
Submitted:
28 August 2023
Posted:
29 August 2023
You are already at the latest version
Abstract
Pancreatic ductal adenocarcinoma (PDAC) a highly malignant neoplasm, is classified as one of the most severe and devastating types of cancer. PDAC is a notable malignancy that exhibits a discouraging prognosis and a rising occurrence. The interplay between diabetes and pancreatic cancer exhibits a reciprocal causation. The identified metabolic disorder has been observed to possess noteworthy consequences on health outcomes, resulting in elevated rates of morbidity. The principal mechanisms involve the suppression of the immune system, the activation of pancreatic stellate cells (PSCs), and the onset of systemic metabolic disease caused by dysfunction of the islets. From this point forward, it is important to recognize that pancreatic cancer-related diabetes (PCRD) has the ability to increase the likelihood of developing pancreatic cancer. This highlights the complex relationship that exists between these two physiological states. Therefore, we investigate into the complex domain of PSCs, elucidating their intricate signaling pathways and the profound influence of chemokines on their behavior and final outcome. In order to surmount the obstacle of drug resistance and eliminate PDAC, researchers have undertaken extensive efforts to explore and cultivate novel natural compounds of the next generation. Additional investigation is necessary in order to comprehensively comprehend the effect of PCRD-mediated apoptosis on the progression and onset of PDAC through the utilization of natural compound. This study aims to examine the potential anticancer properties of natural compound in individuals with diabetes who are undergoing chemotherapy, targeted therapy, or immunotherapy. It is anticipated that these compounds will exhibit increased potency and possess enhanced pharmacological benefits. According to our research findings, it is indicated that naturally derived chemical compounds hold potential in the development of PDAC therapies that are both safe and efficacious.
Keywords:
Pancreatic ductal adenocarcinoma
; pancreatic stellate cells
; pancreatic cancer-related diabetes
; natural product
1. Introduction
Based on estimates, it is anticipated that PDAC, which represents the prevailing manifestation of pancreatic cancer, will ascend to the position of the second most prominent contributor to cancer-associated mortality on a global scale by the year 2030 [1,2]. Furthermore, the delayed detection of PDAC plays a crucial role in contributing to a worse prognosis. The prevalence of metastatic spreading and subsequent infiltration of important arterial structures has led to a significant observation that over 80% of cases lack the suitability for surgical removal of neoplastic growths [3]. Therefore, the discovery of new therapeutic approaches for PDAC is a pressing and unresolved issue in the field of medical research [4]. The gene known as Kirsten rat sarcoma virus (KRAS) is responsible for the synthesis of the Kirsten rat sarcoma viral oncogene homolog, a protein that plays a pivotal role in cellular signaling pathways. It is noteworthy to mention that mutations in the KRAS gene have been detected in roughly 90% of patients diagnosed with PDAC. The aforementioned alterations have an impact on both the control of gene expression and the amplification of gene copies. Furthermore, the onset of cancer requires the presence of genetic alterations in key genes like TP53, CDKN2A, and SMAD4, which are responsible for encoding the tumor suppressor protein p53, cyclin-dependent kinase inhibitor 2A, and SMAD family member 4, respectively [5,6]. In a subset of patients harboring the KRAS mutation, it has been observed that a phenomenon known as concurrent gene amplification occurs, affecting approximately 4% of these individuals [7,8,9]. The BRAF gene, which serves as the downstream signal to the KRAS gene, undergoes mutations in approximately 3-4% of cases independent of KRAS mutations. The given text [9,10], can be interpreted as a numerical representation. In the context of biology, it is important to note Whilst the majority of KRAS mutations lead to a state of constant activity, it is worth noting that a minority of tumors harboring the KRASG12C mutation exhibit nucleotide cycling activity, which holds the potential for targeted intervention [11]. The identification of PDAC subtypes holds significant value in terms of prognostic implications and the potential for subtype-specific therapeutic approaches. However, it is important to note that laboratory models capable of accurately identifying each PDAC subtype require additional prospective validation before they can be routinely integrated into clinical practice [12]. The PDAC microenvironment is distinguished by an abundance of malignant epithelial cells, a substantial presence of stroma primarily composed of immunosuppressive T cells and myelosuppressive-type macrophages (M2), and a state of reduced vascularity [12]. Thus, PSC have a pivotal function in the generation and renewal of the stroma. Upon stimulation by growth factors such as TGFβ1, platelet-derived growth factor (PDGF), and fibroblast growth factor, these cells exhibit the secretion of collagen and other constituents of the extracellular matrix [13,14]. The stroma, which is present in PDAC, a malignancy that originates in the pancreas, can comprise a substantial proportion, varying from 80% to 90%, of the total tumor mass. The stroma has been observed to exhibit a partial association with the proliferation of PSCs. The pancreas is a complex organ in which PSCs play a crucial role as the main progenitors involved in the formation [15,16,17]. Moreover, various stimulus encompasses many states, namely hyperglycemia, obesity, and hyperinsulinemia. Upon initiation, PSCs secrete specific signaling molecules referred to as cytokines, which are involved in facilitating diverse mechanisms within neoplastic cells. The aforementioned mechanisms encompass heightened cellular proliferation and invasion inside tumors, promotion of metastasis, initiation of epithelial-mesenchymal transition (EMT), and acquisition of resistance to chemotherapy [18,19,20,21,22,23,24,25,26,27]. PSCs exhibit a state of inactivity in the normal pancreas and exhibit the presence of alpha-smooth muscle actin (α-SMA) upon activation, leading to their transition into a myofibroblast-like phenotype [28]. The activation of PSCs has a significant impact on the development of PDAC due to their increased production of growth factors and rich extracellular matrix [29,30]. The following section aims to provide a comprehensive overview of the existing knowledge regarding PDAC in light of the PSC hypothesis. This hypothesis was postulated to elucidate the mechanisms by which cancerous tissue contributes to the progression of disease.
PSC have emerged as a promising candidate for stem cell in PDAC
Recent research has conducted an analysis on a specific group of PSCs, revealing a probable resemblance in biological characteristics to pancreatic stem cells [31]. PSCs have been identified to have persistent intercellular communication and to impact the formation of pancreatic cancer cells (PCCs). PSCs have emerged as a crucial element in the physiological processes of PCCs, playing a significant role in tumor advancement, evasion of immune response, and resistance to chemotherapy in PDAC [32,33,34,35]. Glutamine (Gln) plays a pivotal role as a vital nutrient in the process of carcinogenesis, serving as the primary provider of carbon and nitrogen for several metabolic activities [36]. It has been revealed that PCCs and PSCs have Gln metabolism crosstalk. PSCs exhibited a greater level of gene expression for glutamine synthetase (GS) compared to PCCs. Furthermore, the increased expression of GS, whether observed in the tumor or stromal cells, was found to be significantly correlated with an unfavorable prognosis in patients diagnosed with PDAC. In this context, the activation of Wnt signaling triggers a direct interaction between the β-catenin/TCF7 complex and the GS promoter region, resulting in an upregulation of GS expression [37]. Notably, the crucial role of glutamine in modulating redox homeostasis through the synthesis of glutathione (GSH) and generation of nicotinamide adenine dinucleotide phosphate (NADPH) is evident in several populations of cancer stem cells (CSCs). Interestingly, researches have demonstrated that the disruption of glutamine metabolism effectively hinders the process of self-renewal and reduces the expression of genes associated with stemness and pluripotency, achieved by the elevation of intracellular reactive oxygen species (ROS) levels [38,39,40,41,42]. Moreover, IGF2 mRNA translation and mature secretion are fastest and most efficient at high glutamine and glucose concentrations. Controlled secretion secretes IGF2, but unlike insulin, the intracellular pool is insufficient to support secretion and requires constant synthesis. Additionally, glutamine-induced IGF2 release increases Akt signaling. Nutrient-regulated beta cell IGF2 synthesis triggers the beta cell bulk and function-controlling IGF2/IGF1R autocrine loop [43]. Adult tissue-specific stem cells, also known as somatic stem cells, belong to a distinct cell population that exhibits remarkable characteristics such as self-renewal and the capacity to undergo differentiation into specialized cell types with specific functions. Pancreatic stem cells have garnered significant attention in the realm of research over the past decade due to their developmental potential and the advantageous connections they share with other tissue-derived stem cells in terms of developmental biology and anatomy [44,45]. PSCs, are a versatile and multifaceted cell population that can be observed in both the endocrine and exocrine components of pancreatic tissue. These cells make up approximately 7% of the total parenchymal cell population within the pancreas [46]. In recent studies, PSCs have emerged as a promising candidate for the generation of new β-cells, which are responsible for insulin production in the pancreas [47]. PSCs have the ability to undergo activation, resulting in the acquisition of a myofibroblast-like phenotype characterized by the expression of α-smooth muscle actin (α-SMA) as an activation marker protein. This activation process is accompanied by a decrease in the quantity of retinoid-containing fat droplets. Importantly, PSCs are implicated in the pathogenesis of islet fibrosis, a process that significantly contributes to the development of β-cell dysfunction [48]. Activated PSCs exhibit a distinct expression of α-SMA and actively secrete collagen I, collagen III, fibronectin, and various other extracellular matrix (ECM) components. The information included within the aforementioned PSC is effectively depicted in Figure 1. This concerted effort by activated PSCs contributes to the facilitation of pancreatic fibrosis. The identification of lipid droplets, along with the concomitant expression of GFAP, nestin, desmin, and vimentin, serves as a defining characteristic for the quiescent phenotype of PSCs [49]. The current collection of evidence pertaining to the association between PSCs and islets has revealed that PSCs possess the ability to not only stimulate the development of fibrous tissue in islets but also play a role in the endocrine functions associated with pancreatic diseases like glucose intolerance, along with the preservation of islet viability [50,51,52]. PSCs are strategically positioned in close proximity to the basolateral aspect of pancreatic acinar cells. They are primarily found surrounding small pancreatic ducts and blood vessels within the pancreatic tissue [46]. Hence, it is plausible that SP cells, nestin-positive cells, and PSCs could potentially occupy identical spatial positions. Taking into account the distinctive features encompassing the expression of nestin in SP cells and nestin-positive cells, it is plausible to posit that pancreatic stem/progenitor cells (PSCs) could potentially embody a comparable cellular entity within the adult pancreas, fulfilling analogous roles as stem/progenitor cells [53]. A recent discovery has been made regarding the presence of a specific group of cells expressing nestin in pancreatic islets and ducts [54]. The isolated nestin-positive islet-derived progenitor cells (NIPs), generated from adult pancreatic islets, have the ability to undergo differentiation in vitro, resulting in the development of cells exhibiting pancreatic exocrine, endocrine, and hepatic characteristics [55]. The existence of functional glucagon-like peptide-1 receptor (GLP-1Rs) on pancreatic progenitor cells implies a potential direct involvement of GLP-1 in the process of differentiating pancreatic progenitor cells into insulin-producing cells. Differentiation takes place within a specific fraction of NIP cells, which are stimulated to generate insulin. GLP-1 demonstrates a range of actions on β-cells [56,57,58]. The study revealed that the impact of GLP-1 analog on regular pancreatic stellate cells (PSCs) pertains to their capacity to induce the release of inflammatory mediators or promote proliferation. The expression and localization of the GLP-1 receptor (GLP-1R) in both normal pancreas and pancreatitis (AP/CP) tissues [59]. The available body of evidence indicates that there is a limited subset of cancer stem cells (CSCs) present among the PDAC cell population. CSCs are known to have significant involvement in the malignant characteristics of PDAC. Specifically, the identification of pancreatic CSCs is often achieved through the utilization of SP cells, the sphere-formation assay, and various CSC markers [60,61,62]. These elements collectively contribute to our understanding of the pivotal roles played by CSCs in the progression and behavior of PDAC [63]. Thus, PSCs exhibit the expression of many stem cell markers, possess multipotency, and demonstrate the capability to undergo effective differentiation into insulin-producing cells [64,65,66,67]. However, plasticity has the ability to safeguard CSCs and result in resistance. This is exemplified by the increased expression of enzymes and pathways involved in glutamine production, as well as the emergence of CSC subsets that are not reliant on glutamine. Plasticity facilitates the dynamic transition between CSC and non-CSC states, which are associated with distinct metabolic characteristics [68].
Type 2 diabetes is a significant prognostic indicator for pancreatic cancer
According to epidemiological study, individuals diagnosed with type 2 diabetes (T2D) have an increased likelihood of developing several common types of cancer. This analysis focuses on T2D due to the lack of clarity regarding the potential cancer risk linked with type 1 diabetes (T1D). The efficacy of cancer treatments may be impacted by the presence of diabetes. The management of glucose is impacted by cancer and its treatment, and emerging evidence suggests that cancer itself can further complicate the care of individuals with diabetes [69]. There are several biochemical pathways that have been identified as connecting obesity to cancer. These pathways include insulin resistance and anomalies in the IGF-I system, production and transport of sex hormones, subclinical chronic low-grade inflammation and oxidative stress, and alterations in adipokine pathogenesis [70]. Adipose tissue hypertrophy, hyperplasia, or a combination of both can be induced by excessive caloric intake [71]. Hypertrophic adipocytes have the ability to generate and release proinflammatory adipokines and cytokines, which in turn attract macrophages [72]. The presence of elevated macrophages and inflammatory cells leads to the induction of both local and systemic inflammation through the upregulation of cytokine production, specifically interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α) [73]. The cytokines TNF-α have been found to interfere with the signaling pathway of insulin, leading to insulin resistance in adipocytes and an elevated production of free fatty acids (FFAs). The presence of elevated levels of FFAs in non-adipose tissues such as the liver, skeletal muscle, and pancreatic β-cells has been found to be associated with the development of insulin resistance, steatosis, lipotoxicity, and metabolic dysfunction [74]. Notably, the phenomenon of intrapancreatic fat accumulation, which has a direct impact on the macro- and microenvironment of tumors, has been acknowledged as a potential indicator for predicting the extent of suffering experienced by patients with PDAC [75,76,77,78]. The production and release of insulin is exclusively attributed to pancreatic β cells, whereas the synthesis of IGF-1 primarily occurs in the liver. Both IGF-1 and insulin exhibit sequence similarities [79] and have the ability to trigger mitogenic pathways and inhibit apoptosis. The activation of the IGF-1–insulin pathway by obesity is known to induce intracellular signaling through MAPKs or the PI3K–AKT oncogenic cascade, as established in previous studies [79,80,81]. The activation of AKT and ERK signaling pathways by adipocytes leads to the occurrence of KRAS mutations in cancer cells. Leptin, adiponectin, lipocalin-2, and resistin are adipokines that exhibit high expression levels and play a significant role in fostering a proinflammatory milieu that promotes the advancement of cancer [82,83,84]. The available evidence indicates that individuals diagnosed with PDAC and T2D exhibit a higher presence of CD4+, CD68+, and CD8+T cells within their tissues. This implies that the adaptive immunological response of hyperinsulinemia may enhance the ability of tumors to evade the immune system and counteract immunosurveillance [85]. Research findings have shown that CD36 has a role in that infiltrate tumors, leading to the occurrence of lipid peroxidation. Consequently, the efficacy of interferon gamma (IFN-γ), a cytotoxic cytokine with anticancer properties, is diminished [86]. Obesity and T2D are linked by reduced β cell ability to overcome insulin resistance. It also increases systemic inflammation and innate immune response through lipotoxicity and glucotoxicity. IL-1b, TNF-a, IL-6, and IL-10 are increased in T2D [87]. Additional research suggests that β cell loss may be caused by different factors, such as endoplasmic reticulum stress, mitochondrial dysfunction, oxidative stress, inflammation, and islet amyloid buildup [88,89]. The research revealed a decline in cellular function, an increase in fasting blood sugar levels, and a moderate elevation in insulin resistance. Elevated levels of glucagon, somatostatin, and IAPP were seen in patients with PCRD. The restoration of hormone levels to a normal state was observed subsequent to the surgical removal of the tumor. This observation indicates that tumor cells have the ability to modify the amounts of glucagon, somatostatin, and IAPP [90]. The activation of PSCs occurs in response to inflammation and damage of the pancreas. Following the occurrence of an injury, immune cells are recruited to the affected site and subsequently secrete several cytokines, such as IL-1, TNF alpha, and IL-6. Additionally, growth factors including TGF β 1, FGF2, and PDGF angiotensin II are released [91]. The endocrine pancreas is responsible for the production and secretion of many hormones, including pancreatic polypeptide, glucagon, somatostatin, and insulin, which are then released into the bloodstream [92]. The researchers discovered that around 52% of these patients had a recorded medical record of new-onset diabetes, which had been diagnosed within the 2-3 years leading up to the diagnosis of PDAC [93,94]. Patients with PCRD demonstrate a notable diminishment in the dimensions of islets, accompanied by a decline in the quantity of β cells [95,96]. Patients with PDAC may manifest the early signs of diabetes mellitus around 2 to 3 years before their official cancer diagnosis. The specific group of patients under consideration presents the opportunity for early identification and acknowledgment. The potential molecular associations between the mechanisms of carcinogenesis and diabetes may potentially offer a technique to identify early indicators of PDAC. Understanding the mechanics of PCRD holds promise in identifying biomarkers for the early identification of illnesses [95]. In order to gain a comprehensive understanding of the relationship between obesity and cancer, it is imperative to examine the alterations that occur in adipose tissue during weight gain. Therefore, the prevention of obesity-induced activation of the insulin-IGF-1 axis represents a promising therapeutic target for cancer treatment [97]. Furthermore, this particular phenomenon possesses the inherent capability to unveil novel biological pathways, which may be strategically pinpointed for therapeutic interventions, ultimately resulting in improved patient outcomes.
Diabetes management strategies may reduce the risk of pancreatic cancer
Amyloid β (Aβ) is the constituent of the amyloid aggregates found in the brain affected by Alzheimer's disease (AD) [98]. Conversely, the amyloidogenic peptide deposit in the pancreatic islets of Langerhans associated with diabetes is known as islet amyloid polypeptide (IAPP), which is a peptide consisting of 37 amino acids [99,100]. The observed IC50 values of flavonoids containing a catechol moiety were found to be lower compared to those lacking this moiety. It indicates that the specific arrangement and number of hydroxyl groups on the aromatic ring play a critical role in determining the strength of aggregation inhibition. The prevention of amyloid aggregation has the potential to be therapeutically beneficial in addressing age-related conditions such as Alzheimer's disease and type 2 diabetes [101,102,103,104]. Phytochemicals, such as polyphenols and flavonoids, possess significant capabilities in the neutralization of free radicals and mitigation of oxidative harm, rendering them valuable in the management of DM [105]. The utilization of medicinal plant polyphenols has been found to have potential in reducing blood lipids and managing diabetes in individuals with diabetes. In a 9-month randomized, double-blind, placebo-controlled experiment (RCT) involving 240 individuals with prediabetes, it was observed that the group receiving curcumin did not develop type 2 diabetes mellitus (T2DM), while 16.4% of the participants in the placebo group did. Curcumin exhibited a protective effect on islet cells against the development of diabetes [106]. The anti-hyperglycemic effects of these substances are a result of their ability to competitively block α-amylase and α-glucosidase through binding to glucose transporters. By increasing the production of insulin from pancreatic B cells, attaching to receptors, reducing insulin resistance, and enhancing glucose tolerance, these plants may be able to combat diabetes. Improvements in glucose metabolism, B-cell size and activity, and plasma insulin, which decreases blood sugar, are further effects [107]. A randomized placebo-controlled trial was undertaken to investigate the impact of consuming an enriched bread containing a 1:1 combination of (-)-epicatechin and quercetin on the anthropometric and biochemical parameters of the participants. After a period of three months, the intake of enriched bread on a daily basis led to significant reductions in total cholesterol, LDL-cholesterol, total triglycerides, and fasting plasma glucose level [108]. The reduction of ROS formation by curcumin can be attributed to its impact on nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, which leads to an increase in the activity of antioxidant enzymes. Additionally, curcumin is involved in the modulation of the Nrf2-Keap1 pathway [109,110]. The synergistic impact of swertiamarin and quercetin is believed to contribute to the heightened activation of viable β-cells within the islets of Langerhans, leading to a more regulated secretion of insulin. The combination of swertiamarin and quercetin (CSQ) treatment substantially mitigated hyperlipidemia by reducing levels of total cholesterol, triglycerides, and LDL, while simultaneously increasing HDL levels [111]. After conducting a comprehensive scientific study over a duration of ten years, researchers have observed a correlation between the prescription of metformin, an oral biguanide medication, and a reduced occurrence of cancer in individuals diagnosed with type 2 diabetes mellitus (T2DM). This observation was made when comparing these individuals to patients who were prescribed insulin or sulfonylureas [112,113]. Naringenin, a compound found in certain plants, has been observed to exhibit inhibitory effects on the process of gluconeogenesis. Additionally, it has been noted to stimulate the upregulation of adenosine monophosphate-activated protein kinase (AMPK), a key enzyme involved in cellular energy regulation. These properties of naringenin have shown potential in the treatment of diabetes [114]. Numerous studies have provided evidence about the impact of antidiabetic medications (ADMs) on patient survival. However, there remains ongoing discussion and uncertainty surrounding the potential effects of metformin or other ADMs on clinical outcomes specifically in patients with PCRD. The association between DM and unfavorable PDAC outcomes has been shown, leading to uncertainty regarding the impact of ADM on survival. Additionally, it was found that PDAC with DM had higher levels of CA19-9, a biomarker associated with this kind of cancer. Furthermore, the study indicated an inverse relationship between CA19-9 levels and prognosis in PDAC patients [115,116]. Notably, the manifestation of elevated CA19-9 levels was observed as early as two years before the clinical diagnosis. Significantly, CA19-9 has proven to be a valuable tool in providing crucial lead-time for the identification of resectable disease, wherein the implementation of multimodality therapy strategies can result in enhanced long-term survival rates [117,118,119]. As a result, individuals experience relapses due to the presence of cancer stem cells or the development of medication resistance, ultimately resulting in mortality. The early identification of pancreatic cancer is crucial in order to enhance the likelihood of survival. The present investigation into biomarkers in pancreatic cancer (PC) reveals that the serum carbohydrate antigen, CA 19-9, stands as the sole existing biomarker exhibiting an estimated specificity of 90% for PDAC [120]. Consequently, it is imperative to note that there exists a diverse array of pharmaceutical interventions that are considered essential for addressing the condition known as CA19-9. However, it is crucial to emphasize that additional inquiries and research endeavors are imperative in order to attain a comprehensive comprehension of the therapeutic effectiveness of these medications. The study indicated above suggests the potential presence of molecules with anti-anti-type 2 diabetes mellitus (T2DM) properties in natural substances (Table 1).
Herbal medicine in vitro assay
The current focus in the field of global medicine is directed towards the exploration and analysis of traditional medicinal practices prevalent in various regions, such as Korea, China, Japan, Thailand, and India. Traditional Medicine holds a significant and prominent role within the realm of traditional medical practices [121]. Flavonoids that exhibit high activity levels frequently possess a catechol moiety, the functional group whose activity has recently been confirmed in other groups of polyphenolic compounds [122,123,124]. The extension of π-conjugation onto the carbonyl group in the C-ring, facilitated by the C2-C3 double bond, results in a greater radical scavenging capacity for unsaturated flavonoids compared to saturated structures like flavanones [124,125]. This particular study serves to emphasize the significance of the catechol moiety, a chemical structure commonly found in various biological compounds. Furthermore, a multitude of scientific investigations have provided evidence suggesting that the catechol moiety possesses the capacity to assume a crucial function in mitigating potential adverse effects associated with its utilization [126,127,128]. The utilization of antioxidant drugs serves as a viable approach to mitigate the detrimental effects of diseases that are instigated by oxidative stress. The catechol moiety, which is present in various antioxidants such as catecholamines and numerous flavonoids, plays a pivotal role as an antioxidant pharmacophore [124,129]. Naringenin, a polyphenolic flavanone compound, is abundantly present in various medicinal plants as well as citrus fruits such as grapefruits, tomatoes, and cherries. Apoptosis was induced in SNU-213 cells by the upregulation of ASK1, P38, P53, JNK, and ROS [130]. The study demonstrated that the administration of naringenin resulted in a decrease in gemcitabine resistance and a reduction in cancer cell invasion in the AsPC-1 and PANC-1 pancreatic cancer cell lines [131]. The investigation of combination therapy has been conducted with the aim of mitigating medication resistance and enhancing treatment effectiveness. The concurrent treatment of naringenin and hesperetin in BALB/c nude mice resulted in a greater inhibition of cell growth, invasion, and p38 activation in Miapaca-2, PANC-1, and SNU-213 cell lines compared to the individual administration of either drug [132]. The composition of QYHJ consists of a distinctive amalgamation of seven plant species [133]. A network was established to analyze the QYHJ target genes and PDAC target genes. Through this analysis, 11 overlapped genes were identified, which is in partial agreement with our earlier research on immune-related targets of QYHJ in the context of PDAC [134,135,136]. In order to conduct further validation, a set of six key genes was chosen. These genes encompassed many biological processes, including inflammation (HMOX1, ICAM1, VCAM1, and CCL2), oxidative stress (NQO1), and apoptosis (Bcl2) [137]. The study conducted by Cheng et al. provided evidence for the inhibitory effects of metformin on intracellular reactive oxygen species (ROS) levels in pancreatic cancer cell lines. This was achieved by the overexpression of manganese superoxide dismutase (MnSOD) and the downregulation of NADPH oxidase 2 (NOX2) and NADPH oxidase 4 (NOX4) in MiaPaCa-2 and PANC-1 cells [138]. A schematic demonstrating the network of protein-protein interactions (PPI) was created using the 26 possible target genes associated with the anti-prostate cancer (PC) effects of Xiang-lian pill (XLP). The findings indicate that MMP9, CASP8, CASP3, HSP90AA1, CTSB, MMP2, PTGS2, CASP9, IL4, and CTSD are the primary proteins involved in the anti-prostate cancer effects of XLP. These proteins are likely to have crucial functions and can be considered as potential target genes for the active ingredients of XLP [139]. The study revealed that OOE exhibited anti-cancer properties when tested on BxPC3, a cell line associated with pancreatic cancer. The OOE treatment modified the activation of ERK, Src, and STAT3 signaling pathways, thereby regulating the expression of STAT3-downstream genes that are associated with tumor formation. Additionally, the impact of OOE on cell viability, proliferation, and the induction of apoptotic effects and accumulation at the G2/M phase in BxPC3 cells was determined [140]. In MiaPaCa-2 and AsPC-1 pancreatic ductal adenocarcinoma (PDAC) cell lines, thymoquinone decreased proliferation, cell survival, and produced partial G2 cycle arrest and SubG0/G1 arrest and increased p21 mRNA expression, upregulated p53, downregulated HDAC activity, Bcl-2, HDAC 1, 2, and 3, and H4 acetylation were also triggered. Thymoquinone down-regulated Bcl-xL, Bcl-2, XIAP and up-regulated and activated pro-apoptotic molecules like caspases-3, -9, Bax, cytochrome c release, inhibitory tumor growth, Notch1, NICD, PTEN, Akt/mTOR/S6 signaling [141,142,143,144]. Wogonin (WOG), wogonoside, baicalein, and baicalin have been identified as bioactive compounds present in the roots of this particular plant [145]. Wogonin (WOG), wogonoside, baicalein, and baicalin are bioactive chemicals in this plant's roots [149]. PANC-1, Colo-357, and HPCCs4 human pancreatic cancer cells produced ROS after WOG treatment. Beclin-1/PI3K and Akt/ULK1/4E-BP1/CYLD signaling pathways were activated while mTOR was inhibited. WOG and other natural flavones increased p53 expression in Capan-1 and Colo-357 pancreatic cancer cells. These cells also inhibited Mcl-1, CDK-9, c-FLIP, and MDM2 expression [146,147]. The inhibitory effect of berberine (BBR) on cell proliferation was found to be more pronounced in BxPC-3 cells compared to HPDE-E6E7c7 cells. The administration of BBR resulted in a considerable increase in the activity of caspase-3 and -7 in both cell lines [148]. The utilization of natural chemicals is restricted by their inherent low bioavailability in the absence of structural modifications, hence posing challenges in their application [149]. However, it is important to note that natural product-based herbal medicine is currently being recognized as a groundbreaking therapeutic strategy for a multitude of diseases, including cancer. This is primarily due to its exceptional effectiveness and the limited occurrence of adverse reactions [150]. Hence, one can posit that natural products are poised to occupy a central role as a groundbreaking therapeutic approach for the treatment of cancer in the upcoming decade [151]. The aforementioned discovery implies the plausible existence of anti-cancer attributes within natural compounds, thereby promoting a sense of motivation to delve deeper into this field of scientific investigation (Table 2).
Natural compound in clinical trial
Several studies have been conducted, including clinical trials, to assess the effectiveness and safety of natural therapy for PDAC. The phenomenon of medication resistance, whether it is primary or acquired, has the potential to manifest both on-target and off-target [153]. The primary molecular target mutation of a medicine leads to on-target resistance, resulting in a reduction or complete loss of therapeutic effectiveness. Rapid and complex alterations by epigenetic modifications of tumor gene expression profiles contribute to the development of resistance to treatment [154]. European mistletoe (Viscum album L) are commonly employed in combination with traditional cancer treatments or as a standalone alternative therapy for individuals diagnosed with cancer [155]. Recent research has revealed potential implications of mistletoe in the prognosis of individuals diagnosed with colorectal or pancreatic carcinomas [156,157]. The findings of a randomized controlled trial demonstrated a significant increase in overall survival, from 2.7 months to 4.8 months, among patients diagnosed with advanced pancreatic cancer who were not eligible for antineoplastic therapy. This improvement in survival was shown in individuals who received subcutaneous mistletoe treatment, as compared to those who received the standard of supportive care [156,158]. QYHJ has been previously utilized as a supplemental medication in the treatment of PDAC, resulting in enhanced 1-, 3-, and 5-year survival rates among patients, without any apparent adverse effects [133]. Nevertheless, the mechanism by which QYHJ affects PDAC is yet to be fully understood, perhaps due to the intricate nature of the substances involved and their interacting effects within traditional Chinese medicine (TCM) formulae. Therefore, it is imperative to conduct comprehensive pharmacological investigations in the future, which will involve the examination of various combinations and concentrations [137]. The administration of XLP resulted in the inhibition of PC proliferation, while exhibiting little hepatotoxic and renal toxic effects. The pharmacological effects of XLP on PC encompass the modulation of many targets that operate within fat metabolism-related signaling pathways. These effects have significant potential in the regulation of aberrant tumor metabolism. There is a need for conducting extensive randomized controlled studies in order to provide a more robust body of data for the potential utilization of XLP in this setting of PDAC [139]. The administration of a combination of curcuminoids and gemcitabine for a median duration of two weeks in individuals diagnosed with advanced pancreatic cancer did not result in a statistically meaningful therapeutic outcome [159]. Hence, the use of a composite of pharmacological substances, each specifically formulated to target separate pathways, has the potential to provide a synergistic or potentiation effect. It is interesting to acknowledge that both of these occurrences have the potential to yield significant benefits in the context of cancer treatment. Individuals diagnosed with various types of solid tumors, such as breast, colorectal, and stomach cancer, were recruited to participate in a phase II clinical investigation that utilized a double-blind randomized trial methodology. The researchers made a significant observation in which the addition of curcuminoids to chemotherapy led to a decrease in negative side effects and an improvement in the general health of the participants in the study [160,161,162]. However, PDACs patient exhibit a remarkably low sensitivity, with a response rate of less than 5%, to immune checkpoint inhibitors [163,164]. The limited infiltration of T cells in PDAC can be partially attributable to the low number of mutations present in the tumor cells. This results in a dearth of neoantigens, which are molecules that can be recognized by the immune system and trigger strong T cell responses [164,165]. Neoantigens are proteins that are absent in healthy tissues and serve as markers for cancer cells, signaling their foreign nature to T cells. Consequently, PDACs may exhibit weak antigenicity and have a reduced number of infiltrating T cells [166]. CD3+ T cells are of paramount importance in the cellular immune response of the host and can be categorized into two subtypes: CD4+ helper T cells and CD8-+ cytotoxic T cells. The immune responses are enhanced by these cells through the production of lymphatic factors by CD4+ helper T cells, thereby stimulating additional lymphatic cells that are essential for exerting an antitumor effect [167]. Interestingly, there have been reports indicating that Korean Red ginseng possesses immune-modulating capabilities, which may lead to an enhancement in T-cell proliferation. This suggests that it has the potential to modulate cellular immunological responses [168,169]. The study revealed that the concurrent administration of Korean Red ginseng alongside adjuvant chemotherapy led to an increased count of CD4+ lymphocytes and a higher CD4+/CD8+ T lymphocyte ratio following chemotherapy in individuals diagnosed with bile duct or pancreatic cancer. There was no significant difference observed in the prevalence of neutropenia and liver dysfunction between the groups [170]. In addition to enhancing therapeutic efficacy, the utilization of a drug-herb combination concurrently diminishes the dosage magnitude of each individual pharmaceutical agent that is administered. One plausible therapeutic strategy involves the immunomodulation of malignant tumors in PDAC through the activation of D4+/CD8+ T cells. These specialized T cells possess the ability to identify and target neoantigens, which are unique antigens found on cancer cells. The aforementioned environment may be effectively addressed by the utilization of a combined approach involving conventional drugs, hence leading to the attainment of a potent antitumor immunotherapeutic response. The aforementioned clinical trials associated with natural substances are delineated in Table 3, thereby indicating the potential existence of anti-carcinogenic attributes. This, in turn, provides as a motivation for the pursuit of additional investigations within this field of knowledge.
Tendency in FDA-Approved Therapies
Gemcitabine in combination with nanoparticle albumin-bound paclitaxel (NAB-paclitaxel) has been identified as a potential second-line treatment option for patients who have previously undergone FOLFIRINOX (a chemotherapy regimen consisting of leucovorin (LV), 5-fluorouracil (5-FU), irinotecan, and oxaliplatin), possess a favorable comorbidity profile, and express a preference for and receive support for aggressive medical interventions. Patients who meet the criteria of receiving first-line gemcitabine plus NAB-paclitaxel, including a relatively favorable comorbidity profile, patient preference, and a support system for aggressive medical therapy, as well as having access to chemotherapy port and infusion pump management services, may be eligible for second-line therapy with 5-FU plus oxaliplatin, irinotecan, or nanoliposomal irinotecan. Gemcitabine or fluorouracil may serve as viable options for persons with cancer and comorbidities that restrict the use of more intensive treatment regimens, when employed as second-line therapy [171,172,173]. The utilization of nanoliposomal irinotecan (nal-IRI) alongside 5-FU and LV has been recognized as a viable approach in the treatment of PDAC cases that have developed resistance to gemcitabine-based therapy [174]. The therapeutic interventions available for individuals diagnosed with advanced pancreatic cancer (APC) encompass a spectrum of approaches, including the use of gemcitabine as a single agent or the implementation of multi-drug treatment protocols. The selection of a specific therapeutic option is contingent upon various factors such as the patient's age, performance status, presence of other medical diseases, and the preferences of both the patient and their healthcare provider [175]. The treatment landscape for metastatic pancreatic cancer becomes increasingly uncertain beyond first therapy, since patients frequently experience rapid clinical decline and are no longer eligible for further interventions beyond optimal supportive care [176]. Through a comprehensive analysis of publicly accessible bioinformatics data, it has been determined that the administration of FOLFIRINOX has a regulatory effect on tumor immunity. Herein, it was discovered the crucial protein C-X-C motif chemokine 5 (CXCL5), which was found to produce an immunosuppressive milieu in PDAC [177]. The gene CXCL5 is classified as a member of the C-X-C chemokine family, which is widely recognized for its role in attracting granulocytic immune cells by specifically binding to its receptor, C-X-C Chemokine Receptor Type 2 (CXCR2) [178]. The significance of CXCR2 lies in its involvement in the recruitment of tumor-associated neutrophils (TANs), together with CXCL8, CXCL6, and CXCR1. These factors collectively contribute to the suppression of anticancer immune responses [179]. Novel biomarkers, such as KRAS mutations, NTRK1–3 fusions, and BRCA1/2 mutations, have demonstrated potential for targeted therapy in individuals with PDAC. The medications AMG510, entrectinib, larotrectinib, and olaparib continue to be limited in their application and demonstrate suboptimal efficacy [180]. The inhibition of CXCL5 may present a promising option for targeted therapy, and conducting additional research on CXCL5 could facilitate the integration of FOLFIRINOX and immunotherapy [177]. In the field of PDAC, it has become customary to employ maintenance therapy subsequent to the administration of initial combination cytotoxic therapy for specific cohorts of patients. Arm A involved a six-month administration of FOLFIRINOX, while arm B received four months of 5-FU/LV maintenance therapy. Lastly, arm C underwent treatment with gemcitabine and FOLFIRI (5-FU, folic acid, irinotecan) on a bi-monthly basis. The investigation sought to assess the prospective efficacy of maintenance chemotherapy. In arms A, B, and C, the six-month progression-free survival (PFS) rates were observed to be 47%, 44%, and 34%, respectively. Additionally, the median overall survival (OS) durations were found to be 10.1, 11.2, and 7.3 months in arms A, B, and C, respectively [12,181]. The coadministration of gemcitabine with the FOLFIRINOX regimen, consisting of folinic acid, fluorouracil, irinotecan, and oxaliplatin, has been found to augment its efficacy in treating metastatic pancreatic cancer. However, this therapeutic approach has been associated with a notable rise in the incidence of side events and toxicities [175]. As a result, the combination of selective inhibitors and chemotherapeutic agents is increasingly being utilized as a method to alleviate the detrimental impacts on the human body [182,183]. The approval of Olaparib by the FDA is for the purpose of maintenance therapy in patients with PDAC who possess germline BRCA1/2 mutations [184]. The presence of wild-type KRAS in PDAC individuals signifies a distinct subgroup that exhibits an abundance of potentially exploitable oncogenic drivers. These drivers include ERBB inhibitors such as afatinib and MCLA-128, TRK inhibitors like larotrectinib and entrectinib, ALK/ROS inhibitor crizotinib, and the ongoing development of BRAF/MEK inhibitors. Within a limited population of individuals harboring the KRASG12C mutation, there is ongoing exploration into the therapeutic potential of the KRASG12C inhibitor known as AMG510, alongside other pharmacological compounds [12]. It is advisable to do regular examinations for microsatellite instability (MSI) or deficient mismatch repair (dMMR) in patients with PDAC who are eligible for checkpoint inhibitor therapy. While infrequent in PDAC, individuals with tumors exhibiting microsatellite instability-high (MSI-H) may experience significant advantages from the administration of programmed cell death protein (PD-1) inhibitor pembrolizumab. Consequently, it is advised to consider pembrolizumab as a potential treatment option for patients in the second-line or subsequent stages of therapy [171,185]. Pembrolizumab is a pharmacological intervention employed in the treatment of medical diseases defined by mismatch repair deficiency and microsatellite instability [186,187]. As previously elucidated, Table 4 provides a comprehensive summary of the existing pharmacological agents and their corresponding targets employed in the treatment of PDAC. The data presented in Table 4 elucidates the prevailing patterns in the utilization of chemotherapy.
2. Conclusions
As previously elucidated, the timely identification of PDAC holds paramount importance in enhancing the prognosis and overall well-being of afflicted individuals. The intriguing observation that a considerable proportion of patients diagnosed with PDAC present with the onset of diabetes mellitus offers a valuable avenue to identify a subgroup of individuals at heightened risk for developing PDAC [95]. The burgeoning significance of T2D in the pathogenesis of PDAC and the limited yet intriguing evidence regarding their prospective utility as diagnostic indicators and innovative therapeutic agents are indeed captivating. In light of the growing acknowledgement of the stroma's significance in the advancement of cancer, it would be imprudent to disregard the potential impact of stromal factors in the context of PCRD [188]. Such studies are essential for unraveling the underlying mechanisms of PCRD and establishing a comprehensive understanding of the intricate relationship between PCRD and the advancement of cancer [189]. Future research in the field of PDAC treatment should prioritize investigations into the potential synergistic effects of established anti-cancer drugs, as well as studies into the efficacy of natural products in mitigating the adverse effects associated with anti-cancer drug therapies [151]. Thus, naringenin, metformin, mistletoe, Xiang-lian pill (XLP), QYHJ and Korean red ginseng are currently being investigated for their potential therapeutic applications. It is anticipated that through demanding research and experimentation, these treatments may be further developed and contribute to the therapeutic options for cancer patients in the future. Notably, preliminary report presents findings from a randomized clinical trial investigating the effects of red ginseng on immunological control. The results of this study have garnered significant interest, highlighting the need for more research in this area. Such scientific pursuits possess the inherent capacity to unveil hitherto unexplored biomarkers and therapeutic targets, thereby facilitating the prompt detection and improved handling of this profoundly incapacitating ailment. The overarching objective is to effectively identify and encompass a substantial proportion of individuals afflicted with respectable, early stage PDAC who are presently being diagnosed belatedly.
Funding
This research was supported by the Graduate School Innovation Office, Kyung Hee University, a grant from Kyung Hee University in 2023 (KHU-20230914), Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2020R1I1A2066868), the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. 2020R1A5A2019413), a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HF20C0038), and the innovation network support Program through the INNOPOLIS funded by Ministry of Science and ICT (2022-IT-RD-0205-01-101).
References
- Quante, A.S.; Ming, C.; Rottmann, M.; Engel, J.; Boeck, S.; Heinemann, V.; Westphalen, C.B.; Strauch, K. Projections of cancer incidence and cancer-related deaths in Germany by 2020 and 2030. Cancer Med. 2016, 5, 2649–2656. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; et al. Locally advanced pancreatic adenocarcinoma: are we making progress?. Highlights from the "2011 ASCO Annual Meeting". Chicago, IL, USA; June 3-7, 2011. Jop, 2011. 12(4): p. 347-50.
- Thakur, G.; Kumar, R.; Kim, S.-B.; Lee, S.-Y.; Lee, S.-L.; Rho, G.-J. Therapeutic Status and Available Strategies in Pancreatic Ductal Adenocarcinoma. Biomedicines 2021, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, J.D.; et al. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Gong, Y.; Fan, Z.; Luo, G.; Huang, Q.; Deng, S.; Cheng, H.; Jin, K.; Ni, Q.; Yu, X.; et al. Molecular alterations and targeted therapy in pancreatic ductal adenocarcinoma. J. Hematol. Oncol. 2020, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Biankin, A.V.; Waddell, N.; Kassahn, K.S.; Gingras, M.-C.; Muthuswamy, L.B.; Johns, A.L.; Miller, D.K.; Wilson, P.J.; Patch, A.-M.; Wu, J.; et al. Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature 2012, 491, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Yachida, S.; et al. Clinical significance of the genetic landscape of pancreatic cancer and implications for identification of potential long-term survivors. Clin Cancer Res 2012, 18, 6339–6347. [Google Scholar] [CrossRef]
- Lowery, M.A.; Jordan, E.J.; Basturk, O.; Ptashkin, R.N.; Zehir, A.; Berger, M.F.; Leach, T.; Herbst, B.; Askan, G.; Maynard, H.; et al. Real-Time Genomic Profiling of Pancreatic Ductal Adenocarcinoma: Potential Actionability and Correlation with Clinical Phenotype. Clin. Cancer Res. 2017, 23, 6094–6100. [Google Scholar] [CrossRef]
- Waddell, N.; et al. Whole genomes redefine the mutational landscape of pancreatic cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef]
- Lito, P.; Solomon, M.; Li, L.-S.; Hansen, R.; Rosen, N. Allele-specific inhibitors inactivate mutant KRAS G12C by a trapping mechanism. Science 2016, 351, 604–608. [Google Scholar] [CrossRef]
- Singh, R.R.; O’reilly, E.M. New Treatment Strategies for Metastatic Pancreatic Ductal Adenocarcinoma. Drugs 2020, 80, 647–669. [Google Scholar] [CrossRef] [PubMed]
- Masamune, A.; Shimosegawa, T. Signal transduction in pancreatic stellate cells. J. Gastroenterol. 2009, 44, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Erkan, M.; Reiser-Erkan, C.; Michalski, C.W.; Deucker, S.; Sauliunaite, D.; Streit, S.; Esposito, I.; Friess, H.; Kleeff, J. Cancer-Stellate Cell Interactions Perpetuate the Hypoxia-Fibrosis Cycle in Pancreatic Ductal Adenocarcinoma. Neoplasia 2009, 11, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Bachem, M.G.; Schneider, E.; Groß, H.; Weidenbach, H.; Schmid, R.M.; Menke, A.; Siech, M.; Beger, H.; Grünert, A.; Adler, G. Identification, culture, and characterization of pancreatic stellate cells in rats and humans. Gastroenterology 1998, 115, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Apte, M.V.; Wilson, J.S.; Lugea, A.; Pandol, S.J. A Starring Role for Stellate Cells in the Pancreatic Cancer Microenvironment. Gastroenterology 2013, 144, 1210–1219. [Google Scholar] [CrossRef]
- Shields, M.A.; Dangi-Garimella, S.; Redig, A.J.; Munshi, H.G. Biochemical role of the collagen-rich tumour microenvironment in pancreatic cancer progression. Biochem. J. 2011, 441, 541–552. [Google Scholar] [CrossRef]
- Neesse, A.; Algül, H.; A Tuveson, D.; Gress, T.M. Stromal biology and therapy in pancreatic cancer: a changing paradigm. Gut 2015, 64, 1476–1484. [Google Scholar] [CrossRef]
- Waghray, M.; Yalamanchili, M.; di Magliano, M.P.; Simeone, D.M. Deciphering the role of stroma in pancreatic cancer. Curr. Opin. Gastroenterol. 2013, 29, 537–543. [Google Scholar] [CrossRef]
- Apte, M.V.; Haber, P.S.; Darby, S.J.; Rodgers, S.C.; Mccaughan, G.W.; Korsten, M.A.; Pirola, R.C.; Wilson, J.S. Pancreatic stellate cells are activated by proinflammatory cytokines: implications for pancreatic fibrogenesis. Gut 1999, 44, 534–541. [Google Scholar] [CrossRef]
- Incio, J.; et al. Obesity-Induced Inflammation and Desmoplasia Promote Pancreatic Cancer Progression and Resistance to Chemotherapy. Cancer Discov 2016, 6, 852–869. [Google Scholar] [CrossRef]
- Nomiyama, Y.; Tashiro, M.; Yamaguchi, T.; Watanabe, S.; Taguchi, M.; Asaumi, H.; Nakamura, H.; Otsuki, M. High Glucose Activates Rat Pancreatic Stellate Cells Through Protein Kinase C and p38 Mitogen-Activated Protein Kinase Pathway. Pancreas 2007, 34, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-H.; Hong, O.-K.; Kim, J.-W.; Ahn, Y.-B.; Song, K.-H.; Cha, B.-Y.; Son, H.-Y.; Kim, M.-J.; Jeong, I.-K.; Yoon, K.-H. High glucose increases extracellular matrix production in pancreatic stellate cells by activating the renin–angiotensin system. J. Cell. Biochem. 2006, 98, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Ryu, G.R.; Lee, E.; Chun, H.-J.; Yoon, K.-H.; Ko, S.-H.; Ahn, Y.-B.; Song, K.-H. Oxidative stress plays a role in high glucose-induced activation of pancreatic stellate cells. Biochem. Biophys. Res. Commun. 2013, 439, 258–263. [Google Scholar] [CrossRef]
- Kiss, K.; Baghy, K.; Spisák, S.; Szanyi, S.; Tulassay, Z.; Zalatnai, A.; Löhr, J.-M.; Jesenofsky, R.; Kovalszky, I.; Firneisz, G. Chronic Hyperglycemia Induces Trans-Differentiation of Human Pancreatic Stellate Cells and Enhances the Malignant Molecular Communication with Human Pancreatic Cancer Cells. PLOS ONE 2015, 10, e0128059. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Waldron, R.T.; Su, H.-Y.; Moro, A.; Chang, H.-H.; Eibl, G.; Ferreri, K.; Kandeel, F.R.; Lugea, A.; Li, L.; et al. Insulin promotes proliferation and fibrosing responses in activated pancreatic stellate cells. Am. J. Physiol. Liver Physiol. 2016, 311, G675–G687. [Google Scholar] [CrossRef]
- Tang, D.; et al. PSC-derived Galectin-1 inducing epithelial-mesenchymal transition of pancreatic ductal adenocarcinoma cells by activating the NF-κB pathway. Oncotarget 2017, 8, 86488–86502. [Google Scholar] [CrossRef]
- Shi, X.; Wang, M.; Zhang, Y.; Guo, X.; Liu, M.; Zhou, Z.; Zhao, Y.; He, R.; Gao, Y.; Liu, Y.; et al. Hypoxia activated HGF expression in pancreatic stellate cells confers resistance of pancreatic cancer cells to EGFR inhibition. EBioMedicine 2022, 86, 104352. [Google Scholar] [CrossRef]
- Shi, Y.; Gao, W.; Lytle, N.K.; Huang, P.; Yuan, X.; Dann, A.M.; Ridinger-Saison, M.; DelGiorno, K.E.; Antal, C.E.; Liang, G.; et al. Targeting LIF-mediated paracrine interaction for pancreatic cancer therapy and monitoring. Nature 2019, 569, 131–135. [Google Scholar] [CrossRef]
- Biffi, G.; Tuveson, D.A. Diversity and Biology of Cancer-Associated Fibroblasts. Physiol. Rev. 2021, 101, 147–176. [Google Scholar] [CrossRef]
- Augstein, P.M.; Loudovaris, T.; Bandala-Sanchez, E.; Heinke, P.M.; Naselli, G.B.; Lee, L.; Hawthorne, W.J.; Góñez, L.J.; Neale, A.M.B.; Vaillant, F.; et al. Characterization of the Human Pancreas Side Population as a Potential Reservoir of Adult Stem Cells. Pancreas 2018, 47, 25–34. [Google Scholar] [CrossRef]
- Murray, E.R.; Menezes, S.; Henry, J.C.; Williams, J.L.; Alba-Castellón, L.; Baskaran, P.; Quétier, I.; Desai, A.; Marshall, J.J.; Rosewell, I.; et al. Disruption of pancreatic stellate cell myofibroblast phenotype promotes pancreatic tumor invasion. Cell Rep. 2022, 38, 110227. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, Y.; Qian, F. Opportunities and delusions regarding drug delivery targeting pancreatic cancer-associated fibroblasts. Adv. Drug Deliv. Rev. 2021, 172, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; et al. Asporin promotes pancreatic cancer cell invasion and migration by regulating the epithelial-to-mesenchymal transition (EMT) through both autocrine and paracrine mechanisms. Cancer letters 2017, 398, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, H.; Ma, Q.; Huang, R.; Lu, J.; Liang, X.; Liu, X.; Zhang, Z.; Yu, L.; Pang, J.; et al. YAP1-mediated pancreatic stellate cell activation inhibits pancreatic cancer cell proliferation. Cancer Lett. 2019, 462, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Kamphorst, J.J.; Nofal, M.; Commisso, C.; Hackett, S.R.; Lu, W.; Grabocka, E.; Vander Heiden, M.G.; Miller, G.; Drebin, J.A.; Bar-Sagi, D.; et al. Human Pancreatic Cancer Tumors Are Nutrient Poor and Tumor Cells Actively Scavenge Extracellular Protein. Cancer Res. 2015, 75, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, H.; Liu, X.; Guo, W.; Liu, Q.; Chen, L.; Pang, J.; Liu, X.; Li, R.; Tong, W.-M.; et al. Pancreatic stellate cells exploit Wnt/β-catenin/TCF7-mediated glutamine metabolism to promote pancreatic cancer cells growth. Cancer Lett. 2023, 555, 216040. [Google Scholar] [CrossRef]
- Sharif, T.; Dai, C.; Martell, E.; Ghassemi-Rad, M.S.; Hanes, M.R.; Murphy, P.J.; Kennedy, B.E.; Venugopal, C.; Subapanditha, M.K.; Giacomantonio, C.A.; et al. TAp73 Modifies Metabolism and Positively Regulates Growth of Cancer Stem–Like Cells in a Redox-Sensitive Manner. Clin. Cancer Res. 2019, 25, 2001–2017. [Google Scholar] [CrossRef]
- Jaggupilli, A.; Ly, S.; Nguyen, K.; Anand, V.; Yuan, B.; El-Dana, F.; Yan, Y.; Arvanitis, Z.; Piyarathna, D.W.B.; Putluri, N.; et al. Metabolic stress induces GD2+ cancer stem cell-like phenotype in triple-negative breast cancer. Br. J. Cancer 2021, 126, 615–627. [Google Scholar] [CrossRef]
- Mukha, A.; Kahya, U.; Linge, A.; Chen, O.; Löck, S.; Lukiyanchuk, V.; Richter, S.; Alves, T.C.; Peitzsch, M.; Telychko, V.; et al. GLS-driven glutamine catabolism contributes to prostate cancer radiosensitivity by regulating the redox state, stemness and ATG5-mediated autophagy. Theranostics 2021, 11, 7844–7868. [Google Scholar] [CrossRef]
- Yoshikawa, M.; et al. xCT inhibition depletes CD44v-expressing tumor cells that are resistant to EGFR-targeted therapy in head and neck squamous cell carcinoma. Cancer Res 2013, 73, 1855–1866. [Google Scholar] [CrossRef]
- Li, D.; Fu, Z.; Chen, R.; Zhao, X.; Zhou, Y.; Zeng, B.; Yu, M.; Zhou, Q.; Lin, Q.; Gao, W.; et al. Inhibition of glutamine metabolism counteracts pancreatic cancer stem cell features and sensitizes cells to radiotherapy. Oncotarget 2015, 6, 31151–31163. [Google Scholar] [CrossRef] [PubMed]
- Modi, H.; Cornu, M.; Thorens, B. Glutamine Stimulates Biosynthesis and Secretion of Insulin-like Growth Factor 2 (IGF2), an Autocrine Regulator of Beta Cell Mass and Function. J. Biol. Chem. 2014, 289, 31972–31982. [Google Scholar] [CrossRef] [PubMed]
- Bouwens, L.; Houbracken, I.; Mfopou, J.K. The use of stem cells for pancreatic regeneration in diabetes mellitus. Nat. Rev. Endocrinol. 2013, 9, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Schwab, K.E.; Deane, J.A. Endometrial stem/progenitor cells: the first 10 years. Hum Reprod Update 2016, 22, 137–163. [Google Scholar] [CrossRef] [PubMed]
- Apte, M.V.; Pirola, R.C.; Wilson, J.S. Pancreatic stellate cells: a starring role in normal and diseased pancreas. Front. Physiol. 2012, 3, 344. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.-X.; Morahan, G. Pancreatic Stem Cells Remain Unresolved. Stem Cells Dev. 2014, 23, 2803–2812. [Google Scholar] [CrossRef]
- Phillips, P., Pancreatic stellate cells and fibrosis (Pancreatic Cancer and Tumor Microenvironment). Kerala, India: Transworld Research Network, 2012.—Р. 29. 53.
- Nielsen, M.F.B.; Mortensen, M.B.; Detlefsen, S. Identification of markers for quiescent pancreatic stellate cells in the normal human pancreas. Histochem. 2017, 148, 359–380. [Google Scholar] [CrossRef]
- Zha, M.; Li, F.; Xu, W.; Chen, B.; Sun, Z. Isolation and characterization of islet stellate cells in rat. Islets 2014, 6, e28701. [Google Scholar] [CrossRef]
- Zang, G.; Sandberg, M.; Carlsson, P.-O.; Welsh, N.; Jansson, L.; Barbu, A. Activated pancreatic stellate cells can impair pancreatic islet function in mice. Upsala J. Med Sci. 2015, 120, 169–180. [Google Scholar] [CrossRef]
- Lee, E.; Ryu, G.R.; Ko, S.-H.; Ahn, Y.-B.; Song, K.-H. A role of pancreatic stellate cells in islet fibrosis and β-cell dysfunction in type 2 diabetes mellitus. Biochem. Biophys. Res. Commun. 2017, 485, 328–334. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, B.; Li, W.; Zhou, J.; Gao, F.; Wang, X.; Cai, M.; Sun, Z. Pancreatic Stellate Cells: A Rising Translational Physiology Star as a Potential Stem Cell Type for Beta Cell Neogenesis. Front. Physiol. 2019, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Zulewski, H.; Abraham, E.J.; Gerlach, M.J.; Daniel, P.B.; Moritz, W.; Müller, B.; Vallejo, M.; Thomas, M.K.; Habener, J.F. Multipotential Nestin-Positive Stem Cells Isolated From Adult Pancreatic Islets Differentiate Ex Vivo Into Pancreatic Endocrine, Exocrine, and Hepatic Phenotypes. Diabetes 2001, 50, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.J.; et al. Insulinotropic Hormone Glucagon-Like Peptide-1 Differentiation of Human Pancreatic Islet-Derived Progenitor Cells into Insulin-Producing Cells. Endocrinology 2002, 143, 3152–3161. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, G.; A Hussain, M.; Holz, G.G. Glucagon-like peptide 1 stimulates insulin gene promoter activity by protein kinase A-independent activation of the rat insulin I gene cAMP response element. Diabetes 2000, 49, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Buteau, J.; Roduit, R.; Susini, S.; Prentki, M. Glucagon-like peptide-1 promotes DNA synthesis, activates phosphatidylinositol 3-kinase and increases transcription factor pancreatic and duodenal homeobox gene 1 (PDX-1) DNA binding activity in beta (INS-1)-cells. Diabetologia 1999, 42, 856–864. [Google Scholar] [CrossRef]
- Montrose-Rafizadeh, C.; et al. Pancreatic glucagon-like peptide-1 receptor couples to multiple G proteins and activates mitogen-activated protein kinase pathways in Chinese hamster ovary cells. Endocrinology 1999, 140, 1132–1140. [Google Scholar] [CrossRef]
- Nakamura, T.; Ito, T.; Uchida, M.; Hijioka, M.; Igarashi, H.; Oono, T.; Kato, M.; Nakamura, K.; Suzuki, K.; Jensen, R.T.; et al. PSCs and GLP-1R: occurrence in normal pancreas, acute/chronic pancreatitis and effect of their activation by a GLP-1R agonist. Lab. Investig. 2014, 94, 63–78. [Google Scholar] [CrossRef]
- Bhagwandin, V.J.; Shay, J.W. Pancreatic cancer stem cells: Fact or fiction? Biochimica et Biophysica Acta (BBA) Molecular Basis of Disease 2009, 1792, 248–259. [Google Scholar] [CrossRef]
- Hirschmann-Jax, C.; Foster, A. E.; Wulf, G. G.; Nuchtern, J. G.; Jax, T. W.; Gobel, U.; Goodell, M. A.; Brenner, M. K. A distinct "side population" of cells with high drug efflux capacity in human tumor cells. Proc. Natl. Acad. Sci. USA 2004, 101, 14228–14233. [Google Scholar] [CrossRef]
- Wang, Y.; Li, F.; Luo, B.; Wang, X.; Sun, H.; Liu, S.; Cui, Y.; Xu, X. A side population of cells from a human pancreatic carcinoma cell line harbors cancer stem cell characteristics. Neoplasma 2009, 56, 371–378. [Google Scholar] [CrossRef]
- Ishiwata, T.; Matsuda, Y.; Yoshimura, H.; Sasaki, N.; Ishiwata, S.; Ishikawa, N.; Takubo, K.; Arai, T.; Aida, J. Pancreatic cancer stem cells: features and detection methods. Pathol. Oncol. Res. 2018, 24, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Mato, E.; Lucas, M.; Petriz, J.; Gomis, R.; Novials, A. Identification of a pancreatic stellate cell population with properties of progenitor cells: new role for stellate cells in the pancreas. Biochem. J. 2009, 421, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Kordes, C.; Sawitza, I.; Götze, S.; Häussinger, D. Stellate Cells from Rat Pancreas Are Stem Cells and Can Contribute to Liver Regeneration. PLOS ONE 2012, 7, e51878. [Google Scholar] [CrossRef] [PubMed]
- Zha, M.; Xu, W.; Jones, P.M.; Sun, Z. Isolation and characterization of human islet stellate cells. Exp. Cell Res. 2016, 341, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Pang, T.C.Y.; Wilson, J.S.; Apte, M.V. Pancreatic stellate cells: what's new? Curr Opin Gastroenterol 2017, 33, 366–373. [Google Scholar] [CrossRef]
- Pacifico, F.; Leonardi, A.; Crescenzi, E. Glutamine Metabolism in Cancer Stem Cells: A Complex Liaison in the Tumor Microenvironment. Int. J. Mol. Sci. 2023, 24, 2337. [Google Scholar] [CrossRef]
- Joharatnam-Hogan, N.; Morganstein, D.L. Diabetes and cancer: Optimising glycaemic control. J. Hum. Nutr. Diet. 2022, 36, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Jernås, M.; Palming, J.; Sjöholm, K.; Jennische, E.; Svensson, P.-A.; Gabrielsson, B.G.; Levin, M.; Sjögren, A.; Rudemo, M.; Lystig, T.C.; et al. Separation of human adipocytes by size: hypertrophic fat cells display distinct gene expression. FASEB J. 2006, 20, 1540–1542. [Google Scholar] [CrossRef]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory Mechanisms in Obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, J.M.; Stern, J.H.; Scherer, P.E. The cell biology of fat expansion. J. Cell Biol. 2015, 208, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Coe, P.O.; Williams, S.R.; Morris, D.M.; Parkin, E.; Harvie, M.; Renehan, A.G.; O'Reilly, D.A. Development of MR quantified pancreatic fat deposition as a cancer risk biomarker. Pancreatology 2018, 18, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Rebours, V.; Gaujoux, S.; D'Assignies, G.; Sauvanet, A.; Ruszniewski, P.; Lévy, P.; Paradis, V.; Bedossa, P.; Couvelard, A. Obesity and Fatty Pancreatic Infiltration Are Risk Factors for Pancreatic Precancerous Lesions (PanIN). Clin. Cancer Res. 2015, 21, 3522–3528. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.G.; Nguyen, N.N.; Cervantes, A.; Alarcon Ramos, G.C.; Cho, J.; Petrov, M.S. Associations between intra-pancreatic fat deposition and circulating levels of cytokines. Cytokine 2019, 120, 107–114. [Google Scholar] [CrossRef]
- Takahashi, M.; Hori, M.; Ishigamori, R.; Mutoh, M.; Imai, T.; Nakagama, H. Fatty pancreas: A possible risk factor for pancreatic cancer in animals and humans. Cancer Sci. 2018, 109, 3013–3023. [Google Scholar] [CrossRef]
- Pollak, M. Insulin and insulin-like growth factor signalling in neoplasia. Nat. Rev. Cancer 2008, 8, 915–928. [Google Scholar] [CrossRef]
- Fogarty, A.W.; Glancy, C.; Jones, S.; A Lewis, S.; McKeever, T.M.; Britton, J.R. A prospective study of weight change and systemic inflammation over 9 y. Am. J. Clin. Nutr. 2008, 87, 30–35. [Google Scholar] [CrossRef]
- O’brien, K.D.; Brehm, B.J.; Seeley, R.J.; Bean, J.; Wener, M.H.; Daniels, S.; D’alessio, D.A. Diet-Induced Weight Loss Is Associated with Decreases in Plasma Serum Amyloid A and C-Reactive Protein Independent of Dietary Macronutrient Composition in Obese Subjects. J. Clin. Endocrinol. Metab. 2005, 90, 2244–2249. [Google Scholar] [CrossRef]
- Chang, H.-H.; Moro, A.; Takakura, K.; Su, H.-Y.; Mo, A.; Nakanishi, M.; Waldron, R.T.; French, S.W.; Dawson, D.W.; Hines, O.J.; et al. Incidence of pancreatic cancer is dramatically increased by a high fat, high calorie diet in KrasG12D mice. PLOS ONE 2017, 12, e0184455. [Google Scholar] [CrossRef]
- Li, Z.-J.; Dai, H.-Q.; Huang, X.-W.; Feng, J.; Deng, J.-H.; Wang, Z.-X.; Yang, X.-M.; Liu, Y.-J.; Wu, Y.; Chen, P.-H.; et al. Artesunate synergizes with sorafenib to induce ferroptosis in hepatocellular carcinoma. Acta Pharmacol. Sin. 2020, 42, 301–310. [Google Scholar] [CrossRef]
- Turchi, R.; et al. Frataxin deficiency induces lipid accumulation and affects thermogenesis in brown adipose tissue. Cell Death Dis 2020, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; et al. Diabetic Ferroptosis and Pancreatic Cancer: Foe or Friend? Antioxid Redox Signal 2022, 37, 1206–1221. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xiao, L.; Liu, L.; Ye, L.; Su, P.; Bi, E.; Wang, Q.; Yang, M.; Qian, J.; Yi, Q. CD36-mediated ferroptosis dampens intratumoral CD8+ T cell effector function and impairs their antitumor ability. Cell Metab. 2021, 33, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Al-Shukaili, A.; Al-Ghafri, S.; Al-Marhoobi, S.; Al-Abri, S.; Al-Lawati, J.; Al-Maskari, M. Analysis of Inflammatory Mediators in Type 2 Diabetes Patients. Int. J. Endocrinol. 2013, 2013, 976810. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Höppener, J.W.; Ahrén, B.; Lips, C.J. Islet Amyloid and Type 2 Diabetes Mellitus. New Engl. J. Med. 2000, 343, 411–419. [Google Scholar] [CrossRef]
- Permert, J.; Larsson, J.; Westermark, G.T.; Herrington, M.K.; Christmanson, L.; Pour, P.M.; Westermark, P.; Adrian, T.E. Islet Amyloid Polypeptide in Patients with Pancreatic Cancer and Diabetes. New Engl. J. Med. 1994, 330, 313–318. [Google Scholar] [CrossRef]
- Neuzillet, C.; de Gramont, A.; Tijeras-Raballand, A.; de Mestier, L.; Cros, J.; Faivre, S.; Raymond, E. Perspectives of TGF-β inhibition in pancreatic and hepatocellular carcinomas. Oncotarget 2013, 5, 78–94. [Google Scholar] [CrossRef]
- Longnecker, D.S.; Gorelick, F.; Thompson, E.D. Anatomy, histology, and fine structure of the pancreas. The pancreas: an integrated textbook of basic science, medicine, and surgery, 2018: p. 10-23.
- Pannala, R.; Basu, A.; Petersen, G.M.; Chari, S.T. New-onset diabetes: a potential clue to the early diagnosis of pancreatic cancer. Lancet Oncol. 2009, 10, 88–95. [Google Scholar] [CrossRef]
- Sah, R.P.; Nagpal, S.J.S.; Mukhopadhyay, D.; Chari, S.T. New insights into pancreatic cancer-induced paraneoplastic diabetes. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Perera, C.J.; Falasca, M.; Chari, S.T.; Greenfield, J.R.; Xu, Z.; Pirola, R.C.; Wilson, J.S.; Apte, M.V. Role of Pancreatic Stellate Cell-Derived Exosomes in Pancreatic Cancer-Related Diabetes: A Novel Hypothesis. Cancers 2021, 13, 5224. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, S.J.S.; Kandlakunta, H.; Sharma, A.; Sannapaneni, S.; Velamala, P.; Majumder, S.; Matveyenko, A.; Chari, S.T. Endocrinopathy in Pancreatic Cancer Is Characterized by Reduced Islet Size and Density with Preserved Endocrine Composition as Compared to Type 2 Diabetes: Presidential Poster Award. Am. J. Gastroenterol. 2018, 113, S26–S28. [Google Scholar] [CrossRef]
- Park, J.; Morley, T.S.; Kim, M.; Clegg, D.J.; Scherer, P.E. Obesity and cancer—mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 2014, 10, 455–465. [Google Scholar] [CrossRef]
- Haass, C.; Koo, E.H.; Mellon, A.; Hung, A.Y.; Selkoe, D.J. Targeting of cell-surface β-amyloid precursor protein to lysosomes: alternative processing into amyloid-bearing fragments. Nature 1992, 357, 500–503. [Google Scholar] [CrossRef]
- Cooper, G.J.; Willis, A.C.; Clark, A.; Turner, R.C.; Sim, R.B.; Reid, K.B. Purification and characterization of a peptide from amyloid-rich pancreases of type 2 diabetic patients. Proc. Natl. Acad. Sci. 1987, 84, 8628–8632. [Google Scholar] [CrossRef]
- Janson, J.; Laedtke, T.; Parisi, J.E.; O’brien, P.; Petersen, R.C.; Butler, P.C. Increased Risk of Type 2 Diabetes in Alzheimer Disease. Diabetes 2004, 53, 474–481. [Google Scholar] [CrossRef]
- Ben Hmidene, A.; Hanaki, M.; Murakami, K.; Irie, K.; Isoda, H.; Shigemori, H. Inhibitory Activities of Antioxidant Flavonoids from Tamarix gallica on Amyloid Aggregation Related to Alzheimer’s and Type 2 Diabetes Diseases. Biol. Pharm. Bull. 2017, 40, 238–241. [Google Scholar] [CrossRef]
- Sato, M.; Murakami, K.; Uno, M.; Nakagawa, Y.; Katayama, S.; Akagi, K.-I.; Masuda, Y.; Takegoshi, K.; Irie, K. Site-specific Inhibitory Mechanism for Amyloid β42 Aggregation by Catechol-type Flavonoids Targeting the Lys Residues. PEDIATRICS 2013, 288, 23212–23224. [Google Scholar] [CrossRef]
- Sato, M.; Murakami, K.; Uno, M.; Ikubo, H.; Nakagawa, Y.; Katayama, S.; Akagi, K.-I.; Irie, K. Structure–Activity Relationship for (+)-Taxifolin Isolated from Silymarin as an Inhibitor of Amyloid β Aggregation. Biosci. Biotechnol. Biochem. 2013, 77, 1100–1103. [Google Scholar] [CrossRef]
- KURISU, M.; et al. Inhibition of Amyloid β Aggregation by Acteoside, a Phenylethanoid Glycoside. Bioscience Biotechnology and Biochemistry 2013, 77, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Abhari, F.M. The role of plant-derived natural antioxidants in reduction of oxidative stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Luechapudiporn, R.; Phisalaphong, C.; Jirawatnotai, S. Curcumin Extract for Prevention of Type 2 Diabetes. Diabetes Care 2012, 35, 2121–2127. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Liao, Q.; Chen, X.; Peng, C.; Lin, L. The role of irisin in metabolic flexibility: Beyond adipose tissue browning. Drug Discov. Today 2022, 27, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Soto, A.; Chavez-Santoscoy, R.A.; Porras, O.; Hidalgo-Ledesma, M.; Serrano-Medina, A.; Ramírez-Rodríguez, A.A.; Castillo-Martinez, N.A. Epicatechin and quercetin exhibit in vitro antioxidant effect, improve biochemical parameters related to metabolic syndrome, and decrease cellular genotoxicity in humans. Food Res. Int. 2021, 142, 110101. [Google Scholar] [CrossRef] [PubMed]
- Derochette, S.; Franck, T.; Mouithys-Mickalad, A.; Ceusters, J.; Deby-Dupont, G.; Lejeune, J.-P.; Neven, P.; Serteyn, D. Curcumin and resveratrol act by different ways on NADPH oxidase activity and reactive oxygen species produced by equine neutrophils. Chem. Interactions 2013, 206, 186–193. [Google Scholar] [CrossRef]
- Yousefian, M.; Shakour, N.; Hosseinzadeh, H.; Hayes, A.W.; Hadizadeh, F.; Karimi, G. The natural phenolic compounds as modulators of NADPH oxidases in hypertension. Phytomedicine 2018, 55, 200–213. [Google Scholar] [CrossRef]
- Jaishree, V.; Narsimha, S. Swertiamarin and quercetin combination ameliorates hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced type 2 diabetes mellitus in wistar rats. BioMedicine 2020, 130, 110561. [Google Scholar] [CrossRef]
- Evans, J.M.; et al. Metformin and reduced risk of cancer in diabetic patients. Bmj 2005, 330, 1304–1305. [Google Scholar] [CrossRef]
- Li, D.; Yeung, S.J.; Hassan, M.M.; Konopleva, M.; Abbruzzese, J.L. Antidiabetic Therapies Affect Risk of Pancreatic Cancer. Gastroenterology 2009, 137, 482–488. [Google Scholar] [CrossRef]
- Hartogh, D.J.D.; Tsiani, E. Antidiabetic Properties of Naringenin: A Citrus Fruit Polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.A.; Law, R.J.; Frank, R.D.; Bamlet, W.R.; A Burch, P.; Petersen, G.M.; Rabe, K.G.; Chari, S.T. Impact of Diabetes Mellitus on Clinical Outcomes in Patients Undergoing Surgical Resection for Pancreatic Cancer: A Retrospective, Cohort Study. Am. J. Gastroenterol. 2014, 109, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Chari, S.T. Pancreatic Cancer and Diabetes Mellitus. Curr. Treat. Options Gastroenterol. 2018, 16, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.P.; et al. Early detection of pancreatic cancer. Lancet Gastroenterol Hepatol 2020, 5, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Neoptolemos, J.P.; Kleeff, J.; Michl, P.; Costello, E.; Greenhalf, W.; Palmer, D.H. Therapeutic developments in pancreatic cancer: Current and future perspectives. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 333–348. [Google Scholar] [CrossRef]
- Murphy, J.E.; et al. Total Neoadjuvant Therapy With FOLFIRINOX Followed by Individualized Chemoradiotherapy for Borderline Resectable Pancreatic Adenocarcinoma: A Phase 2 Clinical Trial. JAMA Oncol 2018, 4, 963–969. [Google Scholar] [CrossRef]
- Bhat, K.; Wang, F.; Ma, Q.; Li, Q.; Mallik, S.; Hsieh, T.-C.; Wu, E. Advances in Biomarker Research for Pancreatic Cancer. Curr. Pharm. Des. 2012, 18, 2439–2451. [Google Scholar] [CrossRef]
- Lumlerdkij, N.; Tantiwongse, J.; Booranasubkajorn, S.; Boonrak, R.; Akarasereenont, P.; Laohapand, T.; Heinrich, M. Understanding cancer and its treatment in Thai traditional medicine: An ethnopharmacological-anthropological investigation. J. Ethnopharmacol. 2018, 216, 259–273. [Google Scholar] [CrossRef]
- Masek, A.; Chrzescijanska, E.; Latos, M.; Zaborski, M. Influence of hydroxyl substitution on flavanone antioxidants properties. Food Chem. 2017, 215, 501–507. [Google Scholar] [CrossRef]
- Amić, A.; Marković, Z.; Klein, E.; Marković, J.M.D.; Milenković, D. Theoretical study of the thermodynamics of the mechanisms underlying antiradical activity of cinnamic acid derivatives. Food Chem. 2018, 246, 481–489. [Google Scholar] [CrossRef]
- Park, M.N.; Rahman, A.; Rahman, H.; Kim, J.W.; Choi, M.; Kim, J.W.; Choi, J.; Moon, M.; Ahmed, K.R.; Kim, B. Potential Therapeutic Implication of Herbal Medicine in Mitochondria-Mediated Oxidative Stress-Related Liver Diseases. Antioxidants 2022, 11, 2041. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, C.; Haenen, G.; van Acker, F.; van der Vijgh, W.; Bast, A. Flavonoids as peroxynitrite scavengers: the role of the hydroxyl groups. Toxicol. Vitr. 2001, 15, 3–6. [Google Scholar] [CrossRef]
- Van Acker, S.A.B.E.; Van Den Berg, D.-J.; Tromp, M.N.J.L.; Griffioen, D.H.; Van Bennekom, W.P.; Van Der Vijgh, W.J.F.; Bast, A. Structural aspects of antioxidant activity of flavonoids. Free Radic. Biol. Med. 1996, 20, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, M.; Andruniów, T.; Sroka, Z. Flavones' and Flavonols' Antiradical Structure-Activity Relationship-A Quantum Chemical Study. Antioxidants 2020, 9. [Google Scholar] [CrossRef]
- Heijnen, C.G.; Haenen, G.R.; Vekemans, J.A.; Bast, A. Peroxynitrite scavenging of flavonoids: structure activity relationship. Environ. Toxicol. Pharmacol. 2001, 10, 199–206. [Google Scholar] [CrossRef]
- Park, H.J.; Choi, Y.J.; Lee, J.H.; Nam, M.J. Naringenin causes ASK1-induced apoptosis via reactive oxygen species in human pancreatic cancer cells. Food Chem. Toxicol. 2017, 99, 1–8. [Google Scholar] [CrossRef]
- Lou, C.; Zhang, F.; Yang, M.; Zhao, J.; Zeng, W.; Fang, X.; Zhang, Y.; Zhang, C.; Liang, W. Naringenin Decreases Invasiveness and Metastasis by Inhibiting TGF-β-Induced Epithelial to Mesenchymal Transition in Pancreatic Cancer Cells. PLOS ONE 2012, 7, e50956. [Google Scholar] [CrossRef]
- Lee, J.; Kim, D.-H.; Kim, J.H. Combined administration of naringenin and hesperetin with optimal ratio maximizes the anti-cancer effect in human pancreatic cancer via down regulation of FAK and p38 signaling pathway. Phytomedicine 2018, 58, 152762. [Google Scholar] [CrossRef]
- Huang, J.; Guo, W.; Cheung, F.; Tan, H.-Y.; Wang, N.; Feng, Y. Integrating Network Pharmacology and Experimental Models to Investigate the Efficacy of Coptidis and Scutellaria Containing Huanglian Jiedu Decoction on Hepatocellular Carcinoma. Am. J. Chin. Med. 2020, 48, 161–182. [Google Scholar] [CrossRef]
- Li, S.; Zhang, B. Traditional Chinese medicine network pharmacology: theory, methodology and application. Chinese Journal of Natural Medicines 2013, 11, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Network target: a starting point for traditional Chinese medicine network pharmacology. Zhongguo Zhong yao za zhi= Zhongguo zhongyao zazhi= China journal of Chinese materia medica 2011, 36, 2017–2020. [Google Scholar] [PubMed]
- Qian, X.; Chen, Z.; Chen, S.S.; Liu, L.M.; Zhang, A.Q. Integrated Analyses Identify Immune-Related Signature Associated with Qingyihuaji Formula for Treatment of Pancreatic Ductal Adenocarcinoma Using Network Pharmacology and Weighted Gene Co-Expression Network. J. Immunol. Res. 2020, 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.-W.; Xu, P.-L.; Cheng, C.-S.; Jiao, J.-Y.; Wu, Y.; Dong, S.; Xie, J.; Zhu, X.-Y. Integrating network pharmacology and experimental models to investigate the efficacy of QYHJ on pancreatic cancer. J. Ethnopharmacol. 2022, 297, 115516. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-K.; Cheng, K.-C.; Mgbeahuruike, M.O.; Lin, Y.-H.; Wu, C.-Y.; Wang, H.-M.D.; Yen, C.-H.; Chiu, C.-C.; Sheu, S.-J. New Insight into the Effects of Metformin on Diabetic Retinopathy, Aging and Cancer: Nonapoptotic Cell Death, Immunosuppression, and Effects beyond the AMPK Pathway. Int. J. Mol. Sci. 2021, 22, 9453. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Cheng, C.-S.; Xu, P.; Yang, P.; Zhang, K.; Jing, Y.; Chen, Z. Mechanisms of pancreatic tumor suppression mediated by Xiang-lian pill: An integrated in silico exploration and experimental validation. J. Ethnopharmacol. 2022, 298, 115586. [Google Scholar] [CrossRef] [PubMed]
- An, E.-J.; Kim, Y.; Lee, S.-H.; Ko, H.M.; Chung, W.-S.; Jang, H.-J. Anti-Cancer Potential of Oxialis obtriangulata in Pancreatic Cancer Cell through Regulation of the ERK/Src/STAT3-Mediated Pathway. Molecules 2020, 25, 2301. [Google Scholar] [CrossRef]
- Relles, D.; et al. Thymoquinone promotes pancreatic cancer cell death and reduction of tumor size through combined inhibition of histone deacetylation and induction of histone acetylation. Advances in preventive medicine 2016, 2016. [Google Scholar] [CrossRef]
- Mu, H.; et al. Role of NF-κB in the anti-tumor effect of thymoquinone on bladder cancer. Zhonghua Yi Xue Za Zhi 2012, 92, 392–396. [Google Scholar]
- Muralidharan-Chari, V.; Kim, J.; Abuawad, A.; Naeem, M.; Cui, H.; Mousa, S.A. Thymoquinone Modulates Blood Coagulation in Vitro via Its Effects on Inflammatory and Coagulation Pathways. Int. J. Mol. Sci. 2016, 17, 474. [Google Scholar] [CrossRef]
- Yusufi, M.; Banerjee, S.; Mohammad, M.; Khatal, S.; Swamy, K.V.; Khan, E.M.; Aboukameel, A.; Sarkar, F.H.; Padhye, S. Synthesis, characterization and anti-tumor activity of novel thymoquinone analogs against pancreatic cancer. Bioorganic Med. Chem. Lett. 2013, 23, 3101–3104. [Google Scholar] [CrossRef] [PubMed]
- Banik, K.; Khatoon, E.; Harsha, C.; Rana, V.; Parama, D.; Thakur, K.K.; Bishayee, A.; Kunnumakkara, A.B. Wogonin and its analogs for the prevention and treatment of cancer: A systematic review. Phytotherapy Res. 2022, 36, 1854–1883. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Polier, G.; Köhler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and Related Natural Flavones Overcome Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL) Protein Resistance of Tumors by Down-regulation of c-FLIP Protein and Up-regulation of TRAIL Receptor 2 Expression. J. Biol. Chem. 2012, 287, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Polier, G.; Ding, J.; Konkimalla, B.V.; Eick, D.; Ribeiro, N.; Köhler, R.; Giaisi, M.; Efferth, T.; Desaubry, L.; Krammer, P.H.; et al. Wogonin and related natural flavones are inhibitors of CDK9 that induce apoptosis in cancer cells by transcriptional suppression of Mcl-1. Cell Death Dis. 2011, 2, e182. [Google Scholar] [CrossRef] [PubMed]
- Vlavcheski, F.; O’neill, E.J.; Gagacev, F.; Tsiani, E. Effects of Berberine against Pancreatitis and Pancreatic Cancer. Molecules 2022, 27, 8630. [Google Scholar] [CrossRef] [PubMed]
- Kallifatidis, G.; Hoy, J.J.; Lokeshwar, B.L. Bioactive natural products for chemoprevention and treatment of castration-resistant prostate cancer. Semin. Cancer Biol. 2016, 40–41, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Guo, Z.; Zhu, P.; Chen, J.; Huang, Y. Traditional Chinese medicine as a cancer treatment: Modern perspectives of ancient but advanced science. Cancer Med. 2019, 8, 1958–1975. [Google Scholar] [CrossRef]
- Kim, A.; Ha, J.; Kim, J.; Cho, Y.; Ahn, J.; Cheon, C.; Kim, S.-H.; Ko, S.-G.; Kim, B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients 2021, 13, 3801. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Tan, H.; Wang, N.; Chen, L.; Meng, Z.; Chen, Z.; Feng, Y. Functional inhibition of lactate dehydrogenase suppresses pancreatic adenocarcinoma progression. Clin. Transl. Med. 2021, 11, e467. [Google Scholar] [CrossRef]
- Rotow, J.; Bivona, T.G. Understanding and targeting resistance mechanisms in NSCLC. Nat. Rev. Cancer 2017, 17, 637–658. [Google Scholar] [CrossRef]
- Park, J.W.; Han, J.-W. Targeting epigenetics for cancer therapy. Arch. Pharmacal Res. 2019, 42, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Staupe, H.; Buentzel, J.; Keinki, C.; Buentzel, J.; Huebner, J. Systematic analysis of mistletoe prescriptions in clinical studies. J. Cancer Res. Clin. Oncol. 2022, 149, 5559–5571. [Google Scholar] [CrossRef] [PubMed]
- Matthes, H.; Friedel, W.; Bock, P.; Zanker, K. Molecular Mistletoe Therapy: Friend or Foe in Established Anti-Tumor Protocols? A Multicenter, Controlled, Retrospective Pharmaco-Epidemiological Study in Pancreas Cancer. Curr. Mol. Med. 2010, 10, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Friedel, W.E.; Matthes, H.; Bock, P.R.; Zänker, K.S. Systematic evaluation of the clinical effects of supportive mistletoe treatment within chemo- and/or radiotherapy protocols and long-term mistletoe application in nonmetastatic colorectal carcinoma: multicenter, controlled, observational cohort study. J. Soc. Integr. Oncol. 2009, 7, 137–145. [Google Scholar] [PubMed]
- Schad, F.; et al. Intratumoral Mistletoe (Viscum album L) Therapy in Patients with Unresectable Pancreas Carcinoma: A Retrospective Analysis. Integr Cancer Ther 2014, 13, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Epelbaum, R.; Schaffer, M.; Vizel, B.; Badmaev, V.; Bar-Sela, G. Curcumin and Gemcitabine in Patients With Advanced Pancreatic Cancer. Nutr. Cancer 2010, 62, 1137–1141. [Google Scholar] [CrossRef]
- Panahi, Y.; Saadat, A.; Beiraghdar, F.; Nouzari, S.M.H.; Jalalian, H.R.; Sahebkar, A. Antioxidant effects of bioavailability-enhanced curcuminoids in patients with solid tumors: A randomized double-blind placebo-controlled trial. J. Funct. Foods 2014, 6, 615–622. [Google Scholar] [CrossRef]
- Fuchs-Tarlovsky, V. Role of antioxidants in cancer therapy. Nutrition 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Block, K.I.; Koch, A.C.; Mead, M.N.; Tothy, P.K.; Newman, R.A.; Gyllenhaal, C. Impact of antioxidant supplementation on chemotherapeutic efficacy: A systematic review of the evidence from randomized controlled trials. Cancer Treat. Rev. 2007, 33, 407–418. [Google Scholar] [CrossRef]
- Royal, R.E.; Levy, C.; Turner, K.; Mathur, A.; Hughes, M.; Kammula, U.S.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Lowy, I.; et al. Phase 2 Trial of Single Agent Ipilimumab (Anti-CTLA-4) for Locally Advanced or Metastatic Pancreatic Adenocarcinoma. J. Immunother. 2010, 33, 828–833. [Google Scholar] [CrossRef]
- Yarchoan, M.; Hopkins, A.; Jaffee, E.M. Tumor Mutational Burden and Response Rate to PD-1 Inhibition. N. Engl. J. Med. 2017, 377, 2500–2501. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Zhou, X.; Liu, Y.; Das, A.; Vincent, B.G.; Li, J.; Liu, R.; Huang, L. Tumor neoantigen heterogeneity impacts bystander immune inhibition of pancreatic cancer growth. Transl. Oncol. 2020, 13, 100856. [Google Scholar] [CrossRef] [PubMed]
- Rojas, L.A.; Sethna, Z.; Soares, K.C.; Olcese, C.; Pang, N.; Patterson, E.; Lihm, J.; Ceglia, N.; Guasp, P.; Chu, A.; et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature 2023, 618, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-J.; Tao, Z.; Gu, W.; Sun, L.-H. Variation of Blood T Lymphocyte Subgroups in Patients with Non- small Cell Lung Cancer. Asian Pac. J. Cancer Prev. 2013, 14, 4671–4673. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-W.; Min, H.-Y. Ginseng, the 'Immunity Boost': The Effects of Panax ginseng on Immune System. J. Ginseng Res. 2012, 36, 354–368. [Google Scholar] [CrossRef]
- Hwang, I.; Ahn, G.; Park, E.; Ha, D.; Song, J.-Y.; Jee, Y. An acidic polysaccharide of Panax ginseng ameliorates experimental autoimmune encephalomyelitis and induces regulatory T cells. Immunol. Lett. 2011, 138, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-K.; Lee, K.Y.; Kang, J.; Park, J.S.; Jeong, J. Immune-modulating Effect of Korean Red Ginseng by Balancing the Ratio of Peripheral T Lymphocytes in Bile Duct or Pancreatic Cancer Patients With Adjuvant Chemotherapy. Vivo 2021, 35, 1895–1900. [Google Scholar] [CrossRef] [PubMed]
- Sohal, D.P.S.; Kennedy, E.B.; Khorana, A.; Copur, M.S.; Crane, C.H.; Garrido-Laguna, I.; Krishnamurthi, S.; Moravek, C.; O’Reilly, E.M.; Philip, P.A.; et al. Metastatic Pancreatic Cancer: ASCO Clinical Practice Guideline Update. J. Clin. Oncol. 2018, 36, 2545–2556. [Google Scholar] [CrossRef]
- Sohal, D.P.; Mangu, P.B.; Khorana, A.A.; Shah, M.A.; Philip, P.A.; O’reilly, E.M.; Uronis, H.E.; Ramanathan, R.K.; Crane, C.H.; Engebretson, A.; et al. Metastatic Pancreatic Cancer: American Society of Clinical Oncology Clinical Practice Guideline. J. Clin. Oncol. 2016, 34, 2784–2796. [Google Scholar] [CrossRef]
- Shojania, K.G.; Sampson, M.; Ansari, M.T.; Ji, J.; Doucette, S.; Moher, D. How Quickly Do Systematic Reviews Go Out of Date? A Survival Analysis. Ann. Intern. Med. 2007, 147, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.H.; A Tempero, M.; Shan, Y.-S.; Su, W.-C.; Lin, Y.-L.; Dito, E.; Ong, A.; Wang, Y.-W.; Yeh, C.G.; Chen, L.-T. A multinational phase 2 study of nanoliposomal irinotecan sucrosofate (PEP02, MM-398) for patients with gemcitabine-refractory metastatic pancreatic cancer. Br. J. Cancer 2013, 109, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N Engl J Med 2011, 364, 1817–1825. [Google Scholar] [CrossRef]
- Schrag, D.; Archer, L.; Wang, X.; Romanus, D.; Mulcahy, M.; Goldberg, R.; Kindler, H. A patterns-of-care study of post-progression treatment (Rx) among patients (pts) with advanced pancreas cancer (APC) after gemcitabine therapy on Cancer and Leukemia Group B (CALGB) study #80303. J. Clin. Oncol. 2007, 25, 4524. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Shen, M.; Xu, M.-D.; Yu, Z.-Y.; Tao, M. FOLFIRINOX regulated tumor immune microenvironment to extend the survival of patients with resectable pancreatic ductal adenocarcinoma. Gland. Surg. 2020, 9, 2125–2135. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Wu, W.; Wang, B.; Cui, C.; Niu, J.; Chen, J.; Chen, Z.; Liu, Y. CXCL5 Plays a Promoting Role in Osteosarcoma Cell Migration and Invasion in Autocrine- and Paracrine-Dependent Manners. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2017, 25, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Bonavita, O.; Massara, M.; Bonecchi, R. Chemokine regulation of neutrophil function in tumors. Cytokine Growth Factor Rev. 2016, 30, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Klaiber, U.; Hackert, T.; Neoptolemos, J.P. Adjuvant treatment for pancreatic cancer. Transl. Gastroenterol. Hepatol. 2019, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Dahan, L.; Phelip, J.M.; Le Malicot, K.; Williet, N.; Desrame, J.; Volet, J.; Petorin, C.; Malka, D.; Rebischung, C.; Aparicio, T.; et al. FOLFIRINOX until progression, FOLFIRINOX with maintenance treatment, or sequential treatment with gemcitabine and FOLFIRI.3 for first-line treatment of metastatic pancreatic cancer: A randomized phase II trial (PRODIGE 35-PANOPTIMOX). J. Clin. Oncol. 2018, 36, 4000. [Google Scholar] [CrossRef]
- Kamisawa, T.; et al. Pancreatic cancer. Lancet 2016, 388, 73–85. [Google Scholar] [CrossRef]
- Moore, M.J.; Goldstein, D.; Hamm, J.; Figer, A.; Hecht, J.R.; Gallinger, S.; Au, H.J.; Murawa, P.; Walde, D.; Wolff, R.A.; et al. Erlotinib Plus Gemcitabine Compared With Gemcitabine Alone in Patients With Advanced Pancreatic Cancer: A Phase III Trial of the National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol. 2007, 25, 1960–1966. [Google Scholar] [CrossRef]
- Golan, T.; Hammel, P.; Reni, M.; Van Cutsem, E.; Macarulla, T.; Hall, M.J.; Park, J.-O.; Hochhauser, D.; Arnold, D.; Oh, D.-Y.; et al. Maintenance Olaparib for Germline BRCA-Mutated Metastatic Pancreatic Cancer. N. Engl. J. Med. 2019, 381, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Marabelle, A.; Le, D.T.; Ascierto, P.A.; Di Giacomo, A.M.; De Jesus-Acosta, A.; Delord, J.-P.; Geva, R.; Gottfried, M.; Penel, N.; Hansen, A.R.; et al. Efficacy of Pembrolizumab in Patients With Noncolorectal High Microsatellite Instability/Mismatch Repair–Deficient Cancer: Results From the Phase II KEYNOTE-158 Study. J. Clin. Oncol. 2020, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Durham, J.N.; Smith, K.N.; Wang, H.; Bartlett, B.R.; Aulakh, L.K.; Lu, S.; Kemberling, H.; Wilt, C.; Luber, B.S.; et al. Mismatch repair deficiency predicts response of solid tumors to PD-1 blockade. Science 2017, 357, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Ott, P.A.; Bang, Y.-J.; Piha-Paul, S.A.; Razak, A.R.A.; Bennouna, J.; Soria, J.-C.; Rugo, H.S.; Cohen, R.B.; O’Neil, B.H.; Mehnert, J.M.; et al. T-Cell–Inflamed Gene-Expression Profile, Programmed Death Ligand 1 Expression, and Tumor Mutational Burden Predict Efficacy in Patients Treated With Pembrolizumab Across 20 Cancers: KEYNOTE-028. J. Clin. Oncol. 2019, 37, 318–327. [Google Scholar] [CrossRef]
- Karra, P.; Winn, M.; Pauleck, S.; Bulsiewicz-Jacobsen, A.; Peterson, L.; Coletta, A.; Doherty, J.; Ulrich, C.M.; Summers, S.A.; Gunter, M.; et al. Metabolic dysfunction and obesity-related cancer: Beyond obesity and metabolic syndrome. Obesity 2022, 30, 1323–1334. [Google Scholar] [CrossRef]
- Sharma, A.; Kandlakunta, H.; Nagpal, S.J.S.; Feng, Z.; Hoos, W.; Petersen, G.M.; Chari, S.T. Model to Determine Risk of Pancreatic Cancer in Patients With New-Onset Diabetes. Gastroenterology 2018, 155, 730–739. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PSCs engage in intercellular communication with PCCs to cause Wnt activation. The Wnt signaling pathway engages in direct interaction with the β-catenin/TCF7 complex, resulting in the upregulation of GS expression within the promoter region. Glutamine metabolism has been observed to exert inhibitory effects on the processes of self-renewal and the expression of stemness/pluripotent genes [37]. The promotion of oncogenic Akt signaling is facilitated by the release of IGF2 through the action of glutamine. The production of IGF2 by beta cells is regulated by nutrients, which in turn activates the autocrine loop involving IGF2 and IGF1R. This loop plays a crucial role in controlling both the bulk and function of the β cells [43]. The activation of PSCs induces a cellular transformation that resembles the characteristics of myofibroblasts. This transformation is characterized by the production of α-smooth muscle actin (α-SMA), which serves as a protein marker for cellular activation. The process of activation serves to decrease the presence of fat droplets that contain retinoids. PSCs are of utmost importance in the intricate process of islet fibrosis formation, ultimately resulting in the impairment of β-cell functionality [48].
Figure 1.
PSCs engage in intercellular communication with PCCs to cause Wnt activation. The Wnt signaling pathway engages in direct interaction with the β-catenin/TCF7 complex, resulting in the upregulation of GS expression within the promoter region. Glutamine metabolism has been observed to exert inhibitory effects on the processes of self-renewal and the expression of stemness/pluripotent genes [37]. The promotion of oncogenic Akt signaling is facilitated by the release of IGF2 through the action of glutamine. The production of IGF2 by beta cells is regulated by nutrients, which in turn activates the autocrine loop involving IGF2 and IGF1R. This loop plays a crucial role in controlling both the bulk and function of the β cells [43]. The activation of PSCs induces a cellular transformation that resembles the characteristics of myofibroblasts. This transformation is characterized by the production of α-smooth muscle actin (α-SMA), which serves as a protein marker for cellular activation. The process of activation serves to decrease the presence of fat droplets that contain retinoids. PSCs are of utmost importance in the intricate process of islet fibrosis formation, ultimately resulting in the impairment of β-cell functionality [48].
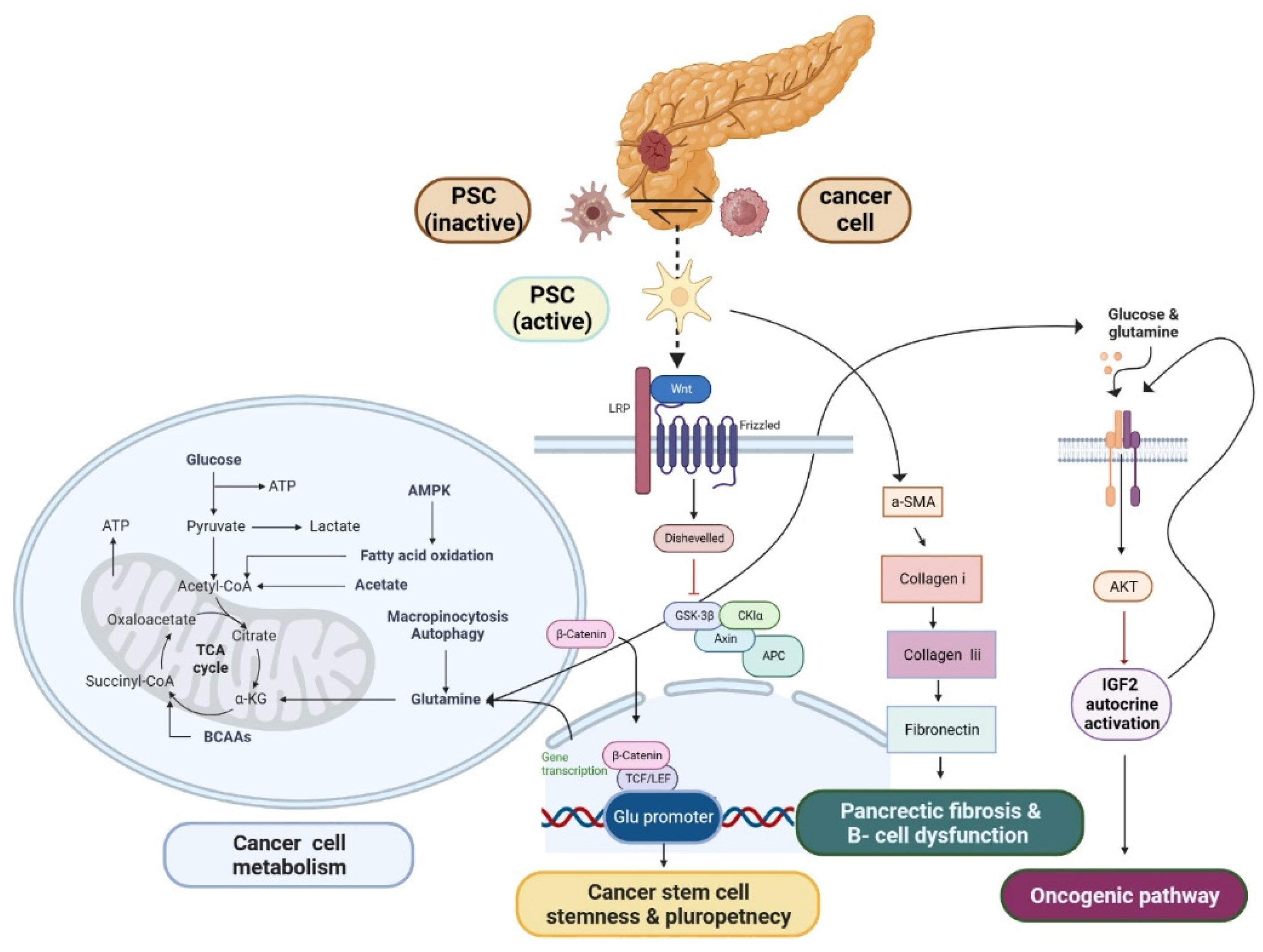

Table 1.
The investigation of the potential anti-type 2 diabetic mellitus (T2DM) properties exhibited by natural compounds.
Table 1.
The investigation of the potential anti-type 2 diabetic mellitus (T2DM) properties exhibited by natural compounds.
| Compound | Mechanism | Experimental model | Reference |
|---|---|---|---|
| Epicatechin and Quercetin (1:1 combination) | reductions in total cholesterol, LDL-cholesterol, total triglycerides, and fasting plasma glucose | randomized placebo-controlled trial | [108] |
| Curcumin | nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, which leads to an increase in the activity of antioxidant enzymes | healthy horses (mixed breeds, mean age 6.2 ± 2.3) | [109,110] |
| Combination of swertiamarin and quercetin (CSQ) | activation of viable β-cells, reducing levels of total cholesterol, triglycerides, and LDL, while simultaneously increasing HDL | diabetic rats | [111] |
| Metformin | insulin or sulfonylureas | patients | [112,113] |
| Naringenin | upregulation of adenosine monophosphate-activated protein kinase (AMPK), a key enzyme involved in cellular energy regulation. | [114] |
Table 2.
The in vitro investigation of the anti-cancer effect of natural products.
| Compound | Mechanism | Experimental model | Reference |
|---|---|---|---|
| Naringenin | Increased Activity of ASK1, P38, P53, JNK, and Reactive Oxygen Species Gemcitabine's ability to prevent cancer drug resistance also limits tumor cell invasion. | SNU-213 cells AsPC-1, PANC-1 |
[130] [131] |
| Combination with naringenin | Effects that suppress cell proliferation, invasion, and p38 signaling | MIA PaCa-2, PANC-1, SNU-213, BALB/c nude mice |
[132]. |
| Metformin | Increased levels of manganese superoxide dismutase (MnSOD) and decreased levels of NADPH oxidase 2 and 4 | MiaPaCa-2 and PANC-1 | [138] |
| QYHJ | Keap1/Nrf2/HO-1/NQO1 axis, p-PI3K/p-AKT/p-mTOR, and p-AKT/mTOR inhibition | PANC-1, MIA PaCa-2 | [137] |
| XLP | A reduction in the activity of MMP2, PTGS2, CASP9, IL4, and CTSD | Mouse PC Panc-02 cells Transfection pCDNA3.1(+)-PTGS2 and pCDNA3.1(+)-NC, Male C57BL/6 mice, 4–6 weeks old | [139,152] |
| Oxalidaceae | ERK/Src/STAT3 | BxPC3 | [140] |
| Thymoquinone | Increased p21 and p53; downregulation of Bcl-xL, Bcl-2, XIAP, Notch1, NICD, PTEN, Akt/mTOR/S6 Signaling; upregulation of caspases-3, -9, Bax, and cytochrome c; G2 cycle arrest and Sub G0/G1 arrest; | MiaPaCa-2, AsPC- 1 | [141,142,143,144]. |
| WOG | p53 expression was stimulated, and the Beclin-1/PI3K, Akt/ULK1/4E-BP1/CYLD, and mTOR pathways were all inhibited. | PANC-1, Colo-357, HPCCs4, Capan-1, Colo-357 |
[146] |
| WOG | Suppressing Mcl-1, CDK-9, c-FLIP, and MDM2 | Capan-1, Colo-357 | [146,147] |
| BBR | Release of cytochrome c and caspase 7 | BxPC-3, HPDE-E6E7c7 cells | [148] |
Table 3.
Investigating the effectiveness of natural products against PDAC in the clinic.
| Compound | Efficacy | Reference |
| Mistletoe (Viscum album L) |
Improve survival rates from the current 2.7 months to 4.8, | [156,158] |
| QYHJ | Improved 1-, 3-, and 5-year survival with no obvious side effects | [139] |
| Combination of Curcuminoids and Gemcitabine | Curcumin increases chemotherapy's efficacy | [159] |
| Curcuminoids | Health benefits, including fewer unwanted consequences, are increasing | [160,161,162] |
| Korean Red ginseng | Elevated CD4+ lymphocytes and a higher CD4+/CD8+ T lymphocyte counts | [168,169] [170] |
Table 4.
An Investigation on the Potential of Drug as a Therapeutic Intervention for PDAC.
| Drug | Target | Characterization | Reference |
|---|---|---|---|
| FOLFIRINOX; Leucovorin (LV), 5-Fluorouracil (5-FU), Irinotecan, and Oxaliplatin | C-X-C motif chemokine 5 (CXCL5) | Influence on tumor immunity regulation | [177]. |
| Olaparib | BRCA1/2 mutations | [184]. | |
| Afatinib and MCLA-128 | PDAC drivers, which are characterized by the presence of wild-type KRAS, include ERBB inhibitors. | ERBB mutations | [12]. |
| larotrectinib and Entrectinib | PDAC drivers include TRK inhibitors, which are activated by the presence of wild-type KRAS. | TRK mutations | [12]. |
| Crizotinib | PDAC drivers with wild-type KRAS include sensitivity to ALK/ROS inhibitors. | ALK/ROS mutations | [12]. |
| AMG510 | KRASG12C mutation | [12]. | |
| Pembrolizumab | Programmed cell death protein (PD-1) inhibitor | Microsatellite instability-high (MSI-H) | [171,185]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



