
Submitted:
24 August 2023
Posted:
25 August 2023
You are already at the latest version
Abstract
Fractures are the most common large organ trauma in humans. The initial inflammatory re-sponse promotes bone healing during the initial post-fracture phase, but chronic and persistent inflammation due to infection or other factors does not contribute to the healing process. The precise mechanisms by which immune cells and their cytokines are regulated in bone healing remain unclear. The use of mesenchymal stem cells (MSCs) for cellular therapy of bones injury is a novel clinical treatment approach. Bone progenitor MSCs not only differentiate into bone, but also interact with the immune system to promote the healing process. We review in vitro and in vivo studies on the role of the immune system and bone marrow MSCs in bone healing and their interactions. A deeper understanding of this paradigm may provide clues to potential therapeu-tic targets in the healing process, thereby improving the reliability and safety of clinical applica-tions of MSCs to promote bone healing.
Keywords:
bone fracture
; MSCs
; immune cells
1. Introduction
In 2019, the results of a European human clinical trial showed that surgical treatment using bioceramics particles combined with autologous mesenchymal stem cells (MSCs) for patients with nonunion of tibia, femur, and humerus is safe and feasible [1]. Another clinical study showed that BM-MSCs enriched by the screen-enrich-combine circulating system (SECCS) for surgical treatment promoted bone regeneration in patients with post-fracture nonunion [2]. As of January 2019, there have been more than 700 clinical trials using MSCs transplants as an alternative to common therapies for better outcomes [3].
MSCs can promote the migration of autologous-BM-MSCs and nerve growth at the fracture site in the treatment of fracture nonunion [4,5], and differentiate to chondrocytes and osteoblasts under certain induction, and also promote the differentiation of osteoblasts [6], thus promoting bone remodeling [7]. Compared with common therapy, the rate of bone healing is greatly accelerated and the success rate of fracture healing is improved, although MSC treatment of fractures is not 100% cured [8]. Calori, G.M., et al. indicated that the healing rate of fracture treated with MSC was close to 90% [9]. The experiment of Chu, W., et al. showed that out of the 16 patients who received mesenchymal stem cell/β-tricalcium phosphate composites(MSC/β-TCP), 15 achieved excellent or good recovery within 2 years, and out of the 23 patients who received porous β-TCP alone, 14 achieved good recovery, and during follow-up, one patient treated with porous β-TCP alone had a Lysholm score of less than 60, indicating poor recovery [10]. The current clinical trial results show that MSCs can be used for fracture healing, and the healing effect is good, moreover the cure rate is also high.
The process of bone regeneration is intricate and well-coordinated, with fracture healing being a multi-stage process. The involvement of immune cells in this process is crucial, although their primary activity occurs during the initial stages of fracture healing (Figure 1) [11]. Throughout different phases of bone healing, cytokines that possess both inflammatory and immunity-regulating functions, such as Interleukin (IL)-1β [12], IL-6 [13,14], IL-17F [12,15], IL-23 [12], and tumor necrosis factor (TNF) [16,17], play significant roles. Acute inflammation following bone injury promotes fracture healing. B cells infiltrate fracture healing tissue in large numbers shortly after injury and differentiate into plasma cells that secrete a large number of factors, including osteoprotegerin, to promote bone healing by inhibiting osteoclastogenesis [18]. Inflammatory signaling by TNF-α, IL-1 and other pro-inflammatory cytokines has also been shown to exert a pro-regenerative function through injury-induced changes in the tissue microenvironment (such as expression of cell surface receptors) [19]. Activation of the inflammatory response drives NF-κB-mediated gene expression to further amplify inflammation [20]. However, chronic long-term inflammation has an inhibitory effect on fracture healing [21]. For example, TNF-α and IL-1 are directly associated with bone destruction in patients with arthritis. In mice with inflammatory arthritis, treatment with TNF-α inhibitors enhanced healing, and lack of IL-1 due to genetic mutations prevented bone and joint disease [22]. It has also been shown that the chronic inflammatory environment caused by peri-implantitis, on the one hand, recruits a large number of neutrophils and induces degradation of the extracellular matrix, leading to more differentiation of MSCs towards adipocytes than osteoblasts, and on the other hand, ROS are involved in the chemotaxis of MSCs towards adipocytes, and eventually the osteolysis process is activated due to the imbalance of osteogenic effects [23]. Immune cells of the innate immune system such as neutrophils, macrophages, monocytes, and natural killer cells are activated and recruited to the fracture site together with a variety of cytokines, thus forming an inflammatory response to create conditions for initial fracture healing. osteoprogenitor mesenchymal stem cells are also involved in this process [24,25].
The immune cells and related cytokines in the acquired immune response also play an important regulatory role in promoting the progress of the healing [26,27]. For example, T helper (Th) 1 cells are polarized by IL-12 and secrete interferon (IFN)-γ, IL-2 and TNF-α, and mainly affect macrophages. For example, TNF-α mediates increased receptor activator of nuclear factor kappa-B ligand (RANKL) expression by macrophages, thereby stimulating osteoclastogenesis [28]. B cells produce approximately 40–60% of the total bone marrow-derived osteoprotegerin (OPG) and thereby inhibit osteoclast differentiation [29]. MSCs participate in both innate and adaptive immunity, and their immunomodulatory functions are exerted mainly via interactions with immune cells through cell-to-cell contact and paracrine activity [30,31,32].
Transplantation of allogeneic bone marrow mesenchymal stem cells into mice has been shown to induce a Th1 type immune response, with significant increases in T cells, B cells, macrophages and IFN-γ, which inhibit bone formation. This was due to significant inhibition of the expression of osteocalcin, runt-related transcription factor 2 (Runx2) and alkaline phosphatase genes in the implants [33]. Liu et al. reported that Pro-inflammatory T cells secrete large amounts of IFN-γ and TNF-α, and induce apoptosis of MSCs through the combined effect of down-regulating RUNX2 pathway and enhancing TNF-α signaling [34]. Some scholars have also found that delayed fracture healing is significantly correlated with the enrichment of CD8+T cells in hematoma [35].This is consistent with these findings in animal studies.
In this review, we discussed the current understanding of the interaction between MSCs and the immune system in fracture repair.
2. Interaction between MSCs and Macrophages
2.1. Effects of macrophages on MSC
Pro-inflammatory cytokines (TNF-α, IL-6, IL-1β, and IFN-γ) and oncostatin M (OSM) stimulate the osteogenic differentiation of MSCs through OSM and nuclear factor-kappa B (NF-κB) pathways [36,37]. In addition, Cu- mesoporous silica nanosphere (MSN)/macrophage conditioned medium (CM) up-regulates OPG in BM-MSCs and down-regulates RANKL to inhibit osteoclast formation [38]. In the co-culture experiment, macrophages phagocytosed by carbon nanohorn-(CNH-) also express OSM, which accelerates the osteogenic differentiation of MSCs through the STAT3 signaling pathway [37]. Lu et al. demonstrated that lipopolysaccharide (LPS)-induced M1 macrophages promote osteogenic formation through the cyclooxygenase-2 (COX2)-prostaglandin E2 (PGE2) pathway [39]. Tu et al. explained the stimulating effect of pro-inflammatory macrophages on MSC osteogenesis from another perspective. IL-23 secreted by macrophages directly induces the bone formation of BM-MSCs by activating STAT3 and β-catenin. When the IL-23 p19 antibody neutralized IL-23 in macrophage CM, the calcium formation and Alkaline Phosphatase (ALP) activity of MSCs were reduced [40].
However, in the study of Gong et al., M2 macrophages enhanced the osteogenic differentiation of BM-MSCs, while M1 macrophages disrupted their osteogenic differentiation. Pro-regenerative cytokines, such as transforming growth factor (TGF)-β, vascular endothelial growth factor (VEGF), and Insulin-like growth factor 1 (IGF-1), are produced by M2 macrophages, and harmful inflammatory cytokines, such as IL-6, IL-12, and TNF-α, are produced by M1 macrophages. It is suspected to be a mechanism that regulates osteogenic differentiation [41]. Meanwhile, TGF-β and VEGF have been shown to have angiogenic activity [42].
2.2. Effects of MSCs on polarization of macrophages
MSCs also affect macrophage subtypes. The implanted MSCs generated a cascade of events, the recruitment of BM-derived specific progenitors with vasculogenic and osteogenic properties, resulting in the mobilization of cells of the innate immune system, such as macrophages, and the induction of their functional switch from a pro-inflammatory to an anti-inflammatory proresolving phenotype. MSCs induce M1 to M2 macrophages is attributed to the activation of the NF-κB pathway by PGE2 secreted by exogenous MSCs [43]. Li et al. also found the transformation from M1 to M2 in the study of bone defect with the bone-inducing material laponite (LAP). Although Lap contributes to bone regeneration, it is still related to inflammation as a foreign body. They found that MSCs transformed Lap-induced M1 macrophages into M2 phenotypes, creating an anti-inflammatory/pro-lytic environment that promotes bone formation [44]. However, the MSCs were not detected 4 weeks after transplantation, suggesting that early MSCs may regulate the polarization of macrophages [45,46]. Related literature reports that macrophages are transformed into M2 type through cytokine-pretreated MSC and IL-4 secreting MSC [47,48]. Although pre-treated BM-MSCs and IL-4 secreting BM-MSCs can promote bone formation, the time of bone regeneration in vitro has a significant impact. After co-culture with macrophages, the pretreated MSC promoted bone regeneration in the early stage (day 3), while the IL-4 secreting MSC played a role in the later stage (day 7). Based on the secretion of IL-4 and PGE2, IL-4 secreting MSCs also have the greater immunomodulatory ability in the transformation from M1 to M2 [49,50]. The interaction between MSC and macrophages is shown in Figure 2.
Macrophages regulate the recruitment and differentiation of the MSCs. On the upper part of the image are shown the best known macrophage pro-inflammatory cytokines, and osteoinductive factors involved in the regulation of MSC functions. MSCs reciprocally regulate macrophage recruitment and function, usually with PGE2 and IL-4 mediated transformation of macrophages from M1 type to M2 type.
3. MSCs inhibit Dendritic cells (DCs)
As an antigen presenting cell(APC), DCs phagocytose and processes antigens before presenting them to T lymphocytes via MHC molecules on the cell surface to activate an acquired immune response [51]. Differentiation of monocytes into DCs secretes IL-12, which can help naive CD4+T cells differentiate into Th1 cells. It has been shown that allogeneic BM-MSCs inhibit the ability of monocytes or CD34+ hematopoietic progenitors to differentiate into DCs, as well as their ability to secrete pro-inflammatory cytokines [52]. Meanwhile, allogeneic BM-MSC can promote the release of anti-inflammatory factors and inhibit the polarization of initial CD4+ lymphocytes into pro-inflammatory cells [53,54]. Similar to T lymphocytes, allogeneic MSCs may also inhibit DC function by means of cell-cell contact [55]. This process, in which MSCs secrete TGF-β1, simultaneously decreases the expression of DC co-stimulatory molecules such as CD80 and CD86 [56,57]. One study found that when autologous or homologous MSCs were co-cultured with dendritic cells, dendritic cells induced differentiation of primitive T cells to classical Treg [56]. Another study found that when monocyte-derived DCs were co-cultured with allogeneic MSCs, DCs were more likely to become myeloid-derived suppressor cell (MDSC)-like phenotypes in response to the growth-regulating oncogene γ (GRO-γ) chemokines secreted by MSCs. Dendritic cells co-cultured with autologous or homozygous MSCs can also induce initial T-cell differentiation to classical Treg [56]. Homozygous MSCs co-cultured with monocyte-derived dendritic cells secrete the growth-regulating oncogene γ (GRO-γ) chemokines, which promotes the development of DCs into a myeloid-derived suppressor cell (MDSC) -like phenotype. GRO-γ–treated MDSCs had a tolerogenic phenotype that was characterized by an increase in the secretion of IL-10 and IL-4, and a reduction in the production of IL-12 and IFN-γ [58]. Recently, it has been proposed that MSC mediates the up-regulation of the suppressor of cytokine signaling 1 (SOCS1) gene through IL-6, which indicates that DC acquires a tolerant phenotype, IL-10 production is significantly increased, and can induce Treg and Th2 differentiation [59].
4. Interaction between MSCs and Natural killer (NK) cells
NK cells belong to the core cells of the natural immune system. NK cells, unlike all other white blood cells, are capable of recognizing and attacking foreign cells, cancer cells and viruses on their own. On one side, secretion of inflammatory cytokines secreted by NK cells can be inhibited by MSCs, such as IFN-γ or TNF-α, on the other side, intercellular factors secreted by MSCs can inhibit NK cell activity [60]. TGF-β and IL-6 produced by activated MSCs limit NK cell effector function but promote NK cell differentiation [61]. In addition, experimental data have suggested that MSCs can improve the function of NK cells after severe injury [62].
NK cells are able to be activated not only by recognizing cells lacking major histocompatibility complex (MHC) Class I surface molecules, but also by cytokines including IL-2 and IL-12 [63]. Activated NK cells induce apoptosis mainly by releasing cytotoxic particles including perforin, NK cytotoxic factor, TNF and so on [63]. Allogeneic BM-MSCs sharply inhibit IL-2-induced resting NK cell proliferation, while had little effect on activated NK cell proliferation [64]. The same study demonstrated that IL-2-activated NK cells initiate NK cell-mediated MSCs cytotoxicity by binding to specific ligands on MSCs, which effectively degrades endogenous and exogenous MSCs. However, IFN-γ can inhibit this cleavage by using up-regulated human leukocyte antigen (HLA) class I molecules [64,65]. A recent study has shown that allogeneic MSCS can adapt their immune behavior in the inflammatory environment by activating toll-like receptor 3 (TLR3) to resist the killing effect of IL-2 activated NK cells [66].
5. Interaction between MSCs and Neutrophils
In the process of healing fractures, MSCs have been found to exert inhibitory effects on various immune cells, with the exception of neutrophils [67]. MSCs inhibit neutrophil apoptosis, alter their chemotaxis, and enhance respiratory burst capacity [68,69,70]. When activated, MSCs work synergistically by producing IL-6, IFN-γ and granulocyte-macrophage colony-stimulating factor (GM-CSF) to effectively delay neutrophil death [71]. Furthermore, MSCs secrete IL-8 and macrophage migration inhibitory factor (MIF) to attract neutrophils and facilitate their infiltration into sites of inflammation. As a result of this recruitment process, the recruited neutrophils exhibit heightened antimicrobial activity and an increased respiratory burst capacity [69,72]. In a corneal injury model and a peritonitis mouse model, TNF alpha induced protein 6(TSG-6) secreted by MSCs effectively inhibits the entry of neutrophils into the injury site, thereby significantly reducing inflammatory responses [73,74]. MSCs also suppress neutrophils through multiple pathways to decrease immune responses and reduce tissue damage [73,75]. Such a biphasic effect indicates the plasticity of immune regulation in MSCs, and we speculate whether MSCs can trigger the necessary inflammatory response while suppressing excessive immune responses and maintaining a local microenvironment conducive to tissue repair.
6. Interactions between MSCs and mast cells (MC)
The functions of MC in bone repair mainly modulate angiogenesis as well as anabolic and catabolic processes during fracture repair and remodeling [76]. It is speculated that MSCs can induce an appropriate inflammatory response while suppressing excessive immune reactions. This implies that during the healing process, MCs may also impact the behavior of MSCs by interacting with specific receptors such as IL-1 receptor (IL-1R), IL-6R, TNF receptor (TNFR), CXCR1, TGF-β receptor 1 (TGFβRI) or basic FGF receptor (bFGFR) [77]. Conversely, MSCs release factors like TGF-β, VEGF or IL-6 which can influence MC function through their respective receptors (TGFβR1/2, VEGF receptor, IL-6R) [78,79]. Meanwhile, mast cell tryptase is an additional angiogenesis factor. Mast cells act at sites of new vessel formation by secreting tryptase, which then functions as a potent angiogenesis factor [42]. Additionally, it is possible that MSCs have a direct impact on MC behavior during the healing of fractures by regulating factors such as MC numbers, cytokine production, mediator release and degranulation. As a result, the influence of MCs on MSCs in bone healing may be attributed to their ability to affect the migration, proliferation and differentiation of MSCs. In summary, while recent studies have identified specific roles for MCs in fracture healing, further investigation is needed to fully understand how these cells interact with MSCs in this context [80].
7. MSCs inhibit B lymphocytes
μMT gene knockout of B lymphocytes may lead to changes in the immune environment of the new bone induction site, thereby stimulating the initial accumulation and proliferation of mesenchymal progenitor cells [81]. As an important part of the acquired immune response system, B cells can differentiate into plasma cells under the stimulation of antigen, which can synthesize and secrete antibodies (immunoglobulin), and mainly perform humoral immunity of the body. Early studies have shown that mouse allogeneic BM-MSCs have an inhibitory effect on the proliferation, activation, and IgG secretion of B cells [82]. The presence of allogeneic BM-MSCs can inhibit the proliferation of by arresting B-lymphocytes in the G (0)/G (1) phase of the cell cycle. Meanwhile, MSCS can also inhibit the proliferation of B lymphocytes by producing soluble factors by transwell experiments [83,84]. Allogeneic BM-MSCs can also change the activation mode of extracellular response kinase 1/2 and p38 mitogen-activated protein kinase pathway, both of which are involved in the survival, activation, and proliferation of B cells [84]. Data from another study suggest that humoral factors released by BM-MCS inhibit terminal differentiation of B cells, possibly by inhibiting the expression of mature protein-1 in B lymphocytes [85]. The inhibition of B cell activation by allogeneic MSC seems to depend on IFN-γ and cell contact through programmed cell death-1/ programmed cell death-Ligand 1 (PD-1/PD-L1) interaction, like the immunosuppression of T lymphocytes by MSC [86]. However, this is not consistent with data showing that MSC promotes B cell proliferation and differentiation in vitro [87].
8. MSCs inhibit T Lymphocytes
In the anabolic and catabolic stages of fracture healing, both innate and adaptive immune processes are essential. In addition to macrophages and neutrophils clearing the damaged site, specific cell-mediated immune function also clears necrotic tissue during the initial inflammatory stage after injury, promotes angiogenesis, and initiates repair [88].
Tregs are known to have a positive effect on fracture healing, because they inhibit the secretion of pro-inflammatory cytokines by activated T cells, which is conducive to the differentiation of MSCs into osteoblasts [89]. Through direct cell-cell contact [90] or paracrine [91], MSCs play a dual role in regulating these different T-cell subpopulations during bone healing. On the one hand, BM- MSCs can inhibit the activation and proliferation of T cells (CD4+ and CD8+ subpopulations) by inducing cell cycle G0 arrest, as shown in the in vitro mixed lymphocyte reaction [92,93,94]. In addition, MSCs stimulate T cell apoptosis through Fas/FasL-dependent pathways [95] and PD-L1 secretion [96]. On the other hand, MSC can not only induce the formation of CD4+CD25+Foxp3+Tregs [97,98] but also can secrete heme oxygenase-1 (HO-1) and IL-10 and up-regulate PD-1 receptors on Tregs triggers the immunosuppressive ability of Tregs [99].
Interestingly, bone fractures cause the immune system to be suppressed [100] and a local increase in the number of induced T regulatory (iTREG) cells that inhibit the active adaptive immune response in the fracture callus [12]. Research has indicated that MSCs are capable of maintaining a state of low immunogenicity [101,102] by producing paracrine factors with immunosuppressive properties [103,104,105] through their direct impact on immune cell populations, such as T cells [106,107]. Such effects indicate that these cells transmit immune tolerance during the early stages of endochondral bone formation and protect developing tissues by inhibiting the allogeneic proliferation of T cells during stem cell recruitment and cartilage formation [12].
9. Effects of B lymphocytes and T lymphocytes on MSCs
B cell-mediated regulation functions in the early stages of the bone healing process, and its absence is associated with delayed healing or non-healing in fractures, and upregulation of Breg function may contribute to improved fracture healing [108]. B cells may also inhibit bone formation by suppressing osteoblast differentiation, for example, in patients with rheumatoid arthritis, B cells have been reported to inhibit osteogenesis via TNF-α and C-C motif chemokine ligand (CCL) 3 [109]. Similarly, the maturation of osteoblasts is hindered by B cells in the presence of colony stimulating factor 3 (G-CSF) during the process of hematopoietic stem cell and progenitor cell homing [80]. MSC-mediated bone regeneration can be partially impeded by CD8+ T cells, while osteogenic differentiation is completely inhibited by CD4+ T cells, however, Treg infusion can eliminate the inhibition of MSCs by activated T cells [34,80]. Pro-inflammatory T cells inhibit bone formation by secreting cytokines that inhibit the RUNX-2 pathway and promote apoptosis of bone marrow-mesenchymal stem cells [34]. Meanwhile, activated T cells induced MSC cell apoptosis via the Fas/FasL and CD40/CD40L pathways [67]. However, contrary to the view that T cells inhibit bone healing, some studies have shown that certain types of T cells are beneficial to osteogenesis. For example, Treg cells can resist this negative effect because Treg cells are able to shed large amounts of TNF receptor superfamily member 1B (TNFRII) and thus inhibit the action of TNF [110] and promoting bone formation. IL-17A secreted by γδT cells shifts the lipid differentiation capacity of MSCs to osteogenic differentiation, thereby improving the quality of bone healing [111]. In addition, TNF-α-stimulated T cells also activate the NF-κB pathway and secrete more CCL5 to recruit MSCs to the site of injury, thus achieving ectopic osteogenesis [112].
10. Effects of pH
Bone injury in the form of fracture or osteotomy can lead to vascular disruption. Vascular disruption following bone injury can lead to an acidic, hypoxic wound environment. This disruption of blood flow results in a hypoxic zone accompanied by local tissue ischemia, decreased pH, and elevated lactate levels within the fracture-healed tissue [113]. Previous research has indicated that the pH level decreases from a normal physiological value of 7.4 to 6.8 during the initial two days of healing, which is known as the inflammatory phase [114]. During this phase, mesenchymal stem cells (MSCs) migrate to the injury site to assist in tissue repair. In view of this, Hazehara-Kunitomo, Y., et al. conducted a study on how brief exposure to acidic conditions (pH 6.8) affects the characteristics of BM-MSCs. Interestingly, their findings revealed that pre-treating BM-MSCs with an acidic pH enhances the expression of specific markers associated with stem cell properties (organic cation/carnitine transporter4 (OCT-4), Nanog homeobox (NANOG)), as well as promoting cell viability and proliferation. However, it was observed that acidic conditions have a negative impact on BM-MSC migration ability. These results suggest that maintaining an acidic pH during the early stages of bone healing is crucial for enhancing BM-MSCs stemness and function [114]. It has been shown that hypoxic cultured hBM-MSCs displayed higher levels of pro-survival, pro-proliferation, pro-migration, vasculogenesis and angiogenesis genes [115]. However, hypoxia inhibits osteogenic differentiation of MSCs [116]. Therefore, acidic, hypoxic conditions may improve the intrinsic properties of these cells, and these findings may further aid in the development of materials and methods for more precise and in situ control of BM-MSC stemness and function, essential for the development of successful cellular therapies.
11. Effects of exosomes on fractures
Exosomes are metastatic "paracrine factors" that act as signaling molecules in the tissue microenvironment [117]. MSC-derived exosomes are extracellular nano-vesicular products derived from MSCs [118]. Exosomes derived from MSC improve wound healing by signaling or by influencing the fate decisions of certain immune cells, thereby promoting the restoration of immune homeostasis or through the attenuation of excessive inflammation [119]. Recently, it has been shown that exosome-rich mineral-doped poly-L-propyleneglycolate acidic scaffolds can improve osteogenesis of human adipose-derived MSCs [120]. There has also been research suggesting that exosomes derived from BM-MSC alleviate bone loss by restoring the function of recipient BM-MSCs and activating Wnt/β-catenin signaling [121]. Hypoxia preconditioning promotes fracture healing by activating hypoxia inducible factor 1, alpha subunit (HIF-1α) to mediate the increase of exosome microRNA 126(mir-126) [122].
12. The effect of immune cells on angiogenesis
Blood vessels in bone tissue play crucial roles in the repair of injuries. The involvement of vascular endothelial cells (ECs) in fracture repairs is facilitated through their interaction with bone cells[123]. In bone tissues, two subtypes of vascular ECs have been identified: type H vascular ECs characterized by high expression of CD31 and EMCN, and type L vascular ECs characterized by low expression of CD31 and EMCN[124]. Although type H ECs constitute only 1.77% of all bone ECs and 0.015% of total bone marrow ECs, they are surrounded by a significant number of bone progenitor cells capable of differentiating into osteoblasts and osteocytes. On the other hand, type L vessels have minimal surrounding bone progenitor cells, suggesting that type H vessels may strongly promote the regeneration process[124,125]. Extensive research has demonstrated that inflammatory responses and neovascularization are critical factors initiating bone regeneration [126,127,128]. The stimulator of interferon genes (STING), an essential factor involved in innate immunity, has been found to be closely associated with angiogenesis. A study indicated that activation of STING leads to a decrease in H-type blood vessels and almost no callus mineralization; however, inhibiting mmSTING can enhance the formation of H-type blood vessels. This suggests that inhibiting STING can accelerate the healing process while promoting the formation of H-type blood vessels simultaneously[129]. Gao et al.'s findings revealed that macrophages/monocytes can differentiate into tar-trate-resistant acid phosphatase (TRAP) mononuclear cells on periosteal surfaces dur-ing bone modeling. These TRAP mononuclear cells secrete platelet-derived growth factor type BB (PDGF-BB), which induces periostin expression from periosteum-derived cells (PDC). Consequently, PDC recruitment occurs on periosteal surfaces to support both type H vessel formation and osteogenesis processes[130].
The increase in regulatory T helper cells and the expression of anti-inflammatory cytokine IL-10 were observed alongside an increase in angiogenic factors (HIF1a and HIF1a regulated genes) specifically within the regenerative bone hematoma, rather than the soft tissue hematoma. These findings suggest that timely resolution of inflammation and early initiation of revascularization are mutually dependent and crucial for promoting a regenerative healing process[128].
13. Application of single cell analysis in fracture repair
While the roles of immune cells and stromal cells in fracture healing have been suggested, there is still limited understanding regarding the variations in their numbers or types during different stages of fracture healing. Zhang et al. utilized single-cell RNA-seq (scRNA-seq) to examine immune cells in a mouse fracture model and discovered significant differences in B cell populations, with fewer B cells observed during tissue formation stage and higher numbers during tissue healing stage. These findings indicate the crucial involvement of B cells in promoting fracture healing. Furthermore, exosomes released by B cells were found to hinder osteoblast differentiation and enhance osteoclast formation, thereby reducing osteogenic activity[131]. Avin et al., through scRNA-seq analysis, compared the proportions of immune cell subtypes and identified higher levels of monocytes and CD14 DCs, as well as lower levels of T cells, myelocytes, and promyelocytes within nonunion bone group. This study also provided valuable insights into gene expression changes from an osteoimmunological perspective. Such discoveries contribute significantly towards comprehending the mechanisms underlying bone nonunion[132]. In order to gain insights into the regulatory factors governing the differentiation of bone mesenchymal cells and their lineage specialization, Sivaraj et al. employed scRNA-seq to investigate the functional characteristics, lineage differentiation potential, and cellular fate transitions of bone mesenchymal stromal cells. Their findings revealed that diaphysis mesenchymal stromal cells, which exhibited limited differentiation capacity, were associated with the sinusoidal vasculature within the bone microenvironment. On the other hand, metaphysis mesenchymal stromal cells displayed multilineage potential for osteogenic, adipogenic, and chondrogenic differentiation. Moreover, PDGFRb signaling and Jun-B transcription factor were identified as key regulators controlling the fate of bone mesenchymal stromal cells during bone formation processes by influencing cell proliferation and differentiation[133]. To enhance fracture healing treatments effectively, future investigations should employ a combination of scRNA-seq with complementary techniques to explore crucial mechanisms underlying immune cell dynamics and variations in stromal cell behavior throughout all stages of bone repair. Additionally, exploring interactions between stromal cells and immune cells during fracture healing may offer valuable insights for identifying novel therapeutic targets.
14. Main limitations and prospects
The potential of MSCs in promoting bone repair has been promising, but there are still several factors that may limit their use and increase variability between studies. These include the source of MSCs, as well as the timing and quantity of implantation. Additionally, it is important to fully characterize the population of MSCs and determine the best approach based on the type and location of bone trauma or defect [134]. In cases where in vitro expansion is necessary for cell transplantation or gene delivery purposes, limitations arise due to lack of standardization and automation in this process. Variability can occur depending on factors such as number of passages, media type, serum usage (including lot-to-lot variations), animal-free media availability or presence of xenogenic proteins which pose a risk for contamination. Similarly to other genetically modified cells, genetic modulation presents safety concerns particularly when viral vectors are used [135]. Intrinsic biological variability between tissue sources, donors, clonal subsets, and single-cell variability, and extrinsically introduced variability through non-standard isolation, selection, and production methods result to MSC heterogeneity. Translation of MSC products beyond clinical trials has been limited due to, among other factors, MSC heterogeneity, complicating the ability to produce reliable, uniform therapy throughout the MSC expansion and banking process [32]. Nevertheless, with the advancement of molecular biology techniques, there is an increasing possibility to utilize engineered MSCs or their secretome in clinical settings for addressing bone defects. The choice of biomaterials also affects treatment outcomes. The body's immune response to bone implants is influenced by a number of factors, including the macroporosity of the biomaterial, surface microstructure, material hardness, and particle size[136,137]. Macrophages are the main effector cells of the immune response to implants, and bone implants affect MSCs mainly in terms of cell viability, adhesion, migration, proliferation and differentiation[138]. The presence of surface roughness in titanium, such as polished, machined, and grit-blasted commercially pure titanium, has been observed to impact the attachment and spreading of immune cells. Over time, macrophage adhesion increases on all surfaces while cell spreading is enhanced with higher surface roughness levels[139]. Furthermore, the roughness of titanium can also influence the production of inflammatory cytokines and chemokines by macrophages. Significant stimulatory effects have been noted on sandblasted and acid-etched surfaces[140], whereas titania nanotube arrays modified surfaces exhibit reduced immune response compared to raw titanium surfaces[141]. It is worth noting that bone's surface roughness measures approximately 32 nm at a nanoscale level, making nanomaterials highly biomimetic[142]. In vitro studies have demonstrated that nanoscale microstructures effectively stimulate human MSCs to produce bone minerals even without osteogenic supplements[143]. Additionally, it has been discovered that macrophages exposed to microstructured topography become activated with both M1 and M2 characteristics instead of nanostructured topography alone[144]. Research also indicates that perios-teal extracellular matrix (PEM) hydrogels promote the recruitment and M2-polarization of macrophages. They further facilitate differentiation of MSCs into endothelial-like cells along with HUVEC tube formation and osteogenic differentiation of MSCs[145]. Therefore, new strategies that reduce MSC heterogeneity by pooled-MSCs, clonal MSCs, priming pretreatment of MSCs, biomaterial-MSC interactions and so on to improve their immunomodulatory potencies will clinically impact[32,146].
In the clinic, because bone marrow-derived MSCs are more difficult to collect and are themselves less abundant, studies of alternative sources have emerged[147,148,149]. Some research results show that adipose MSCs and BM-MSCs have similar immunophenotypes and in vitro differentiation abilities, as well as similar immunomodulatory abilities, and that adipose MSCs are more metabolically active, secrete more cytokines, and are more efficient in their immunomodulatory effects than BM-MSCs. Adipose-derived stem cells (ASCs) can be considered as a good alternative to BM-MSCs for immunomodulatory therapy[150]. ASCs are a significant subset of MSCs[151]. ASCs, derived from the stromal vascular fraction (SVF), are progenitor cells located around blood vessels[152]. Similar to BM-MSCs, ASCs have been extensively studied in the field of bone tissue engineering[153]. ASCs possess the ability to differentiate into multiple cell lineages, including adipocytes, chondrocytes, and osteoblasts. The fate determination of these lineages is regulated by key factors such as Runx2 and Osterix (Sp7 transcription factor)[125,154]. Various signaling pathways play a crucial role in controlling osteogenic differentiation, including bone morphogenetic protein (BMP)[155], Notch[156], Wnt[157] and Hedgehog-signaling[158]. Among these pathways, Wnt signaling pathway acts as a pivotal regulator that directs ASCs differentiation towards osteogenesis by upregulating Runx2 and Osterix expression levels[159]. Numerous studies have explored different substrates to enhance the osteogenic potential of ASCs. These substrates include vitamin D3 [160], alendronate [161], selenium [162], platelet-rich plasma [163], and the inflammatory response [164,165,166,167].
In conclusion, the mechanisms of the immunomodulatory effects of MSCs are complex, and characterizing a single factor is difficult because of the interaction between various factors (Figure 3). More in-depth studies are needed to fully understand and exploit the mechanisms of immunomodulation by MSCs.
Pro-inflammatory cytokines secreted by macrophages (TNF-α, IL-6, IL-1β, IL-23, and IFN-γ) stimulate osteogenic differentiation of MSCs, and TGF-β and VEGF have been shown to have angiogenic activity. PGE2 and IL-4 secreted by MSCs induce the transformation of macrophages from M1 type to M2 type. MSCs affects DCs differentiation and proinflammatory cytokine secre-tion through TGF-β1, IL-6 and GRO-γ. MSCs not only stimulate T cell apoptosis through Fas/FasL pathway and PD-L1 secretion, MSCs can also secrete heme oxygenase-1 (HO-1) and IL-10, up-regulate the PD-1 receptor on Tregs, and trigger the immunosuppressive ability of Tregs. Meanwhile, activated T cells induced MSCs apoptosis via the Fas/FasL and CD40/CD40L pathways. However, Tregs are able to shed large amounts of TNF receptor superfamily member 1B (TNFRII) and thus inhibit the ac-tion of TNF and promoting bone formation, IL-17A secreted by γδT cells shifts the lipid differentiation capacity of MSCs to osteogenic differentiation, there-by improving the quality of bone healing. TGF-β and IL-6 produced by activated MSCs limit NK cell effector function but promote NK cell differentiation. The inhibition of B cell activation by MSC depends on IFN-γ and PD-1/PD-L1.
15. Conclusion
Convincing evidence supports the important role of BM- MSCs in the bone healing process, which is mainly due to their immunomodulatory ability by cell-to-cell contact and secreting paracrine factors. Clinical trials have verified their relative safety and effectiveness, as well as their potential use with cell therapy for bone diseases with potentially inflammatory conditions. However, despite the great progress made since the discovery and characterization of MSCs, it is necessary to evaluate the osteogenic potential of MSCs by selecting donors and/or subpopulations with high osteogenic ability to promote bone recovery and inflammation regulation. Further research will clarify the mechanism of bone marrow MSC immune regulation to promote bone repair.
Author Contributions
Reference collection, Yumei Yang; writing—original draft preparation, Wenjing Xu; writing—review and editing, Na Li; project administration, Jinlian Hua; funding acquisition, Jinlian Hua. All authors have read and agreed to the published version of the manuscript.
Funding
The current study was supported by the National Natural Science Foundation of China (32072806, 32002246), Program of Shaanxi Province Science and Technology Innovation Team (2019TD-036), Program of Shaanxi Province Science and Technology (2022NY-044).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gómez-Barrena, E.; Rosset, P.; Gebhard, F.; Hernigou, P.; Baldini, N.; Rouard, H.; Sensebé, L.; Gonzalo-Daganzo, R.M.; Giordano, R.; Padilla-Eguiluz, N.; et al. Feasibility and safety of treating non-unions in tibia, femur and humerus with autologous, expanded, bone marrow-derived mesenchymal stromal cells associated with biphasic calcium phosphate biomaterials in a multicentric, non-comparative trial. Biomaterials 2019, 196, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chu, W.; Zhuang, Y.; Shi, D.; Tao, H.; Jin, C.; Dai, K.; Zhao, J.; Gan, Y. Bone Mesenchymal Stem Cell-Enriched β-Tricalcium Phosphate Scaffold Processed by the Screen-Enrich-Combine Circulating System Promotes Regeneration of Diaphyseal Bone Non-Union. Cell Transplant 2019, 28, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kandoi, S.; Misra, R.; Vijayalakshmi, S.; Rajagopal, K.; Verma, R.S. The mesenchymal stem cell secretome: A new paradigm towards cell-free therapeutic mode in regenerative medicine. Cytokine Growth Factor Rev 2019, 46, 1–9. [Google Scholar] [CrossRef]
- McGonagle, D.; Baboolal, T.G.; Jones, E. Native joint-resident mesenchymal stem cells for cartilage repair in osteoarthritis. Nat Rev Rheumatol 2017, 13, 719–730. [Google Scholar] [CrossRef]
- Bunpetch, V.; Zhang, Z.Y.; Zhang, X.; Han, S.; Zongyou, P.; Wu, H.; Hong-Wei, O. Strategies for MSC expansion and MSC-based microtissue for bone regeneration. Biomaterials 2019, 196, 67–79. [Google Scholar] [CrossRef]
- Han, L.; Wang, B.; Wang, R.; Gong, S.; Chen, G.; Xu, W. The shift in the balance between osteoblastogenesis and adipogenesis of mesenchymal stem cells mediated by glucocorticoid receptor. Stem Cell Res Ther 2019, 10, 377. [Google Scholar] [CrossRef]
- Huang, S.; Xu, L.; Zhang, Y.; Sun, Y.; Li, G. Systemic and Local Administration of Allogeneic Bone Marrow-Derived Mesenchymal Stem Cells Promotes Fracture Healing in Rats. Cell Transplant 2015, 24, 2643–2655. [Google Scholar] [CrossRef]
- Zimmermann, G.; Wagner, C.; Schmeckenbecher, K.; Wentzensen, A.; Moghaddam, A. Treatment of tibial shaft non-unions: bone morphogenetic proteins versus autologous bone graft. Injury 2009, 40 Suppl 3, S50–53. [Google Scholar] [CrossRef]
- Calori, G.M.; Colombo, M.; Ripamonti, C.; Bucci, M.; Fadigati, P.; Mazza, E.; Mulas, S.; Tagliabue, L. Polytherapy in bone regeneration: clinical applications and preliminary considerations. Int J Immunopathol Pharmacol 2011, 24, 85–90. [Google Scholar] [CrossRef]
- Chu, W.; Wang, X.; Gan, Y.; Zhuang, Y.; Shi, D.; Liu, F.; Sun, Y.; Zhao, J.; Tang, T.; Dai, K. Screen-enrich-combine circulating system to prepare MSC/β-TCP for bone repair in fractures with depressed tibial plateau. Regen Med 2019, 14, 555–569. [Google Scholar] [CrossRef]
- Baht, G.S.; Vi, L.; Alman, B.A. The Role of the Immune Cells in Fracture Healing. Curr Osteoporos Rep 2018, 16, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Al-Sebaei, M.O.; Daukss, D.M.; Belkina, A.C.; Kakar, S.; Wigner, N.A.; Cusher, D.; Graves, D.; Einhorn, T.; Morgan, E.; Gerstenfeld, L.C. Role of Fas and Treg cells in fracture healing as characterized in the fas-deficient (lpr) mouse model of lupus. J Bone Miner Res 2014, 29, 1478–1491. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ricciardi, B.F.; Hernandez-Soria, A.; Shi, Y.; Pleshko Camacho, N.; Bostrom, M.P. Callus mineralization and maturation are delayed during fracture healing in interleukin-6 knockout mice. Bone 2007, 41, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.; Cooney, T.E.; Englund, R.; Lubahn, J.D. Effects of interleukin-6 ablation on fracture healing in mice. J Orthop Res 2011, 29, 1437–1442. [Google Scholar] [CrossRef]
- Nam, D.; Mau, E.; Wang, Y.; Wright, D.; Silkstone, D.; Whetstone, H.; Whyne, C.; Alman, B. T-lymphocytes enable osteoblast maturation via IL-17F during the early phase of fracture repair. PLoS One 2012, 7, e40044. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cho, T.J.; Kon, T.; Aizawa, T.; Tsay, A.; Fitch, J.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Impaired fracture healing in the absence of TNF-alpha signaling: the role of TNF-alpha in endochondral cartilage resorption. J Bone Miner Res 2003, 18, 1584–1592. [Google Scholar] [CrossRef]
- Glass, G.E.; Chan, J.K.; Freidin, A.; Feldmann, M.; Horwood, N.J.; Nanchahal, J. TNF-alpha promotes fracture repair by augmenting the recruitment and differentiation of muscle-derived stromal cells. Proc Natl Acad Sci U S A 2011, 108, 1585–1590. [Google Scholar] [CrossRef]
- Könnecke, I.; Serra, A.; El Khassawna, T.; Schlundt, C.; Schell, H.; Hauser, A.; Ellinghaus, A.; Volk, H.D.; Radbruch, A.; Duda, G.N.; et al. T and B cells participate in bone repair by infiltrating the fracture callus in a two-wave fashion. Bone 2014, 64, 155–165. [Google Scholar] [CrossRef]
- Mountziaris, P.M.; Mikos, A.G. Modulation of the inflammatory response for enhanced bone tissue regeneration. Tissue Eng Part B Rev 2008, 14, 179–186. [Google Scholar] [CrossRef]
- Zhang, M.F.; Wan, S.C.; Chen, W.B.; Yang, D.H.; Liu, W.Q.; Li, B.L.; Aierken, A.; Du, X.M.; Li, Y.X.; Wu, W.P.; et al. Transcription factor Dmrt1 triggers the SPRY1-NF-κB pathway to maintain testicular immune homeostasis and male fertility. Zool Res 2023, 44, 505–521. [Google Scholar] [CrossRef]
- Wei, Y.D.; Du, X.M.; Yang, D.H.; Ma, F.L.; Yu, X.W.; Zhang, M.F.; Li, N.; Peng, S.; Liao, M.Z.; Li, G.P.; et al. Dmrt1 regulates the immune response by repressing the TLR4 signaling pathway in goat male germline stem cells. Zool Res 2021, 42, 14–27. [Google Scholar] [CrossRef] [PubMed]
- O'Keefe, R.J.; Mao, J. Bone tissue engineering and regeneration: from discovery to the clinic--an overview. Tissue Eng Part B Rev 2011, 17, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Bressan, E.; Ferroni, L.; Gardin, C.; Bellin, G.; Sbricoli, L.; Sivolella, S.; Brunello, G.; Schwartz-Arad, D.; Mijiritsky, E.; Penarrocha, M.; et al. Metal Nanoparticles Released from Dental Implant Surfaces: Potential Contribution to Chronic Inflammation and Peri-Implant Bone Loss. Materials (Basel) 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. The biology of fracture healing. An overview for clinicians. Part II. Clin Orthop Relat Res 1989, 294–309. [Google Scholar]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat Rev Nephrol 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The early fracture hematoma and its potential role in fracture healing. Tissue Eng Part B Rev 2010, 16, 427–434. [Google Scholar] [CrossRef]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat Rev Rheumatol 2012, 8, 133–143. [Google Scholar] [CrossRef]
- Lam, J.; Takeshita, S.; Barker, J.E.; Kanagawa, O.; Ross, F.P.; Teitelbaum, S.L. TNF-alpha induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANK ligand. J Clin Invest 2000, 106, 1481–1488. [Google Scholar] [CrossRef]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front Immunol 2014, 5, 511. [Google Scholar] [CrossRef]
- Li, N.; Hua, J. Interactions between mesenchymal stem cells and the immune system. Cell Mol Life Sci 2017, 74, 2345–2360. [Google Scholar] [CrossRef]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol Sci 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Dunn, C.M.; Kameishi, S.; Grainger, D.W.; Okano, T. Strategies to address mesenchymal stem/stromal cell heterogeneity in immunomodulatory profiles to improve cell-based therapies. Acta Biomater 2021, 133, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Dighe, A.S.; Yang, S.; Madhu, V.; Balian, G.; Cui, Q. Interferon gamma and T cells inhibit osteogenesis induced by allogeneic mesenchymal stromal cells. J Orthop Res 2013, 31, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Kikuiri, T.; Akiyama, K.; Chen, C.; Xu, X.; Yang, R.; Chen, W.; Wang, S.; Shi, S. Mesenchymal stem cell-based tissue regeneration is governed by recipient T lymphocytes via IFN-γ and TNF-α. Nat Med 2011, 17, 1594–1601. [Google Scholar] [CrossRef]
- Reinke, S.; Geissler, S.; Taylor, W.R.; Schmidt-Bleek, K.; Juelke, K.; Schwachmeyer, V.; Dahne, M.; Hartwig, T.; Akyüz, L.; Meisel, C.; et al. Terminally differentiated CD8⁺ T cells negatively affect bone regeneration in humans. Sci Transl Med 2013, 5, 177ra136. [Google Scholar] [CrossRef]
- Xue, D.; Chen, E.; Zhong, H.; Zhang, W.; Wang, S.; Joomun, M.U.; Yao, T.; Tan, Y.; Lin, S.; Zheng, Q.; et al. Immunomodulatory properties of graphene oxide for osteogenesis and angiogenesis. Int J Nanomedicine 2018, 13, 5799–5810. [Google Scholar] [CrossRef]
- Hirata, E.; Miyako, E.; Hanagata, N.; Ushijima, N.; Sakaguchi, N.; Russier, J.; Yudasaka, M.; Iijima, S.; Bianco, A.; Yokoyama, A. Carbon nanohorns allow acceleration of osteoblast differentiation via macrophage activation. Nanoscale 2016, 8, 14514–14522. [Google Scholar] [CrossRef]
- Shi, M.; Chen, Z.; Farnaghi, S.; Friis, T.; Mao, X.; Xiao, Y.; Wu, C. Copper-doped mesoporous silica nanospheres, a promising immunomodulatory agent for inducing osteogenesis. Acta Biomater 2016, 30, 334–344. [Google Scholar] [CrossRef]
- Lu, L.Y.; Loi, F.; Nathan, K.; Lin, T.H.; Pajarinen, J.; Gibon, E.; Nabeshima, A.; Cordova, L.; Jämsen, E.; Yao, Z.; et al. Pro-inflammatory M1 macrophages promote Osteogenesis by mesenchymal stem cells via the COX-2-prostaglandin E2 pathway. J Orthop Res 2017, 35, 2378–2385. [Google Scholar] [CrossRef]
- Tu, B.; Liu, S.; Liu, G.; Yan, W.; Wang, Y.; Li, Z.; Fan, C. Macrophages derived from THP-1 promote the osteogenic differentiation of mesenchymal stem cells through the IL-23/IL-23R/β-catenin pathway. Exp Cell Res 2015, 339, 81–89. [Google Scholar] [CrossRef]
- Gong, L.; Zhao, Y.; Zhang, Y.; Ruan, Z. The Macrophage Polarization Regulates MSC Osteoblast Differentiation in vitro. Ann Clin Lab Sci 2016, 46, 65–71. [Google Scholar] [PubMed]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J Investig Dermatol Symp Proc 2000, 5, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tasso, R.; Ulivi, V.; Reverberi, D.; Lo Sicco, C.; Descalzi, F.; Cancedda, R. In vivo implanted bone marrow-derived mesenchymal stem cells trigger a cascade of cellular events leading to the formation of an ectopic bone regenerative niche. Stem Cells Dev 2013, 22, 3178–3191. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, Z.L.; Xiao, M.; Yang, Z.Z.; Peng, M.Z.; Li, C.D.; Zhou, X.J.; Wang, J.W. Impact of bone marrow mesenchymal stem cell immunomodulation on the osteogenic effects of laponite. Stem Cell Res Ther 2018, 9, 100. [Google Scholar] [CrossRef]
- Seebach, E.; Freischmidt, H.; Holschbach, J.; Fellenberg, J.; Richter, W. Mesenchymal stroma cells trigger early attraction of M1 macrophages and endothelial cells into fibrin hydrogels, stimulating long bone healing without long-term engraftment. Acta Biomater 2014, 10, 4730–4741. [Google Scholar] [CrossRef]
- Tour, G.; Wendel, M.; Tcacencu, I. Bone marrow stromal cells enhance the osteogenic properties of hydroxyapatite scaffolds by modulating the foreign body reaction. J Tissue Eng Regen Med 2014, 8, 841–849. [Google Scholar] [CrossRef]
- Lin, T.; Pajarinen, J.; Nabeshima, A.; Lu, L.; Nathan, K.; Jämsen, E.; Yao, Z.; Goodman, S.B. Preconditioning of murine mesenchymal stem cells synergistically enhanced immunomodulation and osteogenesis. Stem Cell Res Ther 2017, 8, 277. [Google Scholar] [CrossRef]
- Wei, F.; Li, Z.; Crawford, R.; Xiao, Y.; Zhou, Y. Immunoregulatory role of exosomes derived from differentiating mesenchymal stromal cells on inflammation and osteogenesis. J Tissue Eng Regen Med 2019, 13, 1978–1991. [Google Scholar] [CrossRef]
- Li, Y.; Yang, C.; Zhao, H.; Qu, S.; Li, X.; Li, Y. New Developments of Ti-Based Alloys for Biomedical Applications. Materials (Basel) 2014, 7, 1709–1800. [Google Scholar] [CrossRef]
- Shin, R.L.; Lee, C.W.; Shen, O.Y.; Xu, H.; Lee, O.K. The Crosstalk between Mesenchymal Stem Cells and Macrophages in Bone Regeneration: A Systematic Review. Stem Cells Int 2021, 2021, 8835156. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Moretta, L. Cellular and molecular interactions of mesenchymal stem cells in innate immunity. Immunol Cell Biol 2013, 91, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Moretta, L. Interactions between mesenchymal stem cells and dendritic cells. Adv Biochem Eng Biotechnol 2013, 130, 199–208. [Google Scholar] [CrossRef]
- Wehner, R.; Taubert, C.; Mende, T.; Gaebler, C.; de Andrade, A.V.; Bornhäuser, M.; Werner, C.; Tonn, T.; Schäkel, K.; Bachmann, M.; et al. Engineered extracellular matrix components do not alter the immunomodulatory properties of mesenchymal stromal cells in vitro. J Tissue Eng Regen Med 2013, 7, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Liu, J.J.; Wu, J.; Zhang, L.L.; Liu, F.; Yin, L.; Zhang, M.M.; Yu, B. Novel mechanism of inhibition of dendritic cells maturation by mesenchymal stem cells via interleukin-10 and the JAK1/STAT3 signaling pathway. PLoS One 2013, 8, e55487. [Google Scholar] [CrossRef] [PubMed]
- Aldinucci, A.; Rizzetto, L.; Pieri, L.; Nosi, D.; Romagnoli, P.; Biagioli, T.; Mazzanti, B.; Saccardi, R.; Beltrame, L.; Massacesi, L.; et al. Inhibition of immune synapse by altered dendritic cell actin distribution: a new pathway of mesenchymal stem cell immune regulation. J Immunol 2010, 185, 5102–5110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.G.; Xu, W.; Sun, L.; Li, W.M.; Li, Q.B.; Zou, P. The characteristics and immunoregulatory functions of regulatory dendritic cells induced by mesenchymal stem cells derived from bone marrow of patient with chronic myeloid leukaemia. Eur J Cancer 2012, 48, 1884–1895. [Google Scholar] [CrossRef]
- Choi, Y.S.; Jeong, J.A.; Lim, D.S. Mesenchymal stem cell-mediated immature dendritic cells induce regulatory T cell-based immunosuppressive effect. Immunol Invest 2012, 41, 214–229. [Google Scholar] [CrossRef]
- Chen, H.W.; Chen, H.Y.; Wang, L.T.; Wang, F.H.; Fang, L.W.; Lai, H.Y.; Chen, H.H.; Lu, J.; Hung, M.S.; Cheng, Y.; et al. Mesenchymal stem cells tune the development of monocyte-derived dendritic cells toward a myeloid-derived suppressive phenotype through growth-regulated oncogene chemokines. J Immunol 2013, 190, 5065–5077. [Google Scholar] [CrossRef]
- Deng, Y.; Yi, S.; Wang, G.; Cheng, J.; Zhang, Y.; Chen, W.; Tai, Y.; Chen, S.; Chen, G.; Liu, W.; et al. Umbilical cord-derived mesenchymal stem cells instruct dendritic cells to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1. Stem Cells Dev 2014, 23, 2080–2092. [Google Scholar] [CrossRef]
- Tang, W.Y.; Liu, J.H.; Peng, C.J.; Liao, Y.; Luo, J.S.; Sun, X.; Tang, Y.L.; Luo, X.Q. Functional Characteristics and Application of Mesenchymal Stem Cells in Systemic Lupus Erythematosus. Arch Immunol Ther Exp (Warsz) 2021, 69, 7. [Google Scholar] [CrossRef]
- Petri, R.M.; Hackel, A.; Hahnel, K.; Dumitru, C.A.; Bruderek, K.; Flohe, S.B.; Paschen, A.; Lang, S.; Brandau, S. Activated Tissue-Resident Mesenchymal Stromal Cells Regulate Natural Killer Cell Immune and Tissue-Regenerative Function. Stem Cell Reports 2017, 9, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Rekasi, H.; Hepner-Schefczyk, M.; Fessmann, K.; Petri, R.M.; Bruderek, K.; Brandau, S.; Jäger, M.; Flohé, S.B. Human mesenchymal stromal/stem cells acquire immunostimulatory capacity upon cross-talk with natural killer cells and might improve the NK cell function of immunocompromised patients. Stem Cell Res Ther 2016, 7, 88. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J Allergy Clin Immunol 2010, 125, S3–23. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Becchetti, S.; Mingari, M.C.; Moretta, L. Mesenchymal stem cell-natural killer cell interactions: evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation. Blood 2006, 107, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Crop, M.J.; Korevaar, S.S.; de Kuiper, R.; JN, I.J.; van Besouw, N.M.; Baan, C.C.; Weimar, W.; Hoogduijn, M.J. Human mesenchymal stem cells are susceptible to lysis by CD8(+) T cells and NK cells. Cell Transplant 2011, 20, 1547–1559. [Google Scholar] [CrossRef]
- Giuliani, M.; Bennaceur-Griscelli, A.; Nanbakhsh, A.; Oudrhiri, N.; Chouaib, S.; Azzarone, B.; Durrbach, A.; Lataillade, J.J. TLR ligands stimulation protects MSC from NK killing. Stem Cells 2014, 32, 290–300. [Google Scholar] [CrossRef]
- Chen, R.; Hao, Z.; Wang, Y.; Zhu, H.; Hu, Y.; Chen, T.; Zhang, P.; Li, J. Mesenchymal Stem Cell-Immune Cell Interaction and Related Modulations for Bone Tissue Engineering. Stem Cells Int 2022, 2022, 7153584. [Google Scholar] [CrossRef]
- Raffaghello, L.; Bianchi, G.; Bertolotto, M.; Montecucco, F.; Busca, A.; Dallegri, F.; Ottonello, L.; Pistoia, V. Human mesenchymal stem cells inhibit neutrophil apoptosis: a model for neutrophil preservation in the bone marrow niche. Stem Cells 2008, 26, 151–162. [Google Scholar] [CrossRef]
- Brandau, S.; Jakob, M.; Hemeda, H.; Bruderek, K.; Janeschik, S.; Bootz, F.; Lang, S. Tissue-resident mesenchymal stem cells attract peripheral blood neutrophils and enhance their inflammatory activity in response to microbial challenge. J Leukoc Biol 2010, 88, 1005–1015. [Google Scholar] [CrossRef]
- Romieu-Mourez, R.; François, M.; Boivin, M.N.; Bouchentouf, M.; Spaner, D.E.; Galipeau, J. Cytokine modulation of TLR expression and activation in mesenchymal stromal cells leads to a proinflammatory phenotype. J Immunol 2009, 182, 7963–7973. [Google Scholar] [CrossRef]
- Cassatella, M.A.; Mosna, F.; Micheletti, A.; Lisi, V.; Tamassia, N.; Cont, C.; Calzetti, F.; Pelletier, M.; Pizzolo, G.; Krampera, M. Toll-like receptor-3-activated human mesenchymal stromal cells significantly prolong the survival and function of neutrophils. Stem Cells 2011, 29, 1001–1011. [Google Scholar] [CrossRef]
- Brandau, S.; Jakob, M.; Bruderek, K.; Bootz, F.; Giebel, B.; Radtke, S.; Mauel, K.; Jäger, M.; Flohé, S.B.; Lang, S. Mesenchymal stem cells augment the anti-bacterial activity of neutrophil granulocytes. PLoS One 2014, 9, e106903. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-κB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Roddy, G.W.; Oh, J.Y.; Lee, R.H.; Bartosh, T.J.; Ylostalo, J.; Coble, K.; Rosa, R.H., Jr.; Prockop, D.J. Action at a distance: systemically administered adult stem/progenitor cells (MSCs) reduce inflammatory damage to the cornea without engraftment and primarily by secretion of TNF-α stimulated gene/protein 6. Stem Cells 2011, 29, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Muschhammer, J.; Qi, Y.; Kügler, A.; de Vries, J.C.; Saffarzadeh, M.; Sindrilaru, A.; Beken, S.V.; Wlaschek, M.; Kluth, M.A.; et al. Suppression of Neutrophil-Mediated Tissue Damage-A Novel Skill of Mesenchymal Stem Cells. Stem Cells 2016, 34, 2393–2406. [Google Scholar] [CrossRef]
- Ragipoglu, D.; Dudeck, A.; Haffner-Luntzer, M.; Voss, M.; Kroner, J.; Ignatius, A.; Fischer, V. The Role of Mast Cells in Bone Metabolism and Bone Disorders. Front Immunol 2020, 11, 163. [Google Scholar] [CrossRef]
- Docheva, D.; Popov, C.; Mutschler, W.; Schieker, M. Human mesenchymal stem cells in contact with their environment: surface characteristics and the integrin system. J Cell Mol Med 2007, 11, 21–38. [Google Scholar] [CrossRef]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-conditioning. Front Immunol 2018, 9, 2837. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol Rev 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Ehnert, S.; Relja, B.; Schmidt-Bleek, K.; Fischer, V.; Ignatius, A.; Linnemann, C.; Rinderknecht, H.; Huber-Lang, M.; Kalbitz, M.; Histing, T.; et al. Effects of immune cells on mesenchymal stem cells during fracture healing. World J Stem Cells 2021, 13, 1667–1695. [Google Scholar] [CrossRef]
- Marusic, A.; Grcevic, D.; Katavic, V.; Kovacic, N.; Lukic, I.K.; Kalajzic, I.; Lorenzo, J.A. Role of B lymphocytes in new bone formation. Lab Invest 2000, 80, 1761–1774. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Han, Q.; Liao, L.; You, S.; Deng, H.; Zhao, R.C. Effects of allogeneic bone marrow-derived mesenchymal stem cells on T and B lymphocytes from BXSB mice. DNA Cell Biol 2005, 24, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V.; et al. Human mesenchymal stem cells modulate B-cell functions. Blood 2006, 107, 367–372. [Google Scholar] [CrossRef]
- Tabera, S.; Pérez-Simón, J.A.; Díez-Campelo, M.; Sánchez-Abarca, L.I.; Blanco, B.; López, A.; Benito, A.; Ocio, E.; Sánchez-Guijo, F.M.; Cañizo, C.; et al. The effect of mesenchymal stem cells on the viability, proliferation and differentiation of B-lymphocytes. Haematologica 2008, 93, 1301–1309. [Google Scholar] [CrossRef]
- Asari, S.; Itakura, S.; Ferreri, K.; Liu, C.P.; Kuroda, Y.; Kandeel, F.; Mullen, Y. Mesenchymal stem cells suppress B-cell terminal differentiation. Exp Hematol 2009, 37, 604–615. [Google Scholar] [CrossRef]
- Schena, F.; Gambini, C.; Gregorio, A.; Mosconi, M.; Reverberi, D.; Gattorno, M.; Casazza, S.; Uccelli, A.; Moretta, L.; Martini, A.; et al. Interferon-γ-dependent inhibition of B cell activation by bone marrow-derived mesenchymal stem cells in a murine model of systemic lupus erythematosus. Arthritis Rheum 2010, 62, 2776–2786. [Google Scholar] [CrossRef] [PubMed]
- Traggiai, E.; Volpi, S.; Schena, F.; Gattorno, M.; Ferlito, F.; Moretta, L.; Martini, A. Bone marrow-derived mesenchymal stem cells induce both polyclonal expansion and differentiation of B cells isolated from healthy donors and systemic lupus erythematosus patients. Stem Cells 2008, 26, 562–569. [Google Scholar] [CrossRef]
- Kon, T.; Cho, T.J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of osteoprotegerin, receptor activator of NF-kappaB ligand (osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J Bone Miner Res 2001, 16, 1004–1014. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, R.; Shi, S. Systemic infusion of mesenchymal stem cells improves cell-based bone regeneration via upregulation of regulatory T cells. Tissue Eng Part A 2015, 21, 498–509. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef]
- Najar, M.; Fayyad-Kazan, H.; Faour, W.H.; Merimi, M.; Sokal, E.M.; Lombard, C.A.; Fahmi, H. Immunological modulation following bone marrow-derived mesenchymal stromal cells and Th17 lymphocyte co-cultures. Inflamm Res 2019, 68, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Klyushnenkova, E.; Mosca, J.D.; Zernetkina, V.; Majumdar, M.K.; Beggs, K.J.; Simonetti, D.W.; Deans, R.J.; McIntosh, K.R. T cell responses to allogeneic human mesenchymal stem cells: immunogenicity, tolerance, and suppression. J Biomed Sci 2005, 12, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.M.; Ritter, T.; Ceredig, R.; Griffin, M.D. Mesenchymal stem cell effects on T-cell effector pathways. Stem Cell Res Ther 2011, 2, 34. [Google Scholar] [CrossRef]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-stem-cell-induced immunoregulation involves FAS-ligand-/FAS-mediated T cell apoptosis. Cell Stem Cell 2012, 10, 544–555. [Google Scholar] [CrossRef]
- Davies, L.C.; Heldring, N.; Kadri, N.; Le Blanc, K. Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression. Stem Cells 2017, 35, 766–776. [Google Scholar] [CrossRef]
- English, K.; Wood, K.J. Mesenchymal stromal cells in transplantation rejection and tolerance. Cold Spring Harb Perspect Med 2013, 3, a015560. [Google Scholar] [CrossRef]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef]
- Yan, Z.; Zhuansun, Y.; Chen, R.; Li, J.; Ran, P. Immunomodulation of mesenchymal stromal cells on regulatory T cells and its possible mechanism. Exp Cell Res 2014, 324, 65–74. [Google Scholar] [CrossRef]
- Meert, K.L.; Ofenstein, J.P.; Sarnaik, A.P. Altered T cell cytokine production following mechanical trauma. Ann Clin Lab Sci 1998, 28, 283–288. [Google Scholar]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef]
- Noël, D.; Djouad, F.; Bouffi, C.; Mrugala, D.; Jorgensen, C. Multipotent mesenchymal stromal cells and immune tolerance. Leuk Lymphoma 2007, 48, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Campioni, D.; Stignani, M.; Melchiorri, L.; Bagnara, G.P.; Bonsi, L.; Alviano, F.; Lanzoni, G.; Moretti, S.; Cuneo, A.; et al. A functional role for soluble HLA-G antigens in immune modulation mediated by mesenchymal stromal cells. Cytotherapy 2008, 10, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Morandi, F.; Raffaghello, L.; Bianchi, G.; Meloni, F.; Salis, A.; Millo, E.; Ferrone, S.; Barnaba, V.; Pistoia, V. Immunogenicity of human mesenchymal stem cells in HLA-class I-restricted T-cell responses against viral or tumor-associated antigens. Stem Cells 2008, 26, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Montespan, F.; Deschaseaux, F.; Sensébé, L.; Carosella, E.D.; Rouas-Freiss, N. Osteodifferentiated mesenchymal stem cells from bone marrow and adipose tissue express HLA-G and display immunomodulatory properties in HLA-mismatched settings: implications in bone repair therapy. J Immunol Res 2014, 2014, 230346. [Google Scholar] [CrossRef] [PubMed]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; De Bruyn, C.; Bron, D.; Toungouz, M.; Lagneaux, L. Immune-related antigens, surface molecules and regulatory factors in human-derived mesenchymal stromal cells: the expression and impact of inflammatory priming. Stem Cell Rev Rep 2012, 8, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Pistoia, V.; Moretta, L. Mesenchymal stem cells: a new strategy for immunosuppression? Trends Immunol 2007, 28, 219–226. [Google Scholar] [CrossRef]
- Sun, G.; Wang, Y.; Ti, Y.; Wang, J.; Zhao, J.; Qian, H. Regulatory B cell is critical in bone union process through suppressing proinflammatory cytokines and stimulating Foxp3 in Treg cells. Clin Exp Pharmacol Physiol 2017, 44, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Meednu, N.; Rosenberg, A.; Rangel-Moreno, J.; Wang, V.; Glanzman, J.; Owen, T.; Zhou, X.; Zhang, H.; Boyce, B.F.; et al. B cells inhibit bone formation in rheumatoid arthritis by suppressing osteoblast differentiation. Nat Commun 2018, 9, 5127. [Google Scholar] [CrossRef]
- van Mierlo, G.J.; Scherer, H.U.; Hameetman, M.; Morgan, M.E.; Flierman, R.; Huizinga, T.W.; Toes, R.E. Cutting edge: TNFR-shedding by CD4+CD25+ regulatory T cells inhibits the induction of inflammatory mediators. J Immunol 2008, 180, 2747–2751. [Google Scholar] [CrossRef]
- Ono, T.; Okamoto, K.; Nakashima, T.; Nitta, T.; Hori, S.; Iwakura, Y.; Takayanagi, H. IL-17-producing γδ T cells enhance bone regeneration. Nat Commun 2016, 7, 10928. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhao, Q.; Gu, B.; Yin, C.; Shen, K.; Tang, H.; Xia, H.; Zhang, X.; Zhao, Y.; Yang, X.; et al. Minimally invasive implantation and decreased inflammation reduce osteoinduction of biomaterial. Theranostics 2020, 10, 3533–3545. [Google Scholar] [CrossRef]
- Spector, J.A.; Mehrara, B.J.; Greenwald, J.A.; Saadeh, P.B.; Steinbrech, D.S.; Bouletreau, P.J.; Smith, L.P.; Longaker, M.T. Osteoblast expression of vascular endothelial growth factor is modulated by the extracellular microenvironment. Am J Physiol Cell Physiol 2001, 280, C72–80. [Google Scholar] [CrossRef] [PubMed]
- Hazehara-Kunitomo, Y.; Hara, E.S.; Ono, M.; Aung, K.T.; Komi, K.; Pham, H.T.; Akiyama, K.; Okada, M.; Oohashi, T.; Matsumoto, T.; et al. Acidic Pre-Conditioning Enhances the Stem Cell Phenotype of Human Bone Marrow Stem/Progenitor Cells. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Elabd, C.; Ichim, T.E.; Miller, K.; Anneling, A.; Grinstein, V.; Vargas, V.; Silva, F.J. Comparing atmospheric and hypoxic cultured mesenchymal stem cell transcriptome: implication for stem cell therapies targeting intervertebral discs. J Transl Med 2018, 16, 222. [Google Scholar] [CrossRef]
- Jadaun, P.K.; Zhang, S.; Koedam, M.; Demmers, J.; Chatterjee, S.; van Leeuwen, J.P.; van der Eerden, B.C. Inhibition of hypoxia-induced Mucin 1 alters the proteomic composition of human osteoblast-produced extracellular matrix, leading to reduced osteogenic and angiogenic potential. J Cell Physiol 2022, 237, 1440–1454. [Google Scholar] [CrossRef]
- Hu, S.; Qiao, L.; Cheng, K. Generation and Manipulation of Exosomes. Methods Mol Biol 2021, 2158, 295–305. [Google Scholar] [CrossRef]
- Tatullo, M.; Marrelli, B.; Zullo, M.J.; Codispoti, B.; Paduano, F.; Benincasa, C.; Fortunato, F.; Scacco, S.; Zavan, B.; Cocco, T. Exosomes from Human Periapical Cyst-MSCs: Theranostic Application in Parkinson's Disease. Int J Med Sci 2020, 17, 657–663. [Google Scholar] [CrossRef]
- Chachques, J.C.; Gardin, C.; Lila, N.; Ferroni, L.; Migonney, V.; Falentin-Daudre, C.; Zanotti, F.; Trentini, M.; Brunello, G.; Rocca, T.; et al. Elastomeric Cardiowrap Scaffolds Functionalized with Mesenchymal Stem Cells-Derived Exosomes Induce a Positive Modulation in the Inflammatory and Wound Healing Response of Mesenchymal Stem Cell and Macrophage. Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Gandolfi, M.G.; Gardin, C.; Zamparini, F.; Ferroni, L.; Esposti, M.D.; Parchi, G.; Ercan, B.; Manzoli, L.; Fava, F.; Fabbri, P.; et al. Mineral-Doped Poly(L-lactide) Acid Scaffolds Enriched with Exosomes Improve Osteogenic Commitment of Human Adipose-Derived Mesenchymal Stem Cells. Nanomaterials (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Zuo, R.; Liu, M.; Wang, Y.; Li, J.; Wang, W.; Wu, J.; Sun, C.; Li, B.; Wang, Z.; Lan, W.; et al. BM-MSC-derived exosomes alleviate radiation-induced bone loss by restoring the function of recipient BM-MSCs and activating Wnt/β-catenin signaling. Stem Cell Res Ther 2019, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, L.; Rong, Y.; Qian, D.; Chen, J.; Zhou, Z.; Luo, Y.; Jiang, D.; Cheng, L.; Zhao, S.; et al. Hypoxic mesenchymal stem cell-derived exosomes promote bone fracture healing by the transfer of miR-126. Acta Biomater 2020, 103, 196–212. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, J.; Jing, W. Motivating role of type H vessels in bone regeneration. Cell Prolif 2020, 53, e12874. [Google Scholar] [CrossRef] [PubMed]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef]
- Guder, C.; Gravius, S.; Burger, C.; Wirtz, D.C.; Schildberg, F.A. Osteoimmunology: A Current Update of the Interplay Between Bone and the Immune System. Front Immunol 2020, 11, 58. [Google Scholar] [CrossRef]
- Sasse, S.; Skorska, A.; Lux, C.A.; Steinhoff, G.; David, R.; Gaebel, R. Angiogenic Potential of Bone Marrow Derived CD133(+) and CD271(+) Intramyocardial Stem Cell Trans- Plantation Post MI. Cells 2019, 9. [Google Scholar] [CrossRef]
- Schmidt-Bleek, K.; Schell, H.; Lienau, J.; Schulz, N.; Hoff, P.; Pfaff, M.; Schmidt, G.; Martin, C.; Perka, C.; Buttgereit, F.; et al. Initial immune reaction and angiogenesis in bone healing. J Tissue Eng Regen Med 2014, 8, 120–130. [Google Scholar] [CrossRef]
- Chen, X.; He, W.; Sun, M.; Yan, Y.; Pang, Y.; Chai, G. STING inhibition accelerates the bone healing process while enhancing type H vessel formation. Faseb j 2021, 35, e21964. [Google Scholar] [CrossRef]
- Gao, B.; Deng, R.; Chai, Y.; Chen, H.; Hu, B.; Wang, X.; Zhu, S.; Cao, Y.; Ni, S.; Wan, M.; et al. Macrophage-lineage TRAP+ cells recruit periosteum-derived cells for periosteal osteogenesis and regeneration. J Clin Invest 2019, 129, 2578–2594. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Wang, G.; Zhang, B.; Wang, C.; Li, D.; Ding, C.; Wei, Q.; Fan, Z.; Tang, H.; et al. Single-Cell RNA Sequencing Reveals B Cells Are Important Regulators in Fracture Healing. Front Endocrinol (Lausanne) 2021, 12, 666140. [Google Scholar] [CrossRef]
- Avin, K.G.; Dominguez, J.M., 2nd; Chen, N.X.; Hato, T.; Myslinski, J.J.; Gao, H.; Liu, Y.; McKinley, T.O.; Brown, K.M.; Moe, S.M.; et al. Single-cell RNAseq provides insight into altered immune cell populations in human fracture nonunions. J Orthop Res 2023, 41, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Sivaraj, K.K.; Jeong, H.W.; Dharmalingam, B.; Zeuschner, D.; Adams, S.; Potente, M.; Adams, R.H. Regional specialization and fate specification of bone stromal cells in skeletal development. Cell Rep 2021, 36, 109352. [Google Scholar] [CrossRef] [PubMed]
- Papaccio, F.; Paino, F.; Regad, T.; Papaccio, G.; Desiderio, V.; Tirino, V. Concise Review: Cancer Cells, Cancer Stem Cells, and Mesenchymal Stem Cells: Influence in Cancer Development. Stem Cells Transl Med 2017, 6, 2115–2125. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.; Santos, S.G.; Gonçalves, R.M.; Teixeira, J.H.; Barbosa, M.A.; Almeida, M.I. Genetically Engineered-MSC Therapies for Non-unions, Delayed Unions and Critical-size Bone Defects. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Mahon, O.R.; Browe, D.C.; Gonzalez-Fernandez, T.; Pitacco, P.; Whelan, I.T.; Von Euw, S.; Hobbs, C.; Nicolosi, V.; Cunningham, K.T.; Mills, K.H.G.; et al. Nano-particle mediated M2 macrophage polarization enhances bone formation and MSC osteogenesis in an IL-10 dependent manner. Biomaterials 2020, 239, 119833. [Google Scholar] [CrossRef]
- Humbert, P.; Brennan, M.; Davison, N.; Rosset, P.; Trichet, V.; Blanchard, F.; Layrolle, P. Immune Modulation by Transplanted Calcium Phosphate Biomaterials and Human Mesenchymal Stromal Cells in Bone Regeneration. Front Immunol 2019, 10, 663. [Google Scholar] [CrossRef]
- Hou, Y.; Cai, K.; Li, J.; Chen, X.; Lai, M.; Hu, Y.; Luo, Z.; Ding, X.; Xu, D. Effects of titanium nanoparticles on adhesion, migration, proliferation, and differentiation of mesenchymal stem cells. Int J Nanomedicine 2013, 8, 3619–3630. [Google Scholar] [CrossRef]
- Takebe, J.; Champagne, C.M.; Offenbacher, S.; Ishibashi, K.; Cooper, L.F. Titanium surface topography alters cell shape and modulates bone morphogenetic protein 2 expression in the J774A.1 macrophage cell line. J Biomed Mater Res A 2003, 64, 207–216. [Google Scholar] [CrossRef]
- Refai, A.K.; Textor, M.; Brunette, D.M.; Waterfield, J.D. Effect of titanium surface topography on macrophage activation and secretion of proinflammatory cytokines and chemokines. J Biomed Mater Res A 2004, 70, 194–205. [Google Scholar] [CrossRef]
- Smith, B.S.; Capellato, P.; Kelley, S.; Gonzalez-Juarrero, M.; Popat, K.C. Reduced in vitro immune response on titania nanotube arrays compared to titanium surface. Biomater Sci 2013, 1, 322–332. [Google Scholar] [CrossRef]
- Mendonça, G.; Mendonça, D.B.; Aragão, F.J.; Cooper, L.F. Advancing dental implant surface technology--from micron- to nanotopography. Biomaterials 2008, 29, 3822–3835. [Google Scholar] [CrossRef]
- Dalby, M.J.; Gadegaard, N.; Tare, R.; Andar, A.; Riehle, M.O.; Herzyk, P.; Wilkinson, C.D.; Oreffo, R.O. The control of human mesenchymal cell differentiation using nanoscale symmetry and disorder. Nat Mater 2007, 6, 997–1003. [Google Scholar] [CrossRef]
- Paul, N.E.; Skazik, C.; Harwardt, M.; Bartneck, M.; Denecke, B.; Klee, D.; Salber, J.; Zwadlo-Klarwasser, G. Topographical control of human macrophages by a regularly microstructured polyvinylidene fluoride surface. Biomaterials 2008, 29, 4056–4064. [Google Scholar] [CrossRef]
- Qiu, P.; Li, M.; Chen, K.; Fang, B.; Chen, P.; Tang, Z.; Lin, X.; Fan, S. Periosteal matrix-derived hydrogel promotes bone repair through an early immune regulation coupled with enhanced angio- and osteogenesis. Biomaterials 2020, 227, 119552. [Google Scholar] [CrossRef]
- Lin, H.; Sohn, J.; Shen, H.; Langhans, M.T.; Tuan, R.S. Bone marrow mesenchymal stem cells: Aging and tissue engineering applications to enhance bone healing. Biomaterials 2019, 203, 96–110. [Google Scholar] [CrossRef]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? Comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev 2012, 21, 2724–2752. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Locatelli, F.; Fibbe, W.E. Mesenchymal stromal cells. Ann N Y Acad Sci 2009, 1176, 101–117. [Google Scholar] [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun Signal 2011, 9, 12. [Google Scholar] [CrossRef]
- Melief, S.M.; Zwaginga, J.J.; Fibbe, W.E.; Roelofs, H. Adipose tissue-derived multipotent stromal cells have a higher immunomodulatory capacity than their bone marrow-derived counterparts. Stem Cells Transl Med 2013, 2, 455–463. [Google Scholar] [CrossRef]
- Ding, H.; Gao, Y.S.; Wang, Y.; Hu, C.; Sun, Y.; Zhang, C. Dimethyloxaloylglycine increases the bone healing capacity of adipose-derived stem cells by promoting osteogenic differentiation and angiogenic potential. Stem Cells Dev 2014, 23, 990–1000. [Google Scholar] [CrossRef]
- Zimmerlin, L.; Donnenberg, V.S.; Pfeifer, M.E.; Meyer, E.M.; Péault, B.; Rubin, J.P.; Donnenberg, A.D. Stromal vascular progenitors in adult human adipose tissue. Cytometry A 2010, 77, 22–30. [Google Scholar] [CrossRef]
- Storti, G.; Scioli, M.G.; Kim, B.S.; Orlandi, A.; Cervelli, V. Adipose-Derived Stem Cells in Bone Tissue Engineering: Useful Tools with New Applications. Stem Cells Int 2019, 2019, 3673857. [Google Scholar] [CrossRef]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J Cell Biochem 2006, 99, 1233–1239. [Google Scholar] [CrossRef]
- Jimi, E.; Hirata, S.; Shin, M.; Yamazaki, M.; Fukushima, H. Molecular mechanisms of BMP-induced bone formation: Cross-talk between BMP and NF-κB signaling pathways in osteoblastogenesis. Japanese Dental ence Review 2010, 46, 33–42. [Google Scholar] [CrossRef]
- Long, F. Building strong bones: molecular regulation of the osteoblast lineage. Nat Rev Mol Cell Biol 2011, 13, 27–38. [Google Scholar] [CrossRef]
- Shafaei, H.; Kalarestaghi, H. Adipose-derived stem cells: An appropriate selection for osteogenic differentiation. J Cell Physiol 2020, 235, 8371–8386. [Google Scholar] [CrossRef]
- Yang, J.; Andre, P.; Ye, L.; Yang, Y.Z. The Hedgehog signalling pathway in bone formation. Int J Oral Sci 2015, 7, 73–79. [Google Scholar] [CrossRef]
- Senarath-Yapa, K.; McArdle, A.; Renda, A.; Longaker, M.T.; Quarto, N. Adipose-derived stem cells: a review of signaling networks governing cell fate and regenerative potential in the context of craniofacial and long bone skeletal repair. Int J Mol Sci 2014, 15, 9314–9330. [Google Scholar] [CrossRef]
- Malladi, P.; Xu, Y.; Yang, G.P.; Longaker, M.T. Functions of vitamin D, retinoic acid, and dexamethasone in mouse adipose-derived mesenchymal cells. Tissue Eng 2006, 12, 2031–2040. [Google Scholar] [CrossRef]
- Wang, C.Z.; Chen, S.M.; Chen, C.H.; Wang, C.K.; Wang, G.J.; Chang, J.K.; Ho, M.L. The effect of the local delivery of alendronate on human adipose-derived stem cell-based bone regeneration. Biomaterials 2010, 31, 8674–8683. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Wang, J.; Liu, Y.; Yu, Q.; Liu, Y.; Deng, N.; Liu, J. Functional Selenium Nanoparticles Enhanced Stem Cell Osteoblastic Differentiation through BMP Signaling Pathways. John Wiley & Sons, Ltd 2014.
- Scioli, M.G.; Bielli, A.; Gentile, P.; Cervelli, V.; Orlandi, A. Combined treatment with platelet-rich plasma and insulin favours chondrogenic and osteogenic differentiation of human adipose-derived stem cells in three-dimensional collagen scaffolds. J Tissue Eng Regen Med 2017, 11, 2398–2410. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J Biol Chem 1999, 274, 10689–10692. [Google Scholar] [CrossRef] [PubMed]
- Herzmann, N.; Salamon, A.; Fiedler, T.; Peters, K. Lipopolysaccharide induces proliferation and osteogenic differentiation of adipose-derived mesenchymal stromal cells in vitro via TLR4 activation. Exp Cell Res 2017, 350, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hess, K.; Ushmorov, A.; Fiedler, J.; Brenner, R.E.; Wirth, T. TNFalpha promotes osteogenic differentiation of human mesenchymal stem cells by triggering the NF-kappaB signaling pathway. Bone 2009, 45, 367–376. [Google Scholar] [CrossRef]
- Fiedler, T.; Salamon, A.; Adam, S.; Herzmann, N.; Taubenheim, J.; Peters, K. Impact of bacteria and bacterial components on osteogenic and adipogenic differentiation of adipose-derived mesenchymal stem cells. Exp Cell Res 2013, 319, 2883–2892. [Google Scholar] [CrossRef]
Figure 1.
Role of immune cells during the four stages of fracture healing: most of the activity of immune cells occurs early in fracture healing, and macrophages accompany almost the entire process of fracture healing.
Figure 1.
Role of immune cells during the four stages of fracture healing: most of the activity of immune cells occurs early in fracture healing, and macrophages accompany almost the entire process of fracture healing.
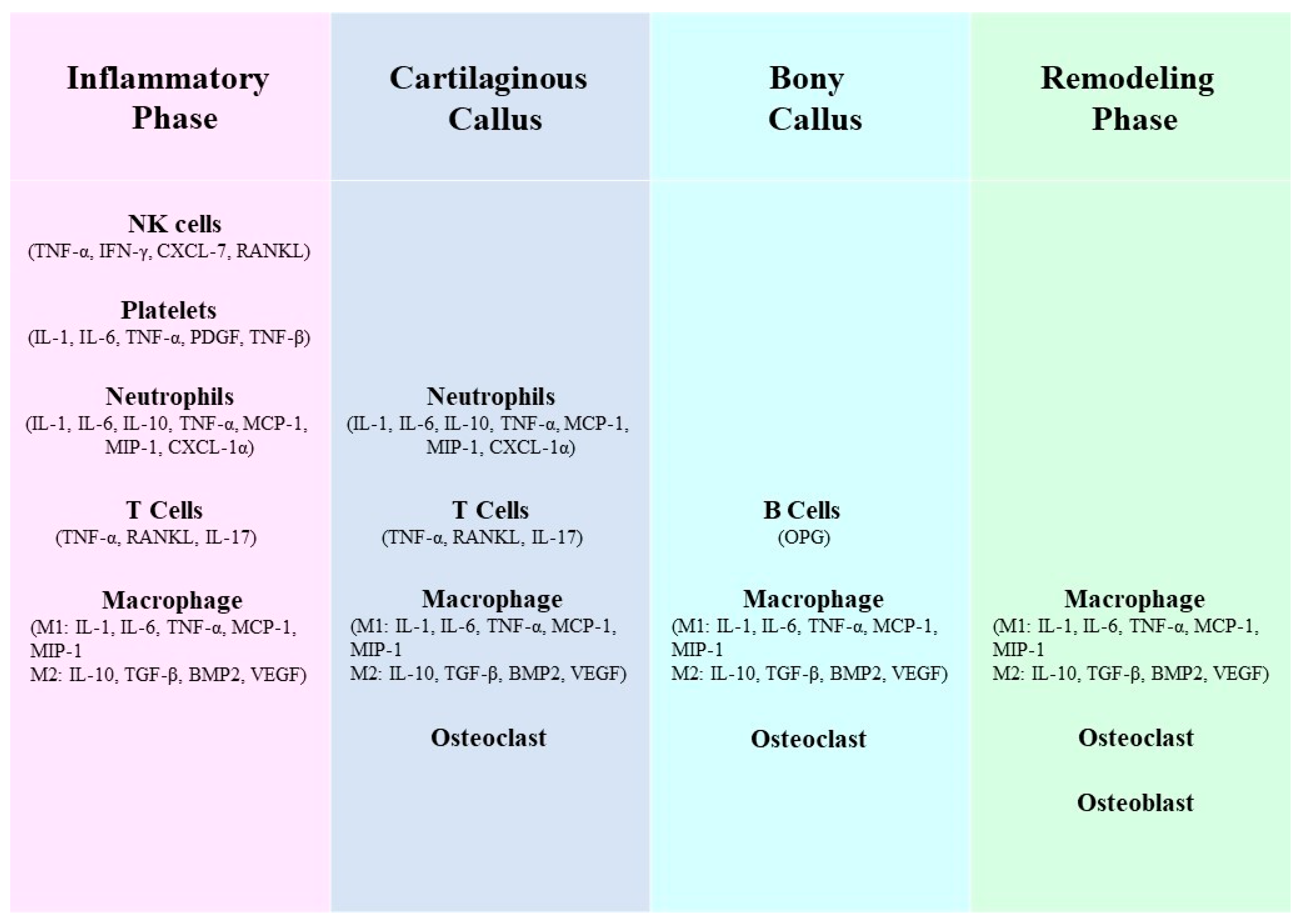
Figure 2.
The interaction between MSCs and Macrophages.
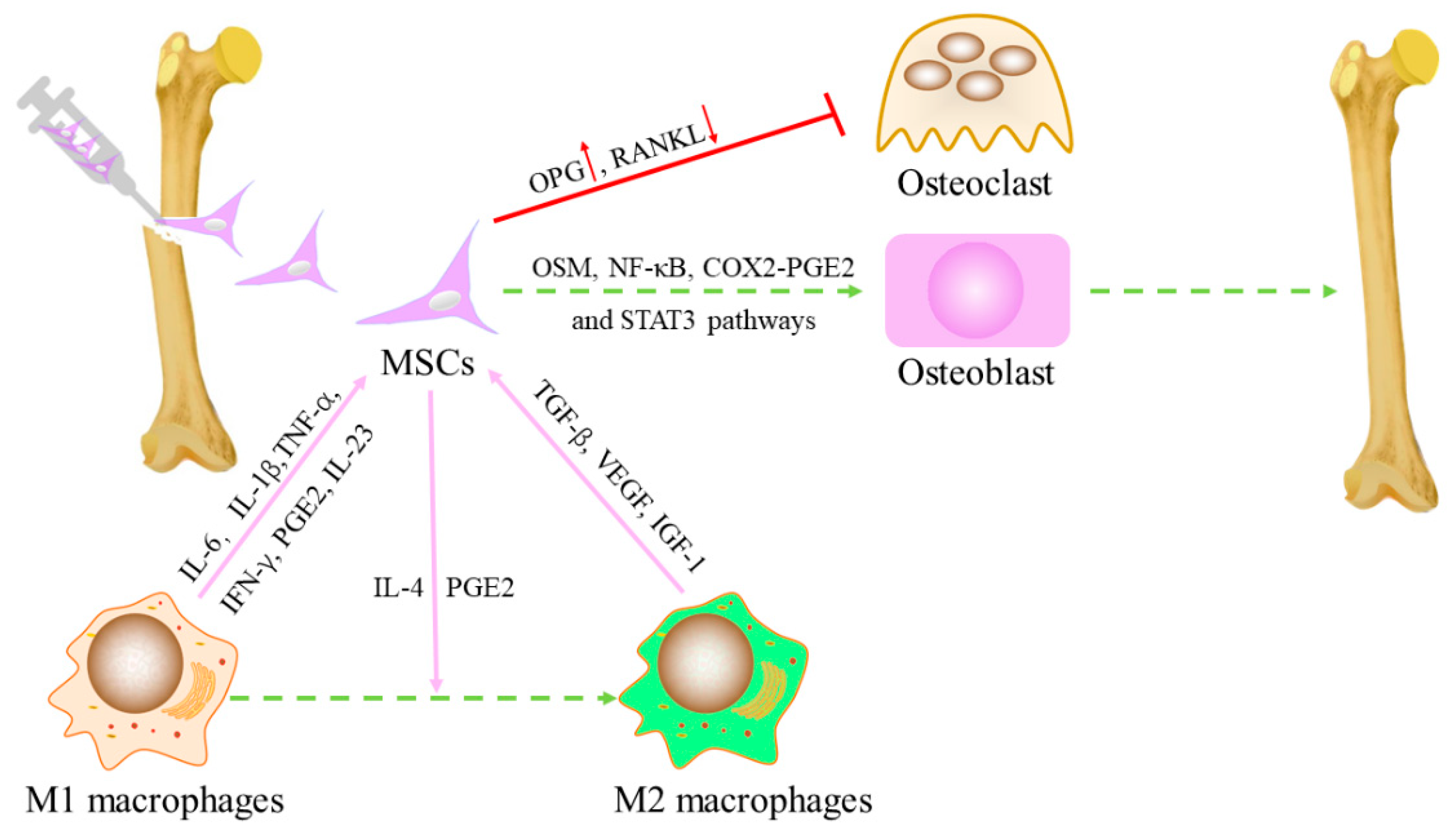
Figure 3.
The mechanism of interaction between MSC and immune cells.
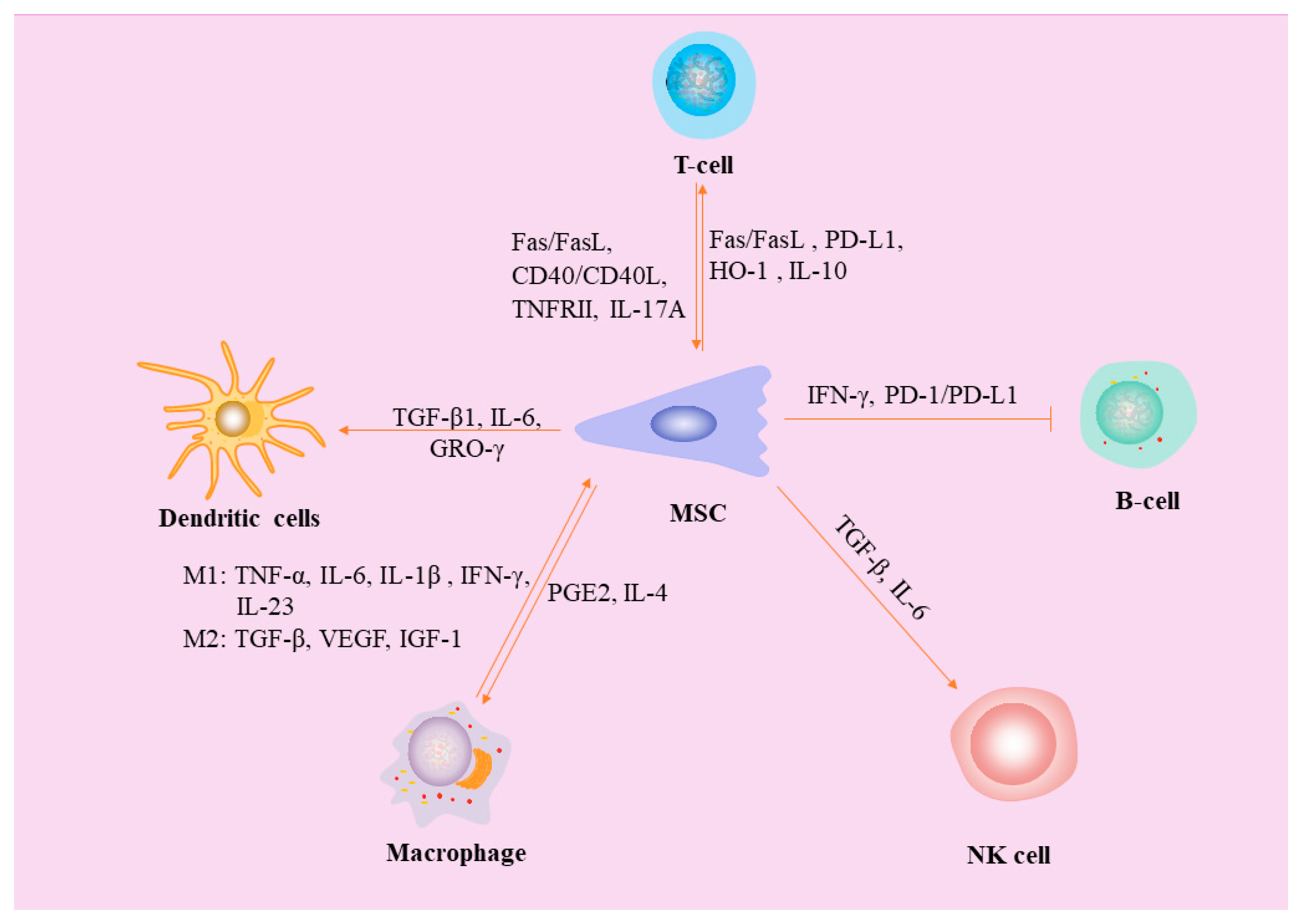
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



