
Submitted:
27 August 2023
Posted:
29 August 2023
You are already at the latest version
Abstract
This review delves into the significance of chemical communication in mammals, with an emphasis on the role of the vomeronasal system in sensing pheromone-mediated interactions. The vomeronasal system shapes a variety of social and sexual behaviors, including reproduction, hierarchy formation, maternal bonding, and species recognition. Intriguingly, this system presents a vast degree of evolutionary variability, more so than variations observed in the olfactory system. This adaptive diversity underscores the challenges in studying the vomeronasal system, emphasizing the need for detailed research into its neuroanatomy and functional morphology. This review traverses an array of topics central to our understanding of chemical communication in mammals. We begin with the foundational role and basic concepts of chemocommunication. This is followed by an in-depth exploration of the olfactory systems. Subsequently, the neuroanatomical underpinnings of the vomeronasal system are juxtaposed against those of the olfactory system. Our focus then shifts to an exhaustive study of the sensory element of the vomeronasal system, the vomeronasal organ, and the integrating center of this information, the accessory olfactory bulb. The discussion extends to secondary projections of both the olfactory and vomeronasal systems in the light of the dual olfactory hypothesis. The review culminates with a detailed look at four specific models of vomeronasal system organization belonging to four evolutionarily distinct mammalian families: rodents, marsupials, herpestids, and bovids. These have been meticulously analyzed by the authors, aiming to spotlight the significant morphofunctional differences stemming from the adaptive changes imprinted on the system.
Keywords:
chemical communication
; olfactory systems
; vomeronasal organ
; accessory olfactory bulb
; dual olfactory hypothesis
; mammalian evolution
1. Introduction
1.1. Chemical Communication in Mammals
Throughout evolution, living organisms have developed intricate systems of chemical communication aimed at interacting with the external environment and enhancing their survival rate [1]. In doing so, they are able to detect a wide array of chemical signals and convert them into sensory information [2]. When these chemical cues are exchanged between different individuals, the substances conveying these messages are termed semiochemicals [3]. These semiochemicals are released through urine, feces, saliva, and secretions from various skin glands, often being deposited in the environment through highly stereotyped behaviors [4]. This form of chemical communication holds a pivotal role, influencing key areas such as social behavior, reproduction, food-seeking, and evasion from potential predators [5]. This is in part because the chemical detection systems project to the limbic system, where processes such as emotions, memory, hunger, sexual instincts, and overall behavior are regulated [6]. Additionally, these semiochemicals operate in tandem with other signals perceived by physical senses (auditory, visual, or tactile), conveying information about species, gender, social status, and physiological and developmental state [7].
Chemical senses emerged in the earliest stages of evolution and are now present in all living organisms, from bacteria to more complex life forms. Hence, all animals are pre-adapted to perceive a variety of chemical signals [8]. However, primitive organisms detect stimuli through specialized cells, while vertebrates have integrated these receptor cells into highly sophisticated organs. If the perception of a specific chemical stimulus enhances survival or reproductive success, the adaptation aims to reduce the sensitivity threshold to this substance and/or increase the expression of genes responsible for its perception [9]. Yet, identifying such a diverse plethora of molecules (essential lipids, lipoproteins, proteins, steroids, alcohols, etc.), spanning a broad range of volatilities and solubilities, demands a vast array of chemoreceptors. Among these, olfactory receptors stand out, phylogenetically present in the olfactory rosette of fish [10] and being highly conserved and expanded in mammals [11]. In fact, the largest superfamily of mammalian genes corresponds to the olfactory receptors. They detect a complex web of odor mixtures, allowing the creation of a dynamic three-dimensional image of the surrounding world over time [12]. Depending on the physicochemical characteristics of the detected signals, such as volatility or spatial dispersion, the captured information will multiply [13].
The olfactory system, recognized as one of the most prominent chemical communication systems alongside taste, is one of the first senses to activate, even allowing for prenatal olfactory learning [14]. However, in mammals, odorous chemical signals have the capability to stimulate various chemosensory structures, extending beyond the olfactory organ: such as the vomeronasal organ, Masera's septal organ, Grüneberg's ganglion, and free nerve endings of the trigeminal nerve, among others.
1.1.1. Types of Chemical Signals
In nature, living organisms maintain a constant degree of communication based on the exchange of multiple chemical signals [15] due to a variety of reasons, primarily associated with their need for food, protection, and reproduction. These chemical signals can be classified into two types: hormones, when they act within the same individual that produces them; or semiochemicals, when they act in an individual different from the signal producer. Hormones are chemical signaling molecules produced by tissues or endocrine glands that control various physiological processes within an organism [16]. In parallel, the chemical substances that different organisms use to communicate with each other and perceive their environment are called semiochemicals, a name derived from the Greek root semeon, meaning mark or signal [17]. This chemical communication is present in microorganisms, plants, and insects [18] as well as in vertebrates [19] and requires specific secretory mechanisms of semiochemicals, which induce changes in the physiology, metabolism, or behavior of the species that receive the chemosensory signals [20].
Semiochemicals are classified into pheromones, if they act upon individuals of the same species, or allelochemicals, if they affect individuals of a different species [21]. In both cases, these chemical signals may consist of a single chemical compound or a mixture of several. Regarding pheromones, they establish chemical communication among members of the same species, regulating aspects such as development, physiological state, social behavior, reproduction, or territorial marking [22]. On the other hand, allelochemicals are further classified, based on the beneficiary of the interaction, into four types: allomones, where the sender benefits from the message; kairomones, where the receiver is the beneficiary; synomones, when both species benefit from the communication, and apneumones, when the chemical signal is emitted by inanimate material and evokes in the receiver an adaptively favorable behavioral or physiological reaction.
Allomones, a term resulting from the combination of the Greek roots allos and hormonas meaning to excite others, are defined as interspecific chemical signals that give an adaptive advantage to the species that produce them [23]. Allomones from many organisms have been described, from lower plants to higher animals, where those emitted by predators primarily attract prey [24,25], and those produced by prey primarily repel predators [26]. For example, venoms or antibiotics are allomones, as they are chemical compounds produced by microorganisms with the aim of inhibiting the growth of other microorganism species. Conversely, a kairomone is an allelochemical that, when in contact with an individual of another species, evokes in the receiver an adaptively favorable behavioral or physiological response, while being unfavorable for the sender [3]. A notable feature of the main categories of chemical messengers is that they are not mutually exclusive, and an example of this occurs with the predator Elatophilus hebraicus, which uses the sex pheromone of its prey Matsucoccus josephi as a kairomone, such that when the predator perceives it, it feels a strong attraction [27]. Another example is the secretion from the mandibular glands of many ants when the nest is disturbed, which acts as a social alarm communication (pheromone), but also has a repelling effect (allomone) against aggressors [28].
Regarding synomones, they can be defined as allelochemicals produced by one organism that, when in contact with an individual of another species, evoke in the receiver a behavior or physiological response that is adaptively favorable for both [21]. Some synomones are repellent, and others are attractive [29], such as repellent molecules that warn of a danger like the toxicity of a plant [30], or chemical signals produced after a parasitic attack aiming to attract the natural predator of the parasite [31].
Lastly, there is a group of allelochemicals originating from inanimate sources, the apneumones, a term formed from the Greek root a-pne meaning breathless or lifeless. The receiver benefits from these molecules, but they differ from kairomones in that the producer cannot experience any disadvantage [32]. However, these apneumones are related to the presence of certain prey in specific environments, which is detrimental to organisms of another species associated with these compounds that might be nearby or on the inanimate material. For example, parasites and predators are drawn by apneumones to inanimate substances related to the presence of their hosts or prey [33].
Finally, it is important to emphasize that the exchange of chemosensory stimuli occurs in both terrestrial and aquatic environments. The most significant physicochemical characteristics of these chemical compounds are their size and polarity, as these are the primary factors determining their volatility in the air and solubility in water, respectively. Thus, in terrestrial environments, substances that act at a distance tend to be small and volatile, while in aquatic environments, where solubility is most relevant, even high molecular weight molecules can act at a distance. Therefore, based on their physicochemical properties, which dictate their potential mobility in different mediums, chemical signals can be classified into four types of compounds: volatile-soluble, volatile-insoluble, non-volatile-soluble, and non-volatile-insoluble [13].
Pheromones
Within semiochemicals, pheromones are compounds that allow for chemical signaling between individuals of the same species. In 1959, the entomologist Martin Luscher and the biochemist Peter Karlson introduced the term “pheromone” by combining the Greek roots pherein (to carry) and hormon (to excite). They defined it as a chemical substance or a mix of chemical substances released by a member of a species that induces specific behavioral responses or physiological changes in other members of the same species upon perception [34]. In fact, the first semiochemical to be isolated and characterized was bombykol, the sexual pheromone of the silkworm moth [35]. Subsequently, two categories of pheromones were defined based on their effect on the benefiting organism: releaser pheromones and primer pheromones. Releaser pheromones trigger immediate behavioral effects in the recipient organisms, whereas primer pheromones induce long-term physiological effects in the recipient individual [36]. Thus, typical releaser pheromones influence aspects related to mating, alarms, trails, and territorial marking. On the other hand, compounds determining the caste of social insects are primer pheromones [37]. Stemming from these discoveries, pheromones have been extensively studied in insects, primarily for their potential in pest control [38]. They have also been the subject of numerous studies in mammals [39].
For decades, the existence of pheromones in mammals has been assumed [40], though some authors have expressed their doubts [41]. Indeed, the concept of a pheromone derives from studies in insects, and the integration of the term in vertebrates has sparked controversy due to the complexity of information received simultaneously through other sensory modalities and the significance of learning in mammals, which can alter behavior, complicating the specific analysis of chemosensory communication [42]. Specifically, in pigs (Sus scrofa), male saliva pheromones are essential for courtship and copulation, but physical stimuli produced by applying pressure on the female back are also necessary [43]. As a result, five operational requirements have been introduced for a substance to be considered a pheromone: chemical simplicity of the signal; high stimulus selectivity with its response; an unequivocal behavioral response from the receiver that is morphologically consistent and functionally apparent; reception specificity based on species, and an unconditional linkage between stimulus and response [44]. In their literature review on chemical signals in terrestrial vertebrates, Apps et al. [45] identified up to 63 mammalian pheromones, such as in mice [46,47,48], hamsters [49], pigs [50], and elephants [51].
Kairomones
Kairomones, a term derived from the Greek root kairo (opportunist), are interspecific allelochemicals that confer an adaptive advantage to the organism receiving the chemical signal, while being disadvantageous to the emitter. Therefore, kairomones are semiochemicals emitted by one species to its own detriment [52]. However, even though kairomones can be detrimental to the emitting organisms, it has been suggested that they could lead to an evolutionary advantage [53]. Although the term kairomone was controversial when introduced, it is now widely accepted [54,55].
Due to the diversity of ecological phenomena associated with kairomones, they can be classified based on various criteria. According to their effect on the benefiting organism, two classes are distinguished: releaser kairomones, which induce an immediate behavioral response, and primer kairomones, which lead to long-term physiological responses in the receiver. They can also be classified based on their function in the receiving individual, resulting in four main groups: foraging kairomones, anti-predator kairomones, sexual kairomones, and aggregation kairomones [56]. Accordingly, foraging kairomones are used to locate food sources [57], anti-predator kairomones to mitigate the negative impact of a natural enemy [58], and sexual kairomones to find mates or for other sexual purposes [59]. Lastly, aggregation kairomones are used by both genders of the receiving species to form aggregations for various purposes, such as optimal exploitation of food resources, mate searching, or defensive reactions. Examples of foraging kairomones include the substances produced by the corn earworm (Heliothis zea), used by the parasite Microplitis croceipes to locate its host [60], or the lactic acid emitted by humans, which attracts the mosquito responsible for transmitting yellow fever, Aedes aegypti [61]. In some cases, kairomones used to find a host are produced by other microorganisms that develop within the host [62]. In the aforementioned cases, the chemical signals aid predators in locating their prey or hosts, but the opposite can also occur. Anti-predator kairomones emitted by predators or parasites are used as alarm signals, inducing defensive behavior in receiving organisms [63]. This is the case with certain plants that recognize kairomones produced by herbivores and emit compounds to repel them or to try to attract their natural enemies [64]. Lastly, sexual kairomones indirectly influence sexual communication and can be used to find mates, like the alcohols released by green leaves when female beetles of the Melolontha spp feed on them, attracting males of the same species [65].
1.1.2. Responsible Exocrine Organs
The independent evolution of multiple semiochemicals in mammals has resulted in a vast diversity of produced compounds as well as specialized secretory glands. These signals are commonly emitted externally via feces, urine, or saliva, but they are also found in other secretions like tear ducts, vaginal secretions [66], or are produced in specific glands like perianal ones [7]. Substances excreted through urine or feces can provide information about the age, identity, or gender of the depositor. In fact, there are body postures or movement patterns associated with urination and defecation, like scattering motions, that facilitate this chemical communication [67]. They also provide information about the location of individuals, especially relevant in various contexts such as social structuring or territorial distribution.
In this way, feces play a significant role in marking and intraspecific chemical communication in many mammals. Some odoriferous compounds originate from anal sac secretions [68], but others are a result of bacterial action on food during digestion [69]. In pigs, several semiochemicals present in maternal feces have been detected that attract piglets, resulting in positive behavioral changes in them, acting as a soothing agent and improving growth rates [70].
Both volatile and non-volatile substances can also be identified that act as chemical signals in the urine of most mammals, as it is a fundamental medium used in animal communication [71]. In guinea pigs, urinary chemical substances are involved in gender discrimination [41]. Additionally, the amino acid felinine, responsible for the characteristic odor of urine in this species and implicated in territorial marking, is found in the urine of domestic cats and other members of the Felidae family [72]. It appears in large amounts in male urine from six months of age and is believed to be a precursor to a pheromone attracting females [73].
Multiple semiochemicals are also found in saliva, as in the case of the boar, which has androgenic steroids in its saliva that act as sexual attractants, and can even stimulate puberty in juvenile females [74]. These compounds are produced in the testicles and transported by the bloodstream to the submaxillary salivary glands [75] where they are emitted in large quantities during copulation.
Similarly, in mammals, chemical stimuli related to sexual behaviors are produced in or near the genitals. For example, the male pig produces a sexual attractant that is part of the preputial secretions [76], while female hamsters attract sexually experienced males with their vaginal secretions [49]. Before copulation, males sniff and lick these secretions since they contain essential chemicals for adequate sexual behavior [77]. A similar phenomenon occurs in primates, where a chemical compound in female vaginal secretions, copulin, triggers copulatory behavior in male conspecifics. Stimulated by estrogens, this semiochemical enhances sexual responsiveness in rhesus monkeys [78]. In other species, like canids [79] or bovids [80], sexually attractive molecules have also been observed in female vaginal secretions.
On the other hand, various cutaneous glands are involved in the production and emission of chemical signals. These exocrine cutaneous glands release their compounds through a duct system, and depending on the type of secretion, they are divided into sweat-producing (sudoriparous) and sebaceous. The sweat glands are related to sweat production and contain hydrophilic molecules, while the sebaceous glands release sebum and lipophilic substances. Consequently, the various cutaneous glands can produce a complex mixture of chemical signals. Moreover, based on the secretion release mechanism, exocrine glands are also classified as apocrine, holocrine, and merocrine or eccrine. In apocrine glands, a portion of the cytoplasm of the cells becomes part of the secretion. In holocrine glands, cells are almost entirely destroyed and become part of the secretion product. Lastly, in merocrine glands, secretion occurs through an exocytosis mechanism, so there is no injury to the secretory cells. Examples of apocrine glands include mammary glands; holocrine examples are sebaceous glands, and merocrine examples include salivary glands.
Several examples of glands producing semiochemicals in mammals are described below. In rabbits, the apocrine glands of the chin are involved in maintaining social status and are used to mark territory by dominant males [81]. Male gerbils use the secretion of an androgen-dependent sebaceous gland to mark territory [82]. Other apocrine sebaceous glands, the tarsal glands of the male deer (Odocoileus hemionus columbianus), secrete a series of molecules, including a semiochemical that causes licking by female congeners [83]. In pronghorn males (Antilocapra americana), a compound produced in the subauricular gland was identified that induces licking, marking, or tapping in other males of the same species [84]. Another specific sebaceous gland is the morillo gland, found exclusively on the snout in capybara males and is related to the production of semiochemicals. These compounds are produced more by dominant males and are used for territorial marking when deposited on vegetation [85]. In the case of guinea pigs (Cavia porcellus), the sebaceous secretion of the perineal gland intervenes in dominance [86]. Similarly, in sugar gliders (Petaurus breviceps), dominant males have a frontal and a sternal gland, with which they mark territory and other members of their family group. Both male and female sugar gliders emit chemical signals through the paracloacal glands [87]. Merocrine or eccrine glands are abundant in humans and primates, but in the rest of the mammals, their location is restricted to the palms and soles. For instance, the Madagascar tenrec (Echinops telfairi) has cutaneous eccrine glands on the footpads, which produce a non-slip secretion containing semiochemicals that inhibit the growth of microorganisms [88]. Lastly, glands producing chemosensory stimuli in minks that become vestigial after weaning have been studied. These are apocrine sweat glands located in the neck and in the perineal and inguinal regions related to maternal recognition of the offspring [89].
1.1.3. Ethology and Scent Marking
In mammals, chemical signals can be emitted passively, without being associated with specific behavior, or they can involve a display of behavioral patterns that facilitate the dispersal of such a signal in the environment. For instance, the male pig passively emits a sexual attractant in its preputial secretions [90]. However, it also exhibits specific behaviors, such as chewing movements that increase the production of salivary secretion. This contains semiochemicals like androstenone or androstenol, produced in the testicles and stored in the parotid gland, which stimulate the immobility reflex in female pigs, facilitating the adoption of the mating posture [91]. These specific markings, termed "scent marking", have been described in numerous mammals, with a wide variety of behavioral patterns characterized in different species. This diversity arises from the various functions that chemical communication might entail [92], the multiple deposition sites of the signal [93], or even the location of the exocrine glands on the body.
Regarding function, initially, a relationship was established between marking and territorial defense. However, this behavior can play other roles in communication. In fact, several authors proposed various functions for chemical marking [94]. Thus, marking can act as a deterrent, avoiding aggression by warning about territory occupation; as a sexual attractant; as an orientation tool; as an indicator of identity, age, or dominance; as an alarm signal for conspecifics or even as an indicator of population size.
Regarding the territorial function, chemical marks define a specific area and prevent intruders from entering or cause their withdrawal, reducing defensive costs. Therefore, animals deposit these compounds at the boundaries of their territory and at strategic locations as a method of territorial dominance. This behavior is also reflected in experimentally studied mice, as subordinate individuals spend less time in parts of a cage treated with dominant male urine (Jones & Nowell, 1974). On the other hand, in rabbits, it has been shown that upon acquiring a dominant status, a male synthesizes 2-phenoxyethanol in its chin gland, which acts as a fixative making its secretions persist in the environment without dissipating [96].
Similarly, marking can have a stimulating or attracting function in the opposite sex, as seen in canids, where females show a higher frequency of urine marking during estrus to attract males [97]. In the case of the striped mongoose (Mungos mungo), a social and cooperative herpestid, both males and females increased the marking frequency during the latter estrus [98]. On the other hand, in rodents, the preputial secretion and urine of males can induce or suppress estrus and ovulation in females, in addition to accelerating female sexual maturity [99,100].
Another function of chemical marks is the spatial orientation of the individuals that produce them within their territory, allowing them a better familiarity with the environment. In this way, species like the slow loris (Nycticebus coucang) use chemical marks to trace paths [101]. Likewise, other mammals, such as coyotes (Canis latrans), mark food or hiding places with urine [102].
Marking behaviors are also used to deposit chemical signals that serve as an indicator of individual identity and provide information about dominance, social rank, sex, or age. In this way, beavers (Castor canadensis) can discriminate between familiar or unknown individuals [103], just like the sugar gliders (Petaurus breviceps papuanus), who are not only able to distinguish both individuals and groups of unknown individuals but also trigger aggressive behaviors in the receiver [87]. This relationship between individual recognition and dominance is reflected in marking patterns. Dominant individuals usually mark more frequently than subdominant or submissive ones and are significantly more likely to adopt specific marking postures, as seen in canids [104]. Likewise, dominant individuals frequently mark in situations where they are intolerant. On the other hand, chemical signals allow recognition between males and females [105] and even induce synchronization of the reproductive state [106].
On occasion, stressed individuals secrete alarm molecules, keeping the members of their group alert. Exposure to these signals produces behavioral changes in the receiver, such as reduced rest, increased heart rate, or stress-induced hyperthermia. This phenomenon has been studied in rats [107]. In cows, the presence of alarm signals in the urine of stressed congeners triggers an increase in cortisol and fearful behaviors [108].
Finally, in certain animals, scent marking has been observed to act as a population self-regulation mechanism, limiting the number of inhabitants before food becomes a limiting factor. This happens in the American beavers (Castor canadensis), who communicate through marking, delineating occupied territory, and preventing further colonization of a particular area. This prevents the population density from increasing above a limit while maintaining a balance between the amount of resources demanded by the residents and the regeneration capacity their habitat can sustain [109].
Regarding the deposition sites of chemical substances, five contexts have been considered: direct release to the surrounding air, deposition on a specific object in the environment, deposition on the substrate, marking on a social partner, and self-marking or self-anointing. We usually associate the emission of chemical signals with specific marking behavior patterns, but the truth is that mammals also have the ability to release chemical compounds without any visible sign of it. For example, many mammal species emit chemical signals when scared, like the black-tailed deer, which secretes chemical compounds in its metatarsal glands when in situations of fear or stress [83].
On the other hand, marking may be associated with certain objects, so animals deposit feces and urine on them or use rubbing movements with their glandular areas to impregnate them with specific secretions. These marking movements directed at objects often involve adopting unusual postures and allow depositing chemical signals at a certain height, improving communicative efficiency [110]. These objects can be part of a complex communication system between individuals of the same species, as seen in the European brown bear (Ursus arctos arctos), which exhibits a rubbing behavior against tree trunks. By standing on their hind legs, they rub their chest, back, and neck while biting and scratching the bark, depositing chemical signals. Males have preferred marking areas, which can be used by a single individual or be communal and used by several males. Places marked by an unknown adult male are avoided by young bears. On the other hand, females mark in areas separate from the males but can also mark on communal trunks along with other males [111]. In the case of goats, when a male reaches sexual maturity, he starts rubbing his head and neck on objects around him as a method to disperse chemical signals with an attractive effect on females [112].
In addition to the deposition of chemical signals through urine or feces in the substrate, mammals show specific marking behaviors on the terrain. In particular, desert-dwelling rodents from the Heteromyidae family perform sand baths [113]. This behavior consists of digging in the substrate and performing a series of specific rubbing movements depending on the species. In this way, the animal grooms itself while spreading chemical secretions in marking areas of the environment, which are sometimes shared. Other mammals have behaviors like scratching the ground, which may involve the deposition of chemical signals produced in the autopodial glands.
Some mammals use marking on a partner or on conspecific individuals through characteristic behaviors. These behaviors may involve friction movements with specific glandular areas on the receiver [87]. The previously mentioned marking of European rabbits by rubbing with their chin on objects or congeners to impregnate them with the secretion of the chin glands allows them to identify members of their social group to establish their territory, thus maintaining the social hierarchy [81,114].
Finally, some mammal species use their own or foreign sources of chemical signals to spread the compounds on their body for various purposes. In this way, the black-tailed deer extends the hind limb to spread the previously described secretions from the tarsal glands over its own head [83]. Likewise, spider monkeys exhibit a self-anointing behavior rubbing their body with a mixture of saliva and plant material [115]. However, in other cases, individuals use foreign chemical signals, like rats, which impregnate themselves with chemical signals from their main predator, the weasel, to mask their own chemical identity and avoid possible attacks [116].
1.1.4. Chemical Signal Detection Systems
Animals are constantly examining their environment for chemical substances that guide them to food sources or favorable habitats. They also investigate other substances that control social interaction and reproductive behavior [117]. These chemical compounds can be detected through direct contact with salivary or nasal secretions, or by monitoring the respiratory air stream through the nasal cavity. In both cases, the chemical signals are perceived by highly specialized detectors, the chemosensory neurons, which are organized into structurally independent subsystems in the nasal cavity. These olfactory subsystems can be divided based on the anatomical location of their sensory neurons, the type of olfactory receptors they express, the signaling mechanisms they use to transduce chemosensory stimuli, the chemical stimuli to which they respond, and the axonal targets of their sensory neurons in regions of the olfactory forebrain [118].
Initially, two chemical perception systems were identified with sensory neurons located in the nasal cavity: the main or olfactory system and the accessory or vomeronasal system. The olfactory neurons constitute the main olfactory epithelium, which lines the convolute surfaces of both the ethmo- and endoturbinates (Figure 1), as well as the caudal part of the nasal septum (Figure 2) [119]. These neurons send their nerve projections to the main olfactory bulb [120,121]. On the other hand, the vomeronasal system presents a highly specialized peripheral sensory structure where the sensory neurons of the vomeronasal epithelium are located, the vomeronasal organ (VNO). The neurosensory information travels through the vomeronasal nerves to the accessory olfactory bulb, an independent structure adjacent to the main olfactory bulb [122,123].
More recently, new sensory systems specialized in the detection of semiochemicals have been discovered in the nasal cavity of mammals, which together are called olfactory subsystems [124]. This is the case of the Grüneberg ganglion, the septal organ, or the solitary chemosensory cells. The Grüneberg ganglion is a structure formed by specific sensory neurons located in the anterior area of the nasal cavity, near the nasal vestibule [125]. On the other hand, the septal organ consists of an isolated area of sensory epithelium located at the base of the nasal septum, ventral to the olfactory epithelium [126]. Finally, the solitary chemosensory cells are distributed in the anterior part of the nasal cavity but are also located in other specific areas such as the entrance of the vomeronasal organ [127]. The first two mentioned subsystems project their neuronal axons to specific areas of the main olfactory bulb, while the axons of the chemosensory cells associated with the VNO are incorporated into the sensitive afferents of the trigeminal nerve [128].
In both the main and the accessory olfactory bulbs, glomeruli are formed in the confluence area of the axonal terminals of the sensory neurons with the dendrites of the second-order neurons with which they establish contact. The vomeronasal glomeruli are part of the accessory olfactory bulb and are clearly differentiated from the glomeruli of the main olfactory bulb by their smaller size and less defined boundaries. However, the glomeruli formed by neurons of the Grüneberg ganglion or the septal organ do not have a clearly differentiated boundary within the main olfactory bulb, although specific areas for each have been described. In fact, a necklace glomerulus complex has been identified in the caudal region of the MOB, consisting of several specific glomerular groups related to the innervation of specific neurosensory cells, such as the cells of the Grüneberg ganglion or cells that express the atypical olfactory receptor guanylyl cyclase type D (GC-D) [129]. Additionally, subbulbar formations have been identified ventral to the accessory olfactory bulb in rats [130], hedgehogs [131], and in lagomorphs - with the latter presenting a particularly complex organization [132] - that could be related to certain specific sensory pathways. The recent characterization of the olfactory limbus - the transition zone between the accessory and main olfactory bulbs - in the fox [133] points to a high morphofunctional complexity in the central integration of chemosensory information.
This wide range of versatile chemodetectors perceives myriads of chemical compounds that vary in their physicochemical properties and function, and that may be involved in controlling multiple behaviors and physiological responses. This multifunctional task is achieved through orchestrated interaction between the various olfactory subsystems, each of which specializes in different functions and uniquely contributes to fulfilling the overwhelming tasks of the sense of smell.
1.1.5. Olfactory Subsystems
In the main olfactory system, molecules are detected through direct contact with the olfactory mucosa of the nasal cavity. The fundamental component of the olfactory mucosa is the olfactory neuroepithelium, comprised of olfactory sensory neurons. These are bipolar cells whose dendrites reach the epithelial surface. Here, long cilia embedded in nasal mucus provide an extensive surface area for interaction with odorants. The process of neurotransduction occurs within these cilia [134]. The ciliary membrane contains receptor proteins and elements of the olfactory transduction machinery, allowing these cellular compartments to act as chemosensory units initiating signal detection.
Olfactory receptors consist of various specialized receptor proteins coupled to the G protein (GPCR) [135]. Each olfactory sensory neuron expresses a unique receptor from a family that encompasses about 1,000 genes. In fact, olfactory receptors are associated with the largest gene superfamily in vertebrates [136]. Additionally, each olfactory receptor interacts with a broad range of chemical compounds, albeit with different affinities. Therefore, a single olfactory receptor can recognize multiple odorants or olfactory chemical signals, and conversely, multiple olfactory receptors can recognize a single odorant. This combinatorial receptor strategy is utilized to encode odor qualities [137].
The axons of the olfactory neuroepithelial cells converge to form the olfactory nerve (first cranial nerve) [138], whose fibers pass through the cribriform plate of the ethmoid bone to establish their first synapse in the glomeruli; spherical structures found on the periphery of the main olfactory bulbs. Anatomically, the olfactory bulbs are rostral extensions of the cerebral hemispheres and as such, belong to the rhinencephalon, the olfactory portion of the telencephalon. They thus constitute the first synaptic and integrative station of the olfactory system. Notably, from a morphofunctional perspective, all neurons expressing the same type of olfactory receptor send their axons to common glomeruli in the main olfactory bulb [139]. From the main olfactory bulbs, information is relayed through the lateral olfactory tract to deeper brain areas [140].
In contrast to the main olfactory system, molecules stimulating the vomeronasal system activate the sensory neurons of the vomeronasal organ, a bilaterally symmetrical tubular structure located dorsally to the floor of the nasal cavity and on both sides of the base of the nasal septum. The characteristics of the vomeronasal epithelium lining the inside of the vomeronasal duct resemble those of the main olfactory epithelium. However, the dendrites of the vomeronasal sensory neurons have microvilli instead of cilia [141]. Both vomeronasal organs are protected by a vomeronasal capsule, which varies in nature depending on the species and is composed of bone, cartilage, or a mixture of both forming specific patterns [142,143]. Caudally, the capsule is typically closed, while rostrally, the vomeronasal organ communicates with the nasal cavity or the nasopalatine canal, depending on the species. In species like rodents, lagomorphs, or some primates, the vomeronasal duct directly opens into the nasal cavity [144], whereas in other species like marsupials, monotremes, or ungulates, the vomeronasal duct opens into the nasopalatine canal, also known as the incisive canal, which connects the oral and nasal cavities [145]. This topographical feature is an initial reflection of the existence among different mammalian species of a wide variability of the vomeronasal organ, at the morphological, topographical, and functional levels. Parallel to the main olfactory system, the axonal terminations of the vomeronasal sensory neurons form the vomeronasal nerve. Subsequently, upon leaving the VNO, they course dorsocaudally to penetrate the cribriform plate of the ethmoid bone and reach the accessory olfactory bulb, constituting its outermost layer, the vomeronasal nerve layer. From the accessory olfactory bulbs, information is transmitted to regions of the central nervous system involved in mediating the pheromonal responses highlighted earlier in this introduction [146].
It has often been hypothesized that the various olfactory subsystems function independently. This idea, in the case of the main olfactory system and the vomeronasal system, is supported by the fact that while olfactory neurons project to the main olfactory bulb, vomeronasal neurons project to the accessory olfactory bulb. Indeed, highly specific afferent patterns have been described in both cases. In the vomeronasal system of rodents, the apical and basal regions of the vomeronasal neuroepithelium project their axons to the anterior and posterior accessory olfactory bulb, respectively [147]. Similarly, several distinct areas of sensory neuroepithelium in the nasal cavity have been identified that correspond to specific regions of the main olfactory bulb. Initially, four zones were established, with no overlap between them [148], but more recent studies discern up to nine zones with variable overlap [149]. Currently, the information gathered on the olfactory and vomeronasal systems supports the notion that they are distinct entities, each with unique structural characteristics and chemosensory tasks. However, they share a number of morphofunctional aspects. Among these are the presence of vomeronasal receptors in the MOE and vice versa [150,151], and the convergence of secondary projections from both systems in the basal telencephalon [152].
The Grüneberg ganglion (Figure 3) consists of a bilateral and compact group of neurons located in the vestibule of the anterior nasal cavity, in a dorsomedial position near the opening of the nostrils [153]. The sensory neurons of the Grüneberg ganglion are embedded in a network of fibroblasts between the nasal septum and the keratinized squamous epithelium permeable to water that lines the nasal vestibule [154]. These neurons, which possess various receptors, lack prominent dendrites or microvilli and do not directly access the nasal lumen, though they have numerous cilia that play a role in sensory transduction [117]. Peripherally, glial cells envelop the neurons, trapping the cilia within the ganglion. By detecting the expression of the olfactory marker protein (OMP), it is observed that their axons project along the nasal septum, enter the cribriform plate of the ethmoid bone, and eventually reach the anterior olfactory nucleus of the telencephalon, without synapsing in the main olfactory bulb. Another peculiarity of the neurons of the Grüneberg ganglion is the absence of projections to the vomeronasal organ [155]. Though its function is not yet fully understood, the ganglion could have a role in thermosensation [156].
The septal organ (Figure 4 and Figure 5) is a bilateral area of sensory neuroepithelium located at the base of the nasal septum, ventral to the main olfactory epithelium and rostral to the choanae [157]. Morphologically, it is composed of basal cells, supporting cells, and ciliated sensory neurons with flattened cell bodies and shortened dendrites [124]. The septal organ projects its neuronal axons to certain glomeruli located in the posterior region of the main olfactory bulb, specifically in the ventromedial zone [158].
On the other hand, although the projection pattern of the septal organ is already defined in newborns, during the first postnatal days, there is a significant increase in the area of the organ [159] and in the number of septal glomeruli [160]. Based on the location of the septal organ, it has been suggested that it may serve an alert function, detecting odors in the environment when breathing is relaxed, and the air stream does not reach the main olfactory epithelium. In fact, the septal organ may respond to some odors with greater sensitivity than the main olfactory epithelium [161], and both share the main olfactory signal transduction pathways. Another suggested possibility is that it might play a role in detecting low volatility compounds transferred by licking, which reach both the vomeronasal organ and the septal organ but not the main olfactory epithelium [162]. Thus, the septal organ could detect both odors and chemical signals with sociosexual influence.
Finally, another independent olfactory subsystem has been identified, comprised of solitary chemosensory cells. The majority of these sensory cells are located in the anterior nasal cavity, but there is also a cluster in the entrance duct of the vomeronasal organ [127], and other populations present in the larynx and in the deeper respiratory tracts [163]. At the same time, fibers from the trigeminal nerve are observed near the solitary chemosensory cells, indicating sensory information transmission to these fibers. The upper respiratory tract is continuously assaulted by harmful substances and xenobiotics carried by the inspiratory air flow that are detected by the trigeminal nerve, which evokes protective reflexes such as sneezing, apnea, and local inflammation of the mucosa. Likewise, certain inhaled pathogens and irritants stimulate the solitary chemosensory cells, which help enhance the chemical response capabilities of the trigeminal nerve. Additionally, solitary chemosensory cells play a significant role in regulating the access of chemical substances to the vomeronasal organ [128], thus providing a mechanism to early identify potential environmental irritants. The fact that anosmic individuals or animals lacking functional olfactory systems still retain the ability to detect a variety of chemical irritants through the trigeminal system confirms the crucial role played by this system in nasal chemosensation [164].
2. Vomeronasal System
The vomeronasal system (VNS) or accessory olfactory system is specialized in detecting chemical signals, primarily pheromones, kairomones, and molecules from the major histocompatibility complex. It consists of a set of anatomically and histologically distinguishable structures from the main olfactory system. It is present in most reptiles [165,166] and amphibians [167], but it is particularly developed in mammals, in which this chemosensory system comprises three main components: the vomeronasal organ (VNO) (Figure 6), which acts as the peripheral chemoreceptor organ detecting chemical signals; the vomeronasal nerve, transmitting information to the brain; and the accessory olfactory bulb (AOB), the first neural center where vomeronasal afferent information is processed and integrated before heading to specific areas of the CNS [168].
The significant functional diversity of the vomeronasal system is expressed in the existence of three distinct subpopulations of vomeronasal sensory neurons. Each is associated with a specific family of chemosensory receptors: vomeronasal type 1 receptors (V1R), vomeronasal type 2 receptors (V2R), and formyl peptide receptors (FPR) [169]. However, it is crucial to note that not all mammalian species have functional receptors in all three families, indicating a diversity in chemical signal detection and specialized adaptation based on the biological and ecological needs of each species. This phenomenon underscores the richness and versatility of the VNS in detecting and processing chemical signals, highlighting the importance of chemical communication in mammalian life.
Identifying these receptor families was essential, and the study of G-proteins was crucial. Initially, by analyzing the expression of Gαo and Gαi2 proteins in rats, it was observed that the nerve endings of the vomeronasal neuroreceptor cells of the AOB were organized into two complementary regions [170]. Subsequently, both G-proteins involvement in the transduction chain of vomeronasal neurons was determined, and the V1R receptor family in mice was discovered [171]. At that time, the expression pattern of the V1R receptors matched that of the Gαi2 protein. However, the second family of vomeronasal receptors was not identified until two years later when three separate studies examining the Gαo protein expression in the VNO simultaneously demonstrated the existence of the second vomeronasal receptor family, V2R [172,173,174]. Finally, the third family of vomeronasal receptors, formylated peptide receptors (FPR), which coexpress with both G-proteins (Gαi2 and Gαo), was identified [175]. On the other hand, each neuronal population maintains a specific projection pattern to the AOB. Despite the fewer types of vomeronasal receptors compared to olfactory receptors, the vomeronasal receptor neurons projection pattern to multiple AOB glomeruli seem to be more complex than those in the primary olfactory system [176].
There is a general consensus that the VNS is primarily responsible for perceiving pheromones, although it also perceives other non-pheromonal chemical signals, such as kairomones, which mediate defensive behavior [177], and other types of chemical signals vital for tracking prey and attack behavior, as seen in reptiles [178] and urodele amphibians [179]. In certain mammals, such as the gray short-tailed opossum, the VNS also influences food preference [180].
Chemical stimuli found in urine deposits, vaginal secretions, odorous gland secretions, or saliva can be investigated by direct contact. However, specific behaviors are also used to facilitate the entry of non-volatile substances into the VNO, such as facial grooming and "flehmen." The "flehmen" behavior is seen in ungulates and felines and consists of adopting a specific facial posture with the head tilted back, the mouth slightly open, the upper lip everted, and the neck extended for a few seconds [181,182]. It usually occurs after contact with biological secretions from conspecifics, and males exhibit this behavior more frequently [183,184].
Historical interest in the vomeronasal system began with the discovery of the VNO by the Danish anatomist Ludvig Levis Jacobson in the early 19th century, who described its key macroscopic features in a broad range of non-human mammals [185]. While previous illustrations showed the supposed location of the VNO in human nasal septum drawings [186], Jacobson reported this structure absence in Homo sapiens. However, the human VNO was later discovered in embryos [187] and a detailed histological description was then carried out both in fetuses and adults [188]. Although Jacobson also contemplated the hypothesis of a possible sensory function of the organ, he mainly suggested a secretory role. The histological contributions of Balogh [189], Klein [190], and Piana [191] revived the hypothesis of the sensory function, but only by the end of the 19th century, the availability of the Golgi technique definitively showed the morphological similarity of the neurons of the olfactory and vomeronasal epithelium of the snake, thus establishing the sensory function of the VNO [192]. On the other hand, approximately half a century after the discovery of the VNO, the AOB was identified in sheep by Balogh [189] using traceability and dissection of the vomeronasal nerve. However, the term AOB was coined by Von Gudden [193] following his studies on the vomeronasal nerves in rabbits. Later on, Santiago Ramón y Cajal provided a detailed, accurate, and specific description of the AOB in various mammals, revealing its laminar architecture and the presence of different cell types [194]. It was the North American neuroanatomist Rollo McCotter [122] who established, in a broad range of species, the different nature of the olfactory and vomeronasal nerves and their respective destinations in the MOB and AOB. Finally, in the second half of the 20th century, a clear relationship between the VNS and reproductive behavior was established [195,196] leading to the seminal work of Powers & Winans [197] which convincingly demonstrated the critical role that the VNO plays in rodent reproduction.
Evolutionarily, the vomeronasal system has been linked to the transition of vertebrates to terrestrial environments; however, recent evidence suggests that a precursor SVN exists in teleost fish and its evolutionary origin predates the divergence between teleosts and tetrapods [10,198]. Added to this is the unique case of several species of lungfish that have a vomeronasal system homologous to mammals, showcasing a defined vomeronasal organ and an accessory olfactory bulb [199,200,201,202]. Hence, the significance of chemical communication has been a constant throughout the evolutionary history of vertebrates, resulting in significant morphofunctional variations among the chemosensory systems of different species. Specifically, the shift from aquatic to terrestrial life led to changes that significantly impacted pheromonal communication in vertebrates. This arose from the transformation of its key chemical property from solubility to volatility, a process that altered pheromone release mechanisms, accompanied by morphological and physiological changes in the sensory organs [203].
Subsequently, some terrestrial animal species returned to aquatic environments, such as cetaceans, which underwent drastic changes in their olfactory morphology during this migration. While their terrestrial relatives, including hippos, exhibit a defined VNO [204,205], no VNO has been found in any cetacean [206]. In contrast, toothed whales also lost their main olfactory system [207]. However, sea snakes, which evolved from terrestrial tetrapod reptiles, feature a functional and well-developed underwater VNS, while losing their main olfactory system [208]. In snakes, the vomeronasal system is predominantly considered the major chemosensory system [209,210,211,212]. The failure of their olfactory system to adapt to aquatic life, in contrast to the successful adaptation of their vomeronasal system, underscores the importance and development of the VNS in these reptiles. Other primarily aquatic reptiles, such as sea turtles, have a well-developed VNS [213]. However, while some alligator and crocodile embryos show VNS, it regresses to be absent in adulthood [214,215]. Yet, most reptiles [216] and amphibians [217] have a functional VNS.
Regarding airborne vertebrates, there is no evidence of pheromonal communication in most birds due to the absence of their VNS [218]. The same is observed in many bats, though certain species possess a particularly well-developed VNS [219]. Finally, the vast majority of terrestrial mammals have a functional VNS, and some, like rodents, lagomorphs, or marsupials, have an especially developed VNS. Semi-aquatic mammals like the capybara [220], hippopotamus [221], beaver [222], and platypus also exhibit a VNS [223].
Among primates, it is believed that the last common ancestor with a functional vomeronasal system might have been small, arboreal, and nocturnal [224,225]. Without adequate light, vision is limited, heightening the reliance on olfactory signals [226]. Presently, primates can be classified into strepsirrhines and haplorhines, based on the presence or absence of a rhinarium: a moist, hairless skin area around the nostrils seen in some mammals. Strepsirrhines, which include lemurs and lorises, possess a rhinarium and are nocturnal, in addition to having a highly developed VNS [227]. On the other hand, haplorhines lack a rhinarium and are mostly diurnal. They include New World monkeys or platyrrhines, and Old World monkeys or catarrhines, among which are the great apes and humans. Regarding the vomeronasal system, platyrrhine monkeys have a well-developed VNO [228]. In parallel, the VNO of catarrhine primates is generally absent; however, a rudimentary VNO in the postnatal stage has been observed in certain chimpanzee and human individuals. In both species, their VNOs have a superior position and specific histological features differing from those displayed by prosimians and New World primates [229].
For humans, it was traditionally thought that their VNS degenerated towards the end of fetal life, so only occasionally could vestigial remains be found in adults. This was consistent with a diurnal catarrhine primate. Also, the specific features of its VNO in adults increased the complexity of identifying such structures. Currently, the functionality of the human VNS during the prenatal phase is recognized, though its vestigial presence in adults and its level of functionality in the postnatal phase remain subjects of debate [230,231,232,233].
2.1. Anatomy of the Vomeronasal Organ
The vomeronasal organ comprises two tubular structures located bilaterally at the base of the anterior nasal septum. Both organs have a single point of communication with the exterior which, depending on the species, can either be located in the nasopalatine or incisive duct - a conduit that connects the oral and nasal cavities through the palatine fissure - or directly in the nasal cavity [234].
Each organ consists of two clearly differentiated elements: the vomeronasal duct, which forms the lumen of the organ and is lined by a pseudostratified columnar epithelium, and the vomeronasal capsule, a rigid and protective envelope of either bony or cartilaginous nature, depending on the species [142]. Associated with the vomeronasal duct is the parenchyma, tissue responsible for the organ function, consisting of an accumulation of soft tissue associated with the duct, rich in glands, vessels, nerves, and connective tissue [235].
Upon making a transverse cut in the central part of the organ, the typical crescent shape of the duct lumen at that level is observed. Internally, the duct is lined throughout its surface by two distinguishable epithelia. On its lateral side, it has a pseudostratified and ciliated respiratory epithelium, while on its medial part, the vomeronasal sensory epithelium (Figure 7) is located [236]. This sensory epithelium consists of a thin layer of basal cells, a broad central layer of bipolar neuroreceptor cells, and an outer layer of supporting cells that sustain the dendritic processes of the neuroreceptor cells (Figure 8). These dendritic processes project towards the lumen. At this level, they form microvilli that contain the vomeronasal receptors, which are responsible for recognizing the molecules involved in chemocommunication [237,238].
The nerve fascicles of the parenchyma are constituted by the coalescence into bundles of the axonal processes of the neuroreceptor cells, which converge from the vomeronasal epithelium. These nerve bundles are located in the dorsal and medial areas of the VNO, although occasionally they can also run on the ventral or lateral side of the duct. In turn, these bundles come together to form the vomeronasal nerve, which courses dorsocaudally in the submucosa of the nasal septum [239]. After passing through the cribriform plate of the ethmoid on its medial part, the vomeronasal nerve projects towards the anterior area of the telencephalon to synapse in the accessory olfactory bulb [240].
In the central region of the VNO, both the glandular and vascular components of the parenchyma are predominantly situated in the lateral portion of the parenchyma. Meanwhile, the glandular tissue is also concentrated in the caudal part of the VNO. The vomeronasal glands (Figure 9) are responsible for secreting mucus that is discharged into the lumen of the duct, either at its commissures or in the central part of the respiratory epithelium [241]. The mucus facilitates the entry and exit of molecules within the duct, but it also plays a crucial role in vomeronasal perireceptor processes. Histologically, these glands are distinguished by their acinar or tubular morphology, and by their serous or mucoid secretions, respectively [242,243]. Concerning the vascular tissue of the VNO, only arteries of significant caliber can be found in its posterior region; however, it has large veins that run parallel to the vomeronasal duct on the lateral flank [244]. This distinct morphological organization is important as it is involved in the vascular pumping mechanism that is activated to allow the entry of chemical stimuli into the organ. Specifically, when the vessels of the parenchyma contract, the lumen of the duct expands, creating a vacuum effect that draws chemical molecules into the duct. Similarly, when the vessels dilate, a constriction of the duct occurs, expelling the content of the duct outward. This vascular pumping mechanism is known as the vomeronasal pump [245,246,247].
The vomeronasal capsule provides structural support for the proper functioning of the vascular pump, counteracting the potential collapse of the parenchyma that the increase in negative pressure might induce, while simultaneously safeguarding the encapsulated structures. Its morphology not only varies along the VNO, but there are also notable differences among various species, both in its nature and its configuration [142]. More typically, towards the front, the capsule is shaped like an incomplete ring with a lateral opening associated with the outlet of the vomeronasal duct. In the central region, the ring becomes complete, but posteriorly the capsule separates again, adopting a "J" shape, with a dorsolateral slit allowing the passage of the vomeronasal nerves. As we progress caudally, on occasions, the ring closes entirely. Its nature can be bony, cartilaginous, or a combination of both in different patterns, making it one of the VNO components with the most diverse design from a comparative anatomy standpoint.
Regarding the sensory transduction of the VNO, in the most studied model, the murine, the neuroreceptor cells are organized into different subpopulations throughout the vomeronasal epithelium, following a clearly defined zoning. In this way, the V1R receptors, linked to the Gαi2 protein, are confined to the neurons whose cell bodies are located in the apical part of the epithelium, while the neurons expressing the V2R receptors, linked to the Gαo protein, locate their cell bodies in the basal layer of the vomeronasal sensory epithelium [248]. In both cases, each neuroreceptor cell exclusively expresses a single receptor among the total population of vomeronasal receptors [173]. Formyl peptide receptors co-express with both G proteins in the two neural layers. On the other hand, it has been demonstrated that the peptides of the major histocompatibility complex (MHC) can perceive non-volatile molecules, both in the olfactory and vomeronasal epithelium of mammals. This suggests a general role of MHC peptides in chemical communication, even in species lacking a functional VNO [249,250].
The transduction cascade of virtually all vomeronasal neurons converges in most species on the activation of a member of the transient receptor potential (TRP) channel family, the TRPC2, which is expressed in both neural layers of the VNO [251]. It was observed that the protein resides in the microvilli of the sensory neurons and coincides with the expression of both Gai2 and Gao [238]. In fact, the genomic sequence of the intact TRPC2 gene is considered a predictive marker of the functionality of the vomeronasal organ [218]. When the Trpc2 gene was knocked out in mice, its pivotal role in vomeronasal-mediated behavior became evident. These modified mice exhibited reduced aggressive responses and did not differentiate between males and females, displaying sexual behaviors towards both. This suggests that these mice are unable to discern the gender of their counterparts due to a lack of olfactory signal transduction through VSNs [252,253]. In humans, the loss of a functional TRPC2 gene was confirmed; however, the presence of functional V1R receptors has been observed in the olfactory mucosa [254]. It should also be noted that chemical communication in vertebrates originated long before the development of the VNO, and pheromone detection can also be mediated by other olfactory organs [255].
While initially, it was thought that all mammals expressed both families of vomeronasal receptors, the absence of the V2R family in the vomeronasal system of various species was subsequently described. Therefore, based on the presence or absence of the two neuronal subpopulations linked to the V1R and V2R vomeronasal receptors, two vomeronasal transduction models were identified: the segregated model and the uniform model. In the segregated model, both types of vomeronasal receptors coexist, while in the uniform model the V2R receptor is absent, and only V1R type neurons are expressed. Recent research on the Tammar wallaby (Notamacropus eugenii) concluded that macropods could present a third vomeronasal transduction model, consisting only of V2R neurons, as they did not find expression of the Gαi2 protein in the VNO or in the AOB [256].
A segregated-type vomeronasal system has been characterized in certain species of rodents [257], lagomorphs [143], and marsupials [258], while the rest of the studied mammals, such as ungulates, carnivores, or primates, fall within the uniform model [259,260,261].
The neurochemical study of the organ allows obtaining characteristics about its functionality using G proteins, but other markers are also used that provide specific information about this structure. The calcium-binding proteins calbindin (CB) and calretinin (CR) are used to identify neuroactive substances, but also to differentiate cell populations or to define the morphology of vomeronasal neuroreceptor cells [262]. On the other hand, the GAP-43 marker shows neuronal growth and is expressed in the nerve bundles present in the organ [263]. These markers are complemented with routine stains (hematoxylin-eosin) or specific stains (Gallego trichrome), which display the structural characteristics of the VNO. Likewise, using PAS (Periodic acid Schiff) and Alcian blue stains, the nature of the vomeronasal glands can be identified, staining their secretions with neutral and acid mucopolysaccharides, respectively [264].
Likewise, in the study of the VNO, lectins such as Ulex europaeus agglutinin (UEA), Bandeiraea simplicifolia isolectin B4 (BSI-B4), or Lycopersicon esculentum agglutinin (LEA) have been frequently employed, as some are specific to the vomeronasal and/or olfactory system in various mammals [265]. Thus, it is possible to differentiate neuronal populations and study the morphology of the neuroreceptor cells in the sensory epithelium of the VNO. On the other hand, when expressed in the neuronal axons of these cells, it becomes easier to identify the nerve bundles of the VNO and track the vomeronasal nerve by analyzing its topography and anatomical relationships.
Over the past decade, genomic studies have paved the way for groundbreaking insights into the VNS. To date, only a limited number of links between chemical ligands, vomeronasal receptors, and behavior have been clarified. This is primarily due to the challenges encountered when dealing with expansive, homologous gene families that have a high degree of sequence similarity. Nonetheless, when examining mouse strains with mutations in genes linked to ligand-VR signal transduction, the role played by the VNO in various social behaviors can be studied in a more specific way. These behaviors include male-to-male and maternal aggression, sexual allure, lordosis, selective pregnancy termination, and interspecies reactions such as aversion and defensive behaviors [266].
One significant discovery has been the broad expansion and diversification of the vomeronasal receptor gene families, V1Rs and V2Rs [267,268]. High-throughput sequencing methodologies, such as RNA-Seq, have enabled researchers to identify and catalog an extensive array of VNRs in various species, showcasing a vast diversity that arguably correlates with species-specific pheromonal communication [11,266,269]. Another groundbreaking revelation is the functional differentiation of V1R and V2R receptors, elucidated through transcriptomic analyses. While V1Rs primarily respond to small volatile molecules, V2Rs are more attuned to larger, peptide-based cues [270].
Comparative genomics indicate that the retention or loss of specific vomeronasal receptors often aligns with the ecological and social structures of the species in question. For instance, species with more intricate social hierarchies or mating systems tend to possess a richer repertoire of functional VNRs [271]. Moreover, the transcriptome analysis of the VNO in mice under different physiological and environmental conditions showed notable variations in the expression of vomeronasal receptors even among individuals of the same species [272,273,274].
The morphological diversity of the VNO among different orders of mammals is also notable, and indeed, since the advent of Darwin theory of evolution, the morphological configuration of the VNO has been used as a phylogenetic classification method [275,276,277]. A significant portion of the studies on the SVN (secondary vomeronasal organ) have been conducted in rodents, specifically in myomorphic species like the laboratory rat (Rattus norvegicus domestica) or the mouse (Mus musculus), which have been used as a general SVN model in rodents (Figure 10). Both species display a highly developed vomeronasal organ, with a large neuroreceptor epithelium, of considerable thickness and cell density [278]. The vascular pump consists mainly of a large central vein, which is surrounded by other smaller-diameter veins [279]. The vomeronasal capsule is made up of a thin bone layer in adults; however, in newborn rats, this covering is cartilaginous [235]. A thoroughly investigated aspect in the rodent VNO is the presence of postnatal proliferation in the vomeronasal epithelium. The distribution of these proliferating cells varies with age. In neonatal rats, such cells are distributed fairly evenly across the sensory epithelium. However, starting from P21, the majority of these cells tend to cluster near the boundaries with the non-sensory epithelium [280]. These findings have been more recently corroborated in mice using bromodeoxyuridine (BrdU) immunohistochemistry [281].
Other myomorphic rodents display specific morphological features in their VNO, but in general, all show a comparable level of development. For instance, the Gambian rat (Cricetomys gambianus) displays certain ossification areas in its cartilaginous vomeronasal capsule [282]. Among the sciuromorphic rodents, the daurian ground squirrel has an exceptionally voluminous vomeronasal vein, which, together with other smaller-caliber veins, constitutes the vascular pump. It also displays a sensory epithelium with significant thickness, and its vomeronasal capsule is exclusively cartilaginous [283]. In hystricomorphic rodents, the long-tailed chinchilla (Chinchilla lanigera) showed considerable variation in the primary features of its VNO. Its sensory epithelium shifts from a medial location at the front of the VNO to an entirely dorsal position at the back. Similarly, the vomeronasal capsule in this species consists of cartilaginous tissue at its anterior level, but this shifts dorsally as we move caudally, eventually being completely replaced by a bone layer that encases the VNO at its back [284].
Concerning the vascular pump, the VNO of the chinchilla has several significant-diameter veins, but features a dominant central vein [285]. Similarly, within the hystricomorphs, African mole rats displayed unique characteristics concerning their vomeronasal organ. In these eusocial rodents, there is hardly any postnatal growth in their vomeronasal neuroepithelium [286]; however, all the components of their VNO exhibit significant and functional development. Thus, it has a sensory epithelium of substantial thickness, and laterally displays large venous sinuses. Transversely, the vomeronasal duct takes on the typical “J” or crescent shape. The parenchyma is protected by a cartilaginous capsule and is also reinforced by an external bone layer that occupies a ventral and/or lateral position depending on the species [287].
As with rodents, lagomorphs have a extensively developed vomeronasal organ (Figure 11). The rabbit (Oryctolagus cuniculus) has a double vomeronasal capsule, made up of an external bone covering and an internal cartilaginous lining [288,289]. Its sensory epithelium is comparably thick to that of rodents, but its vascular pump comprises multiple large-caliber veins that provide a powerful suction capability to the VNO in this species [143,290].
Marsupials also display a remarkable development of the vomeronasal organ, suggesting that, as in rodents and lagomorphs, pheromonal communication plays an essential role in these mammals. The Tammar wallaby (Notamacropus eugenii), a macropodid of the order Diprotodontia, sports a complete cartilaginous capsule and a densely populated and extensive neuroreceptor epithelium. The vascular tissue enabling the pumping mechanism consists of numerous large-caliber veins, which are mostly distributed on the medial side, but voluminous veins also appear on the medial face of the vomeronasal duct [291]. In the order Didelphimorphia, the gray short-tailed opossum (Monodelphis domestica) presents certain unique features. Its vomeronasal capsule is cartilaginous, although at the caudal level, it is replaced by a bony capsule, which encircles a parenchyma very rich in glandular tissue. Its vascular pump is made up of a large main vessel which are located laterally to the vomeronasal duct, and other smaller vessels in the ventromedial and lateral areas. Its sensory epithelium is very thick and is characterized by a rosette-shaped structure that divides said epithelium in half at the caudal part of the organ. This configuration corresponds to the fusion of the main vomeronasal glandular secretion duct with the neuroepithelium, through its opening into the lumen of the vomeronasal duct [292].
In species of the order Monotremata, the VNO shows notable development. Studies on newborn individuals of the platypus and echidna determined that both possess a thick sensory epithelium and a cartilaginous capsule. In contrast, both showed few veins of modest caliber forming their vascular pump [293,294].
Within the order Eulipotyphla, the studied species display a highly developed VNO. The African pygmy hedgehog (Atelerix albiventris) has a cartilaginous vomeronasal capsule and a neuroreceptor epithelium of moderate thickness, which extends beyond the usual boundaries, occupying almost the entirety of the lumen in front of the respiratory epithelium. The parenchyma contains mucous and serous glands, and a network of venous sinuses that extend around the entire vomeronasal duct. Through a cross-section, the lumen of its VNO shows a circular shape at the front, which becomes more oval as we move caudally [295]. On the other hand, the VNO of the dasyurid marsupial Antechinus subtropicus displayed significant differences compared to the African pygmy hedgehog. In this species, the capsule is also cartilaginous, but it features a primary vein lateral to the vomeronasal duct, along with other secondary veins located both laterally and medially to it, forming its vascular pump. Additionally, its vomeronasal neuroepithelium has a notably greater thickness compared to the African pygmy hedgehog; however, its extension in front of the respiratory epithelium corresponds with the usual pattern, and its lumen shape, when observed in cross-section, is the typical "J" or crescent shape [296].
Mammals of the order Carnivora, both felines and canines, have shown moderate development of their VNO (Figure 12). Studies on the dog [260,297] domestic cat (Felis silvestris catus) [298], brown bear (Ursus arctos) [299], and European ferret (Mustela putorius) [300] revealed that all share the main structural characteristics of the VNO, such as a cartilaginous capsule, the distribution of vessels around the vomeronasal duct forming the vascular pump, and a broad sensory neuroepithelium, albeit thinner than mammals of the orders previously described. However, the red fox (Vulpes vulpes) represents an exception among domestic canines. In this species, the vomeronasal epithelium is not only more developed but also displays the expression of G proteins associated with both V1R and V2R receptors [127].
Artiodactyls exhibit a high level of VNO development. Their cartilaginous capsule is complex and developed, displaying numerous morphological differences between species. The length of the sensory vomeronasal epithelium tends to be longer compared to species of the aforementioned orders; however, its thickness is notably inferior, as seen in carnivores. Still, it maintains the three cellular layers that make up the neuroepithelium, which are organized with a clearly defined boundary. Vessels in the vomeronasal parenchyma are numerous, of medium caliber, and are uniformly arranged around the vomeronasal duct [301]. Regarding the morphology that the vomeronasal duct exhibits in cross-section, artiodactyls usually have an oval shape, unlike the usual crescent or "J" shape. Variations also exist between different species like cow [145,302], sheep [303], goat [304,305], deer [306], moose [307], giraffe [308], and duike [282].
Strepsirrhine primates, which are mostly nocturnal, have a highly developed VNO compared to haplorhine primates, who tend to be diurnal and have a modestly developed VNO. Both have a cartilaginous capsule surrounding the vomeronasal parenchyma; however, the thickness of the vomeronasal epithelium is significantly greater in strepsirrhines. Similarly, the vascular pump in strepsirrhines has numerous large-caliber vessels around the entire vomeronasal duct, in contrast to the few vessels typically displayed by the VNO of haplorhine primates [309].
Among the various bats, the development of the VNO is extremely variable. From species that show a total absence of their VNO even in the embryonic phase, to other species that demonstrate astonishing development of this sensitive structure. Likewise, many other bat groups represent intermediate levels of development [219].
2.2. Neuroanatomy of the Accessory Olfactory Bulb
The accessory olfactory bulb (AOB), the first neuronal integration center of the vomeronasal sensory system, is located in the olfactory areas of the forebrain of certain vertebrate species; what we know as the basal rhinencephalon. In mammals, it is usually located in a dorsocaudal position relative to the main olfactory bulb (MOB) [310].
Macroscopically, in most species, the AOB is a relatively difficult structure to discern. Although in most cases it gives rise to a convex prominence in the dorsal transition between the main olfactory bulb and the olfactory tract, its boundaries with these structures are very diffuse. However, histologically, it is easily recognizable due to its laminar morphology. Although this laminar pattern is essentially analogous to that of the MOB, there are significant variations between different groups of mammals. Some species have clearly defined lamination and high cellularity, while others have much less defined layer boundaries (Figure 13 and Figure 14). Thus, from superficial to deep, the AOB layers are:
1. Vomeronasal nerve layer: It consists of bundles of unmyelinated fibers, originating from vomeronasal receptor neurons, surrounded by numerous glial cells, both astrocytic and ensheathing glia [311,312]. Whereas in the MOB of mammals, each olfactory axon projects to a single glomerulus [139,313], in the AOB, fibers branch out and distribute to more than one glomerulus [314].
2. Glomerular layer: Comprised of spherical structures called glomeruli, these result from synaptic contact between axonal terminations of vomeronasal nerve fibers and the apical dendrites of mitral cells. These glomeruli show a relatively acellular texture rich in neuropil but are bounded by a narrow band of periglomerular cells, especially in their deepest part. Their degree of definition is lower than that observed in the main olfactory bulb.
3. External plexiform / Mitral / Internal plexiform: It is at the level of the three central layers of the accessory olfactory bulb where the greatest differences lie, both compared to the MOB and in the interspecific comparisons that can be made at the level of the AOB itself. The differentiation of an external plexiform layer -formed by the dendrites of the mitral cells and granular cells, as well as by tufted cells and other neuronal types -, a mitral layer - formed by a linear band that includes the somas of the mitral cells, the second neuron of the olfactory pathway - and an internal plexiform layer - containing the axons of the mitral cells -, is constant in the organization of the MOB in mammals. However, in the case of the AOB, the existence of plexiform layers has only been proposed in those species where the degree of lamination is highest, mainly rodents, but the issue is controversial. Thus, Cajal [194] concluded that the plexiform layers were absent; however, classic comparative studies of the olfactory system [315,316] included the external plexiform in their description of the AOB but not the internal plexiform. Meisami and Bhatnagar [310] in their exhaustive bibliographic review state that it is inappropriate to extrapolate data from one species to another, indicating that in mammals with large and well-developed AOBs, both plexiform layers exist, although recognizing that the boundaries of these layers are not recognizable with Nissl staining, requiring more specific stains such as cytochrome oxidase or Galyas staining. However, Salazar et al. [239], in their exhaustive study of the mouse AOB, concluded the impossibility of determining the presence of plexiform layers, as well as differentiating mitral cells from tufted cells, coining the term mitral/tufted layer. In their subsequent study of the cat AOB, Salazar & Sánchez-Quinteiro [317] opted for the term mitral/tufted/plexiform layer which more accurately encompasses the fusion of the three AOB structures equivalent to the three inner layers of the MOB. In later works, this group leaned towards a simpler name: mitral/plexiform layer, which we adopted throughout this work. It is important to clarify that the morphology of the main cells of this mitral/plexiform layer rarely corresponds to the typically mitral morphology observed in the mitral layer of the MOB. This has led certain authors to prefer to avoid the term mitral when describing this layer, as is the case with Larriva-Sahd [318] and Villamayor et al. [132] in their respective studies of the AOB in rats and rabbits. In both cases, they opted for the terms "outer cell layer" to designate the set of the mitral/plexiform layer and "principal cells" to designate the projection neurons that constitute this layer. While undoubtedly a more accurate name in terms of morphological reality, the direct functional equivalence between the mitral cells of the MOB and the principal cells of the AOB (projection neurons in both cases) advises maintaining the term mitral cells to designate the principal cells of the AOB. Regarding the use of the term plexiform, Larriva-Sahd [318] considers that, in contrast to the plexus arrangement of the proximal processes of the MOB mitral cells, in the case of the AOB these processes adopt a radial arrangement, resulting in a dense and convoluted dendritic frame, very different from the parallel fibrous texture that forms the plexiform layer of the MOB. The set of radial dendritic arborizations, together with the proximal processes of granular cells and other interneurons, make up a neuropil rather than a plexiform layer. Another interesting aspect that affects the stratification of the AOB is the topography of the lateral olfactory tract (LOT), which can be located either through or below the accessory bulb. Thus, in broad groups of mammals including rodents, insectivores, and primates, the broad axonal bundles that make up the dorsal component of the LOT run through the innermost area of the mitral/plexiform layer or outer cell layer [319].
4. Granule cell layer: It is composed of granular neurons, which lack axons and accumulate compactly. These cells are GABAergic and presumably inhibitory [320]. However, they are not a homogeneous population in terms of the expression of peptidic neuromodulators [321]. Their basal dendrites receive centrifugal impulses from the amygdala, whereas their apical dendrites interact with the basal dendrites of the mitral cells [322]. On the other hand, it has been observed that the number of granular cells is lower in domestic animals and animals kept in zoological facilities [323]. Furthermore, neuronal proliferation has been found in the AOB of adult rats, with most of these new cells being located in the granular layer [324]. Contrary to what one might assume, most of the proliferating neurons present in the OB do not derive from the subventricular zone of the olfactory bulb but from slow-dividing cells that might correspond to the population of resident neural stem cells. These could generate neurons that are incorporated into the OB circuits in vivo [325].
Projection neurons of the olfactory bulb send axons through the lateral olfactory tract (TOL), giving it a laminar structure. Thus, in embryonic stages, the TOL consists of three layers: superficial, middle, and deep. The axons from the BOA are the first to mature, so they are located in the deep layer, whereas the axons from the MOB constitute the middle and superficial strata. The middle layer is composed of mature axons, while the superficial layer consists of newly generated axons that are added, forming a series of stacked axonal laminae at different maturation stages. Initially, the superficial lamina is proportionally larger than the middle lamina, but throughout development, the middle lamina increases, while the superficial lamina drastically decreases [326]. This segregation of both pathways in the TOL, the vomeronasal and olfactory pathways, can be seen in adult animals like rabbits or rats [327,328].
Throughout all the layers that make up the BOA, different cell types coexist. From superficial to deep, we can find periglomerular cells, superficial short-axon neurons, mitral cells, round projection cells, tufted cells, external granular cells, dwarf cells, polygonal neurons, and internal granular cells.
Periglomerular cells (PG) are small short-axon neurons found at the base of the glomeruli in both the AOB and the BOP, although they are more numerous in the latter. In the AOB, these cells are primarily GABAergic and have inhibitory functions, possibly playing a modulatory role [329]. There are two classes of PG: amacrine cells, meaning they lack axons, and interneurons. Both have a small fusiform soma, from which they send one or two dendrites to one or several glomeruli. In them, their dendrites branch and originate a dense dendritic plexus. On the other hand, the axon of the interneurons distributes within the neuropil adjacent to the glomeruli.
Superficial short-axon cells are morphologically similar to PG but have some differences. They are larger in size and are located superficially in the external plexiform layer. They have a thick main dendrite that rises to enter a single glomerulus, where it establishes a dendritic network. They also emit other dendrites that distribute ventrally between the somas of PG and other superficial short-axon cells. Their axon interacts with the somas of adjacent PG [311].
Mitral cells, also called principal cells, are the second neuron in the olfactory and vomeronasal sensory pathways. In the BOP, they are distributed linearly, forming a dense layer of mitral cells. In the AOB, this layer transforms into a diffuse mitral cell band integrated with both plexiform layers, constituting a single mitral-plexiform layer, also called the external cell layer. The cytoarchitecture of the accessory olfactory bulb has been addressed in depth in the rat through the study carried out with the Golgi technique by Larriva-Sahd [318]. This study shows that mitral cells have two types of dendrites: glomerular dendrites (thick) and accessory dendrites (thin). Glomerular dendrites are multiple and extend to the glomeruli located in their respective half of the AOB, according to the anteroposterior axis [330]. The shape of the soma of the mitral cells depends on the number of glomerular dendrites it comprises, so its morphology will be oval, triangular, or polyhedral, presenting two, three, or more primary dendrites, respectively. The axons of these neurons leave the AOB caudally [318].
Another cell type described in rats are the round projection cells, which have an oval shape and paired dendrites [318]. Their axon emerges from the base of a proximal dendrite and extends towards the lateral olfactory tract. Also, from their axon emerge one or two extensions that flow into the soma or dendritic processes of a neighboring cell.
Tufted cells are triangular in shape, although they can have a fusiform morphology. Their size is slightly smaller compared to mitral cells, but they have a similar general organization of their dendrites. However, tufted cells have a single glomerular dendrite that attaches to a single glomerulus. In these cells, the axon usually arises from the base of a non-glomerular dendrite, although it occasionally originates directly from the soma. Then, the axon descends ventrally, traveling a broad zigzag path to end near dwarf cells, external granular cells, or mitral cells.
In the granular layer, closer to the surface, we find the external granular cells, neurons with round or elliptical somas that have two or three dendrites. The primary dendrites tend to be short, while the secondary branches are longer and branch at their ends. Additionally, their dendrites show the presence of numerous dendritic gemmules. These cells lack an axon, so they are included in the category of amacrine cells. They sometimes integrate into the TOL fibers [318].
In the deeper part of the granular layer are the internal granular cells, which have round or triangular somas and also lack an axon [331]. Their dendritic processes consist of a thick ascending dendrite and one or two sets of short, thin branches. Like the external granular cells, they have gemmules on their ascending dendrites. Internal granular cells communicate with the dendrites of tufted cells and mitral cells but also with the somas of certain periglomerular cells and short axon surface neurons [318,332].
In a more internal layer of the granular layer, we find the dwarf cells and the polygonal neurons. Dwarf cells are interneurons without axons that have a very small, spherical soma, and their dendritic tree lacks gemmules [318]. Polygonal neurons are small fusiform bipolar neurons that have paired dendrites. Their axons branch out, extending through the neuropil, interacting with external granular cells and dwarf cells.
Moreover, in the accessory olfactory bulb, different types of glial cells can be found. The radial glia cells, like the olfactory envelope glial cells, are the most common, but astrocytes and oligodendrocytes are also observed [333]
Regarding the basic circuit that sensory information follows in the AOB, there are functionally three main neuronal components that are activated sequentially through vomeronasal stimulation: the axons of the vomeronasal sensory neurons, the mitral cells, and the granular cells [334]. The axons of the sensory vomeronasal neurons, present in the apical layer of the vomeronasal epithelium (V1R), are distributed in the anterior zone of the AOB [335], while the sensory vomeronasal neurons of the basal layer (V2R) project their axons to the caudal half of it [172]. On the other hand, although mitral cells receive information from several glomeruli, they connect only with axons of sensory vomeronasal neurons of the same type (V1R or V2R). Thus, a convergence of specific sensory inputs occurs in a small population of mitral cells. In the MOB, a much stricter convergence pattern occurs since the information received by each mitral cell comes from a single glomerulus [314].
Morphologically, the AOB shows significant differences among the multiple species of mammals. In those animal groups where this structure is most developed, clear lamination can be identified. However, as we have mentioned, this generally does not allow differentiation of the external plexiform, mitral, and internal plexiform layers, these forming a broad layer (mitral plexiform/external cellular), consisting of projection cells (mitral/main) distributed within a neuropil. On the other hand, species with a less sophisticated AOB have a very reduced mitral/plexiform layer in which it even becomes difficult to discriminate the projection cells.
The case of the platypus (Ornithorhynchus anatinus) is unique among mammals, as in this species the AOB and MOB are of similar size. Likewise, both the platypus and the echidna (Tachyglossus aculeatus), both monotremes, have well-differentiated layers forming the laminar structure of the AOB [294].
Rodents also possess a AOB with a high degree of development; however, as has been described, both the rat [318,336] and the mouse [239,337] display poorly differentiated boundaries between the external plexiform, mitral, and internal plexiform layers. Nevertheless, both murine species have a thick mitral-plexiform layer containing a dense network of mitral and tufted cells. In the case of lagomorphs, the AOB has extensive development, and specifically, the rabbit (Oryctolagus cuniculus) exhibits a laminar configuration highlighting a broad mitral-plexiform layer formed by three main types of cells: large, tufted, and rounded cells [132].
Among the different species of bats, there is significant variability in the development of their AOB. Generally, they present only four distinguishable layers; however, in some cases, such as Glossophaga soricina, Frahm & Bhatnagar [338] suggested the presence of the plexiform layers (internal and external) and the mitral cell layer. The rest of the studied mammalian orders show the plexiform layers and the mitral layer merged into a single mitral-plexiform layer. Marsupials possess a sophisticated AOB; however, their lamination is restricted to four layers. Despite this, their mitral-plexiform layer has a considerable thickness. These characteristics have been observed in the gray short-tailed opossum (Monodelphis domestica) [339] and in the Tammar wallaby (Notamacropus eugenii) [256]
Carnivores generally display a surprisingly moderate development of their AOB, with domestic ones being the most studied (Figure 15). There are notable differences between felids and canids in this regard. Although both exhibit four layers in their lamination, felids show a more pronounced development of the mitral-plexiform layer [260,317]. However, in the red fox (Vulpes vulpes), a higher degree of AOB differentiation than in dogs has been observed, exhibiting a thick glomerular layer. Additionally, in the latter species, the imaginary lines separating different strata follow irregular trajectories, to the extent that the mitral-plexiform layer shows prominent extensions into the glomerular layer [340]. This raises the hypothesis of a possible involution of the vomeronasal system as a consequence of selection pressure and crossbreeding associated with the domestication of dogs. It is noteworthy how differences in the organization of the olfactory bulb between domestic and wild canids are not so pronounced at the level of the main olfactory bulb. However, recent neuroanatomical observations have identified subtle morphological and neurochemical differences between the MOB of wolves and foxes and the MOB of dogs [341]. Despite this fact, there is a growing interest in exploring the clinical implications of the VNO in domestic dogs. This has been bolstered by the optimization of an MRI protocol for in vivo visualization of the VNO [342], and the recent identification of a case of canine vomeronasal agenesis, which manifested significant behavioral disorders, such as an inability for sexual discrimination of conspecifics and reduced sexual behavior [343]. Moreover, the use of pheromonotherapy, like the application of dog appeasing pheromone (DAP) to mitigate symptoms of separation anxiety, including house soiling, vocalizations, and damage [344,345], along with the deepening in the links between VNO inflammation and aggressive behaviors in canids, felids, and livestock [346,347,348,349], has further developed this interest in the dog VNS.
Within the carnivores, the Mustelidae family has been the subject of extensive study, as the characterization of its AOB remains unresolved. Whereas the mink (Mustela vison) shows a well-developed VNO [350], the characterization of its AOB has proved challenging. Despite efforts, a precise and consistent morphological definition has not been possible using traditional neuroanatomical techniques [351]. However, the AOB has been studied more successfully in the ferret using immunohistochemical markers and lectins, which have revealed a poorly developed AOB [300].
The studied artiodactyls species such as the sheep (Ovis orientalis aries), the Siberian roe deer (Capreolus pygargus), or the common hippopotamus (Hippopotamus amphibius) have a moderate development of the AOB (Figure 16), higher than carnivores, as they also exhibit four distinct layers forming their laminar organization [299,306,352] .
Regarding the neurochemical study of the AOB, the use of Gαi2 and Gαo proteins has garnered much attention, as both proteins indicate the expression of vomeronasal receptor families V1R and V2R, respectively, establishing an anteroposterior zonation in species that exhibit both receptors. This delineates the AOB organization into two areas: an anterior region rich in Gαi2 and a posterior region rich in Gαo [169]. Thus, the analysis of both G proteins allows assessment if the studied species fits within a uniform model, or conversely, exhibits a segregated pattern. For the Gαi2 marker, labeling is restricted to the neural and glomerular layers of the AOB, while the Gαo marker is expressed in both the neural and glomerular layers as well as the other strata of the AOB [353].
On the other hand, various immunohistochemical markers have been utilized in AOB studies. Calcium-binding proteins such as calbindin (CB), calretinin (CR), parvalbumin (PV), neurocalcin (NC), and secretagogin are typically expressed in certain layers and cell populations, both in the AOB and BOP [339,354,355]. Additionally, CR labels mitral cells and can be expressed in atypical BOP glomeruli, typically located near the AOB [356]. The glial fibrillary acidic protein (GFAP) is used to generally identify the glial component of the AOB, and specifically astrocytes and ensheathing cells [357]. The microtubule-associated protein 2 (MAP2) is primarily expressed in the dendrites of mitral cells [358], leading to intense labeling in the external plexiform and mitral layers, but can also be expressed in the internal plexiform and granular layers. The growth-associated protein 43 (GAP-43), also termed plasticity protein, is expressed in neuronal growth cones during development [359]. In the AOB, its immunolabeling is restricted to the neural, glomerular, and granular layers [132]. The olfactory marker protein (OMP) is involved in signal transduction and is used to evaluate neuronal maturity [360] of the olfactory system. In the AOB the expression of this marker is confined to the vomeronasal nerve and glomerular layers. Various markers in the OB are also employed to identify neuronal populations expressing different types of neuronal receptors, such as GABA receptors, dopaminergic receptors [361], cholinergic receptors [362], adrenergic receptors [363], and serotonergic receptors [364]. Additionally, immunohistochemical studies of various neuropeptides in the AOB, such as substance P, cholecystokinin, or neurotensin [365], and neuronal markers like PGP9.5 [366] or doublecortin (DCX) [367] are commonly conducted.
Another technique commonly used in the characterization of the AOB is histochemical labeling with lectins. The labeling pattern produced by each lectin may vary between the different species studied, although the Ulex europaeus agglutinin (UEA), Bandeiraea simplicifolia isolectin B4 (BSI-B4), and Lycopersicon esculentum agglutinin (LEA) are regularly expressed in the nervous and glomerular layers. Specifically, UEA allows differentiation of the anteroposterior zonation of the AOB in species of the segregated model, as it is expressed more intensely in the anterior region (VR1) [147]. In contrast, Vicia villosa agglutinin (VVA) shows more intense labeling in the posterior part of the AOB [368]. In this way, different neuronal populations can be identified. Many other lectins are also used in the study of the AOB, such as soybean lectin (SBA), wheat germ lectin (WGA), and Dolichos biflorus agglutinin (DBA) [369,370].
2.3. Olfactory Pathways
Regarding the vomeronasal information flow, vomeronasal sensory neurons send the sensory information received from the vomeronasal receptor epithelium to the mitral cells of the AOB [371]. In contrast, olfactory information is captured in the main olfactory epithelium and later travels through the olfactory nerves to the MOB. Then, these stimuli are transmitted through various projections to different brain areas.
The application of neuronal tracer studies to the olfactory pathways allowed the discrimination of the existence of segregated and parallel projections from the main and accessory olfactory bulbs [372,373]. This was known as the dual olfactory system hypothesis, a formulation demonstrated for the first time using the Fink-Heimer technique in rabbits [374]. Although this hypothesis is still currently valid, it has been refined over the past decades, especially regarding the study of secondary and tertiary projections, proving the existence of neuronal communication between both pathways.
The efferences of the MOB project ipsilaterally to the anterior olfactory nucleus, tenia tecta, olfactory tubercle, piriform cortex, lateral and medial areas of the entorhinal cortex, and lateral amygdaloid nuclei [375,376]. Among the tertiary projections of the pathway, the communication of the entorhinal cortex with the hippocampus and the piriform cortex and lateral amygdala with the hypothalamus and mediodorsal thalamic nucleus stands out [377,378]. It has recently been shown that there is a direct projection of olfactory sensory information to the anterior subdivision of the medial amygdala, a structure traditionally involved in mediating pheromonal information. This supports the hypothesis that both olfactory systems act collaboratively and not differentially in the control of socio-sexual and anti-predator behaviors [379].
Unlike the main olfactory system, efferent projections from the AOB bypass the thalamocortical axis, distributing ipsilaterally to third-order limbic system nuclei, such as the accessory olfactory tract nucleus, the terminal stria nucleus, and the medial and posteromedial cortical amygdaloid nuclei (together forming the vomeronasal amygdala) [373,380,381,382,383]. The medial amygdala is strongly interconnected with other structures that receive vomeronasal information from the AOB. This suggests that the information detected by the vomeronasal system undergoes complex intrinsic processing before being transmitted to other structures [384]. The final processing center between vomeronasal information and effectors is the hypothalamus [385]. In this way, the VNS is directly involved in the activity of sex hormones and can facilitate the development of aggressive, defensive, or reproductive behaviors [386]. It is important to note that olfactory and vomeronasal projections show some degree of overlap in the amygdaloid cortex and medial amygdala, suggesting that there are anatomical pathways that allow extensive integration of olfactory and vomeronasal information [152,384,387].
Additionally, the MOB and AOB differ in their centrifugal afferent connections [388]. The MOB receives massive cholinergic and GABAergic projections from the basal forebrain, which mainly originate in the nucleus of the horizontal branch of Broca diagonal band and the magnocellular preoptic nucleus [389]. Cholinergic signaling within the OB modulates olfactory learning and memory, odor discrimination, odor habituation, and social interactions [390]. From a comparative neuroanatomy perspective, the study by Liberia et al. [391] indicates that the synaptic connectivity of the afferent cholinergic circuits is highly conserved in the OB of macrosmatic and microsmatic mammals. Regarding the GABAergic afferents of the basal forebrain, they innervate all the layers of the MOB at least as densely as cholinergic axons, but only a few studies have examined their function in odor processing [392].
The AOB receives significant afferents from a wide range of nerve centers. The bed nucleus of the stria terminalis and the VN amygdala project reciprocally to the AOB, thus forming a feedback circuit [393]. Tracer studies have revealed that both feedback projections to the AOB are topographically organized and use different neurotransmitters [394]. Specifically, GABAergic projections from the bed nucleus terminate in the outer cell layer, while glutamatergic projections from the amygdala are directed to the inner layer of granule cells. A significant number of these feedback neurons in both areas express estrogen receptors ER-α, linking the animal endocrine state with integration in the AOB. The relevance of this pathway was subsequently confirmed from different morphofunctional approaches [395,396].
Other important afferents for the function of the AOB come from noradrenergic structures of the brainstem, which play a crucial role in the formation of olfactory memory [397]. This comes into play, for example, during mating, in which stimulation of the vaginocervical zone leads to sustained increases in the levels of noradrenaline in the AOB, which persist for about 4 hours [398]. This time span establishes a crucial phase in which noradrenaline generates plastic changes in the intensity of dendrodendritic synaptic connections [399]. Additionally, the diagonal band of Broca is the origin of numerous cholinergic fibers involved in increasing the excitability of granule cells [400], while from the raphe, abundant serotonergic fibers are sent to both olfactory bulbs that act similarly to the cholinergic ones [364].
In summary, the olfactory and vomeronasal systems represent two separate systems but functionally interconnected, in charge of processing olfactory and pheromonal stimuli, respectively. These systems differ in their afferent and efferent connections, morphology, physiology, and functional implications, but both contribute to the complex sensory processing of olfactory and pheromonal information that leads to behavior (Wilson & Mainen, 2006). The increasing evidence suggests the existence of a more complex and intricate relationship between these systems than previously assumed. Recognizing and understanding the complexities of this interaction will provide a clearer picture of how odors and pheromones shape animal behavior.
3. Specific study of the Vomeronasal System
The study of the morphological features of the VNS has been utilized as a phylogenetic classification method at both neuroanatomical [144] and genetic levels [402]. It is also indispensable for fully understanding the physiology, evolution, and functional implications of the VNS in mammals. For this reason, to conclude this review, we have included a final section where, based on our group observations, we exemplify in four mammal species from evolutionarily distinct families the vast morphofunctional differences among different mammal groups. This not only pertains to the organization of the vomeronasal organ and the accessory olfactory bulb but also reveals how each species has adapted to various environments, social and reproductive behaviors, feeding habits, and predators, leading to different configurations of the system.
The species selected for this section of the review are the capybara (Hydrochoerus hydrochaeris), a hystricomorph rodent; the Bennett's wallaby (Notamacropus rufogriseus), a macropod diprotodont; the meerkat (Suricata suricatta), a herpestid carnivore; and the dama gazelle (Nanger dama), an artiodactyl from the bovid family.
3.1. Capybara (Hydrochoerus Hydrochaeris)
There is limited information available on the vomeronasal system (VNS) of rodents, beyond the extensively studied laboratory rodents: rat (Rattus norvegicus domestica) and mouse (Mus musculus). There is scant data regarding wild rodents. The interest in comparative analysis with wild rodents stems from the fact that most studies on the VNS have been conducted on laboratory rodent strains, exposed to artificial selection that does not reflect the selection pressures in the wild. Consequently, these laboratory strains exhibit significant genetic and behavioral differences compared to wild rodent models [403]. Moreover, these species can have significant disparities with wild rodents due to the domestication process, as seen in canids, where artificial selection on domestic dogs presumably plays a pivotal role in the regression of their VNS [340]. Lastly, the capybara is a semi-aquatic mammal, and studying it helps expand the limited neuroanatomical information on the VNS in such species (Figure 17). This characterization is vital since semi-aquatic habits could influence the organization and functioning of chemosensory systems [404].
A unique feature of the capybara that adds particular interest to its study is the degree of development its VNS reaches in newborn individuals. While studies like the VNS examination of the Asian elephant (Elephas maximus) showed that at 6 weeks of age, the VNS already has morphological similarities with adults [405], many studies detail the VNS development throughout the embryonic process without clarifying the maturity and functionality level of the VNS at birth [406] – a time that implies numerous transformations involving chemical communication. Hence, the capybara constitutes an excellent model for studying chemical communication development in precocial species.
The expression of G protein subunits in the adult capybara VNS is relevant for understanding the VNS role in this early life stage. The capybara aligns with other studied hystricomorphs like chinchillas, guinea pigs, or degus in having a clear anteroposterior zoning in the AOB [257,284,407]. However, in these species, unlike laboratory rodents, no apical-basal zoning has been found at the VNO level. It is striking that other non-rodent species with a segregated model, like rabbits or wallabies, also lack this apical-basal zoning, suggesting this is a highly specific trait for laboratory rodents [408]. The immunopositivity of both G proteins throughout the neuroepithelium of the VNO, the vomeronasal nerves, and the neural and glomerular layers of the newborn capybara AOB indicates the VNS full maturity at birth [409]. A similar early prenatal development pattern in the VNS of other precocial mammals, like pigs [410,411,412] and sheep [352], was found, but it was determined during the fetal period, and in neither case was the expression of G proteins in the VNS evaluated. Notably, the anteroposterior zoning is also evidenced with the UEA lectin, which stains the anterior area more intensely. Although this pattern aligns with findings in mice, a recent study on UEA lectin staining in the mouse olfactory system could only identify this zoning in three of the sixteen subjects studied [221]. The authors concluded that these individual differences might be caused by the presence or absence of signals activating the receptor cells or reflect the difference between wild and laboratory rodents. The presence of anteroposterior zoning marked by UEA in capybaras appears to support this theory.
Another differential aspect is that the newborn capybara exhibits a bias towards a more prominent caudal AOB. This characteristic was previously described in adult individuals [413]. These results, combined with recent observations on the morphometric parameters of the AOB in two degu species with contrasting social habits, suggest that some AOB structural features are tied to the species lifestyle and arise during an early ontogeny stage [414].
The early functionality of the VNO and AOB in capybara was confirmed in the VNO, where calcium-binding protein markers, calretinin, and calbindin stained the nervous bundles of the parenchyma and almost all sensory neuroepithelial cells, although different cell populations were evidenced. Something similar happens in shrews, where calretinin produces intense staining in almost all receptor cells and vomeronasal nerves of prenatal individuals [415]. Likewise, the robust expression of GFAP in astrocytes from the neural and glomerular layers of the capybara AOB contrasts with the reduced astrocytic development observed at this early stage in other species, like newborn opossums [416]. The expression of olfactory marker protein (OMP) is, however, very reduced in this species AOB, especially when compared to the MOB. This has been observed in rodents like mice or rats [248] and in marsupials like opossums [360].
In addition to the cited neurochemical aspects, the capybara has exclusive morphological features, such as the nature of the capsule protecting both VNOs, the dorsal location of the VNO in the nasal cavity over the incisive bone palatal process, the high degree of morphological differentiation of the AOB at this early stage, and finally, the presence of a migratory stream from the VNO neuroepithelium towards the vomeronasal nerves.
The dorsal location of both VNOs is a result of them resting on the palatine process of the incisive bone, and not on the vomer bone, as is the case in most species [409]. This topography is only comparable to that described in rabbits [143], where the organs also rest on the palatine process, although in this case, the bony projection is smaller, resulting in a lesser dorsal displacement of both VNOs. While in most species the nature of the vomeronasal capsule is exclusively cartilaginous, there are species such as the mouse where the capsule is formed by a thin bony lamina [417] or like lagomorphs that have a cartilaginous capsule externally reinforced by a bony envelope [143]. In the capybara, the capsule is initially cartilaginous, but from the central area of the organ it is progressively replaced from ventral to dorsal by a bony lamina that ends up completely enveloping the VNO parenchyma in its caudal zone [409].
The secretion from the vomeronasal glands into the vomeronasal duct plays a crucial role in vomeronasal peri-receptor processes [242]. The capybara VNO shows significant development of glandular tissue, especially at the caudal end of the organ [409]. However, in the central part of the duct, the presence of glands is moderate. This pattern is analogous to that found in other rodents such as rats, guinea pigs [418], or mice [241]. Likewise, the glandular nature of the capybara VNO is both PAS+ and AA+, contrasting with most of the studied rodents, which only express positivity for PAS in glandular secretion [284,419,420]. This notable variation in gland characteristics within the same order might be due to the capybara adaptation to its semi-aquatic nature, which could require a specific pheromone-receptor interaction environment.
A unique point, for which, to our knowledge, there is no other example in the literature, is the profuse cellular migration that occurs from the sensory epithelium of the VNO to the nerve bundles of the vomeronasal nerves (Figure 18) [409]. This cellular migration is immunolabelled with anti-GAP-43 and histochemically labelled with UEA and LEA lectins but immunonegative when labelled with anti-LHRH. This indicates that the migration is not analogous to that described in prenatal rat and mouse fetuses, which occurs from the vomeronasal part of the olfactory placode to the hypothalamus and is mainly composed of LHRH positive cells [421,422,423,424]. It is challenging to hypothesize the importance of these cells in the capybara, as this discovery is unprecedented in both olfactory and vomeronasal nerves.
Regarding the morphology of the capybara AOB (Figure 19), it presents a distinctly differentiated laminar pattern [409], similar to that found in other rodents [417], lagomorphs [132], marsupials [425], and prosimians [426]. However, among all these cases, the capybara is the species that has a more distinctly organized mitral layer.
3.2. Bennett’s Wallaby (Notamacropus Rufogriseus)
The extensive study of the vomeronasal system of Bennett's wallaby (Notamacropus eugenii) conducted by Schneider et al. [256,291,293,427] proposed a new and surprising processing model in the VNS of mammals. Specifically, the investigation into the presence of the G protein subunits, Gαi2 and Gαo -associated with the vomeronasal receptor families V1R and V2R, respectively [428]- concluded that the Tammar wallaby only expresses V2R receptors, and therefore, does not fit into either of the two existing models: the uniform model (species that express V1R receptors: Soricidae, Hyracoidea, Perissodactyla, Artiodactyla, Carnivora or Primates; [429]) and the segregated model (species that express both V1R and V2R receptors: Rodentia, Lagomorpha, Afrosoricida and Didelphimorphia;[220]).
Aware of the disruption that Schneider et al. [256] findings pose to the established knowledge on vomeronasal transduction, the authors of the study cautioned that their results should be taken with care. In fact, the observation that the anti-Gαo protein was only expressed in a subpopulation of neuroreceptor cells of the vomeronasal epithelium and not in the entire population of vomeronasal neurons was somewhat atypical, as in the previously described uniform model, all vomeronasal neurons are marked with Gαi2. To explain this low number of Gαo-positive cells, the authors hypothesized a reduced affinity of the antibody used. However, there are no other examples in the literature of any similar selectivity deficit in response to the use of anti-Gαo antibodies.
More critically, the immunolabeling of the AOB in the Tammar wallaby with the Gαo marker has only been carried out in this study [256], so there are no other references that support or refute the results. The staining protocol followed by these authors resulted in a very weak labeling pattern. It was also restricted to a small area in the superficial layers of the AOB, where projections from the vomeronasal neuroreceptor cells are received. In contrast, in all other studied species, both from the segregated and the uniform models, the Gαo and Gαi2 markers are expressed throughout the thickness of the superficial layers. This includes the neural and glomerular layers. Regrettably, the authors did not specify the antibody used that showed a negative pattern to the Gαi2 marker both in the VNO and the AOB, without providing information about the commercial source or the batch number of the antibody, preventing any comparison with other studies on the expression of the Gαi2 protein in the VNS that used the same antibody.
The existence of this third model could only be contrasted with the study of the VNS of Bennett's wallaby (Notamacropus rufogriseus) conducted by Torres et al. [430]. The immunohistochemical study of the G proteins in the VNS of Bennett's wallaby revealed a canonical labeling pattern for both Gαi2 and Gαo in the VNO and AOB of all the samples studied. The labeling pattern observed in the vomeronasal neuroepithelium of Bennett's wallaby using the Gαo marker [430] was identical to that described in the Tammar wallaby [256], in which only a small fraction of neuroreceptor cells was marked. However, while Schneider et al. did not obtain a positive Gαi2 immunolabeling in the VNO, Torres et al. [430] observed a large number of αi2 immunopositive cells in the vomeronasal neuroepithelium. Additionally, the strong immunopositivity identified in the vomeronasal axons, both in the proper lamina and in the nasal mucosa, confirmed the neuroreceptor nature of these cells [430]. At the AOB level, a clear anteroposterior zoning pattern typical of mammalian species belonging to the segregated model is observed. Vomeronasal axons reaching the anterior zone of the AOB only expressed Gαi2, while those reaching the posterior zone only expressed the Gαo subunit.
The use of the UEA lectin (Ulex europaeus) provided further evidence of the segregation of vomeronasal information in the wallaby, revealing selective labeling of the anterior zone of the AOB. The affinity of the UEA lectin for the anterior zone of the AOB has been reported in all species belonging to the segregation model in which this histochemical marker has been investigated, including the hamster [369], the mouse [147,417,431,432], the rat [264], and the capybara [409]. However, studies that employed UEA labeling in species belonging to the uniform model, including the pig [433], the cat [317], the dog [260], and the goat [434], have not shown any evidence of zonation in the AOB in any case.
Beyond the zonation, the main morphological and histological features of the wallaby VNS show the opening of the VNO to the nasopalatine duct, the semilunar shape of the vomeronasal duct, the stratification and cellularity of the neuroreceptor epithelium, and the presence of numerous PAS-positive vomeronasal glands in the lateral parenchyma (Figure 20, Figure 21, Figure 22 and Figure 23). Likewise, typical of these species is the arrangement of large blood vessels around the vomeronasal duct along its medial and lateral planes and a profuse, non-myelinated dorsal, medial, ventral, and ventrolateral innervation [291] responsible for transmitting the information collected by the vomeronasal sensory neuroepithelium to the AOB. This arrangement of the vomeronasal axons in the parenchyma is atypical, as it has not been previously described in the VNO of any other non-macropodid mammalian species. Another feature common to the VNO of both macropodids, observed by Schneider et al. [291] and [277] in the Tammar wallaby, is that the caudal vomeronasal capsule does not enclose the VNO parenchyma, so the posterior portion of the VNO is free from the surrounding cartilage. A similar finding has been reported in other marsupials, such as Notoryctes [435] and Caenolestes [436].
Regarding the lamination of the AOB in macropodids, it is characterized by a well-defined lamination, with an organization of mitral cells in a distinct mitral/plexiform layer [430]. Such features appear comparable to those described in species with highly developed AOBs, as seen in rodents [310] and lagomorphs [132]. The lectin LEA, in contrast to UEA, yields a non-zonal staining pattern, similar to that observed for OMP, a marker for mature olfactory and vomeronasal cells [437], and identical to the LEA pattern seen in the rabbit AOB [132]. Anti-GAP-43 confirms the presence of growing axons in the Bennett's wallaby AOB, with no observed differences between the anterior and posterior zones. However, anti-GFAP displayed more pronounced staining of the glial components in the posterior portion of the AOB compared to the anterior segment. Such a pattern has not been previously reported in other studies of this marker in the mammalian AOB [239,438], necessitating further examination in future research to interpret its implications.
Calbindin (CB) and calretinin (CR) are expressed throughout the VNS of the wallaby, comprising the neuroepithelium of the VNO (Figure 22), vomeronasal nerves, and the AOB. For both markers, VNO immunostaining encompasses the soma and dendrites in a pattern akin to that described in mice [262]. The distribution of CB and CR staining in the Bennett's wallaby AOB is concentrated in the vomeronasal fibers and glomeruli, labelling the neural and glomerular layers in a manner analogous to other species, like the rabbit [132] or capybara [409]. In other marsupials, such as the opossum, significant differences in CB and CR staining patterns in the AOB were observed when compared to the Bennett's wallaby pattern. In the opossum AOB, CB-labeled neurons were present in all layers, excluding the nervous layer and periglomerular cells. CR in the opossum follows a pattern similar to that in the wallaby; however, in the opossum, CR also labels mitral cells and distinguishes an anteroposterior zonation. This is evidenced by the presence of a more intense staining in the posterior zone of the AOB compared to its anterior part [339].
In short, the VNS of macropods displays a degree of differentiation and histochemical and neurochemical diversity comparable to species with a more developed VNS. The existence of the intermediate third type in the processing of vomeronasal information reported in the Tammar wallaby (Notamacropus eugenii) is not supported by our histochemical and immunohistochemical findings in the Bennett's wallaby (Notamacropus rufogriseus).
3.3. Meerkat (Suricata Suricatta) (Figure 24)
To the best of our knowledge, there is no existing neuroanatomical information on the VNO of the Herpestidae family beyond the recent neuroanatomical study of both the AOB and MOB of the meerkat [439]. Within this group, the meerkat holds an emblematic position. It is a gregarious species where marked dominance is evident, as one male and one female monopolize reproduction in groups of up to forty individuals [440], living primarily in dark burrows they dig themselves. This likely translates into a heightened development of their chemical perception senses. In fact, it has been determined that these animals can recognize kinship through the individual scent of each member [441,442]
The meerkat AOB displays specific inherent features, which is unsurprising considering the vast genomic and morphological diversity described for this structure in other mammals [443,444]. It is intriguing to compare the meerkat AOB with that of other Carnivora species, such as canids and felids, whose VNOs have been deeply studied. The cat (Felis silvestris catus) and the dog (Canis lupus familiaris) provide striking examples of the vast disparity in the degree of differentiation that can exist in the AOB structural organization. The cat AOB is well developed, macroscopically discernible, and shows clear layer definition [317]. In contrast, the limited differentiation observed in the dog AOB is striking [260,438,445]. In the meerkat, similar to canids, the AOB is hard to identify macroscopically. However, microscopically, the laminar pattern in meerkats is more defined than in dogs, more closely resembling the pattern described in cats. Moreover, the meerkat AOB includes well-defined glomeruli, a broad plexiform mitral layer with relatively numerous principal cells, and granular cells organized in clusters. These similarities are not unexpected since meerkats belong to the Feliformia suborder. However, the layer thickness, the degree of glomerular differentiation, and the number of cells found in the meerkat AOB seem to be inferior to those observed in cats.
Figure 24.
A. The vomeronasal organ of the meerkat. Transverse decalcified histological section stained with PAS. B. Magnification of the are boxed in A. NVN. Vomeronasal nerves; RM. Respiratory epithelium; V. Vomer; VNC. Vomeronasal cartilage; VNG. Vomeronasal glands.
Figure 24.
A. The vomeronasal organ of the meerkat. Transverse decalcified histological section stained with PAS. B. Magnification of the are boxed in A. NVN. Vomeronasal nerves; RM. Respiratory epithelium; V. Vomer; VNC. Vomeronasal cartilage; VNG. Vomeronasal glands.
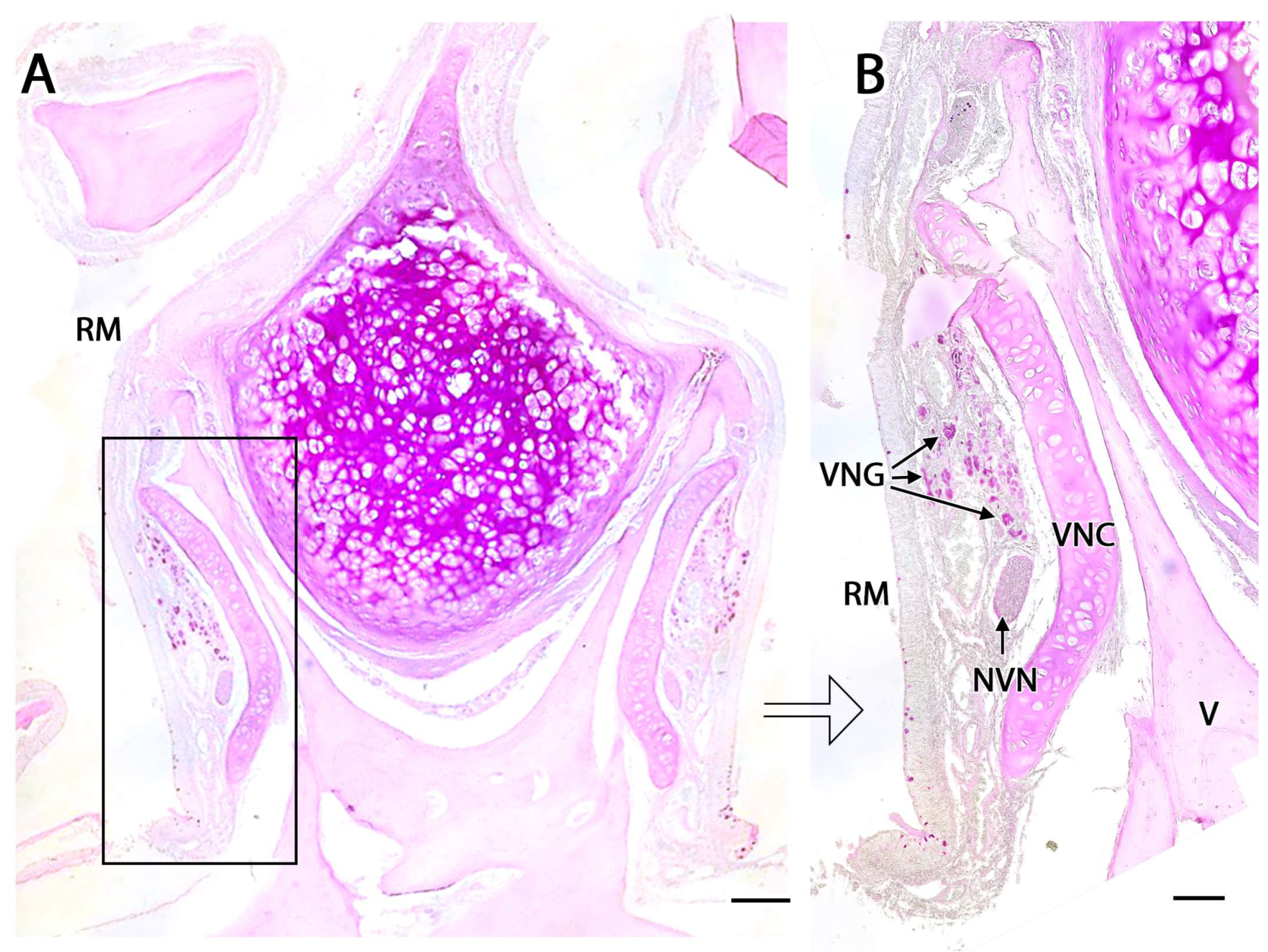
These observations gain greater significance in the context of the debate over the taxonomic classification of the meerkat. Its placement within the feliforms has wavered between the Herpestidae and Viverridae families [446]. Currently, the meerkat is considered a herpestid, alongside a wide range of mongoose species that share a broad spectrum of social complexities with it. The extensive morphological diversity observed in the VNO across species has served as a fruitful tool for establishing phylogenetic relationships among various mammalian species and identifying evolutionary pathways [447]. Despite the advent of genomic techniques, which have enabled population-level genetic variability characterization at a resolution previously unimaginable, the study of morphological features remains a critical element in mammalian taxonomic classification [448]. The VNO continues to be an important reference for these taxonomic studies [277,449]. Future research on the neuroanatomical and neurochemical characteristics of the meerkat VNO and AOB, combined with further observations in other viverrid and herpestid species, may help clarify this phylogenetic debate.
Regarding the expression of the G protein subunits, Gαi2 and Gαo, in the AOB, this aligns the meerkat with the group of mammals that have lost the expression of the V2R receptor family. It also tests positive for OMP, albeit with a slightly weaker expression in the AOB compared to the MOB. This pattern has been similarly reported in leporids [132], didelphids [450], and canids [340]. Anti-MAP-2 (Microtubule Associated Protein 2) and SMI-32 (Neurofilament Protein Marker 32) are routinely used to label neuronal somas in the OB. MAP-2 is mainly expressed in the mitral plexiform layer and in the granular stratum of the AOB, while SMI-32 is not expressed in the AOB, despite inducing a strong reaction in the mitral cells of the MOB.
Among the calcium-binding proteins, ant-CB and anti-CR immunostain the nervous stratum and in the neuropil of all glomeruli. However, periglomerular cells (PGC) are not marked by either anti-CR or anti-CB, which might be related to the low number of PGC detected in the Nissl stain. In contrast, the CR immunostaining pattern in the MOB glomeruli reveals a striking and intense immunostaining of the PGC. However, in the neuropil, the intensity of anti-CR staining is very weak, except for an isolated subpopulation of independent glomeruli located near the AOB, which has strong immunoreactivity. These atypical glomeruli might be involved in processing chemical signals from the vomeronasal nerve, as hypothesized for the olfactory limbus glomeruli in mice [130] and foxes [133].
Although the gorse lectin, UEA, has been employed as a specific marker for the global vomeronasal pathway (VNO, vomeronasal nerves, and AOB) in adult mice [417,451] and dogs, it does not produce any positive labeling in the meerkat AOB. Conversely, the lectin LEA stains the neural and glomerular layers in both the AOB and MOB.
3.4. Dama Gazelle (Nanger Dama)
The dama gazelle is a wild bovid about which only recent information regarding its vomeronasal system has been obtained, and it additionally holds the interest of being an endangered species. In certain species with a well-characterized VNS, methods related to this system are already employed to enhance reproduction, such as the use of pheromones, which help control stress levels and improve fertility [452]. While there currently exists a vast application field for bovine pheromones [453,454], their potential application in the reproduction of the dama gazelle necessitates a prior neuroanatomical and morphofunctional characterization of its vomeronasal organ.
A morphological feature of the VNO in the dama gazelle (Figure 25), common to most Ruminantia, is its remarkable length. However, the implications of this trait have only been thoroughly investigated in the dama gazelle, wherein immunohistochemical and lectin-histochemical characteristics of the vomeronasal duct and the vomeronasal nerves were evaluated throughout the VNO entire extent [455]. Beyond this fact, it is significant that in this Antilopinae species, the vomeronasal duct opens into the incisive duct, into which it incorporates from its medial side, both sharing the same cartilaginous capsule, which opens into a functional incisive papilla. This fact is especially relevant, as it is not a trait common to all Antilopinae; for instance, the three species of alcelaphine antelopes studied by [456] lack an incisive papilla.
The serial histological study of the VNO (Figure 26) has allowed the determination of the differences along the rostral-caudal axis of the organ concerning the epithelial lining of the vomeronasal duct. Among the most relevant aspects, it is striking how there is a narrowing in its lumen coinciding with the more central levels; however, in the final third of the organ, there is a remarkable increase in the development of the parenchyma and the size of the duct, which retains its medial sensory lining and lateral respiratory lining. The access of pheromones to this extensive caudal chamber requires the presence of a robust vascular pump, responsible for mobilizing pheromones from the external environment to the sensory cells of the VNO. This pump is driven by vasomotor movements, which allow the suction of stimulating substances into the vomeronasal organ and actively expel the content of the vomeronasal duct. These mechanisms are activated by fibers that run through the nasopalatine nerve and cause the constriction of the blood vessels within the VNO capsule [245]. After constriction, the volume of blood in the cavernous tissue is reduced, creating a pressure differential that expands the VNO lumen and extracts fluid from the region surrounding the duct opening [457]. Stimuli enter the vomeronasal organ in solution via the mucus stream that passes to the vomeronasal duct from the incisive duct. Functional vomeronasal stimuli must be molecules soluble in that mucus or substances that become soluble when binding to carrier molecules that may be secreted by the vomeronasal glands [458].
The access of pheromones is determined not only by the action of the pump but also by the physicochemical properties of the molecules. It seems logical to suggest that a diffusion gradient of substances is established along the duct, with only the most soluble substances in the glandular secretion that fills the vomeronasal duct reaching the caudal end, leaving the less soluble ones in the anterior part. It is consistent to hypothesize that the specificity of the vomeronasal receptors along the vomeronasal duct adapts to the differential migration of the substance mixture along the duct, just as compounds are specifically eluted along a chromatography column.
Given the significant variations observed in the structure of the vomeronasal duct and its epithelial lining, the neurochemical study of the VNO has been conducted not only in the central part of the VNO - the most typical - but has been extended throughout the VNO length. The study of G protein expression showed immunopositivity for both markers (Gαi2 and Gαo) in the vomeronasal epithelium. The serial immunohistochemical study throughout the entire vomeronasal duct confirmed that the expression of both markers is not constant in the VNO, as the large dilated caudal segment, despite having a well-characterized neuroepithelium, lacks immunolabeling for both proteins. This suggests that the receptor type expressed in this caudal third of the vomeronasal duct does not correspond to either V1R or V2R, suggesting that the molecules detected by these two receptor types do not reach the caudal end of the organ. This hypothesis implies that only highly soluble compounds in the vomeronasal mucosa and probably low molecular weight can be detected in the caudal VNO, and likely by another type of receptors not yet described. The absence of vomeronasal receptor expression in such an extensive segment of the VNO could explain why previous G protein expression studies in the VNO of ruminants have not found positivity for the Gαo marker, as has been the case for the goat [459], sheep [303], and sika deer [460].
It is worth noting that [429], in their study of the cow AOB, did not find positivity for Gαo, nor was it found in the dog AOB [260] or fox [340]. However, this does not exclude the expression of V2R receptors in the VNO, as it is plausible that the information from these receptors may project to other areas of the olfactory bulb, not necessarily to the AOB. In fact, it has recently been observed in the fox olfactory bulb that the expression of Gαo linked to the vomeronasal nerves does not project to the AOB, but to the transition zone located between the main olfactory bulb and the accessory, an area known as the olfactory limb [133].
The use of an additional range of neuronal markers, such as PGP 9.5 and specific neuronal enolase (EN), showed that the expression of these markers extends to the caudal sensory epithelium area, even though neither the analyzed G proteins nor the OMP marker is expressed there. This finding provides evidence that the histologically described neuroepithelial nature of this long caudal stretch of the vomeronasal duct is correct. On the other hand, it suggests that the type of receptors that are expressed differs from those that belong to the V1R and V2R families. Other markers, such as calcium-binding proteins, however, have differential expression. For example, antibodies against CB and CR mark a subpopulation of vomeronasal receptor neurons in the dama gazelle vomeronasal duct. As with G proteins and OMP, the expression of calcium-binding proteins did not occur in the caudal area of the sensory epithelium (Figure 27).
In summary, the research on the vomeronasal organ of the dama gazelle, besides providing valuable information about the configuration and function of this structure in this endangered species, could be used to design and implement programs based on the use of pheromones to enhance reproductive success and increase genetic diversity in captive populations. Such programs can be critical for the species survival. Furthermore, the discovery in this species of notable differences in the organization of the vomeronasal duct and the expression of neuronal markers along the rostro-caudal axis of the organ underscores the need to consider such differences when conducting future studies on the VNO of other species.
4. Conclusions
Throughout this review, our objective has been to provide a comprehensive and precise portrayal of the crucial role that chemical communication plays in mammals, specifically focusing on the detection of pheromone-mediated communication through the vomeronasal system. Due to comprehensive research and the accumulation of scientific knowledge, we have gained a profound understanding of the significant impact exerted by the vomeronasal system on animal behavior. This intricate sensory system profoundly influences several aspects of social and sexual interactions, encompassing reproductive processes, the formation of hierarchies, maternal bonding and intraspecies recognition. Furthermore, it is evident that there is a notable range of variability resulting from evolutionary adaptations within this particular system, surpassing the documented variances in the olfactory system. The extensive range of adaptive variability is a challenge in the examination of the vomeronasal system. Therefore, it is vital to direct further research efforts towards a deeper neuroanatomical and morphofunctional understanding of the system. The comprehensive understanding of the impact of pheromones on the physiology and welfare of animals and humans necessitates the pursuit of such an undertaking. Recognizing the intricacies of the vomeronasal system is paramount in the realm of biological research. By delving deeper into its evolutionary origins, functional roles, and neuroanatomical intricacies, we not only gain invaluable insights into its unique position in mammalian communication but also unveil its broader implications in social and reproductive behaviors. The culmination of such basic research has profound potential; it can set the stage for leveraging pheromones as powerful tools. By harnessing this knowledge, we open doors to innovative approaches in enhancing the well-being and health of both animals and humans.
Author Contributions
Conceptualization, M.V.T., I.O.-L. and P.S.-Q.; writing—original draft preparation, M.V.T., I.O.-L. and P.S.-Q.; writing—review and editing, M.V.T., I.O.-L. and P.S.-Q.; supervision, M.V.T. and P.S.-Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research has received no external funding.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stevens, M. Sensory Ecology, Behaviour, and Evolution; Oxford University Press: Oxford, UK, 2013; ISBN 978-0-19-960177-6. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behaviour: Communication by Smell and Taste; Cambridge University Press: Cambridge, UK ; New York, NY, USA, 2003; ISBN 978-0-521-48068-0. [Google Scholar]
- Wood, W.F. Chemical Ecology: Chemical Communication in Nature. J. Chem. Educ. 1983, 60, 531. [Google Scholar] [CrossRef]
- Castañeda, M. de L.A.; Martínez-Gómez, M.; Guevara-Guzmán, R.; Hudson, R. Chemical communication in domestic mammals. Vet. Mex. 2007, 38, 105–123. [Google Scholar]
- Kokocińska-Kusiak, A.; Woszczyło, M.; Zybala, M.; Maciocha, J.; Barłowska, K.; Dzięcioł, M. Canine Olfaction: Physiology, Behavior, and Possibilities for Practical Applications. Animals 2021, 11, 2463. [Google Scholar] [CrossRef]
- Iovino, M.; Messana, T.; Iovino, E.; De Pergola, G.; Guastamacchia, E.; Giagulli, V.A.; Triggiani, V. Neuroendocrine Mechanisms Involved in Male Sexual and Emotional Behavior. EMIDDT 2019, 19, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Beruter, J.; Beauchamp, G.K.; Muetterties, E.L. Complexity of Chemical Communication in Mammals: Urinary Components Mediating Sex Discrimination by Male Guinea Pigs. Biochem. Biophys. Res. Commun. 1973, 53, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. Chemical Communication within Animal Species. In Chemical Ecology; Academic: New York, NY, USA, 1970; pp. 133–155. [Google Scholar]
- Surov, A.V.; Maltsev, A.N. Analysis of Chemical Communication in Mammals: Zoological and Ecological Aspects. Biol. Bull. Russ. Acad. Sci. 2016, 43, 1175–1183. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Arana, Álvaro. J.; Coppel, C.; Ortiz-Leal, I.; Torres, M.V.; Sanchez-Quinteiro, P.; Sánchez, L. A Comprehensive Structural, Lectin and Immunohistochemical Characterization of the Zebrafish Olfactory System. Sci. Rep. 2021, 11, 8865. [Google Scholar] [CrossRef]
- Saraiva, L.R.; Ahuja, G.; Ivandic, I.; Syed, A.S.; Marioni, J.C.; Korsching, S.I.; Logan, D.W. Molecular and Neuronal Homology between the Olfactory Systems of Zebrafish and Mouse. Sci. Rep. 2015, 5, 11487. [Google Scholar] [CrossRef]
- Meister, M. On the Dimensionality of Odor Space. eLife 2015, 4, e07865. [Google Scholar] [CrossRef]
- Mollo, E.; Garson, M.J.; Polese, G.; Amodeo, P.; Ghiselin, M.T. Taste and Smell in Aquatic and Terrestrial Environments. Nat. Prod. Rep. 2017, 34, 496–513. [Google Scholar] [CrossRef]
- Hepper, P.G.; Wells, D.L. Perinatal Olfactory Learning in the Domestic Dog. Chem. Sens. 2006, 31, 207–212. [Google Scholar] [CrossRef]
- Weissburg, M.J.; Ferner, M.C.; Pisut, D.P.; Smee, D.L. Ecological Consequences of Chemically Mediated Prey Perception. J. Chem. Ecol. 2002, 28, 1953–1970. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, D.A.; Lewis, W.J. Terminology of Chemical Releasing Stimuli in Intraspecific and Interspecific Interactions. J. Chem. Ecol. 1976, 2, 211–220. [Google Scholar] [CrossRef]
- Law, J.H.; Regnier, F.E. Pheromones. Annu. Rev. Biochem. 1971, 40, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects 2019, 10, 441. [Google Scholar] [CrossRef]
- Burger, B.V. Mammalian Semiochemicals. In The Chemistry of Pheromones and Other Semiochemicals II; Schulz, S., Ed.; Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2004; Volume 240, pp. 231–278. ISBN 978-3-540-21308-6. [Google Scholar]
- Apfelbach, R.; Parsons, M.H.; Soini, H.A.; Novotny, M.V. Are Single Odorous Components of a Predator Sufficient to Elicit Defensive Behaviors in Prey Species? Front. Neurosci. 2015, 9. [Google Scholar] [CrossRef]
- Dicke, M.; Sabelis, M.W. Infochemical Terminology: Based on Cost-Benefit Analysis Rather than Origin of Compounds? Funct. Ecol. 1988, 2, 131. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Behavior. In Chemical Communication in Crustaceans; Breithaupt, T., Thiel, M., Eds.; Springer: New York, NY, USA, 2010; pp. 23–38. ISBN 978-0-387-77100-7. [Google Scholar]
- Sbarbati, A.; Osculati, F. Allelochemical Communication in Vertebrates: Kairomones, Allomones and Synomones. Cells Tissues Organs 2006, 183, 206–219. [Google Scholar] [CrossRef]
- Blum, M.S. Semiochemical Parsimony in the Arthropoda. Annu. Rev. Entomol. 1996, 41, 353–374. [Google Scholar] [CrossRef]
- Gallie, D.R.; Chang, S.C. Signal Transduction in the Carnivorous Plant Sarracenia Purpurea (Regulation of Secretory Hydrolase Expression during Development and in Response to Resources). Plant Physiol. 1997, 115, 1461–1471. [Google Scholar] [CrossRef]
- Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, I.S. The Effects of Predator Odors in Mammalian Prey Species: A Review of Field and Laboratory Studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [Google Scholar] [CrossRef]
- Dunkelblum, E.; Mendel, Z.; Gries, G.; Gries, R.; Zegelman, L.; Hassner, A.; Mori, K. Antennal Response and Field Attraction of the Predator Elatophilus Hebraicus (Hemiptera: Anthocoridae) to Sex Pheromones and Analogues of Three Matsucoccus Spp. (Homoptera: Matsucoccidae). Bioorganic Med. Chem. 1996, 4, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Cavill, G.W.K.; Robertson, P.L. Ant Venoms, Attractants, and Repellents: Secretions Are Used by Ants in Attack and Defense and as Chemical Messengers in Their Social Organization. Science 1965, 149, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Mattiacci, L.; Dicke, M.; Posthumus, M.A. Induction of Parasitoid Attracting Synomone in Brussels Sprouts Plants by Feeding OfPieris Brassicae Larvae: Role of Mechanical Damage and Herbivore Elicitor. J. Chem. Ecol. 1994, 20, 2229–2247. [Google Scholar] [CrossRef] [PubMed]
- Stowe, M.K.; Turlings, T.C.; Loughrin, J.H.; Lewis, W.J.; Tumlinson, J.H. The Chemistry of Eavesdropping, Alarm, and Deceit. Proc. Natl. Acad. Sci. USA 1995, 92, 23–28. [Google Scholar] [CrossRef]
- Han, B.; Chen, Z. Behavioral and Electrophysiological Responses of Natural Enemies to Synomones from Tea Shoots and Kairomones from Tea Aphids, Toxoptera Aurantii. J. Chem. Ecol. 2002, 28, 2203–2219. [Google Scholar] [CrossRef]
- Pernaa, J.; Aksela, M. Learning Organic Chemistry through a Study of Semiochemicals. J. Chem. Educ. 2011, 88, 1644–1647. [Google Scholar] [CrossRef]
- Kasinger, H.; Bauer, B.; Denzinger, J. The Meaning of Semiochemicals to the Design of Self-Organizing Systems. In Proceedings of the 2008 Second IEEE International Conference on Self-Adaptive and Self-Organizing Systems, Venezia, Italy, October 2008; pp. 139–148. [Google Scholar]
- Karlson, P.; Lüscher, M. ‘Pheromones’: A New Term for a Class of Biologically Active Substances. Nature 1959, 183, 55–56. [Google Scholar] [CrossRef]
- Butenandt, A.; Beckmann, R.; Stamm, D.; Hecker, E. Uber Den Sexsual-Lockstoff Des Seidenspinners Bombyx Mori. Reindarstellung Und Konstitution. Z. Naturforschg. 1959, 14b, 283–284. [Google Scholar]
- Wilson, E.O. Chemical Communication in the Social Insects: Insect Societies Are Organized Principally by Complex Systems of Chemical Signals. Science 1965, 149, 1064–1071. [Google Scholar] [CrossRef]
- Conte, Y.L.; Hefetz, A. Primer Pheromones in Social Hymenoptera. Annu. Rev. Entomol. 2008, 53, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Cork, A.; Kamal, N.; Alam, S.; Choudhury, J.; Talekar, N. Pheromones and Their Applications to Insect Pest Control. Bangl J. Entomol. 2003, 13, 1–13. [Google Scholar]
- Liberles, S.D. Mammalian Pheromones. Annu. Rev. Physiol. 2014, 76, 151–175. [Google Scholar] [CrossRef]
- Brennan, P.A.; Zufall, F. Pheromonal Communication in Vertebrates. Nature 2006, 444, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, G.K.; Doty, R.L.; Moulton, D.G.; Mugford, R.A. The Pheromone Concept in Mammalian Chemical Communication: A Critique. In Mammalian Olfaction, Reproductive Processes, and Behavior; Elsevier: Amsterdam, The Netherlands, 1976; pp. 143–160. ISBN 978-0-12-221250-5. [Google Scholar]
- Brown, R.E. Mammalian Social Odors: A Critical Review. In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 1979; Volume 10, pp. 103–162. ISBN 978-0-12-004510-5. [Google Scholar]
- Booth, W.D.; Signoret, J.P. Olfaction and Reproduction in Ungulates. Oxf. Rev. Reprod. Biol. 1992, 14, 263–301. [Google Scholar]
- Schaal, B.; Coureaud, G.; Langlois, D.; Giniès, C.; Sémon, E.; Perrier, G. Chemical and Behavioural Characterization of the Rabbit Mammary Pheromone. Nature 2003, 424, 68–72. [Google Scholar] [CrossRef]
- Apps, P.J.; Weldon, P.J.; Kramer, M. Chemical Signals in Terrestrial Vertebrates: Search for Design Features. Nat. Prod. Rep. 2015, 32, 1131–1153. [Google Scholar] [CrossRef]
- Ferrero, D.M.; Moeller, L.M.; Osakada, T.; Horio, N.; Li, Q.; Roy, D.S.; Cichy, A.; Spehr, M.; Touhara, K.; Liberles, S.D. A Juvenile Mouse Pheromone Inhibits Sexual Behaviour through the Vomeronasal System. Nature 2013, 502, 368–371. [Google Scholar] [CrossRef]
- Lin, D.Y.; Zhang, S.-Z.; Block, E.; Katz, L.C. Encoding Social Signals in the Mouse Main Olfactory Bulb. Nature 2005, 434, 470–477. [Google Scholar] [CrossRef]
- Novotny, M.; Harvey, S.; Jemiolo, B.; Alberts, J. Synthetic Pheromones That Promote Inter-Male Aggression in Mice. Proc. Natl. Acad. Sci. USA 1985, 82, 2059–2061. [Google Scholar] [CrossRef]
- Singer, A.G.; Agosta, W.C.; O’Connell, R.J.; Pfaffmann, C.; Bowen, D.V.; Field, F.H. Dimethyl Disulfide: An Attractant Pheromone in Hamster Vaginal Secretion. Science 1976, 191, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Dorries, K.M.; Adkins-regan, E.; Halpern, B.P. Olfactory Sensitivity to the Pheromone, Androstenone, Is Sexually Dimorphic in the Pig. Physiol. Behav. 1995, 57, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.E.L.; Lazar, J.; Greenwood, D.R. Olfactory Adventures of Elephantine Pheromones. Biochem. Soc. Trans. 2003, 31, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.L.; Eisner, T.; Whittaker, R.H. Allomones and Kairomones: Transspecific Chemical Messengers. BioScience 1970, 20, 21–22. [Google Scholar] [CrossRef]
- Lass, S.; Bittner, K. Facing Multiple Enemies: Parasitised Hosts Respond to Predator Kairomones. Oecologia 2002, 132, 344–349. [Google Scholar] [CrossRef]
- Weldon, P.J. In Defense of ?Kairomone? As a Class of Chemical Releasing Stimuli. J. Chem. Ecol. 1980, 6, 719–725. [Google Scholar] [CrossRef]
- Pasteels, J.M. Is Kairomone a Valid and Useful Term? J. Chem. Ecol. 1982, 8, 1079–1081. [Google Scholar] [CrossRef]
- Ruther, J.; Meiners, T.; Steidle, J.L.M. Rich in Phenomena-Lacking in Terms. A Classification of Kairomones. Chemoecology 2002, 12, 161–167. [Google Scholar] [CrossRef]
- Ayelo, P.M.; Pirk, C.W.W.; Yusuf, A.A.; Chailleux, A.; Mohamed, S.A.; Deletre, E. Exploring the Kairomone-Based Foraging Behaviour of Natural Enemies to Enhance Biological Control: A Review. Front. Ecol. Evol. 2021, 9, 641974. [Google Scholar] [CrossRef]
- Schonewolf, K.W.; Bell, R.; Rypstra, A.L.; Persons, M.H. Field Evidence of an Airborne Enemy-Avoidance Kairomone in Wolf Spiders. J. Chem. Ecol. 2006, 32, 1565–1576. [Google Scholar] [CrossRef]
- Reinecke, A.; Ruther, J.; Tolasch, T.; Francke, W.; Hilker, M. Alcoholism in Cockchafers: Orientation of Male Melolontha Melolontha towards Green Leaf Alcohols. Naturwissenschaften 2002, 89, 265–269. [Google Scholar] [CrossRef]
- Jones, R.L.; Lewis, W.J.; Bowman, M.C.; Beroza, M.; Bierl, B.A. Host-Seeking Stimulant for Parasite of Corn Earworm: Isolation, Identification, and Synthesis. Science 1971, 173, 842–843. [Google Scholar] [CrossRef] [PubMed]
- Acree, F.; Turner, R.B.; Gouck, H.K.; Beroza, M.; Smith, N. L-Lactic Acid: A Mosquito Attractant Isolated from Humans. Science 1968, 161, 1346–1347. [Google Scholar] [CrossRef] [PubMed]
- Thibout, E.; Guillot, J.F.; Ferary, S.; Limouzin, P.; Auger, J. Origin and Identification of Bacteria Which Produce Kairomones in the Frass OfAcrolepiopsis Assectella (Lep., Hyponomeutoidea). Experientia 1995, 51, 1073–1075. [Google Scholar] [CrossRef] [PubMed]
- Schoeppner, N.M.; Relyea, R.A. Damage, Digestion, and Defence: The Roles of Alarm Cues and Kairomones for Inducing Prey Defences: Damage, Digestion, and Defence. Ecol. Lett. 2005, 8, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Kugimiya, S.; Takabayashi, J.; Bohlmann, J. Herbivore-Induced Defense Response in a Model Legume. Two-Spotted Spider Mites Induce Emission of ( E )- β -Ocimene and Transcript Accumulation of ( E )- β -Ocimene Synthase in Lotus Japonicus. Plant Physiol. 2004, 135, 1976–1983. [Google Scholar] [CrossRef]
- Reinecke, A.; Ruther, J.; Mayer, C.J.; Hilker, M. Optimized Trap Lure for Male Melolontha Cockchafers. J. Appl. Entomol. 2006, 130, 171–176. [Google Scholar] [CrossRef]
- Briand, L.; Trotier, D.; Pernollet, J.-C. Aphrodisin, an Aphrodisiac Lipocalin Secreted in Hamster Vaginal Secretions. Peptides 2004, 25, 1545–1552. [Google Scholar] [CrossRef]
- Altmann, D.; Sorensen, L. Zoo Behaviour Science in the Research Centre for Vertebrate Studies of the Academy of Sciences of the German Democratic Republic (in the Animal Park of Berlin). Appl. Anim. Behav. Sci. 1987, 18, 67–81. [Google Scholar] [CrossRef]
- Salazar, I.; Fdez de Troconiz, P.; Prieto, M.D.; Cifuentes, J.M.; Quinteiro, P.S. Anatomy and Cholinergic Innervation of the Sinus Paranalis in Dogs. Anatom Histol. Embryol. 1996, 25, 49–53. [Google Scholar] [CrossRef]
- Martín, J.; Barja, I.; López, P. Chemical Scent Constituents in Feces of Wild Iberian Wolves (Canis Lupus Signatus). Biochem. Syst. Ecol. 2010, 38, 1096–1102. [Google Scholar] [CrossRef]
- Aviles-Rosa, E.O.; Surowiec, K.; McGlone, J. Identification of Faecal Maternal Semiochemicals in Swine (Sus Scrofa) and Their Effects on Weaned Piglets. Sci. Rep. 2020, 10, 5349. [Google Scholar] [CrossRef] [PubMed]
- Ewer, R.F. Ethology of Mammals; Springer US: Boston, MA, USA, 1968; ISBN 978-1-4899-4658-4. [Google Scholar]
- Bradshaw, J.W.S.; Cameron-Beaumont, C.L. The Signaling Repertoire of the Domestic Cat and Its Undomesticated Relatives. In The Domestic Cat: The Biology of Its Behaviour; Cambridge University Press: Cambridge, UK, 2000; pp. 67–93. [Google Scholar]
- Miyazaki, M.; Yamashita, T.; Suzuki, Y.; Saito, Y.; Soeta, S.; Taira, H.; Suzuki, A. A Major Urinary Protein of the Domestic Cat Regulates the Production of Felinine, a Putative Pheromone Precursor. Chem. Biol. 2006, 13, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Stefańczyk-Krzymowska, S.; Krzymowski, T.; Wasowska, B.; Jana, B.; Słomiński, J. Intramuscular Injections of Male Pheromone 5 Alpha-Androstenol Change the Secretory Ovarian Function in Gilts during Sexual Maturation. Reprod. Biol. 2003, 3, 241–257. [Google Scholar] [PubMed]
- Booth, W.D. Sexual Dimorphism Involving Steroidal Pheromones and Their Binding Protein in the Submaxillary Salivary Gland of the Göttingen Miniature Pig. J. Endocrinol. 1984, 100, 195. [Google Scholar] [CrossRef]
- Pietras, R.J. Sex Pheromone Production by Preputial Gland: The Regulatory Role of Estrogen. Chem. Sens. 1981, 6, 391–408. [Google Scholar] [CrossRef]
- Murphy, M.R.; Schneider, G.E. Olfactory Bulb Removal Eliminates Mating Behavior in the Male Golden Hamster. Science 1970, 167, 302–304. [Google Scholar] [CrossRef]
- Michael, R.P.; Keverne, E.B. Primate Sex Pheromones of Vaginal Origin. Nature 1970, 225, 84–85. [Google Scholar] [CrossRef]
- Goodwin, M.; Gooding, K.M.; Regnier, F. Sex Pheromone in the Dog. Science 1979, 203, 559–561. [Google Scholar] [CrossRef]
- Rivard, G.; Klemm, W.R. Two Body Fluids Containing Bovine Estrous Pheromone(s). Chem. Sens. 1989, 14, 273–279. [Google Scholar] [CrossRef]
- Hayes, R.A.; Richardson, B.J.; Claus, S.C.; Wyllie, S.G. Semiochemicals and Social Signaling in the Wild European Rabbit in Australia: II. Variations in Chemical Composition of Chin Gland Secretion across Sampling Sites. J. Chem. Ecol. 2002, 28, 2613–2625. [Google Scholar] [CrossRef] [PubMed]
- Thiessen, D.D.; Friend, H.C.; Lindzey, G. Androgen Control of Territorial Marking in the Mongolian Gerbil. Science 1968, 160, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Müller-Schwarze, D. Pheromones in Black-Tailed Deer (Odocoileus Hemionus Columbianus). Anim. Behav. 1971, 19, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Müller-Schwarze, D.; Müller-Schwarze, C.; Singer, A.G.; Silverstein, R.M. Mammalian Pheromone: Identification of Active Component in the Subauricular Scent of the Male Pronghorn. Science 1974, 183, 860–862. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Krantz, K.; Aplin, R.T. Behavioural, Anatomical and Chemical Aspects of Scent Marking amongst Capybaras (Hydrochoerus Hydrochaeris ) (Rodentia: Caviomorpha). J. Zool. 1984, 202, 341–360. [Google Scholar] [CrossRef]
- Berüter, J.; Beauchamp, G.K.; Muetterties, E.L. Mammalian Chemical Communication: Perineal Gland Secretion of the Guinea Pig. Physiol. Zool. 1974, 47, 130–136. [Google Scholar] [CrossRef]
- Schultze-Westrum, T. Innerartliche Verständigung durch Düfte beim Gleitbeutler Petaurus breviceps papuanus Thomas (Marsupialia, Phalangeridae). Z. Vergl. Physiol. 1965, 50, 151–220. [Google Scholar] [CrossRef]
- Stumpf, P.; K�nzle, H.; Welsch, U. Cutaneous Eccrine Glands of the Foot Pads of the Small Madagascan Tenrec ( Echinops Telfairi, Insectivora, Tenrecidae): Skin Glands in a Primitive Mammal. Cell Tissue Res. 2004, 315, 59–70. [Google Scholar] [CrossRef]
- Yager, J.A.; Hunter, D.B.; Wilson, M.R.; Allen, O.B. A Source of Cutaneous Maternal Semiochemicals in the Mink? Experientia 1988, 44, 79–81. [Google Scholar] [CrossRef]
- Izard, M.K. Pheromones and Reproduction in Domestic Animals. In Pheromones and Reproduction in Mammals; eBook: 1983.
- Melrose, D.R.; Reed, H.C.B.; Patterson, R.L.S. Androgen Steroids Associated with Boar Odour as an Aid to the Detection of Oestrus in Pig Artificial Insemination. Br. Vet. J. 1971, 127, 497–502. [Google Scholar] [CrossRef]
- Gosling, L.M. A Reassessment of the Function of Scent Marking in Territories. Z. Für Tierpsychol. 1982, 60, 89–118. [Google Scholar] [CrossRef]
- Eisenberg, J.F.; Kleiman, D.G. Olfactory Communication in Mammals. Annu. Rev. Ecol. Syst. 1972, 3, 1–32. [Google Scholar] [CrossRef]
- Johnson, R.P. Scent Marking in Mammals. Anim. Behav. 1973, 21, 521–535. [Google Scholar] [CrossRef]
- Jones, R.B.; Nowell, N.W. A Comparison of the Aversive and Female Attractant Properties of Urine from Dominant and Subordinate Male Mice. Anim. Learn. Behav. 1974, 2, 141–144. [Google Scholar] [CrossRef]
- Hayes, R.A.; Richardson, B.J.; Wyllie, S.G. To Fix or Not to Fix: The Role of 2-Phenoxyethanol in Rabbit, Oryctolagus Cuniculus, Chin Gland Secretion. J. Chem. Ecol. 2003, 29, 1051–1064. [Google Scholar] [CrossRef]
- Wirant, S.C.; Halvorsen, K.T.; McGuire, B. Preliminary Observations on the Urinary Behaviour of Female Jack Russell Terriers in Relation to Stage of the Oestrous Cycle, Location, and Age. Appl. Anim. Behav. Sci. 2007, 106, 161–166. [Google Scholar] [CrossRef]
- Müller, C.A.; Manser, M.B. Scent-Marking and Intrasexual Competition in a Cooperative Carnivore with Low Reproductive Skew: Scent-Marking and Intrasexual Competition. Ethology 2008, 114, 174–185. [Google Scholar] [CrossRef]
- Bronson, F.H.; Whitten, W.K. Oestrus-Accelerating Pheromone of Mice: Assay, Androgen-Dependency and Presence in Bladder Urine. Reproduction 1968, 15, 131–134. [Google Scholar] [CrossRef]
- Bronson, F.H.; Caroom, D. Preputial Gland of the Male Mouse: Attractant Function. Reproduction 1971, 25, 279–282. [Google Scholar] [CrossRef]
- Seitz, E. Die Bedeutung Geruchlicher Orientierung Beim Plumplori Nycticebus Coucang Boddaert 1785 (Prosimii, Lorisidae). Z. Für Tierpsychol. 1969, 26, 73–103. [Google Scholar] [CrossRef]
- Harrington, F.H. Urine Marking at Food and Caches in Captive Coyotes. Can. J. Zool. 1982, 60, 776–782. [Google Scholar] [CrossRef]
- Sun, L.; Müller-Schwarze, D. Sibling Recognition in the Beaver: A Field Test for Phenotype Matching. Anim. Behav. 1997, 54, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Jordan, N.R.; Golabek, K.A.; Apps, P.J.; Gilfillan, G.D.; McNutt, J.W. Scent-Mark Identification and Scent-Marking Behaviour in African Wild Dogs (Lycaon Pictus ). Ethology 2013, 119, 644–652. [Google Scholar] [CrossRef]
- Ferkin, M.H.; Johnston, R.E. Meadow Voles, Microtus Pennsylvanicus, Use Multiple Sources of Scent for Sex Recognition. Anim. Behav. 1995, 49, 37–44. [Google Scholar] [CrossRef]
- Hradecký, P. Possible Pheromonal Regulation of Reproduction in Wild Carnivores. J. Chem. Ecol. 1985, 11, 241–250. [Google Scholar] [CrossRef]
- Kiyokawa, Y.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Modulatory Role of Testosterone in Alarm Pheromone Release by Male Rats. Horm. Behav. 2004, 45, 122–127. [Google Scholar] [CrossRef]
- Boissy, A.; Terlouw, C.; Le Neindre, P. Presence of Cues from Stressed Conspecifics Increases Reactivity to Aversive Events in Cattle: Evidence for the Existence of Alarm Substances in Urine. Physiol. Behav. 1998, 63, 489–495. [Google Scholar] [CrossRef]
- Aleksiuk, M. Scent-Mound Communication, Territoriality, and Population Regulation in Beaver (Castor Canadensis Kuhl). J. Mammal. 1968, 49, 759–762. [Google Scholar] [CrossRef]
- Bartecki, U.; Heymann, E.W. Field Observations on Scent-marking Behaviour in Saddle-back Tamarins, Saguinus Fuscicollis (Callitrichidae, Primates). J. Zool. 1990, 220, 87–99. [Google Scholar] [CrossRef]
- Tschanz, B.; Meyer-Holzapfel, M.; Bachmann, S. Das Informationssystem Bei Braunbären1. Z. Für Tierpsychol. 2010, 27, 47–72. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Iwata, E.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Regional Differences of Pheromone Production in the Sebaceous Glands of Castrated Goats Treated with Testosterone. J. Vet. Med. Sci. 2000, 62, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, J.F. A Comparative Study of Sandbathing Behavior in Heteromyid Rodents. Behav. 1963, 22, 16–23. [Google Scholar] [CrossRef]
- Hayes, R.A.; Richardson, B.J.; Wyllie, S.G. Semiochemicals and Social Signaling in the Wild European Rabbit in Australia: I. Scent Profiles of Chin Gland Secretion from the Field. J. Chem. Ecol. 2002, 28, 363–384. [Google Scholar] [CrossRef]
- Laska, M.; Bauer, V.; Salazar, L.T.H. Self-Anointing Behavior in Free-Ranging Spider Monkeys (Ateles Geoffroyi) in Mexico. Primates 2007, 48, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Stoddart, D.M.; Ding, H.; Zhang, J. Self-Anointing Behavior in the Rice-Field Rat, Rattus Rattoides. J. Mammal. 1995, 76, 1238–1241. [Google Scholar] [CrossRef]
- Breer, H.; Fleischer, J.; Strotmann, J. Signaling in the Chemosensory Systems: The Sense of Smell: Multiple Olfactory Subsystems. Cell. Mol. Life Sci. 2006, 63, 1465–1475. [Google Scholar] [CrossRef]
- Munger, S.D. Noses within Noses. Nature 2009, 459, 521–522. [Google Scholar] [CrossRef]
- Barrios, A.W.; Sanchez Quinteiro, P.; Salazar, I. The Nasal Cavity of the Sheep and Its Olfactory Sensory Epithelium. Microsc. Res. Tech. 2014, 77, 1052–1059. [Google Scholar] [CrossRef]
- Cajal, Sr. Inducciones Fisiológicas de La Morfología y Conexiones de Las Neuronas. Arch. Pedagog. Y Cienc. Afines 1906, 1, 216–236. [Google Scholar]
- Salazar, I.; Sanchez-Quinteiro, P.; Barrios, A.W.; López Amado, M.; Vega, J.A. Anatomy of the Olfactory Mucosa. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 164, pp. 47–65. ISBN 978-0-444-63855-7. [Google Scholar]
- McCotter, R.E. The Connection of the Vomeronasal Nerves with the Accessory Olfactory Bulb in the Opossum and Other Mammals. Anat. Rec. 1912, 6, 299–318. [Google Scholar] [CrossRef]
- Salazar, I.; Barrios, A.W.; Sánchez-Quinteiro, P. Revisiting the Vomeronasal System From an Integrated Perspective. Anat. Rec. 2016, 299, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Barrios, A.W.; Núñez, G.; Sanchez Quinteiro, P.; Salazar, I. Anatomy, Histochemistry, and Immunohistochemistry of the Olfactory Subsystems in Mice. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Breer, H. The Grueneberg Ganglion: A Novel Sensory System in the Nose. Histol. Histopathol. 2010, 909–915. [Google Scholar] [CrossRef]
- Storan, M.J.; Key, B. Septal Organ of Grüneberg Is Part of the Olfactory System. J. Comp. Neurol. 2006, 494, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; López-Beceiro, A.; Sanchez-Quinteiro, P. The Vomeronasal Organ of Wild Canids: The Fox (Vulpes Vulpes) as a Model. J. Anat. 2020, 237, 890–906. [Google Scholar] [CrossRef]
- Ogura, T.; Krosnowski, K.; Zhang, L.; Bekkerman, M.; Lin, W. Chemoreception Regulates Chemical Access to Mouse Vomeronasal Organ: Role of Solitary Chemosensory Cells. PLoS ONE 2010, 5, e11924. [Google Scholar] [CrossRef]
- Juilfs, D.M.; Fülle, H.J.; Zhao, A.Z.; Houslay, M.D.; Garbers, D.L.; Beavo, J.A. A Subset of Olfactory Neurons That Selectively Express CGMP-Stimulated Phosphodiesterase (PDE2) and Guanylyl Cyclase-D Define a Unique Olfactory Signal Transduction Pathway. Proc. Natl. Acad. Sci. USA 1997, 94, 3388–3395. [Google Scholar] [CrossRef]
- Larriva-Sahd, J. Cytological Organization of the Alpha Component of the Anterior Olfactory Nucleus and Olfactory Limbus. Front. Neuroanat. 2012, 6. [Google Scholar] [CrossRef]
- Valverde, F.; López-Mascaraque, L.; De Carlos, J.A. Structure of the Nucleus Olfactorius Anterior of the Hedgehog (Erinaceus Europaeus). J. Comp. Neurol. 1989, 279, 581–600. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Cifuentes, J.M.; Quintela, L.; Barcia, R.; Sanchez-Quinteiro, P. Structural, Morphometric and Immunohistochemical Study of the Rabbit Accessory Olfactory Bulb. Brain Struct. Funct. 2020, 225, 203–226. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Vargas-Barroso, V.; Fidalgo, L.E.; López-Beceiro, A.M.; Larriva-Sahd, J.A.; Sánchez-Quinteiro, P. The Olfactory Limbus of the Red Fox (Vulpes Vulpes). New Insights Regarding a Noncanonical Olfactory Bulb Pathway. Front. Neuroanat. 2023, 16, 1097467. [Google Scholar] [CrossRef]
- Menco, B. Ultrastructural Studies on Membrane, Cytoskeletal, Mucous, and Protective Compartments in Olfaction. Microsc. Res. Tech. 1992, 22, 215–224. [Google Scholar] [CrossRef]
- Dryer, L.; Berghard, A. Odorant Receptors: A Plethora of G-Protein-Coupled Receptors. Trends Pharmacol. Sci. 1999, 20, 413–417. [Google Scholar] [CrossRef]
- Zhang, X.; Firestein, S. The Olfactory Receptor Gene Superfamily of the Mouse. Nat. Neurosci. 2002, 5, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial Receptor Codes for Odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Crespo, C.; Liberia, T.; Blasco-Ibáñez, J.M.; Nácher, J.; Varea, E. Cranial Pair I: The Olfactory Nerve: THE OLFACTORY NERVE. Anat. Rec. 2019, 302, 405–427. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P.; Wang, F.; Dulac, C.; Chao, S.K.; Nemes, A.; Mendelsohn, M.; Edmondson, J.; Axel, R. Visualizing an Olfactory Sensory Map. Cell 1996, 87, 675–686. [Google Scholar] [CrossRef]
- Price, J.L.; Powell, T.P. Certain Observations on the Olfactory Pathway. J. Anat. 1971, 110, 105–126. [Google Scholar]
- Höfer, D.; Shin, D.; Drenckhahn, D. Identification of Cytoskeletal Markers for the Different Microvilli and Cell Types of the Rat Vomeronasal Sensory Epithelium. J. Neurocytol. 2000, 29, 147–156. [Google Scholar] [CrossRef]
- Salazar, I.; Quinteiro, P.S.; Cifuentes, J.M. Comparative Anatomy of the Vomeronasal Cartilage in Mammals: Mink, Cat, Dog, Pig, Cow and Horse. Ann. Anat. Anat. Anz. 1995, 177, 475–481. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Cifuentes, J.M.; Fdz. -de-Troconiz, P.; Sanchez-Quinteiro, P. Morphological and Immunohistochemical Study of the Rabbit Vomeronasal Organ. J. Anat. 2018, 233, 814–827. [Google Scholar] [CrossRef]
- Wöhrmann-Repenning, A. Comparative anatomical studies of the vomeronasal complex and the rostral palate of various mammals. Gegenbaurs Morphol. Jahrb. 1984, 130, 501–530. [Google Scholar]
- Salazar, I.; Sánchez-Quinteiro, P.; Alemañ, N.; Prieto, D. Anatomical, Immnunohistochemical and Physiological Characteristics of the Vomeronasal Vessels in Cows and Their Possible Role in Vomeronasal Reception. J. Anat. 2008, 212, 686–696. [Google Scholar] [CrossRef]
- Wysocki, C.J. Neurobehavioral Evidence for the Involvement of the Vomeronasal System in Mammalian Reproduction. Neurosci. Biobehav. Rev. 1979, 3, 301–341. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez Quinteiro, P. Differential Development of Binding Sites for Four Lectins in the Vomeronasal System of Juvenile Mouse: From the Sensory Transduction Site to the First Relay Stage. Brain Res. 2003, 979, 15–26. [Google Scholar] [CrossRef]
- Ressler, K.J.; Sullivan, S.L.; Buck, L.B. A Zonal Organization of Odorant Receptor Gene Expression in the Olfactory Epithelium. Cell 1993, 73, 597–609. [Google Scholar] [CrossRef]
- Zapiec, B.; Mombaerts, P. The Zonal Organization of Odorant Receptor Gene Choice in the Main Olfactory Epithelium of the Mouse. Cell Rep. 2020, 30, 4220–4234. [Google Scholar] [CrossRef]
- Trinh, K.; Storm, D.R. Vomeronasal Organ Detects Odorants in Absence of Signaling through Main Olfactory Epithelium. Nat. Neurosci. 2003, 6, 519–525. [Google Scholar] [CrossRef]
- Hohenbrink, P.; Mundy, N.I.; Zimmermann, E.; Radespiel, U. First Evidence for Functional Vomeronasal 2 Receptor Genes in Primates. Biol. Lett. 2013, 9, 20121006. [Google Scholar] [CrossRef]
- Pro-Sistiaga, P.; Mohedano-Moriano, A.; Ubeda-Bañon, I.; Del Mar Arroyo-Jimenez, M.; Marcos, P.; Artacho-Pérula, E.; Crespo, C.; Insausti, R.; Martinez-Marcos, A. Convergence of Olfactory and Vomeronasal Projections in the Rat Basal Telencephalon. J. Comp. Neurol. 2007, 504, 346–362. [Google Scholar] [CrossRef]
- Grüneberg, H. A Ganglion Probably Belonging to the N. Terminalis System in the Nasal Mucosa of the Mouse. Z. Anat. Entwickl. Gesch. 1973, 140, 39–52. [Google Scholar] [CrossRef]
- Fleischer, J.; Schwarzenbacher, K.; Breer, H. Expression of Trace Amine-Associated Receptors in the Grueneberg Ganglion. Chem. Sens. 2007, 32, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Schwarzenbacher, K.; Besser, S.; Hass, N.; Breer, H. Olfactory Receptors and Signalling Elements in the Grueneberg Ganglion. J. Neurochem. 2006, 98, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.; Pyrski, M.; Biel, M.; Leinders-Zufall, T.; Zufall, F. Grueneberg Ganglion Neurons Are Finely Tuned Cold Sensors. J. Neurosci. 2010, 30, 7563–7568. [Google Scholar] [CrossRef] [PubMed]
- Rodolfo-Masera, T. Su l’esistenza Di Un Particolare Organo Olfattivo Nel Setto Nasale Della Cavia e Di Altri Roditori. Arch. Ital. Anat. Embriol. 1943, 48, 157–213. [Google Scholar]
- Tian, H.; Ma, M. Molecular Organization of the Olfactory Septal Organ. J. Neurosci. 2004, 24, 8383–8390. [Google Scholar] [CrossRef] [PubMed]
- 159. Weiler, E.; Farbman, A.I. The Septal Organ of the Rat During Postnatal Development. Chem. Sens. 2003, 28, 581–593. [Google Scholar] [CrossRef]
- Ma, M.; Grosmaitre, X.; Iwema, C.L.; Baker, H.; Greer, C.A.; Shepherd, G.M. Olfactory Signal Transduction in the Mouse Septal Organ. J. Neurosci. 2003, 23, 317–324. [Google Scholar] [CrossRef]
- Marshall, D.A.; Maruniak, J.A. Masera’s Organ Responds to Odorants. Brain Res. 1986, 366, 329–332. [Google Scholar] [CrossRef]
- Wysocki, C.J.; Wellington, J.L.; Beauchamp, G.K. Access of Urinary Nonvolatiles to the Mammalian Vomeronasal Organ. Science 1980, 207, 781–783. [Google Scholar] [CrossRef]
- Tizzano, M.; Cristofoletti, M.; Sbarbati, A.; Finger, T.E. Expression of Taste Receptors in Solitary Chemosensory Cells of Rodent Airways. BMC Pulm. Med. 2011, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Moulton, D.G. Chemosensitivity of Rat Nasal Trigeminal Receptors. Physiol. Behav. 1982, 28, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Filoramo, N.I.; Schwenk, K. The Mechanism of Chemical Delivery to the Vomeronasal Organs in Squamate Reptiles: A Comparative Morphological Approach. J. Exp. Zool. 2009, 311A, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Kaneoya, Y.; Tonomori, W.; Kitayama, C. Histological Features and Gα olf Expression Patterns in the Nasal Cavity of Sea Turtles. J. Anat. [CrossRef]
- Reiss, J.O.; Eisthen, H.L. Comparative Anatomy and Physiology of Chemical Senses in Amphibians. In Sensory Evolution on the Threshold: Adaptations in Secondarily Aquatic Vertebrates; University of California Press: Berkeley, CA, USA, 2008; ISBN 978-0-520-25278-3. [Google Scholar]
- Monti-Bloch, L.; Jennings-White, C.; Berliner, D.L. The Human Vomeronasal System: A Review. Ann. NY Acad. Sci. 1998, 855, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Tirindelli, R. Coding of Pheromones by Vomeronasal Receptors. Cell Tissue Res. 2021, 383, 367–386. [Google Scholar] [CrossRef]
- Shinohara, H.; Asano, T.; Kato, K. Differential Localization of G-Proteins Gi and Go in the Accessory Olfactory Bulb of the Rat. J. Neurosci. 1992, 12, 1275–1279. [Google Scholar] [CrossRef]
- Dulac, C.; Axel, R. A Novel Family of Genes Encoding Putative Pheromone Receptors in Mammals. Cell 1995, 83, 195–206. [Google Scholar] [CrossRef]
- Herrada, G.; Dulac, C. A Novel Family of Putative Pheromone Receptors in Mammals with a Topographically Organized and Sexually Dimorphic Distribution. Cell 1997, 90, 763–773. [Google Scholar] [CrossRef]
- Matsunami, H.; Buck, L.B. A Multigene Family Encoding a Diverse Array of Putative Pheromone Receptors in Mammals. Cell 1997, 90, 775–784. [Google Scholar] [CrossRef]
- Ryba, N.J.P.; Tirindelli, R. A New Multigene Family of Putative Pheromone Receptors. Neuron 1997, 19, 371–379. [Google Scholar] [CrossRef]
- Rivière, S.; Challet, L.; Fluegge, D.; Spehr, M.; Rodriguez, I. Formyl Peptide Receptor-like Proteins Are a Novel Family of Vomeronasal Chemosensors. Nature 2009, 459, 574–577. [Google Scholar] [CrossRef]
- Brennan, P.A. The Vomeronasal System. CMLS Cell. Mol. Life Sci. 2001, 58, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Papes, F.; Logan, D.W.; Stowers, L. The Vomeronasal Organ Mediates Interspecies Defensive Behaviors through Detection of Protein Pheromone Homologs. Cell 2010, 141, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.; Kubie, J.L. The Role of the Ophidian Vomeronasal System in Species-Typical Behavior. Trends Neurosci. 1984, 7, 472–477. [Google Scholar] [CrossRef]
- Placyk, Jr., J. S.; Graves, B.M. Prey Detection by Vomeronasal Chemoreception in a Plethodontid Salamander. J. Chem. Ecol. 2002, 28, 1017–1036. [Google Scholar] [CrossRef]
- Halpern, M.; Daniels, Y.; Zuri, I. The Role of the Vomeronasal System in Food Preferences of the Gray Short-Tailed Opossum, Monodelphis Domestica. Nutr. Metab. 2005, 2, 6. [Google Scholar] [CrossRef]
- Dagg, A.I.; Taub, A. Flehmen 1970, 34, 686–695. 34. [CrossRef]
- Estes, R.D. The Role of the Vomeronasal Organ in Mammalian Reproduction. Mammalia 1972, 36. [Google Scholar] [CrossRef]
- Hart, B.L. Flehmen Behavior and Vomeronasal Organ Function. In Chemical Signals in Vertebrates 3; Müller-Schwarze, D., Silverstein, R.M., Eds.; Springer: Boston, MA USA, 1983; pp. 87–103. ISBN 978-1-4757-9652-0. [Google Scholar]
- Hart, L.A.; Hart, B.L. Flehmen, Osteophagia, and Other Behaviors of Giraffes (Giraffa Giraffa Angolensis): Vomeronasal Organ Adaptation. Animals 2023, 13, 354. [Google Scholar] [CrossRef]
- Jacobson, L. Anatomisk Beskrivelse over et myt Organ i Huusdyrenes Naese. Vet. =Selsk. Skr. 1813, 2, 209–246. [Google Scholar]
- Ruysch, F. Thesaurus Anatomicus Tertius; Amstelaedami: Joannen Wolters 1703: 2013; pp. 48–49.
- Dursy, E. Zur Entwicklungsgeschichte des Kopfes des Menschen und der höheren Wirbelthiere: Mit Holzschnitten und einem Atlas von neun Kupfertafeln mit erklärendem Texte; Laupp, 1869.
- Kölliker, A. Über Die Jacobson’schen Organe Des Menschen. In Festschrift zu dem 40jährign Professoren-Jubiläum des Herrn Franz von Rinecker 31 März 1877; Leipzig, 1877; pp. 3–11.
- Balogh, C. Proceedings of the “Kaiserlichen Akademie der Wissenschaften Wien”; 1860; pp. 449–476.
- Klein, E. Memoirs: Contributions to the Minute Anatomy of the Nasal Mucous Membrane. J. Cell Sci. 1881, s2–s21, 98–113. [Google Scholar] [CrossRef]
- Piana, G. Contribuzioni Alla Conoscenza Della Strutture e Della Funzione Dell’ Organo Di Jacobson. Deusch. Zeitsch. F. Tiermed. 1882, 7, 325. [Google Scholar]
- Retzius, G. Die Riechzellen Der Ophidier in Der Riechschleimhaut Und Im Jacobson’schen Organ. Biol. Untersuch. Neue Folge 1894, 48–51. [Google Scholar]
- Gudden, B.V. Experimentaluntersuchungen Über Das Peripherische Und Centrale Nervensystem. Arch. Psychiatr. Nervenkr. 1870, 2, 693–723. [Google Scholar] [CrossRef]
- Ramón y Cajal, S.R. Textura Del Lobulo Olfativo Accesorio. Rev. Micros. 1902, 1, 141–150. [Google Scholar]
- Vandenbergh, J.G. Male Odor Accelerates Female Sexual Maturation in Mice. Endocrinology 1969, 84, 658–660. [Google Scholar] [CrossRef]
- Whitten, W.K. Modification of the Oestrous Cycle of the Mouse by External Stimuli Associated with the Male. J. Endocrinol. 1956, 13, 399–404. [Google Scholar] [CrossRef]
- Powers, J.B.; Winans, S.S. Vomeronasal Organ: Critical Role in Mediating Sexual Behavior of the Male Hamster. Science 1975, 187, 961–963. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Origin and Evolution of the Vertebrate Vomeronasal System Viewed through System-Specific Genes. Bioessays 2006, 28, 709–718. [Google Scholar] [CrossRef]
- González, A. Lungfishes, like Tetrapods, Possess a Vomeronasal System. Front. Neuroanat. 2010. [Google Scholar] [CrossRef]
- Nakamuta, S.; Nakamuta, N.; Taniguchi, K.; Taniguchi, K. Histological and Ultrastructural Characteristics of the Primordial Vomeronasal Organ in Lungfish. Anat. Rec. 2012, 295, 481–491. [Google Scholar] [CrossRef]
- Nakamuta, S.; Yamamoto, Y.; Miyazaki, M.; Sakuma, A.; Nikaido, M.; Nakamuta, N. Type 1 Vomeronasal Receptors Expressed in the Olfactory Organs of Two African Lungfish, Protopterus Annectens and Protopterus Amphibius. J. Comp. Neurol. 2023, 531, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Wittmer, C.; Nowack, C. Epithelial Crypts: A Complex and Enigmatic Olfactory Organ in African and South American Lungfish (Lepidosireniformes, Dipnoi): WITTMER and NOWACK. J. Morphol. 2017, 278, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Swaney, W.T.; Keverne, E.B. The Evolution of Pheromonal Communication. Behav. Brain Res. 2009, 200, 239–247. [Google Scholar] [CrossRef]
- Pihlström, H. Comparative Anatomy and Physiology of Chemical Senses in Aquatic Mammals. In Sensory Evolution on the Threshold: Adaptations in Secondarily Aquatic Vertebrates; Thewissen, J.G.M., Ed.; University of California Press: Berkeley, CA, USA, 2008; ISBN 978-0-520-25278-3. [Google Scholar]
- Suzuki, H.; Nishida, H.; Kondo, H.; Yoda, R.; Iwata, T.; Nakayama, K.; Enomoto, T.; Wu, J.; Moriya-Ito, K.; Miyazaki, M.; et al. A Single Pheromone Receptor Gene Conserved across 400 Million Years of Vertebrate Evolution. Mol. Biol. Evol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Breathnach, A.S. The Cetacean Central Nervous System. Biol. Rev. 1960, 35, 187–230. [Google Scholar] [CrossRef]
- McGowen, M.R.; Clark, C.; Gatesy, J. The Vestigial Olfactory Receptor Subgenome of Odontocete Whales: Phylogenetic Congruence between Gene-Tree Reconciliation and Supermatrix Methods. Syst. Biol. 2008, 57, 574–590. [Google Scholar] [CrossRef]
- Kishida, T. Olfaction of Aquatic Amniotes. Cell Tissue Res. 2021, 383, 353–365. [Google Scholar] [CrossRef]
- Halpern, M. Nasal Chemical Senses in Reptiles: Structure and Function. In Hormones, Brain, and Behaviour. Biology of the Reptilia; University of Chicago Press: Chicago, CA, USA, 1992; Volume 18, pp. 423–523. [Google Scholar]
- Schwenk, K. Of Tongues and Noses: Chemoreception in Lizards and Snakes. Trends Ecol. Evol. 1995, 10, 7–12. [Google Scholar] [CrossRef]
- Kondoh, D.; Yamamoto, Y.; Nakamuta, N.; Taniguchi, K.; Taniguchi, K. Lectin Histochemical Studies on the Olfactory Epithelium and Vomeronasal Organ in the Japanese Striped Snake, Elaphe Quadrivirgata. J. Morphol. 2010, 271, 1197–1203. [Google Scholar] [CrossRef]
- Kondoh, D.; Yamamoto, Y.; Nakamuta, N.; Taniguchi, K.; Taniguchi, K. Seasonal Changes in the Histochemical Properties of the Olfactory Epithelium and Vomeronasal Organ in the Japanese Striped Snake, Elaphe Quadrivirgata: Seasonal Changes in Snake Olfactory Organs. Anat. Histol. Embryol. 2012, 41, 41–53. [Google Scholar] [CrossRef]
- Hatanaka, T.; Matsuzaki, O. Odor Responses of the Vomeronasal System in Reeve’s Turtle, Geoclemys Reevesii. Brain Behav. Evol. 1993, 41, 183–186. [Google Scholar] [CrossRef]
- Parsons, T.S. Studies on the Comparative Embryology of the Reptilian Nose; Bull. Mus. Comp. Zool. Harvard College; 1959; Volume 120.
- Parsons, T.S. Evolution of the Nasal Structure in the Lower Tetrapods. Am. Zool. 1967, 7, 397–413. [Google Scholar] [CrossRef]
- Houck, L.D. Pheromone Communication in Amphibians and Reptiles. Annu. Rev. Physiol. 2009, 71, 161–176. [Google Scholar] [CrossRef]
- Eisthen, H.L. Presence of the Vomeronasal System in Aquatic Salamanders. Phil. Trans. R. Soc. Lond. B 2000, 355, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.; Antunes, A. Vomeronasal Receptors in Vertebrates and the Evolution of Pheromone Detection. Annu. Rev. Anim. Biosci. 2017, 5, 353–370. [Google Scholar] [CrossRef]
- Bhatnagar, K.P.; Meisami, E. Vomeronasal Organ in Bats and Primates: Extremes of Structural Variability and Its Phylogenetic Implications. Microsc. Res. Tech. 1998, 43, 465–475. [Google Scholar] [CrossRef]
- Suárez, R.; Fernández-Aburto, P.; Manger, P.R.; Mpodozis, J. Deterioration of the Gαo Vomeronasal Pathway in Sexually Dimorphic Mammals. PLoS ONE 2011, 6, e26436. [Google Scholar] [CrossRef]
- Kondoh, D.; Watanabe, K.; Nishihara, K.; Ono, Y.S.; Nakamura, K.G.; Yuhara, K.; Tomikawa, S.; Sugimoto, M.; Kobayashi, S.; Horiuchi, N.; et al. Histological Properties of Main and Accessory Olfactory Bulbs in the Common Hippopotamus. Brain Behav. Evol. 2017, 90, 224–231. [Google Scholar] [CrossRef]
- Tomiyasu, J.; Korzekwa, A.; Kawai, Y.K.; Robstad, C.A.; Rosell, F.; Kondoh, D. The Vomeronasal System in Semiaquatic Beavers. J. Anat. 2022, 241, 809–819. [Google Scholar] [CrossRef]
- Grus, W.E.; Shi, P.; Zhang, J. Largest Vertebrate Vomeronasal Type 1 Receptor Gene Repertoire in the Semiaquatic Platypus. Mol. Biol. Evol. 2007, 24, 2153–2157. [Google Scholar] [CrossRef]
- Cartmill, M. Rethinking Primate Origins: The Characteristic Primate Traits Cannot Be Explained Simply as Adaptations to Arboreal Life. Science 1974, 184, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Heesy, C.P.; Ross, C.F. Evolution of Activity Patterns and Chromatic Vision in Primates: Morphometrics, Genetics and Cladistics. J. Hum. Evol. 2001, 40, 111–149. [Google Scholar] [CrossRef]
- Wang, G.; Shi, P.; Zhu, Z.; Zhang, Y. -p. More Functional V1R Genes Occur in Nest-Living and Nocturnal Terricolous Mammals. Genome Biol. Evol. 2010, 2, 277–283. [Google Scholar] [CrossRef]
- Garrett, E.C.; Dennis, J.C.; Bhatnagar, K.P.; Durham, E.L.; Burrows, A.M.; Bonar, C.J.; Steckler, N.K.; Morrison, E.E.; Smith, T.D. The Vomeronasal Complex of Nocturnal Strepsirhines and Implications for the Ancestral Condition in Primates: Vomeronasal Complex of Nocturnal Strepsirhines. Anat. Rec. 2013, 296, 1881–1894. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.D.; Garrett, E.C.; Bhatnagar, K.P.; Bonar, C.J.; Bruening, A.E.; Dennis, J.C.; Kinznger, J.H.; Johnson, E.W.; Morrison, E.E. The Vomeronasal Organ of New World Monkeys (Platyrrhini). Anat. Rec. 2011, 294, 2158–2178. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.D.; Siegel, M.I.; Bhatnagar, K.P. Reappraisal of the Vomeronasal System of Catarrhine Primates: Ontogeny, Morphology, Functionality, and Persisting Questions. Anat. Rec. 2001, 265, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M. Human Vomeronasal Organ Function: A Critical Review of Best and Worst Cases. Chem. Senses 2001, 26, 433–445. [Google Scholar] [CrossRef]
- Zhang, J.; Webb, D.M. Evolutionary Deterioration of the Vomeronasal Pheromone Transduction Pathway in Catarrhine Primates. Proc. Natl. Acad. Sci. USA 2003, 100, 8337–8341. [Google Scholar] [CrossRef]
- Witt, M.; Hummel, T. Vomeronasal Versus Olfactory Epithelium: Is There a Cellular Basis for Human Vomeronasal Perception. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 2006; Volume 248, pp. 209–259. ISBN 978-0-12-364652-1. [Google Scholar]
- Smith, T.D.; Laitman, J.T.; Bhatnagar, K.P. The Shrinking Anthropoid Nose, the Human Vomeronasal Organ, and the Language of Anatomical Reduction: The Shrinking Anthropoid Nose. Anat. Rec. 2014, 297, 2196–2204. [Google Scholar] [CrossRef]
- Negus, V.E. The Organ of Jacobson. J. Anat. 1956, 90, 515–519. [Google Scholar]
- Salazar, I.; Sanchez-Quinteiro, P. Supporting Tissue and Vasculature of the Mammalian Vomeronasal Organ: The Rat as a Model. Microsc. Res. Tech. 1998, 41, 492–505. [Google Scholar] [CrossRef]
- Døving, K.B.; Trotier, D. Structure and Function of the Vomeronasal Organ. J. Exp. Biol. 1998, 201, 2913–2925. [Google Scholar] [CrossRef] [PubMed]
- Menco, B.Ph.M. Ultrastructural Aspects of Olfactory Signaling. Chem. Senses 1997, 22, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Menco, B.Ph.M.; Carr, V. McM.; Ezeh, P.I.; Liman, E.R.; Yankova, M.P. Ultrastructural Localization of G-Proteins and the Channel Protein TRP2 to Microvilli of Rat Vomeronasal Receptor Cells. J. Comp. Neurol. 2001, 438, 468–489. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sanchez-Quinteiro, P.; Cifuentes, J.M.; De Troconiz, P.F. General Organization of the Perinatal and Adult Accessory Olfactory Bulb in Mice. Anat. Rec. Part. A Discov. Mol. Cell. Evol. Biol. 2006, 288A, 1009–1025. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, L.D.; Reiss, J.O.; Pozzi, A.G. Olfactory Subsystems in the Peripheral Olfactory Organ of Anuran Amphibians. Cell Tissue Res. 2021, 383, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, A.S.; Kühnel, W. Morphological Evidence for a Direct Innervation of the Mouse Vomeronasal Glands. Cell Tissue Res. 1987, 247. [Google Scholar] [CrossRef]
- Takami, S.; Getchell, M.L.; Getchell, T.V. Resolution of Sensory and Mucoid Glycoconjugates with Terminal ?-Galactose Residues in the Mucomicrovillar Complex of the Vomeronasal Sensory Epithelium by Dual Confocal Laser Scanning Microscopy. Cell Tissue Res. 1995, 280, 211–216. [Google Scholar] [CrossRef]
- Kondoh, D.; Tomiyasu, J.; Itakura, R.; Sugahara, M.; Yanagawa, M.; Watanabe, K.; Alviola, P.A.; Yap, S.A.; Cosico, E.A.; Cruz, F.A.; et al. Comparative Histological Studies on Properties of Polysaccharides Secreted by Vomeronasal Glands of Eight Laurasiatheria Species. Acta Histochem. 2020, 122, 151515. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez Quinteiro, P.; Cifuentes, J.M.; Fernández, P.; Lombardero, M. Distribution of the Arterial Supply to the Vomeronasal Organ in the Cat. Anat. Rec. 1997, 247, 129–136. [Google Scholar] [CrossRef]
- Meredith, M.; O’Connell, R.J. Efferent Control of Stimulus Access to the Hamster Vomeronasal Organ. J. Physiol. 1979, 286, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Eccles, R. Autonomic Innervation of the Vomeronasal Organ of the Cat. Physiol. Behav. 1982, 28, 1011–1015. [Google Scholar] [CrossRef] [PubMed]
- Meredith, M. Chronic Recording of Vomeronasal Pump Activation in Awake Behaving Hamsters. Physiol. Behav. 1994, 56, 345–354. [Google Scholar] [CrossRef]
- Halpern, M.; Shapiro, L.S.; Jia, C. Heterogeneity in the Accessory Olfactory System. Chem. Senses 1998, 23, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Leinders-Zufall, T.; Lane, A.P.; Puche, A.C.; Ma, W.; Novotny, M.V.; Shipley, M.T.; Zufall, F. Ultrasensitive Pheromone Detection by Mammalian Vomeronasal Neurons. Nature 2000, 405, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Spehr, M.; Kelliher, K.R.; Li, X.-H.; Boehm, T.; Leinders-Zufall, T.; Zufall, F. Essential Role of the Main Olfactory System in Social Recognition of Major Histocompatibility Complex Peptide Ligands. J. Neurosci. 2006, 26, 1961–1970. [Google Scholar] [CrossRef]
- Liman, E.R.; Corey, D.P.; Dulac, C. TRP2: A Candidate Transduction Channel for Mammalian Pheromone Sensory Signaling. Proc. Natl. Acad. Sci. USA 1999, 96, 5791–5796. [Google Scholar] [CrossRef]
- Leypold, B.G.; Yu, C.R.; Leinders-Zufall, T.; Kim, M.M.; Zufall, F.; Axel, R. Altered Sexual and Social Behaviors in Trp2 Mutant Mice. Proc. Natl. Acad. Sci. USA 2002, 99, 6376–6381. [Google Scholar] [CrossRef]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of Sex Discrimination and Male-Male Aggression in Mice Deficient for TRP2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef]
- Rodriguez, I.; Greer, C.A.; Mok, M.Y.; Mombaerts, P. A Putative Pheromone Receptor Gene Expressed in Human Olfactory Mucosa. Nat. Genet. 2000, 26, 18–19. [Google Scholar] [CrossRef]
- Baum, M.J.; Kelliher, K.R. Complementary Roles of the Main and Accessory Olfactory Systems in Mammalian Mate Recognition. Annu. Rev. Physiol. 2009, 71, 141–160. [Google Scholar] [CrossRef]
- Schneider, N.Y.; Fletcher, T.P.; Shaw, G.; Renfree, M.B. Goα Expression in the Vomeronasal Organ and Olfactory Bulb of the Tammar Wallaby. Chem. Senses 2012, 37, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Suárez, R.; Mpodozis, J. Heterogeneities of Size and Sexual Dimorphism between the Subdomains of the Lateral-Innervated Accessory Olfactory Bulb (AOB) of Octodon Degus (Rodentia: Hystricognathi). Behav. Brain Res. 2009, 198, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.; Shapiro, L.S.; Jia, C. Differential Localization of G Proteins in the Opossum Vomeronasal System. Brain Res. 1995, 677, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Trask, B.J. V2R Gene Families Degenerated in Primates, Dog and Cow, but Expanded in Opossum. Trends Genet. 2007, 23, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P. Morphological and Immunohistochemical Features of the Vomeronasal System in Dogs. Anat. Rec. 2013, 296, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Moriya-Ito, K.; Hayakawa, T.; Suzuki, H.; Hagino-Yamagishi, K.; Nikaido, M. Evolution of Vomeronasal Receptor 1 (V1R) Genes in the Common Marmoset (Callithrix Jacchus). Gene 2018, 642, 343–353. [Google Scholar] [CrossRef]
- Kishimoto, J.; Keverne, E.B.; Emson, P.C. Calretinin, Calbindin-D28k and Parvalbumin-like Immunoreactivity in Mouse Chemoreceptor Neurons. Brain Res. 1993, 610, 325–329. [Google Scholar] [CrossRef]
- Verhaagen, J.; Oestreicher, A.; Gispen, W.; Margolis, F. The Expression of the Growth Associated Protein B50/GAP43 in the Olfactory System of Neonatal and Adult Rats. J. Neurosci. 1989, 9, 683–691. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez Quinteiro, P. Lectin Binding Patterns in the Vomeronasal Organ and Accessory Olfactory Bulb of the Rat. Anat. Embryol. 1998, 198, 331–339. [Google Scholar] [CrossRef]
- Ichikawa, M.; Osada, T.; Ikai, A. Bandeiraea Simplicifolia Lectin I and Vicia Villosa Agglutinin Bind Specifically to the Vomeronasal Axons in the Accessory Olfactory Bulb of the Rat. Neurosci. Res. 1992, 13, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Soria, X.; Levitin, M.O.; Saraiva, L.R.; Logan, D.W. The Olfactory Transcriptomes of Mice. PLoS Genet. 2014, 10, e1004593. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Zhang, J. Comparative Genomic Analysis Identifies an Evolutionary Shift of Vomeronasal Receptor Gene Repertoires in the Vertebrate Transition from Water to Land. Genome Res. 2007, 17, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Francia, S.; Silvotti, L.; Ghirardi, F.; Catzeflis, F.; Percudani, R.; Tirindelli, R. Evolution of Spatially Coexpressed Families of Type-2 Vomeronasal Receptors in Rodents. Genome Biol. Evol. 2015, 7, 272–285. [Google Scholar] [CrossRef]
- Yohe, L.R.; Davies, K.T.J.; Rossiter, S.J.; Dávalos, L.M. Expressed Vomeronasal Type-1 Receptors (V1rs) in Bats Uncover Conserved Sequences Underlying Social Chemical Signaling. Genome Biol. Evol. 2019, 11, 2741–2749. [Google Scholar] [CrossRef] [PubMed]
- Haga, S.; Hattori, T.; Sato, T.; Sato, K.; Matsuda, S.; Kobayakawa, R.; Sakano, H.; Yoshihara, Y.; Kikusui, T.; Touhara, K. The Male Mouse Pheromone ESP1 Enhances Female Sexual Receptive Behaviour through a Specific Vomeronasal Receptor. Nature 2010, 466, 118–122. [Google Scholar] [CrossRef]
- Silva, L.; Mendes, T.; Antunes, A. Acquisition of Social Behavior in Mammalian Lineages Is Related with Duplication Events of FPR Genes. Genomics 2020, 112, 2778–2783. [Google Scholar] [CrossRef]
- Oboti, L.; Ibarra-Soria, X.; Pérez-Gómez, A.; Schmid, A.; Pyrski, M.; Paschek, N.; Kircher, S.; Logan, D.W.; Leinders-Zufall, T.; Zufall, F.; et al. Pregnancy and Estrogen Enhance Neural Progenitor-Cell Proliferation in the Vomeronasal Sensory Epithelium. BMC Biol. 2015, 13, 104. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Gullón, J.; Quintela, L.; Sánchez-Quinteiro, P.; Martínez, P.; Robledo, D. Sex Separation Unveils the Functional Plasticity of the Vomeronasal Organ in Rabbits. Front. Mol. Neurosci. 2022, 15, 1034254. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Robledo, D.; Fernández, C.; Gullón, J.; Quintela, L.; Sánchez-Quinteiro, P.; Martínez, P. Analysis of the Vomeronasal Organ Transcriptome Reveals Variable Gene Expression Depending on Age and Function in Rabbits. Genomics 2021, 113, 2240–2252. [Google Scholar] [CrossRef]
- Broom, R. A Contribution to the Comparative Anatomy of the Mammalian Organ of Jacobson. Trans. R. Soc. Edinb. 1900, 39, 231–255. [Google Scholar] [CrossRef]
- Wöhrmann-Repenning, A. [Phylogenetic aspects of the Jacobson’s organ and nasopalatine duct topography in insectivores, primates, Tupaia and Didelphis]. Anat. Anz. 1984, 157, 137–149. [Google Scholar] [PubMed]
- Sánchez-Villagra, M.R. Ontogenetic and Phylogenetic Transformations of the Vomeronasal Complex and Nasal Floor Elements in Marsupial Mammals. Zool. J. Linn. Soc. 2001, 131, 459–479. [Google Scholar] [CrossRef]
- Vaccarezza, O.L.; Sepich, L.N.; Tramezzani, J.H. The Vomeronasal Organ of the Rat. J. Anat. 1981, 132, 167–185. [Google Scholar] [PubMed]
- Mechin, V.; Pageat, P.; Teruel, E.; Asproni, P. Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice. Animals 2021, 11, 1211. [Google Scholar] [CrossRef]
- Weiler, E.; McCulloch, M.A.; Farbman, A.I. Proliferation in the Vomeronasal Organ of the Rat during Postnatal Development. Eur. J. Neurosci. 1999, 11, 700–711. [Google Scholar] [CrossRef]
- Giacobini, P.; Benedetto, A.; Tirindelli, R.; Fasolo, A. Proliferation and Migration of Receptor Neurons in the Vomeronasal Organ of the Adult Mouse. Dev. Brain Res. 2000, 123, 33–40. [Google Scholar] [CrossRef]
- Ibokwe, C.; Okpe, G.C. Morphological Studies Of Vomeronasal Organ In The Wild Juvenile Red-Flanked Duiker Cephalophus Rufilatus (GRAY 1864). Anim. Res. Int. 2009, 6. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Yang, C.; He, Y. Histological Structure of the Vomeronasal Organ and Accessory Olfactory Bulb and the Seasonal Changes of Olfactory Bulb C-Fos Expression in Spermophilus Dauricus. Acta Theriol. Sin. 2021, 41, 685. [Google Scholar] [CrossRef]
- Oikawa, T.; Shimamura, K.; Saito, T.R.; Taniguchi, K. Fine Structure of the Vomeronasal Organ in the Chinchilla (Chinchilla Laniger). Exp. Anim. 1994, 43, 487–497. [Google Scholar] [CrossRef]
- Jurcisek, J.A.; Durbin, J.E.; Kusewitt, D.F.; Bakaletz, L.O. Anatomy of the Nasal Cavity in the Chinchilla. Cells Tissues Organs 2003, 174, 136–152. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.D.; Alport, L.J.; Burrows, A.M.; Bhatnagar, K.P.; Dennis, J.C.; Tuladhar, P.; Morrison, E.E. Perinatal Size and Maturation of the Olfactory and Vomeronasal Neuroepithelia in Lorisoids and Lemuroids. Am. J. Primatol. 2007, 69, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.C.; Stilwell, N.K.; Smith, T.D.; Park, T.J.; Bhatnagar, K.P.; Morrison, E.E. Is the Mole Rat Vomeronasal Organ Functional? Anat. Rec. 2020, 303, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Luckhaus, G. [Light and electron microscopic findings in the epithelial lamina of the vomeronasal organ of the rabbit]. Anat. Anz. 1969, 124, 477–489. [Google Scholar]
- Mahdy, E.; behery, E.; Mohamed, S. Comparative Morpho-Histological Analysis on the Vomeronasal Organ and the Accessory Olfactory Bulb in Balady Dogs (Canis Familiaris) and New Zealand Rabbits (Oryctolagus Cuniculus). J. Adv. Vet. Anim. Res. 2019, 6, 506. [Google Scholar] [CrossRef]
- Elgayar, S.A.M.; Eltony, S.A.; Othman, M.A. Morphology of Non-Sensory Epithelium during Post-Natal Development of the Rabbit Vomeronasal Organ. Anat. Histol. Embryol. 2014, 43, 282–293. [Google Scholar] [CrossRef]
- Schneider, N.Y.; Fletcher, T.P.; Shaw, G.; Renfree, M.B. The Vomeronasal Organ of the Tammar Wallaby. J. Anat. 2008, 213, 93–105. [Google Scholar] [CrossRef]
- Poran, N.S. Vomeronasal Organ and Its Associated Structures in the OpossumMonodelphis Domestica. Microsc. Res. Tech. 1998, 43, 500–510. [Google Scholar] [CrossRef]
- Schneider, N.Y. The Development of the Olfactory Organs in Newly Hatched Monotremes and Neonate Marsupials: Olfaction in Monotremes and Marsupials. J. Anat. 2011, 219, 229–242. [Google Scholar] [CrossRef]
- Ashwell, K.W.S. Development of the Olfactory Pathways in Platypus and Echidna. Brain Behav. Evol. 2012, 79, 45–56. [Google Scholar] [CrossRef]
- Kondoh, D.; Tanaka, Y.; Kawai, Y.K.; Mineshige, T.; Watanabe, K.; Kobayashi, Y. Morphological and Histological Features of the Vomeronasal Organ in African Pygmy Hedgehog (Atelerix Albiventris). Animals 2021, 11, 1462. [Google Scholar] [CrossRef]
- Aland, R.C.; Gosden, E.; Bradley, A.J. Seasonal Morphometry of the Vomeronasal Organ in the Marsupial Mouse, Antechinus Subtropicus. J. Morphol. 2016, 277, 1517–1530. [Google Scholar] [CrossRef]
- Dennis, J.C.; Allgier, J.G.; Desouza, L.S.; Eward, W.C.; Morrison, E.E. Immunohistochemistry of the Canine Vomeronasal Organ. J. Anat. 2003, 203, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sanchez Quinteiro, P.; Cifuentes, J.M.; Garcia Caballero, T. The Vomeronasal Organ of the Cat. J. Anat. 1996, 188 (Pt. 2), 445–454. [Google Scholar]
- Tomiyasu, J.; Kondoh, D.; Sakamoto, H.; Matsumoto, N.; Sasaki, M.; Kitamura, N.; Haneda, S.; Matsui, M. Morphological and Histological Features of the Vomeronasal Organ in the Brown Bear. J. Anat. 2017, 231, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kelliher, K.R.; Baum, M.J.; Meredith, M. The Ferret’s Vomeronasal Organ and Accessory Olfactory Bulb: Effect of Hormone Manipulation in Adult Males and Females. Anat. Rec. 2001, 263, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Lombardero, M.; Sánchez-Quinteiro, P.; Roel, P.; Cifuentes, J.M. Origin and Regional Distribution of the Arterial Vessels of the Vomeronasal Organ in the Sheep. A Methodological Investigation with Scanning Electron Microscopy and Cutting-Grinding Technique. Ann. Anat. Anat. Anz. 1998, 180, 181–187. [Google Scholar] [CrossRef]
- Salazar, I.; Quinteiro, P.S.; Cifuentes, J.M. The Soft-Tissue Components of the Vomeronasal Organ in Pigs, Cows and Horses. Anat. Histol. Embryol. 1997, 26, 179–186. [Google Scholar] [CrossRef]
- Salazar, I.; Quinteiro, P.S.; Alemañ, N.; Cifuentes, J.M.; Troconiz, P.F. Diversity of the Vomeronasal System in Mammals: The Singularities of the Sheep Model. Microsc. Res. Tech. 2007, 70, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, M.; Shin, T.; Kang, M.S. Fine Structure of the Vomeronasal Sensory Epithelium of Korean Goats (Capra Hircus). J. Reprod. Dev. 1999, 45, 81–89. [Google Scholar] [CrossRef]
- Yang, W.; Choi, Y.; Park, C.; Lee, K.-H.; Ahn, M.; Kang, W.; Heo, S.-D.; Kim, J.; Shin, T. Histological and Lectin Histochemical Studies in the Vomeronasal Organ of the Korean Black Goat, Capra Hircus Coreanae. Acta Histochem. 2021, 123, 151684. [Google Scholar] [CrossRef]
- Park, C.; Ahn, M.; Lee, J.-Y.; Lee, S.; Yun, Y.; Lim, Y.-K.; Taniguchi, K.; Shin, T. A Morphological Study of the Vomeronasal Organ and the Accessory Olfactory Bulb in the Korean Roe Deer, Capreolus Pygargus. Acta Histochem. 2014, 116, 258–264. [Google Scholar] [CrossRef]
- Vedin, V.; Eriksson, B.; Berghard, A. Organization of the Chemosensory Neuroepithelium of the Vomeronasal Organ of the Scandinavian Moose Alces Alces. Brain Res. 2010, 1306, 53–61. [Google Scholar] [CrossRef]
- Kondoh, D.; Nakamura, K.G.; Ono, Y.S.; Yuhara, K.; Bando, G.; Watanabe, K.; Horiuchi, N.; Kobayashi, Y.; Sasaki, M.; Kitamura, N. Histological Features of the Vomeronasal Organ in the Giraffe, Giraffa Camelopardalis: KONDOH et al. Microsc. Res. Tech. 2017, 80, 652–656. [Google Scholar] [CrossRef]
- Smith, T.D.; Bhatnagar, K.P.; Shimp, K.L.; Kinzinger, J.H.; Bonar, C.J.; Burrows, A.M.; Mooney, M.P.; Siegel, M.I. Histological Definition of the Vomeronasal Organ in Humans and Chimpanzees, with a Comparison to Other Primates. Anat. Rec. 2002, 267, 166–176. [Google Scholar] [CrossRef]
- Meisami, E.; Bhatnagar, K.P. Structure and Diversity in Mammalian Accessory Olfactory Bulb. Microsc. Res. Tech. 1998, 43, 476–499. [Google Scholar] [CrossRef]
- Takami, S.; Graziadei, P.P.C. Morphological Complexity of the Glomerulus in the Rat Accessory Olfactory Bulb-a Golgi Study. Brain Res. 1990, 510, 339–342. [Google Scholar] [CrossRef]
- Nakajima, M.; Tsuruta, M.; Mori, H.; Nishikawa, C.; Okuyama, S.; Furukawa, Y. A Comparative Study of Axon-Surrounding Cells in the Two Nasal Nerve Tracts from Mouse Olfactory Epithelium and Vomeronasal Organ. Brain Res. 2013, 1503, 16–23. [Google Scholar] [CrossRef]
- Vassar, R.; Chao, S.K.; Sitcheran, R.; Nunez, J.M.; Vosshall, L.B.; Axel, R. Topographic Organization of Sensory Projections to the Olfactory Bulb. Cell 1994, 79, 981–991. [Google Scholar] [CrossRef]
- Del Punta, K.; Puche, A.; Adams, N.C.; Rodriguez, I.; Mombaerts, P. A Divergent Pattern of Sensory Axonal Projections Is Rendered Convergent by Second-Order Neurons in the Accessory Olfactory Bulb. Neuron 2002, 35, 1057–1066. [Google Scholar] [CrossRef]
- Crosby, E.C.; Humphrey, T. Studies of the Vertebrate Telencephalon. II. The Nuclear Pattern of the Anterior Olfactory Nucleus, Tuberculum Olfactorium and the Amygdaloid Complex in Adult Man. J. Comp. Neurol. 1941, 74, 309–352. [Google Scholar] [CrossRef]
- Allison, A.C. The Structure of the Olfactory Bulb and Its Relationship to the Olfactory Pathways in the Rabbit and the Rat. J. Comp. Neurol. 1953, 98, 309–353. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P. A Detailed Morphological Study of the Vomeronasal Organ and the Accessory Olfactory Bulb of Cats. Microsc. Res. Tech. 2011, 74, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Larriva-Sahd, J. The Accessory Olfactory Bulb in the Adult Rat: A Cytological Study of Its Cell Types, Neuropil, Neuronal Modules, and Interactions with the Main Olfactory System. J. Comp. Neurol. 2008, 510, 309–350. [Google Scholar] [CrossRef]
- Switzer, III. , R.C.; Johnson, J.I.; Kirsch, J.A.W. Phylogeny Through Brain Traits. Brain Behav. Evol. 1980, 17, 339–363. [Google Scholar] [CrossRef]
- Baker, H.; Towle, A.C.; Margolis, F.L. Differential Afferent Regulation of Dopaminergic and GABAergic Neurons in the Mouse Main Olfactory Bulb. Brain Res. 1988, 450, 69–80. [Google Scholar] [CrossRef]
- Gouda, M.; Matsutani, S.; Senba, E.; Tohyama, M. Peptidergic Granule Cell Populations in the Rat Main and Accessory Olfactory Bulb. Brain Res. 1990, 512, 339–342. [Google Scholar] [CrossRef]
- McLean, J.H.; Shipley, M.T. Neuroanatomical Substrates of Olfaction. In Science of Olfaction; Serby, M.J., Chobor, K.L., Eds.; Springer: New York, NY, USA, 1992; pp. 126–171. ISBN 978-1-4612-7690-6. [Google Scholar]
- Frahm, H.D.; Stephan, H.; Baron, G. Comparison of Brain Structure Volumes in Insectivora and Primates. V. Area Striata (AS). J. Hirnforsch. 1984, 25, 537–557. [Google Scholar]
- Bonfanti, L.; Peretto, P.; Merighi, A.; Fasolo, A. Newly-Generated Cells from the Rostral Migratory Stream in the Accessory Olfactory Bulb of the Adult Rat. Neuroscience 1997, 81, 489–502. [Google Scholar] [CrossRef]
- Defteralı, Ç.; Moreno-Estellés, M.; Crespo, C.; Díaz-Guerra, E.; Díaz-Moreno, M.; Vergaño-Vera, E.; Nieto-Estévez, V.; Hurtado-Chong, A.; Consiglio, A.; Mira, H.; et al. Neural Stem Cells in the Adult Olfactory Bulb Core Generate Mature Neurons in Vivo. Stem Cells 2021, 39, 1253–1269. [Google Scholar] [CrossRef]
- Inaki, K.; Nishimura, S.; Nakashiba, T.; Itohara, S.; Yoshihara, Y. Laminar Organization of the Developing Lateral Olfactory Tract Revealed by Differential Expression of Cell Recognition Molecules. J. Comp. Neurol. 2004, 479, 243–256. [Google Scholar] [CrossRef]
- Broadwell, R.D. Olfactory Relationships of the Telencephlaon and Diencephalon in the Rabbit. 1. An Autoradiographic Study of the Efferent Connections of the Main and Accessory Olfactory Bulbs. J. Comp. Neurol. 1975, 163, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Schwob, J.E.; Price, J.L. The Development of Axonal Connections in the Central Olfactory System of Rats. J. Comp. Neurol. 1984, 223, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Takami, S.; Fernandez, G.D.; Graziadei, P.P.C. The Morphology of GABA-Immunoreactive Neurons in the Accessory Olfactory Bulb of Rats. Brain Res. 1992, 588, 317–323. [Google Scholar] [CrossRef]
- Takami, S.; Graziadei, P.P.C. Light Microscopic Golgi Study of Mitral/Tufted Cells in the Accessory Olfactory Bulb of the Adult Rat. J. Comp. Neurol. 1991, 311, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Hinds, J.W. Autoradiographic Study of Histogenesis in the Mouse Olfactory Bulb I. Time of Origin of Neurons and Neuroglia. J. Comp. Neurol. 1968, 134, 287–304. [Google Scholar] [CrossRef]
- Rall, W.; Shepherd, G.M.; Reese, T.S.; Brightman, M.W. Dendrodendritic Synaptic Pathway for Inhibition in the Olfactory Bulb. Exp. Neurol. 1966, 14, 44–56. [Google Scholar] [CrossRef]
- Martín-López, E.; Corona, R.; López-Mascaraque, L. Postnatal Characterization of Cells in the Accessory Olfactory Bulb of Wild Type and Reeler Mice. Front. Neuroanat. 2012, 6. [Google Scholar] [CrossRef]
- Jia, C.; Chen, W.R.; Shepherd, G.M. Synaptic Organization and Neurotransmitters in the Rat Accessory Olfactory Bulb. J. Neurophysiol. 1999, 81, 345–355. [Google Scholar] [CrossRef]
- Del Punta, K. Sequence Diversity and Genomic Organization of Vomeronasal Receptor Genes in the Mouse. Genome Res. 2000, 10, 1958–1967. [Google Scholar] [CrossRef]
- Segovia, S.; Orensanz, L.M.; Valencia, A.; Guillamón, A. Effects of Sex Steroids on the Development of the Accessory Olfactory Bulb in the Rat: A Volumetric Study. Dev. Brain Res. 1984, 16, 312–314. [Google Scholar] [CrossRef]
- Yokosuka, M. Histological Properties of the Glomerular Layer in the Mouse Accessory Olfactory Bulb. Exp. Anim. 2012, 61, 13–24. [Google Scholar] [CrossRef]
- Frahm, H.D.; Bhatnagar, K.P. Comparative Morphology of the Accessory Olfactory Bulb in Bats. J. Anat. 1980, 130, 349–365. [Google Scholar]
- Jia, C.; Halpern, M. Calbindin D28k, Parvalbumin, and Calretinin Immunoreactivity in the Main and Accessory Olfactory Bulbs of the Gray Short-Tailed Opossum,Monodelphis Domestica. J. Morphol. 2004, 259, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Can Domestication Shape Canidae Brain Morphology? The Accessory Olfactory Bulb of the Red Fox as a Case in Point. Ann. Anat. Anat. Anz. 2022, 240, 151881. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; López-Callejo, L.N.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox. Animals 2022, 12, 1079. [Google Scholar] [CrossRef] [PubMed]
- Dzięcioł, M.; Podgórski, P.; Stańczyk, E.; Szumny, A.; Woszczyło, M.; Pieczewska, B.; Niżański, W.; Nicpoń, J.; Wrzosek, M.A. MRI Features of the Vomeronasal Organ in Dogs (Canis Familiaris). Front. Vet. Sci. 2020, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Muñiz-de Miguel, S.; Barreiro-Vázquez, J.D.; Sánchez-Quinteiro, P.; Ortiz-Leal, I.; González-Martínez, Á. Behavioural Disorder in a Dog with Congenital Agenesis of the Vomeronasal Organ and the Septum Pellucidum. Vet. Rec. Case Rep. 2023, 11, e571. [Google Scholar] [CrossRef]
- Pageat, P.; Gaultier, E. Current Research in Canine and Feline Pheromones. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 187–211. [Google Scholar] [CrossRef]
- Puglisi, I.; Masucci, M.; Cozzi, A.; Teruel, E.; Navarra, M.; Cirmi, S.; Pennisi, M.G.; Siracusa, C. Effects of a Novel Gel Formulation of Dog Appeasing Pheromone (DAP) on Behavioral and Physiological Stress Responses in Dogs Undergoing Clinical Examination. Animals 2022, 12, 2472. [Google Scholar] [CrossRef]
- Asproni, P.; Cozzi, A.; Verin, R.; Lafont-Lecuelle, C.; Bienboire-Frosini, C.; Poli, A.; Pageat, P. Pathology and Behaviour in Feline Medicine: Investigating the Link between Vomeronasalitis and Aggression. J. Feline Med. Surg. 2016, 18, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Asproni, P.; Mainau, E.; Cozzi, A.; Carreras, R.; Bienboire-Frosini, C.; Teruel, E.; Pageat, P. Is There a Link between Vomeronasalitis and Aggression in Stable Social Groups of Female Pigs? Animals 2022, 12, 303. [Google Scholar] [CrossRef] [PubMed]
- Mechin, V.; Asproni, P.; Bienboire-Frosini, C.; Cozzi, A.; Chabaud, C.; Arroub, S.; Mainau, E.; Nagnan-Le Meillour, P.; Pageat, P. Inflammation Interferes with Chemoreception in Pigs by Altering the Neuronal Layout of the Vomeronasal Sensory Epithelium. Front. Vet. Sci. 2022, 9, 936838. [Google Scholar] [CrossRef]
- Mechin, V.; Pageat, P.; Boutry, M.; Teruel, E.; Portalier, C.; Asproni, P. Does the Environmental Air Impact the Condition of the Vomeronasal Organ? A Mouse Model for Intensive Farming. Animals 2023, 13, 1902. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Cifuentes, J.M.; Quinteiro, P.S.; Caballero, G. The Vomeronasal System of the Mink, Mustela Vison. I. The Vomeronasal Organ. Funct. Dev. Morphol. 1994, 4, 113–117. [Google Scholar] [PubMed]
- Salazar, I.; Quinteiro, P.S.; Cifuentes, J.M.; Lombardero, M. The Accessory Olfactory Bulb of the Mink, Mustela Vison: A Morphological and Lectin Histochemical Study. Anatom Histol. Embryol. 1998, 27, 297–300. [Google Scholar] [CrossRef]
- Salazar, I.; Lombardero, M.; Alemañ, N.; Sánchez Quinteiro, P. Development of the Vomeronasal Receptor Epithelium and the Accessory Olfactory Bulb in Sheep. Microsc. Res. Tech. 2003, 61, 438–447. [Google Scholar] [CrossRef]
- Tirindelli, R.; Mucignat-Caretta, C.; Ryba, N.J.P. Molecular Aspects of Pheromonal Communication via the Vomeronasal Organ of Mammals. Trends Neurosci. 1998, 21, 482–486. [Google Scholar] [CrossRef]
- Alonso, J.R.; Briñón, J.G.; Crespo, C.; Bravo, I.G.; Arévalo, R.; Aijón, J. Chemical Organization of the Macaque Monkey Olfactory Bulb: II. Calretinin, Calbindin D-28k, Parvalbumin, and Neurocalcin Immunoreactivity: Calcium-Binding Proteins in the Monkey OB. J. Comp. Neurol. 2001, 432, 389–407. [Google Scholar] [CrossRef]
- Briñón, J.G.; Weruaga, E.; Crespo, C.; Porteros, A.; Arévalo, R.; Aijón, J.; Alonso, J.R. Calretinin-, Neurocalcin-, and Parvalbumin-Immunoreactive Elements in the Olfactory Bulb of the Hedgehog (Erinaceus Europaeus). J. Comp. Neurol. 2001, 429, 554–570. [Google Scholar] [CrossRef]
- Weruaga, E. A Sexually Dimorphic Group of Atypical Glomeruli in the Mouse Olfactory Bulb. Chem. Senses 2001, 26, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Smithson, L.J.; Kawaja, M.D. A Comparative Examination of Biomarkers for Olfactory Ensheathing Cells in Cats and Guinea Pigs. Brain Res. 2009, 1284, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Dehmelt, L.; Halpain, S. The MAP2/Tau Family of Microtubule-Associated Proteins. Genome Biol. 2005, 6, 204. [Google Scholar] [CrossRef]
- Benowitz, L.I.; Perrone-Bizzozero, N.I.; Neve, R.L.; Rodriguez, W. Chapter 26 GAP-43 as a Marker for Structural Plasticity in the Mature CNS. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 1990; Volume 86, pp. 309–320. ISBN 978-0-444-81121-9. [Google Scholar]
- Shapiro, L.S.; Roland, R.M.; Halpern, M. Development of Olfactory Marker Protein and N-CAM Expression in Chemosensory Systems of the Opossum,Monodelphis Domestica. J. Morphol. 1997, 234, 109–129. [Google Scholar] [CrossRef]
- Mugnaini, E.; Oertel, W.H.; Wouterlood, F.F. Immunocytochemical Localization of GABA Neurons and Dopamine Neurons in the Rat Main and Accessory Olfactory Bulbs. Neurosci. Lett. 1984, 47, 221–226. [Google Scholar] [CrossRef]
- Le Jeune, H.; Aubert, I.; Jourdan, F.; Quirion, R. Comparative Laminar Distribution of Various Autoradiographic Cholinergic Markers in Adult Rat Main Olfactory Bulb. J. Chem. Neuroanat. 1995, 9, 99–112. [Google Scholar] [CrossRef]
- Woo, C.C.; Leon, M. Distribution and Development of ?-Adrenergic Receptors in the Rat Olfactory Bulb. J. Comp. Neurol. 1995, 352, 1–10. [Google Scholar] [CrossRef]
- Huang, Z.; Thiebaud, N.; Fadool, D.A. Differential Serotonergic Modulation across the Main and Accessory Olfactory Bulbs: Differential Serotonin Modulation in Olfactory System. J. Physiol. 2017, 595, 3515–3533. [Google Scholar] [CrossRef]
- Matsutani, S.; Senba, E.; Tohyama, M. Neuropeptide- and Neurotransmitter-Related Immunoreactivities in the Developing Rat Olfactory Bulb. J. Comp. Neurol. 1988, 272, 331–342. [Google Scholar] [CrossRef]
- Lundberg, L.-M.; Alm, P.; Wharton, J.; Polak, J.M. Protein Gene Product 9.5 (PGP 9.5): A New Neuronal Marker Visualizing the Whole Uterine Innervation and Pregnancy-Induced and Developmental Changes in the Guinea Pig. Histochemistry 1988, 90, 9–17. [Google Scholar] [CrossRef]
- Nacher, J.; Crespo, C.; McEwen, B.S. Doublecortin Expression in the Adult Rat Telencephalon: Doublecortin Expression in the Adult Rat. Eur. J. Neurosci. 2001, 14, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Takami, S.; Graziadei, P.P.C.; Ichikawa, M. The Differential Staining Patterns of Two Lectins in the Accessory Olfactory Bulb of the Rat. Brain Res. 1992, 598, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Nii, Y.; Ogawa, K. Subdivisions of the Accessory Olfactory Bulb, as Demonstrated by Lectin-Histochemistry in the Golden Hamster. Neurosci. Lett. 1993, 158, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.S.; Halpern, M.; Ee, P.-L. Lectin Histochemical Identification of Carbohydrate Moieties in Opossum Chemosensory Systems during Development, with Special Emphasis on VVA-Identified Subdivisions in the Accessory Olfactory Bulb. J. Morphol. 1995, 224, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Hovis, K.R.; Ramnath, R.; Dahlen, J.E.; Romanova, A.L.; LaRocca, G.; Bier, M.E.; Urban, N.N. Activity Regulates Functional Connectivity from the Vomeronasal Organ to the Accessory Olfactory Bulb. J. Neurosci. 2012, 32, 7907–7916. [Google Scholar] [CrossRef]
- Raisman, G. An Experimental Study of the Projection of the Amygdala to the Accessory Olfactory Bulb and Its Relationship to the Concept of a Dual Olfactory System. Exp. Brain Res. 1972, 14. [Google Scholar] [CrossRef]
- Scalia, F.; Winans, S.S. The Differential Projections of the Olfactory Bulb and Accessory Olfactory Bulb in Mammals. J. Comp. Neurol. 1975, 161, 31–55. [Google Scholar] [CrossRef]
- Winans, S.S.; Scalia, F. Amygdaloid Nucleus: New Afferent Input from the Vomeronasal Organ. Science 1970, 170, 330–332. [Google Scholar] [CrossRef]
- Shipley, M.T.; Adamek, G.D. The Connections of the Mouse Olfactory Bulb: A Study Using Orthograde and Retrograde Transport of Wheat Germ Agglutinin Conjugated to Horseradish Peroxidase. Brain Res. Bull. 1984, 12, 669–688. [Google Scholar] [CrossRef]
- Dulac, C.; Wagner, S. Genetic Analysis of Brain Circuits Underlying Pheromone Signaling. Annu. Rev. Genet. 2006, 40, 449–467. [Google Scholar] [CrossRef]
- Price, J.L. Beyond the Primary Olfactory Cortex: Olfactory-Related Areas in the Neocortex, Thalamus and Hypothalamus. Chem. Senses 1985, 10, 239–258. [Google Scholar] [CrossRef]
- Courtiol, E.; Wilson, D.A. The Olfactory Thalamus: Unanswered Questions about the Role of the Mediodorsal Thalamic Nucleus in Olfaction. Front. Neural Circuits 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Cádiz-Moretti, B.; Otero-García, M.; Martínez-García, F.; Lanuza, E. Afferent Projections to the Different Medial Amygdala Subdivisions: A Retrograde Tracing Study in the Mouse. Brain Struct. Funct. 2016, 221, 1033–1065. [Google Scholar] [CrossRef]
- Devor, M. Fiber Trajectories of Olfactory Bulb Efferents in the Hamster. J. Comp. Neurol. 1976, 166, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Marcos, A. On the Organization of Olfactory and Vomeronasal Cortices. Prog. Neurobiol. 2009, 87, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Castellanos, N.; Pardo-Bellver, C.; Martínez-García, F.; Lanuza, E. The Vomeronasal Cortex - Afferent and Efferent Projections of the Posteromedial Cortical Nucleus of the Amygdala in Mice. Eur. J. Neurosci. 2014, 39, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Stowers, L.; Liberles, S.D. State-Dependent Responses to Sex Pheromones in Mouse. Curr. Opin. Neurobiol. 2016, 38, 74–79. [Google Scholar] [CrossRef]
- Pardo-Bellver, C.; Cádiz-Moretti, B.; Novejarque, A.; Martínez-García, F.; Lanuza, E. Differential Efferent Projections of the Anterior, Posteroventral, and Posterodorsal Subdivisions of the Medial Amygdala in Mice. Front. Neuroanat. 2012, 6. [Google Scholar] [CrossRef]
- Lo, L.; Anderson, D.J. A Cre-Dependent, Anterograde Transsynaptic Viral Tracer for Mapping Output Pathways of Genetically Marked Neurons. Neuron 2011, 72, 938–950. [Google Scholar] [CrossRef]
- Trotier, D. Vomeronasal Organ and Human Pheromones. Eur. Ann. Otorhinolaryngol. Head. Neck Dis. 2011, 128, 184–190. [Google Scholar] [CrossRef]
- Cadiz, B.; Martinez-Garcia, F.; Lanuza, E. Neural Substrate to Associate Odorants and Pheromones: Convergence of Projections from the Main and Accessory Olfactory Bulbs in Mice. In Chemical Signals in Vertebrates 12; 2013; pp. 3–16. ISBN 978-1-4614-5926-2.
- Mohrhardt, J.; Nagel, M.; Fleck, D.; Ben-Shaul, Y.; Spehr, M. Signal Detection and Coding in the Accessory Olfactory System. Chem. Senses 2018, 43, 667–695. [Google Scholar] [CrossRef] [PubMed]
- Záborszky, L.; Carlsen, J.; Brashear, H.R.; Heimer, L. Cholinergic and GABAergic Afferents to the Olfactory Bulb in the Rat with Special Emphasis on the Projection Neurons in the Nucleus of the Horizontal Limb of the Diagonal Band: CHOLINERGIC AND GABAERGIC PROJECTIONS. J. Comp. Neurol. 1986, 243, 488–509. [Google Scholar] [CrossRef] [PubMed]
- De Saint Jan, D. Target-Specific Control of Olfactory Bulb Periglomerular Cells by GABAergic and Cholinergic Basal Forebrain Inputs. eLife 2022, 11, e71965. [Google Scholar] [CrossRef] [PubMed]
- Liberia, T.; Blasco-Ibáñez, J.M.; Nácher, J.; Varea, E.; Lanciego, J.L.; Crespo, C. Synaptic Connectivity of the Cholinergic Axons in the Olfactory Bulb of the Cynomolgus Monkey. Front. Neuroanat. 2015, 9. [Google Scholar] [CrossRef]
- Böhm, E.; Brunert, D.; Rothermel, M. Input Dependent Modulation of Olfactory Bulb Activity by HDB GABAergic Projections. Sci. Rep. 2020, 10, 10696. [Google Scholar] [CrossRef]
- Gomez, D.M.; Newman, S.W. Differential Projections of the Anterior and Posterior Regions of the Medial Amygdaloid Nucleus in the Syrian Hamster. J. Comp. Neurol. 1992, 317, 195–218. [Google Scholar] [CrossRef]
- Fan, S.; Luo, M. The Organization of Feedback Projections in a Pathway Important for Processing Pheromonal Signals. Neuroscience 2009, 161, 489–500. [Google Scholar] [CrossRef]
- Inbar, T.; Davis, R.; Bergan, J.F. A Sex-specific Feedback Projection from Aromatase-expressing Neurons in the Medial Amygdala to the Accessory Olfactory Bulb. J. Comp. Neurol. 2022, 530, 648–655. [Google Scholar] [CrossRef]
- Pardo-Bellver, C.; Vila-Martin, M.E.; Martínez-Bellver, S.; Villafranca-Faus, M.; Teruel-Sanchis, A.; Savarelli-Balsamo, C.A.; Drabik, S.M.; Martínez-Ricós, J.; Cervera-Ferri, A.; Martínez-García, F.; et al. Neural Activity Patterns in the Chemosensory Network Encoding Vomeronasal and Olfactory Information in Mice. Front. Neuroanat. 2022, 16, 988015. [Google Scholar] [CrossRef]
- Keverne, E.B.; Brennan, P.A. Olfactory Recognition Memory. J. Physiol. -Paris. 1996, 90, 399–401. [Google Scholar] [CrossRef]
- Brennan, P.A.; Kendrick, K.M.; Keverne, E.B. Neurotransmitter Release in the Accessory Olfactory Bulb during and after the Formation of an Olfactory Memory in Mice. Neuroscience 1995, 69, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.A. The Nose Knows Who’s Who: Chemosensory Individuality and Mate Recognition in Mice. Horm. Behav. 2004, 46, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Kaba, H. Muscarinic Receptor Type 1 (M1) Stimulation, Probably through KCNQ/Kv7 Channel Closure, Increases Spontaneous GABA Release at the Dendrodendritic Synapse in the Mouse Accessory Olfactory Bulb. Brain Res. 2010, 1339, 26–40. [Google Scholar] [CrossRef]
- Wilson, R.I.; Mainen, Z.F. Early Events in Olfactory Processing. Annu. Rev. Neurosci. 2006, 29, 163–201. [Google Scholar] [CrossRef] [PubMed]
- Grus, W.E.; Zhang, J. Origin of the Genetic Components of the Vomeronasal System in the Common Ancestor of All Extant Vertebrates. Mol. Biol. Evol. 2009, 26, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Hurst, J.L.; Barnard, C.J. Comparing Behaviour in Wild and Laboratory Strains of the House Mouse: Levels of Comparison and Functional Inference. Behav. Process. 1994, 32, 79–86. [Google Scholar] [CrossRef]
- Catania, K.C. Underwater “sniffing” by Semi-Aquatic Mammals. Nature 2006, 444, 1024–1025. [Google Scholar] [CrossRef]
- Johnson, E.W.; Rasmussen, L. Morphological Characteristics of the Vomeronasal Organ of the Newborn Asian Elephant (Elephas Maximus). Anat. Rec. 2002, 267, 252–259. [Google Scholar] [CrossRef]
- Garrosa, M.; Gayoso, M.J.; Esteban, F.J. Prenatal Development of the Mammalian Vomeronasal Organ. Microsc. Res. Tech. 1998, 41, 456–470. [Google Scholar] [CrossRef]
- Sugai, T.; Sugitani, M.; Onoda, N. Subdivisions of the Guinea-Pig Accessory Olfactory Bulb Revealed by the Combined Method with Immunohistochemistry, Electrophysiological, and Optical Recordings. Neuroscience 1997, 79, 871–885. [Google Scholar] [CrossRef]
- Fieni, F. Apical and Basal Neurones Isolated from the Mouse Vomeronasal Organ Differ for Voltage-Dependent Currents. J. Physiol. 2003, 552, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. The Vomeronasal System of the Newborn Capybara: A Morphological and Immunohistochemical Study. Sci. Rep. 2020, 10, 13304. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Lombardero, M.; Cifuentes, J.M.; Quinteiro, P.S.; Aleman, N. Morphogenesis and Growth of the Soft Tissue and Cartilage of the Vomeronasal Organ in Pigs. J. Anat. 2003, 202, 503–514. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez Quinteiro, P.; Lombardero, M.; Aleman, N.; Fernández de Trocóniz, P. The Prenatal Maturity of the Accessory Olfactory Bulb in Pigs. Chem. Senses 2004, 29, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, W.; Jeong, C.; Kim, H.; Taniguchi, K.; Shin, T. Developmental Changes Affecting Lectin Binding in the Vomeronasal Organ of Domestic Pigs, Sus Scrofa. Acta Histochem. 2012, 114, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Suárez, R.; Santibáñez, R.; Parra, D.; Coppi, A.A.; Abrahão, L.M.B.; Sasahara, T.H.C.; Mpodozis, J. Shared and Differential Traits in the Accessory Olfactory Bulb of Caviomorph Rodents with Particular Reference to the Semiaquatic Capybara: The AOB of Capybaras and Other Caviomorphs. J. Anat. 2011, 218, 558–565. [Google Scholar] [CrossRef]
- Fernández-Aburto, P.; Delgado, S.E.; Sobrero, R.; Mpodozis, J. Can Social Behaviour Drive Accessory Olfactory Bulb Asymmetries? Sister Species of Caviomorph Rodents as a Case in Point. J. Anat. 2020, 236, 612–621. [Google Scholar] [CrossRef]
- Malz, C.R.; Knabe, W.; Kuhn, H.-J. Calretinin Immunoreactivity in the Prenatally Developing Olfactory Systems of the Tree Shrew Tupaia Belangeri. Anat. Embryol. 2002, 205, 83–97. [Google Scholar] [CrossRef]
- Brunjes, P.C.; Jazaeri, A.; Sutherland, M.J. Olfactory Bulb Organization and Development in Monodelphis Domestica (Grey Short-Tailed Opossum). J. Comp. Neurol. 1992, 320, 544–554. [Google Scholar] [CrossRef]
- Salazar, I.; Sanchez-Quinteiro, P.; Lombardero, M.; Cifuentes, J.M. Histochemical Identification of Carbohydrate Moieties in the Accessory Olfactory Bulb of the Mouse Using a Panel of Lectins. Chem. Senses 2001, 26, 645–652. [Google Scholar] [CrossRef]
- Bojsen-Møller, F. Topography of the Nasal Glands in Rats and Some Other Mammals. Anat. Rec. 1964, 150, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Pastor, L.M.; Frutos, M.J.; Graa, L.; Ramos, D.; Gallego-Huidobro, J.; Calvo, A. Histochemical Study of Glycoconjugates in the Nasal Mucosa of the Rat and Guinea Pig. Histochem. J. 1992, 24, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Roslinski, D.L.; Bhatnagar, K.P.; Burrows, A.M.; Smith, T.D. Comparative Morphology and Histochemistry of Glands Associated with the Vomeronasal Organ in Humans, Mouse Lemurs, and Voles. Anat. Rec. 2000, 260, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Origin of Luteinizing Hormone-Releasing Hormone Neurons. Nature 1989, 338, 161–164. [Google Scholar] [CrossRef]
- Schwanzel-Fukuda, M.; Pfaff, D.W. The Migration of Luteinizing Hormone-Releasing Hormone (LHRH) Neurons from the Medial Alfactory Placode into the Medial Basal Forebrain. Experientia 1990, 46, 956–962. [Google Scholar] [CrossRef]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Migration of LHRH-Immunoreactive Neurons from the Olfactory Placode Rationalizes Olfacto-Hormonal Relationships. J. Steroid Biochem. Mol. Biol. 1991, 39, 565–572. [Google Scholar] [CrossRef]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Luteinizing Hormone-Releasing Hormone (LHRH) and Neural Cell Adhesion Molecule (NCAM)-Immunoreactivity in Development of the Forebrain and Reproductive System. Ann Endocrinol (Paris) 1994, 55, 235–241. [Google Scholar]
- Skeen, L.C.; Hall, W.C. Efferent Projections of the Main and the Accessory Olfactory Bulb in the Tree Shrew (Tupaia Glis). J. Comp. Neurol. 1977, 172, 1–35. [Google Scholar] [CrossRef]
- Stephan, H. Der Bulbus Olfatorius Accessorius Bei Insektivoren Und Primaten. Cells Tissues Organs 1965, 62, 215–253. [Google Scholar] [CrossRef]
- Schneider, N.Y.; Fletcher, T.P.; Shaw, G.; Renfree, M.B. The Olfactory System of the Tammar Wallaby Is Developed at Birth and Directs the Neonate to Its Mother’s Pouch Odours. REPRODUCTION 2009, 138, 849–857. [Google Scholar] [CrossRef]
- Wettschureck, N.; Offermanns, S. Mammalian G Proteins and Their Cell Type Specific Functions. Physiol. Rev. 2005, 85, 1159–1204. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Kawai, Y.K.; Watanabe, K.; Muranishi, Y. Artiodactyl Livestock Species Have a Uniform Vomeronasal System with a Vomeronasal Type 1 Receptor (V1R) Pathway. Tissue Cell 2022, 77, 101863. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Does a Third Intermediate Model for the Vomeronasal Processing of Information Exist? Insights from the Macropodid Neuroanatomy. Brain Struct. Funct. 2022, 227, 881–899. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Kamikawa, A.; Sasaki, M.; Kitamura, N. Localization of A1-2 Fucose Glycan in the Mouse Olfactory Pathway. Cells Tissues Organs 2017, 203, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.-A.; Niedermeier, S.; Claassen, L.; Popp, A. Comparative Lectin Histochemistry on the Murine Respiratory Tract and Primary Olfactory Pathway Using a Fully Automated Staining Procedure. Acta Histochem. 2022, 124, 151877. [Google Scholar] [CrossRef]
- Salazar, I.; Sanchez-Quinteiro, P.; Lombardero, M.; Cifuentes, J.M. A Descriptive and Comparative Lectin Histochemical Study of the Vomeronasal System in Pigs and Sheep. J. Anat. 2000, 196, 15–22. [Google Scholar] [CrossRef]
- Mogi, K.; Sakurai, K.; Ichimaru, T.; Ohkura, S.; Mori, Y.; Okamura, H. Structure and Chemical Organization of the Accessory Olfactory Bulb in the Goat. Anat. Rec. 2007, 290, 301–310. [Google Scholar] [CrossRef]
- Sweet, G. Memoirs: Contributions to Our Knowledge of the Anatomy of Notoryctes Typhlops, Stirling. J. Cell Sci. 1906, s2-50, 547–572. [Google Scholar] [CrossRef]
- Broom, R. On the Organ of Jacobson and Some Other Structures in the Nose of Cænolestes. By R. BROOM, D.Sc., P.R.S., C.M.Z.S. Proc. Zool. Soc. Lond. 1926, 96, 419–424. [Google Scholar] [CrossRef]
- Bock, P.; Rohn, K.; Beineke, A.; Baumgärtner, W.; Wewetzer, K. Site-Specific Population Dynamics and Variable Olfactory Marker Protein Expression in the Postnatal Canine Olfactory Epithelium. J. Anat. 2009, 215, 522–535. [Google Scholar] [CrossRef]
- Salazar, I.; Cifuentes, J.M.; Quinteiro, P.S.; Caballero, T.G. Structural, Morphometric, and Immunohistological Study of the Accessory Olfactory Bulb in the Dog. Anat. Rec. 1994, 240, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.V.; Ortiz-Leal, I.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata Suricatta). Animals 2021, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- O’Riain, M.J.; Bennett, N.C.; Brotherton, P.N.M.; McIlrath, G.; Clutton-Brock, T.H. Reproductive Suppression and Inbreeding Avoidance in Wild Populations of Co-Operatively Breeding Meerkats ( Suricata Suricatta ). Behav. Ecol. Sociobiol. 2000, 48, 471–477. [Google Scholar] [CrossRef]
- Leclaire, S.; Nielsen, J.F.; Thavarajah, N.K.; Manser, M.; Clutton-Brock, T.H. Odour-Based Kin Discrimination in the Cooperatively Breeding Meerkat. Biol. Lett. 2013, 9, 20121054. [Google Scholar] [CrossRef] [PubMed]
- Leclaire, S.; Jacob, S.; Greene, L.K.; Dubay, G.R.; Drea, C.M. Social Odours Covary with Bacterial Community in the Anal Secretions of Wild Meerkats. Sci. Rep. 2017, 7, 3240. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Rapid Turnover and Species-Specificity of Vomeronasal Pheromone Receptor Genes in Mice and Rats. Gene 2004, 340, 303–312. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez-Quinteiro, P. The Risk of Extrapolation in Neuroanatomy: The Case of the Mammalian Accessory Olfactory Bulb. Front. Neuroanat. 2009, 3. [Google Scholar] [CrossRef]
- Nakajima, T.; Sakaue, M.; Kato, M.; Saito, S.; Ogawa, K.; Taniguchi, K. Immunohistochemical and Enzyme-Histochemical Study on the Accessory Olfactory Bulb of the Dog. Anat. Rec. 1998, 252, 393–402. [Google Scholar] [CrossRef]
- Hughes, L. Meerkats. Essential Wildlife; Character-19 eds., 2020.
- Broom, R. On the Organ of Jacobson in the Hyrax. J. Anat. Physiol. 1898, 32, 709–713. [Google Scholar]
- Yohe, L.R.; Krell, N.T. An Updated Synthesis of and Outstanding Questions in the Olfactory and Vomeronasal Systems in Bats: Genetics Asks Questions Only Anatomy Can Answer. Anat. Rec. 2023, ar.25290. [Google Scholar] [CrossRef]
- Wöhrmann-Repenning, A. Functional Aspects of the Vomeronasal Complex in Mammals. Zool. Jb. Anat. 1991, 121, 71–80. [Google Scholar]
- Shnayder, L.; Schwanzel-Fukuda, M.; Halpern, M. Differential OMP Expression in Opossum Accessory Olfactory Bulb: NeuroReport. 1993, 5, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Sasaki, M.; Kitamura, N. Age-Dependent Decrease in Glomeruli and Receptor Cells Containing A1–2 Fucose Glycan in the Mouse Main Olfactory System but Not in the Vomeronasal System. Cell Tissue Res. 2018, 373, 361–366. [Google Scholar] [CrossRef]
- Tiwari, R.P.; Ahmed, A.; Mishra, G.K. J. Vet. Sci. Tech. 2014, 15–20.
- Rekwot, P.I.; Ogwu, D.; Oyedipe, E.O.; Sekoni, V.O. The Role of Pheromones and Biostimulation in Animal Reproduction. Anim. Reprod. Sci. 2001, 65, 157–170. [Google Scholar] [CrossRef]
- Landaeta-Hernández, A.J.; Ungerfeld, R.; Chenoweth, P.J. Biostimulation and Pheromones in Livestock: A Review. Anim. Reprod. Sci. 2023, 248, 107154. [Google Scholar] [CrossRef]
- Torres, M.V.; Ortiz-Leal, I.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Immunohistological Study of the Unexplored Vomeronasal Organ of an Endangered Mammal, the Dama Gazelle (Nanger Dama ). Microsc. Res. Tech. 2023, 86, 1206–1233. [Google Scholar] [CrossRef]
- Hart, B.L.; Hart, L.A.; Maina, J.N. Alteration in Vomeronasal System Anatomy in Alcelaphine Antelopes: Correlation with Alteration in Chemosensory Investigation. Physiol. Behav. 1988, 42, 155–162. [Google Scholar] [CrossRef]
- Meredith, M.; Marques, D.M.; O’Connell, R.J.; Stern, F.L. Vomeronasal Pump: Significance for Male Hamster Sexual Behavior. Science 1980, 207, 1224–1226. [Google Scholar] [CrossRef]
- Krishna, N.S.R.; Getchell, M.L.; Margolis, F.L.; Getchell, T.V. Differential Expression of Vomeromodulin and Odorant-Binding Protein, Putative Pheromone and Odorant Transporters, in the Developing Rat Nasal Chemosensory Mucosae. J. Neurosci. Res. 1995, 40, 54–71. [Google Scholar] [CrossRef]
- Takigami, S.; Mori, Y.; Ichikawa, M. Projection Pattern of Vomeronasal Neurons to the Accessory Olfactory Bulb in Goats. Chem. Senses 2000, 25, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Akaogi, S.; Nakamuta, S.; Tsujimoto, T.; Nakamuta, N. Characteristics of Olfactory Organs in Sika Deer (Cervus Nippon). Jpn. J. Zoo. Wildl. Med. 2019, 24, 115–122. [Google Scholar] [CrossRef]
Figure 1.
Dissection of the turbinate complex in the dog. The difference between the red respiratory mucosa (RM) and the brown olfactory mucosa (OM) is observed. 1. Dorsal turbinate, 2. Ventral turbinate; 3. Endoturbinate; 4. Ectoturbinates; 5. Olfactory bulb; 6. Telencephalon frontal lobe.
Figure 1.
Dissection of the turbinate complex in the dog. The difference between the red respiratory mucosa (RM) and the brown olfactory mucosa (OM) is observed. 1. Dorsal turbinate, 2. Ventral turbinate; 3. Endoturbinate; 4. Ectoturbinates; 5. Olfactory bulb; 6. Telencephalon frontal lobe.
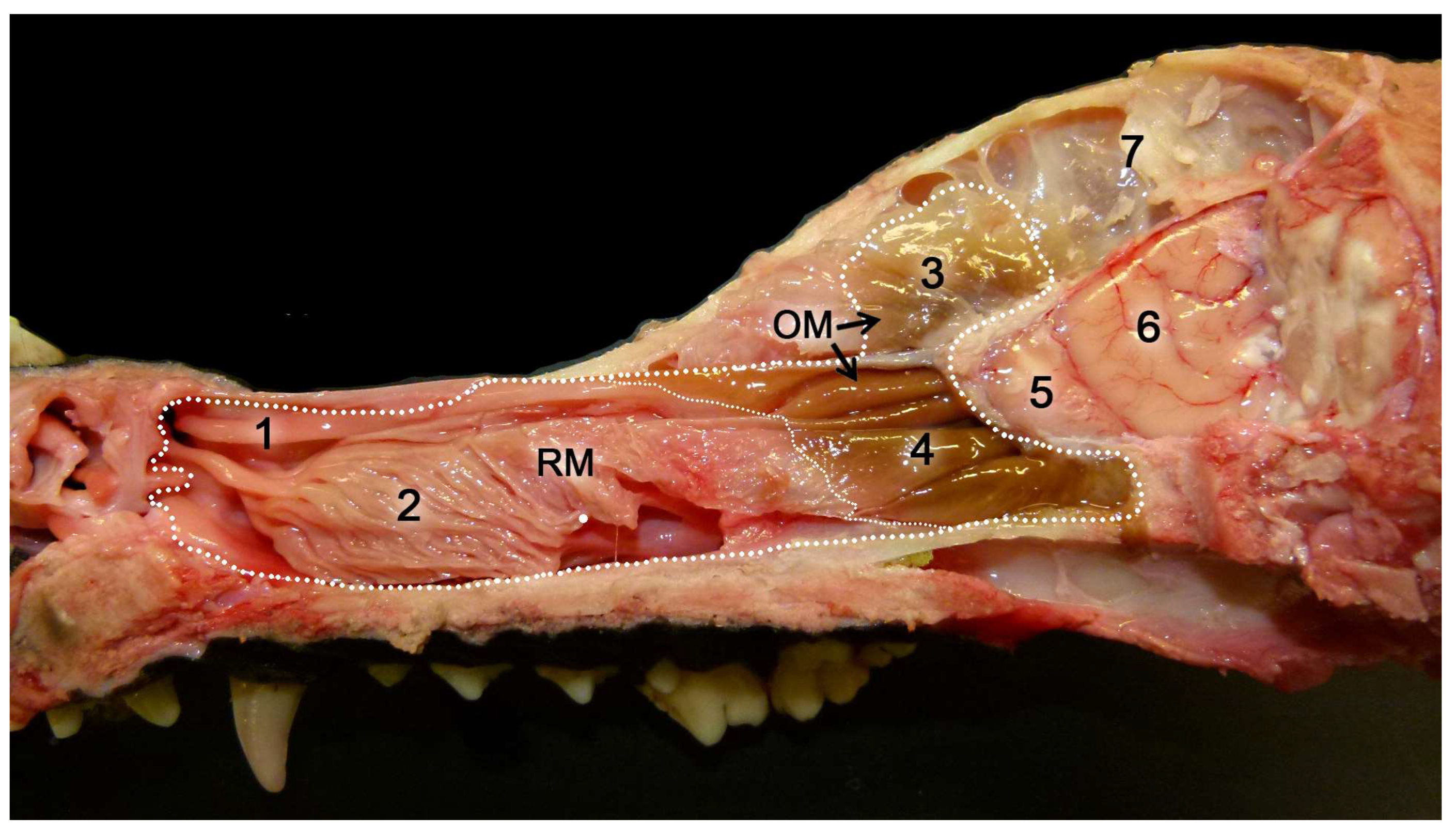
Figure 2.
Lateral view of the nasal septum of a dog showing the projection area of the vomeronasal organ (open arrowheads) and the vomeronasal nerves (arrows). FS, Frontal sinus; 1. Olfactory bulb, 2. Olfactory peduncle; 3. Telencephalon.
Figure 2.
Lateral view of the nasal septum of a dog showing the projection area of the vomeronasal organ (open arrowheads) and the vomeronasal nerves (arrows). FS, Frontal sinus; 1. Olfactory bulb, 2. Olfactory peduncle; 3. Telencephalon.
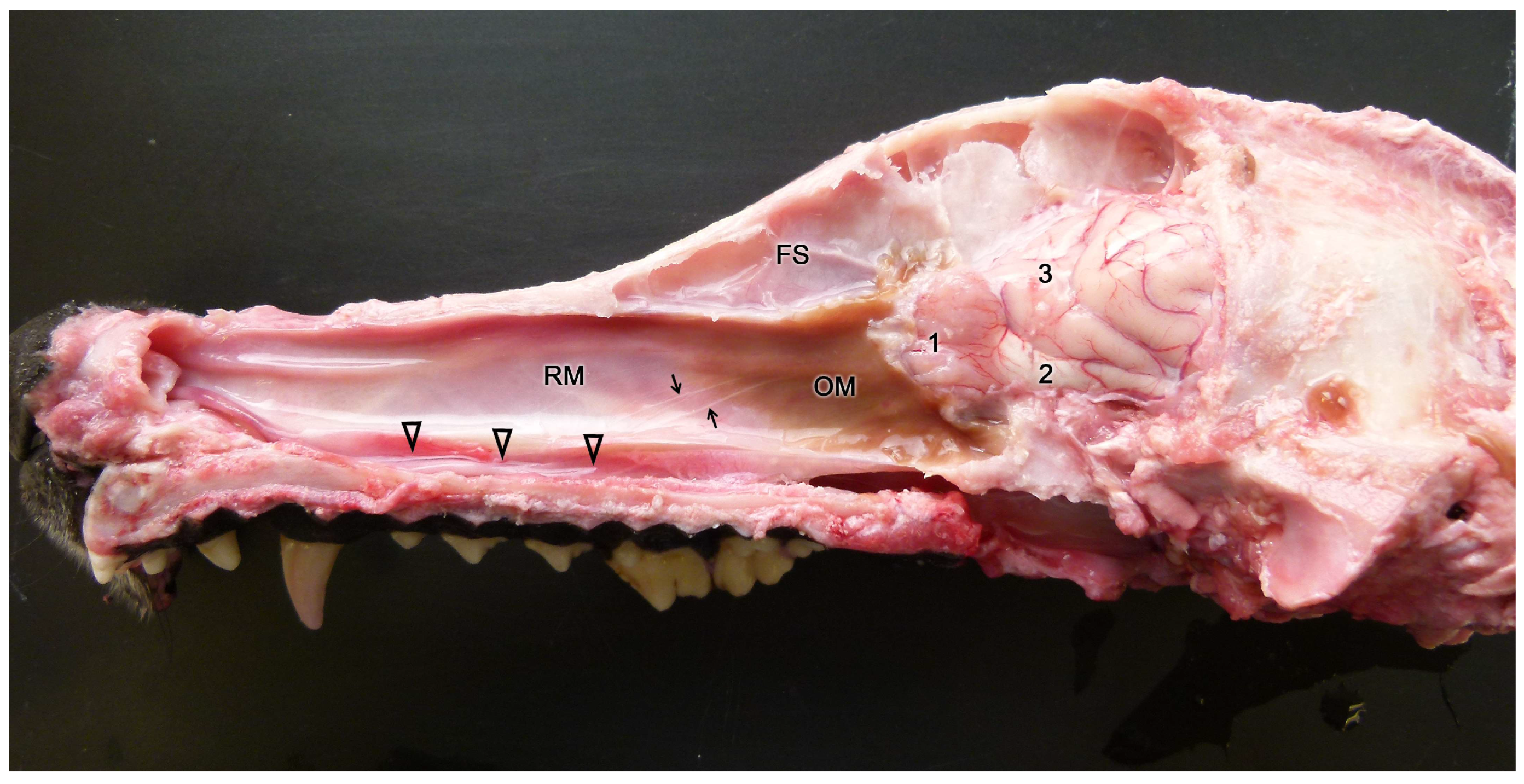
Figure 3.
Grüneberg ganglion of the mouse. Transverse section of the dorsal part of the nasal vestibule (NV) immunostained with anti-OMP showing immunopositive ganglion cells (arrowheads). NVC, Cartilage of the nasal vestibule.
Figure 3.
Grüneberg ganglion of the mouse. Transverse section of the dorsal part of the nasal vestibule (NV) immunostained with anti-OMP showing immunopositive ganglion cells (arrowheads). NVC, Cartilage of the nasal vestibule.
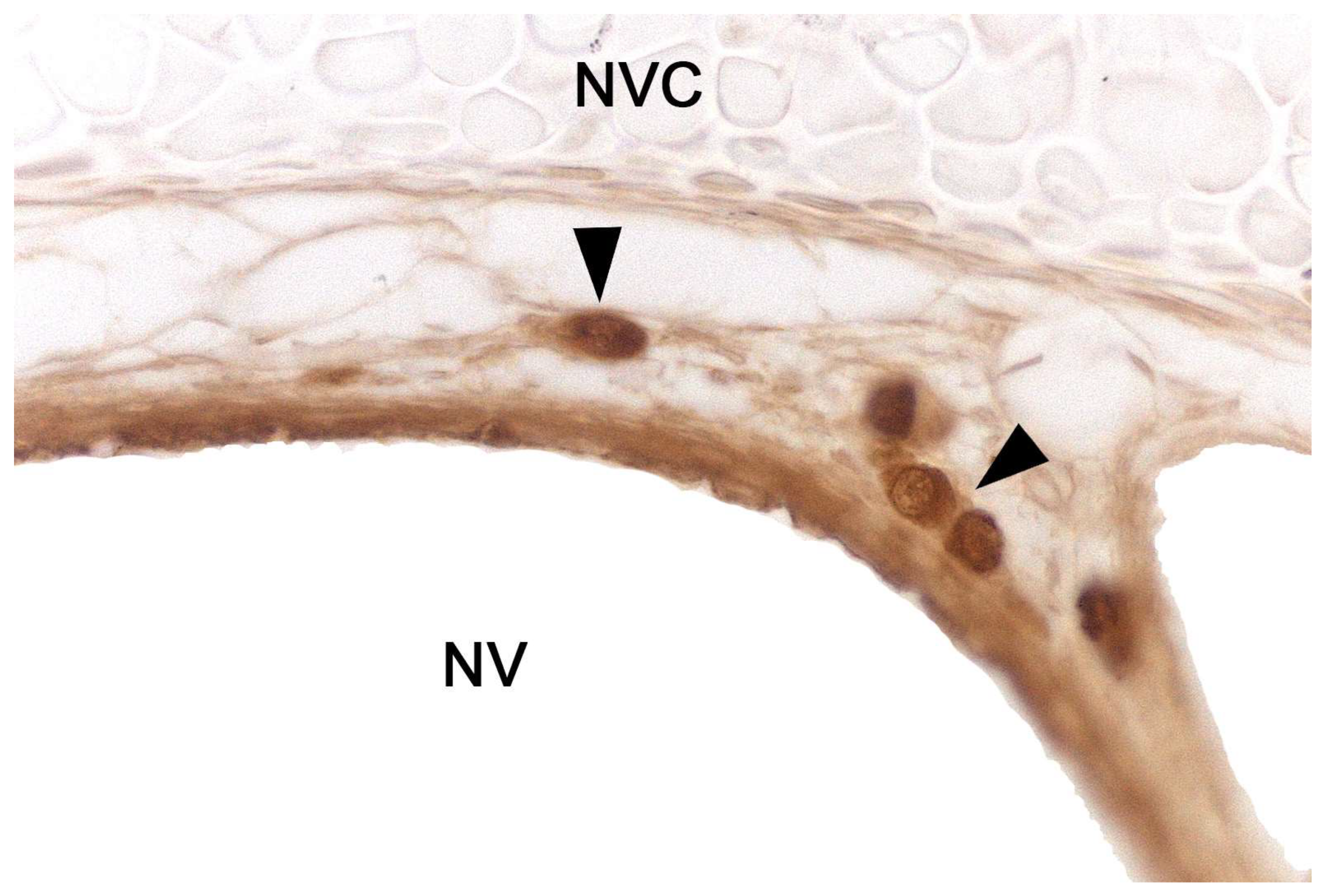
Figure 4.
Septal organ of the mouse. Decalcified transverse section of the nasal cavity immunostained with anti-OMP. The immunopositive olfactory epithelium (OE) lines the roof of the nasal cavity, the nasal septum, the dorsal part of the ventral turbinate (VN) and the branches of the olfactory nerve (nI). On the basal part of the nasal septum a patch of OMP-positive epithelium corresponding to the septal organ (SO) can be observed). VN, vomeronasal nerves.
Figure 4.
Septal organ of the mouse. Decalcified transverse section of the nasal cavity immunostained with anti-OMP. The immunopositive olfactory epithelium (OE) lines the roof of the nasal cavity, the nasal septum, the dorsal part of the ventral turbinate (VN) and the branches of the olfactory nerve (nI). On the basal part of the nasal septum a patch of OMP-positive epithelium corresponding to the septal organ (SO) can be observed). VN, vomeronasal nerves.
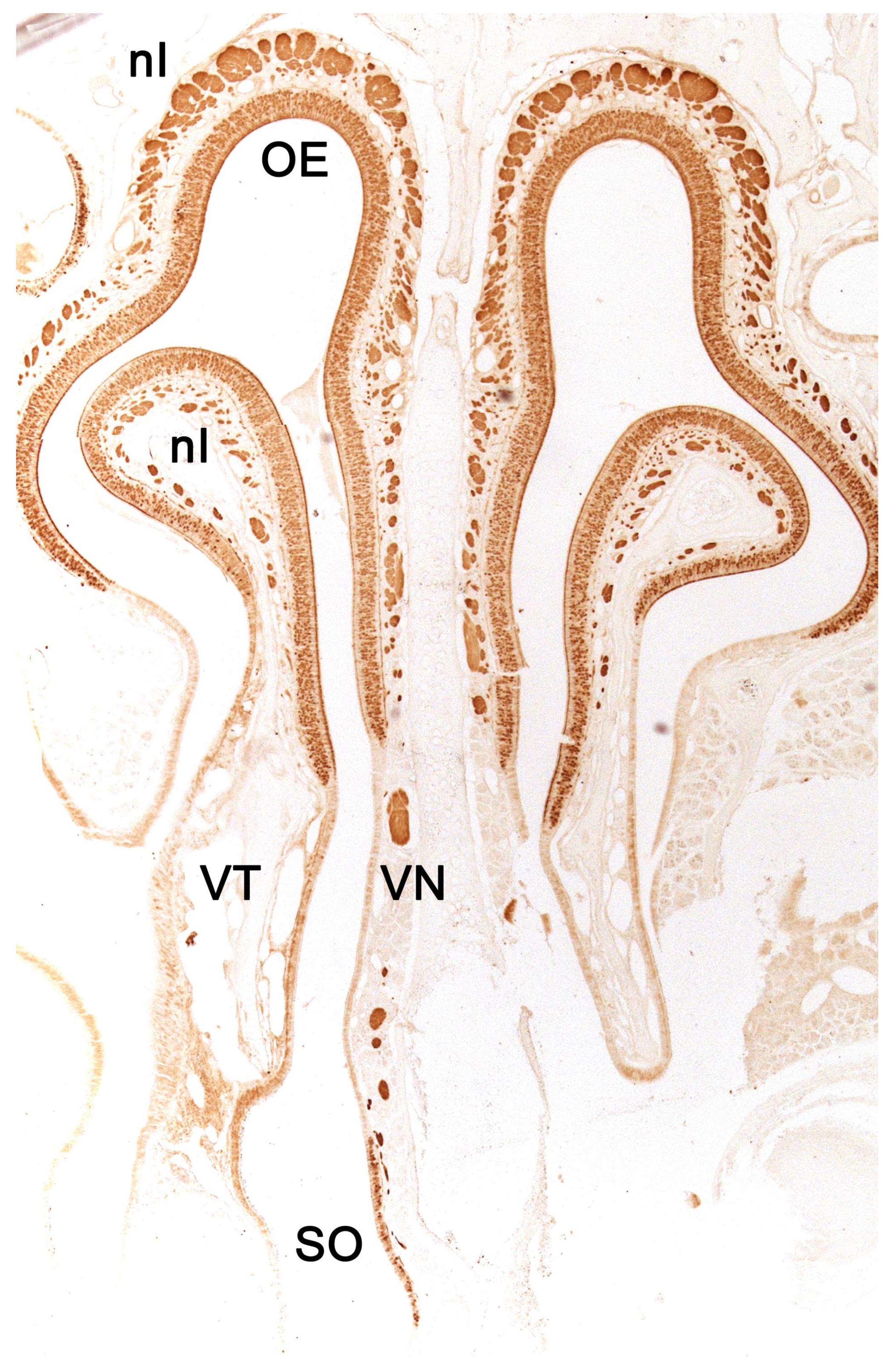
Figure 5.
Septal organ of the mouse. View at higher magnification of the septal organ (SO) area showed in the Figure 4. Decalcified transverse section of the nasal cavity immunostained with anti-OMP. Immunopositive neuroreceptor cells and the branches of the septal olfactory axons (SON) can be observed. VN, vomeronasal nerves.
Figure 5.
Septal organ of the mouse. View at higher magnification of the septal organ (SO) area showed in the Figure 4. Decalcified transverse section of the nasal cavity immunostained with anti-OMP. Immunopositive neuroreceptor cells and the branches of the septal olfactory axons (SON) can be observed. VN, vomeronasal nerves.
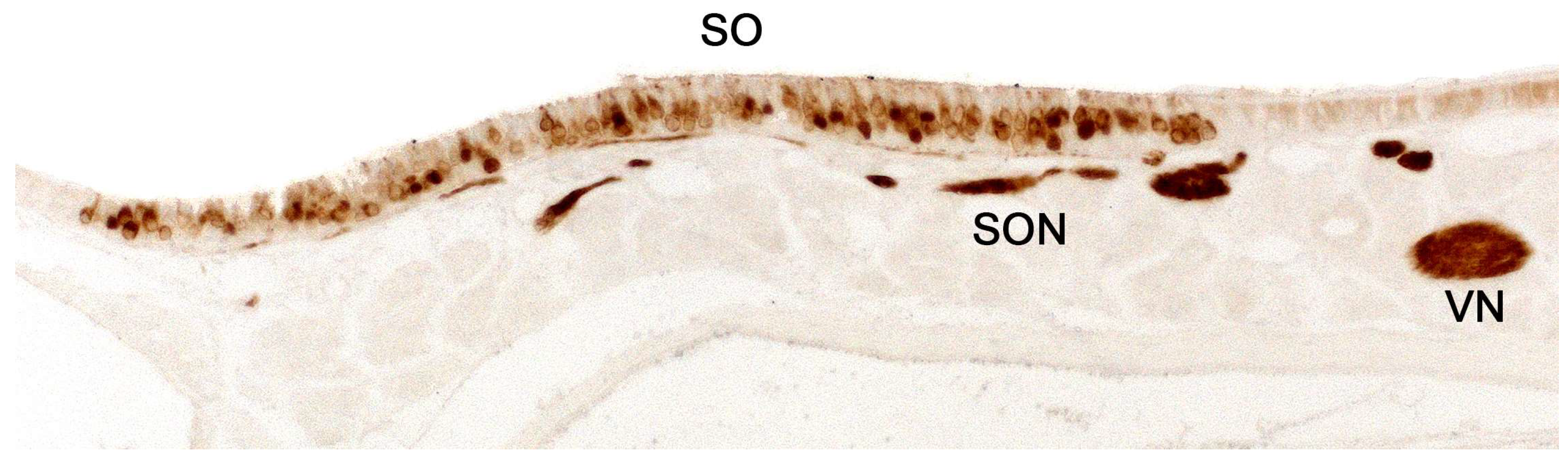
Figure 6.
Vomeronasal organ of the horse. Autofluorescence transversal section of the VNO in which all components of the organ are identified. Aa, Artery; RE, Respiratory epithelium; RM, Respiratory mucosa; SE, Sensory epithelium; VC, Vomeronasal cartilage; VG, Vomeronasal glands; VN, Vomeronasal nerves; Vv, Veins. Nuclear contrast: TOPRO-3 iodide.
Figure 6.
Vomeronasal organ of the horse. Autofluorescence transversal section of the VNO in which all components of the organ are identified. Aa, Artery; RE, Respiratory epithelium; RM, Respiratory mucosa; SE, Sensory epithelium; VC, Vomeronasal cartilage; VG, Vomeronasal glands; VN, Vomeronasal nerves; Vv, Veins. Nuclear contrast: TOPRO-3 iodide.
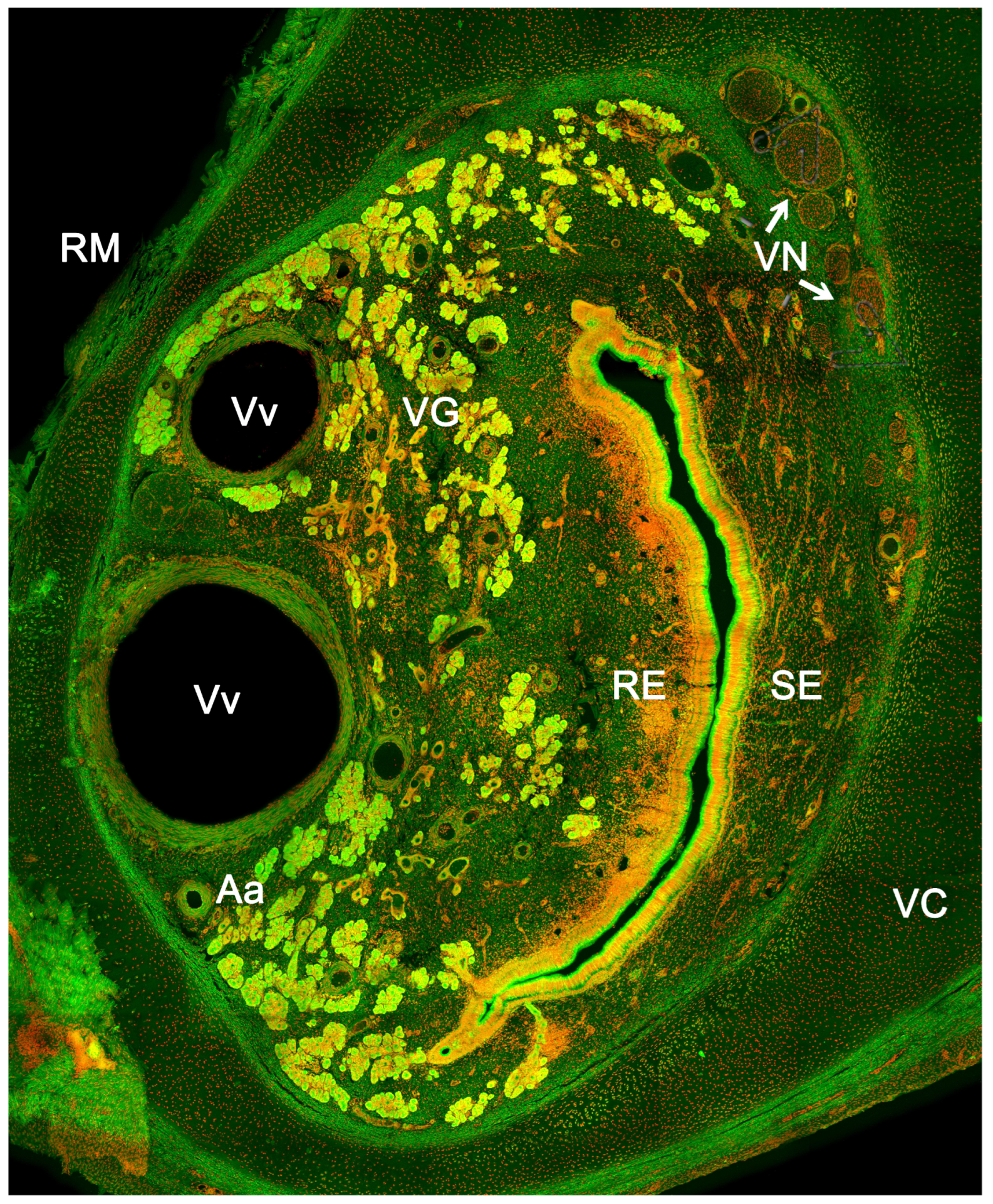
Figure 7.
Sensory epithelium of the dama gazelle vomeronasal organ stained with hematoxylin-eosin. 1. Basal cells; 2. Neuroreceptor cells; 3. Sustentacular cells; 4. Mucomicrovillar complex; 5. Vomeronasal nerve axons; 6. Veins; 7. Collagen fibers.
Figure 7.
Sensory epithelium of the dama gazelle vomeronasal organ stained with hematoxylin-eosin. 1. Basal cells; 2. Neuroreceptor cells; 3. Sustentacular cells; 4. Mucomicrovillar complex; 5. Vomeronasal nerve axons; 6. Veins; 7. Collagen fibers.
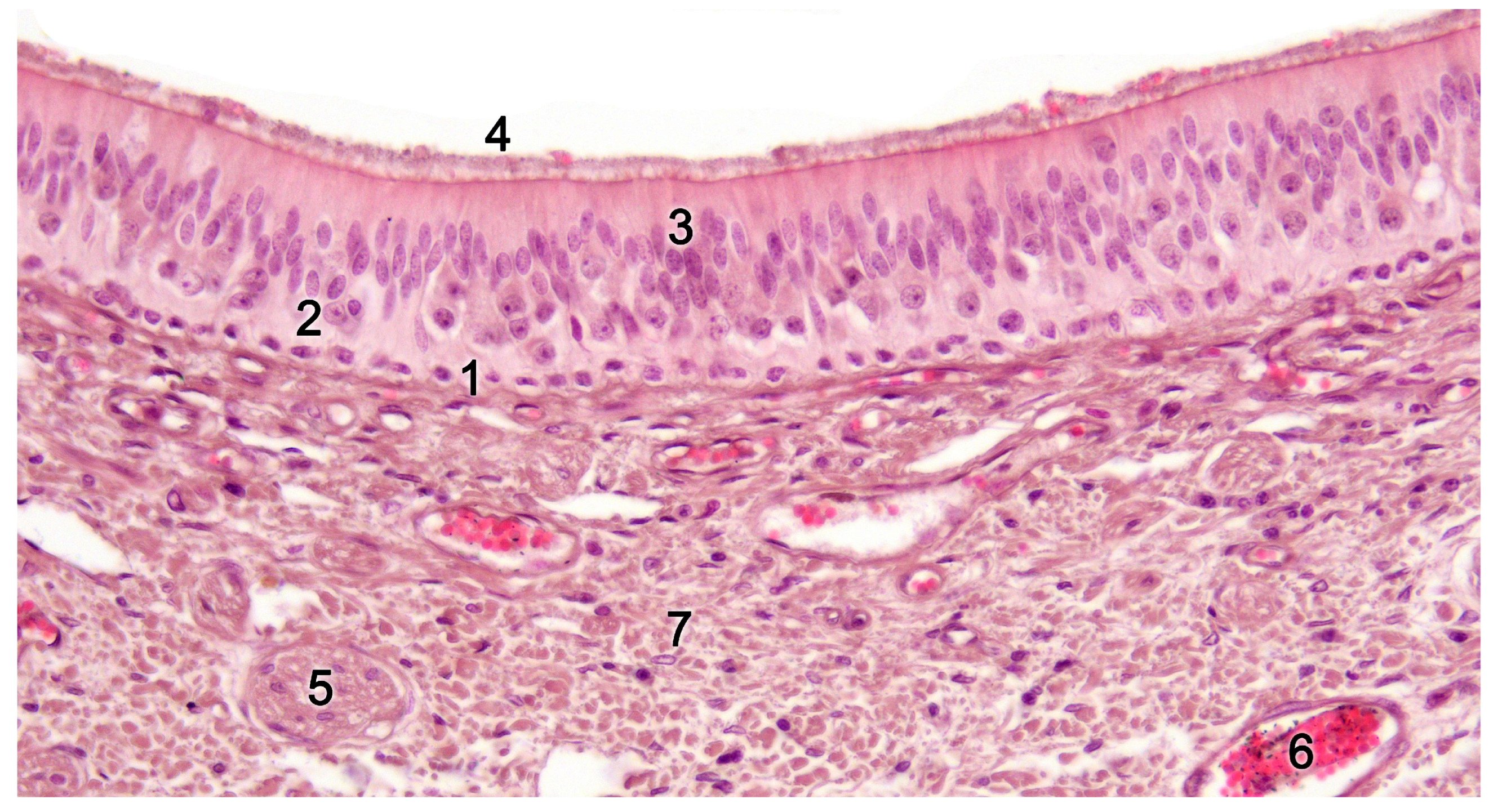
Figure 8.
Vomeronasal organ of the horse. Autofluorescence confocal microscopy of the sensory epithelium (SE) allows for the clear differentiation of the zones corresponding to the main strata: Basal (1); Neuroreceptor (2); Sustentacular (3); Mucomicrovillar (4). RE, Respiratory epithelium; Red nuclear contrast: TOPRO-3 iodide.
Figure 8.
Vomeronasal organ of the horse. Autofluorescence confocal microscopy of the sensory epithelium (SE) allows for the clear differentiation of the zones corresponding to the main strata: Basal (1); Neuroreceptor (2); Sustentacular (3); Mucomicrovillar (4). RE, Respiratory epithelium; Red nuclear contrast: TOPRO-3 iodide.
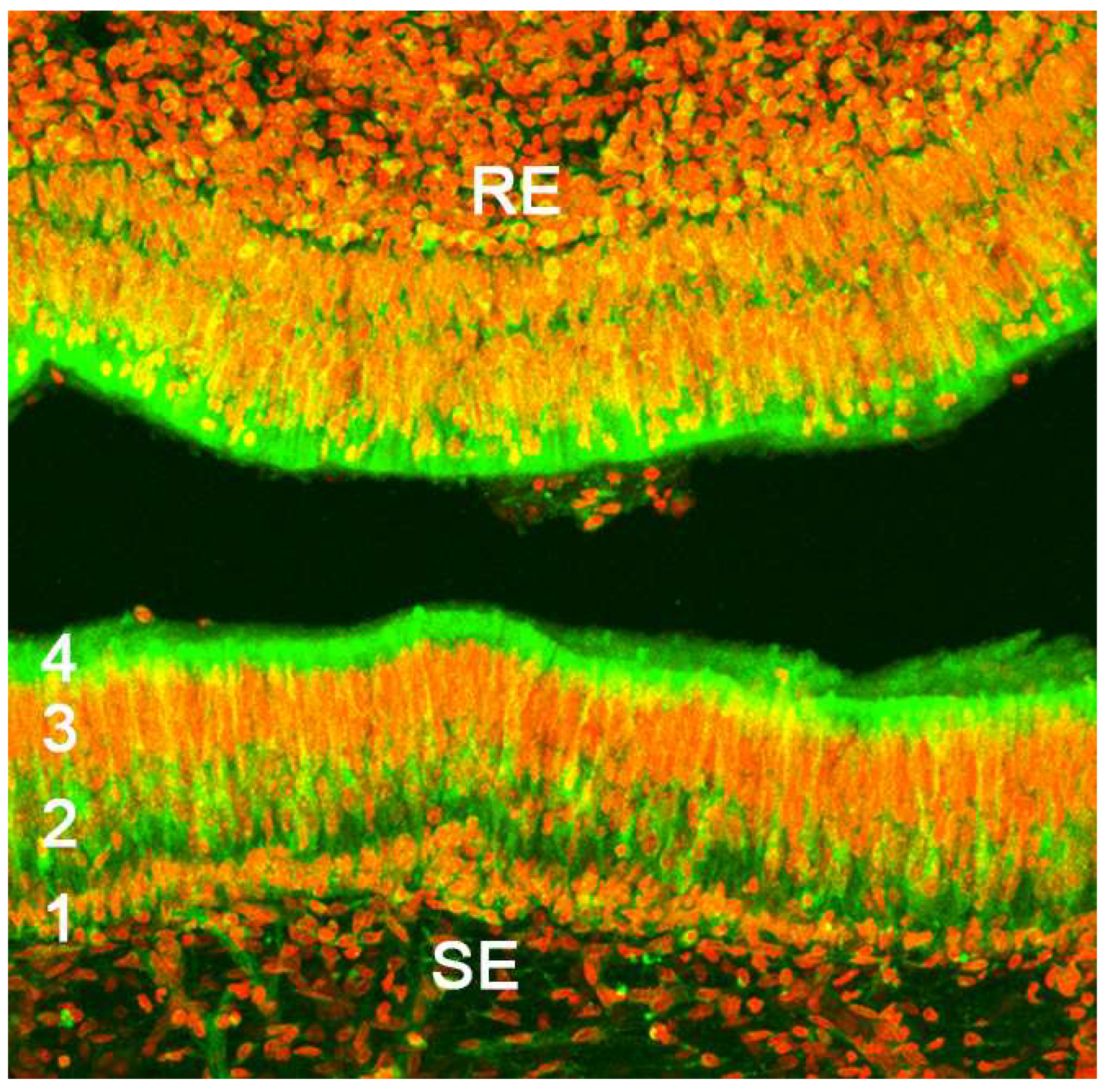
Figure 9.
Vomeronasal gland of the cow stained with PAS-Alcianblue-hematoxylin staining. Both the glandular acini and the duct are stained.
Figure 9.
Vomeronasal gland of the cow stained with PAS-Alcianblue-hematoxylin staining. Both the glandular acini and the duct are stained.
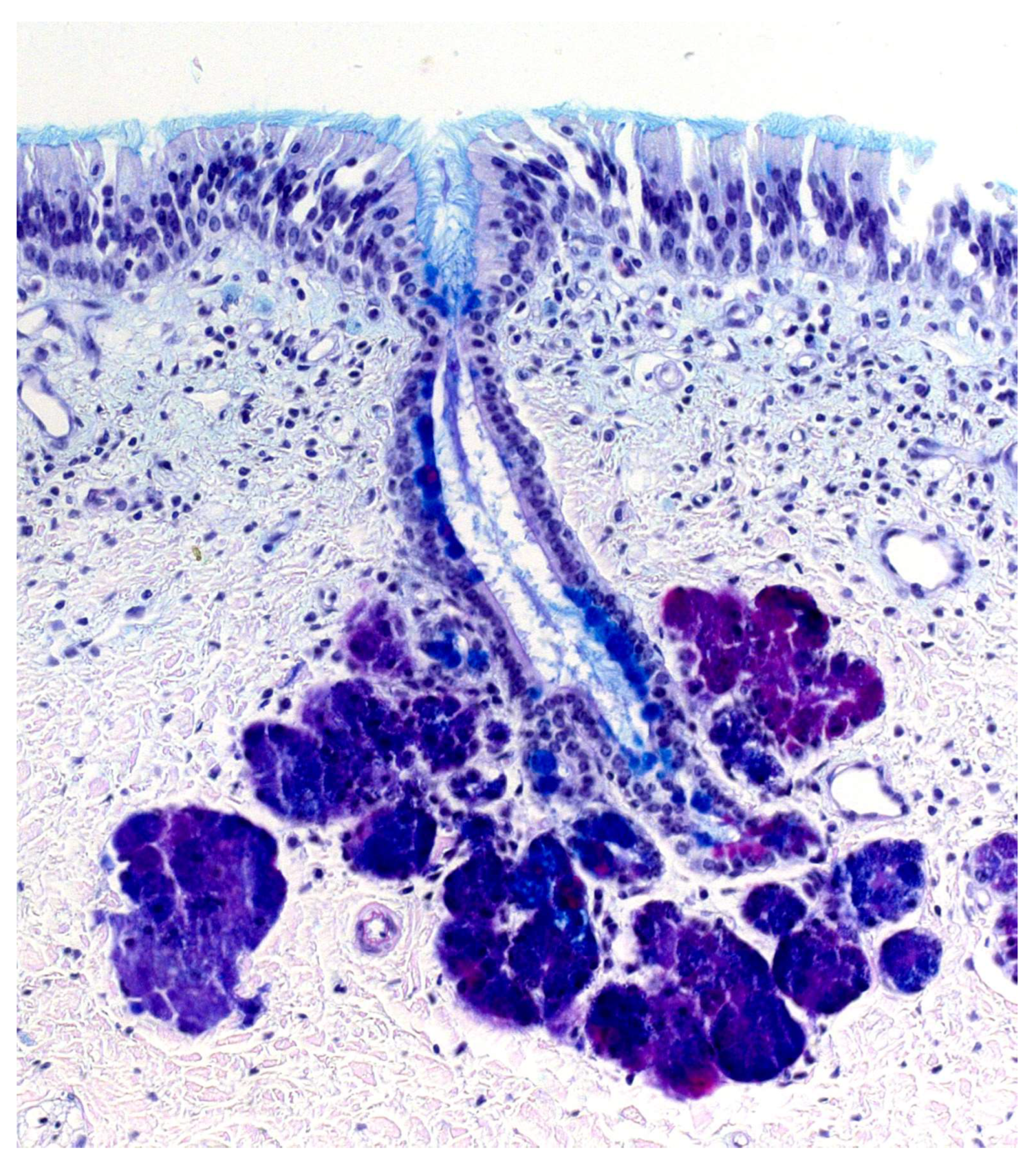
Figure 10.
Sagittal section of the nose of a fetal mouse. The vomeronasal organ is located in the base of the nasal septum. A functional opening to the nasal cavity (NC) can be observed The NC is lined by olfactory epithelium (OE). Hematoxylin-eosin staining.
Figure 10.
Sagittal section of the nose of a fetal mouse. The vomeronasal organ is located in the base of the nasal septum. A functional opening to the nasal cavity (NC) can be observed The NC is lined by olfactory epithelium (OE). Hematoxylin-eosin staining.
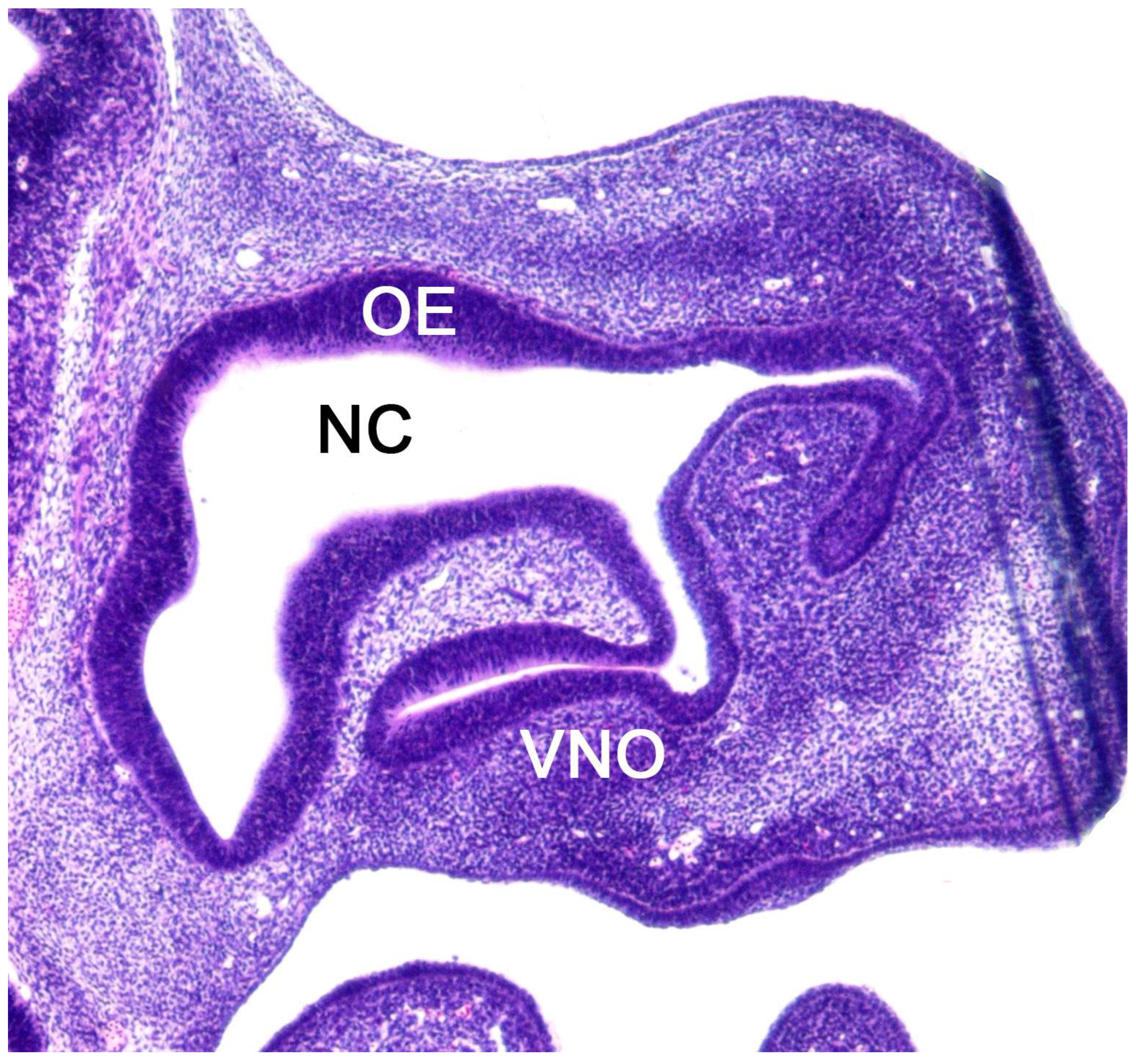
Figure 11.
The vomeronasal organ of the rabbit. Hematoxylin-eosin staining. Transverse section of the head showing the nasal septum (1) with the VNOs located over the palatine processes of the incisive bone (2). The J-shaped cartilaginous envelope of the VNO (3) is covered by a thin bony layer (4). The ventral recess of the nasal cavity (5) is shaped by the cartilage of the incisive duct (6). 7. Ventral turbinate; 8. Palate.
Figure 11.
The vomeronasal organ of the rabbit. Hematoxylin-eosin staining. Transverse section of the head showing the nasal septum (1) with the VNOs located over the palatine processes of the incisive bone (2). The J-shaped cartilaginous envelope of the VNO (3) is covered by a thin bony layer (4). The ventral recess of the nasal cavity (5) is shaped by the cartilage of the incisive duct (6). 7. Ventral turbinate; 8. Palate.
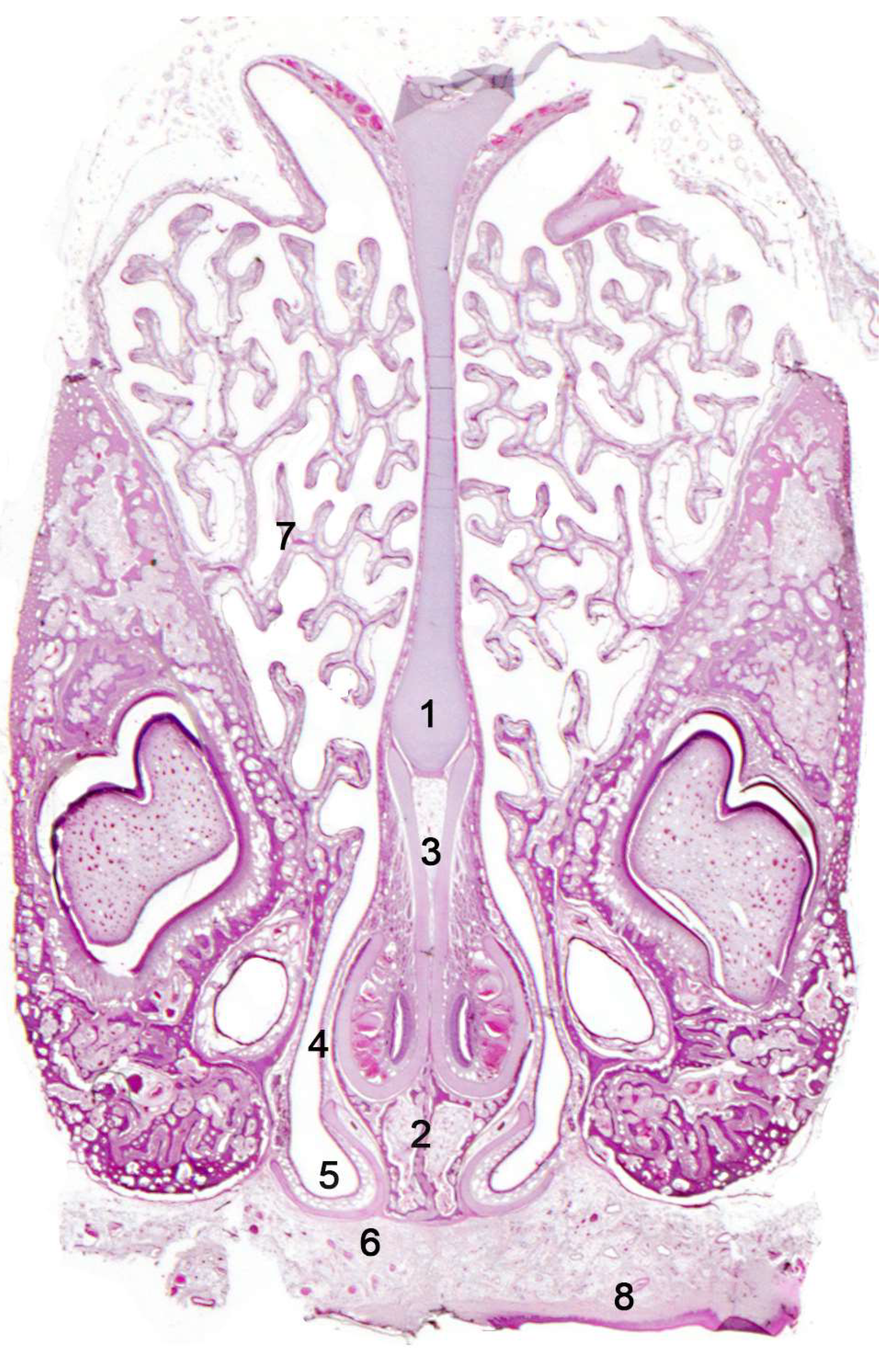
Figure 12.
The vomeronasal organ of the fox. Transverse section of the base of the nasal septum (NS) showing the topographic relationships of the vomeronasal organs (arrows) with the ventral recesses of the nasal cavity (arrowheads). CT, Canine tooth; SP, Soft palate; Vm, Vomer bone.
Figure 12.
The vomeronasal organ of the fox. Transverse section of the base of the nasal septum (NS) showing the topographic relationships of the vomeronasal organs (arrows) with the ventral recesses of the nasal cavity (arrowheads). CT, Canine tooth; SP, Soft palate; Vm, Vomer bone.
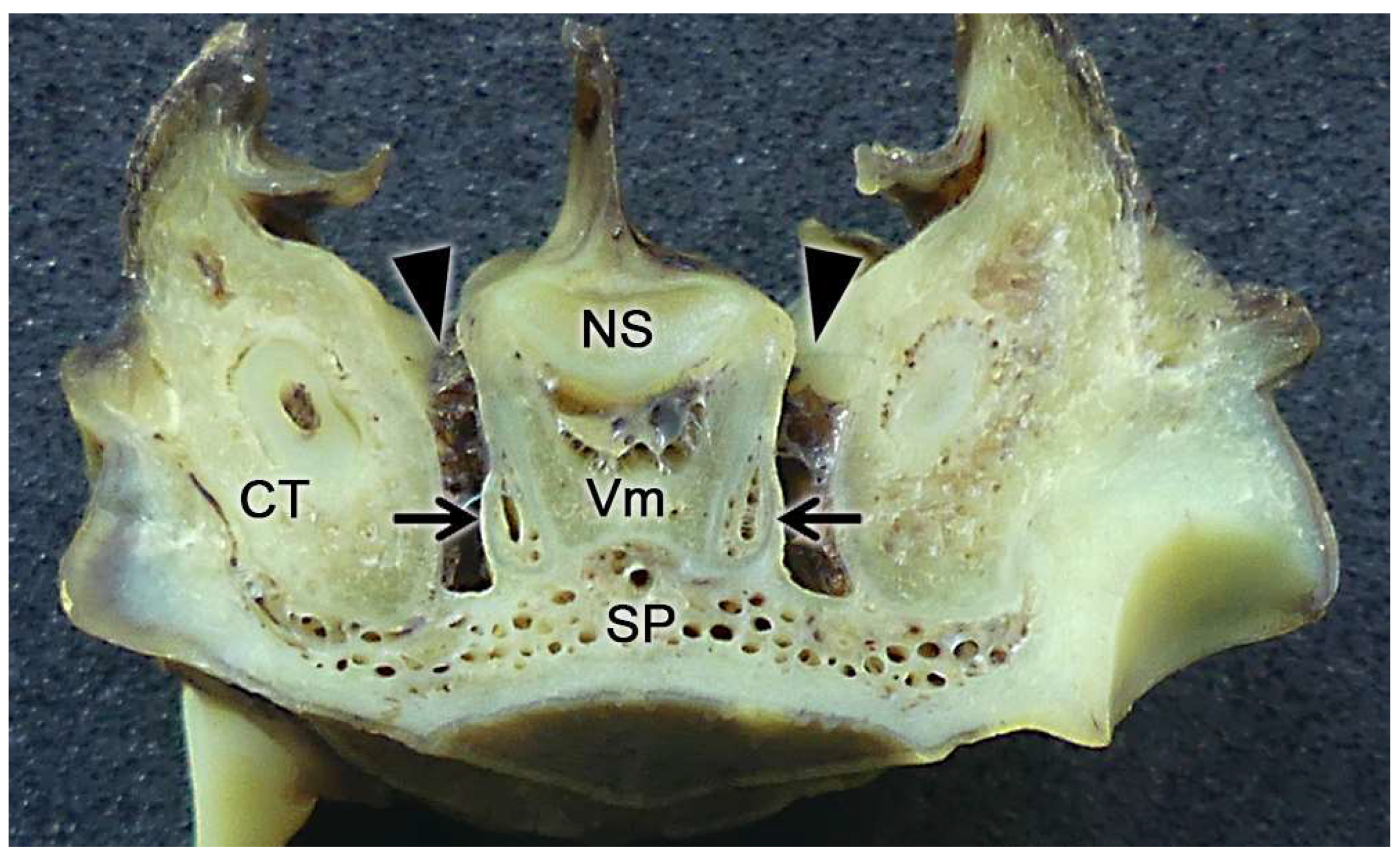
Figure 13.
The accessory olfactory bulb of the postnatal mouse. Confocal double immunofluorescence of a sagittal section stained with anti-MAP2 (green) and GFAP (red) showing the lamination of the AOB. 1. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral plexiform layer; 4. Somas of mitral cells appear as unstained circular structures; 5. Lateral olfactory trac; 6. Granular cells layer. FL, Frontal lobe.
Figure 13.
The accessory olfactory bulb of the postnatal mouse. Confocal double immunofluorescence of a sagittal section stained with anti-MAP2 (green) and GFAP (red) showing the lamination of the AOB. 1. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral plexiform layer; 4. Somas of mitral cells appear as unstained circular structures; 5. Lateral olfactory trac; 6. Granular cells layer. FL, Frontal lobe.
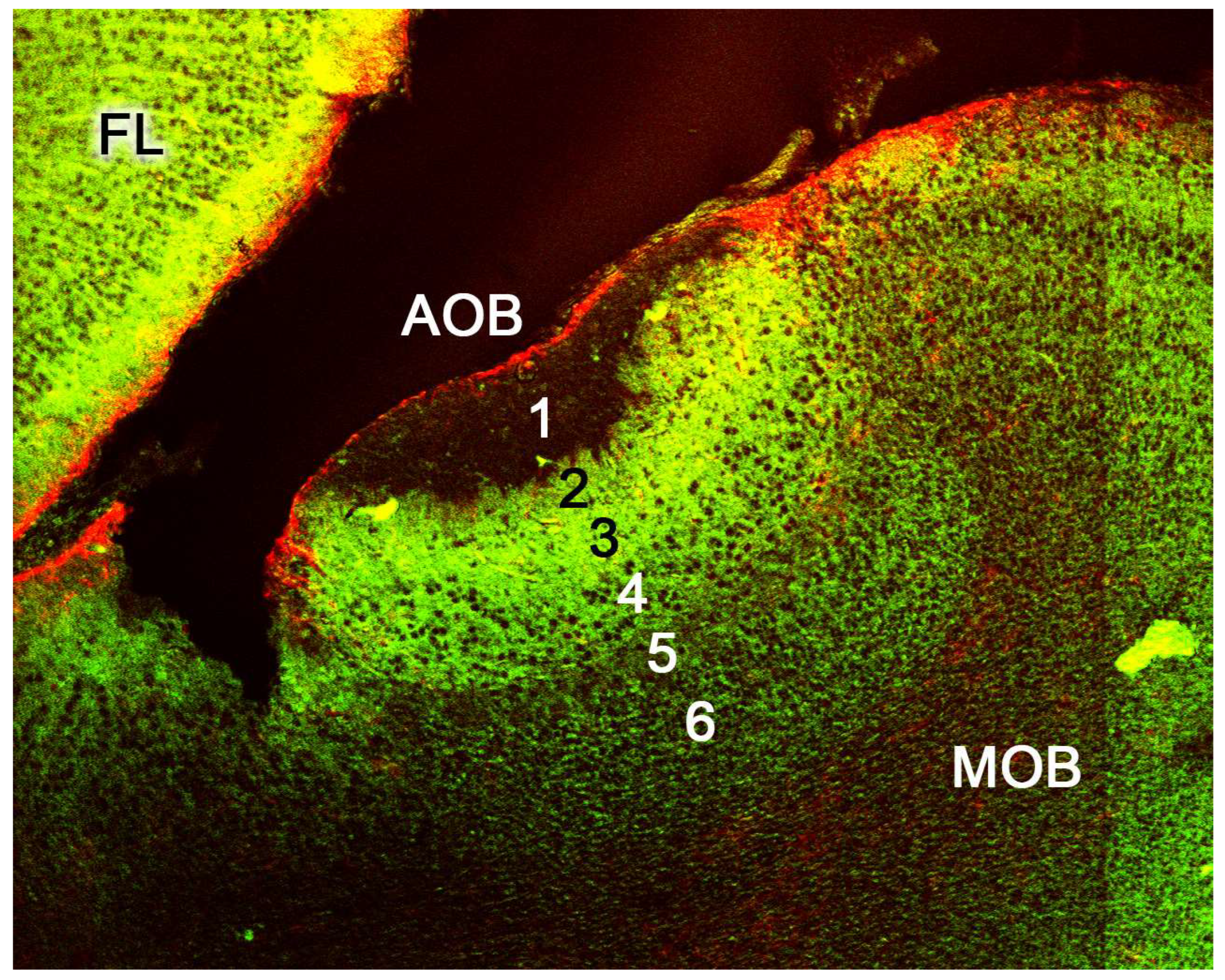
Figure 14.
The accessory olfactory bulb of the mouse. Immunohistochemical staining against MAP2. FL. Frontal lobe; GlL. Glomerular layer; GrL. Granular layer; MPL; Mitral-plexiform layer: VNL. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral plexiform layer; 4. Somas of mitral cells appear as unstained circular structures; 5. Lateral olfactory trac; 6. Granular cells layer. FL, Frontal lobe.
Figure 14.
The accessory olfactory bulb of the mouse. Immunohistochemical staining against MAP2. FL. Frontal lobe; GlL. Glomerular layer; GrL. Granular layer; MPL; Mitral-plexiform layer: VNL. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral plexiform layer; 4. Somas of mitral cells appear as unstained circular structures; 5. Lateral olfactory trac; 6. Granular cells layer. FL, Frontal lobe.
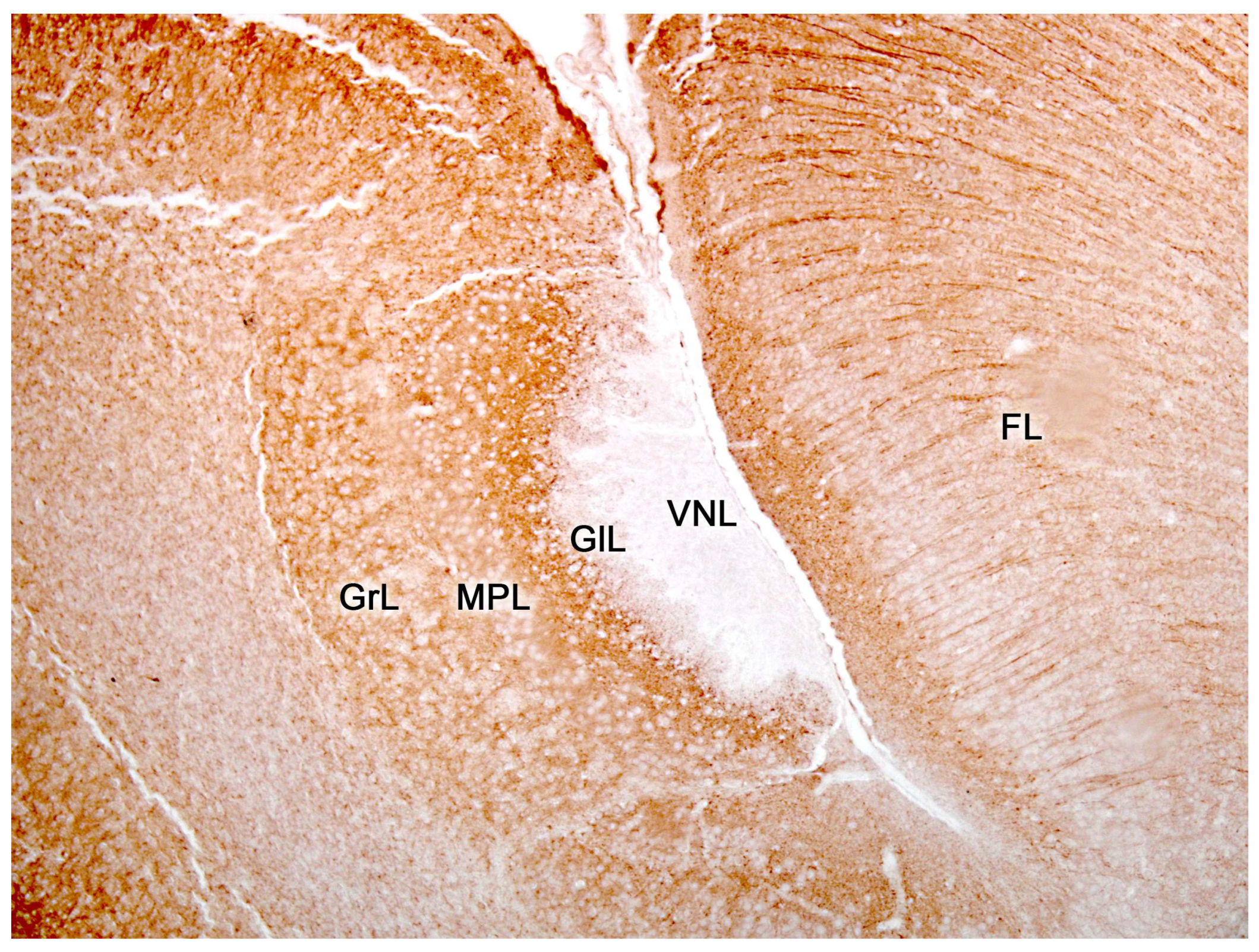
Figure 15.
The accessory olfactory bulb of the cat. Nissl staining of a histological transverse section. The red box shows the topographic relationship of the olfactory bulb and the telencephalon frontal lobe. At higher magnification the yellow area containing the AOB allows to differentiate the lanination of the AOB. 1. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral-plexiform layer; 4. Granular cell layer.
Figure 15.
The accessory olfactory bulb of the cat. Nissl staining of a histological transverse section. The red box shows the topographic relationship of the olfactory bulb and the telencephalon frontal lobe. At higher magnification the yellow area containing the AOB allows to differentiate the lanination of the AOB. 1. Vomeronasal nerve layer; 2. Glomerular layer; 3. Mitral-plexiform layer; 4. Granular cell layer.
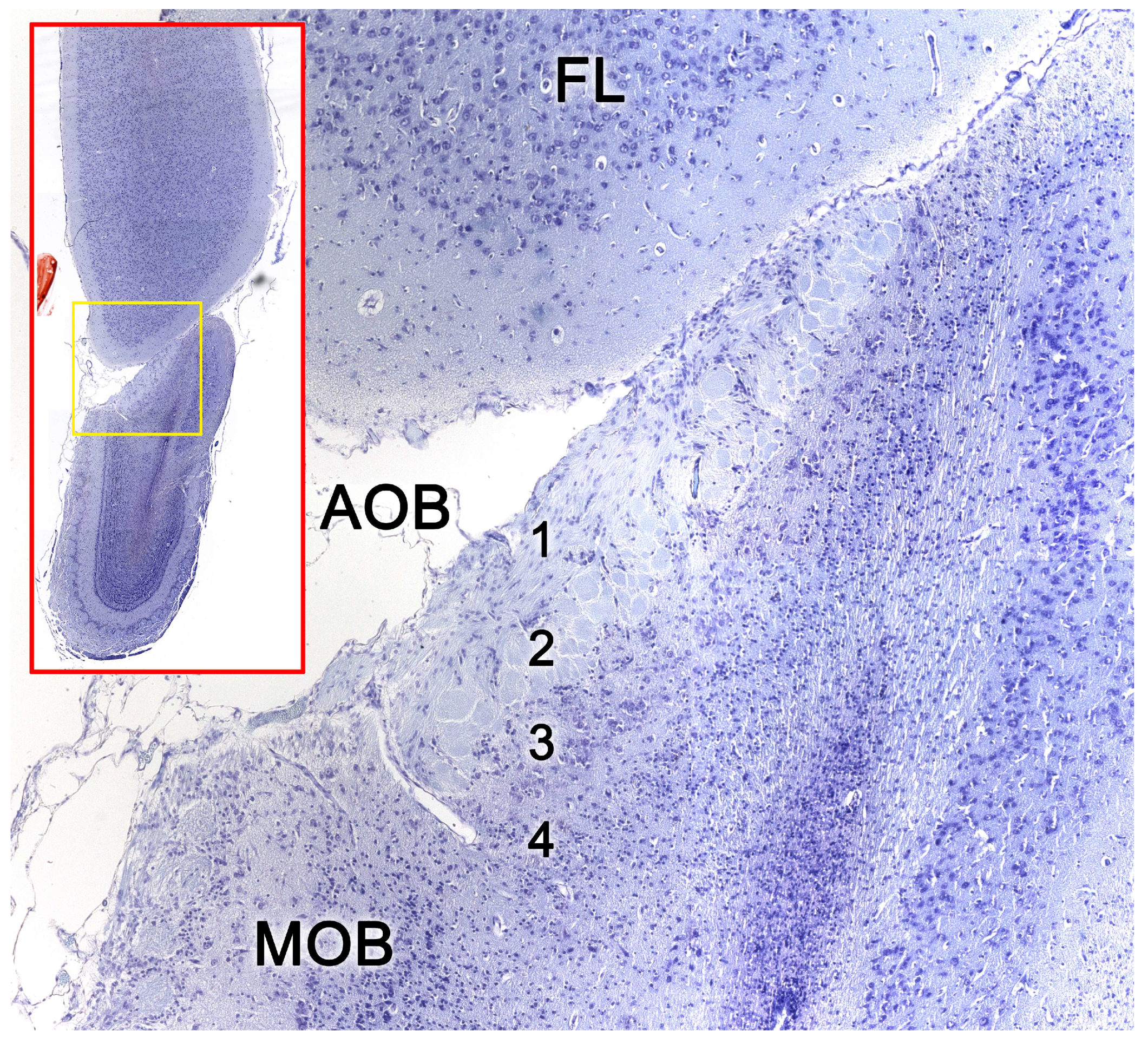
Figure 16.
The accessory olfactory bulb of the sheep. Consecutive Nissl (A) and Tolivia (B) stainings of a sagittal section. The red box shows the topographic relationship of the olfactory bulb and the telencephalon frontal lobe. At higher magnification the yellow area containing the AOB allows to differentiate the lanination of the AOB. 1. Duramater; 2. Vomeronasal nerve layer; 3. Glomerular layer; 4. Mitral-plexiform layer; 5. Lateral olfactory tract; 6. Granular cell layer.
Figure 16.
The accessory olfactory bulb of the sheep. Consecutive Nissl (A) and Tolivia (B) stainings of a sagittal section. The red box shows the topographic relationship of the olfactory bulb and the telencephalon frontal lobe. At higher magnification the yellow area containing the AOB allows to differentiate the lanination of the AOB. 1. Duramater; 2. Vomeronasal nerve layer; 3. Glomerular layer; 4. Mitral-plexiform layer; 5. Lateral olfactory tract; 6. Granular cell layer.
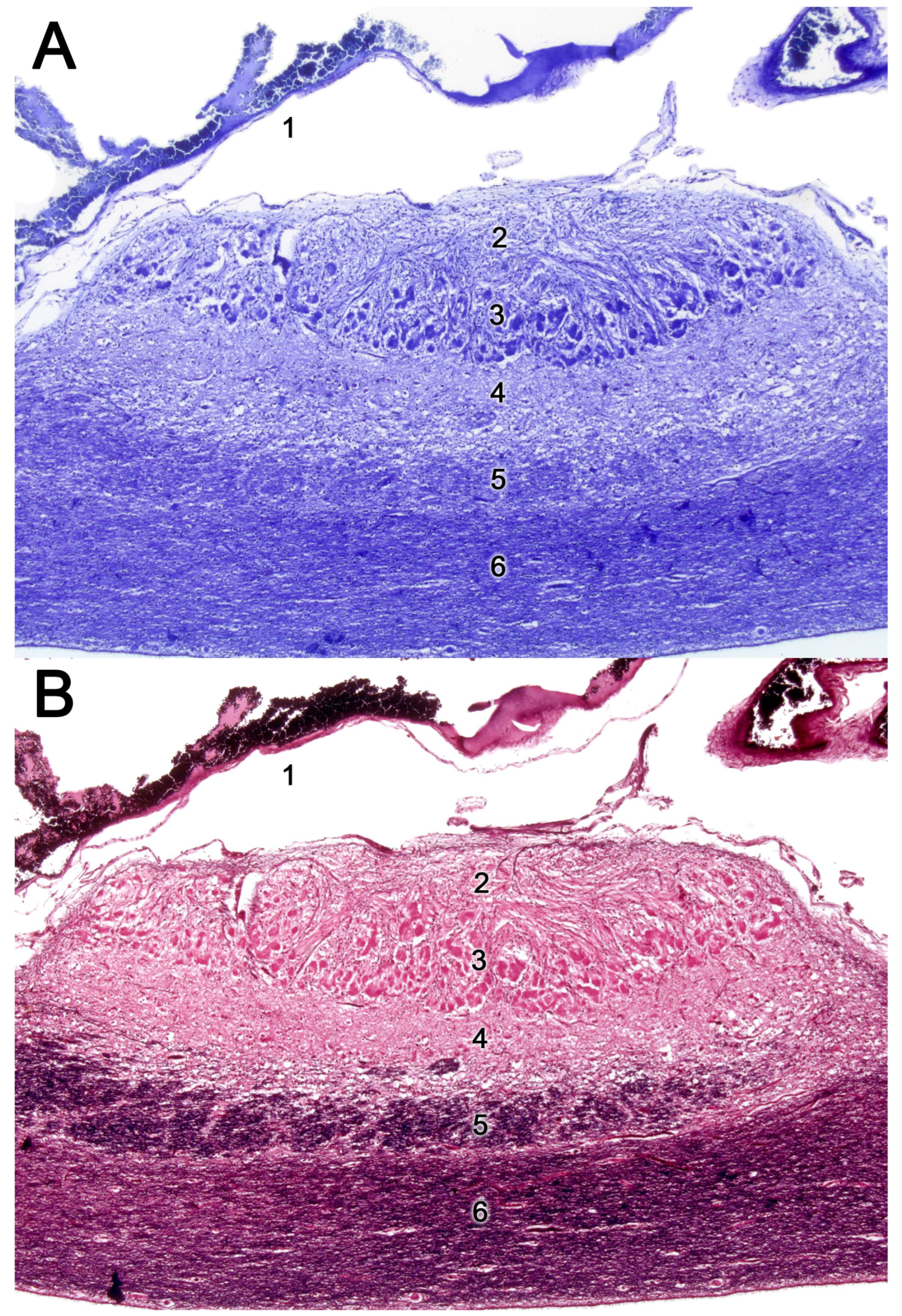
Figure 17.
The vomeronasal organ of the capybara. Transverse decalcified histological section stained with hematoxylin-eosin. RE. Respiratory epithelium; SE. Sensory epithelium; VC. Vomeronasal cartilage; VD. Vomeronasal duct; VG. Vomeronasal glands; VD. Vomeronasal duct; Vm. Vomer bone; Vv. Veins.
Figure 17.
The vomeronasal organ of the capybara. Transverse decalcified histological section stained with hematoxylin-eosin. RE. Respiratory epithelium; SE. Sensory epithelium; VC. Vomeronasal cartilage; VD. Vomeronasal duct; VG. Vomeronasal glands; VD. Vomeronasal duct; Vm. Vomer bone; Vv. Veins.
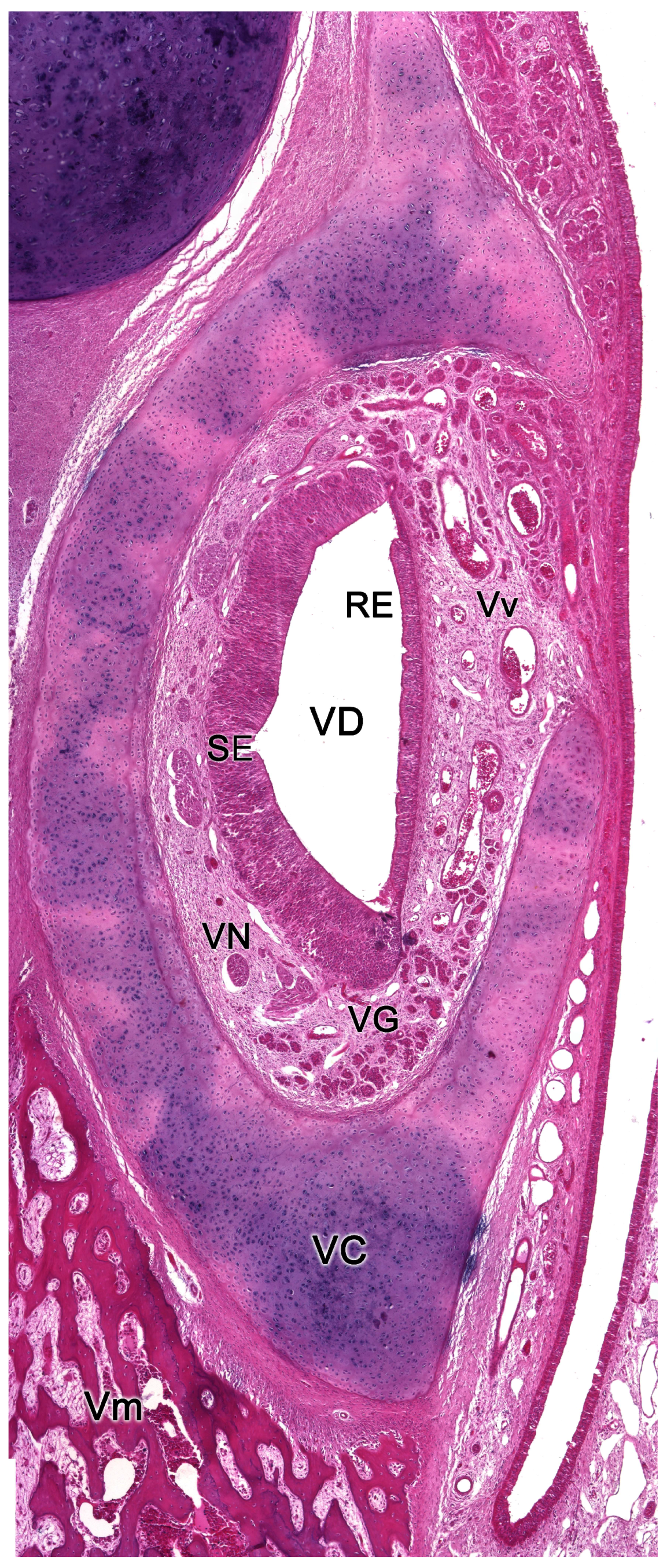
Figure 18.
Histological section of the capybara VNO stained with hematoxylin-eosin. A migratory stream (arrows) departing from the sensory epithelium (SE) to the vomeronasal axons (VN) can be observed.
Figure 18.
Histological section of the capybara VNO stained with hematoxylin-eosin. A migratory stream (arrows) departing from the sensory epithelium (SE) to the vomeronasal axons (VN) can be observed.
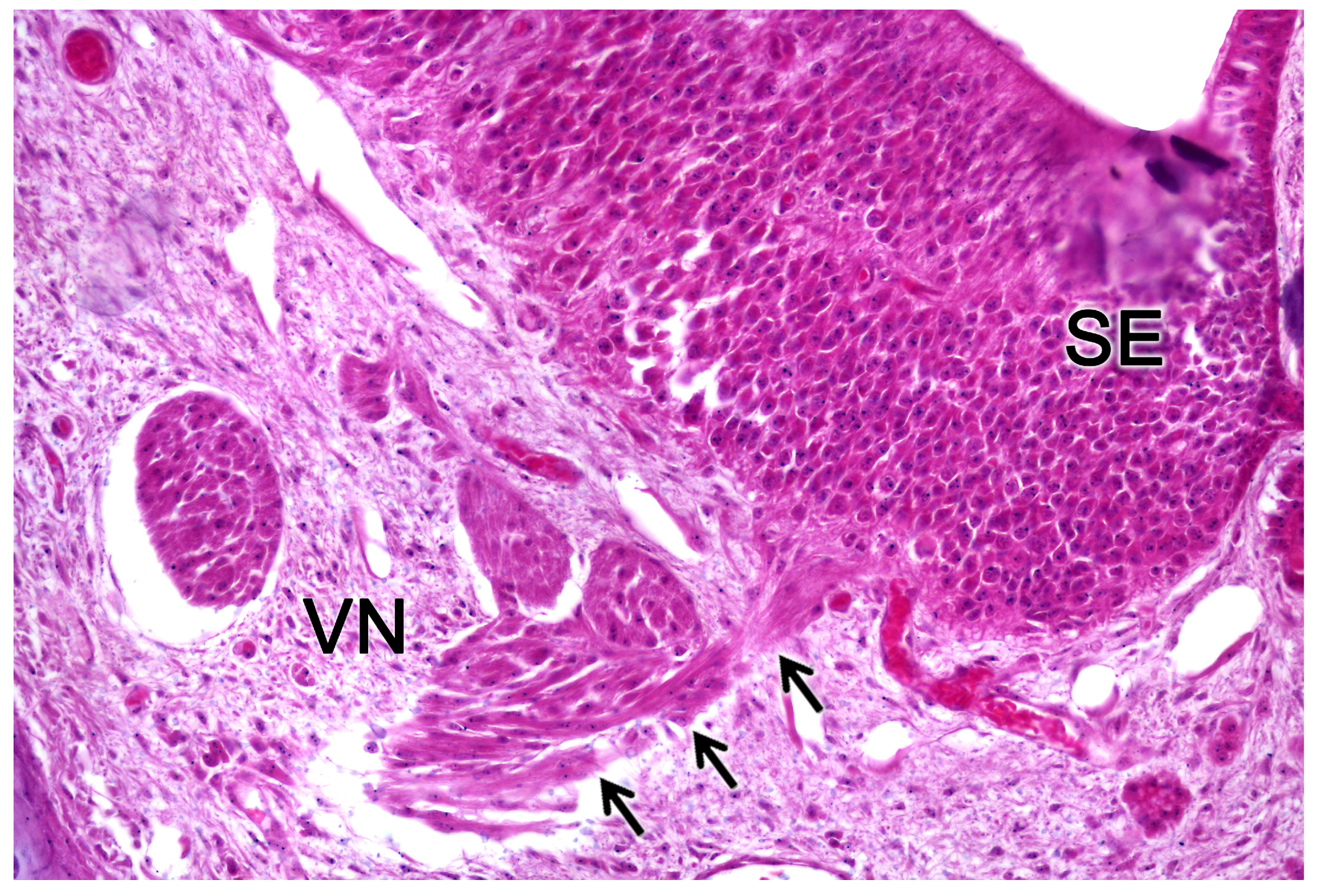
Figure 19.
Immunohistochemical staining of the capybara AOB labelled with anti-GAP43 shows strong immunolabelling in the superficial layers: Vomeronasal nerve (VNL) and glomerular (GlL) layers. GrL. Granular layer; LOT. Lateral olfactory tract; LV; lateral ventricle; MPL. Mitral-plexiform layer; WM. White matter.
Figure 19.
Immunohistochemical staining of the capybara AOB labelled with anti-GAP43 shows strong immunolabelling in the superficial layers: Vomeronasal nerve (VNL) and glomerular (GlL) layers. GrL. Granular layer; LOT. Lateral olfactory tract; LV; lateral ventricle; MPL. Mitral-plexiform layer; WM. White matter.
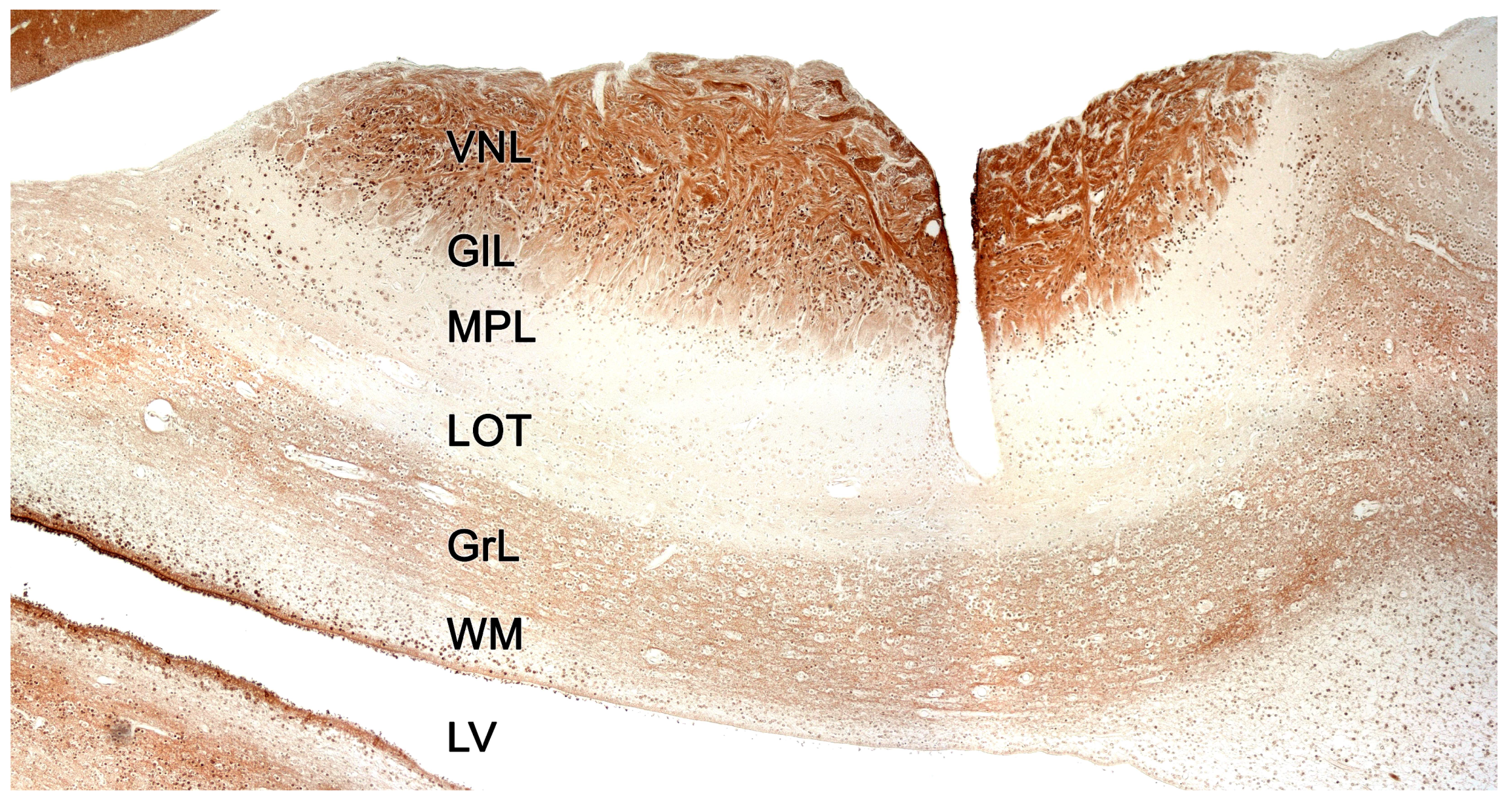
Figure 20.
Hematoxylin-eosin histological staining of the wallaby vomeronasal duct. RE. Respiratory epithelium; SE. Sensory epithelium; VN. Vomeronasal nerve.
Figure 20.
Hematoxylin-eosin histological staining of the wallaby vomeronasal duct. RE. Respiratory epithelium; SE. Sensory epithelium; VN. Vomeronasal nerve.
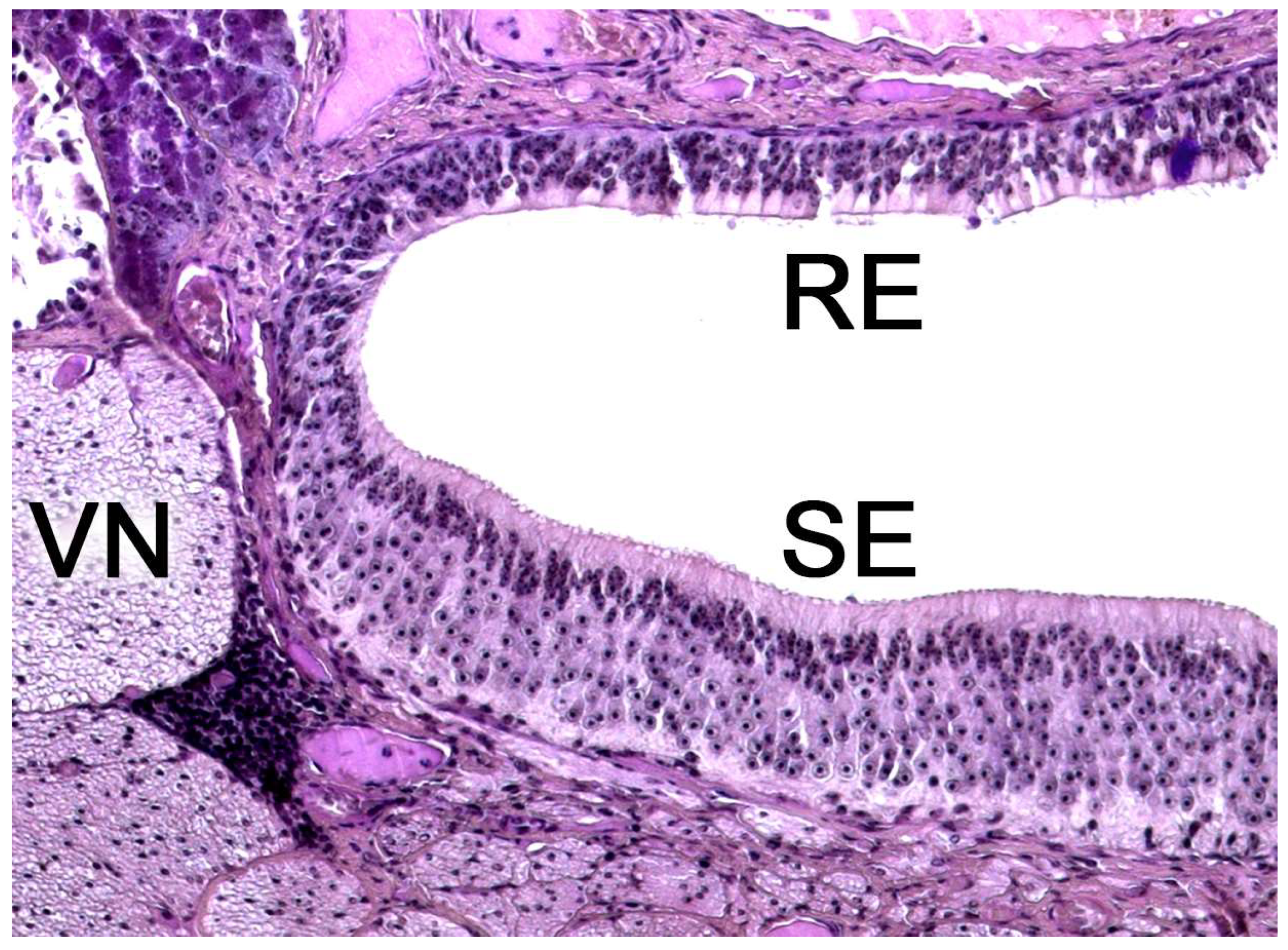
Figure 21.
PAS staining of the wallaby VNO showing a high density of PAS+ glandular acini (VG) n the dorsolateral part of the parenchyma. The vomeronasal duct contains PAS+ secretion. RE. Respiratory epithelium; RM. Respiratory mucosa; SE. Sensory epithelium; VN. Vomeronasal nerve; Vv. Veins.
Figure 21.
PAS staining of the wallaby VNO showing a high density of PAS+ glandular acini (VG) n the dorsolateral part of the parenchyma. The vomeronasal duct contains PAS+ secretion. RE. Respiratory epithelium; RM. Respiratory mucosa; SE. Sensory epithelium; VN. Vomeronasal nerve; Vv. Veins.
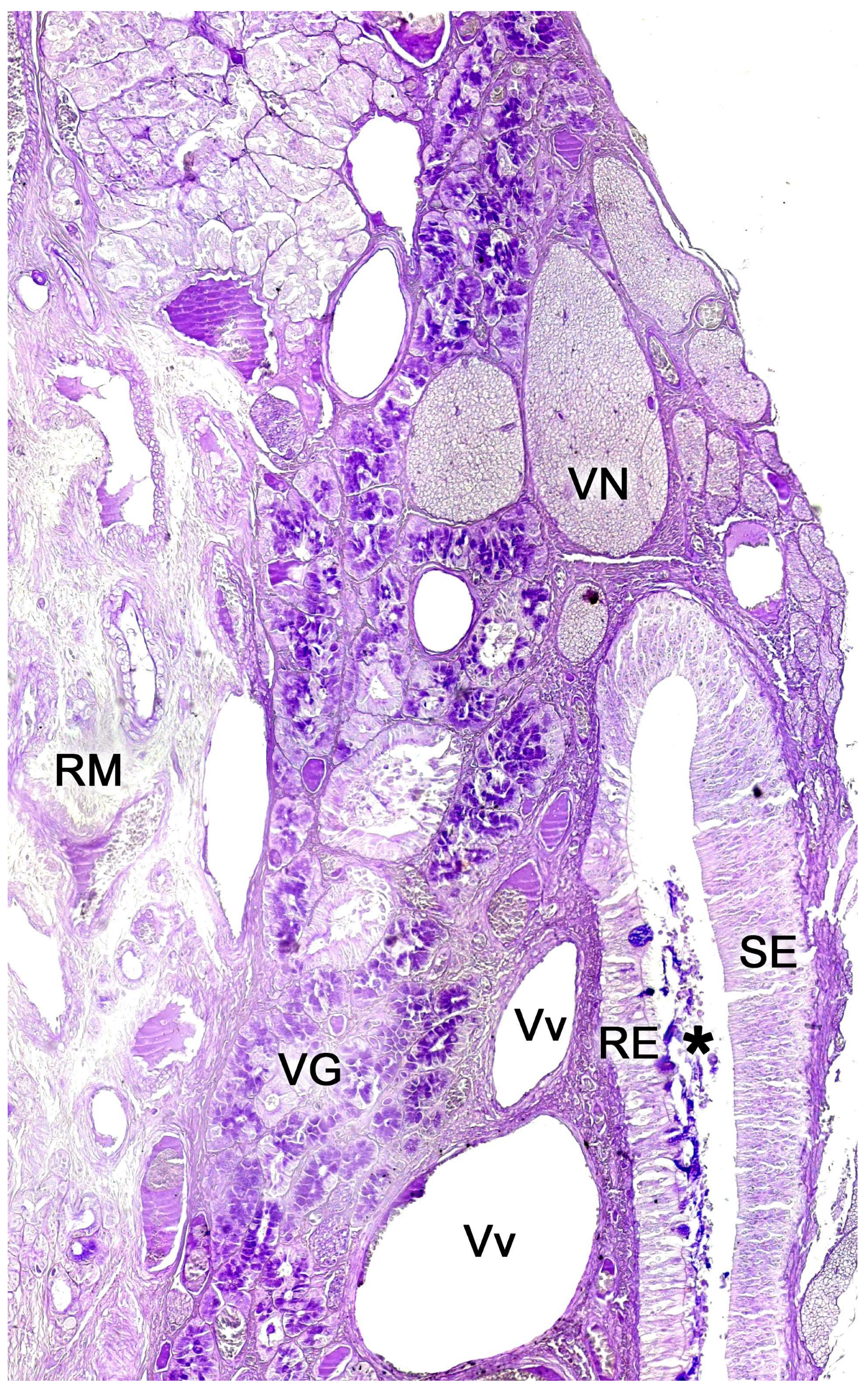
Figure 22.
Anti-calretinin immunostaining of the wallaby SE of the VNO. Neuroreceptor cells somata (open arrowheads); Neuroreceptor cells dendritic knobs (arrowheads). Intraepithelial blood vessel (asterisk).
Figure 22.
Anti-calretinin immunostaining of the wallaby SE of the VNO. Neuroreceptor cells somata (open arrowheads); Neuroreceptor cells dendritic knobs (arrowheads). Intraepithelial blood vessel (asterisk).
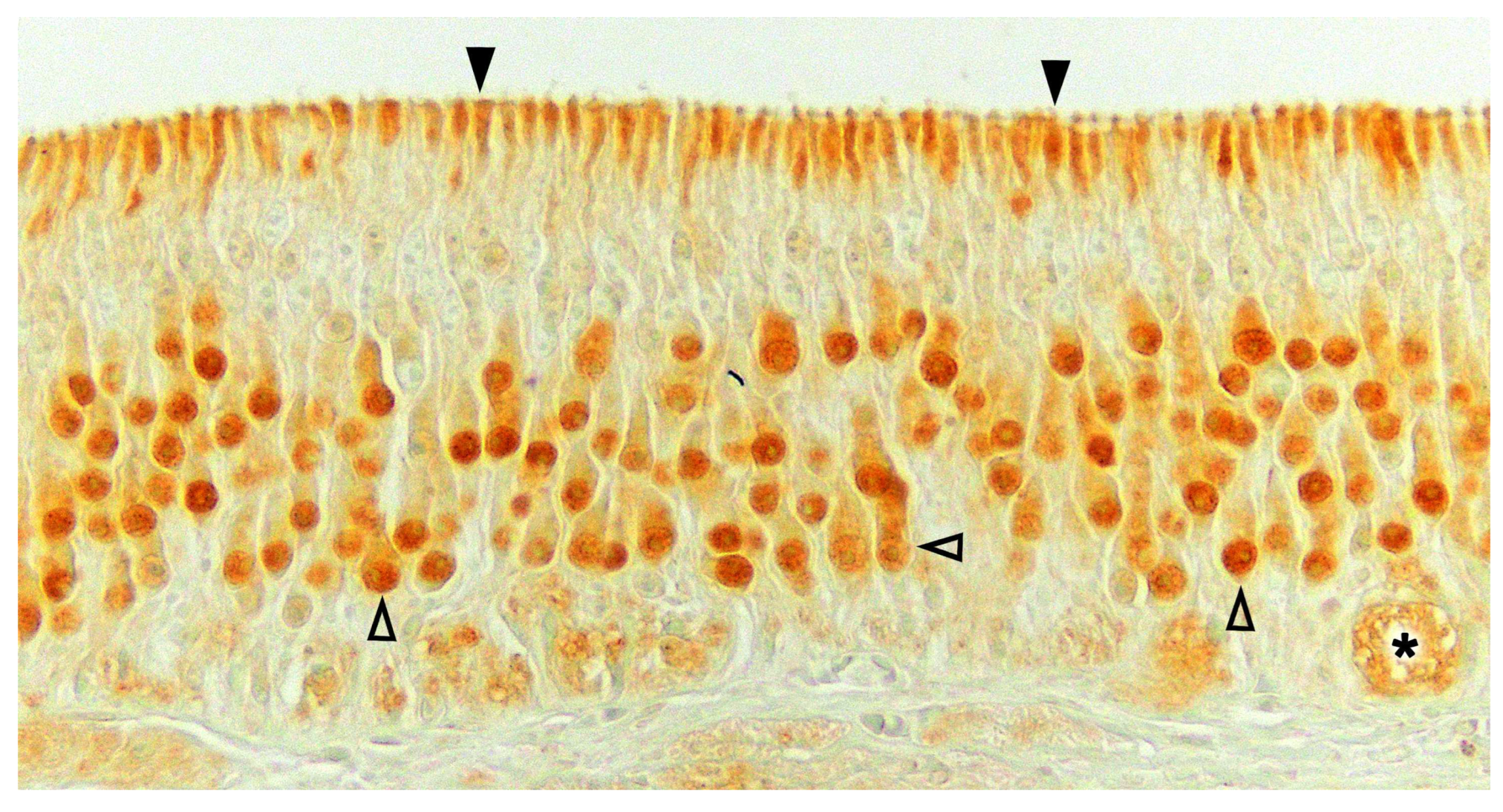
Figure 23.
UEA histochemical labelling of the wallaby VNO sensory epithelium. Neuroreceptor cells somata (open arrowheads); Neuroreceptor cells dendritic knobs (arrowheads). Dendritic processes of the neuroreceptor cells (arrows); Intraepithelial blood vessel (asterisk).
Figure 23.
UEA histochemical labelling of the wallaby VNO sensory epithelium. Neuroreceptor cells somata (open arrowheads); Neuroreceptor cells dendritic knobs (arrowheads). Dendritic processes of the neuroreceptor cells (arrows); Intraepithelial blood vessel (asterisk).

Figure 25.
Histological transverse section of the VNO of the dama gazelle showing its main components. NCd. Nasal caudal nerve; RE. Respiratory epithelium; RM. Respiratory mucosa; SE. Sensory epithelium; VD. Vomeronasal duct; VG. Vomeronasal glands; VN. Vomeronasal nerve; Vv. Veins.
Figure 25.
Histological transverse section of the VNO of the dama gazelle showing its main components. NCd. Nasal caudal nerve; RE. Respiratory epithelium; RM. Respiratory mucosa; SE. Sensory epithelium; VD. Vomeronasal duct; VG. Vomeronasal glands; VN. Vomeronasal nerve; Vv. Veins.
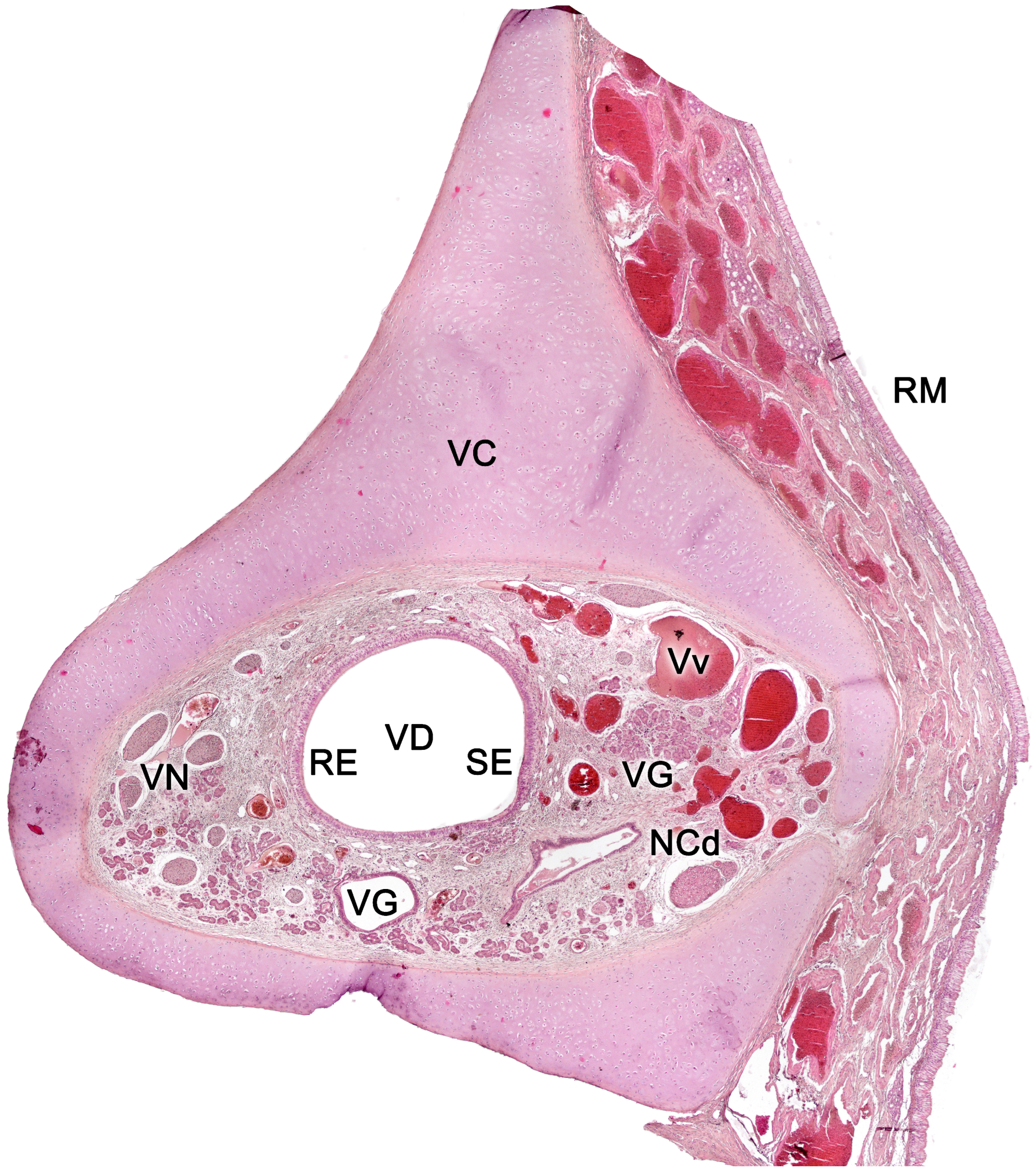
Figure 26.
Hematoxilin-eosin staining of the dama gazelle VNO sensory epithelium. Neuroreceptor cells (arrowhead); Sustentacular cells (white arrowheads): Basal cells (asterisk); MMC. Mucomicrovillar complex; SE. Sensory epithelium; VN. Vomeronasal nerve; Vv. Veins.
Figure 26.
Hematoxilin-eosin staining of the dama gazelle VNO sensory epithelium. Neuroreceptor cells (arrowhead); Sustentacular cells (white arrowheads): Basal cells (asterisk); MMC. Mucomicrovillar complex; SE. Sensory epithelium; VN. Vomeronasal nerve; Vv. Veins.
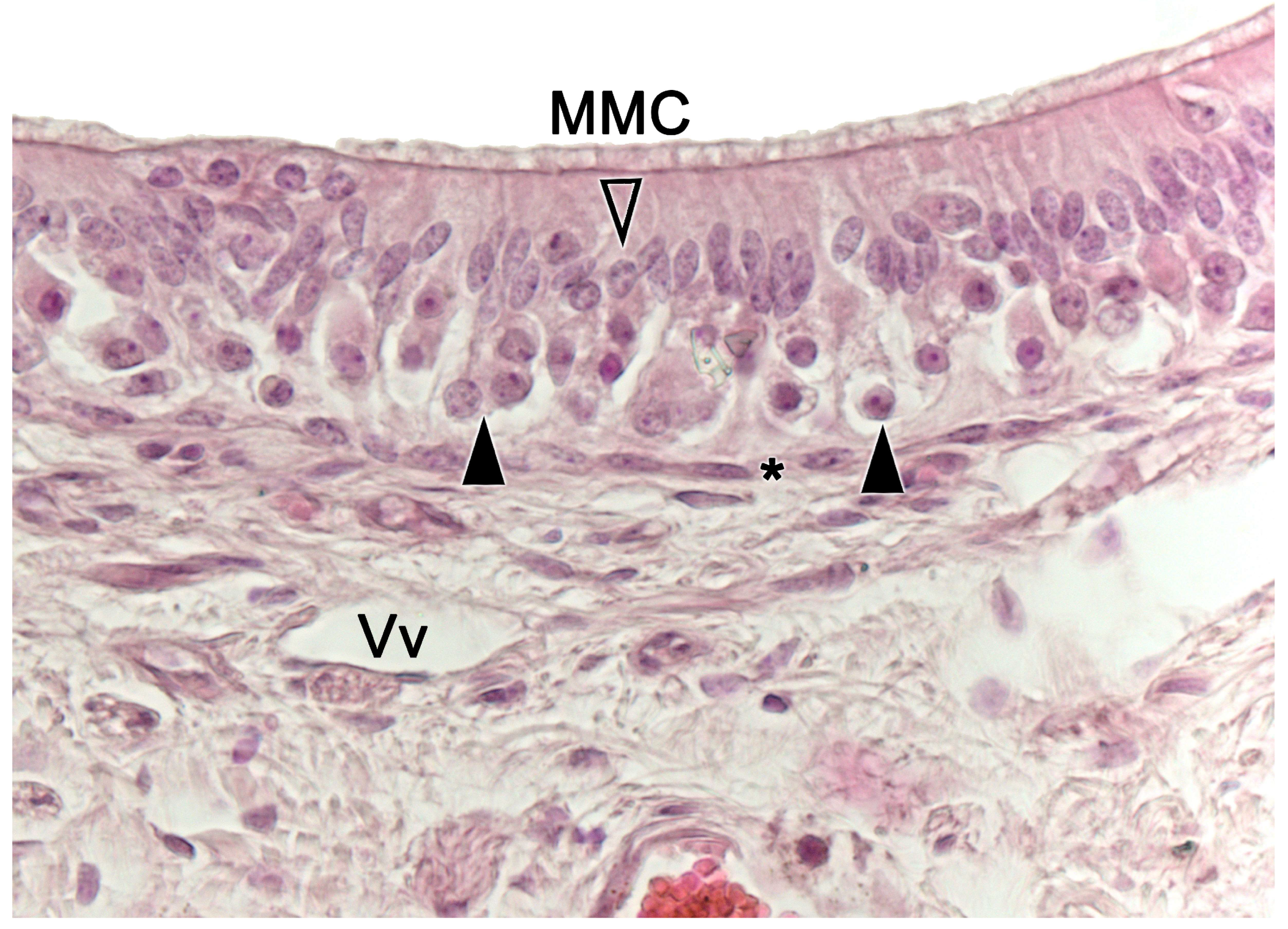
Figure 27.
PGP9.5 immunostaining of the SE of the dama gazelle VNO showing the morphology and distribution of the neuroreceptor somata, and their dendritic knobs and processes.
Figure 27.
PGP9.5 immunostaining of the SE of the dama gazelle VNO showing the morphology and distribution of the neuroreceptor somata, and their dendritic knobs and processes.
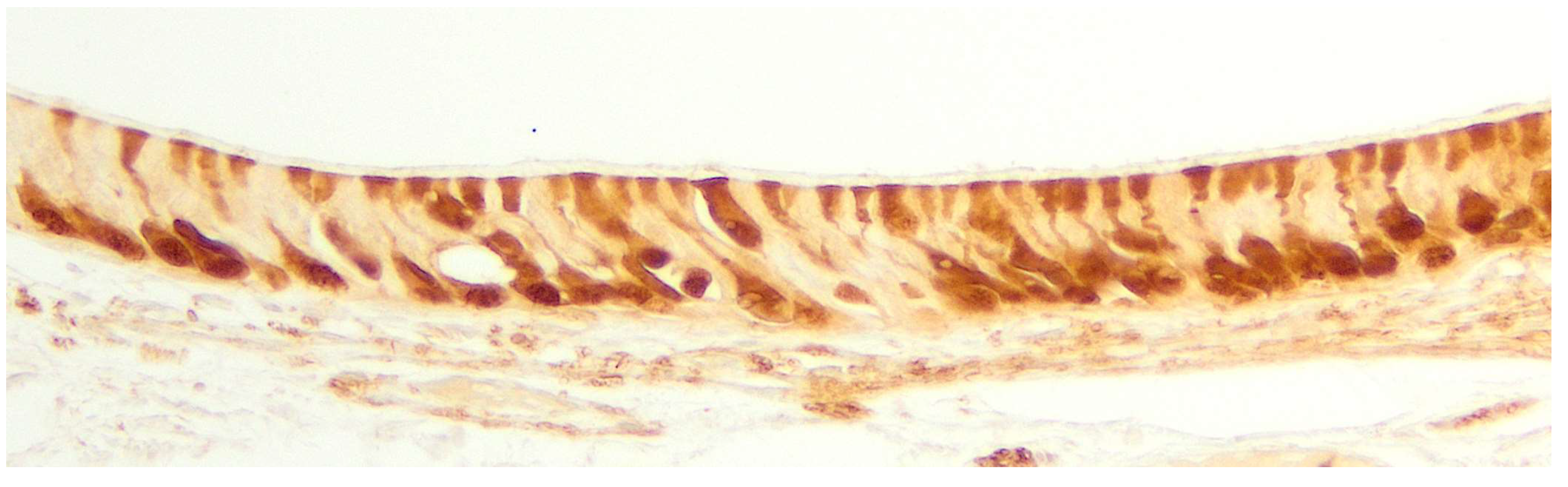
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



