate of the rats Figure 4.
Figure 4.
Rat follows up during the treatment (B) and gestational period (A).
2- Histological analysis
a. Ovary of rat offspring
Atresia in secondary and antral follicles was observed by eosin staining. Histological analysis of rat ovary was assessed in control group (n=5), CPF group (n=5), HFD group (n=5) and Coex group (n=5). Statistical data show a significant difference in atretic follicle number between CPF group and control group (p=0,0008) Figure 5. Follicles with fractured oocytes were observed in some sections of CPF exposed group and Coex group Figure 6.
Figure 5.
Measurement of atretic follicle number in the ovary of the rat offspring.
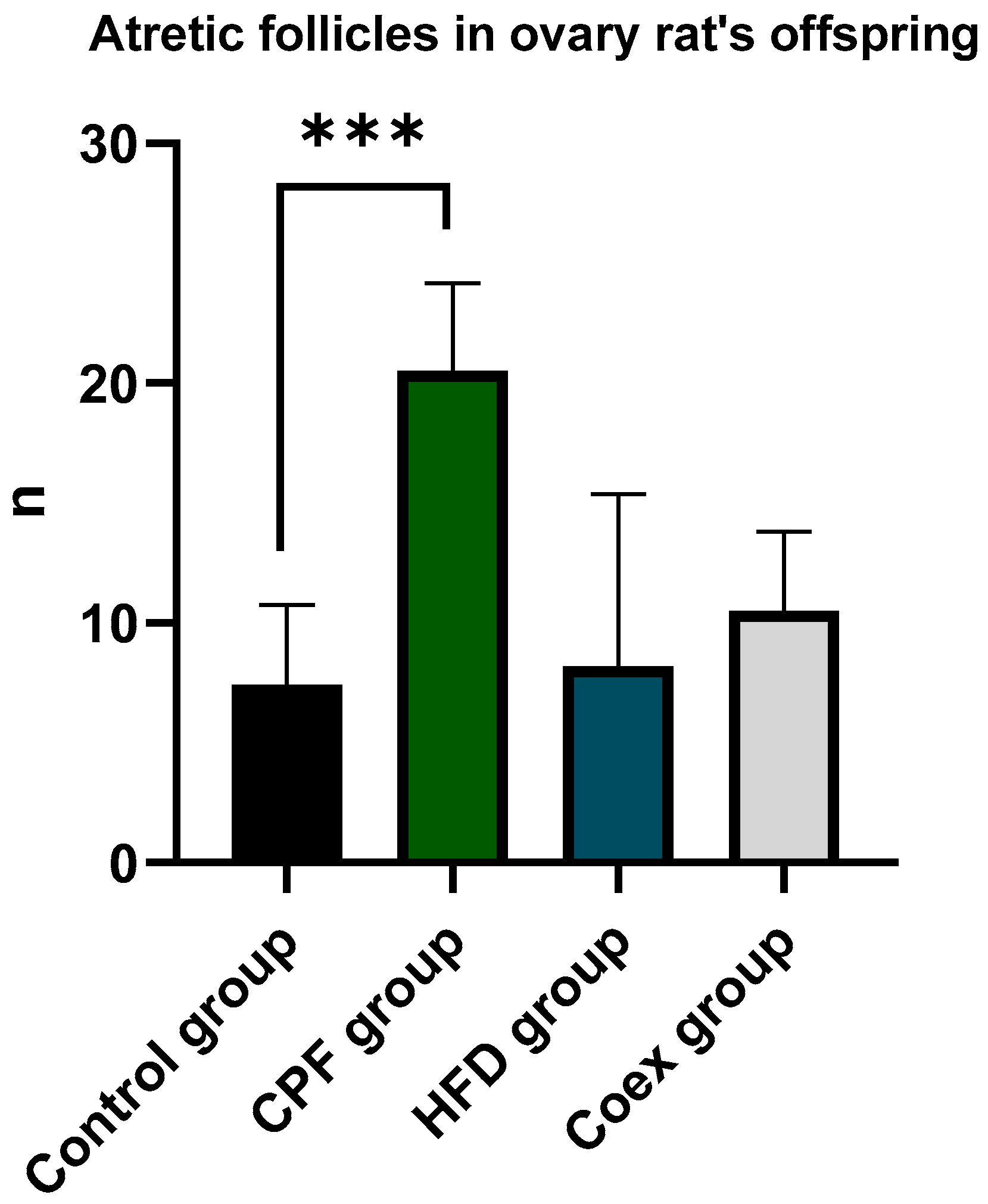
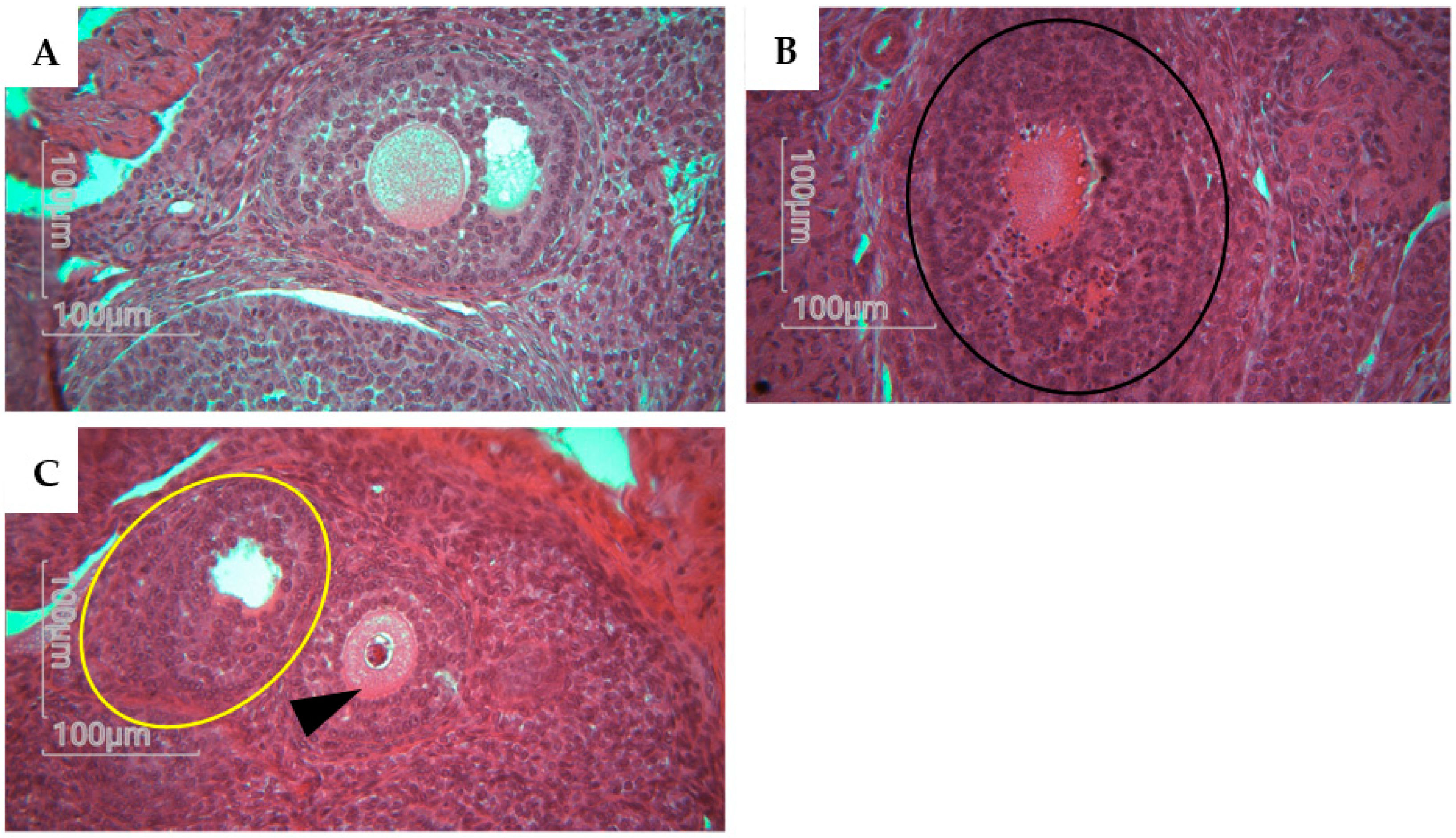
Figure 6.
Histological analysis of rat offsping follicules showing alterations Magnification X40. A represents a normal morphology of antral follicle with normal oocytes and normal cumulus cells and theca cells. B represents an atresia in follicle with color change to dark red and absence of oocyte (yellow cercle). C represents a degenerative follicle, with no oocytes (black cercle) and a follicle with fractured oocyte that colored into dark red (head arrow).
Figure 6.
Histological analysis of rat offsping follicules showing alterations Magnification X40. A represents a normal morphology of antral follicle with normal oocytes and normal cumulus cells and theca cells. B represents an atresia in follicle with color change to dark red and absence of oocyte (yellow cercle). C represents a degenerative follicle, with no oocytes (black cercle) and a follicle with fractured oocyte that colored into dark red (head arrow).
b. Testis of rat offspring
Based on the histological analysis of testis from rat offspring’s the control group displayed typical and normal histological features of the seminiferous tubes (STs). However, in contrast, the CPF and the HFD subgroups exhibited a clear desquamation of the seminiferous epithelium Figure 6. This desquamation resulted in the release of cellular debris and a large number of germinal lineage cells, predominantly primary spermatocytes, Cell in degeneration and round spermatids presented in the tubular lumen (Figure 8 B, C, D). The Coex group presents a disorganized structure of ST (Figure 7 D).
Figure 7.
Histological testicular section of rat offsprings exposed to different treatments (Magnification X 10). A represents the normal morphology of rat testis. B represents a testis morphology of CPF exposed group, The observation reveals a disordered structure of ST, a notable absence of germ cells and a partial reduction in the ST. C represents the HFD exposed group, note reduction of some germ cells. D represents the coexposed group, the germ cells found in the tubular lumen.
Figure 7.
Histological testicular section of rat offsprings exposed to different treatments (Magnification X 10). A represents the normal morphology of rat testis. B represents a testis morphology of CPF exposed group, The observation reveals a disordered structure of ST, a notable absence of germ cells and a partial reduction in the ST. C represents the HFD exposed group, note reduction of some germ cells. D represents the coexposed group, the germ cells found in the tubular lumen.
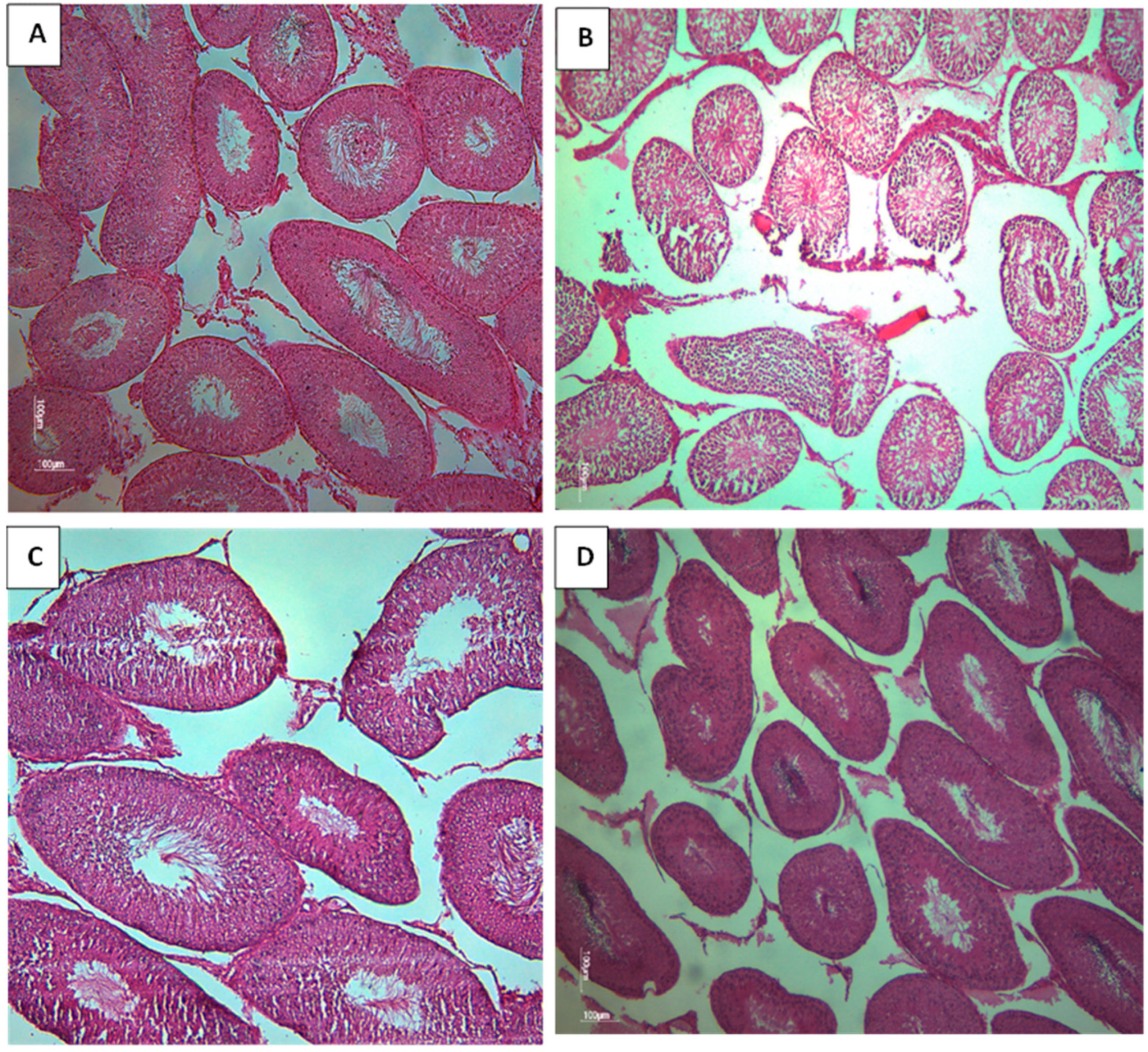
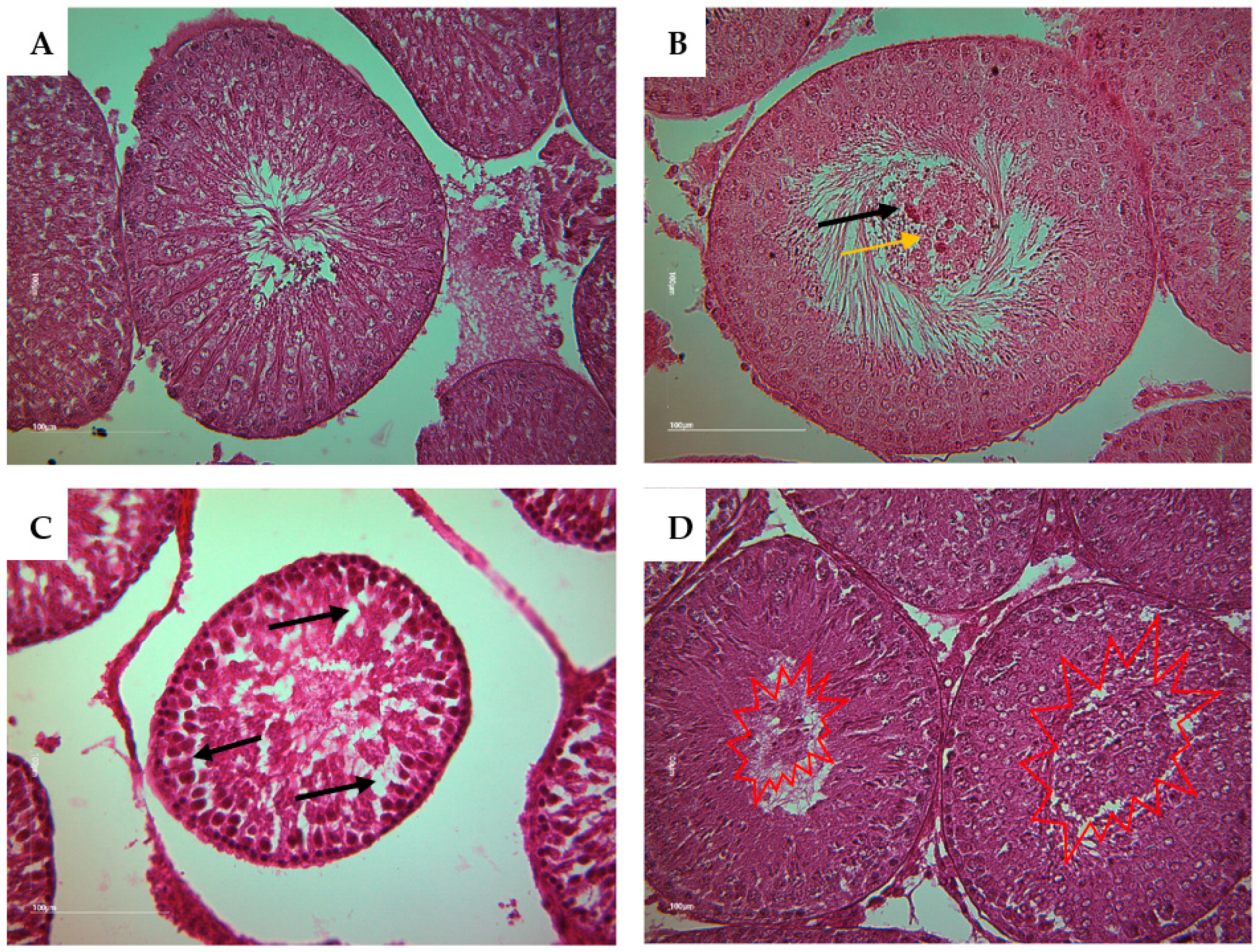
Figure 8.
Histological detection of tubular section alteration of rat offsprings (Magnification X 40). A show a normal ST containing various cell types including spermatogonia Ad, spermatogonia B, primary spermatocytes, round and elongated spermatids with Sertoli cells and Leydig cells. B shows the presence of some germinal lineage cells in the tubular lumen including primary spermatocytes (yellow arrow) and cell in degeneration (black arrow). C shows an abnormal structure of ST with partial depletion or an absence of some germ cell (black arrow). D shows an accumulation of a large quantity of germinal lineage cells in the tubular lumen including cells in degeneration, round spermatids and primary spermatocytes (red explosion).
Figure 8.
Histological detection of tubular section alteration of rat offsprings (Magnification X 40). A show a normal ST containing various cell types including spermatogonia Ad, spermatogonia B, primary spermatocytes, round and elongated spermatids with Sertoli cells and Leydig cells. B shows the presence of some germinal lineage cells in the tubular lumen including primary spermatocytes (yellow arrow) and cell in degeneration (black arrow). C shows an abnormal structure of ST with partial depletion or an absence of some germ cell (black arrow). D shows an accumulation of a large quantity of germinal lineage cells in the tubular lumen including cells in degeneration, round spermatids and primary spermatocytes (red explosion).
Morphometric analysis of tubule sections did not exhibit significant statistical differences in ST area between the different groups, as indicated in Table 1.

Table 1.
Area (μm2) of seminiferous tubule sections observed in rat offspring in different groups; Control, CFP group, HFD group and Coexp group.
Table 1.
Area (μm2) of seminiferous tubule sections observed in rat offspring in different groups; Control, CFP group, HFD group and Coexp group.
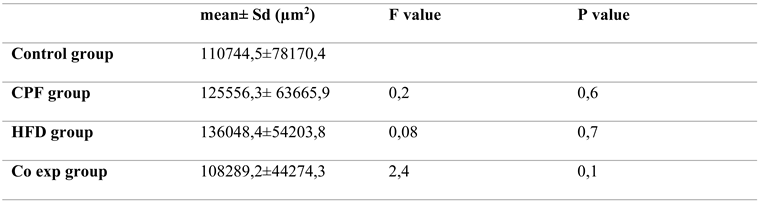
Values are expressed in mean ± standard deviation; Two- Way ANOVA tests; significance (p≤0.05).
3- Kisspeptin and GnRHR immunodetection
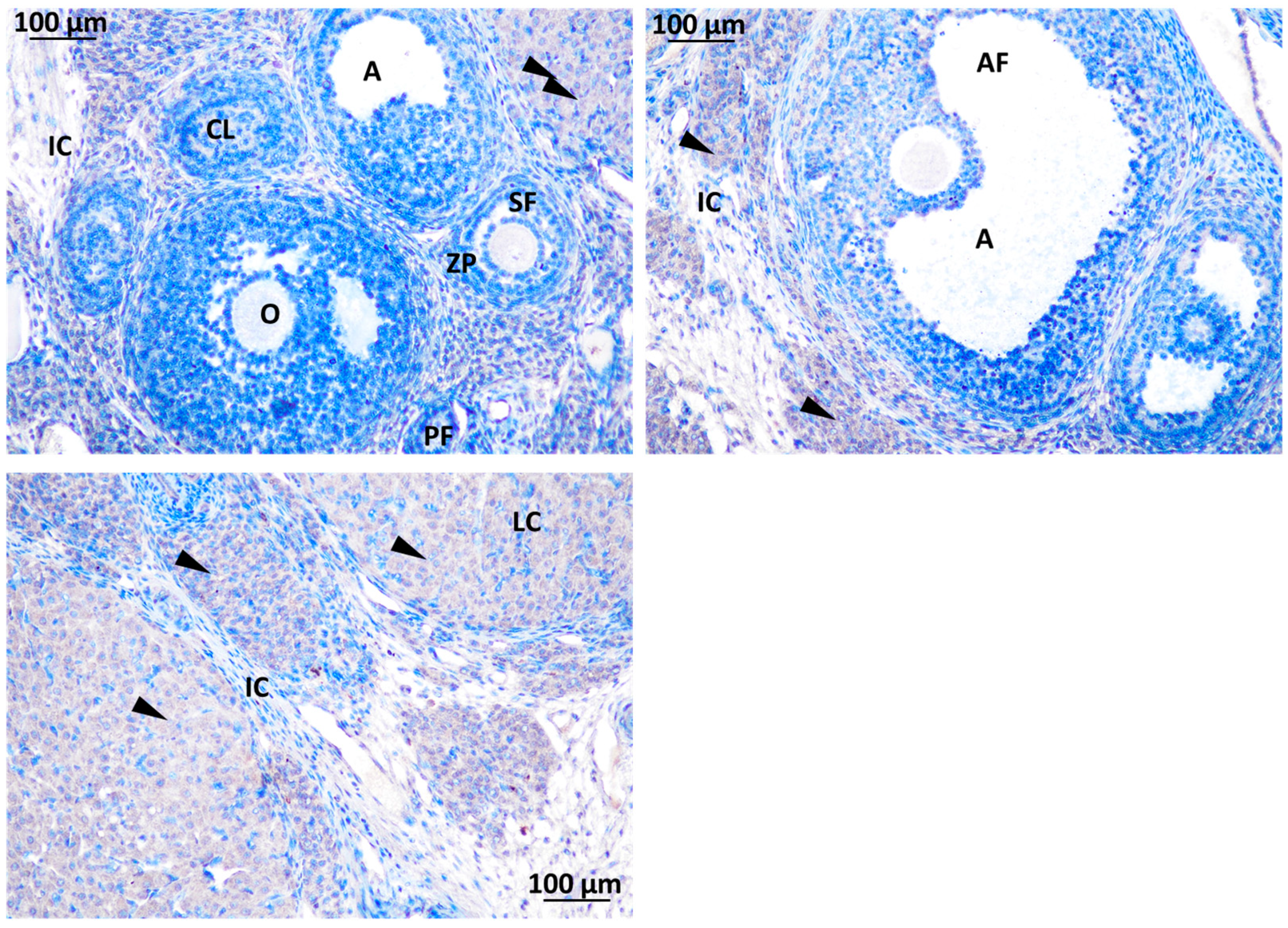
a. Localization of immunoreactive kisspeptin and GnRHR in the ovary of rat offspring
The expression of Kisspeptin was detected in different localizations within the rat ovary. Strong immunoreactivity for GnRH was exhibited in the CL: corpus luteum, LC: luteal cells, O: oocyte, TC: theca cell of atretic follicle and in the cytoplasm of GC: granulosa cell. No detection was observed in interstitial tissue and theca cells of secondary follicle Figure 9.
Figure 9.
immunodetection of Kisspeptin in rat ovary offspring. IC: interstitial cell, CL: corpus luteum cell, A: antrum, O: oocyte, ZP: zona pellucida, SF: secondary follicle, PF: primary follicle, AF: antral follicle, CO: cumulus oophorus, TC: theca cell, GC: granulosa cell. The first section of the Figure shows the atretic follicle (surrounded in red) Magnification X40.
Figure 9.
immunodetection of Kisspeptin in rat ovary offspring. IC: interstitial cell, CL: corpus luteum cell, A: antrum, O: oocyte, ZP: zona pellucida, SF: secondary follicle, PF: primary follicle, AF: antral follicle, CO: cumulus oophorus, TC: theca cell, GC: granulosa cell. The first section of the Figure shows the atretic follicle (surrounded in red) Magnification X40.
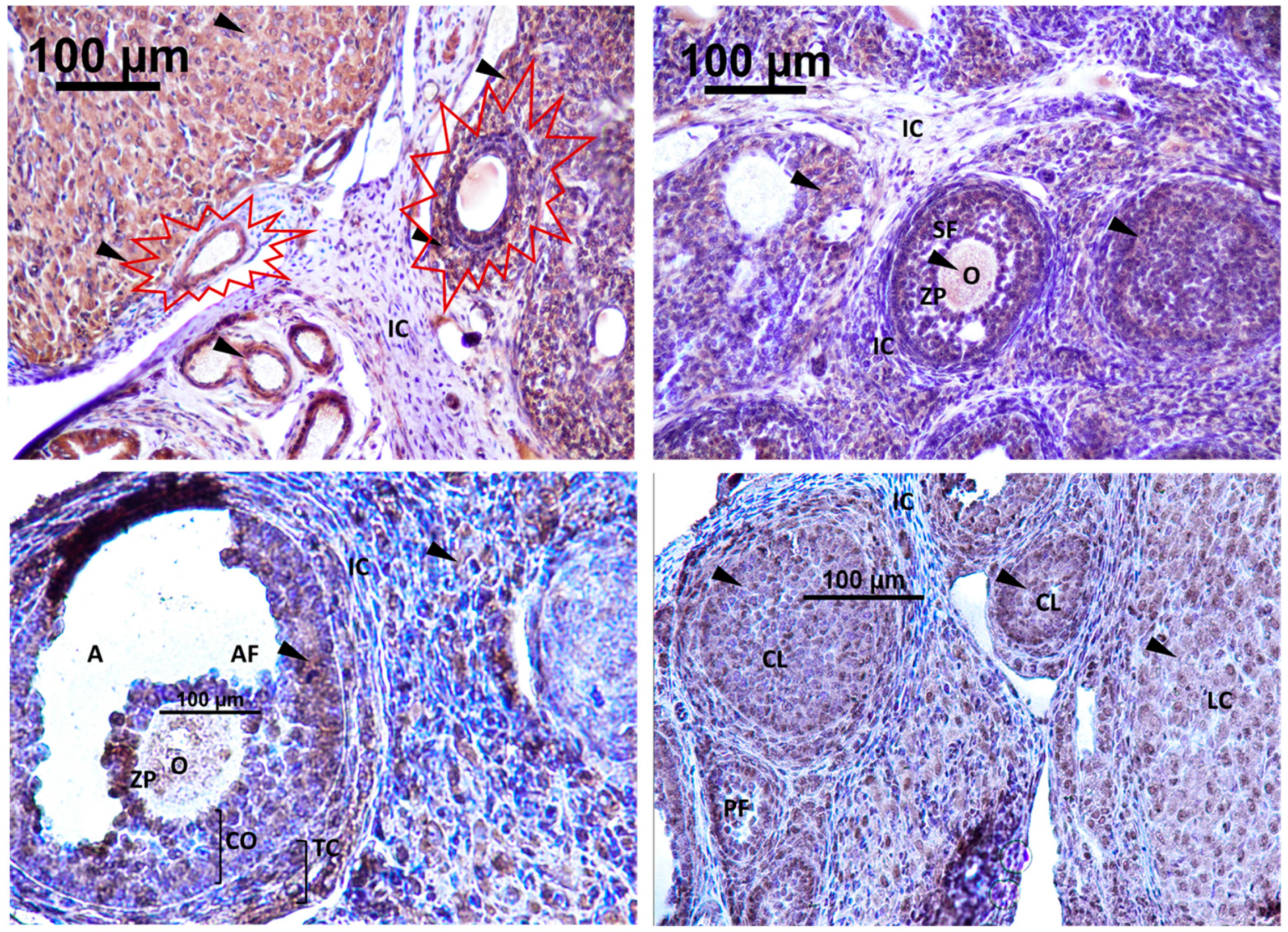
Strong immunodetection for GnRHR was noted in the interstitial cell (IC) and O: oocyte within the rat ovary offspring Figure 9. No detection of GnRHR in SF: secondary follicle, PF: primary follicle, AF: antral follicle, CO: cumulus oophorus, TC: theca cell, GC: granulosa cell Figure 10.
Figure 10.
immunodetection of GnRHR in rat ovary offspring; IC: interstitial cell, CL: corpus luteum, A: antrum, O: oocyte, ZP: zona pellucida, SF: secondary follicle, PF: primary follicle, AF: antral follicle Magnification X40.
Figure 10.
immunodetection of GnRHR in rat ovary offspring; IC: interstitial cell, CL: corpus luteum, A: antrum, O: oocyte, ZP: zona pellucida, SF: secondary follicle, PF: primary follicle, AF: antral follicle Magnification X40.
b. Localization of immunoreactive kisspeptin and GnRHR in the testis of rat offspring
Immunodetection of kisspeptin has revealed its presence in specific areas within the testis of rat offspring. Kisspeptin was observed in two main locations in the testis: Interstitial Leydig Cells (LC) in the interstitial spaces between the seminiferous tubules suggesting a potential role in regulating testosterone production and Inside the Seminiferous Tubules within the seminiferous tubules, indicates a possible direct influence on spermatogenesis Figure 11.
Figure 11.
immunodetection of Kisspeptin in seminiferous tubules of rat offsprings testis; S.Ad: spermatogonia Ad, SpI: spermatocyte I, SCN: Sertoli cell nuclei, ST: seminiferous tube, S.B: Spermatogonia B, Sp C: Spermatogonia C.
Figure 11.
immunodetection of Kisspeptin in seminiferous tubules of rat offsprings testis; S.Ad: spermatogonia Ad, SpI: spermatocyte I, SCN: Sertoli cell nuclei, ST: seminiferous tube, S.B: Spermatogonia B, Sp C: Spermatogonia C.
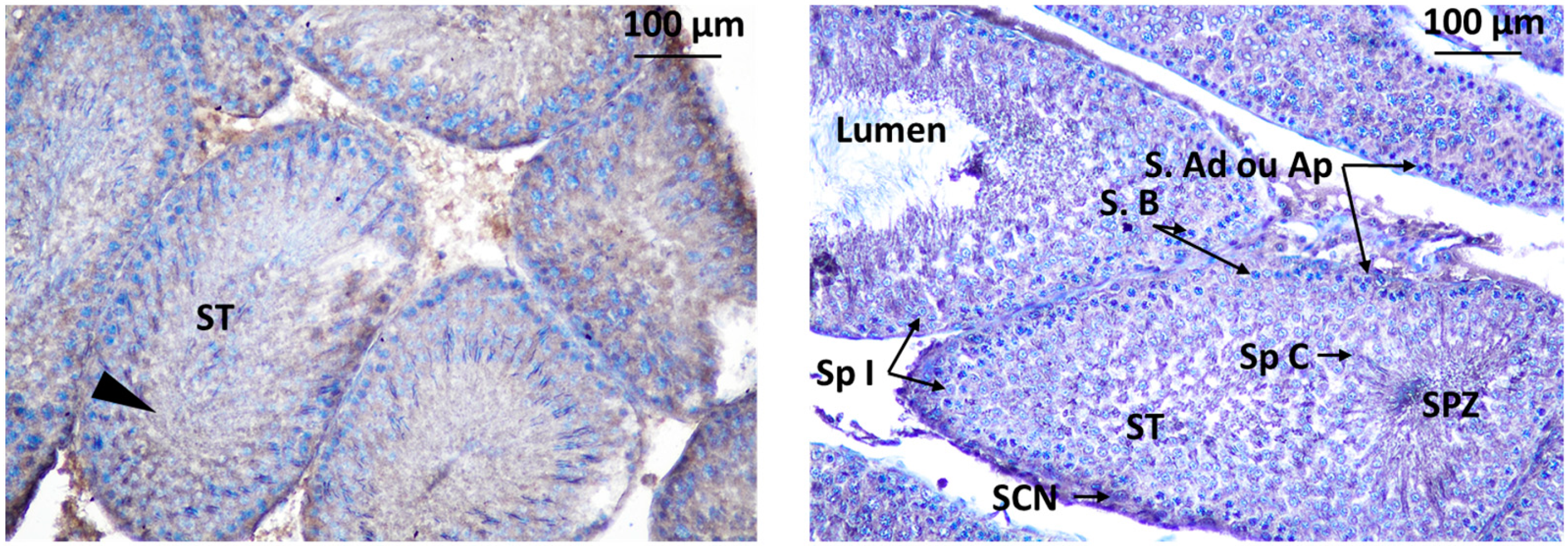
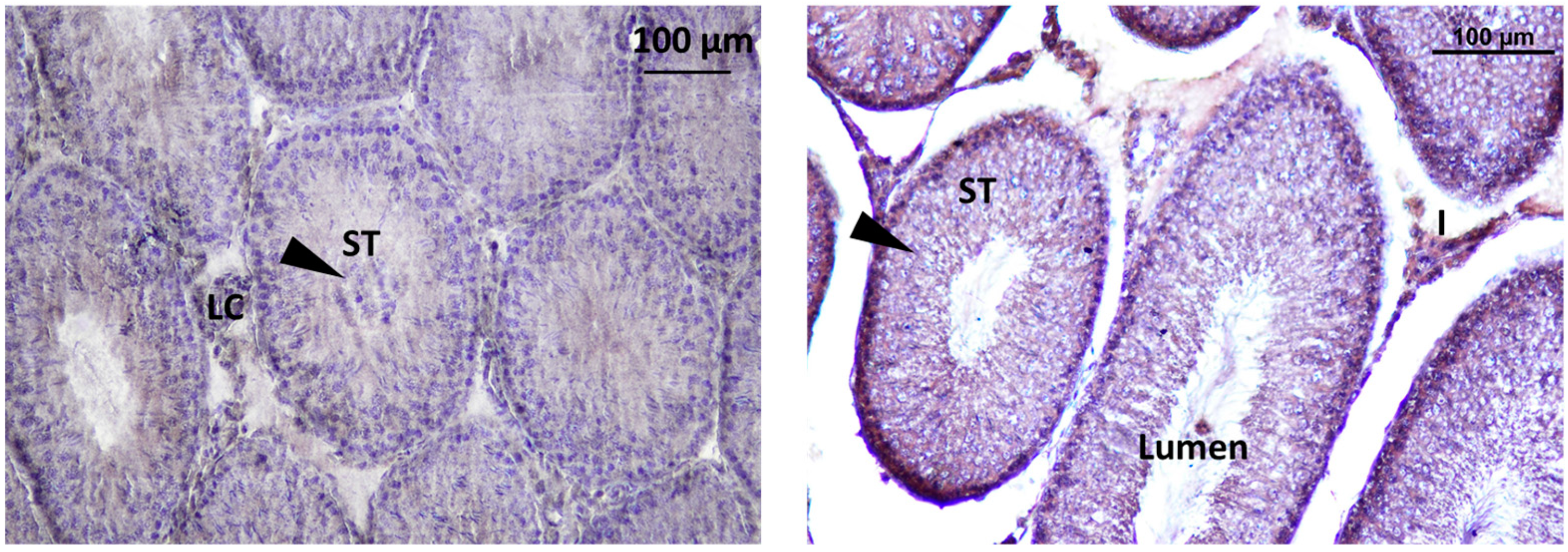
Localization of GnRHR was observed in the interstitial Leydig cells (LC) and inside the seminiferous tubule. Localization of immunoreactive kisspeptin and GnRHR in the testis of rat offsprings Figure 12.
Figure 12.
immunodetection of Kisspeptin in seminiferous tubules of rat offsprings testis; LC: Leydig cell, ST: seminiferous tube, I: intertubular.
Figure 12.
immunodetection of Kisspeptin in seminiferous tubules of rat offsprings testis; LC: Leydig cell, ST: seminiferous tube, I: intertubular.
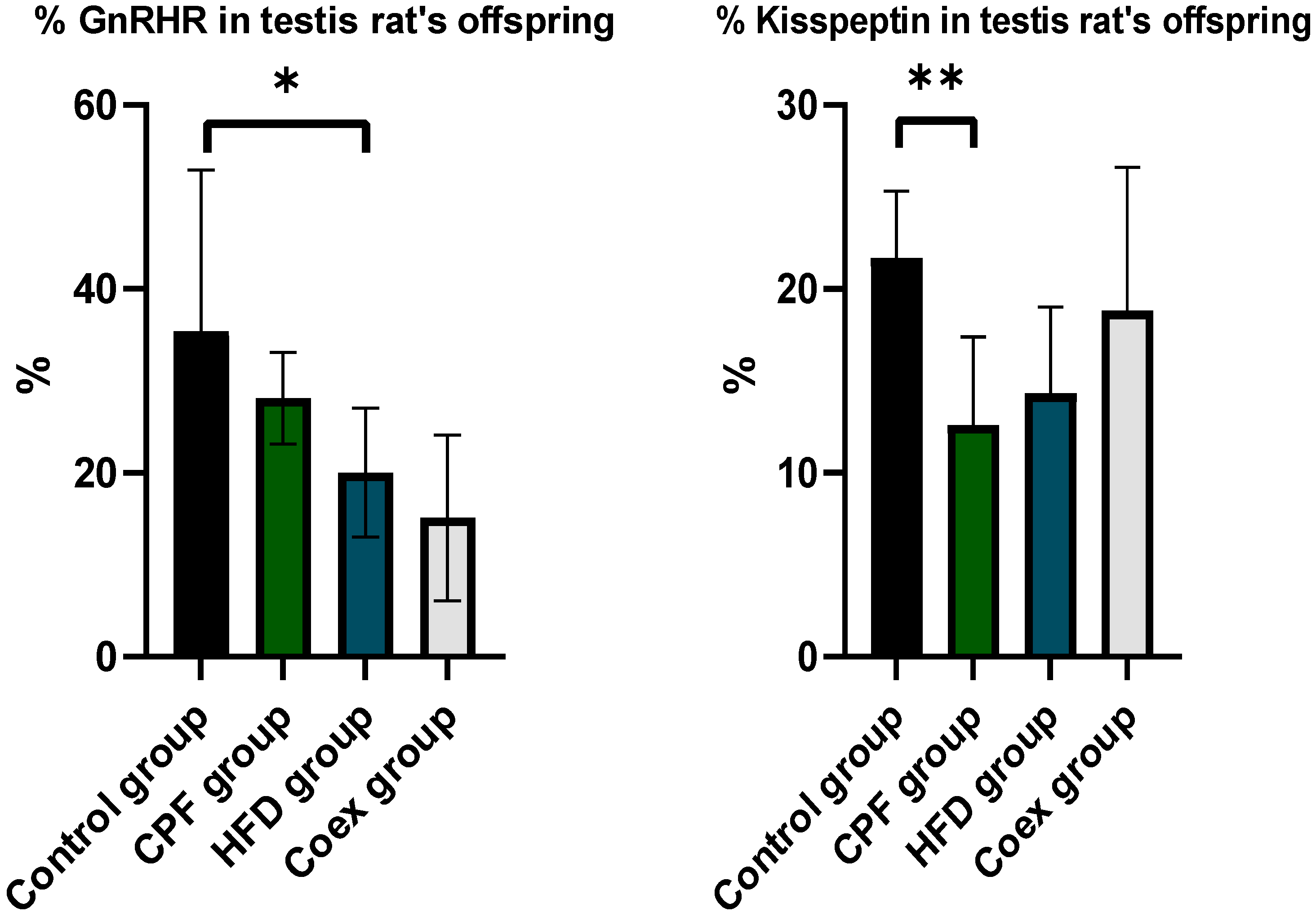
4- In silico quantification of the Expression of Kisspeptin and GnRHR
The levels of Kisspeptin and GnRHR expression in the reproductive organs (testis and ovary) of rat offsprings were measured using HistoQuest®.
The statistical analysis revealed a significant decrease in the percentage of GnRHR (P=0.01) in the offspring from the high-fat diet (HFD) group compared to the control group. However, there were no significant differences observed between the control group, the group exposed to CPF (chlorpyrifos, a pesticide), and the coexposure group (p<=0,05) (Figure 13 A).
The data analysis demonstrated a significant reduction in the percentage of Kisspeptin (P=0.002) in the testis rat offspring exposed to CPF, compared to the control group. No significant difference found in the other groups (p<=0,05) (Figure 13 B).
The quantification of levels of Kisspeptin and GnRHR in ovary rat offspring show no significant difference between the different groups (p<=0,05).
Figure 13.
Effect of different treatments on GnRHR and kisspeptin expression in testis rat Offspring.
Figure 13.
Effect of different treatments on GnRHR and kisspeptin expression in testis rat Offspring.
The comparison of the number of cells in ovary rat offspring revealed a significant decrease in cell number shown in the CPF group compared to the control group (p=0,002). No significant difference found in the other groups (p<=0,05) Figure 14.
Data analysis of testis’ cell number in different treated group show no significant differences in cell number between the groups (p<=0,05).
Figure 11.
Effect of Different Treatments on the number of cell in ovary rat Offspring.
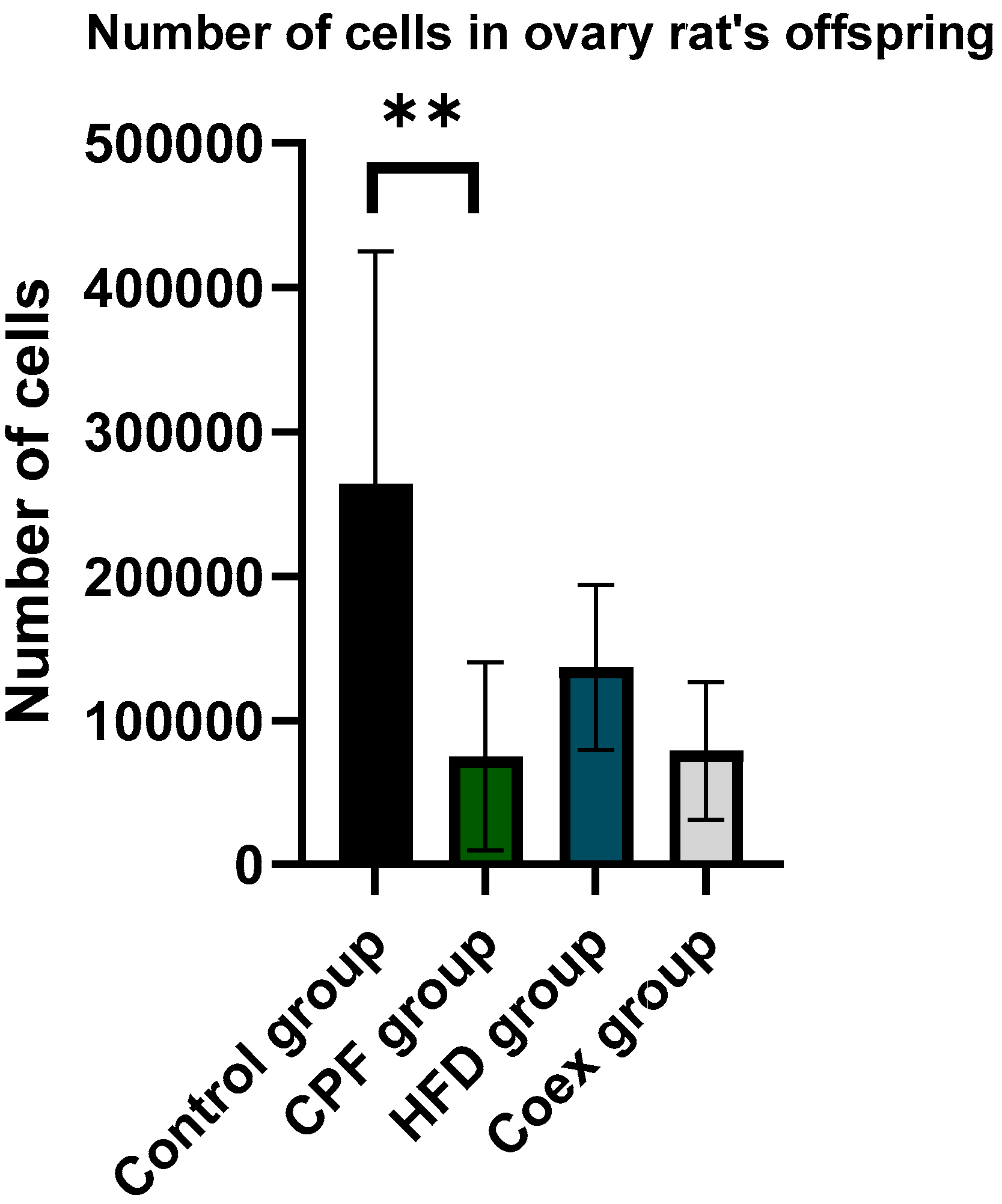
4. Discussion
The present study investigates the adverse effects of chlorpyrifos pesticide and high fat diet on fertility and reproduction function in rat offspring. Chlorpyrifos pesticide was chosen in this study because it is widely used in agriculture especially in France and it’s considered as an endocrine disruptor [9,10]. In vivo studies have explored the effects of CPF treatment on body weight and birth weight, some showing an increase in body weight and birth weight with CPF exposure, while others have shown a decrease [18,49].
The current study revealed a significant gain in body weight from PND1 to PND60 with the highest body weight observed in HFD group. The intergroup comparison at PND30 and PND60 shows a significantly lower body weight in the CPF-treated groups compared to the other groups. Similar results were observed by Condette et al. 2014, which revealed that rats exposed to CPF exhibited a notable increase in weight (averaging 13 g; p<0.01) and length (averaging 2 cm; p<0.01) compared to non-exposed groups [50]. In addition, the body weight of rat offspring decreases in the co-exposed (Coex) group compared to the HFD group at PND 15 and PND 30. This finding suggests the toxic effect of CPF. Several studies corroborate the observed results [18,45]. Similarly, Akhtar et al. 2009 revealed a significant body weight decrease at high dose (9 mg kg-1d -1) of chlorpyrifos [12]. Conversely, Silva in 2021 reported that male exposed to CPF showed increased body weights [51]. The perinatal exposure to CPF cause an indirect toxicity of rat offspring by leading to a hormonal disorder through the endocrine disrupting property [52]. CPF can block or alter growth hormones including GnRH which contribute to a development decline [53]. The morphological analysis of rat testis shows an abnormal testis structure including a depletion of seminiferous epithelium and some germinal lineage cells and absence specially in CPF exposed group and HFD exposed group. Similar outcome was found in the study of spooner in 2015 [54,55]. It seems that exposure to CPF may have detrimental effects on spermatogenesis and the production of viable sperm [46].
No significant difference in seminiferous tube area was observed but histopathological sign was detected. This alteration is defined by the presence of some germinal lineage cells in the tubular lumen including primary spermatocytes and cells in degeneration and an accumulation of large quantities of germinal lineage cells in the tubular lumen. The study of Gabriel et al. 2014 confirms the current results, they reported that oral ingestion of Permethrin lead to abnormalities such as disruption of the normal architecture, decrease of mature sperm cells, and reduction of luminal diameter and reduced interstitial spaces [56]. Histological analysis of ovary rat offspring shows significant increase in atretic follicle in CPF exposed group compared to control group. It seems that CPF disrupts the folliculogenesis and leads to follicle and oocyte degeneration [57]. Several studies demonstrate similar result [58,59]
Our study demonstrated the immunodetection of kisspeptin and GnRHR for the first time in the reproductive organs of rat offspring. This finding suggests the presence and potential role of kisspeptin and GnRHR in the reproductive development of rats as reported in other studies [31,34,35,42]. In ovary rat offspring, the immunodetection of kisspeptin was found in the different stages of follicles (from primordial to antral follicle) and in the corpus luteum cells. Indeed, the results of the study strongly suggest that kisspeptin plays a significant role in the regulation of folliculogenesis and suggests its involvement in regulating the development and maturation of ovarian follicles [60]. Similarly, the immunodetection of kisspeptin in testis rat offspring’s including seminiferous tubes and Leydig cells reveal the function of kisspeptin in spermatogenesis, the initiation of puberty, regulation of the male reproductive system, and overall control of testosterone secretion [30,61].
This study is the first to assess the impact of pesticide exposure and HFD on kisspeptin and GnRHR presence in the reproductive organs of rat offspring. Our findings demonstrate a reduction in the expression of GnRHR in the testis of rat offspring’s exposed to HFD. In the context of a high-fat diet, excessive intake of dietary fats can lead to an increased release of leptin, a hormone produced by adipose (fat) cells in response to the body’s energy stores [62]. Leptin receptors are present on kisspeptin neurons in the hypothalamus. When leptin binds to these receptors, it stimulates the activity of kisspeptin neurons, leading to an increase in kisspeptin expression. Kisspeptin, in turn, stimulates the release of GnRH from the hypothalamus [63]. However, over time, the constant exposure to high fat diet leads to an increase of leptin expression and as a result the increase of GnRH [63]. It’s widely known that the responsiveness of cells to specific signaling hormones includes different mechanisms to regulate homeostasis [64]. These mechanisms include receptor downregulation and receptor desensitization [65]. It seems that as a response to excessive stimulation of GnRH after the mother’s high fat diet exposure, testis cells applied a downregulation mechanism to reduce the number of GnRHR receptors Figure 12.
Figure 12.
GnRHR downregulation as a response to High fat diet (personal source).
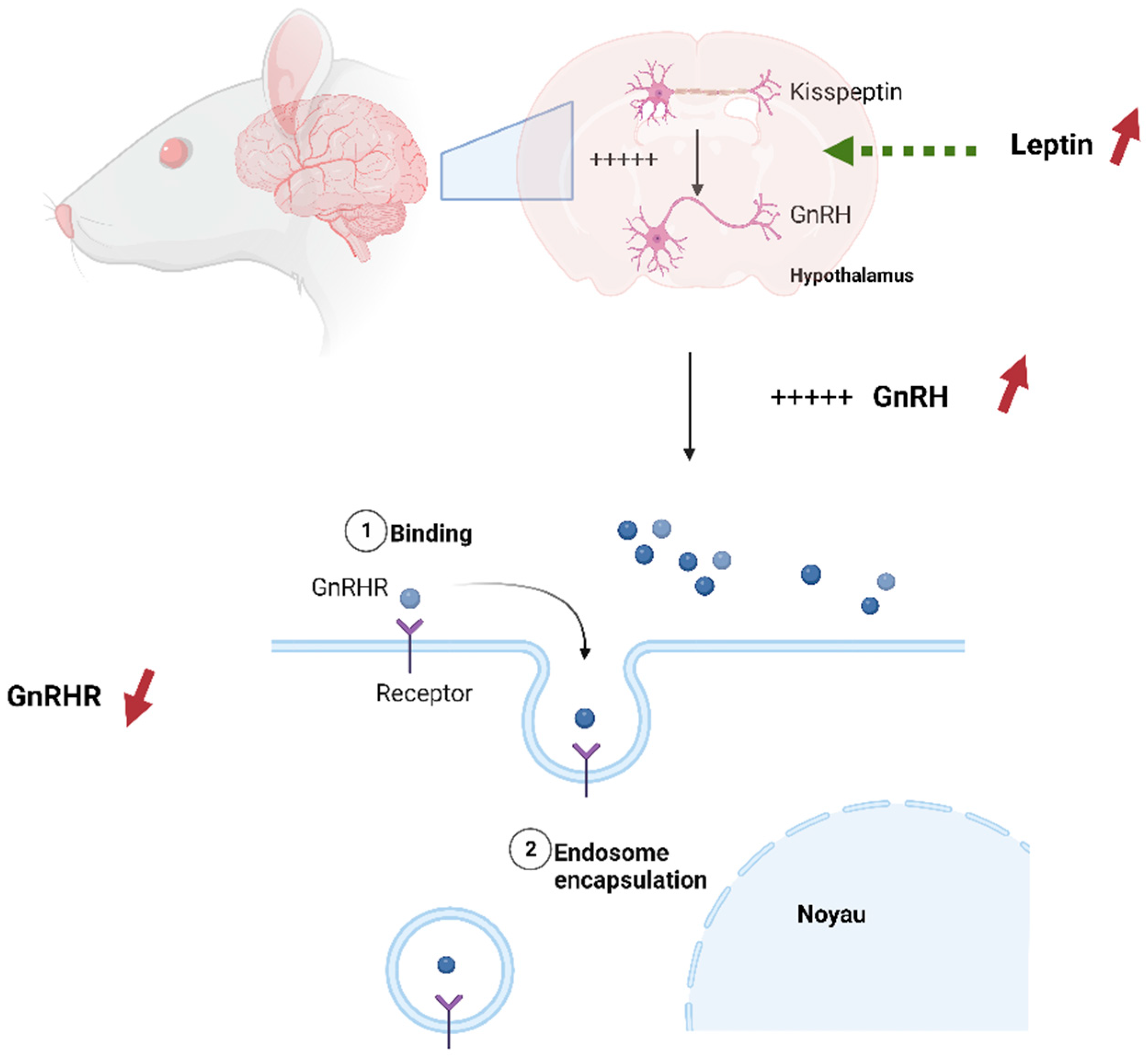
Hiller-Sturmhöfel et Bartke, 1998 defined this process by internalization of some receptors, effectively removing them from the cell surface [64]. According to CPF exposure, our result shows a negative impact of the expression of kisspeptin in the rat offspring’s testis. Chlorpyrifos, as an endocrine disruptor, alters the expression of extrahypothalamic kisspeptin in rat ‘s testis. This result is similar to the study of Johanna that reported the decrease of kisspeptin expression in cells exposed to dioxin [66,67]. Conversely it was revealed that Kiss1 expression was not affected by endocrine disruptors [68].
In ovary rat offspring, the comparison of kisspeptin and GnRHR does not exhibit persistent changes after perinatal exposure to CPF, HFD and coex. Whereas, several studies have shown a negative link between pesticide exposure and kisspeptin expression [30,36,69,70].
The strategy of this study is a chronic low dose perigestational exposure to understand the indirect effect on the progeny. Thus, our study highlights the concept “window of vulnerability” of the “Developmental Origin of Health and Disease” [17]. In other terms, exposure to food contaminants such as pesticides during the first 1000 days of life increases vulnerability to chronic diseases [71].
In the current study, the perinatal exposure of female rats to chlorpyrifos and/or a high-fat diet had no effect on the expression level of kisspeptin and GnRH receptor in the female offspring, however, it significantly impacted the male offspring. The differential response of the male and female offspring to the exposure raises questions about the underlying mechanisms and biological differences between the sexes [72]. It may indicate that there are sex-specific pathways or hormone signaling networks involved in the regulation of kisspeptin and GnRH receptor expression [73]. The DOHaD hypothesis suggests that early life exposures lead to the development of chronic diseases such as obesity, diabetes, cardiovascular disease, and even certain neurodevelopmental disorders [22]. These periods of development (prenatal and early postnatal periods) are considered as critical windows of vulnerability[20]. During these windows, cells and tissues are rapidly differentiating and developing, making them more susceptible to disruptions caused by environmental exposures [20]. The chronic low-dose exposure to CPF can have indirect effects on offspring when it occurs through maternal exposure during pregnancy[18]. CPF can cross the placental barrier and reach the developing fetus and leads to an indirect health alteration of offspring which can develop in long-term health Impact[74]. In addition, our findings suggest the implication of non-genetic mechanisms “epigenetics” that might explain these changes in male offspring in response to maternal environment during early development and postnatal period [21]. Exposure to CPF during pregnancy can also influence gene expression patterns in the fetus through epigenetic mechanisms [21]. Understanding these sex-specific differences could be crucial for developing targeted interventions or therapies to mitigate the potential negative impacts of environmental factors on reproductive health [75].
A significant severe reduction in cell number was demonstrated in the offspring’s ovary of the rat exposed to CPF. It seems that the CPF induces an oxidative stress in ovary tissue leading to an increase in ROS levels within the ovarian tissue. These ROS are highly reactive and can cause damage to cellular components, such as lipids, proteins, and DNA. As a result, the delicate balance within the ovarian cells is disrupted. Apoptosis process is initiated as a protective response to eliminate cells that are severely affected by the ROS-induced damage[76,77,78,79].
5. Conclusions
The present study reveals that perinatal exposure of female rats to chlorpyrifos and/or a high-fat diet induce a histopathological impact on reproductive organs (testis and ovary) in the offspring. Notably, the presence level of kisspeptin and GnRH receptor in testis rat offspring declined in CPF group and HFD group respectively indicating that offspring’s testis is more sensitive to obesity and pesticide exposure than offspring’s ovary. This study required us to better understand the link between environmental factors, obesity, and reproductive function.
6. Limitations and perspectives
To investigate the impact of pesticide exposure and a high-fat diet on reproductive organ expression, we used an animal model. While it can provide valuable insights, there may be differences between rats and humans, making it necessary to interpret the results with caution when applying them to humans. The study exposes rats to a specific dose of the pesticide chlorpyrifos (CPF) and a high-fat diet (HFD). However, it is important to note that the chosen exposure levels may not accurately reflect real-world exposure scenarios for humans, which can vary significantly. In fact, exposure to pesticide residues is not limited to one pesticide but a mixture of organophosphates, carbamates, pyrethroids, etc. In consequence, it is of real importance to study the effect of a cocktail of the common pesticides residues we are orally exposed and evaluate the impact compared to that of one pesticide[80].
We aim to use in silico analysis software to compare the expression of kisspeptin and GnRHR in different groups. While this approach can provide valuable insights, it relies on computational models and may have limitations in accurately capturing the complexity of biological systems. Understanding the impact of pesticide exposure and dietary factors on reproductive organ expression may have implications for reproductive health. The findings of this study may contribute to the development of preventive strategies or interventions to mitigate the potential negative effects on reproductive function.
Author Contributions
“Conceptualization, H.KC. and M.B.; methodology, M.L.; software, M.L., IL.; validation, A.C., H.KC. and M.L.; formal analysis, V.B.; investigation, N.D.; resources, S.T.; data curation, N.D.; writing—original draft preparation, M.L.; writing—review and editing, H.KC. and M.BK.; visualization, M.A. and H.BA.; supervision, H.KC. and M.BK.; project administration, H.KC.; funding acquisition, M.BK. All authors have read and agreed to the published version of the manuscript.”
Funding
“The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.”
Institutional Review Board Statement
The study protocol obtained approval from the Regional Directorate for Health, Animal and Environment Protection, an accredited organization in France. Additionally, it was also approved by the French Ministry of Research under the reference number APAFIS#8207-2016121322563594 v2.
Data Availability Statement
Not applicable.
Acknowledgments
the authors thank the MP3CV laboratory for providing Tissuefax, HistoQuest software to assess the insilico quantification.
Conflicts of Interest
The authors have no conflict of interest to be declared.
References
- Cooper TG, Noonan E, von Eckardstein S, Auger J, Baker HWG, Behre HM, et al. World Health Organization reference values for human semen characteristics*‡. Human Reproduction Update. 2010;16:231–45. [CrossRef]
- Baker KC, Steiner AZ, Hansen KR, Barnhart KT, Cedars MI, Legro RS, et al. Poor reproducibility of percentage of normally shaped sperm using the World Health Organization Fifth Edition strict grading criteria. F S Rep. 2022;3:110–5. [CrossRef]
- Agarwal A, Mulgund A, Hamada A, Chyatte MR. A unique view on male infertility around the globe. Reproductive Biology and Endocrinology. 2015;13:37. [CrossRef]
- Infertility around the globe: new thinking on gender, reproductive technologies and global movements in the 21st century | Human Reproduction Update | Oxford Academic [Internet]. [cited 2022 Sep 12]. Available from: https://academic.oup.com/humupd/article/21/4/411/683746.
- Noppakun K, Juntarawijit C. Association between pesticide exposure and obesity: A cross-sectional study of 20,295 farmers in Thailand. F1000Res. 2022;10:445.
- Djekkoun N, Lalau J-D, Bach V, Depeint F, Khorsi-Cauet H. Chronic oral exposure to pesticides and their consequences on metabolic regulation: role of the microbiota. Eur J Nutr. 2021;60:4131–49. [CrossRef]
- Segal TR, Giudice LC. Before the beginning: environmental exposures and reproductive and obstetrical outcomes. Fertility and Sterility. 2019;112:613–21. [CrossRef]
- Lahimer M, Capelle S, Lefranc E, Cabry R, Montjean D, Bach V, et al. Effect of pesticide exposure on human sperm characteristics, genome integrity, and methylation profile analysis. Environ Sci Pollut Res [Internet]. 2023 [cited 2023 Jun 22]; Available from: https://doi.org/10.1007/s11356-023-27695-7. [CrossRef]
- Travaux et implication de l’Anses sur les perturbateurs endocriniens [Internet]. Anses - Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail. 2019 [cited 2023 Apr 7]. Available from: https://www.anses.fr/fr/content/travaux-et-implication-de-lanses-sur-les-perturbateurs-endocriniens.
- US EPA O. Overview of Endocrine Disruption [Internet]. 2015 [cited 2023 Apr 10]. Available from: https://www.epa.gov/endocrine-disruption/overview-endocrine-disruption.
- U.S. Environmental Protection Agency | US EPA [Internet]. [cited 2023 Feb 25]. Available from: https://www.epa.gov/.
- Akhtar N, Srivastava MK, Raizada RB. Assessment of chlorpyrifos toxicity on certain organs in rat, Rattus norvegicus. 2009;
- Wołejko E, Łozowicka B, Jabłońska-Trypuć A, Pietruszyńska M, Wydro U. Chlorpyrifos Occurrence and Toxicological Risk Assessment: A Review. Int J Environ Res Public Health. 2022;19:12209. [CrossRef]
- Greer JB, Magnuson JT, Hester K, Giroux M, Pope C, Anderson T, et al. Effects of Chlorpyrifos on Cholinesterase and Serine Lipase Activities and Lipid Metabolism in Brains of Rainbow Trout (Oncorhynchus mykiss). Toxicol Sci. 2019;172:146–54. [CrossRef]
- Abou Diwan M, Lahimer M, Bach V, Gosselet F, Khorsi-Cauet H, Candela P. Impact of Pesticide Residues on the Gut-Microbiota–Blood–Brain Barrier Axis: A Narrative Review. International Journal of Molecular Sciences. 2023;24:6147. [CrossRef]
- Colović MB, Krstić DZ, Lazarević-Pašti TD, Bondžić AM, Vasić VM. Acetylcholinesterase inhibitors: pharmacology and toxicology. Curr Neuropharmacol. 2013;11:315–35.
- Grandjean P, Barouki R, Bellinger DC, Casteleyn L, Chadwick LH, Cordier S, et al. Life-Long Implications of Developmental Exposure to Environmental Stressors: New Perspectives. Endocrinology. 2015;156:3408–15. [CrossRef]
- Djekkoun N, Depeint F, Guibourdenche M, El Khayat El Sabbouri H, Corona A, Rhazi L, et al. Chronic Perigestational Exposure to Chlorpyrifos Induces Perturbations in Gut Bacteria and Glucose and Lipid Markers in Female Rats and Their Offspring. Toxics. 2022;10:138. [CrossRef]
- Mandy M, Nyirenda M. Developmental Origins of Health and Disease: the relevance to developing nations. Int Health. 2018;10:66–70. [CrossRef]
- Selevan SG, Kimmel CA, Mendola P. Identifying critical windows of exposure for children’s health. Environ Health Perspect. 2000;108:451–5. [CrossRef]
- Safi-Stibler S, Gabory A. Epigenetics and the Developmental Origins of Health and Disease: Parental environment signalling to the epigenome, critical time windows and sculpting the adult phenotype. Seminars in Cell & Developmental Biology. 2020;97:172–80. [CrossRef]
- Lacagnina S. The Developmental Origins of Health and Disease (DOHaD). Am J Lifestyle Med. 2019;14:47–50. [CrossRef]
- Montjean D, Neyroud A-S, Yefimova MG, Benkhalifa M, Cabry R, Ravel C. Impact of Endocrine Disruptors upon Non-Genetic Inheritance. International Journal of Molecular Sciences. 2022;23:3350. [CrossRef]
- Authority (EFSA) EFS, Carrasco Cabrera L, Medina Pastor P. The 2019 European Union report on pesticide residues in food. EFSA Journal. 2021;19:e06491. [CrossRef]
- Marques-Pinto A, Carvalho D. Human infertility: are endocrine disruptors to blame? Endocr Connect. 2013;2:R15–29. [CrossRef]
- Padda J, Khalid K, Moosa A, Syam M, Kakani V, Imdad U, et al. Role of Kisspeptin on Hypothalamic-Pituitary-Gonadal Pathology and Its Effect on Reproduction. Cureus. 13:e17600. [CrossRef]
- Rønnekleiv OK, Kelly MJ. Kisspeptin Excitation of GnRH Neurons. Adv Exp Med Biol. 2013;784:113–31. [CrossRef]
- Prashar V, Arora T, Singh R, Sharma A, Parkash J. Hypothalamic Kisspeptin Neurons: Integral Elements of the GnRH System. Reprod Sci. 2023;30:802–22. [CrossRef]
- Han S-K, Gottsch ML, Lee KJ, Popa SM, Smith JT, Jakawich SK, et al. Activation of Gonadotropin-Releasing Hormone Neurons by Kisspeptin as a Neuroendocrine Switch for the Onset of Puberty. J Neurosci. 2005;25:11349–56. [CrossRef]
- Feng J, Xu R, Li Y, Zhou Q, Song G, Deng Y, et al. The effect of high-fat diet and exercise on KISS-1/GPR54 expression in testis of growing rats. Nutrition & Metabolism. 2021;18:1. [CrossRef]
- Wang J-Y, Hsu M-C, Tseng T-H, Wu L-S, Yang K-T, Chiu C-H. Kisspeptin expression in mouse Leydig cells correlates with age. Journal of the Chinese Medical Association. 2015;78:249–57. [CrossRef]
- Bhattacharya M, Babwah AV. Kisspeptin: Beyond the Brain. Endocrinology. 2015;156:1218–27. [CrossRef]
- UENOYAMA Y, PHENG V, TSUKAMURA H, MAEDA K. The roles of kisspeptin revisited: inside and outside the hypothalamus. J Reprod Dev. 2016;62:537–45. [CrossRef]
- Rajin TR, Patra MK, Sheikh PA, Singh AK, Mishra GK, Karikalan M, et al. Expression of kisspeptin and its receptor in different functional classes of ovarian follicle in the buffalo (Bubalus bubalis). Theriogenology. 2022;179:87–96. [CrossRef]
- Tanyapanyachon P, Amelkina O, Chatdarong K. The expression of kisspeptin and its receptor in the domestic cat ovary and uterus in different stages of the ovarian cycle. Theriogenology. 2018;117:40–8. [CrossRef]
- Zeydabadi Nejad S, Ramezani Tehrani F, Zadeh-Vakili A. The Role of Kisspeptin in Female Reproduction. Int J Endocrinol Metab. 2017;15:e44337.
- Szeliga A, Meczekalski B. Kisspeptin Modulation of Reproductive Function. Endocrines. 2022;3:367–74. [CrossRef]
- Xie Q, Kang Y, Zhang C, Xie Y, Wang C, Liu J, et al. The Role of Kisspeptin in the Control of the Hypothalamic-Pituitary-Gonadal Axis and Reproduction. Front Endocrinol (Lausanne). 2022;13:925206. [CrossRef]
- Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Shagoury JK, et al. The GPR54 gene as a regulator of puberty. N Engl J Med. 2003;349:1614–27. [CrossRef]
- Abdelkareem AO, Gebril SM, AbdelHafez FF, Terry J, Bedaiwy MA. Kisspeptin and kisspeptin receptor immunoreactivity in euploid and aneuploid choriodecidual tissues of recurrent pregnancy losses. F S Sci. 2023;4:56–64. [CrossRef]
- SENGUPTA A, CHAKRABARTI N, SRIDARAN R. Presence of Immunoreactive Gonadotropin Releasing Hormone (GnRH) and Its Receptor (GnRHR) in Rat Ovary During Pregnancy. Mol Reprod Dev. 2008;75:1031–44. [CrossRef]
- Ciaramella V, Chianese R, Pariante P, Fasano S, Pierantoni R, Meccariello R. Expression Analysis of Gnrh1 and Gnrhr1 in Spermatogenic Cells of Rat. International Journal of Endocrinology. 2015;2015:e982726. [CrossRef]
- Palmon A, Ben Aroya N, Tel-Or S, Burstein Y, Fridkin M, Koch Y. The gene for the neuropeptide gonadotropin-releasing hormone is expressed in the mammary gland of lactating rats. Proc Natl Acad Sci U S A. 1994;91:4994–6. [CrossRef]
- Cheung LWT, Wong AST. Gonadotropin-releasing hormone: GnRH receptor signaling in extrapituitary tissues. FEBS J. 2008;275:5479–95. [CrossRef]
- El Khayat El Sabbouri H, Gay-Quéheillard J, Joumaa WH, Delanaud S, Guibourdenche M, Darwiche W, et al. Does the perigestational exposure to chlorpyrifos and/or high-fat diet affect respiratory parameters and diaphragmatic muscle contractility in young rats? Food Chem Toxicol. 2020;140:111322. [CrossRef]
- Elbetieha A, Da’as SI, Khamas W, Darmani H. Evaluation of the toxic potentials of cypermethrin pesticide on some reproductive and fertility parameters in the male rats. Arch Environ Contam Toxicol. 2001;41:522–8. [CrossRef]
- Dudás J, Dietl W, Romani A, Reinold S, Glueckert R, Schrott-Fischer A, et al. Nerve Growth Factor (NGF)—Receptor Survival Axis in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2018;19:1771. [CrossRef]
- Huang Y, Sun W, Gao F, Ma H, Yuan T, Liu Z, et al. Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats. Biology. 2023;12:760. [CrossRef]
- Acker CI, Nogueira CW. Chlorpyrifos acute exposure induces hyperglycemia and hyperlipidemia in rats. Chemosphere. 2012;89:602–8. [CrossRef]
- Condette CJ, Khorsi-Cauet H, Morlière P, Zabijak L, Reygner J, Bach V, et al. Increased Gut Permeability and Bacterial Translocation after Chronic Chlorpyrifos Exposure in Rats. PLOS ONE. 2014;9:e102217. [CrossRef]
- Silva MH. Chlorpyrifos and Δ9 Tetrahydrocannabinol exposure and effects on parameters associated with the endocannabinoid system and risk factors for obesity. Curr Res Toxicol. 2021;2:296–308. [CrossRef]
- Travaux et implication de l’Anses sur les perturbateurs endocriniens [Internet]. Anses - Agence nationale de sécurité sanitaire de l’alimentation, de l’environnement et du travail. 2019 [cited 2023 Apr 10]. Available from: https://www.anses.fr/fr/content/travaux-et-implication-de-lanses-sur-les-perturbateurs-endocriniens.
- Tzoupis H, Nteli A, Androutsou M-E, Tselios T. Gonadotropin-Releasing Hormone and GnRH Receptor: Structure, Function and Drug Development. Curr Med Chem. 2020;27:6136–58. [CrossRef]
- Miranda-Spooner M, Paccola CC, Neves FMO, de Oliva SU, Miraglia SM. Late reproductive analysis in rat male offspring exposed to nicotine during pregnancy and lactation. Andrology. 2016;4:218–31. [CrossRef]
- Russell LD, Ettlin RA, Hikim APS, Clegg ED. Histological and Histopathological Evaluation of the Testis. International Journal of Andrology. 1993;16:83–83. [CrossRef]
- Omotoso G, Onanuga I, Ibrahim R. Histological effects of permethrin insecticide on the testis of adult wistar rats. Ibnosina Journal of Medicine and Biomedical Sciences. 2014;06:125–9. [CrossRef]
- Merviel P, Cabry R, Chardon K, Haraux E, Scheffler F, Mansouri N, et al. Impact of oocytes with CLCG on ICSI outcomes and their potential relation to pesticide exposure. J Ovarian Res. 2017;10:42. [CrossRef]
- Gupta RK, Miller KP, Babus JK, Flaws JA. Methoxychlor Inhibits Growth and Induces Atresia of Antral Follicles through an Oxidative Stress Pathway. Toxicological Sciences. 2006;93:382–9. [CrossRef]
- Corriero A, Zupa R, Mylonas CC, Passantino L. Atresia of ovarian follicles in fishes, and implications and uses in aquaculture and fisheries. Journal of Fish Diseases. 2021;44:1271–91. [CrossRef]
- Gaytan F, Garcia-Galiano D, Dorfman MD, Manfredi-Lozano M, Castellano JM, Dissen GA, et al. Kisspeptin Receptor Haplo-insufficiency Causes Premature Ovarian Failure Despite Preserved Gonadotropin Secretion. Endocrinology. 2014;155:3088–97. [CrossRef]
- Meccariello R. The Kisspeptin System in Male Reproduction. Endocrines. 2022;3:168–74. [CrossRef]
- Izadi V, Saraf-Bank S, Azadbakht L. Dietary intakes and leptin concentrations. ARYA Atheroscler. 2014;10:266–72.
- Childs GV, Odle AK, MacNicol MC, MacNicol AM. The Importance of Leptin to Reproduction. Endocrinology. 2020;162:bqaa204. [CrossRef]
- Hiller-Sturmhöfel S, Bartke A. The Endocrine System. Alcohol Health Res World. 1998;22:153–64.
- Shankaran H, Wiley HS, Resat H. Receptor downregulation and desensitization enhance the information processing ability of signalling receptors. BMC Syst Biol. 2007;1:48. [CrossRef]
- Mueller JK, Heger S. Endocrine disrupting chemicals affect the Gonadotropin releasing hormone neuronal network. Reproductive Toxicology. 2014;44:73–84. [CrossRef]
- Roepke TA, Sadlier NC. REPRODUCTIVE TOXICOLOGY: Impact of endocrine disruptors on neurons expressing GnRH or kisspeptin and pituitary gonadotropins. Reproduction. 2021;162:F131–45. [CrossRef]
- Dickerson SM, Cunningham SL, Gore AC. Prenatal PCBs disrupt early neuroendocrine development of the rat hypothalamus. Toxicology and Applied Pharmacology. 2011;252:36–46. [CrossRef]
- Xu R, Feng J, Liang C, Song G, Yan Y. Effects of high-fat diet and treadmill running on the hypothalamic Kiss-1-GPR54 signaling pathway in male growing rats. Hormones (Athens). 2022;21:641–52. [CrossRef]
- Liu R, Li H, Zhang D, Lv Y, Lin X, Zhang H. Expression of Kisspeptin/kiss1r System is Down-regulated in the Hypothalamic Arcuate Nucleus of Pubertal Male Rats with High-fat-diet. Journal of Reproduction and Contraception. 2014;25:1–11. [CrossRef]
- Alabduljabbar S, Zaidan SA, Lakshmanan AP, Terranegra A. Personalized Nutrition Approach in Pregnancy and Early Life to Tackle Childhood and Adult Non-Communicable Diseases. Life (Basel). 2021;11:467. [CrossRef]
- Yehuda R, Lehrner A. Intergenerational transmission of trauma effects: putative role of epigenetic mechanisms. World Psychiatry. 2018;17:243–57. [CrossRef]
- Lee EB, Dilower I, Marsh CA, Wolfe MW, Masumi S, Upadhyaya S, et al. Sexual Dimorphism in Kisspeptin Signaling. Cells. 2022;11:1146. [CrossRef]
- Dugershaw BB, Aengenheister L, Hansen SSK, Hougaard KS, Buerki-Thurnherr T. Recent insights on indirect mechanisms in developmental toxicity of nanomaterials. Part Fibre Toxicol. 2020;17:31. [CrossRef]
- Sommer M, Mmari K. Addressing Structural and Environmental Factors for Adolescent Sexual and Reproductive Health in Low- and Middle-Income Countries. Am J Public Health. 2015;105:1973–81. [CrossRef]
- Chargui I, Grissa I, Bensassi F, Hrira MY, Haouem S, Haouas Z, et al. Oxidative Stress, Biochemical and Histopathological Alterations in the Liver and Kidney of Female Rats Exposed to Low Doses of Deltamethrin (DM): A Molecular Assessment. Biomedical and Environmental Sciences. 2012;25:672–83. [CrossRef]
- 77. El-Din MAE-DS, Ghareeb AE-WE, El-Garawani IM, El-Rahman HAA. Induction of apoptosis, oxidative stress, hormonal, and histological alterations in the reproductive system of thiamethoxam-exposed female rats. Environ Sci Pollut Res. 2023;30:77917–30. [CrossRef]
- Watermann B, Grote K, Gnass K, Kolodzey H, Thomsen A, Appel KE, et al. Histological alterations in ovaries of pubertal female rats induced by triphenyltin. Experimental and Toxicologic Pathology. 2008;60:313–21. [CrossRef]
- Grewal KK, Sandhu GS, Kaur R, Brar RS, Sandhu HS. Toxic Impacts of Cypermethrin on Behavior and Histology of Certain Tissues of Albino Rats. Toxicol Int. 2010;17:94–8. [CrossRef]
- Lukowicz C, Ellero -Simatos Sandrine, R égnier M, Polizzi A, Lasserre F, Montagner A, et al. Metabolic Effects of a Chronic Dietary Exposure to a Low-Dose Pesticide Cocktail in Mice: Sexual Dimorphism and Role of the Constitutive Androstane Receptor. Environmental Health Perspectives. 126:067007. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



